Акрокантозавр
| Акрокантозавр | |
|---|---|

| |
| Установленный скелет (NCSM 14345) в Музее естественных наук Северной Каролины . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Кархародонтозавриды |
| Род: | † Акрокантозавр Стовалл и Лэнгстон , 1950 г. |
| Типовой вид | |
| † Акрокантозавр атокензис Стовалл и Лэнгстон, 1950 г.
| |
| Синонимы | |
| |
Акрокантозавр ( / ˌ æ k r oʊ ˌ k æ n θ ə ˈ s ɔːr ə s / AK -roh- KAN -thə- SOR -əs ; букв. « высокиберистая ящерица ) — род динозавров кархародонтозавридных » , существовавший в то, что сейчас является Северной Америкой, во время аптского и раннего альбского этапов раннего мела , от 113 до 110 миллионов лет назад. Как и большинство родов динозавров, Acrocanthosaurus содержит только один вид — A. atokensis . Он имел ареал по всему континенту, и ископаемые останки были известны в США штатах Оклахома , Техас и Вайоминг на западе и в Мэриленде на востоке.
Акрокантозавр был двуногим хищником . Как следует из названия, он наиболее известен высокими нервными отростками на многих позвонках , которые, скорее всего, поддерживают гребень мышц на шее, спине и бедрах животного. Акрокантозавр был одним из крупнейших тероподов: самый крупный из известных экземпляров достигал 11–11,5 метров (36–38 футов) в длину и весил примерно 4,4–6,6 метрических тонны (4,9–7,3 коротких тонны). Большие следы теропода, обнаруженные в Техасе, возможно, были оставлены акрокантозавром , хотя прямой связи с останками скелетов нет.
Недавние открытия прояснили многие детали его анатомии, что позволило провести специализированные исследования, посвященные структуре его мозга и функциям передних конечностей. Акрокантозавр был крупнейшим тероподом в своей экосистеме и, вероятно, высшим хищником , который охотился на зауроподов , орнитоподов и анкилозавров .
Открытие и присвоение имени
[ редактировать ]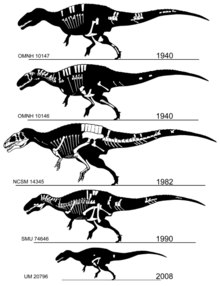
Акрокантозавр назван в честь его высоких нервных шипов, от греческого ɑκρɑ / akra («высокий»), ɑκɑνθɑ / akantha («шип» или «позвоночник») и σɑʊρος / sauros («ящерица»). [ 1 ] Существует один названный вид ( A. atokensis ), названный в честь округа Атока в Оклахоме , где были найдены оригинальные экземпляры. Название было придумано в 1950 году американскими палеонтологами Дж. Уиллисом Стоваллом и Ванном Лэнгстоном-младшим. [ 2 ] Лэнгстон предложил название «Aracanthus atokaensis» для рода и вида в своей неопубликованной магистерской диссертации 1947 года . [ 3 ] [ 4 ] название было изменено на Acrocanthosaurus atokensis . но для официальной публикации [ 2 ]
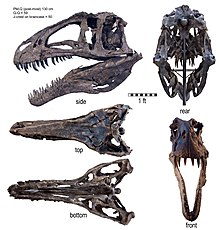
Голотип 10146 и OMNH 10147), обнаруженные в начале 1940-х годов и описанные и паратип ( OMNH одновременно в 1950 году, состоят из двух частичных скелетов и куска материала черепа из формации Антлерс в Оклахоме. [ 2 ] Два гораздо более полных экземпляра были описаны в 1990-х годах. Первый ( SMU 74646) представляет собой частичный скелет, в котором отсутствует большая часть черепа, обнаруженный в формации Твин-Маунтинс в Техасе и в настоящее время являющийся частью коллекции Музея науки и истории Форт-Уэрта . [ 5 ] Еще более полный скелет ( NCSM 14345, по прозвищу «Фрэн») был обнаружен в формации Антлерс в Оклахоме Сефисом Холлом и Сидом Лавом, подготовлен Институтом Блэк-Хиллз в Южной Дакоте и сейчас хранится в Музее естественных наук Северной Каролины . наук в Роли . Образец является самым крупным и включает единственные известные целые череп и переднюю конечность. Скелетные элементы OMNH 10147 почти такого же размера, как и сопоставимые кости в NCSM 14345, что указывает на животное примерно такого же размера, тогда как голотип и SMU 74646 значительно меньше. [ 6 ]
Присутствие акрокантозавра в формации клеверли было установлено в 2012 году описанием еще одного частичного скелета, UM 20796. Экземпляр, состоящий из частей двух позвонков, частичных лобковых костей , бедренной кости , частичной малоберцовой кости и фрагментов, представляет собой ювенильную особь. животное. произошел из костного ложа в бассейне Бигхорн на северо-центральной части Вайоминга и был найден возле лопатки зауропосейдона Он . Ряд других фрагментарных останков теропод из формации также может принадлежать Acrocanthosaurus , который может быть единственным крупным тероподом в формации Клеверли. [ 7 ]
В отличие от многих других родов динозавров, а тем более крупных теропод, акрокантозавр населял как западные, так и восточные регионы североамериканского континента. Присутствие этого рода в формации Арундел в Мэриленде (примерно одновременной с западными формациями) уже давно подозревалось, поскольку зубы у нее были почти идентичны зубам акрокантозавра, ранее известным из этой формации. [ 8 ] В 2024 году неполный скелет теропода (USNM 466054) из формации Арундел был идентифицирован как скелет полувзрослого акрокантозавра , известного как A. cf. atokensis , что стало первой точной находкой этого рода в восточной части Северной Америки. Этот скелет, наиболее полностью известный экземпляр теропода из формации, несмотря на его фрагментарный характер, до этого исследования был ранее идентифицирован как орнитомимозавр , а также представляет собой самую маленькую известную особь этого рода. [ 9 ]
Акрокантозавр может быть известен по менее полным останкам за пределами Оклахомы, Техаса, Вайоминга и Мэриленда. зуб из южной Аризоны . К роду отнесли [ 10 ] и соответствующие следы зубов были обнаружены на костях зауроподов из той же области. [ 11 ] Многие другие зубы и кости из различных геологических формаций на западе Соединенных Штатов также были отнесены к акрокантозавру , но большинство из них были идентифицированы ошибочно; [ 12 ] однако существует некоторое разногласие с этой оценкой в отношении окаменелостей из формации Клеверли. [ 7 ]
Описание
[ редактировать ]
Акрокантозавр был одним из крупнейших известных теропод: предполагаемая длина черепа 1,23–1,29 м (4,0–4,2 фута) и длина тела 11–11,5 м (36–38 футов) на основе самого крупного известного экземпляра (NCSM 14345). . [ 13 ] [ 6 ] [ 14 ] На основе различных методов исследователи получили оценки массы тела этого экземпляра от 4,4 до 6,6 метрических тонн (от 4,9 до 7,3 коротких тонны). [ 13 ] [ 15 ] [ 16 ]
Череп
[ редактировать ]
Череп акрокантозавра , как и большинства других аллозавроидов , был длинным, низким и узким. Отверстие для уменьшения веса перед глазницей ( antorbital fenestra ) было довольно большим, более четверти длины черепа и двух третей его высоты. Внешняя поверхность верхней челюсти (кость верхней челюсти) и верхняя поверхность носовой кости на нёбе не имели такой грубой текстуры, как у гиганотозавра или кархародонтозавра . Длинные, низкие гребни отходили от носовых костей, идя вдоль каждой стороны морды от ноздри обратно к глазу, где они переходили на слезные кости . [ 6 ] Это характерная особенность всех аллозавроидов. [ 17 ] В отличие от аллозавра , на слезной кости перед глазом не было заметного гребня. Слезные и заглазничные кости встретились, образуя толстую бровь над глазом, как это видно у кархародонтозаврид и неродственных им абелизаврид . На каждой стороне верхней челюсти расположено девятнадцать изогнутых зазубренных зубов , но количество зубов на нижней челюсти не опубликовано. Зубы акрокантозавра были шире, чем у кархародонтозавра , и не имели морщинистой текстуры, характерной для кархародонтозаврид. Дентария гиганотозавра (несущая зубы кость нижней челюсти) была квадратной по переднему краю, как у , и неглубокой, в то время как остальная часть челюсти позади нее стала очень глубокой. У акрокантозавра и гиганотозавра был общий толстый горизонтальный гребень на внешней поверхности надугловой кости нижней челюсти, под сочленением с черепом. [ 6 ]
Посткраниальный скелет
[ редактировать ]
Наиболее примечательной особенностью акрокантозавра был ряд высоких нервных отростков, расположенных на позвонках шеи, спины, бедер и верхней части хвоста, которые могли более чем в 2,5 раза превышать высоту позвонков, от которых они отходили. [ 2 ] У других динозавров также были высокие шипы на спине, иногда значительно выше, чем у акрокантозавра . Например, африканский род Spinosaurus имел шипы высотой почти 2 м (6,6 фута), что примерно в 11 раз выше тел его позвонков. [ 18 ] Нижние шипы акрокантозавра имели крепления для мощных мышц, подобных тем, что есть у современного бизона , вероятно, образующие высокий толстый гребень на его спине. [ 2 ] Функция шипов остается неизвестной, хотя они, возможно, участвовали в общении , хранении жира , контроле мышц или температуры . Все его шейные (шея) и спинные (спинные) позвонки имели заметные углубления ( плевроцели по бокам ), а хвостовые (хвостовые) позвонки имели более мелкие. Это больше похоже на кархародонтозаврид, чем на аллозавра . [ 5 ]
Помимо позвонков, акрокантозавр имел типичный аллозавроидный скелет. Акрокантозавр был двуногим, с длинным тяжелым хвостом, уравновешивающим голову и тело, удерживая центр тяжести над бедрами. Его передние конечности были относительно короче и крепче, чем у аллозавра , но в остальном были похожи: на каждой руке было по три когтистых пальца. В отличие от многих более мелких и быстро бегающих динозавров, его бедренная кость была длиннее голени и плюсневых костей . [ 5 ] [ 6 ] предполагая, что акрокантозавр не был быстрым бегуном. [ 19 ] Неудивительно, что кости задних ног акрокантозавра были пропорционально более крепкими, чем у его меньшего родственника аллозавра . Ступни у него имели по четыре пальца, хотя, как это типично для теропод, первый был значительно меньше остальных и не соприкасался с землей. [ 5 ] [ 6 ]
Классификация и систематика
[ редактировать ]
Акрокантозавр отнесен к надсемейству Allosauroidea внутри инфраотряда Tetanurae . Это надсемейство, помимо других особенностей, характеризуется парными гребнями на носовой и слезной костях в верхней части морды и высокими нервными отростками на шейных позвонках. [ 17 ] Первоначально он был отнесен к семейству Allosauridae вместе с Allosaurus . [ 2 ] эта договоренность также подтверждается исследованиями, проведенными еще в 2000 году. [ 6 ] Большинство исследований показали, что он является членом родственного семейства Carcharodontosauridae. [ 17 ] [ 20 ] [ 21 ]
На момент открытия акрокантозавр и большинство других крупных теропод были известны лишь по фрагментарным останкам, что привело к весьма разнообразным классификациям этого рода. Дж. Уиллис Стовалл и Ванн Лэнгстон-младший сначала отнесли его к «Antrodemidae», эквиваленту Allosauridae , перенес его в Megalosauridae , таксон мусорной корзины . Альфред Шервуд Ромер но в 1956 году [ 22 ] По мнению других авторов, длинные шипы на его позвонках предполагали родство со спинозавром . [ 23 ] [ 24 ] Эта интерпретация акрокантозавра как спинозавра сохранялась до 1980-х годов. [ 25 ] и был повторен в полутехнических книгах о динозаврах того времени. [ 26 ] [ 27 ]
высокие шипованные позвонки из раннего мела Англии Когда-то считалось, что очень похожи на позвонки акрокантозавра . [ 28 ] а в 1988 году Грегори С. Пол назвал их вторым видом рода, A. altispinax . [ 29 ] Эти кости первоначально были отнесены к Altispinax , английскому тероподу, известному только по зубам, и это отнесение привело по крайней мере к одному автору, предположившему, что Altispinax сам по себе был синонимом Acrocanthosaurus . [ 28 ] Эти позвонки позже были отнесены к новому роду Becklespinax , отдельному как от Acrocanthosaurus , так и от Altispinax . [ 30 ]
Большинство кладистических анализов, включая акрокантозавра, показали, что это кархародонтозавр, обычно занимающий базальное положение относительно кархародонтозавра из Африки и гиганотозавра из Южной Америки . [ 17 ] [ 5 ] [ 31 ] Его часто считали родственным таксоном столь же базального Eocarcharia , также из Африки. Неовенатора , обнаруженного в Англии, часто считают еще более базальным кархародонтозавридом или базальным членом сестринской группы под названием Neovenatoridae . [ 19 ] [ 21 ] Это говорит о том, что семейство зародилось в Европе , а затем расселилось по южным континентам (объединенным в то время как суперконтинент Гондвана ). Если бы акрокантозавр был кархародонтозавридом, то его распространение также произошло бы в Северной Америке. [ 5 ] Все известные кархародонтозавриды жили в раннем и среднем меловом периоде. [ 17 ] В 2011 году Оливер Раухут назвал новый род динозавров-теропод из формации Тендагуру юрского периода в Танзании именем Veterupristisaurus и обнаружил, что он является родственным таксоном Acrocanthosaurus , что еще больше подтверждает его позицию как кархародонтозаврид. [ 32 ] Следующая кладограмма по Novas et al., 2013 показывает место Acrocanthosaurus среди Carcharodontosauridae. [ 33 ]

Кау (2024) получил аналогичные результаты для акрокантозавра . [ 34 ] Его результаты показаны ниже.
| Кархародонтозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Рост и долголетие
[ редактировать ]
Судя по особенностям костей голотипов OMNH 10146 и NCSM 14345, подсчитано, что акрокантозавру потребовалось не менее 12 лет, чтобы полностью вырасти. Это число могло быть намного выше, поскольку в процессе ремоделирования костей и роста костномозговой полости некоторые линии Харриса были утрачены. Если принять во внимание эти линии, акрокантозавру потребовалось 18–24 года, чтобы стать зрелым. [ 7 ]
Сила укуса
[ редактировать ]Сила укуса акрокантозавра была изучена и сравнена с силой укуса 33 других динозавров Сакамото и др. (2022). Согласно результатам, его сила укуса в передней части челюстей составила 8266 ньютонов, а сила заднего укуса оценивалась в 16894 ньютона. [ 35 ]
Функция передних конечностей
[ редактировать ]Как и у большинства других нептичьих теропод, передние конечности акрокантозавра не соприкасались с землей и не использовались для передвижения; вместо этого они выполняли хищническую функцию. Открытие полной передней конечности (NCSM 14345) позволило провести первый анализ функции и диапазона движений передней конечности у акрокантозавра . [ 36 ] В ходе исследования изучались поверхности костей, которые могли бы сочленяться с другими костями, чтобы определить, насколько далеко суставы могут двигаться без вывихов . Во многих суставах кости не прилегали друг к другу точно, что указывает на наличие значительного количества хряща в суставах, как это наблюдается у многих современных архозавров . Среди других результатов исследование показало, что в положении покоя передние конечности свисали с плеч, плечевая кость была слегка наклонена назад, локоть согнут, а когти были направлены медиально (внутрь). [ 36 ] Плечо акрокантозавра было ограничено в диапазоне движений по сравнению с плечом человека. Рука не могла совершать полный круг, но могла отводиться (отклоняться назад) на 109 ° от вертикали, так что плечевая кость могла фактически быть слегка наклонена вверх. Выдвижение (поворот вперед) было ограничено всего 24 ° от вертикали. Рука не могла достичь вертикального положения при приведении (повороте вниз), но могла отводить (покачиваться вверх) на 9° выше горизонтали. Движения в локтях также были ограничены по сравнению с людьми: общий диапазон движений составлял всего 57°. Рука не могла ни полностью выпрямиться (выпрямиться), ни сильно согнуться (согнуться), а плечевая кость не могла даже образовать прямой угол с предплечьем. Лучевая (скручивания) , и локтевая кости (кости предплечья) сомкнуты вместе, так что не было возможности пронации или супинации как в человеческих предплечьях. [ 36 ]

Ни одна из запястных костей (костей запястья) не прилегала друг к другу точно, что позволяет предположить наличие большого количества хрящей в запястье, которые могли бы сделать его жестким. Все пальцы могли переразгибаться (наклоняться назад), пока почти не касались запястья. При сгибании средний палец сходится к первому пальцу, а третий палец скручивается внутрь. На первом пальце руки был самый большой коготь, который был постоянно согнут так, что загибался назад к нижней стороне руки. Точно так же средний коготь мог быть постоянно согнутым, тогда как третий коготь, также самый маленький, мог как сгибаться, так и разгибаться. [ 36 ] После определения диапазона движений в суставах передних конечностей исследование продолжило выдвигать гипотезу о хищнических повадках акрокантозавра . Передние конечности не могли сильно качнуться вперед и не могли даже почесать собственную шею животного. Следовательно, маловероятно, что они использовались при первоначальном захвате добычи, а акрокантозавр, вероятно, вел рот во время охоты. С другой стороны, передние конечности были способны очень сильно прижиматься к телу. После того, как добыча была схвачена челюстями, мускулистые передние конечности могли втянуться, плотно прижимая добычу к телу и не позволяя ей сбежать. Когда животное-жертва попыталось оторваться, оно лишь еще больше пронзилось бы постоянно согнутыми когтями первых двух пальцев. Чрезвычайная сверхрастяжимость пальцев могла быть адаптацией, позволяющей акрокантозавру удерживать борющуюся добычу, не опасаясь вывиха. Как только добыча оказалась прижатой к телу, акрокантозавр, возможно, расправился с ней челюстями. Другая возможность заключается в том, что Акрокантозавр держал добычу в челюстях, неоднократно втягивая передние конечности, нанося когтями большие порезы. [ 36 ] Другие менее вероятные теории предполагают, что диапазон движений передних конечностей позволяет хвататься за бок зауропода и цепляться за него, чтобы опрокинуть зауроподов меньшего роста, хотя это маловероятно из-за того, что у акрокантозавра довольно прочная структура ног по сравнению с другими подобными структурами. тероподы.
Строение мозга и внутреннего уха
[ редактировать ]
В 2005 году ученые реконструировали эндокаст (копию) акрокантозавра полости черепа с помощью компьютерной томографии (КТ) для анализа пространств внутри черепной коробки голотипа (OMNH 10146). была бы заполнена мозговыми оболочками и спинномозговой жидкостью При жизни большая часть этого пространства , помимо самого мозга, . Однако общие особенности головного мозга и черепно-мозговых нервов можно определить по эндокастам и сравнить с другими тероподами, для которых были созданы эндокасты. Хотя мозг похож на мозг многих теропод, он больше всего похож на мозг аллозавроидов. Он больше всего похож на мозг Кархародонтозавра и Гиганотозавра, а не на мозг Аллозавра или Синраптора , что подтверждает гипотезу о том, что Акрокантозавр был кархародонтозавридом. [ 37 ]
Мозг имел слегка сигмовидную форму (S-образную форму), без особого расширения полушарий головного мозга , больше напоминал крокодиловый, чем птичий . Это соответствует общему консерватизму мозга нецелурозавровых теропод . У акрокантозавра были большие и выпуклые обонятельные луковицы , что указывало на хорошее обоняние . Реконструкция полукружных каналов уха, контролирующих равновесие , показывает, что голова удерживалась под углом 25° ниже горизонтали. Это определяли путем ориентации эндокаста так, чтобы латеральный полукружный канал был параллелен земле, как это обычно бывает, когда животное находится в позе тревоги. [ 37 ]
Возможные следы
[ редактировать ]
В формации Глен Роуз в центральном Техасе сохранилось множество следов динозавров, в том числе отпечатки больших трехпалых теропод. [ 38 ] Самая известная из этих троп была обнаружена вдоль реки Пэлюкси в государственном парке «Долина динозавров» , часть которой сейчас выставлена в Американском музее естественной истории в Нью-Йорке . [ 39 ] хотя в литературе описано несколько других мест по всему штату. [ 40 ] [ 41 ] Невозможно сказать, какое животное оставило отпечатки, поскольку со следами не было связано никаких ископаемых костей. Однако ученые уже давно считают вероятным, что следы принадлежат акрокантозавру . [ 42 ] Исследование 2001 года сравнило следы Глен Роуз со стопами различных крупных теропод, но не смогло с уверенностью отнести их к какому-либо конкретному роду. Тем не менее, исследование отметило, что размеры и форма следов находились в пределах, ожидаемых для акрокантозавра . Поскольку формация Глен-Роуз близка к формациям Антлерс и Твин-Маунтинс как по географическому положению, так и по геологическому возрасту, а единственным крупным тероподом, известным из этих формаций, является акрокантозавр , исследование пришло к выводу, что акрокантозавр, скорее всего, оставил следы. [ 43 ]
Знаменитая тропа Глен Роуз, выставленная в Нью-Йорке, включает следы теропод, принадлежащие нескольким особям, которые двигались в том же направлении, что и до двенадцати динозавров -зауроподов . Отпечатки теропод иногда находят поверх следов зауроподов, что указывает на то, что они образовались позже. Это было выдвинуто как доказательство того, что небольшая преследовала стадо стая акрокантозавров зауроподов. [ 39 ] Хотя эта гипотеза интересна и правдоподобна, ее трудно доказать, и существуют другие объяснения. Например, несколько одиночных теропод могли пройти в одном направлении в разное время после прохождения зауроподов, создавая впечатление стаи, преследующей свою добычу. То же самое можно сказать и о предполагаемом «стаде» зауроподов, которые также могли или не могли двигаться как группа. [ 44 ] В месте, где он пересекает путь одного из зауроподов, на одной из троп теропод отсутствует след, что было названо доказательством нападения. [ 45 ] Однако другие ученые сомневаются в справедливости этой интерпретации, поскольку зауропод не изменил походку, как можно было бы ожидать, если бы крупный хищник висел на его боку. [ 44 ]
Патология
[ редактировать ]
На черепе Acrocanthosaurus atokensis голотипа виден светлый экзостотический материал на чешуйчатой кости . Нервный отросток одиннадцатого позвонка был сломан и зажил, а нервный отросток третьего хвостового позвонка имел необычную крючковидную структуру. [ 46 ]
Палеоэкология
[ редактировать ]
Определенные окаменелости акрокантозавра были обнаружены в формации Твин-Маунтинс на севере Техаса, формации Антлерс на юге Оклахомы, формации Клеверли в северо-центральном Вайоминге и формации Арундел в Мэриленде. Эти геологические образования не были датированы радиометрически , но ученые использовали биостратиграфию, чтобы оценить их возраст. Судя по изменениям в аммонитов таксонах , граница между аптским и альбским этапами раннего мела была расположена в пределах формации Глен-Роуз в Техасе, которая может содержать следы акрокантозавра и находится чуть выше формации Твин-Маунтинс. Это указывает на то, что формация гор-близнецов полностью лежит в пределах аптского этапа, который длился от 125 до 112 миллионов лет назад. [ 47 ] Формация Антлерс содержит окаменелости дейнониха и тенонтозавра , двух родов динозавров, также обнаруженных в формации Клеверли, которая была радиометрически датирована аптским и альбским этапами, что позволяет предположить аналогичный возраст рогов. [ 48 ] Следовательно, акрокантозавр, скорее всего, существовал между 125 и 100 миллионами лет назад. [ 17 ]
В это время территория, сохранившаяся в формациях «Горы-близнецы» и «Антлерс», представляла собой большую пойму , которая впадала в мелкое внутреннее море . Несколько миллионов лет спустя это море расширилось на север, став Западным внутренним морским путем и разделив Северную Америку на две части почти на протяжении всего позднего мела . Присутствие акрокантозавра в формации Арундел позволяет предположить, что ему удалось распространиться по континенту до того, как морской путь смог помешать ему. [ 9 ] Формация Глен Роуз представляет собой прибрежную среду, где, возможно, следы акрокантозавра сохранились в илистых отмелях вдоль древней береговой линии. Поскольку акрокантозавр был крупным хищником, ожидается, что он имел обширный ареал обитания и обитал в самых разных средах обитания. [ 43 ] Потенциальные жертвы включают зауроподов, таких как Астродон. [ 49 ] или, возможно, даже огромный Завропосейдон , [ 50 ] а также крупных орнитоподов, таких как Тенонтозавр . [ 51 ] Меньший теропод Deinonychus также бродил по этой территории, но его длина составляла 3 м (10 футов), скорее всего, он составлял лишь минимальную конкуренцию или даже пищу для акрокантозавра . [ 48 ]
Ссылки
[ редактировать ]- ^ Лидделл, Генри Джордж; Роберт Скотт (1980). Греко-английский лексикон, сокращенное издание . Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-910207-5 .
- ^ Перейти обратно: а б с д и ж Стовалл, Дж. Уиллис ; Лэнгстон, Ванн . (1950). « Acrocanthosaurus atokensis , новый род и вид нижнемеловых теропод из Оклахомы». Американский натуралист из Мидленда . 43 (3): 696–728. дои : 10.2307/2421859 . JSTOR 2421859 .
- ^ Лэнгстон, Ванн Р. (1947). Новый род и вид меловых динозавров-теропод из Тринити округа Атока, Оклахома. Неопубликованная кандидатская диссертация . Университет Оклахомы.
- ^ Чаплевский, Николас Дж.; Чифелли, Ричард Л.; Лэнгстон, Ванн Р. младший (1994). «Каталог типов и фигурных ископаемых позвоночных. Музей естественной истории Оклахомы». Специальное издание Геологической службы Оклахомы . 94 (1): 1–35.
- ^ Перейти обратно: а б с д и ж Харрис, Джеральд Д. (1998). «Повторный анализ Acrocanthosaurus atokensis , его филогенетического статуса и палеобиологических последствий, основанный на новом образце из Техаса». Бюллетень Музея естественной истории и науки Нью-Мексико . 13 :1–75.
- ^ Перейти обратно: а б с д и ж г Карри, Филип Дж .; Карпентер, Кеннет (2000). «Новый экземпляр Acrocanthosaurus atokensis (Theropoda, Dinosauria) из нижнемеловой формации Антлерс (нижний мел, апт) Оклахомы, США» . Геодиверситас . 22 (2): 207–246. Архивировано из оригинала 14 ноября 2007 года.
- ^ Перейти обратно: а б с Д'Эмик, Майкл Д.; Мелстром, Киган М.; Эдди, Дрю Р. (2012). «Палеобиология и географический ареал крупнотелого мелового динозавра-теропода Acrocanthosaurus atokensis ». Палеогеография, Палеоклиматология, Палеоэкология . 333–334: 13–23. Бибкод : 2012PPP...333...13D . дои : 10.1016/j.palaeo.2012.03.003 .
- ^ Липка, Томас Р. (1998). «Сродство загадочных теропод фации Арунделской глины (апт), Потомакская формация, Атлантическая прибрежная равнина Мэриленда». В Лукасе, Спенсер Г .; Киркланд, Джеймс И .; Эстеп, JW (ред.). Нижне- и среднемеловые наземные экосистемы . Музея естественной истории и науки Нью-Мексико Бюллетень 14 . стр. 229–234.
- ^ Перейти обратно: а б Каррано, Мэтью Т. (1 мая 2024 г.). «Первая окончательная запись акрокантозавра (Theropoda: Carcharodontosauridae) в нижнем мелу восточной части Северной Америки» . Меловые исследования . 157 : 105814. Бибкод : 2024CrRes.15705814C . дои : 10.1016/j.cretres.2023.105814 . ISSN 0195-6671 .
- ^ Раткевич, Рональд П. (1997). «Останки динозавров южной Аризоны». В Вольберге, Дональд Л.; Стамп, Эдвард; Розенберг, Гэри (ред.). Dinofest International: Материалы симпозиума, проведенного в Университете штата Аризона . Филадельфия: Академия естественных наук. ISBN 978-0-935868-94-4 .
- ^ Раткевич, Рональд П. (1998). «Новый меловой брахиозавридный динозавр, Sonorasaurus thompsoni gen. et sp. nov., из Аризоны». Журнал Академии наук Аризоны и Невады . 31 (1): 71–82.
- ^ Харрис, Джеральд Д. (1998). «Большие тероподы раннего мела в Северной Америке». В Лукасе, Спенсер Г .; Киркланд, Джеймс И .; Эстеп, JW (ред.). Нижне- и среднемеловые наземные экосистемы . Музея естественной истории и науки Нью-Мексико Бюллетень 14 . стр. 225–228.
- ^ Перейти обратно: а б Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 102. ИСБН 978-1-78684-190-2 . OCLC 985402380 .
- ^ Канале, Дж.И.; Апестегия, С.; Галлина, Пенсильвания; Митчелл, Дж.; Смит, Северная Дакота; Каллен, ТМ; Шинья, А.; Халуза, А.; Джанекини, ФА; Маковицкий, П.Дж. (7 июля 2022 г.). «Новый гигантский плотоядный динозавр демонстрирует конвергентные эволюционные тенденции в уменьшении рук теропод» . Современная биология . 32 (14): 3195–3202.e5. Бибкод : 2022CBio...32E3195C . дои : 10.1016/j.cub.2022.05.057 . ПМИД 35803271 .
- ^ Снайвли, Э.; О'Брайен, Х.; Хендерсон, DM; Мэллисон, Х.; Сурринг, Луизиана; Бернс, Мэн; Хольц, ТР; Младший, Рассел, AP; Уитмер, LM; Карри, Пи Джей; Хартман, SA; Коттон, младший (2019). «Меньшая вращательная инерция и большие мышцы ног указывают на более быстрые повороты у тираннозаврид, чем у других крупных теропод» . ПерДж . 7 : е6432. дои : 10.7717/peerj.6432 . ПМК 6387760 . ПМИД 30809441 .
- ^ Кампионе, Николас Э.; Эванс, Дэвид С. (2020). «Точность и точность оценки массы тела нептичьих динозавров» . Биологические обзоры . 95 (6): 1759–1797. дои : 10.1111/brv.12638 . ISSN 1469-185Х . ПМИД 32869488 . S2CID 221404013 .
- ^ Перейти обратно: а б с д и ж Хольц, Томас Р .; Мольнар, Ральф Э .; Карри, Филип Дж. (2004). «Базальная столбняк». В Вейшампеле, Дэвид Б .; Додсон, Питер ; Осмольска, Гальшка (ред.). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. стр. 71–110. ISBN 978-0-520-24209-8 .
- ^ Мольнар, Ральф Э .; Курзанов Сергей М.; Дун Чжимин (1990). «Карнозаврия» В Вейшампеле, Дэвид Б .; Додсон, Питер ; Осмольска, Гальшка (ред.). Динозаврия (Первое изд.). Беркли: Издательство Калифорнийского университета. стр. 100-1 169–209. ISBN 978-0-520-06727-1 .
- ^ Перейти обратно: а б Нэйш, Даррен ; Хатт, Стивен; Мартилл, Дэвид М. (2001). «Ящеротазовые динозавры 2: Тероподы». Динозавры острова Уайт . Лондон: Палеонтологическая ассоциация. стр. 242–309. ISBN 978-0-901702-72-2 .
- ^ Брусатте, Стивен Л.; Бенсон, Роджер Б.Дж.; Чуре, Дэниел Дж.; Сюй, Син; Салливан, Корвин; Хоун, Дэвид ВЕ (2009). «Первый окончательный кархародонтозаврид (Dinosauria: Theropoda) из Азии и запоздалое восхождение тираннозаврид» (PDF) . Naturwissenschaften . 96 (9): 1051–8. Бибкод : 2009NW.....96.1051B . дои : 10.1007/s00114-009-0565-2 . hdl : 20.500.11820/33528c2e-0c9c-4160-8693-984f077ee5d0 . ПМИД 19488730 . S2CID 25532873 .
- ^ Перейти обратно: а б Бенсон, Роджер Б.Дж.; Каррано, Мэтью Т.; Брусатте, Стивен Л. (2009). «Новая клада архаичных крупнотелых хищных динозавров (Theropoda: Allosauroidea), дожившая до позднего мезозоя» (PDF) . Naturwissenschaften . 97 (1): 71–8. Бибкод : 2010NW.....97...71B . дои : 10.1007/s00114-009-0614-x . ПМИД 19826771 . S2CID 22646156 .
- ^ Ромер, Альфред С. (1956). Остеология рептилий . Чикаго: Издательство Чикагского университета. п. 772 стр. ISBN 978-0-89464-985-1 .
- ^ Уокер, Алик Д. (1964). «Триасовые рептилии района Элгина: орнитозухи и происхождение карнозавров» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 248 (744): 53–134. Бибкод : 1964RSPTB.248...53W . дои : 10.1098/rstb.1964.0009 .
- ^ Ромер, Альфред С. (1966). Палеонтология позвоночных (Третье изд.). Чикаго: Издательство Чикагского университета. п. 468 стр . ISBN 978-0-7167-1822-2 .
- ^ Кэрролл, Роберт Л. (1988). Палеонтология и эволюция позвоночных . WH Фриман и компания. ISBN 978-0-7167-1822-2 .
- ^ Ламберт, Дэвид; Группа диаграмм (1983). «Спинозавриды» . Полевое руководство по динозаврам . Нью-Йорк: Книги Эйвона. стр. 84–85 . ISBN 978-0-380-83519-5 .
- ^ Норман, Дэвид Б. (1985). «Карнозавры». Иллюстрированная энциклопедия динозавров: оригинальный и убедительный взгляд на жизнь в царстве динозавров . Нью-Йорк: Crescent Books. стр. 62–67. ISBN 978-0-517-46890-6 .
- ^ Перейти обратно: а б Перенасыщение, Дональд Ф. (1982). Новый словарь динозавров . Секаукус, Нью-Джерси: Citadel Press. стр. 39, 48 . ISBN 978-0-8065-0782-8 .
- ^ Пол, Грегори С. (1988). «Род Акрокантозавр » . Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 314–315 . ISBN 978-0-671-61946-6 .
- ^ Ольшевский, Георгий (1991). Пересмотр параинфракласса Archosauria Cope, 1869 г., исключая Advanced Crocodylia . Сан-Диего: публикации, требующие исследования. п. 196 стр.
- ^ Эдди, Дрю Р.; Кларк, Джулия А. (2011). Фарке, Эндрю (ред.). «Новая информация о краниальной анатомии Acrocanthosaurus atokensis и ее значении для филогении Allosauroidea (Dinosauria: Theropoda)» . ПЛОС ОДИН . 6 (3): e17932. Бибкод : 2011PLoSO...617932E . дои : 10.1371/journal.pone.0017932 . ПМК 3061882 . ПМИД 21445312 .
- ^ Раухут, Оливер В.М. (2011). «Тероподовые динозавры поздней юры Тендагуру (Танзания)» . Специальные статьи по палеонтологии . 86 : 195–239.
- ^ Новас, Фернандо Э. (2013). «Эволюция хищных динозавров в меловой период: данные из Патагонии». Меловые исследования . 45 : 174–215. Бибкод : 2013CrRes..45..174N . дои : 10.1016/j.cretres.2013.04.001 . hdl : 11336/102037 .
- ^ Кау, Андреа (2024). «Единая основа макроэволюции хищных динозавров» . Бюллетень Итальянского палеонтологического общества . 63 (1): 1–19. дои : 10.4435/BSPI.2024.08 . ISSN 0375-7633 .
- ^ Сакамото, Манабу (12 июля 2022 г.). «Оценка силы укуса у вымерших динозавров с использованием филогенетически предсказанных физиологических площадей поперечного сечения приводящих мышц челюсти» . ПерДж . 10 : е13731. дои : 10.7717/peerj.13731 . ISSN 2167-8359 . ПМЦ 9285543 . ПМИД 35846881 .
- ^ Перейти обратно: а б с д и Сентер, Фил; Робинс, Джеймс Х. (2005). «Амплитуда движений передней конечности динозавра-теропода Acrocanthosaurus atokensis и последствия для хищнического поведения» . Журнал зоологии . 266 (3): 307–318. дои : 10.1017/S0952836905006989 .
- ^ Перейти обратно: а б Францоза, Джонатан; Роу, Тимоти. (2005). «Краниальная эндокаст мелового динозавра-теропода Acrocanthosaurus atokensis » . Журнал палеонтологии позвоночных . 25 (4): 859–864. doi : 10.1671/0272-4634(2005)025[0859:CEOTCT]2.0.CO;2 . S2CID 10173542 .
- ^ «Засуха в Техасе обнаружила следы динозавров 113 миллионов лет назад» .
- ^ Перейти обратно: а б Берд, Роланд Т. (1941). «Динозавр заходит в музей». Естественная история . 43 : 254–261.
- ^ Роджерс, Джек В. (2002). «Следы динозавров теропод в формации Глен Роуз нижнего мела (альб), округ Кинни, Техас». Техасский научный журнал . 54 (2): 133–142.
- ^ Хоторн, Дж. Майкл; Бонем, Рена М.; Фарлоу, Джеймс О .; Джонс, Джеймс О. (2002). «Ихнология, стратиграфия и палеосреда места следов динозавров на водосбросе озера Берн, юго-центральный Техас». Техасский научный журнал . 54 (4): 309–324.
- ^ Лэнгстон, Ванн (1974). «Команчские четвероногие, немлекопитающие». Геонауки и человек . 3 : 77–102.
- ^ Перейти обратно: а б Фарлоу, Джеймс О. (2001). « Акрокантозавр и создатель следов команчейских крупных теропод» . В Танке, Даррен ; Карпентер, Кен (ред.). Жизнь мезозойских позвоночных . Блумингтон: Издательство Университета Индианы. стр. 408–427 . ISBN 978-0-253-33907-2 .
- ^ Перейти обратно: а б Локли, Мартин Г. (1991). Отслеживание динозавров: новый взгляд на древний мир . Кембридж: Издательство Кембриджского университета. п. 252 стр. ISBN 978-0-521-39463-5 .
- ^ Томас, Дэвид А.; Фарлоу, Джеймс О. (1997). «Отслеживание нападения динозавра». Научный американец . 266 (6): 48–53. Бибкод : 1997SciAm.277f..74T . doi : 10.1038/scientificamerican1297-74 .
- ^ Молнар, Р.Э., 2001, Палеопатология теропод: обзор литературы: В: Жизнь мезозойских позвоночных, под редакцией Танке, Д.Х., и Карпентера, К., Indiana University Press, стр. 337–363.
- ^ Джейкобс, Луи Л.; Винклер, Дейл А.; Марри, Патрик А. (1991). «О возрасте и корреляции млекопитающих Тринити, ранний мел Техаса, США». Информационные бюллетени по стратиграфии . 24 (1–2): 35–43. дои : 10.1127/номер/24/1991/35 .
- ^ Перейти обратно: а б Бринкман, Дэниел Л.; Чифелли, Ричард Л.; Чаплевский, Николас Дж. (1998). «Первое появление Deinonychus antirhopus (Dinosauria: Theropoda) из формации Антлерс (нижний мел: апт – альб) Оклахомы» . Бюллетень геологической службы Оклахомы . 146 : 1–27.
- ^ Роуз, Питер Дж. (2007). «Новый титанозаврообразный зауропод (Dinosauria: Saurischia) из раннего мела центрального Техаса и его филогенетические взаимоотношения» . Электронная палеонтология . 10 (2): 65с. [опубликовано в Интернете]
- ^ Ведель, Мэтью Дж.; Чифелли, Ричард Л.; Кент Сандерс, Р. (2000). « Sauroposeidon proteles , новый зауропод из раннего мела Оклахомы» (PDF) . Журнал палеонтологии позвоночных . 20 (1): 109–114. doi : 10.1671/0272-4634(2000)020[0109:SPANSF]2.0.CO;2 . S2CID 55987496 .
- ^ Винклер, Дейл А.; Марри, Патрик А.; Джейкобс, Луи Л. (1997). «Новый вид тенонтозавра (Dinosauria: Ornithopoda) из раннего мела Техаса» . Журнал палеонтологии позвоночных . 17 (2): 330–348. Бибкод : 1997JVPal..17..330W . дои : 10.1080/02724634.1997.10010978 . Архивировано из оригинала 27 сентября 2007 года.















- Кархародонтозавриды
- Альбийская жизнь
- Аптская жизнь
- Раннемеловые динозавры Северной Америки.
- Ископаемые тропы
- Клеверная фауна
- Палеонтология в Мэриленде
- Палеонтология в Оклахоме
- Палеонтология в Вайоминге
- Палеонтология в Техасе
- Ископаемые таксоны описаны в 1950 году.
- Таксоны, названные Дж. Уиллисом Стоваллом
- Таксон, названный Ванном Лэнгстоном младшим.
- Высшие хищники
- Символы Оклахомы