Фагоциты

Фагоциты — это клетки , которые защищают организм, поглощая вредные инородные частицы, бактерии , а также мертвые или умирающие клетки. Их название происходит от греческого phagein , «есть» или «пожирать», и «-cyte», суффикса в биологии, обозначающего «клетку», от греческого kutos , «полый сосуд». [ 1 ] Они необходимы для борьбы с инфекциями и для последующего иммунитета . [ 2 ] Фагоциты играют важную роль во всем животном мире. [ 3 ] и высоко развиты у позвоночных. [ 4 ] В одном литре человеческой крови содержится около шести миллиардов фагоцитов. [ 5 ] Их открыл в 1882 году Илья Ильич Мечников во время изучения морских звезд личинок . [ 6 ] Мечников был удостоен Нобелевской премии по физиологии и медицине 1908 года. За свое открытие [ 7 ] Фагоциты встречаются у многих видов; некоторые амебы ведут себя как фагоциты-макрофаги, что позволяет предположить, что фагоциты появились на ранних стадиях эволюции жизни. [ 8 ]
Фагоциты человека и других животных называются «профессиональными» или «непрофессиональными» в зависимости от того, насколько они эффективны при фагоцитозе . [ 9 ] К профессиональным фагоцитам относятся многие типы лейкоцитов (такие как нейтрофилы , моноциты , макрофаги , тучные клетки и дендритные клетки ). [ 10 ] Основное различие между профессиональными и непрофессиональными фагоцитами заключается в том, что профессиональные фагоциты имеют на своей поверхности молекулы, называемые рецепторами , которые могут обнаруживать вредные объекты, такие как бактерии, которые обычно не встречаются в организме. Непрофессиональные фагоциты не имеют эффективных фагоцитарных рецепторов, например, для опсонинов . [ 11 ] Фагоциты играют решающую роль в борьбе с инфекциями, а также в поддержании здоровья тканей путем удаления мертвых и умирающих клеток, срок службы которых истек. [ 12 ]
Во время инфекции химические сигналы привлекают фагоциты к местам проникновения возбудителя в организм. Эти химические вещества могут поступать из бактерий или других уже присутствующих фагоцитов. Фагоциты перемещаются методом, называемым хемотаксисом . Когда фагоциты вступают в контакт с бактериями, рецепторы на поверхности фагоцита связываются с ними. Это связывание приведет к поглощению бактерий фагоцитом. [ 13 ] Некоторые фагоциты убивают попавший в организм патоген с помощью окислителей и оксида азота . [ 14 ] После фагоцитоза макрофаги и дендритные клетки также могут участвовать в презентации антигена — процессе, при котором фагоцит перемещает части проглоченного материала обратно на свою поверхность. Этот материал затем отображается другим клеткам иммунной системы. организма Некоторые фагоциты затем попадают в лимфатические узлы и отображают материал лейкоцитам, называемым лимфоцитами . Этот процесс важен для формирования иммунитета. [ 15 ] и многие патогены разработали методы уклонения от атак фагоцитов. [ 2 ]
История
[ редактировать ]
Русский зоолог Илья Ильич Мечников (1845–1916) впервые признал, что специализированные клетки участвуют в защите от микробных инфекций. [ 16 ] В 1882 году он изучал подвижные (свободно движущиеся) клетки личинок морских звезд , полагая, что они важны для иммунной защиты животных. Чтобы проверить свою идею, он вставил небольшие шипы мандаринового в личинки дерева. Через несколько часов он заметил, что шипы окружили подвижные клетки. [ 16 ] Мечников отправился в Вену и поделился своими идеями с Карлом Фридрихом Клаусом , который предложил название «фагоцит» (от греческих слов phagein , что означает «есть или пожирать», и kutos , что означает «полый сосуд»). [ 1 ] ) для клеток, которые наблюдал Мечников. [ 17 ]
Год спустя Мечников изучал пресноводного ракообразного под названием дафния , крошечное прозрачное животное, которое можно рассматривать прямо под микроскопом. Он обнаружил, что споры грибков, атаковавшие животное, уничтожались фагоцитами. Он распространил свои наблюдения на лейкоциты млекопитающих и обнаружил, что бактерия Bacillus anthracis может быть поглощена и уничтожена фагоцитами - процесс, который он назвал фагоцитозом . [ 18 ] Мечников предположил, что фагоциты являются основной защитой от вторгающихся организмов. [ 16 ]
В 1903 году Альмрот Райт обнаружил, что фагоцитоз усиливается специфическими антителами , которые он назвал опсонинами , от греческого opson — «приправа или приправа». [ 19 ] Мечников был удостоен (совместно с Паулем Эрлихом ) Нобелевской премии по физиологии и медицине 1908 года за работы по фагоцитам и фагоцитозу. [ 7 ]
Хотя важность этих открытий постепенно получила признание в начале двадцатого века, сложные взаимоотношения между фагоцитами и всеми другими компонентами иммунной системы не были известны до 1980-х годов. [ 20 ]
Фагоцитоз
[ редактировать ]![Мультфильм: 1. Частица изображена овалом, а поверхность фагоцита - прямой линией. Различные более мелкие фигуры находятся на линии и овале. 2. Меньшие частицы на каждой поверхности соединяются. 3. Линия теперь вогнутая и частично охватывает овал.[21]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/ba/Phagocytosis_in_three_steps.png/220px-Phagocytosis_in_three_steps.png)
Фагоцитоз — это процесс поглощения таких частиц, как бактерии, инвазивные грибы , паразиты, мертвые клетки-хозяева , а также клеточный и инородный мусор. клеткой [ 22 ] Он включает в себя цепочку молекулярных процессов. [ 23 ] [ 24 ] Фагоцитоз происходит после того, как инородное тело, например, бактериальная клетка, связалось с молекулами, называемыми «рецепторами», которые находятся на поверхности фагоцита. Затем фагоцит растягивается вокруг бактерии и поглощает ее. Фагоцитоз бактерий нейтрофилами человека занимает в среднем девять минут. [ 25 ] Попав внутрь этого фагоцита, бактерия попадает в отсек, называемый фагосомой . В течение одной минуты фагосома сливается либо с лизосомой , либо с гранулой, образуя фаголизосому . Затем бактерия подвергается множеству механизмов уничтожения. [ 26 ] и через несколько минут умирает. [ 25 ] Дендритные клетки и макрофаги действуют не так быстро, и фагоцитоз в этих клетках может длиться многие часы. Макрофаги едят медленно и неопрятно; они поглощают огромное количество материала и часто выделяют непереваренную часть обратно в ткани. Этот мусор служит сигналом для привлечения большего количества фагоцитов из крови. [ 27 ] Фагоциты обладают ненасытным аппетитом; ученые даже кормили макрофаги железными опилками , а затем использовали небольшой магнит, чтобы отделить их от других клеток. [ 28 ]

Фагоцит имеет на своей поверхности множество типов рецепторов, которые используются для связывания материала. [ 2 ] К ним относятся опсониновые рецепторы, рецепторы-мусорщики и Toll-подобные рецепторы . Рецепторы опсонина усиливают фагоцитоз бактерий, покрытых к иммуноглобулину G (IgG) антителами или комплементом . «Комплемент» — это название сложной серии белковых молекул, обнаруженных в крови, которые разрушают клетки или помечают их для разрушения. [ 29 ] Рецепторы-мусорщики связываются с большим количеством молекул на поверхности бактериальных клеток, а Toll-подобные рецепторы, названные так из-за их сходства с хорошо изученными рецепторами плодовых мух, которые кодируются геном Toll, связываются с более специфическими молекулами, включая чужеродная ДНК и РНК. [ 30 ] Связывание с Toll-подобными рецепторами увеличивает фагоцитоз и заставляет фагоцит выделять группу гормонов, вызывающих воспаление . [ 2 ]
Способы убийства
[ редактировать ]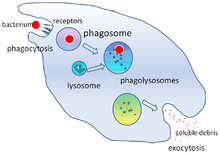
Уничтожение микробов является важнейшей функцией фагоцитов, которая осуществляется либо внутри фагоцита ( внутриклеточное уничтожение), либо вне фагоцита ( внеклеточное уничтожение). [ 31 ]
Кислородозависимый внутриклеточный
[ редактировать ]Когда фагоцит поглощает бактерии (или любой материал), его потребление кислорода увеличивается. Увеличение потребления кислорода, называемое респираторным взрывом , приводит к образованию реактивных кислородсодержащих молекул, обладающих противомикробным действием. [ 32 ] Соединения кислорода токсичны как для захватчика, так и для самой клетки, поэтому они хранятся в отсеках внутри клетки. Этот метод уничтожения вторгшихся микробов с использованием активных кислородсодержащих молекул называется кислородзависимым внутриклеточным уничтожением, которое существует двух типов. [ 14 ]
Первый тип — кислородзависимое образование супероксида . [ 2 ] Это богатое кислородом вещество, убивающее бактерии. [ 33 ] Супероксид преобразуется в пероксид водорода и синглетный кислород с помощью фермента, называемого супероксиддисмутазой . Супероксиды также реагируют с перекисью водорода с образованием гидроксильных радикалов , которые помогают убить вторгшийся микроб. [ 2 ]
Второй тип предполагает использование фермента миелопероксидазы из гранул нейтрофилов. [ 34 ] Когда гранулы сливаются с фагосомой, в фаголизосому высвобождается миелопероксидаза, и этот фермент использует перекись водорода и хлор для создания гипохлорита — вещества, используемого в домашнем отбеливателе . Гипохлорит чрезвычайно токсичен для бактерий. [ 2 ] Миелопероксидаза содержит гемовый пигмент, который отвечает за зеленый цвет выделений, богатых нейтрофилами, таких как гной и инфицированная мокрота . [ 35 ]
Кислороднезависимый внутриклеточный
[ редактировать ]
Фагоциты также могут убивать микробы кислороднезависимыми методами, но они не так эффективны, как кислородзависимые. Существует четыре основных типа. бактерии Первый использует электрически заряженные белки, которые повреждают мембрану . Второй тип использует лизоцимы; эти ферменты разрушают клеточную стенку бактерий . Третий тип использует лактоферрины , которые присутствуют в гранулах нейтрофилов и удаляют необходимое железо из бактерий. [ 36 ] Четвертый тип использует протеазы и гидролитические ферменты ; эти ферменты используются для переваривания белков уничтоженных бактерий. [ 37 ]
внеклеточный
[ редактировать ]Интерферон-гамма , который когда-то назывался фактором активации макрофагов, стимулирует макрофаги вырабатывать оксид азота . Источником интерферона-гамма может быть CD4. + Т-клетки , CD8 + Т-клетки , естественные клетки-киллеры , В-клетки , естественные Т-клетки-киллеры , моноциты, другие макрофаги или дендритные клетки. [ 38 ] Затем оксид азота высвобождается из макрофага и из-за своей токсичности убивает микробы вблизи макрофага. [ 2 ] Активированные макрофаги продуцируют и секретируют фактор некроза опухоли . Этот цитокин — класс сигнальных молекул [ 39 ] — убивает раковые клетки и клетки, зараженные вирусами, и помогает активировать другие клетки иммунной системы. [ 40 ]
При некоторых заболеваниях, например, редкой хронической гранулематозной болезни , эффективность фагоцитов снижается, и проблемой являются рецидивирующие бактериальные инфекции. [ 41 ] При этом заболевании наблюдается аномалия, затрагивающая различные элементы кислородзависимого уничтожения. Другие редкие врожденные аномалии, такие как синдром Чедиака-Хигаси , также связаны с дефектным уничтожением проглоченных микробов. [ 42 ]
Вирусы
[ редактировать ]Вирусы могут размножаться только внутри клеток и проникать в них, используя многие рецепторы, участвующие в иммунитете. Попав внутрь клетки, вирусы используют ее биологические механизмы в своих целях, заставляя клетку создавать сотни идентичных копий самих себя. Хотя фагоциты и другие компоненты врожденной иммунной системы могут в ограниченной степени контролировать вирусы, как только вирус оказывается внутри клетки, адаптивные иммунные реакции, особенно лимфоциты, становятся более важными для защиты. [ 43 ] В местах вирусных инфекций лимфоциты часто значительно превосходят по численности все остальные клетки иммунной системы; это часто встречается при вирусном менингите . [ 44 ] Зараженные вирусом клетки, убитые лимфоцитами, выводятся из организма фагоцитами. [ 45 ]
Роль в апоптозе
[ редактировать ]У животного клетки постоянно умирают. Баланс между делением клеток и гибелью клеток сохраняет количество клеток относительно постоянным у взрослых. [ 12 ] Клетка может умереть двумя способами: некрозом или апоптозом. В отличие от некроза, который часто возникает в результате заболевания или травмы, апоптоз — или запрограммированная гибель клеток — является нормальной здоровой функцией клеток. Организму ежедневно приходится избавляться от миллионов мертвых или умирающих клеток, и фагоциты играют в этом процессе решающую роль. [ 46 ]
Умирающие клетки, которые проходят заключительные стадии апоптоза. [ 47 ] отображают молекулы, такие как фосфатидилсерин , на поверхности клеток для привлечения фагоцитов. [ 48 ] Фосфатидилсерин обычно обнаруживается на цитозольной поверхности плазматической мембраны, но во время апоптоза перераспределяется на внеклеточную поверхность белком, известным как скрамблаза . [ 49 ] [ 50 ] Эти молекулы маркируют клетку для фагоцитоза клетками, обладающими соответствующими рецепторами, такими как макрофаги. [ 51 ] Удаление отмирающих клеток фагоцитами происходит упорядоченно, не вызывая воспалительной реакции , и является важной функцией фагоцитов. [ 52 ]
Взаимодействие с другими клетками
[ редактировать ]Фагоциты обычно не связаны с каким-либо конкретным органом , а перемещаются по организму, взаимодействуя с другими фагоцитирующими и нефагоцитарными клетками иммунной системы. Они могут взаимодействовать с другими клетками, производя химические вещества, называемые цитокинами , которые привлекают другие фагоциты к месту инфекции или стимулируют спящие лимфоциты . [ 53 ] Фагоциты являются частью врожденной иммунной системы , с которой рождаются животные, включая человека. Врожденный иммунитет очень эффективен, но неспецифичен, поскольку не делает различий между различными типами захватчиков. С другой стороны, адаптивная иммунная система челюстных позвоночных — основа приобретенного иммунитета — высокоспециализирована и может защитить практически от любого типа захватчика. [ 54 ] Адаптивная иммунная система зависит не от фагоцитов, а от лимфоцитов, которые производят защитные белки, называемые антителами , которые метят захватчиков для уничтожения и предотвращают заражение клеток вирусами. [ 55 ] Фагоциты, в частности дендритные клетки и макрофаги, стимулируют лимфоциты к выработке антител посредством важного процесса, называемого презентацией антигена . [ 56 ]
Презентация антигена
[ редактировать ]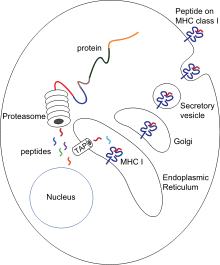
Презентация антигена — это процесс, при котором некоторые фагоциты перемещают части поглощенных материалов обратно на поверхность своих клеток и «презентуют» их другим клеткам иммунной системы. [ 57 ] Существуют две «профессиональные» антигенпредставляющие клетки: макрофаги и дендритные клетки. [ 58 ] После поглощения чужеродные белки ( антигены ) расщепляются на пептиды внутри дендритных клеток и макрофагов. клетки Эти пептиды затем связываются с гликопротеинами главного комплекса гистосовместимости (MHC), которые переносят пептиды обратно на поверхность фагоцита, где они могут быть «презентированы» лимфоцитам. [ 15 ] организма Зрелые макрофаги не уходят далеко от места заражения, но дендритные клетки могут достигать лимфатических узлов , где находятся миллионы лимфоцитов. [ 59 ] Это повышает иммунитет, поскольку лимфоциты реагируют на антигены, представленные дендритными клетками, так же, как и в месте первоначальной инфекции. [ 60 ] Но дендритные клетки также могут разрушать или усмирять лимфоциты, если они распознают компоненты тела хозяина; это необходимо для предотвращения аутоиммунных реакций. Этот процесс называется толерантностью. [ 61 ]
Иммунологическая толерантность
[ редактировать ]Дендритные клетки также способствуют иммунологической толерантности, [ 62 ] который останавливает тело от нападения на себя. Первый тип толерантности — центральная толерантность , возникающая в тимусе. Т-клетки, которые слишком сильно связываются (через свои Т-клеточные рецепторы) с аутоантигеном (представленным дендритными клетками на молекулах MHC), погибают. Второй тип иммунологической толерантности — периферическая толерантность . Некоторые аутореактивные Т-клетки покидают тимус по ряду причин, в основном из-за отсутствия экспрессии некоторых аутоантигенов в тимусе. Другой тип Т-клеток; Т-регуляторные клетки могут подавлять аутореактивные Т-клетки на периферии. [ 63 ] Когда иммунологическая толерантность снижается, могут возникнуть аутоиммунные заболевания . [ 64 ]
Профессиональные фагоциты
[ редактировать ]
Фагоциты человека и других челюстных позвоночных делятся на «профессиональные» и «непрофессиональные» группы в зависимости от эффективности их участия в фагоцитозе. [ 9 ] Профессиональные фагоциты представляют собой миелоидные клетки , в состав которых входят моноциты , макрофаги , нейтрофилы , тканевые дендритные клетки и тучные клетки . [ 10 ] В одном литре человеческой крови содержится около шести миллиардов фагоцитов. [ 5 ]
Активация
[ редактировать ]Все фагоциты, и особенно макрофаги, существуют в степени готовности. Макрофаги обычно относительно дремлют в тканях и медленно размножаются. В этом полупокоящемся состоянии они удаляют мертвые клетки-хозяева и другой неинфекционный мусор и редко принимают участие в презентации антигена. Но во время инфекции они получают химические сигналы — обычно гамма-интерферон — которые увеличивают выработку ими молекул MHC II и подготавливают их к представлению антигенов. В этом состоянии макрофаги являются хорошими антигенпрезентаторами и киллерами. Если они получают сигнал непосредственно от захватчика, они становятся «гиперактивными», перестают размножаться и концентрируются на убийстве. Их размер и скорость фагоцитоза увеличиваются — некоторые становятся достаточно большими, чтобы поглотить вторгшихся простейших . [ 65 ]
В крови нейтрофилы неактивны, но движутся с высокой скоростью. Когда они получают сигналы от макрофагов в местах воспаления, они замедляются и покидают кровь. В тканях они активируются цитокинами и прибывают на место боя готовые убивать. [ 66 ]
Миграция
[ редактировать ]
При возникновении инфекции подается химический сигнал «SOS», привлекающий к месту заражения фагоциты. [ 67 ] Эти химические сигналы могут включать белки вторгшихся бактерий, пептиды системы свертывания крови , продукты комплемента и цитокины, выделяемые макрофагами, расположенными в ткани вблизи места инфекции. [ 2 ] Другая группа химических аттрактантов — это цитокины , которые рекрутируют нейтрофилы и моноциты из крови. [ 13 ]
Чтобы добраться до места заражения, фагоциты покидают кровоток и попадают в пораженные ткани. Сигналы инфекции заставляют эндотелиальные клетки, выстилающие кровеносные сосуды, вырабатывать белок под названием селектин , к которому нейтрофилы прилипают, проходя мимо. Другие сигналы, называемые вазодилататорами, ослабляют соединения, соединяющие эндотелиальные клетки, позволяя фагоцитам проходить через стенку. Хемотаксис — это процесс, при котором фагоциты следуют по «запаху» цитокинов к инфицированному участку. [ 2 ] Нейтрофилы перемещаются через органы, выстланные эпителиальными клетками, к местам инфекции, и хотя это важный компонент борьбы с инфекцией, сама миграция может привести к симптомам, похожим на болезнь. [ 68 ] Во время инфекции миллионы нейтрофилов рекрутируются из крови, но через несколько дней они погибают. [ 69 ]
Моноциты
[ редактировать ]
Моноциты развиваются в костном мозге и достигают зрелости в крови. Зрелые моноциты имеют крупные гладкие дольчатые ядра и обильную цитоплазму , содержащую гранулы. Моноциты поглощают чужеродные или опасные вещества и представляют антигены другим клеткам иммунной системы. Моноциты образуют две группы: циркулирующую группу и маргинальную группу, которые остаются в других тканях (около 70% находятся в маргинальной группе). Большинство моноцитов покидают кровоток через 20–40 часов и попадают в ткани и органы, превращаясь при этом в макрофаги. [ 70 ] или дендритные клетки в зависимости от сигналов, которые они получают. [ 71 ] В одном литре человеческой крови содержится около 500 миллионов моноцитов. [ 5 ]
Макрофаги
[ редактировать ]Зрелые макрофаги не путешествуют далеко, а стоят на страже тех участков тела, которые подвергаются воздействию внешнего мира. Там они действуют как сборщики мусора, антигенпредставляющие клетки или свирепые убийцы, в зависимости от получаемых сигналов. [ 72 ] Они происходят из моноцитов, гранулоцитарных стволовых клеток или клеточного деления ранее существовавших макрофагов. [ 73 ] Макрофаги человека имеют около 21 микрометра . диаметр [ 74 ]

Этот тип фагоцитов не имеет гранул, но содержит множество лизосом . Макрофаги обнаруживаются по всему телу почти во всех тканях и органах (например, микроглиальные клетки в мозге и альвеолярные макрофаги в легких ), где они молча подстерегают. Местоположение макрофага может определять его размер и внешний вид. Макрофаги вызывают воспаление посредством производства интерлейкина-1 , интерлейкина-6 и TNF-альфа . [ 75 ] Макрофаги обычно обнаруживаются только в тканях и редко обнаруживаются в кровообращении. По оценкам, продолжительность жизни тканевых макрофагов составляет от четырех до пятнадцати дней. [ 76 ]
Макрофаги могут быть активированы для выполнения функций, которые покоящийся моноцит не может выполнить. [ 75 ] Т-хелперные клетки (также известные как эффекторные Т-клетки или Т - клетки), подгруппа лимфоцитов, ответственны за активацию макрофагов. Клетки Th и 1 активируют макрофаги путем передачи сигнала IFN-гамма демонстрации белка- лиганда CD40 . [ 77 ] Другие сигналы включают TNF-альфа и липополисахариды бактерий. [ 75 ] Клетки Th 1 могут рекрутировать другие фагоциты в место инфекции несколькими способами. Они секретируют цитокины, которые действуют на костный мозг , стимулируя выработку моноцитов и нейтрофилов, а также секретируют некоторые цитокины , которые отвечают за миграцию моноцитов и нейтрофилов из кровотока. [ 78 ] Th 1 происходят в результате Клетки дифференцировки CD4 + Т-клетки, как только они ответили на антиген во вторичных лимфоидных тканях . [ 75 ] Активированные макрофаги играют важную роль в разрушении опухоли , производя TNF-альфа, IFN-гамма, оксид азота, активные соединения кислорода, катионные белки и гидролитические ферменты. [ 75 ]
Нейтрофилы
[ редактировать ]
Нейтрофилы обычно обнаруживаются в кровотоке и представляют собой наиболее распространенный тип фагоцитов, составляя от 50% до 60% от общего количества циркулирующих лейкоцитов. [ 79 ] В одном литре человеческой крови содержится около пяти миллиардов нейтрофилов. [ 5 ] которые имеют диаметр около 10 микрометров [ 80 ] и живут всего около пяти дней. [ 40 ] После получения соответствующих сигналов им требуется около тридцати минут, чтобы выйти из крови и добраться до места заражения. [ 81 ] Они свирепые пожиратели и быстро поглощают захватчиков, покрытых антителами и комплементом , а также поврежденные клетки или клеточный мусор. Нейтрофилы не возвращаются в кровь; они превращаются в гнойные клетки и умирают. [ 81 ] Зрелые нейтрофилы меньше моноцитов и имеют сегментированное ядро , состоящее из нескольких отделов; каждый участок соединен нитями хроматина — нейтрофилы могут иметь 2–5 сегментов. Нейтрофилы обычно не покидают костный мозг до зрелости, но во время инфекции предшественники нейтрофилов, называемые метамиелоцитами , миелоцитами и промиелоцитами . высвобождаются [ 82 ]
Внутриклеточные гранулы нейтрофилов человека давно известны своими белковоразрушающими и бактерицидными свойствами. [ 83 ] Нейтрофилы могут секретировать продукты, которые стимулируют моноциты и макрофаги. Секреция нейтрофилов усиливает фагоцитоз и образование активных соединений кислорода, участвующих во внутриклеточном уничтожении. [ 84 ] Секреции первичных гранул нейтрофилов стимулируют фагоцитоз бактерий, покрытых IgG -антителами. [ 85 ] При встрече с бактериями, грибами или активированными тромбоцитами они образуют паутинные структуры хроматина, известные как внеклеточные ловушки нейтрофилов (NET). Состоящие в основном из ДНК, NETs вызывают смерть в результате процесса, называемого нетозом: после того, как патогены попадают в ловушку NETs, они погибают за счет окислительных и неокислительных механизмов. [ 86 ]
Дендритные клетки
[ редактировать ]
Дендритные клетки – это специализированные антигенпрезентирующие клетки, имеющие длинные отростки, называемые дендритами. [ 87 ] которые помогают поглотить микробы и других захватчиков. [ 88 ] [ 89 ] Дендритные клетки присутствуют в тканях, контактирующих с внешней средой, главным образом в коже, внутренней оболочке носа, легких, желудке и кишечнике. [ 90 ] После активации они созревают и мигрируют в лимфоидные ткани, где взаимодействуют с Т- и В-клетками, инициируя и управляя адаптивным иммунным ответом. [ 91 ] Зрелые дендритные клетки активируют Т-хелперы и цитотоксические Т-клетки . [ 92 ] Активированные Т-хелперы взаимодействуют с макрофагами и В-клетками, активируя их по очереди. Кроме того, дендритные клетки могут влиять на тип вырабатываемого иммунного ответа; когда они попадают в лимфоидные области, где содержатся Т-клетки, они могут активировать Т-клетки, которые затем дифференцируются в цитотоксические Т-клетки или Т-хелперы. [ 88 ]
Тучные клетки
[ редактировать ]Тучные клетки имеют Toll-подобные рецепторы и взаимодействуют с дендритными клетками, В-клетками и Т-клетками, помогая опосредовать адаптивные иммунные функции. [ 93 ] Тучные клетки экспрессируют молекулы MHC класса II и могут участвовать в презентации антигена; однако роль тучных клеток в презентации антигена не очень хорошо изучена. [ 94 ] Тучные клетки могут поглощать и убивать грамотрицательные бактерии (например, сальмонеллу ), а также перерабатывать их антигены. [ 95 ] Они специализируются на переработке фимбриальных белков на поверхности бактерий, которые участвуют в адгезии к тканям. [ 96 ] [ 97 ] В дополнение к этим функциям тучные клетки вырабатывают цитокины, вызывающие воспалительную реакцию. [ 98 ] Это жизненно важная часть уничтожения микробов, поскольку цитокины привлекают больше фагоцитов к месту инфекции. [ 95 ] [ 99 ]
| Основное расположение | Разнообразие фенотипов |
|---|---|
| Кровь | нейтрофилы, моноциты |
| Костный мозг | макрофаги, моноциты, синусоидальные клетки , выстилающие клетки |
| Костная ткань | остеокласты |
| Кишечные и кишечные пейеровые бляшки | макрофаги |
| Соединительная ткань | гистиоциты , макрофаги, моноциты, дендритные клетки |
| Печень | Купферовы клетки , моноциты |
| Легкое | самовоспроизводящиеся макрофаги, моноциты, тучные клетки, дендритные клетки |
| Лимфоидная ткань | свободные и фиксированные макрофаги и моноциты, дендритные клетки |
| Нервная ткань | клетки микроглии ( CD4 + ) |
| Селезенка | свободные и фиксированные макрофаги, моноциты, синусоидальные клетки |
| Тимус | свободные и фиксированные макрофаги и моноциты |
| Кожа | резидентные клетки Лангерганса , другие дендритные клетки, обычные макрофаги, тучные клетки |
Непрофессиональные фагоциты
[ редактировать ]Умирающие клетки и чужеродные организмы потребляются клетками, отличными от «профессиональных» фагоцитов. [ 101 ] Эти клетки включают эпителиальные клетки , эндотелиальные клетки , фибробласты и мезенхимальные клетки. Их называют непрофессиональными фагоцитами, чтобы подчеркнуть, что, в отличие от профессиональных фагоцитов, фагоцитоз не является их основной функцией. [ 102 ] Например, фибробласты, которые могут фагоцитировать коллаген в процессе восстановления рубцов, также предпримут попытки поглотить инородные частицы. [ 103 ]
Непрофессиональные фагоциты более ограничены, чем профессиональные фагоциты, в типе частиц, которые они могут поглощать. Это связано с отсутствием у них эффективных фагоцитарных рецепторов, в частности опсонинов , которые представляют собой антитела и комплемент, присоединяемые к захватчикам иммунной системой. [ 11 ] Кроме того, большинство непрофессиональных фагоцитов не производят активные кислородсодержащие молекулы в ответ на фагоцитоз. [ 104 ]
| Основное расположение | Разнообразие фенотипов |
|---|---|
| Кровь, лимфа и лимфатические узлы | Лимфоциты |
| Кровь, лимфа и лимфатические узлы | NK- и LGL-клетки (крупные гранулярные лимфоциты) |
| Кровь | Эозинофилы и базофилы [ 105 ] |
| Кожа | Эпителиальные клетки |
| Печень | Гепатоциты [ 106 ] |
| Кровеносные сосуды | Эндотелиальные клетки |
| Соединительная ткань | Фибробласты |
Уклонение и устойчивость патогена
[ редактировать ]
Патогену удается заразить организм только в том случае, если он может преодолеть его защиту. Патогенные бактерии и простейшие разработали множество методов сопротивления атакам фагоцитов, и многие из них действительно выживают и размножаются внутри фагоцитирующих клеток. [ 107 ] [ 108 ]
Избегание контакта
[ редактировать ]Существует несколько способов, которыми бактерии избегают контакта с фагоцитами. Во-первых, они могут расти в местах, куда фагоциты не способны добраться (например, на поверхности неповрежденной кожи). Во-вторых, бактерии могут подавлять воспалительную реакцию ; без этого ответа на инфекцию фагоциты не могут реагировать адекватно. В-третьих, некоторые виды бактерий могут подавлять способность фагоцитов перемещаться к месту инфекции, нарушая хемотаксис. [ 107 ] В-четвертых, некоторые бактерии могут избегать контакта с фагоцитами, обманывая иммунную систему, заставляя ее «думать», что бактерии — это «собственные». Treponema pallidum — бактерия, вызывающая сифилис , — прячется от фагоцитов, покрывая свою поверхность фибронектином . [ 109 ] который естественным образом вырабатывается организмом и играет решающую роль в заживлении ран . [ 110 ]
Как избежать поглощения
[ редактировать ]Бактерии часто производят капсулы из белков или сахаров, которые покрывают их клетки и препятствуют фагоцитозу. [ 107 ] Некоторыми примерами являются капсула K5 и антиген O75 O, обнаруженный на поверхности Escherichia coli . [ 111 ] и экзополисахаридные капсулы Staphylococcus epidermidis . [ 112 ] Streptococcus pneumoniae образует несколько типов капсул, которые обеспечивают разные уровни защиты. [ 113 ] и стрептококки группы А продуцируют белки, такие как белок М и фимбриальные белки, чтобы блокировать поглощение. Некоторые белки препятствуют приему пищи, связанному с опсонином; Золотистый стафилококк вырабатывает белок А, блокирующий рецепторы антител, что снижает эффективность опсонинов. [ 114 ] Энтеропатогенные виды рода Yersinia связываются с помощью фактора вирулентности YopH с рецепторами фагоцитов, откуда влияют на способность клеток осуществлять фагоцитоз. [ 115 ]
Выживание внутри фагоцита
[ редактировать ]
Бактерии разработали способы выживания внутри фагоцитов, где они продолжают уклоняться от иммунной системы. [ 116 ] Чтобы безопасно проникнуть внутрь фагоцита, они экспрессируют белки, называемые инвазинами . Находясь внутри клетки, они остаются в цитоплазме и избегают токсичных химических веществ, содержащихся в фаголизосомах. [ 117 ] Некоторые бактерии предотвращают слияние фагосомы и лизосомы с образованием фаголизосомы. [ 107 ] Другие патогены, такие как Leishmania сильно модифицированную вакуоль , которая помогает им персистировать и размножаться. , создают внутри фагоцита [ 118 ] Некоторые бактерии способны жить внутри фаголизосомы. золотистый стафилококк Например, вырабатывает ферменты каталазу и супероксиддисмутазу , которые расщепляют химические вещества, такие как перекись водорода, вырабатываемые фагоцитами для уничтожения бактерий. [ 119 ] Бактерии могут покинуть фагосому до образования фаголизосомы: Listeria monocytogenes может проделать отверстие в стенке фагосомы с помощью ферментов, называемых листериолизин О и фосфолипаза С. [ 120 ] M.tuberculosis инфицирует нейтрофилы , которые, в свою очередь, поглощаются макрофагами и тем самым заражают и последние. [ 121 ] M. leprae инфицирует макрофаги , шванновские клетки и нейтрофилы . [ 121 ]
Убийство
[ редактировать ]Бактерии разработали несколько способов уничтожения фагоцитов. [ 114 ] К ним относятся цитолизины , образующие поры в клеточных мембранах фагоцитов, стрептолизины и лейкоцидины , вызывающие разрыв гранул нейтрофилов и выделение токсичных веществ. [ 122 ] [ 123 ] и экзотоксины , которые уменьшают запас АТФ в фагоцитах , необходимый для фагоцитоза. После проглатывания бактерия может убить фагоцит, высвобождая токсины, которые проходят через мембрану фагосомы или фаголизосомы и поражают другие части клетки. [ 107 ]
Нарушение клеточной сигнализации
[ редактировать ]
Некоторые стратегии выживания часто включают в себя разрушение цитокинов и других методов передачи сигналов клетками, чтобы предотвратить реакцию фагоцита на инвазию. [ 124 ] Простейшие паразиты Toxoplasma gondii , Trypanosoma cruzi и Leishmania заражают макрофаги, и каждый из них имеет уникальный способ их приручения. [ 124 ] Некоторые виды Leishmania изменяют передачу сигналов инфицированных макрофагов, подавляют выработку цитокинов и микробицидных молекул — оксида азота и активных форм кислорода — и нарушают презентацию антигена. [ 125 ]
Повреждение хозяина фагоцитами
[ редактировать ]Макрофаги и нейтрофилы, в частности, играют центральную роль в воспалительном процессе, высвобождая белки и низкомолекулярные медиаторы воспаления, которые контролируют инфекцию, но могут повредить ткани хозяина. В общем, фагоциты стремятся уничтожить патогены, поглощая их и подвергая воздействию ряда токсичных химикатов внутри фаголизосомы . Если фагоциту не удается поглотить свою цель, эти токсичные агенты могут высвободиться в окружающую среду (действие, называемое «разочарованным фагоцитозом»). Поскольку эти агенты также токсичны для клеток-хозяев, они могут нанести серьезный ущерб здоровым клеткам и тканям. [ 126 ]
Когда нейтрофилы высвобождают содержимое своих гранул в почках , содержимое гранул (активные кислородные соединения и протеазы) разрушает внеклеточный матрикс клеток-хозяев и может вызвать повреждение клубочковых клеток, влияя на их способность фильтровать кровь и вызывая изменения формы. Кроме того, фосфолипазы продукты (например, лейкотриены ) усиливают повреждение. Это высвобождение веществ способствует хемотаксису большего количества нейтрофилов к месту инфекции, и клетки клубочков могут быть дополнительно повреждены молекулами адгезии во время миграции нейтрофилов. Повреждение клубочковых клеток может привести к почечной недостаточности . [ 127 ]
Нейтрофилы также играют ключевую роль в развитии большинства форм острого повреждения легких . [ 128 ] Здесь активированные нейтрофилы выделяют содержимое своих токсичных гранул в легочную среду. [ 129 ] Эксперименты показали, что снижение количества нейтрофилов уменьшает последствия острого повреждения легких. [ 130 ] но лечение путем ингибирования нейтрофилов клинически нереалистично, поскольку оно сделает хозяина уязвимым для инфекции. [ 129 ] В печени повреждение нейтрофилов может способствовать дисфункции и повреждению в ответ на выброс эндотоксинов , вырабатываемых бактериями, сепсис , травму, алкогольный гепатит , ишемию и гиповолемический шок , возникающий в результате острого кровоизлияния . [ 131 ]
Химические вещества, выделяемые макрофагами, также могут повредить ткани хозяина. TNF-α — это важное химическое вещество, которое выделяется макрофагами и вызывает свертывание крови в мелких сосудах, предотвращая распространение инфекции. [ 132 ] Если бактериальная инфекция распространяется в кровь, TNF-α высвобождается в жизненно важные органы, что может вызвать расширение сосудов и уменьшение объема плазмы ; за этим, в свою очередь, может последовать септический шок . Во время септического шока выброс TNF-α вызывает закупорку мелких сосудов, снабжающих кровью жизненно важные органы, и органы могут выйти из строя. Септический шок может привести к смерти. [ 13 ]
Эволюционное происхождение
[ редактировать ]
Фагоцитоз является обычным явлением и, вероятно, появился на ранних стадиях эволюции . [ 133 ] сначала развиваются у одноклеточных эукариот. [ 134 ] Амебы — это одноклеточные протисты , которые отделились от дерева и превратились в многоклеточных животных вскоре после дивергенции растений, и они разделяют многие специфические функции с фагоцитирующими клетками млекопитающих. [ 134 ] Например, Dictyostelium discoideum — амеба, живущая в почве и питающаяся бактериями. Подобно фагоцитам животных, он поглощает бактерии путем фагоцитоза преимущественно через Toll-подобные рецепторы и имеет другие биологические функции, общие с макрофагами. [ 135 ] Dictyostelium discoideum социален; при голодании он агрегирует, образуя мигрирующий псевдоплазмодий или слизняк . Этот многоклеточный организм в конечном итоге образует плодовое тело со спорами , устойчивыми к опасностям окружающей среды. До образования плодовых тел клетки в течение нескольких дней мигрируют как слизнеобразный организм. В это время воздействие токсинов или бактериальных патогенов может поставить под угрозу выживание вида из-за ограничения производства спор. Некоторые из амеб поглощают бактерии и токсины, циркулируя внутри слизняка, и в конечном итоге эти амебы умирают. Они генетически идентичны другим амебам слизняка; их самопожертвование для защиты других амеб от бактерий похоже на самопожертвование фагоцитов, наблюдаемое в иммунной системе высших позвоночных. Эта древняя иммунная функция социальных амеб предполагает эволюционно консервативный клеточный механизм поиска пищи, который мог быть адаптирован к защитным функциям задолго до диверсификации амеб в более высокие формы. [ 136 ] Фагоциты встречаются во всем животном мире. [ 3 ] от морских губок до насекомых и низших и высших позвоночных. [ 137 ] [ 138 ] Способность амеб различать свое и чужое является ключевой и лежит в основе иммунной системы многих видов амеб. [ 8 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Литтл С., Фаулер Х.В., Коулсон Дж. (1983). Краткий Оксфордский словарь английского языка . Издательство Оксфордского университета (Издательство Гильдии). стр. 1566–67.
- ^ Jump up to: а б с д и ж г час я дж Делвес и др. 2006 , стр. 2–10.
- ^ Jump up to: а б Делвес и др. 2006 , с. 250
- ^ Делвес и др. 2006 , с. 251
- ^ Jump up to: а б с д Хоффбранд, Петтит и Мосс 2005 , с. 331
- ↑ Илья Мечников , получено 28 ноября 2008 г. Из Нобелевских лекций по физиологии и медицине 1901–1921 гг ., Elsevier Publishing Company, Амстердам, 1967. Архивировано 22 августа 2008 г., в Wayback Machine.
- ^ Jump up to: а б Шмальштиг, ФК; А. С. Гольдман (2008). «Илья Ильич Мечников (1845–1915) и Пауль Эрлих (1854–1915): столетие Нобелевской премии 1908 года по физиологии и медицине». Журнал медицинской биографии . 16 (2): 96–103. дои : 10.1258/jmb.2008.008006 . ПМИД 18463079 . S2CID 25063709 .
- ^ Jump up to: а б Джейнвей, Глава: Эволюция врожденной иммунной системы. получено 20 марта 2009 г.
- ^ Jump up to: а б Эрнст и Стендаль 2006 , стр. 186.
- ^ Jump up to: а б Робинсон и Бэбкок 1998 , с. 187 и Ernst & Stendahl 2006 , стр. 7–10.
- ^ Jump up to: а б Эрнст и Стендаль 2006 , стр. 10.
- ^ Jump up to: а б Томпсон CB (1995). «Апоптоз в патогенезе и лечении заболеваний». Наука . 267 (5203): 1456–62. Бибкод : 1995Sci...267.1456T . дои : 10.1126/science.7878464 . ПМИД 7878464 . S2CID 12991980 .
- ^ Jump up to: а б с Джейнвей, Глава: Индуцированные врожденные реакции на инфекцию.
- ^ Jump up to: а б Фанг ФК (октябрь 2004 г.). «Противомикробные активные формы кислорода и азота: концепции и противоречия». Нат. Преподобный Микробиол . 2 (10): 820–32. дои : 10.1038/nrmicro1004 . ПМИД 15378046 . S2CID 11063073 .
- ^ Jump up to: а б Делвес и др. 2006 , стр. 172–84.
- ^ Jump up to: а б с Кауфманн С.Х. (2019). «Взросление иммунологии» . Границы в иммунологии . 10 :684. дои : 10.3389/fimmu.2019.00684 . ПМЦ 6456699 . ПМИД 31001278 .
- ^ Атерман К. (1 апреля 1998 г.). «Медали, воспоминания — и Мечников» . Дж. Леукок. Биол . 63 (4): 515–17. дои : 10.1002/jlb.63.4.515 . ПМИД 9544583 . S2CID 44748502 .
- ^ «Илья Мечников» . Нобелевский фонд . Проверено 19 декабря 2014 г.
- ^ Делвес и др. 2006 , с. 263
- ^ Робинсон и Бэбкок 1998 , с. VII
- ^ Эрнст и Стендаль 2006 , стр. 6.
- ^ Эрнст и Стендаль 2006 , стр. 4.
- ^ Эрнст и Стендаль 2006 , с. 78
- ^ Фельдман М.Б., Вяс Дж.М., Мансур М.К. (май 2019 г.). «Для этого нужна деревня: фагоциты играют центральную роль в иммунитете к грибкам» . Семинары по клеточной биологии и биологии развития . 89 : 16–23. дои : 10.1016/j.semcdb.2018.04.008 . ПМК 6235731 . ПМИД 29727727 .
- ^ Jump up to: а б Хэмптон МБ, Виссерс МБ, Уинтерборн CC (февраль 1994 г.). «Единый анализ для измерения скорости фагоцитоза и уничтожения бактерий нейтрофилами» . Дж. Леукок. Биол . 55 (2): 147–52. дои : 10.1002/jlb.55.2.147 . ПМИД 8301210 . S2CID 44911791 . Архивировано из оригинала 28 декабря 2012 года . Проверено 19 декабря 2014 г.
- ^ Делвес и др. 2006 , стр. 6–7.
- ^ Sompayrac 2019 , с. 2
- ^ Sompayrac 2019 , с. 2
- ^ Sompayrac 2019 , стр. 13–16.
- ^ Фройнд И., Эйгенброд Т., Хелм М., Далпке А.Х. (январь 2019 г.). «Модификации РНК модулируют активацию врожденных Toll-подобных рецепторов» . Гены . 10 (2): 92. doi : 10.3390/genes10020092 . ПМК 6410116 . ПМИД 30699960 .
- ^ Дейл, округ Колумбия, Боксер Л, Лайлз У.К. (август 2008 г.). «Фагоциты: нейтрофилы и моноциты» . Кровь . 112 (4): 935–45. дои : 10.1182/кровь-2007-12-077917 . ПМИД 18684880 . S2CID 746699 .
- ^ Дальгрен, К; А. Карлссон (17 декабря 1999 г.). «Дыхательный взрыв нейтрофилов человека». Журнал иммунологических методов . 232 (1–2): 3–14. дои : 10.1016/S0022-1759(99)00146-5 . ПМИД 10618505 .
- ^ Шатвелл, КП; А.В. Сигал (1996). «НАДФН-оксидаза». Международный журнал биохимии и клеточной биологии . 28 (11): 1191–95. дои : 10.1016/S1357-2725(96)00084-2 . ПМИД 9022278 .
- ^ Клебанов С.Дж. (1999). «Миелопероксидаза». Учеб. доц. Являюсь. Врачи . 111 (5): 383–89. дои : 10.1111/paa.1999.111.5.383 . ПМИД 10519157 .
- ^ Мейер К.К. (сентябрь 2004 г.). «Нейтрофилы, миелопероксидаза и бронхоэктатическая болезнь при муковисцидозе: зеленый цвет вреден». Дж. Лаб. Клин. Мед . 144 (3): 124–26. дои : 10.1016/j.lab.2004.05.014 . ПМИД 15478278 .
- ^ Хоффбранд, Петтит и Мосс 2005 , стр. 118
- ^ Делвес и др. 2006 , стр. 6–10.
- ^ Шредер К., Герцог П.Дж., Раваси Т., Хьюм Д.А. (февраль 2004 г.). «Интерферон-гамма: обзор сигналов, механизмов и функций». Дж. Леукок. Биол . 75 (2): 163–89. дои : 10.1189/jlb.0603252 . ПМИД 14525967 . S2CID 15862242 .
- ^ Делвес и др. 2006 , с. 188
- ^ Jump up to: а б Сомпайрак 2019 , с. 136
- ^ Липу Х.Н., Ахмед Т.А., Али С., Ахмед Д., Вакар М.А. (сентябрь 2008 г.). «Хроническая гранулематозная болезнь». Джей Пак Мед Ассоц . 58 (9): 516–18. ПМИД 18846805 .
- ^ Каплан Дж., Де Доменико I, Уорд Д.М. (январь 2008 г.). «Синдром Чедиака-Хигаси». Курс. Мнение. Гематол . 15 (1): 22–29. doi : 10.1097/MOH.0b013e3282f2bcce . ПМИД 18043242 . S2CID 43243529 .
- ^ Sompayrac 2019 , с. 7
- ^ де Алмейда С.М., Ногейра М.Б., Рабони С.М., Видаль Л.Р. (октябрь 2007 г.). «Лабораторная диагностика лимфоцитарного менингита» . Браз Джей Заразить Дис . 11 (5): 489–95. дои : 10.1590/s1413-86702007000500010 . ПМИД 17962876 .
- ^ Sompayrac 2019 , с. 22
- ^ Sompayrac 2019 , с. 68
- ^ «Апоптоз» . Интернет-словарь Мерриам-Вебстера . Проверено 19 декабря 2014 г.
- ^ Ли М.О., Саркисян М.Р., Мехал В.З., Ракич П., Флавелл Р.А. (ноябрь 2003 г.). «Фосфатидилсериновый рецептор необходим для очистки апоптотических клеток». Наука . 302 (5650): 1560–63. дои : 10.1126/science.1087621 . ПМИД 14645847 . S2CID 36252352 . (Для онлайн-доступа необходима бесплатная регистрация)
- ^ Нагата С., Сакураги Т., Сегава К. (декабрь 2019 г.). «Флиппаза и скрамблаза для воздействия фосфатидилсерина» . Современное мнение в иммунологии . 62 : 31–38. дои : 10.1016/j.coi.2019.11.009 . ПМИД 31837595 .
- ^ Ван Икс (2003). «Поглощение трупа клетки, опосредованное фосфатидилсериновым рецептором C. elegans через CED-5 и CED-12» . Наука . 302 (5650): 1563–1566. Бибкод : 2003Sci...302.1563W . дои : 10.1126/science.1087641 . PMID 14645848 . S2CID 25672278 . [ постоянная мертвая ссылка ] (Для онлайн-доступа необходима бесплатная регистрация)
- ^ Сэвилл Дж., Грегори С., Хаслетт С. (2003). «Съешь меня или умри». Наука . 302 (5650): 1516–17. дои : 10.1126/science.1092533 . hdl : 1842/448 . ПМИД 14645835 . S2CID 13402617 .
- ^ Чжоу Цз, Юй Х (октябрь 2008 г.). «Созревание фагосом при удалении апоптотических клеток: рецепторы лидируют» . Тенденции клеточной биологии . 18 (10): 474–85. дои : 10.1016/j.tcb.2008.08.002 . ПМК 3125982 . ПМИД 18774293 .
- ^ Sompayrac 2019 , с. 3
- ^ Sompayrac 2019 , с. 4
- ^ Sompayrac 2019 , стр. 27–35.
- ^ Делвес и др. 2006 , стр. 171–184.
- ^ Делвес и др. 2006 , стр. 456.
- ^ Ли Т., МакГиббон А. (2004). «Антигенпрезентирующие клетки (АПК)» . Университет Далхаузи . Архивировано из оригинала 12 января 2008 года . Проверено 19 декабря 2014 г.
- ^ Делвес и др. 2006 , с. 161
- ^ Sompayrac 2019 , с. 8
- ^ Делвес и др. 2006 , стр. 237–242.
- ^ Ланге С., Дюрр М., Достер Х., Мелмс А., Бишоф Ф. (2007). «Взаимодействия дендритных клеток и регуляторных Т-клеток контролируют самонаправленный иммунитет». Иммунол. Клеточная Биол . 85 (8): 575–81. дои : 10.1038/sj.icb.7100088 . ПМИД 17592494 . S2CID 36342899 .
- ^ Штайнман, Ральф М. (2004). «Дендритные клетки и иммунная толерантность» . Университет Рокфеллера. Архивировано из оригинала 11 марта 2009 года . Проверено 19 декабря 2014 г.
- ^ Романьяни, С (2006). «Иммунологическая толерантность и аутоиммунитет». Внутренняя и неотложная медицина . 1 (3): 187–96. дои : 10.1007/BF02934736 . ПМИД 17120464 . S2CID 27585046 .
- ^ Sompayrac 2019 , стр. 16–17.
- ^ Sompayrac 2019 , стр. 18–19.
- ^ Делвес и др. 2006 , с. 6
- ^ Зен К., Паркос, Калифорния (октябрь 2003 г.). «Лейкоцитарно-эпителиальные взаимодействия». Курс. Мнение. Клеточная Биол . 15 (5): 557–64. дои : 10.1016/S0955-0674(03)00103-0 . ПМИД 14519390 .
- ^ Sompayrac 2019 , с. 18
- ^ Хоффбранд, Петтит и Мосс 2005 , стр. 117
- ^ Делвес и др. 2006 , стр. 1–6.
- ^ Sompayrac 2019 , с. 136
- ^ Такахаси К., Наито М., Такея М. (июль 1996 г.). «Развитие и гетерогенность макрофагов и родственных им клеток по путям их дифференцировки». Патол. Межд . 46 (7): 473–85. дои : 10.1111/j.1440-1827.1996.tb03641.x . ПМИД 8870002 . S2CID 6049656 .
- ^ Кромбах Ф., Мюнцинг С., Аллмелинг А.М., Герлах Дж.Т., Бер Дж., Дёргер М. (сентябрь 1997 г.). «Размер клеток альвеолярных макрофагов: межвидовое сравнение» . Окружающая среда. Перспектива здоровья . 105 (Приложение 5): 1261–63. дои : 10.2307/3433544 . JSTOR 3433544 . ПМК 1470168 . ПМИД 9400735 .
- ^ Jump up to: а б с д и Делвес и др. 2006 , стр. 31–36.
- ^ Эрнст и Стендаль 2006 , стр. 8.
- ^ Делвес и др. 2006 , с. 156
- ^ Делвес и др. 2006 , с. 187
- ^ Ствртинова, Вера; Ян Якубовский и Иван Хулин (1995). «Нейтрофилы, центральные клетки при остром воспалении» . Патофизиология воспаления и лихорадки: принципы заболевания . Вычислительный центр Словацкой академии наук: Академическая электронная пресса. ISBN 978-80-967366-1-4 . Архивировано из оригинала 31 декабря 2010 года . Проверено 19 декабря 2014 г.
- ^ Делвес и др. 2006 , с. 4
- ^ Jump up to: а б Сомпайрак 2019 , с. 18
- ^ Линдеркамп О., Рюф П., Бреннер Б., Гулбинс Э., Ланг Ф. (декабрь 1998 г.). «Пассивная деформируемость зрелых, незрелых и активных нейтрофилов у здоровых и септических новорожденных» . Педиатр. Рез . 44 (6): 946–50. дои : 10.1203/00006450-199812000-00021 . ПМИД 9853933 .
- ^ Паолетти, Нотарио и Рицевути 1997 , стр. 62
- ^ Зенлайн О., Кенне Э., Роциус П., Эрикссон Э.Э., Линдбом Л. (январь 2008 г.). «Продукты секреции нейтрофилов регулируют антибактериальную активность моноцитов и макрофагов» . Клин. Эксп. Иммунол . 151 (1): 139–45. дои : 10.1111/j.1365-2249.2007.03532.x . ПМК 2276935 . ПМИД 17991288 .
- ^ Зенлайн О., Кай-Ларсен Ю., Фритьоф Р. (октябрь 2008 г.). «Белки первичных гранул нейтрофилов HBP и HNP1-3 усиливают бактериальный фагоцитоз человеческими и мышиными макрофагами» . Дж. Клин. Инвестируйте . 118 (10): 3491–502. дои : 10.1172/JCI35740 . ПМК 2532980 . ПМИД 18787642 .
- ^ Папаянопулос V (февраль 2018 г.). «Нейтрофильные внеклеточные ловушки в иммунитете и заболеваниях». Обзоры природы. Иммунология . 18 (2): 134–147. дои : 10.1038/nri.2017.105 . ПМИД 28990587 . S2CID 25067858 .
- ^ Штейнман Р.М., Кон З.А. (1973). «Идентификация нового типа клеток в периферических лимфоидных органах мышей. I. Морфология, количественный анализ, распределение в тканях» . Дж. Эксп. Мед . 137 (5): 1142–62. дои : 10.1084/jem.137.5.1142 . ПМК 2139237 . ПМИД 4573839 .
- ^ Jump up to: а б Штейнман, Ральф. «Дендритные клетки» . Университет Рокфеллера. Архивировано из оригинала 27 июня 2009 года . Проверено 19 декабря 2014 г.
- ^ Гермонпре П., Валладо Дж., Зитвогель Л., Тери С., Амигорена С. (2002). «Презентация антигена и стимуляция Т-клеток дендритными клетками». Энн. Преподобный Иммунол . 20 :621–67. doi : 10.1146/annurev.immunol.20.100301.064828 . ПМИД 11861614 .
- ^ Хоффбранд, Петтит и Мосс 2005 , стр. 134
- ^ Саллюсто Ф, Ланзавеккья А (2002). «Поучительная роль дендритных клеток в ответах Т-клеток» . Артрит Рес . 4 (Приложение 3): S127–32. дои : 10.1186/ar567 . ПМК 3240143 . ПМИД 12110131 .
- ^ Sompayrac 2019 , стр. 45–46.
- ^ Новак Н., Бибер Т., Пэн В.М. (2010). «Сеть рецепторов, подобных иммуноглобулину E-Toll» . Международный архив аллергии и иммунологии . 151 (1): 1–7. дои : 10.1159/000232565 . ПМИД 19672091 . Проверено 19 декабря 2014 г.
- ^ Калесников Дж., Галли С.Дж. (ноябрь 2008 г.). «Новые разработки в биологии тучных клеток» . Природная иммунология . 9 (11): 1215–23. дои : 10.1038/ni.f.216 . ПМЦ 2856637 . ПМИД 18936782 .
- ^ Jump up to: а б Малавия Р., Авраам С.Н. (февраль 2001 г.). «Модуляция тучовыми клетками иммунного ответа на бактерии». Иммунол. Преподобный . 179 : 16–24. дои : 10.1034/j.1600-065X.2001.790102.x . ПМИД 11292019 . S2CID 23115222 .
- ^ Коннелл И., Агаче В., Клемм П., Шембри М., Мэрилд С., Сванборг К. (сентябрь 1996 г.). «Экспрессия фимбрионов типа 1 повышает вирулентность Escherichia coli для мочевыводящих путей» . Учеб. Натл. акад. наук. США . 93 (18): 9827–32. Бибкод : 1996PNAS...93.9827C . дои : 10.1073/pnas.93.18.9827 . ПМК 38514 . ПМИД 8790416 .
- ^ Малавия Р., Твестен, штат Нью-Джерси, Росс Э.А., Авраам С.Н., Пфайфер Дж.Д. (февраль 1996 г.). «Тучные клетки перерабатывают бактериальные антигены посредством фагоцитов для презентации MHC класса I Т-клеткам» . Дж. Иммунол . 156 (4): 1490–96. дои : 10.4049/jimmunol.156.4.1490 . ПМИД 8568252 . S2CID 7917861 . Проверено 19 декабря 2014 г.
- ^ Тейлор М.Л., Меткалф Д.Д. (2001). «Тучные клетки при аллергии и защите хозяина». Аллергия Астма Учебник . 22 (3): 115–19. дои : 10.2500/108854101778148764 . ПМИД 11424870 .
- ^ Урб М., Шеппард, округ Колумбия (2012). «Роль тучных клеток в защите от патогенов» . ПЛОС Патогены . 8 (4): e1002619. дои : 10.1371/journal.ppat.1002619 . ПМЦ 3343118 . ПМИД 22577358 .
- ^ Jump up to: а б Паолетти, Нотарио и Ричевути 1997 , с. 427
- ^ Бирге Р.Б., Укер Д.С. (июль 2008 г.). «Врожденный апоптотический иммунитет: успокаивающее прикосновение смерти» . Гибель клеток отличается . 15 (7): 1096–1102. дои : 10.1038/cdd.2008.58 . ПМИД 18451871 .
- ^ Кузине С., Сехас Э., Шиттни Дж., Деплазес П., Вебер Р., Циммерли С. (декабрь 2000 г.). «Фагоцитарное поглощение Encephalitozoon cuniculi непрофессиональными фагоцитами» . Заразить. Иммунитет . 68 (12): 6939–45. дои : 10.1128/IAI.68.12.6939-6945.2000 . ПМК 97802 . ПМИД 11083817 .
- ^ Сигал Дж., Ли В., Арора П.Д., Макки М., Дауни Дж., Маккаллох Калифорния (январь 2001 г.). «Участие актиновых нитей и интегринов на этапе связывания при фагоцитозе коллагена фибробластами человека». Журнал клеточной науки . 114 (Часть 1): 119–129. дои : 10.1242/jcs.114.1.119 . ПМИД 11112696 .
- ^ Рабинович М. (март 1995 г.). «Профессиональные и непрофессиональные фагоциты: введение». Тенденции клеточной биологии . 5 (3): 85–87. дои : 10.1016/S0962-8924(00)88955-2 . ПМИД 14732160 .
- ^ Лин А, Лоре К (2017). «Гранулоциты: новые члены семейства антигенпредставляющих клеток» . Границы в иммунологии . 8 : 1781. дои : 10.3389/fimmu.2017.01781 . ПМЦ 5732227 . ПМИД 29321780 .
- ^ Дэвис С.П., Терри Л.В., Уилкинсон А.Л., Стаматаки З. (2020). «Межклеточные структуры в печени: история четырех Е» . Границы в иммунологии . 11 : 650. дои : 10.3389/fimmu.2020.00650 . ПМЦ 7247839 . ПМИД 32528462 .
- ^ Jump up to: а б с д и Тодар, Кеннет. «Механизмы бактериальной патогенности: защита бактерий от фагоцитов» . 2008 год . Проверено 19 декабря 2014 г.
- ^ Александр Дж., Сатоскар А.Р., Рассел Д.Г. (сентябрь 1999 г.). «Виды Leishmania: модели внутриклеточного паразитизма» . Дж. Клеточная наука . 112 (18): 2993–3002. дои : 10.1242/jcs.112.18.2993 . ПМИД 10462516 . Проверено 19 декабря 2014 г.
- ^ Челли Дж., Финли Б.Б. (май 2002 г.). «Бактериальное избегание фагоцитоза». Тенденции Микробиол . 10 (5): 232–37. дои : 10.1016/S0966-842X(02)02343-0 . ПМИД 11973157 .
- ^ Валеник Л.В., Ся ХК, Шварцбауэр Дж.Э. (сентябрь 2005 г.). «Фрагментация фибронектина способствует опосредованному альфа4бета1-интегрином сокращению временной матрицы фибрин-фибронектин». Экспериментальные исследования клеток . 309 (1): 48–55. doi : 10.1016/j.yexcr.2005.05.024 . ПМИД 15992798 .
- ^ Бернс С.М., Халл С.И. (август 1999 г.). «Потеря устойчивости к проглатыванию и фагоцитарному уничтожению мутантами O (-) и K (-) уропатогенного штамма Escherichia coli O75: K5» . Заразить. Иммунитет . 67 (8): 3757–62. дои : 10.1128/IAI.67.8.3757-3762.1999 . ПМК 96650 . ПМИД 10417134 .
- ^ Вуонг С., Коцианова С., Войич Дж.М. (декабрь 2004 г.). «Решающая роль модификации экзополисахаридов в формировании бактериальной биопленки, уклонении от иммунитета и вирулентности» . Ж. Биол. Хим . 279 (52): 54881–86. дои : 10.1074/jbc.M411374200 . ПМИД 15501828 .
- ^ Мелин М., Ярва Х., Сиира Л., Мери С., Кайхти Х., Вякевяйнен М. (февраль 2009 г.). « Капсульный серотип 19F Streptococcus pneumoniae более устойчив к отложению С3 и менее чувствителен к опсонофагоцитозу, чем серотип 6В» . Заразить. Иммунитет . 77 (2): 676–84. дои : 10.1128/IAI.01186-08 . ПМК 2632042 . ПМИД 19047408 .
- ^ Jump up to: а б Фостер Ти Джей (декабрь 2005 г.). «Уклонение от иммунитета стафилококков». Нат. Преподобный Микробиол . 3 (12): 948–58. дои : 10.1038/nrmicro1289 . ПМИД 16322743 . S2CID 205496221 .
- ^ Феллман М., Делей Ф., МакГи К. (февраль 2002 г.). «Устойчивость к фагоцитозу иерсиний». Международный журнал медицинской микробиологии . 291 (6–7): 501–9. дои : 10.1078/1438-4221-00159 . ПМИД 11890550 .
- ^ Сансонетти П. (декабрь 2001 г.). «Фагоцитоз бактериальных патогенов: последствия для реакции хозяина». Семин. Иммунол . 13 (6): 381–90. дои : 10.1006/smim.2001.0335 . ПМИД 11708894 .
- ^ Дерш П., Исберг Р.Р. (март 1999 г.). «Участок белка инвазина Yersinia pseudotuberculosis усиливает опосредованное интегрином поглощение клетками млекопитающих и способствует самоассоциации» . ЭМБО Дж . 18 (5): 1199–1213. дои : 10.1093/emboj/18.5.1199 . ПМЦ 1171211 . ПМИД 10064587 .
- ^ Антуан Ж.К., Прина Э., Ланг Т., Курре Н. (октябрь 1998 г.). «Биогенез и свойства паразитофорных вакуолей, содержащих Leishmania в мышиных макрофагах». Тенденции Микробиол . 6 (10): 392–401. дои : 10.1016/S0966-842X(98)01324-9 . ПМИД 9807783 .
- ^ Дас Д., Саха СС, Бишайи Б (июль 2008 г.). «Внутриклеточная выживаемость Staphylococcus aureus : корреляция продукции каталазы и супероксиддисмутазы с уровнями воспалительных цитокинов». Воспаление. Рез . 57 (7): 340–49. дои : 10.1007/s00011-007-7206-z . ПМИД 18607538 . S2CID 22127111 .
- ^ Хара Х, Кавамура И, Номура Т, Томинага Т, Цучия К, Мицуяма М (август 2007 г.). «Цитолизин-зависимый выход бактерии из фагосомы необходим, но недостаточен для индукции иммунного ответа Th1 против инфекции Listeria monocytogenes: особая роль листериолизина О определяется заменой гена цитолизина» . Заразить. Иммунитет . 75 (8): 3791–3801. дои : 10.1128/IAI.01779-06 . ЧВК 1951982 . ПМИД 17517863 .
- ^ Jump up to: а б Паркер Х.А., Форрестер Л., Калдор К.Д., Дикерхоф Н., Хэмптон М.Б. (2021). «Антимикробная активность нейтрофилов в отношении микобактерий» . Границы в иммунологии . 12 : 782495. дои : 10.3389/fimmu.2021.782495 . ПМЦ 8732375 . ПМИД 35003097 .
- ^ Датта В., Мысковский С.М., Квинн Л.А., Чием Д.Н., Варки Н., Кансал Р.Г., Котб М., Низет В. (май 2005 г.). «Мутационный анализ стрептококкового оперона группы А, кодирующего стрептолизин S, и его роль вирулентности при инвазивной инфекции». Мол. Микробиол . 56 (3): 681–95. дои : 10.1111/j.1365-2958.2005.04583.x . ПМИД 15819624 . S2CID 14748436 .
- ^ Ивацуки К., Ямасаки О., Моризан С., Ооно Т. (июнь 2006 г.). «Стафилококковые кожные инфекции: инвазия, уклонение и агрессия». Дж. Дерматол. Наука 42 (3): 203–14. дои : 10.1016/j.jdermsci.2006.03.011 . ПМИД 16679003 .
- ^ Jump up to: а б Денкерс Э.Ю., Батчер Б.А. (январь 2005 г.). «Саботаж и эксплуатация макрофагов, паразитируемых внутриклеточными простейшими». Тенденции Паразитол . 21 (1): 35–41. дои : 10.1016/j.pt.2004.10.004 . ПМИД 15639739 .
- ^ Грегори DJ, Оливье М (2005). «Подрыв передачи сигналов клетки-хозяина простейшим паразитом Leishmania ». Паразитология . 130 Приложение: С27–35. дои : 10.1017/S0031182005008139 . ПМИД 16281989 . S2CID 24696519 .
- ^ Паолетти стр. 426–30
- ^ Хайнцельманн М., Мерсер-Джонс М.А., Пассмор Дж.К. (август 1999 г.). «Нейтрофилы и почечная недостаточность». Являюсь. Дж. Почки Дис . 34 (2): 384–99. дои : 10.1016/S0272-6386(99)70375-6 . ПМИД 10430993 .
- ^ Ли В.Л., врач Дауни (февраль 2001 г.). «Активация нейтрофилов и острое повреждение легких». Curr Opin Crit Care . 7 (1): 1–7. дои : 10.1097/00075198-200102000-00001 . ПМИД 11373504 . S2CID 24164360 .
- ^ Jump up to: а б Мораес Т.Дж., Журавска Дж.Х., Дауни Г.П. (январь 2006 г.). «Содержимое нейтрофильных гранул в патогенезе повреждения легких». Курс. Мнение. Гематол . 13 (1): 21–27. дои : 10.1097/01.moh.0000190113.31027.d5 . ПМИД 16319683 . S2CID 29374195 .
- ^ Авраам Э. (апрель 2003 г.). «Нейтрофилы и острое повреждение легких». Крит. Уход Мед . 31 (4 приложения): S195–99. дои : 10.1097/01.CCM.0000057843.47705.E8 . ПМИД 12682440 . S2CID 4004607 .
- ^ Ричевути Г (декабрь 1997 г.). «Повреждение тканей хозяина фагоцитами». Энн. Н-Й акад. Наука . 832 (1): 426–48. Бибкод : 1997NYASA.832..426R . дои : 10.1111/j.1749-6632.1997.tb46269.x . ПМИД 9704069 . S2CID 10318084 .
- ^ Чарли Б., Риффо С., Ван Рит К. (октябрь 2006 г.). «Врожденный и адаптивный иммунный ответ свиней на грипп и коронавирусную инфекцию» . Энн. Н-Й акад. Наука . 1081 (1): 130–36. Бибкод : 2006NYASA1081..130C . дои : 10.1196/анналы.1373.014 . hdl : 1854/LU-369324 . ПМК 7168046 . ПМИД 17135502 .
- ^ Sompayrac 2019 , с. 2
- ^ Jump up to: а б Коссон П., Солдати Т. (июнь 2008 г.). «Ешь, убей или умри: когда амеба встречается с бактериями». Курс. Мнение. Микробиол . 11 (3): 271–76. дои : 10.1016/j.mib.2008.05.005 . ПМИД 18550419 .
- ^ Боззаро С., Буччи С., Штайнерт М. (2008). «Фагоцитоз и взаимодействие хозяин-патоген у Dictyostelium с взглядом на макрофаги». Международное обозрение клеточной и молекулярной биологии . Том. 271. С. 253–300. дои : 10.1016/S1937-6448(08)01206-9 . ISBN 978-0-12-374728-0 . ПМИД 19081545 . S2CID 7326149 .
- ^ Чен Г, Жученко О, Куспа А (август 2007 г.). «Иммуноподобная активность фагоцитов у социальной амебы» . Наука 317 (5838): 678–81. Бибкод : 2007Sci...317..678C . дои : 10.1126/science.1143991 . ПМК 3291017 . ПМИД 17673666 .
- ^ Делвес и др. 2006 , стр. 251–252.
- ^ Ханингтон ПК, Тэм Дж., Катценбек Б.А., Хитчен С.Дж., Барреда Д.Р., Белошевич М. (апрель 2009 г.). «Развитие макрофагов карповых рыб». Дев. Комп. Иммунол . 33 (4): 411–29. дои : 10.1016/j.dci.2008.11.004 . ПМИД 19063916 .
Библиография
[ редактировать ]- Дельвес, П.Дж.; Мартин, С.Дж.; Бертон, ДР; Ройт, И.М. (2006). Основная иммунология Ройта (11-е изд.). Молден, Массачусетс: Blackwell Publishing. ISBN 978-1-4051-3603-7 .
- Эрнст, доктор медицинских наук; Стендаль, О., ред. (2006). Фагоцитоз бактерий и бактериальная патогенность . Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-84569-4 . Веб-сайт
- Хоффбранд, А.В.; Петтит, Дж. Э.; Мосс, ПАУ (2005). Основная гематология (4-е изд.). Лондон: Blackwell Science. ISBN 978-0-632-05153-3 .
- Паолетти, Р.; Нотарио, А.; Ричевути, Г., ред. (1997). Фагоциты: биология, физиология, патология и фармакотерапия . Нью-Йорк: Нью-Йоркская академия наук. ISBN 978-1-57331-102-1 .
- Робинсон, JP; Бэбкок, Г. Ф., ред. (1998). Функция фагоцитов. Руководство для исследований и клинической оценки . Нью-Йорк: Уайли-Лисс. ISBN 978-0-471-12364-4 .
- Сомпайрак, Л. (2019). Как работает иммунная система (6-е изд.). Молден, Массачусетс: Blackwell Publishing. ISBN 978-1-119-54212-4 .
Внешние ссылки
[ редактировать ]- Фагоциты Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Лейкоциты, поглощающие бактерии