Инфузория
| Инфузория Временной диапазон:
| |
|---|---|
| |
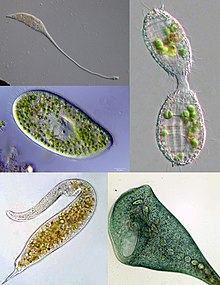
| Некоторые примеры разнообразия инфузорий. По часовой стрелке сверху слева: Лакримария , Колепс , Стентор , Дилептус , Парамеций. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | ЦАРЬ |
| Клэйд : | САР |
| Клэйд : | Альвеолата |
| Тип: | Цилиофора Дофлейн , 1901 г., исправление. |
| Подтипы и классы [ 1 ] | |
|
См. текст для подклассов. | |
| Синонимы | |
| |
Инфузории характеризующихся представляют собой группу альвеолятов, наличием волосообразных органелл, называемых ресничками , которые по структуре идентичны эукариотическим жгутикам , но в целом короче и присутствуют в гораздо большем количестве, с волнистым рисунком, отличным от жгутиков. Реснички встречаются у всех членов группы (хотя у своеобразных Suctoria они есть только в течение части жизненного цикла ) и по-разному используются при плавании, ползании, прикреплении, питании и ощущениях.
Инфузории — важная группа простейших , распространенная практически везде, где есть вода — в озерах, прудах, океанах, реках и почвах, включая бескислородные и обедненные кислородом среды обитания. [ 2 ] Описано около 4500 уникальных свободноживущих видов, а потенциальное число современных видов оценивается в 27 000–40 000. [ 3 ] В это число входят многие эктосимбиотические и эндосимбиотические виды, а также некоторые облигатные и условно- патогенные паразиты . Виды инфузорий имеют размер от всего лишь 10 мкм у некоторых колподий до целых 4 мм в длину у некоторых гелеид и включают некоторые из наиболее морфологически сложных простейших. [ 4 ] [ 5 ]
В большинстве систем систематики « Ciliophora » причисляется к типу. [ 6 ] под любым из нескольких королевств , включая Хромисту , [ 7 ] Протиста [ 8 ] или простейшие . [ 9 ] В некоторых старых системах классификации, таких как влиятельные таксономические работы Альфреда Каля , реснитчатые простейшие относят к классу « Ciliata ». [ 10 ] [ 11 ] (термин, который также может относиться к роду рыб ). В таксономической схеме, одобренной Международным обществом протистологов , которая исключает формальные обозначения рангов , такие как «тип» и «класс», «Ciliophora» является таксоном без ранга внутри Alveolata . [ 12 ] [ 13 ]
Клеточная структура
[ редактировать ]
Ядра
[ редактировать ]В отличие от большинства других эукариот , инфузории имеют два разных типа ядер : крошечное диплоидное микроядро («генеративное ядро», несущее зародышевую линию клетки) и большое амплиплоидное макронуклеус («вегетативное ядро», которое заботится о общей клеточной регуляции, выражающей фенотип организма). [ 14 ] [ 15 ] Последний генерируется из микроядра путем амплификации генома и тяжелого редактирования. Микроядро передает свой генетический материал потомству, но не экспрессирует свои гены. Макронуклеус обеспечивает малую ядерную РНК для вегетативного роста. [ 16 ] [ 15 ]
Деление макронуклеуса происходит у большинства видов инфузорий, за исключением представителей класса Karyorelictea, у которых макронуклеусы заменяются каждый раз при делении клетки. [ 17 ] Макронуклеарное деление осуществляется путем амитоза , а сегрегация хромосом происходит в результате процесса, механизм которого неизвестен. [ 15 ] Через определенное количество поколений (200–350 у Paramecium aurelia и до 1500 у Tetrahymena [ 17 ] ) клетка проявляет признаки старения, и макронуклеусы должны регенерироваться из микроядер. Обычно это происходит после конъюгации , после которой из постконъюгального микронуклеуса образуется новый макронуклеус. [ 15 ]
Цитоплазма
[ редактировать ]Пищевые вакуоли образуются в результате фагоцитоза и обычно следуют определенному пути через клетку, поскольку их содержимое переваривается и расщепляется лизосомами , поэтому вещества, содержащиеся в вакуоли, затем становятся достаточно маленькими, чтобы диффундировать через мембрану пищевой вакуоли в клетку. Все, что остается в пищевой вакуоли к моменту достижения цитопрокта ( анальной поры ), высвобождается путем экзоцитоза . У большинства инфузорий также есть одна или несколько заметных сократительных вакуолей , которые собирают воду и вытесняют ее из клетки для поддержания осмотического давления или для поддержания ионного баланса. У некоторых родов, таких как Paramecium , они имеют характерную форму звезды, каждая точка которой представляет собой собирательную трубку.
Специализированные структуры инфузорий
[ редактировать ]
Преимущественно реснички тела расположены в моно- и дикинетидах , которые включают соответственно одну и две кинетосомы (базальные тельца), каждая из которых может поддерживать ресничку. Они расположены в ряды, называемые кинетиями , которые идут от передней части клетки к задней. Тело и оральные кинетиды составляют инфрацилиатуру — организацию, уникальную для инфузорий и важную для их классификации, и включают различные фибриллы и микротрубочки, участвующие в координации ресничек. У некоторых форм встречаются и телесные поликинетиды, например, у спиротрихов , у которых они обычно образуют щетинки, называемые усами .
Инфрацилиатура — один из основных компонентов клеточной коры . Другие — это альвеолы , небольшие пузырьки под клеточной мембраной, которые прилегают к ней, образуя пленку, поддерживающую форму клетки, которая варьируется от гибкой и сократительной до жесткой. многочисленные митохондрии и экструсомы Также обычно присутствуют . Наличие альвеол, строение ресничек, форма митоза и различные другие детали указывают на тесное родство инфузорий, Apicomplexa и динофлагеллят . Эти внешне несходные группы составляют альвеолаты .
Кормление
[ редактировать ]Большинство инфузорий являются гетеротрофами , питающимися более мелкими организмами, такими как бактерии и водоросли , а детрит заносится в ротовую бороздку (рот) модифицированными ротовыми ресничками. Обычно это включает ряд мембран слева от рта и околоротовую мембрану справа, обе из которых возникают из поликинетид , групп многих ресничек вместе с соответствующими структурами. Пища продвигается ресничками через ротовую пору в пищевод, где образуются пищевые вакуоли.
Многие виды также являются миксотрофами , сочетая фаготрофию и фототрофию посредством клептопластики или симбиоза с фотосинтезирующими микробами. [ 18 ] [ 19 ]
Было замечено, что инфузория Halteria питается хлорвирусами . [ 20 ]
Однако методы кормления значительно различаются. Некоторые инфузории лишены рта и питаются путем всасывания ( осмотрофия ), другие являются хищными и питаются другими простейшими и в частности другими инфузориями. Некоторые инфузории паразитируют на животных , хотя известно, что только один вид, Balantidium coli , вызывает заболевания у человека. [ 21 ]
Репродукция и сексуальные явления
[ редактировать ]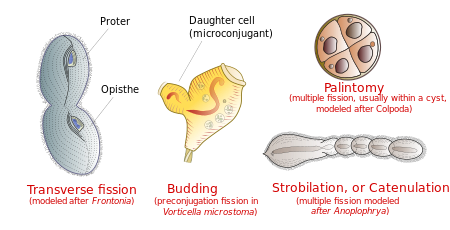
Воспроизведение
[ редактировать ]Инфузории размножаются бесполым путем , путем различных видов деления . [ 17 ] Во время деления микронуклеус подвергается митозу , а макронуклеус удлиняется и подвергается амитозу (за исключением кариореликтовых инфузорий, макронуклеусы которых не делятся). Затем клетка делится на две части, и каждая новая клетка получает копию микронуклеуса и макронуклеуса.

Обычно клетка делится поперечно: передняя половина инфузории ( протер ) образует один новый организм, а задняя половина ( описта ) образует другой. Однако в некоторых группах инфузорий встречаются и другие типы деления. К ним относятся почкование (появление мелких ресничных потомков, или «роевиков», из тела зрелого родителя); стробилация (множественные деления по телу клетки, образующие цепочку новых организмов); и палинтомия (множественные деления, обычно внутри кисты ). [ 22 ]
Деление может происходить спонтанно, как часть вегетативного клеточного цикла . Альтернативно оно может протекать в результате самооплодотворения ( автогамия ), [ 23 ] или это может следовать за конъюгацией , сексуальным феноменом, при котором инфузории совместимых типов спаривания обмениваются генетическим материалом. Хотя конъюгацию иногда называют формой размножения, она не связана напрямую с репродуктивными процессами и не приводит напрямую к увеличению числа отдельных инфузорий или их потомства. [ 24 ]
Спряжение
[ редактировать ]- Обзор
Конъюгация инфузорий — это половой феномен, который приводит к генетической рекомбинации и ядерной реорганизации внутри клетки. [ 24 ] [ 22 ] Во время конъюгации две инфузории совместимого типа спаривания образуют мостик между своими цитоплазмами . Микроядра подвергаются мейозу , макронуклеусы исчезают, и происходит обмен гаплоидными микроядрами по мостику. У некоторых инфузорий (перитрихов, хонотрихов и некоторых суккторий ) конъюгирующие клетки постоянно сливаются, и один конъюгант поглощается другим. [ 21 ] [ 25 ] Однако в большинстве групп инфузорий клетки после конъюгации разделяются, и обе образуют новые макроядра из своих микроядер. [ 26 ] За конъюгацией и автогамией всегда следует деление. [ 22 ]
У многих инфузорий, например Paramecium , конъюгирующие партнеры (гамонты) сходны или неразличимы по размеру и форме. Это называется «изогамонтическим» сопряжением. В некоторых группах партнеры различаются по размеру и форме. Это называется «анизогамонтной» конъюгацией. у сидячих перитрихов Например, один половой партнер (микроконъюгант) маленький и подвижный, а другой (макроконъюгант) крупный и сидячий . [ 24 ]
- Стадии конъюгации

Стадии конъюгации у Paramecium caudatum

У Paramecium caudatum этапы конъюгации следующие (см. схему справа):
- Совместимые спаривающиеся штаммы встречаются и частично сливаются.
- Микроядра подвергаются мейозу, образуя четыре гаплоидных микроядра на клетку.
- Три из этих микроядер распадаются. Четвертый подвергается митозу.
- Две клетки обмениваются микроядрами.
- Затем клетки разделяются.
- Микроядра в каждой клетке сливаются, образуя диплоидное микроядро.
- Митоз происходит трижды, в результате чего образуется восемь микроядер.
- Четыре новых микроядра трансформируются в макронуклеусы, а старый макронуклеус распадается.
- Бинарное деление происходит дважды, в результате чего образуются четыре идентичные дочерние клетки.
Перестройки ДНК (скремблирование генов)
[ редактировать ]Инфузории содержат два типа ядер: соматические « макронуклеусы » и зародышевые « микронуклеусы ». Во время полового размножения (конъюгации) передается только ДНК в микроядре. С другой стороны, только ДНК в макронуклеусе активно экспрессируется и влияет на фенотип организма. Макронуклеарная ДНК происходит из микроядерной ДНК путем поразительно обширной перестройки и амплификации ДНК.
Макронуклеус начинается как копия микронуклеуса. Микроядерные хромосомы фрагментируются на множество более мелких частей и амплифицируются, образуя множество копий. Образующиеся макронуклеарные хромосомы часто содержат только один ген . У Tetrahymena микроядро имеет 10 хромосом (по пять на гаплоидный геном), тогда как макронуклеус имеет более 20 000 хромосом. [ 27 ]
Кроме того, микроядерные гены прерываются многочисленными «внутренними элиминированными последовательностями» (IES). Во время развития макронуклеуса IES удаляются, а оставшиеся сегменты гена, макронуклеарные последовательности (MDS), соединяются вместе, образуя рабочий ген. Tetrahymena содержит около 6000 IES, и во время этого процесса удаляется около 15% микроядерной ДНК. Процесс управляется малыми РНК и эпигенетическими метками хроматина . [ 27 ]
У спиротриховых инфузорий (таких как Oxytricha ) этот процесс еще более сложен из-за «перемешивания генов»: MDS в микроядре часто располагаются в другом порядке и ориентации, чем в макронуклеарном гене, поэтому помимо делеции происходит инверсия ДНК. и транслокация необходимы для «расшифровки». Этот процесс управляется длинными РНК, происходящими из родительского макронуклеуса. Более 95% микроядерной ДНК удаляется во время развития макроядер спиротрихов. [ 27 ]
Старение
[ редактировать ]В клональных популяциях Paramecium старение происходит в течение последующих поколений, что приводит к постепенной потере жизнеспособности, если только клеточная линия не оживляется путем конъюгации или автогамии . У Paramecium тетраурелия клонально стареющая линия теряет жизнеспособность и погибает примерно после 200 делений, если клеточная линия не омолаживается путем конъюгации или самооплодотворения. Основы клонального старения были разъяснены экспериментами по трансплантации Ауфдерхайда в 1986 году. [ 28 ] который продемонстрировал, что макронуклеус, а не цитоплазма, ответственен за клональное старение. Дополнительные эксперименты Смита-Зоннеборна, [ 29 ] Холмс и Холмс, [ 30 ] и Гилли и Блэкберн [ 31 ] продемонстрировали, что во время клонального старения повреждение ДНК резко увеличивается. Таким образом, повреждение ДНК, по-видимому, является причиной старения P. Tetraurelia .
Ископаемая запись
[ редактировать ]До недавнего времени самыми старыми известными окаменелостями инфузорий были тинтинниды ордовикского периода . В 2007 году Ли и др. опубликовал описание ископаемых инфузорий из формации Доушантуо около 580 миллионов лет назад, в эдиакарский период . В их число входили два типа тинтиннид и возможный предок сукториан. [ 32 ] Ископаемая вортицелла была обнаружена внутри кокона пиявки триасового периода , около 200 миллионов лет назад. [ 33 ]
Филогения
[ редактировать ]По данным филогенетического анализа 2016 г. [ 1 ] Mesodiniea неизменно считается сестринской группой для всех других инфузорий. выделяют две большие подгруппы Кроме того, внутри подтипа Intramacronucleata : SAL ( Spirotrichea + Armophorea + Litostomatea ) и CONthreeP или Ventrata ( Colpodea + Oligohymenophorea + Nassophorea + Phyllopharyngea + Plagiopylea + Prostomatea ). [ 1 ] Класс Protocruziea является сестринской группой Ventrata / CONthreeP . Класс Cariacotrichea был исключен из анализа, но первоначально он был установлен в составе Intramacronucleata. [ 1 ] .
Одонтостоматиды . были идентифицированы в 2018 году [ 34 ] как отдельный класс Odontostomatea , родственный Armophorea .
| Цилиофора | |
Классификация
[ редактировать ]
Для инфузорий было предложено несколько различных схем классификации. Следующая схема основана на молекулярно- филогенетическом анализе до четырех генов из 152 видов, представляющих 110 семейств: [ 1 ]
- Класс Mesodiniea (например, Mesodinium )
Подтип Постцилиодесматофора
[ редактировать ]- Класс Heterotrichea (например, Stentor )
- Класс Кариореликтея
Подтип Intramacronucleata
[ редактировать ]
- Класс Армофория
- Класс Одонтостоматеа. [ 34 ] (например, дискоморфелла , сапродиниум )
- Класс Cariacotrichea (только один вид — Cariacothrix caudata ).
- Класс Муранотрихея
- Класс Параблефариммея
- Класс Colpoda (например, Colpoda )
- Класс Литостоматы
- Подкласс Haptoria (например, Didinium )
- Подкласс Ринхостоматия
- Подкласс Trichostomatia (например, Balantidium )
- Класс Насофореи
- Класс Филлофарингеи
- Подкласс Хонотрихия
- Подкласс Циртофория
- Подкласс Ринходии
- Подкласс Suctoria (например, Podophyra )
- Подкласс Сингимения
- Класс Олигогименофореи
- Подкласс Апостоматия
- Подкласс Астоматия
- Подкласс Hymenostomatia (например, Tetrahymena )
- Подкласс Peniculia (например, Paramecium )
- Подкласс Peritrichia (например, Vorticella )
- Подкласс Scuticociliatia
- Класс Плагиопилеи
- Class Prostomatea (e.g. Coleps )
- Класс Протокрузия
- Класс Спиротрихеи
- Подкласс Хореотрихия
- Подкласс Эуплотия
- Подкласс Гипотрихия
- Подкласс Ликнофория
- Подкласс Олиготрихия
- Подкласс Факодинидеа
- Подкласс Протогипотрихии
Другой
[ редактировать ]Некоторые старые классификации включали Opalinidae в состав инфузорий. Принципиальным отличием многоресничных жгутиконосцев (например, гемимастигид , Стефанопогона , Мультицилий , опалинов ) от инфузорий является наличие макронуклеусов только у инфузорий. [ 35 ]
Патогенность
[ редактировать ]Единственным представителем отдела инфузорий, патогенным для человека, является Balantidium coli . [ 36 ] [ 37 ] который вызывает заболевание балантидиаз . Он не патогенен для домашних свиней, являющихся основным резервуаром этого патогена. [ 38 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Гао, Фэн; Уоррен, Алан; Чжан, Цяньцянь; Гонг, Цзюнь; Мяо, Мяо; Солнце, Пинг; Сюй, Дапэн; Хуан, Цзе; Йи, Чжэньчжэнь (29 апреля 2016 г.). «Эволюционная гипотеза ресничных простейших, основанная на всех данных, с пересмотренной классификацией типа Ciliophora (Eukaryota, Alveolata)» . Научные отчеты . 6 : 24874. Бибкод : 2016NatSR...624874G . дои : 10.1038/srep24874 . ISSN 2045-2322 . ПМЦ 4850378 . ПМИД 27126745 .
- ^ Роттерова, Дж.; Эджкомб, вице-президент; Чепичка, И.; Бейнарт, Р. (2022). «Анаэробные инфузории как модельная группа для изучения симбиозов в обедненных кислородом средах» . Журнал эукариотической микробиологии . 69 (5): e12912. дои : 10.1111/jeu.12912 . ПМИД 35325496 . S2CID 247677842 .
- ^ Фойснер, В.; Хоксворт, Дэвид, ред. (2009). Разнообразие и географическое распространение протистов . Темы биоразнообразия и охраны природы. Том. 8. Спрингер Нидерланды. п. 111. дои : 10.1007/978-90-481-2801-3 . ISBN 9789048128006 .
- ^ Нильсен, Торкель Гиссель; Кьёрбо, Томас (1994). «Регуляция биомассы и продукции зоопланктона в прибрежной экосистеме умеренного климата. 2. Инфузории» . Лимнология и океанография . 39 (3): 508–519. Бибкод : 1994LimOc..39..508N . дои : 10.4319/lo.1994.39.3.0508 .
- ^ Линн, Денис (2008). Ресничные простейшие, 3-е издание . Спрингер. стр. 129 . ISBN 978-1-4020-8238-2 .
- ^ «Отчет ИТИС» . Интегрированная таксономическая информационная система . Проверено 11 декабря 2014 г.
- ^ Кавальер-Смит, Томас (01 января 2018 г.). «Королевство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения» . Протоплазма . 255 (1): 297–357. дои : 10.1007/s00709-017-1147-3 . ISSN 1615-6102 . ПМЦ 5756292 . ПМИД 28875267 .
- ^ И З, Сун В, Клэмп Дж. С., Чен З, Гао С, Чжан Ц (декабрь 2008 г.). «Пересмотр систематических взаимоотношений внутри отряда Euplotida (Protista, Ciliophora) с использованием новых последовательностей гена, кодирующего рРНК малых субъединиц, и тестирование использования комбинированных наборов данных для построения филогении комплекса Diophrys». Мол. Филогенет. Эвол . 50 (3): 599–607. дои : 10.1016/j.ympev.2008.12.006 . ПМИД 19121402 .
- ^ Мяо М., Сун В., Чэнь З. и др. (2007). «Уникальная эуплотидная инфузория, Gastrocirrus (Protozoa, Ciliophora): оценка ее филогенетического положения, сделанная на основе последовательности гена малой субъединицы рРНК». Дж. Эукариот. Микробиол . 54 (4): 371–8. дои : 10.1111/j.1550-7408.2007.00271.x . ПМИД 17669163 . S2CID 25977768 .
- ^ Альфред Каль (1930). Urtiere или Protozoa I: Инфузория или Ciliata — Том I, общий раздел и простоматы .
- ^ «Медицинское определение CILIATA» . www.merriam-webster.com . Проверено 22 декабря 2017 г.
- ^ Адл, Сина М.; Басс, Дэвид; Лейн, Кристофер Э.; Лукеш, Юлиус; Шох, Конрад Л.; Смирнов Алексей; Агата, Сабина; Берни, Седрик; Браун, Мэтью В.; Бурки, Фабьен; Карденас, Пако (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ISSN 1550-7408 . ПМК 6492006 . ПМИД 30257078 .
- ^ Адл, Сина М.; и др. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 .
- ^ Райков, И.Б. (1969). «Макронуклеус инфузорий» . Исследования в области протозоологии . 3 :4–115. ISBN 9781483186146 .
- ^ Перейти обратно: а б с д Арчибальд, Джон М.; Симпсон, Аластер ГБ; Сламовиц, Клаудио Х., ред. (2017). Справочник протистов (2-е изд.). Международное издательство Спрингер. п. 691. ИСБН 978-3-319-28147-6 .
- ^ Прескотт, DM (июнь 1994 г.). «ДНК мерцательных простейших» . Микробиологические обзоры . 58 (2): 233–267. дои : 10.1128/мр.58.2.233-267.1994 . ISSN 0146-0749 . ПМК 372963 . ПМИД 8078435 .
- ^ Перейти обратно: а б с Х., Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и справочник по литературе . Нью-Йорк: Спрингер. п. 324. ИСБН 9781402082382 . ОСЛК 272311632 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Эстебан, ГФ; Фенчел, Т.; Финли, Би Джей (2010). «Миксотрофия у инфузорий» . Протист . 161 (5): 621–641. дои : 10.1016/j.protis.2010.08.002 . ПМИД 20970377 .
- ^ Альтенбургер, А.; Блоссом, HE; Гарсия-Куэтос, Л.; Якобсен, Х.Х.; Карстенсен, Дж.; Лундхольм, Н.; Хансен, П.Дж.; Харагути, Л.; Харагути, Л. (2020). «Диморфизм криптофитов - случай Teleaulax amphioxeia/Plagioselmis prolonga и его экологические последствия» . Достижения науки . 6 (37). Бибкод : 2020SciA....6.1611A . дои : 10.1126/sciadv.abb1611 . ПМК 7486100 . ПМИД 32917704 .
- ^ Делонг, Джон П.; Ван Эттен, Джеймс Л.; Аль-Амели, Зейна; Агаркова Ирина В.; Дуниган, Дэвид Д. (3 января 2023 г.). «Потребление вирусов возвращает энергию в пищевые цепи» . Труды Национальной академии наук . 120 (1): e2215000120. Бибкод : 2023PNAS..12015000D . дои : 10.1073/pnas.2215000120 . ПМЦ 9910503 . ПМИД 36574690 .
- ^ Перейти обратно: а б Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и справочник по литературе (3-е изд.). Спрингер. стр. 58 . ISBN 978-1-4020-8238-2 .
1007/978-1-4020-8239-9
- ^ Перейти обратно: а б с Х., Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и справочник по литературе . Нью-Йорк: Спрингер. п. 23. ISBN 9781402082382 . ОСЛК 272311632 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бергер Дж. Д. (октябрь 1986 г.). «Автогамия у Paramecium. Приверженность мейозу на каждой стадии клеточного цикла». Эксп. Сотовый Res . 166 (2): 475–85. дои : 10.1016/0014-4827(86)90492-1 . ПМИД 3743667 .
- ^ Перейти обратно: а б с Райков, И.Б. (1972). «Ядерные явления при конъюгации и автогамии у инфузорий» . Исследования в области протозоологии . 4 : 149.
- ^ Финли, Гарольд Э. «Конъюгация микростомы Vorticella ». Труды Американского микроскопического общества (1943): 97–121.
- ^ «Знакомство с реснитчатыми» . Проверено 16 января 2009 г.
- ^ Перейти обратно: а б с Мотидзуки, Кадзуфуми (2010). «Перестройки ДНК, направляемые некодирующими РНК у инфузорий» . Междисциплинарные обзоры Wiley: РНК . 1 (3): 376–387. дои : 10.1002/wrna.34 . ПМЦ 3746294 . ПМИД 21956937 .
- ^ Ауфдерхайде, Карл Дж. (1986). «Клональное старение Paramecium Tetraurelia . II. Доказательства функциональных изменений макронуклеуса с возрастом». Механизмы старения и развития . 37 (3): 265–279. дои : 10.1016/0047-6374(86)90044-8 . ПМИД 3553762 . S2CID 28320562 .
- ^ Смит-Зоннеборн, Дж. (1979). «Репарация ДНК и обеспечение долголетия Paramecium тетраурелия». Наука . 203 (4385): 1115–1117. Бибкод : 1979Sci...203.1115S . дои : 10.1126/science.424739 . ПМИД 424739 .
- ^ Холмс, Джордж Э.; Холмс, Норрин Р. (июль 1986 г.). «Накопление повреждений ДНК при старении Paramecium Tetraurelia ». Молекулярная и общая генетика . 204 (1): 108–114. дои : 10.1007/bf00330196 . ПМИД 3091993 . S2CID 11992591 .
- ^ Гилли, Дэвид; Блэкберн, Элизабет Х. (1994). «Отсутствие укорочения теломер во время старения у Paramecium » (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 91 (5): 1955–1958. Бибкод : 1994ПНАС...91.1955Г . дои : 10.1073/пнас.91.5.1955 . ПМЦ 43283 . ПМИД 8127914 .
- ^ Ли, К.-В.; и др. (2007). «Ресничные простейшие из докембрийской формации Душантуо, Вэньгань, Южный Китай». Геологическое общество, Лондон, специальные публикации . 286 (1): 151–156. Бибкод : 2007GSLSP.286..151L . дои : 10.1144/SP286.11 . S2CID 129584945 .
- ^ Бомфлер, Бенджамин; Керп, Ганс; Тейлор, Томас Н.; Моэструп, Эйвинд; Тейлор, Эдит Л. (18 декабря 2012 г.). «Кокон триасовой пиявки из Антарктиды содержит ископаемое животное-колокольчик» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (51): 20971–20974. Бибкод : 2012PNAS..10920971B . дои : 10.1073/pnas.1218879109 . ISSN 1091-6490 . ПМК 3529092 . ПМИД 23213234 .
- ^ Перейти обратно: а б ФЕРНАНДЕС, Ноэми М.; Виццони, Винисиус Ф.; БОРХЕС, Барбара ду Н.; СОАРЕС, Карлос АГ; да Силва-Нето, Инасио Д.; Пайва, Тьяго да С. (2018), «Молекулярная филогения и сравнительная морфология указывают на то, что одонтостоматиды (Alveolata, Ciliophora) образуют отдельный таксон классового уровня, родственный Armophorea» , Molecular Phylogenetics and Evolution , 126 : 382–389, doi : 10.1016 /j.ympev.2018.04.026 , ISSN 1055-7903 , PMID 29679715 , S2CID 5032558
- ^ Кавалер-Смит, Т. (2000). Мегаэволюция жгутиковых: основа диверсификации эукариот. В: Ледбитер, BSC, Грин, Дж.К. (ред.). Жгутиконосцы. Единство, многообразие и эволюция . Лондон: Тейлор и Фрэнсис, стр. 361–390, с. 362, [1] .
- ^ «Балантидиаз» . DPDx — Лабораторная идентификация паразитарных заболеваний, вызывающих обеспокоенность общественного здравоохранения . Центры по контролю и профилактике заболеваний. 2013.
- ^ Рамачандран, Амби (23 мая 2003 г.). "Введение" . Паразит: Balantidium coli Болезнь: Балантидиаз . Стэнфордский университет. Архивировано из оригинала 14 апреля 2013 года . Проверено 3 сентября 2018 г.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Шистер, Фредерик Л. и Линн Рамирес-Авила (октябрь 2008 г.). «Текущий мировой статус Balantidium coli » . Обзоры клинической микробиологии . 21 (4): 626–638. дои : 10.1128/CMR.00021-08 . ПМК 2570149 . ПМИД 18854484 .
Дальнейшее чтение
[ редактировать ]- Линн, Денис Х. (2008). Ресничные простейшие: характеристика, классификация и справочник по литературе . Нью-Йорк: Спрингер. ISBN 9781402082382 . ОСЛК 272311632 .
- Хаусманн, Клаус; Брэдбери, Филлис С., ред. (1996). Инфузории: клетки как организмы . Штутгарт: Густав Фишер Верлаг. ISBN 978-3437250361 . OCLC 34782787 .
- Ли, Джон Дж.; Лидейл, Гордон Ф.; Брэдбери, Филлис С., ред. (2000). Иллюстрированный справочник по простейшим: организмы, традиционно называемые простейшими, или недавно открытые группы (2-е изд.). Лоуренс, Канзас: Общество протозоологов. ISBN 9781891276224 . OCLC 49191284 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Ciliophora, на Викискладе?
СМИ, связанные с Ciliophora, на Викискладе?