Генетический код

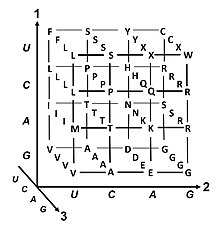
Генетический код — это набор правил, используемых живыми клетками для перевода информации, закодированной в генетическом материале ( последовательности ДНК или РНК, состоящие из триплетов нуклеотидов или кодонов ), в белки . Трансляция осуществляется рибосомой , которая связывает протеиногенные аминокислоты в порядке, заданном информационной РНК (мРНК), используя молекулы транспортной РНК (тРНК) для переноса аминокислот и считывания трех нуклеотидов мРНК за раз. Генетический код всех организмов очень похож и может быть выражен в простой таблице из 64 записей.
Кодоны определяют, какая аминокислота будет добавлена следующей во время биосинтеза белка . За некоторыми исключениями, [1] трехнуклеотидный кодон в последовательности нуклеиновой кислоты определяет одну аминокислоту. Подавляющее большинство генов кодируются по единой схеме (см. таблицу кодонов РНК ). Эту схему часто называют каноническим или стандартным генетическим кодом или просто генетическим кодом, хотя существуют варианты кодов (например, в митохондриях ).
История [ править ]
Попытки понять, как кодируются белки, начались после открытия структуры ДНК в 1953 году. Ключевые первооткрыватели, английский биофизик Фрэнсис Крик и американский биолог Джеймс Уотсон , работавшие вместе в Кавендишской лаборатории Кембриджского университета, выдвинули гипотезу, что информация течет от ДНК и что существует связь между ДНК и белками. [2] Советско-американский физик Джордж Гамов первым предложил работоспособную схему синтеза белка из ДНК. [3] Он постулировал, что для кодирования 20 стандартных аминокислот, используемых живыми клетками для построения белков, необходимо использовать наборы из трех оснований (триплетов), что позволило бы использовать максимум 4 3 = 64 аминокислоты. [4] Он назвал это взаимодействие ДНК и белка (исходный генетический код) «алмазным кодом». [5]
В 1954 году Гамов создал неформальную научную организацию « Клуб связей РНК» , как предложил Уотсон, для учёных разных убеждений, интересовавшихся тем, как белки синтезируются из генов. Однако в клубе могло быть только 20 постоянных членов, представляющих каждую из 20 аминокислот; и еще четыре почетных члена, представляющих четыре нуклеотида ДНК. [6]
Первый научный вклад клуба, позже отмеченный как «одна из самых важных неопубликованных статей в истории науки». [7] и «самая известная неопубликованная статья в анналах молекулярной биологии», [8] был сделан Криком. Крик представил машинописную статью под названием «О вырожденных шаблонах и гипотезе адаптера: заметка для клуба связей РНК». [9] членам клуба в январе 1955 года, что, как вспоминал Уотсон, «полностью изменило наше представление о синтезе белка». [10] Гипотеза утверждает, что триплетный код не передавался аминокислотам, как думал Гамов, а переносился другой молекулой — адаптером, который взаимодействует с аминокислотами. [8] Позже адаптер был идентифицирован как тРНК. [11]
Кодоны [ править ]
Эксперимент Крика , Бреннера, Барнетта и Уоттса-Тобина впервые продемонстрировал, что кодоны состоят из трех оснований ДНК.
Маршалл Ниренберг и Дж. Генрих Маттеи первыми раскрыли природу кодона в 1961 году. [12] Они использовали бесклеточную систему для трансляции полиурациловой последовательности РНК (т.е. UUUUU...) и обнаружили, что синтезированный ими полипептид состоит только из аминокислоты фенилаланина . [13] Таким образом, они пришли к выводу, что кодон UUU соответствует аминокислоте фенилаланину.
За этим последовали эксперименты в Северо Очоа лаборатории , которые продемонстрировали, что полиадениновая последовательность РНК (ААААА...), кодирующая полипептид полилизин . [14] и что последовательность полицитозиновой РНК (CCCCC...) кодирует полипептид полипролин . [15] Следовательно, кодон AAA указывает на аминокислоту лизин , а кодон CCC указывает на аминокислоту пролин . Затем с использованием различных сополимеров было определено большинство оставшихся кодонов.
Последующая работа Хар Гобинда Хораны определила остальную часть генетического кода. Вскоре после этого Роберт Холли определил структуру транспортной РНК (тРНК), молекулы-адаптера, которая облегчает процесс трансляции РНК в белок. Эта работа была основана на более ранних исследованиях Очоа, в результате которых последний получил Нобелевскую премию по физиологии и медицине в 1959 году за работы по энзимологии синтеза РНК. [16]
Продолжая эту работу, Ниренберг и Филип Ледер раскрыли тройную природу кода и расшифровали его кодоны. В этих экспериментах различные комбинации мРНК пропускались через фильтр, содержащий рибосомы — компоненты клеток, которые переводят РНК в белок. Уникальные триплеты способствуют связыванию специфических тРНК с рибосомой. Ледер и Ниренберг в своих экспериментах смогли определить последовательности 54 из 64 кодонов. [17] Корана, Холли и Ниренберг получили за свою работу Нобелевскую премию (1968 г.). [18]
Три стоп-кодона были названы первооткрывателями Ричардом Эпштейном и Чарльзом Стейнбергом. «Янтарь» был назван в честь их друга Харриса Бернштейна, чья фамилия по-немецки означает «янтарь». [19] Два других стоп-кодона были названы «охра» и «опал», чтобы сохранить тему «названий цветов».
генетические коды (синтетическая биология Расширенные )
В широкой академической аудитории широко принята концепция эволюции генетического кода от исходного и неоднозначного генетического кода к четко определенному («замороженному») коду с репертуаром из 20 (+2) канонических аминокислот. [20] Однако существуют разные мнения, концепции, подходы и идеи, и это лучший способ изменить это экспериментальным путем. Предлагаются даже модели, предсказывающие «точки входа» для вторжения синтетических аминокислот в генетический код. [21]
С 2001 года 40 неприродных аминокислот были добавлены в белки путем создания уникального кодона (перекодирования) и соответствующей пары транспортная РНК:аминоацил-тРНК-синтетаза, чтобы кодировать их с разнообразными физико-химическими и биологическими свойствами для использования в качестве инструмент для изучения структуры и функции белка или для создания новых или улучшенных белков. [22] [23]
Х. Мураками и М. Сисидо расширили некоторые кодоны до четырех и пяти оснований. Стивен А. Беннер сконструировал функциональный 65-й ( in vivo ) кодон. [24]
В 2015 году Н. Будиса , Д. Зёлль и соавт. сообщили о полной замене всех 20 899 остатков триптофана (кодонов UGG) на неприродный тиенопиррол-аланин в генетическом коде бактерии Escherichia coli . [25]
В 2016 году был создан первый стабильный полусинтетический организм. Это была (одноклеточная) бактерия с двумя синтетическими основаниями (называемыми X и Y). Основания пережили деление клеток. [26] [27]
В 2017 году исследователи из Южной Кореи сообщили, что создали мышь с расширенным генетическим кодом, которая может производить белки с неприродными аминокислотами. [28]
В мае 2019 года исследователи сообщили о создании нового штамма Syn61 бактерии Escherichia coli . Этот штамм имеет полностью синтетический геном, который рефакторингован (расширены все перекрытия), перекодирован (полностью исключено использование трех из 64 кодонов) и дополнительно модифицирован для удаления теперь ненужных тРНК и факторов высвобождения. Он полностью жизнеспособен и растет в 1,6 раза медленнее, чем его аналог дикого типа «MDS42». [29] [30]
Особенности [ править ]
Рамка чтения [ править ]
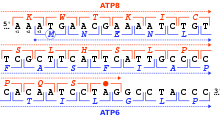
Рамка считывания определяется исходным тройком нуклеотидов, с которого начинается трансляция. Он устанавливает рамку для серии последовательных неперекрывающихся кодонов, которая известна как « открытая рамка считывания » (ORF). Например, строка 5'-AAATGAACG-3' (см. рисунок), если читать ее с первой позиции, содержит кодоны AAA, TGA и ACG; если читать со второй позиции, он содержит кодоны AAT и GAA; и если читать с третьей позиции, он содержит кодоны ATG и AAC. Таким образом, каждую последовательность можно прочитать в направлении 5' → 3' в трех рамках считывания , каждая из которых дает, возможно, отдельную аминокислотную последовательность: в данном примере Lys (K)-Trp (W)-Thr (T), Asn(N)-Glu(E) или Met(M)-Asn(N) соответственно (при трансляции с митохондриальным кодом позвоночных). Когда ДНК двухцепочечная, шесть возможных рамок считывания : три в прямой ориентации на одной цепи и три в обратной ориентации на противоположной цепи. определяются [32] : 330 Рамки, кодирующие белок, определяются стартовым кодоном , обычно первым кодоном AUG (ATG) в последовательности РНК (ДНК).
У эукариот ORF в экзонах часто прерываются интронами .
Стартовые и стоп-кодоны [ править ]
Трансляция начинается с кодона инициации цепи или стартового кодона . Одного стартового кодона недостаточно для начала процесса. близлежащие последовательности, такие как последовательность Шайна-Дальгарно в E. coli и факторы инициации Для начала трансляции также необходимы . Наиболее распространенным стартовым кодоном является AUG, который читается как метионин или формилметионин (у бактерий, митохондрий и пластид). Альтернативные стартовые кодоны в зависимости от организма включают «GUG» или «UUG»; эти кодоны обычно представляют собой валин и лейцин соответственно, но как стартовые кодоны они транслируются как метионин или формилметионин. [33]
Три стоп-кодона имеют названия: UAG — янтарь , UGA — опал (иногда его также называют умброй ) и UAA — охра . Стоп-кодоны также называют «терминирующими» или «нонсенс-кодонами». Они сигнализируют о высвобождении образующегося полипептида из рибосомы, поскольку ни одна родственная тРНК не имеет антикодонов, комплементарных этим стоп-сигналам, что позволяет вместо этого фактору высвобождения связываться с рибосомой. [34]
Эффект мутаций [ править ]

В процессе репликации ДНК иногда возникают ошибки при полимеризации второй цепи. Эти ошибки, мутации организма , могут повлиять на фенотип , особенно если они происходят в кодирующей последовательности белка гена. Частота ошибок обычно составляет 1 ошибку на каждые 10–100 миллионов оснований - из-за способности ДНК-полимеразы «корректировать» . [36] [37]
Миссенс-мутации и нонсенс-мутации являются примерами точковых мутаций , которые могут вызывать генетические заболевания, такие как серповидно-клеточная анемия и талассемия соответственно. [38] [39] [40] Клинически важные миссенс-мутации обычно изменяют свойства кодируемого аминокислотного остатка в основном, кислотном, полярном или неполярном состояниях, тогда как нонсенс-мутации приводят к образованию стоп-кодона . [32]
Мутации, которые нарушают последовательность рамки считывания путем инделирования ( инсерции или делеции ) некратных трех нуклеотидных оснований, известны как мутации сдвига рамки считывания . Эти мутации обычно приводят к трансляции, совершенно отличной от исходной, и, вероятно, приводят стоп-кодона , который усекает белок. к считыванию [41] Эти мутации могут нарушать функцию белка и поэтому редко встречаются в in vivo последовательностях, кодирующих белок . Одна из причин, по которой наследование мутаций сдвига рамки считывания встречается редко, заключается в том, что, если транслируемый белок необходим для роста в условиях селективного давления, с которым сталкивается организм, отсутствие функционального белка может привести к смерти до того, как организм станет жизнеспособным. [42] Мутации сдвига рамки считывания могут привести к тяжелым генетическим заболеваниям, таким как болезнь Тея-Сакса . [43]
Хотя большинство мутаций, изменяющих последовательности белков, вредны или нейтральны, некоторые мутации имеют преимущества. [44] Эти мутации могут позволить мутантному организму противостоять определенным стрессам окружающей среды лучше, чем организмы дикого типа , или быстрее размножаться. В этих случаях мутация будет иметь тенденцию становиться более распространенной в популяции посредством естественного отбора . [45] Вирусы , использующие РНК в качестве генетического материала, имеют высокую скорость мутаций. [46] что может быть преимуществом, поскольку благодаря этому эти вирусы быстро эволюционируют и, таким образом, уклоняются от защитных реакций иммунной системы . [47] В больших популяциях организмов, размножающихся бесполым путем, например, E. coli , могут одновременно возникать множественные полезные мутации. Это явление называется клональной интерференцией и вызывает конкуренцию между мутациями. [48]
Вырождение [ править ]

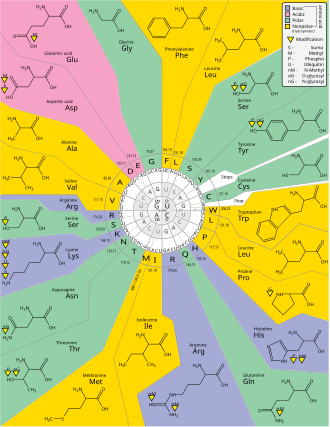
Вырожденность – это избыточность генетического кода. Этот термин дали Бернфилд и Ниренберг. Генетический код имеет избыточность, но не имеет двусмысленности ( в таблицах кодонов полную корреляцию см. ниже). Например, хотя кодоны GAA и GAG определяют глутаминовую кислоту (избыточность), ни один из них не указывает на другую аминокислоту (нет двусмысленности). Кодоны, кодирующие одну аминокислоту, могут различаться в любом из трех положений. Например, аминокислота лейцин обозначается YUR CUN обозначения или кодонами (UUA, UUG, CUU, CUC, CUA или CUG) (разница в первом или третьем положении указывается с использованием IUPAC ), а аминокислота серин определяется кодонами UC N или AG Y (UCA, UCG, UCC, UCU, AGU или AGC) (разница в первой, второй или третьей позиции). [49] Практическим следствием избыточности является то, что ошибки в третьем положении триплетного кодона вызывают только молчащую мутацию или ошибку, которая не влияет на белок, поскольку гидрофильность или гидрофобность поддерживаются за счет эквивалентной замены аминокислот; например, кодон NUN (где N = любой нуклеотид) имеет тенденцию кодировать гидрофобные аминокислоты. NCN дает аминокислотные остатки небольшого размера и умеренной гидропатичности ; NAN кодирует гидрофильные остатки среднего размера. Генетический код настолько хорошо структурирован в отношении гидропатичности, что математический анализ ( разложение по сингулярным значениям ) 12 переменных (4 нуклеотида x 3 позиции) дает замечательную корреляцию (C = 0,95) для прогнозирования гидропатичности кодируемой аминокислоты непосредственно из триплетная нуклеотидная последовательность, без трансляции. [50] [51] Обратите внимание, что в таблице ниже восемь аминокислот вообще не подвергаются мутациям в третьем положении кодона, тогда как на рисунке выше мутация во втором положении, вероятно, вызовет радикальное изменение физико-химических свойств кодона. кодируемая аминокислота.Тем не менее, в глобальном масштабе изменения в первой позиции кодонов важнее, чем изменения во второй позиции. [52] Причина может заключаться в том, что смена заряда (от положительного заряда к отрицательному или наоборот) может произойти только при мутациях в первой позиции определенных кодонов, но не при изменении второй позиции любого кодона. Такое изменение заряда может иметь драматические последствия для структуры или функции белка. Этот аспект, возможно, был в значительной степени недооценен в предыдущих исследованиях. [52]
кодонов Предвзятость использования
Частота кодонов, также известная как ошибка использования кодонов , может варьироваться от вида к виду, что имеет функциональные последствия для контроля трансляции . Кодон варьируется в зависимости от организма; например, наиболее распространенным пролиновым кодоном в E. coli является CCG, тогда как у людей это наименее используемый пролиновый кодон. [53]
Таблица частот кодонов генома человека [54] |
|---|
генетические Альтернативные коды
Нестандартные аминокислоты [ править ]
В некоторых белках стандартные стоп-кодоны заменяются нестандартными аминокислотами в зависимости от связанных с ними сигнальных последовательностей в информационной РНК. Например, UGA может кодировать селеноцистеин , а UAG — пирролизин . Селеноцистеин стал рассматриваться как 21-я аминокислота, а пирролизин — как 22-я. [55] И селеноцистеин, и пирролизин могут присутствовать в одном и том же организме. [55] Хотя генетический код обычно фиксирован в организме, ахейный прокариот Acetohalobium arabaticum может расширять свой генетический код с 20 до 21 аминокислоты (за счет включения пирролизина) в различных условиях роста. [56]
Вариации [ править ]
Первоначально существовал простой и широко распространенный аргумент о том, что генетический код должен быть универсальным, а именно, что любое изменение генетического кода будет смертельным для организма (хотя Крик заявлял, что вирусы являются исключением). Это известно как аргумент «замороженной случайности» в пользу универсальности генетического кода. Однако в своей основополагающей статье о происхождении генетического кода в 1968 году Фрэнсис Крик все же заявил, что универсальность генетического кода во всех организмах является недоказанным предположением и, вероятно, в некоторых случаях не соответствует действительности. Он предсказал, что «код универсален (одинаков для всех организмов) или почти универсален». [58] Первая вариация была обнаружена в 1979 году исследователями, изучающими митохондриальные гены человека . [59] После этого было обнаружено множество небольших вариантов, [60] включая различные альтернативные митохондриальные коды. [61] Эти минорные варианты, например, включают трансляцию кодона UGA как триптофана у видов Mycoplasma и трансляцию CUG как серина, а не лейцина у дрожжей «клады CTG» (таких как Candida albicans ). [62] [63] [64] Поскольку вирусы должны использовать тот же генетический код, что и их хозяева, модификации стандартного генетического кода могут помешать синтезу или функционированию вирусного белка. Однако такие вирусы, как тотивирусы, адаптировались к модификации генетического кода хозяина. [65] У бактерий и архей общими стартовыми кодонами являются GUG и UUG. В редких случаях некоторые белки могут использовать альтернативные стартовые кодоны. [60] Удивительно, но вариации в интерпретации генетического кода существуют также в генах человека, кодируемых ядерным кодом: в 2016 году исследователи, изучающие трансляцию малатдегидрогеназы, обнаружили, что примерно в 4% мРНК, кодирующих этот фермент, стоп-кодон естественным образом используется для кодирования аминокислоты триптофан и аргинин. [66] Этот тип перекодирования индуцируется контекстом стоп-кодона с высокой читаемостью. [67] и это называется функциональным трансляционным чтением . [68]
Несмотря на эти различия, все известные природные коды очень похожи. Механизм кодирования одинаков для всех организмов: трехосновные кодоны, тРНК , рибосомы, однонаправленное чтение и трансляция отдельных кодонов в отдельные аминокислоты. [69] Самые крайние вариации встречаются у некоторых инфузорий, где значение стоп-кодонов зависит от их положения в мРНК. Находясь близко к 3'-концу, они действуют как терминаторы, тогда как во внутренних положениях они либо кодируют аминокислоты, как у Condylostoma magnum. [70] или вызвать сдвиг рамки рибосомы , как в Euplotes . [71]
Происхождение и вариации генетического кода, включая механизмы, лежащие в основе эволюции генетического кода, широко изучены. [72] [73] и были проведены некоторые исследования по экспериментальному развитию генетического кода некоторых организмов. [74] [75] [76] [77]
Вывод [ править ]
Варианты генетических кодов, используемых организмом, можно сделать путем идентификации высококонсервативных генов, закодированных в этом геноме, и сравнения использования его кодонов с аминокислотами в гомологичных белках других организмов. Например, программа FACIL выводит генетический код, определяя, какие аминокислоты в гомологичных белковых доменах чаще всего совпадают с каждым кодоном. Полученные вероятности аминокислот (или стоп-кодонов) для каждого кодона отображаются в логотипе генетического кода. [57]
По состоянию на январь 2022 года наиболее полное исследование генетических кодов провели Шульгина и Эдди, которые проверили 250 000 геномов прокариот с помощью своего инструмента Codetta. Этот инструмент использует подход, аналогичный FACIL, с более крупной базой данных Pfam . Несмотря на то, что NCBI уже предоставил 27 таблиц перевода, авторы смогли найти 5 новых вариантов генетического кода (подтвержденных мутациями тРНК) и исправить несколько неправильных атрибуций. [78] Кодетта позже была использована для анализа изменений генетического кода инфузорий . [79]
Происхождение [ править ]
Генетический код — ключевая часть истории жизни , согласно одной из версий, самовоспроизводящиеся молекулы РНК предшествовали жизни, какой мы ее знаем. Это гипотеза мира РНК . Согласно этой гипотезе, любая модель возникновения генетического кода тесно связана с моделью перехода от рибозимов (ферментов РНК) к белкам как основным ферментам в клетках. В соответствии с гипотезой мира РНК, молекулы транспортной РНК, по-видимому, возникли раньше современных аминоацил-тРНК-синтетаз , поэтому последняя не может быть частью объяснения ее закономерностей. [80]
Гипотетический случайно возникший генетический код дополнительно мотивирует биохимическую или эволюционную модель его происхождения. Если бы аминокислоты были случайным образом распределены по триплетным кодонам, их было бы 1,5 × 10. 84 возможные генетические коды. [81] : 163 Это число определяется путем расчета количества способов, которыми 21 элемент (20 аминокислот плюс одна остановка) может быть помещен в 64 ячейки, при этом каждый элемент используется хотя бы один раз. [82] Однако распределение назначений кодонов в генетическом коде неслучайно. [83] В частности, генетический код объединяет определенные назначения аминокислот.
Аминокислоты, которые имеют один и тот же путь биосинтеза, как правило, имеют одинаковое первое основание в своих кодонах. Это может быть эволюционным реликтом раннего, более простого генетического кода с меньшим количеством аминокислот, который позже развился, чтобы кодировать больший набор аминокислот. [84] Это также могло отражать стерические и химические свойства, которые оказали другое влияние на кодон в ходе его эволюции. Аминокислоты со схожими физическими свойствами также имеют тенденцию иметь схожие кодоны. [85] [86] уменьшение проблем, вызванных точечными мутациями и неправильным переводом. [83]
Учитывая неслучайную схему кодирования генетического триплета, обоснованная гипотеза происхождения генетического кода могла бы учитывать множество аспектов таблицы кодонов, таких как отсутствие кодонов для D-аминокислот, паттерны вторичных кодонов для некоторых аминокислот, ограничение синонимичных позиции на третье положение, небольшой набор всего из 20 аминокислот (вместо числа, приближающегося к 64) и связь паттернов стоп-кодонов с паттернами кодирования аминокислот. [87]
Три основные гипотезы касаются происхождения генетического кода. Многие модели относятся к одному из них или к гибриду: [88]
- Случайное замораживание: генетический код был создан случайным образом. Например, ранние тРНК -подобные рибозимы могли иметь различное сродство к аминокислотам, при этом кодоны, возникающие из другой части рибозима, демонстрировали случайную изменчивость. Как только было бы закодировано достаточное количество пептидов , любое серьезное случайное изменение в генетическом коде стало бы смертельным; поэтому он стал «замороженным». [89]
- Стереохимическое сродство: генетический код является результатом высокого сродства между каждой аминокислотой и ее кодоном или антикодоном; последний вариант подразумевает, что молекулы пре-тРНК по этому сродству соответствуют соответствующим аминокислотам. Позже в ходе эволюции это совпадение постепенно было заменено совпадением по аминоацил-тРНК-синтетазам. [87] [90] [91]
- Оптимальность: генетический код продолжал развиваться после своего первоначального создания, так что текущий код максимизирует некоторую функцию приспособленности , обычно какую-то минимизацию ошибок. [87] [88] [92]
Гипотезы рассматривают различные сценарии: [93]
- Химические принципы регулируют специфическое взаимодействие РНК с аминокислотами. Эксперименты с аптамерами показали, что некоторые аминокислоты обладают избирательным химическим сродством к своим кодонам. [94] Эксперименты показали, что из 8 протестированных аминокислот 6 демонстрируют некоторую ассоциацию триплета РНК с аминокислотами. [81] [91]
- Биосинтетическая экспансия. Генетический код вырос из более простого более раннего кода в процессе «биосинтетического расширения». Первобытная жизнь «открыла» новые аминокислоты (например, как побочные продукты метаболизма ) и позже включила некоторые из них в механизм генетического кодирования. [95] Хотя было обнаружено много косвенных доказательств того, что в прошлом использовалось меньше типов аминокислот, [96] Точные и подробные гипотезы о том, какие аминокислоты в каком порядке вошли в код, являются спорными. [97] [98] Однако несколько исследований показали, что Gly, Ala, Asp, Val, Ser, Pro, Glu, Leu, Thr могут принадлежать к группе аминокислот раннего присоединения, тогда как Cys, Met, Tyr, Trp, His, Phe могут принадлежать к группе аминокислот раннего присоединения. к группе аминокислот позднего добавления. [99] [100] [101] [102]
- Естественный отбор привел к присвоению кодонов генетического кода, которые минимизируют последствия мутаций . [103] Недавняя гипотеза [104] предполагает, что триплетный код произошел от кодов, в которых использовались более длинные, чем триплетные, кодоны (например, квадруплетные кодоны). Более продолжительное декодирование, чем триплетное, увеличит избыточность кодонов и будет более устойчивым к ошибкам. Эта особенность могла бы обеспечить точное декодирование в отсутствие сложных механизмов трансляции, таких как рибосома , например, до того, как клетки начали производить рибосомы.
- Информационные каналы. Теоретико-информационные подходы моделируют процесс трансляции генетического кода в соответствующие аминокислоты как подверженный ошибкам информационный канал. [105] Собственный шум (то есть ошибка) в канале ставит перед организмом фундаментальный вопрос: как можно сконструировать генетический код, способный противостоять шуму? [106] при этом точно и эффективно переводя информацию? Эти «скоростного искажения» модели [107] предполагают, что генетический код возник в результате взаимодействия трех конфликтующих эволюционных сил: потребности в разнообразных аминокислотах, [108] для устойчивости к ошибкам [103] и при минимальных затратах ресурсов. Код возникает при переходе, когда отображение кодонов на аминокислоты становится неслучайным. Появление кода определяется топологией , определяемой вероятными ошибками, и связано с проблемой раскраски карты . [109]
- Теория игр: модели, основанные на сигнальных играх, сочетают в себе элементы теории игр, естественного отбора и информационных каналов. Такие модели использовались, чтобы предположить, что первые полипептиды, вероятно, были короткими и выполняли неферментативную функцию. Теоретико-игровые модели предположили, что организация цепочек РНК в клетках могла быть необходима для предотвращения «обманчивого» использования генетического кода, то есть предотвращения подавления мира РНК древним эквивалентом вирусов. [110]
- Стоп-кодоны. Кодоны для остановки трансляции также представляют собой интересный аспект проблемы происхождения генетического кода. В качестве примера решения проблемы эволюции стоп-кодонов было высказано предположение, что стоп-кодоны таковы, что они с наибольшей вероятностью рано прекратят трансляцию в случае ошибки сдвига кадра . [111] Напротив, некоторые стереохимические молекулярные модели объясняют происхождение стоп-кодонов как «неопределяемые». [87]
См. также [ править ]
Ссылки [ править ]
- ^ Туранов А.А., Лобанов А.В., Фоменко Д.Е., Моррисон Х.Г., Согин М.Л., Клобутчер Л.А., Хэтфилд Д.Л., Гладышев В.Н. (январь 2009 г.). «Генетический код поддерживает целевую вставку двух аминокислот с помощью одного кодона» . Наука . 323 (5911): 259–61. дои : 10.1126/science.1164748 . ПМК 3088105 . ПМИД 19131629 .
- ^ Уотсон, доктор медицинских наук; Крик, FH (30 мая 1953 г.). «Генетическое значение структуры дезоксирибонуклеиновой кислоты» . Природа . 171 (4361): 964–967. Бибкод : 1953Natur.171..964W . дои : 10.1038/171964b0 . ISSN 0028-0836 . ПМИД 13063483 . S2CID 4256010 .
- ^ Стегманн, Ульрих Э. (1 сентября 2016 г.). « Переосмысление« генетического кодирования »: анализ фактического использования» . Британский журнал философии науки . 67 (3): 707–730. дои : 10.1093/bjps/axv007 . ISSN 0007-0882 . ПМЦ 4990703 . ПМИД 27924115 .
- ^ Крик, Фрэнсис (10 июля 1990 г.). «Глава 8: Генетический код» . Что за безумная погоня: личный взгляд на научные открытия . Основные книги. стр. 89–101. ISBN 9780465091386 . OCLC 1020240407 . [ постоянная мертвая ссылка ]
- ^ Хейс, Брайан (1998). «Информатика: изобретение генетического кода» . Американский учёный . 86 (1): 8–14. дои : 10.1511/1998.17.3338 . ISSN 0003-0996 . JSTOR 27856930 . S2CID 121907709 .
- ^ Штраус, Бернард С. (1 марта 2019 г.). «Мартинас Йчас: «Архивариус» Галстук-клуба РНК» . Генетика . 211 (3): 789–795. дои : 10.1534/genetics.118.301754 . ISSN 1943-2631 . ПМК 6404253 . ПМИД 30846543 .
- ^ «Фрэнсис Крик — Профили в результатах научного поиска» . Profiles.nlm.nih.gov . Проверено 21 июля 2022 г.
- ^ Jump up to: Перейти обратно: а б Фрай, Майкл (2022). «Гипотеза об адаптере Крика и открытие транспортной РНК: эксперимент, превосходящий теоретические предсказания» . Философия, теория и практика биологии . 14 . дои : 10.3998/ptpbio.2628 . ISSN 2475-3025 . S2CID 249112573 .
- ^ Крик, Фрэнсис (1955). «О вырожденных шаблонах и гипотезе адаптера: заметка для Клуба связей РНК» . Национальная медицинская библиотека . Проверено 21 июля 2022 г.
- ^ Уотсон, Джеймс Д. (2007). Избегайте скучных людей: уроки из жизни в науке . Издательство Оксфордского университета. п. 112. ИСБН 978-0-19-280273-6 . OCLC 47716375 .
- ^ Барчишевская, Мирослава З.; Перриг, Патрик М.; Барчишевский, Ян (2016). «тРНК — золотой стандарт молекулярной биологии» . Молекулярные биосистемы . 12 (1): 12–17. дои : 10.1039/c5mb00557d . ПМИД 26549858 .
- ^ Янофски, Чарльз (9 марта 2007 г.). «Установление тройной природы генетического кода» . Клетка . 128 (5): 815–818. дои : 10.1016/j.cell.2007.02.029 . ПМИД 17350564 . S2CID 14249277 .
- ^ Ниренберг М.В., Маттеи Дж.Х. (октябрь 1961 г.). «Зависимость бесклеточного синтеза белка в E. coli от встречающихся в природе или синтетических полирибонуклеотидов» . Труды Национальной академии наук Соединенных Штатов Америки . 47 (10): 1588–602. Бибкод : 1961PNAS...47.1588N . дои : 10.1073/pnas.47.10.1588 . ПМК 223178 . ПМИД 14479932 .
- ^ Гарднер Р.С., Вахба А.Дж., Базилио С., Миллер Р.С., Лендьел П., Спейер Дж.Ф. (декабрь 1962 г.). «Синтетические полинуклеотиды и код аминокислоты. VII» . Труды Национальной академии наук Соединенных Штатов Америки . 48 (12): 2087–94. Бибкод : 1962PNAS...48.2087G . дои : 10.1073/pnas.48.12.2087 . ПМК 221128 . ПМИД 13946552 .
- ^ Вахба А.Дж., Гарднер Р.С., Базилио С., Миллер Р.С., Спейер Дж.Ф., Лендьел П. (январь 1963 г.). «Синтетические полинуклеотиды и код аминокислоты. VIII» . Труды Национальной академии наук Соединенных Штатов Америки . 49 (1): 116–22. Бибкод : 1963PNAS...49..116W . дои : 10.1073/pnas.49.1.116 . ПМК 300638 . ПМИД 13998282 .
- ^ «Нобелевская премия по физиологии и медицине 1959 года» (пресс-релиз). Шведская королевская академия наук. 1959 год . Проверено 27 февраля 2010 г.
Нобелевская премия по физиологии и медицине 1959 года была присуждена совместно Северо Очоа и Артуру Корнбергу «за открытие механизмов биологического синтеза рибонуклеиновой кислоты и дезоксирибонуклеиновой кислоты».
- ^ Ниренберг М., Ледер П., Бернфилд М., Бримакомб Р., Трупин Дж., Роттман Ф., О'Нил С. (май 1965 г.). «Кодовые слова РНК и синтез белка, VII. Об общей природе кода РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 53 (5): 1161–8. Бибкод : 1965ПНАС...53.1161Н . дои : 10.1073/pnas.53.5.1161 . ПМК 301388 . ПМИД 5330357 .
- ^ «Нобелевская премия по физиологии и медицине 1968 года» (пресс-релиз). Шведская королевская академия наук. 1968 год . Проверено 27 февраля 2010 г.
Нобелевская премия по физиологии и медицине 1968 года была присуждена совместно Роберту Холли, Хару Гобинду Хоране и Маршаллу Ниренбергу «за интерпретацию генетического кода и его функции в синтезе белка».
- ^ Эдгар Б. (октябрь 2004 г.). «Геном бактериофага Т4: археологические раскопки» . Генетика . 168 (2): 575–82. дои : 10.1093/генетика/168.2.575 . ПМЦ 1448817 . ПМИД 15514035 .
- ^ Будиса, Недилько (23 декабря 2005 г.). Книга в онлайн-библиотеке Wiley . дои : 10.1002/3527607188 . ISBN 9783527312436 .
- ^ Кубышкин В.; Будиса, Н. (2017). «Синтетическое отчуждение микробных организмов с помощью инженерии генетического кода: почему и как?». Биотехнологический журнал . 12 (8): 1600097. doi : 10.1002/biot.201600097 . ПМИД 28671771 .
- ^ Се Дж., Шульц П.Г. (декабрь 2005 г.). «Добавление аминокислот в генетический репертуар». Современное мнение в области химической биологии . 9 (6): 548–54. дои : 10.1016/j.cbpa.2005.10.011 . ПМИД 16260173 .
- ^ Ван Ц, Пэрриш А.Р., Ван Л. (март 2009 г.). «Расширение генетического кода для биологических исследований» . Химия и биология . 16 (3): 323–36. doi : 10.1016/j.chembiol.2009.03.001 . ПМК 2696486 . ПМИД 19318213 .
- ^ Саймон М. (7 января 2005 г.). Новые вычисления: акцент на биоинформатике . Springer Science & Business Media. стр. 105–106. ISBN 978-0-387-22046-8 .
- ^ Хёсл, МГ; Оем, С.; Дуркин, П.; Дармон, Э.; Пейл, Л.; Аэрни, Х.-Р.; Раппсилбер, Дж .; Райнхарт, Дж.; Лич, Д.; Зёлль, Д.; Будиса, Н. (2015). «Химическая эволюция бактериального протеома» . Angewandte Chemie, международное издание . 54 (34): 10030–10034. дои : 10.1002/anie.201502868 . ПМЦ 4782924 . ПМИД 26136259 . НИХМСИД: NIHMS711205
- ^ «Создан первый стабильный полусинтетический организм | KurzweilAI» . www.kurzweilai.net . 3 февраля 2017 года . Проверено 9 февраля 2017 г.
- ^ Чжан Ю, Лэмб Б.М., Фельдман А.В., Чжоу А.С., Лавернь Т., Ли Л., Ромесберг Ф.Е. (февраль 2017 г.). «Полусинтетический организм, созданный для стабильного расширения генетического алфавита» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (6): 1317–1322. Бибкод : 2017PNAS..114.1317Z . дои : 10.1073/pnas.1616443114 . ПМК 5307467 . ПМИД 28115716 .
- ^ Хан С., Ян А., Ли С., Ли Х.В., Пак CB, Пак HS (февраль 2017 г.). «Расширение генетического кода Mus musculus» . Природные коммуникации . 8 : 14568. Бибкод : 2017NatCo...814568H . дои : 10.1038/ncomms14568 . ПМК 5321798 . ПМИД 28220771 .
- ^ Циммер, Карл (15 мая 2019 г.). «Ученые создали бактерии с синтетическим геномом. Является ли это искусственной жизнью? Важным событием для синтетической биологии является то, что колонии кишечной палочки процветают благодаря ДНК, созданной с нуля людьми, а не природой» . Нью-Йорк Таймс . Архивировано из оригинала 2 января 2022 года . Проверено 16 мая 2019 г.
- ^ Фреденс, Юлиус; и др. (15 мая 2019 г.). «Тотальный синтез Escherichia coli с перекодированным геномом» . Природа . 569 (7757): 514–518. Бибкод : 2019Natur.569..514F . дои : 10.1038/s41586-019-1192-5 . ПМК 7039709 . ПМИД 31092918 . S2CID 205571025 .
- ^ Митохондрии Homo sapiens , полный геном. «Пересмотренная Кембриджская эталонная последовательность (rCRS): доступ NC_012920» , Национальный центр биотехнологической информации . Проверено 27 декабря 2017 г.
- ^ Jump up to: Перейти обратно: а б Кинг Р.С., Маллиган П., Стэнсфилд В. (10 января 2013 г.). Генетический словарь . ОУП США. п. 608. ИСБН 978-0-19-976644-4 .
- ^ Туриоль С., Борнс С., Боннал С., Одижье С., Пратс Х., Пратс А.С., Вагнер С. (2003). «Поколение разнообразия изоформ белка путем альтернативной инициации трансляции кодонов, отличных от AUG» . Биология клетки . 95 (3–4): 169–78. дои : 10.1016/S0248-4900(03)00033-9 . ПМИД 12867081 .
- ^ Малой С (29 ноября 2003 г.). «Как бессмысленные мутации получили свои названия» . Курс микробной генетики . Государственный университет Сан-Диего . Проверено 10 марта 2010 г.
- ^ Ссылки на изображение можно найти на странице Wikimedia Commons по адресу: Commons:File:Notablemutations.svg#References .
- ^ Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К. и др., ред. (2000). «Спонтанные мутации» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Фрейзингер Э., Гроллман А.П., Миллер Х., Кискер С. (апрель 2004 г.). «Толерантность (не) к повреждениям позволяет лучше понять точность репликации ДНК» . Журнал ЭМБО . 23 (7): 1494–505. дои : 10.1038/sj.emboj.7600158 . ПМК 391067 . ПМИД 15057282 .
- ^ Бойле, С; Ванде Вельде, центральный; Кливленд, Д.В. (2006). «АЛС: заболевание двигательных нейронов и их ненейрональных соседей» . Нейрон . 52 (1): 39–59. дои : 10.1016/j.neuron.2006.09.018 . ПМИД 17015226 .
- ^ Чанг Дж.К., Кан Ю.В. (июнь 1979 г.). «бета-0-талассемия, бессмысленная мутация у человека» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (6): 2886–9. Бибкод : 1979PNAS...76.2886C . дои : 10.1073/pnas.76.6.2886 . ПМЦ 383714 . ПМИД 88735 .
- ^ Бойле С., Ванде Вельде С., Кливленд Д.В. (октябрь 2006 г.). «АЛС: заболевание двигательных нейронов и их ненейрональных соседей» . Нейрон . 52 (1): 39–59. дои : 10.1016/j.neuron.2006.09.018 . ПМИД 17015226 .
- ^ Исбрандт Д., Хопвуд Дж. Дж., фон Фигура К., Петерс С. (1996). «Две новые мутации сдвига рамки считывания, вызывающие преждевременное появление стоп-кодонов у пациента с тяжелой формой синдрома Марото-Лами» . Человеческая мутация . 7 (4): 361–3. doi : 10.1002/(SICI)1098-1004(1996)7:4<361::AID-HUMU12>3.0.CO;2-0 . ПМИД 8723688 . S2CID 22693748 .
- ^ Ворона Дж. Ф. (1993). «Что мы знаем о частоте спонтанных мутаций человека?». Экологический и молекулярный мутагенез . 21 (2): 122–9. Бибкод : 1993EnvMM..21..122C . дои : 10.1002/em.2850210205 . ПМИД 8444142 . S2CID 32918971 .
- ^ Льюис Р. (2005). Генетика человека: концепции и приложения (6-е изд.). Бостон, Массачусетс: МакГроу Хилл. стр. 227–228. ISBN 978-0-07-111156-0 .
- ^ Сойер С.А., Парш Дж., Чжан З., Хартл Д.Л. (апрель 2007 г.). «Распространенность положительного отбора среди замен почти нейтральных аминокислот у дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (16): 6504–10. Бибкод : 2007PNAS..104.6504S . дои : 10.1073/pnas.0701572104 . ПМЦ 1871816 . ПМИД 17409186 .
- ^ Мосты КР (2002). «Малярия и красная клетка» . Гарвард . Архивировано из оригинала 27 ноября 2011 года.
- ^ Дрейк Дж.В., Холланд Дж.Дж. (ноябрь 1999 г.). «Скорость мутаций среди РНК-вирусов» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (24): 13910–3. Бибкод : 1999PNAS...9613910D . дои : 10.1073/pnas.96.24.13910 . ПМК 24164 . ПМИД 10570172 .
- ^ Холланд Дж., Шпиндлер К., Городиски Ф., Грабау Э., Никол С., ВандеПол С. (март 1982 г.). «Быстрая эволюция геномов РНК». Наука . 215 (4540): 1577–85. Бибкод : 1982Sci...215.1577H . дои : 10.1126/science.7041255 . ПМИД 7041255 .
- ^ де Виссер Дж. А., Розен Д. Е. (апрель 2006 г.). «Клональное вмешательство и периодический отбор новых полезных мутаций в Escherichia coli» . Генетика . 172 (4): 2093–100. doi : 10.1534/genetics.105.052373 . ПМЦ 1456385 . ПМИД 16489229 .
- ^ Уотсон, Джеймс Д. (2008). Молекулярная биология гена . Пирсон/Бенджамин Каммингс. ISBN 978-0-8053-9592-1 . : 102–117 : 521–522
- ^ Мишель-Байерле, Мария Элизабет (1990). Реакционные центры фотосинтезирующих бактерий: Feldafing-II-Meeting . Спрингер-Верлаг. ISBN 978-3-540-53420-4 .
- ^ Фюллен Г., Юван, округ Колумбия (1994). «Генетические алгоритмы и рекурсивный ансамблевый мутагенез в белковой инженерии». Комплексность Интернэшнл 1.
- ^ Jump up to: Перейти обратно: а б Фрике, Маркус (2019). «Глобальное значение вторичных структур РНК в последовательностях, кодирующих белки» . Биоинформатика . 35 (4): 579–583. doi : 10.1093/биоинформатика/bty678 . ПМЦ 7109657 . ПМИД 30101307 . S2CID 51968530 .
- ^ «Таблица частот использования кодонов (диаграмма) - Genscript» . www.genscript.com . Проверено 4 февраля 2022 г.
- ^ «Таблица использования кодонов» . www.kazusa.or.jp .
- ^ Jump up to: Перейти обратно: а б Чжан Ю., Баранов П.В., Аткинс Дж.Ф., Гладышев В.Н. (май 2005 г.). «Пирролизин и селеноцистеин используют разные стратегии декодирования» . Журнал биологической химии . 280 (21): 20740–51. дои : 10.1074/jbc.M501458200 . ПМИД 15788401 .
- ^ Прат Л., Хайнеманн И.Ю., Эрни Х.Р., Райнхарт Дж., О'Донохью П., Сёлль Д. (декабрь 2012 г.). «Расширение генетического кода бактерий в зависимости от источника углерода» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (51): 21070–5. Бибкод : 2012PNAS..10921070P . дои : 10.1073/pnas.1218613110 . ПМК 3529041 . ПМИД 23185002 .
- ^ Jump up to: Перейти обратно: а б Дутил Б.Е., Юргеленайте Р., Шкларчик Р., ван Хиюм С.А., Харханги Х.Р., Шмид М., де Вильд Б., Франсойс К.Дж., Стунненберг Х.Г., Строус М., Джеттен М.С., Оп ден Кэмп Х.Дж., Хюйнен М.А. (июль 2011 г.). «FACIL: быстрый и точный вывод генетического кода и логотип» . Биоинформатика . 27 (14): 1929–33. doi : 10.1093/биоинформатика/btr316 . ПМК 3129529 . ПМИД 21653513 .
- ^ Фрэнсис Крик, 1968. «Происхождение генетического кода». Дж. Мол. Биол.
- ^ Баррелл Б.Г., Банкир А.Т., Друэн Дж. (1979). «Другой генетический код в митохондриях человека». Природа . 282 (5735): 189–194. Бибкод : 1979Natur.282..189B . дои : 10.1038/282189a0 . ПМИД 226894 . S2CID 4335828 . ( [1] )
- ^ Jump up to: Перейти обратно: а б Эльжановский А., Остелл Дж. (7 апреля 2008 г.). «Генетические коды» . Национальный центр биотехнологической информации (NCBI) . Проверено 10 марта 2010 г.
- ^ Джукс Т.Х., Осава С. (декабрь 1990 г.). «Генетический код в митохондриях и хлоропластах». Эксперименты . 46 (11–12): 1117–26. дои : 10.1007/BF01936921 . ПМИД 2253709 . S2CID 19264964 .
- ^ Фитцпатрик Д.А., Лог М.Э., Стаджич Дж.Е., Батлер Дж. (1 января 2006 г.). «Филогения грибов, основанная на 42 полных геномах, полученных в результате супердерева и комбинированного анализа генов» . Эволюционная биология BMC . 6:99 . дои : 10.1186/1471-2148-6-99 . ПМК 1679813 . ПМИД 17121679 .
- ^ Сантос М.А., Туите М.Ф. (май 1995 г.). «Кодон CUG у Candida albicans декодируется in vivo как серин, а не лейцин» . Исследования нуклеиновых кислот . 23 (9): 1481–6. дои : 10.1093/нар/23.9.1481 . ПМК 306886 . ПМИД 7784200 .
- ^ Батлер Г., Расмуссен М.Д., Лин М.Ф. и др. (июнь 2009 г.). «Эволюция патогенности и полового размножения в восьми геномах Candida» . Природа . 459 (7247): 657–62. Бибкод : 2009Natur.459..657B . дои : 10.1038/nature08064 . ПМЦ 2834264 . ПМИД 19465905 .
- ^ Тейлор DJ, Баллинджер М.Дж., Боуман С.М., Брюэнн Дж.А. (2013). «Коэволюция вируса-хозяина в условиях модифицированного ядерного генетического кода» . ПерДж . 1 : е50. дои : 10.7717/peerj.50 . ПМЦ 3628385 . ПМИД 23638388 .
- ^ Хофхейс Дж., Шуерен Ф., Нётцель С., Лингнер Т., Гертнер Дж., Ян О., Томс С. (2016). «Функциональное расширение малатдегидрогеназы выявляет модификацию генетического кода» . Открытая Биол . 6 (11): 160246. doi : 10.1098/rsob.160246 . ПМК 5133446 . ПМИД 27881739 .
- ^ Шурен Ф., Лингнер Т., Джордж Р., Хофхейс Дж., Гартнер Дж., Томс С. (2014). «Пероксисомальная лактатдегидрогеназа генерируется в результате трансляционного чтения у млекопитающих» . электронная жизнь . 3 : e03640. doi : 10.7554/eLife.03640 . ПМЦ 4359377 . ПМИД 25247702 .
- ^ Ф. Шурен и С. Томс (2016). «Функционально-трансляционное прочтение: взгляд на системную биологию» . ПЛОС Генетика . 12 (8): e1006196. дои : 10.1371/journal.pgen.1006196 . ПМЦ 4973966 . ПМИД 27490485 .
- ^ Кубышкин В., Асеведо-Роча К.Г., Будиса Н. (февраль 2018 г.). «Об универсальных кодирующих событиях в биогенезе белков» . Биосистемы . 164 : 16–25. Бибкод : 2018BiSys.164...16K . doi : 10.1016/j.biosystems.2017.10.004 . ПМИД 29030023 .
- ^ Хифи С.М., Мариотти М., Гладышев В.Н., Аткинс Дж.Ф., Баранов П.В. (ноябрь 2016 г.). «Новые варианты генетического кода инфузорий, включая переназначение всех трех стоп-кодонов на смысловые кодоны в Condylostoma magnum » . Молекулярная биология и эволюция . 33 (11): 2885–2889. дои : 10.1093/molbev/msw166 . ПМК 5062323 . ПМИД 27501944 .
- ^ Лобанов А.В., Хифи С.М., Туранов А.А., Геращенко М.В., Пуччарелли С., Деварадж Р.Р. и др. (январь 2017 г.). «Позиционно-зависимое завершение и широко распространенный обязательный сдвиг рамки в Euplotes переводе » . Структурная и молекулярная биология природы . 24 (1): 61–68. дои : 10.1038/nsmb.3330 . ПМЦ 5295771 . ПМИД 27870834 .
- ^ Кунин Е.В., Новожилов А.С. (февраль 2009 г.). «Происхождение и эволюция генетического кода: универсальная загадка» . ИУБМБ Жизнь . 61 (2): 91–111. дои : 10.1002/iub.146 . ПМЦ 3293468 . ПМИД 19117371 .
- ^ Сенгупта С., Хиггс П.Г. (июнь 2015 г.). «Пути эволюции генетического кода в древних и современных организмах». Журнал молекулярной эволюции . 80 (5–6): 229–243. Бибкод : 2015JMolE..80..229S . дои : 10.1007/s00239-015-9686-8 . ПМИД 26054480 . S2CID 15542587 .
- ^ Се Дж., Шульц П.Г. (август 2006 г.). «Химический набор инструментов для белков - расширенный генетический код». Nature Reviews Молекулярно-клеточная биология . 7 (10): 775–782. дои : 10.1038/nrm2005 . ПМИД 16926858 . S2CID 19385756 .
- ^ Нойманн Х., Ван К., Дэвис Л., Гарсиа-Алай М., Чин Дж.В. (март 2010 г.). «Кодирование нескольких неприродных аминокислот посредством эволюции рибосомы, декодирующей квадруплет». Природа . 18 (464): 441–444. дои : 10.1038/nrm2005 . ПМИД 16926858 . S2CID 19385756 .
- ^ Лю CC, Шульц П.Г. (2010). «Добавление новых химических элементов в генетический код». Ежегодный обзор биохимии . 79 : 413–444. doi : 10.1146/annurev.biochem.052308.105824 . ПМИД 20307192 .
- ^ Чин Дж.В. (февраль 2014 г.). «Расширение и перепрограммирование генетического кода клеток и животных». Ежегодный обзор биохимии . 83 : 379–408. doi : 10.1146/annurev-biochem-060713-035737 . ПМИД 24555827 .
- ^ Шульгина Ю.; Эдди, СР (9 ноября 2021 г.). «Вычислительный экран для поиска альтернативных генетических кодов в более чем 250 000 геномах» . электронная жизнь . 10 . дои : 10.7554/eLife.71402 . ПМЦ 8629427 . ПМИД 34751130 .
- ^ Чен, В; Гэн, Ю; Чжан, Б; Ян, Ю; Чжао, Ф; Мяо, М. (4 апреля 2023 г.). «Стоп или нет: полногеномное профилирование переназначенных стоп-кодонов у инфузорий» . Молекулярная биология и эволюция . 40 (4). дои : 10.1093/molbev/msad064 . ПМК 1008964 . ПМИД 36952281 .
- ^ Рибас де Пуплана Л., Тернер Р.Дж., Стир Б.А., Шиммель П. (сентябрь 1998 г.). «Происхождение генетического кода: тРНК старше их синтетаз?» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (19): 11295–300. Бибкод : 1998PNAS...9511295D . дои : 10.1073/pnas.95.19.11295 . ПМК 21636 . ПМИД 9736730 .
- ^ Jump up to: Перейти обратно: а б Ярус, Майкл (2010). Жизнь из мира РНК: Внутренний предок . Издательство Гарвардского университета. ISBN 978-0-674-05075-4 .
- ^ «Функция Mathematica для # возможного расположения элементов в корзинах? – Онлайн-группы технических дискуссий — Сообщество Wolfram» . Community.wolfram.com . Проверено 3 февраля 2017 г.
- ^ Jump up to: Перейти обратно: а б Фриланд С.Дж., Херст Л.Д. (сентябрь 1998 г.). «Генетический код – один на миллион». Журнал молекулярной эволюции . 47 (3): 238–48. Бибкод : 1998JMolE..47..238F . дои : 10.1007/PL00006381 . ПМИД 9732450 . S2CID 20130470 .
- ^ Тейлор Ф.Дж., Коутс Д. (1989). «Код внутри кодонов». Биосистемы . 22 (3): 177–87. Бибкод : 1989BiSys..22..177T . дои : 10.1016/0303-2647(89)90059-2 . ПМИД 2650752 .
- ^ Ди Джулио М (октябрь 1989 г.). «Расширение, достигаемое за счет минимизации полярных расстояний в ходе эволюции генетического кода». Журнал молекулярной эволюции . 29 (4): 288–93. Бибкод : 1989JMolE..29..288D . дои : 10.1007/BF02103616 . ПМИД 2514270 . S2CID 20803686 .
- ^ Вонг Дж.Т. (февраль 1980 г.). «Роль минимизации химических расстояний между аминокислотами в эволюции генетического кода» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (2): 1083–6. Бибкод : 1980PNAS...77.1083W . дои : 10.1073/pnas.77.2.1083 . ПМЦ 348428 . ПМИД 6928661 .
- ^ Jump up to: Перейти обратно: а б с д Эривес А (август 2011 г.). «Модель ферментов протоантикодонов РНК, требующих гомохиральности L-аминокислот» . Журнал молекулярной эволюции . 73 (1–2): 10–22. Бибкод : 2011JMolE..73...10E . дои : 10.1007/s00239-011-9453-4 . ПМЦ 3223571 . ПМИД 21779963 .
- ^ Jump up to: Перейти обратно: а б Фриланд С.Дж., Найт Р.Д., Ландвебер Л.Ф., Херст Л.Д. (апрель 2000 г.). «Ранняя фиксация оптимального генетического кода» . Молекулярная биология и эволюция . 17 (4): 511–18. doi : 10.1093/oxfordjournals.molbev.a026331 . ПМИД 10742043 .
- ^ Крик Ф.Х. (декабрь 1968 г.). «Происхождение генетического кода». Журнал молекулярной эволюции . 38 (3): 367–79. дои : 10.1016/0022-2836(68)90392-6 . ПМИД 4887876 . S2CID 4144681 .
- ^ Хопфилд Джей-Джей (1978). «Происхождение генетического кода: проверяемая гипотеза, основанная на структуре, последовательности и кинетической корректуре тРНК» . ПНАС . 75 (9): 4334–4338. Бибкод : 1978PNAS...75.4334H . дои : 10.1073/pnas.75.9.4334 . ПМК 336109 . ПМИД 279919 .
- ^ Jump up to: Перейти обратно: а б Ярус М., Видманн Дж. Дж., Найт Р. (ноябрь 2009 г.). «Связывание РНК-аминокислот: стереохимическая эра генетического кода» . Журнал молекулярной эволюции . 69 (5): 406–29. Бибкод : 2009JMolE..69..406Y . дои : 10.1007/s00239-009-9270-1 . ПМИД 19795157 .
- ^ Браун, Шон М.; Ворачек, Вацлав; Фриланд, Стивен (5 апреля 2023 г.). «Как будет выглядеть алфавит чужеродных аминокислот и почему?». Астробиология . 23 (5): 536–549. Бибкод : 2023AsBio..23..536B . дои : 10.1089/ast.2022.0107 . ПМИД 37022727 . S2CID 257983174 .
- ^ Найт Р.Д., Фриланд С.Дж., Ландвебер Л.Ф. (июнь 1999 г.). «Отбор, история и химия: три лица генетического кода» . Тенденции биохимических наук . 24 (6): 241–7. дои : 10.1016/S0968-0004(99)01392-4 . PMID 10366854 .
- ^ Найт Р.Д., Ландвебер Л.Ф. (сентябрь 1998 г.). «Рифма или причина: взаимодействия РНК-аргинина и генетический код» . Химия и биология . 5 (9): 215–20 р. дои : 10.1016/S1074-5521(98)90001-1 . ПМИД 9751648 .
- ^ Сенгупта С., Хиггс П.Г. (2015). «Пути эволюции генетического кода у древних и современных организмов». Журнал молекулярной эволюции . 80 (5–6): 229–243. Бибкод : 2015JMolE..80..229S . дои : 10.1007/s00239-015-9686-8 . ПМИД 26054480 . S2CID 15542587 .
- ^ Брукс DJ, Фреско JR, Леск AM, Сингх М (октябрь 2002 г.). «Эволюция частот аминокислот в белках с течением времени: предполагаемый порядок введения аминокислот в генетический код» . Молекулярная биология и эволюция . 19 (10): 1645–55. doi : 10.1093/oxfordjournals.molbev.a003988 . ПМИД 12270892 .
- ^ Амирновин Р. (май 1997 г.). «Анализ метаболической теории происхождения генетического кода». Журнал молекулярной эволюции . 44 (5): 473–6. Бибкод : 1997JMolE..44..473A . дои : 10.1007/PL00006170 . ПМИД 9115171 . S2CID 23334860 .
- ^ Роннеберг Т.А., Ландвебер Л.Ф., Фриланд С.Дж. (декабрь 2000 г.). «Проверка биосинтетической теории генетического кода: факт или артефакт?» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (25): 13690–5. Бибкод : 2000PNAS...9713690R . дои : 10.1073/pnas.250403097 . ЧВК 17637 . ПМИД 11087835 .
- ^ Трифонов, Эдвард Н. (сентябрь 2009 г.). «Происхождение генетического кода и самых ранних олигопептидов» . Исследования в области микробиологии . 160 (7): 481–486. дои : 10.1016/j.resmic.2009.05.004 . ПМИД 19524038 .
- ^ Хиггс, Пол Г.; Пудриц, Ральф Э. (июнь 2009 г.). «Термодинамическая основа синтеза пребиотических аминокислот и природа первого генетического кода» . Астробиология . 9 (5): 483–490. arXiv : 0904.0402 . Бибкод : 2009AsBio...9..483H . дои : 10.1089/ast.2008.0280 . ISSN 1531-1074 . ПМИД 19566427 . S2CID 9039622 .
- ^ Халиотис, Анаргирос; Властаридис, Панайотис; Моссиалос, Димитрис; Ибба, Майкл; Беккер, Хьюберт Д.; Статопулос, Константинос; Амуциас, Григориос Д. (17 февраля 2017 г.). «Сложная эволюционная история аминоацил-тРНК-синтетаз» . Исследования нуклеиновых кислот . 45 (3): 1059–1068. дои : 10.1093/nar/gkw1182 . ISSN 0305-1048 . ПМК 5388404 . ПМИД 28180287 .
- ^ Нтунтуми, Криса; Властаридис, Панайотис; Моссиалос, Димитрис; Статопулос, Константинос; Илиопулос, Иоаннис; Промпонас, Василиос; Оливер, Стивен Дж; Амуциас, Григорис Д. (4 ноября 2019 г.). «Области низкой сложности в белках прокариот выполняют важные функциональные роли и высоко консервативны» . Исследования нуклеиновых кислот . 47 (19): 9998–10009. дои : 10.1093/nar/gkz730 . ISSN 0305-1048 . ПМК 6821194 . ПМИД 31504783 .
- ^ Jump up to: Перейти обратно: а б Фриланд С.Дж., Ву Т., Кельманн Н. (октябрь 2003 г.). «Дело об ошибке минимизации стандартного генетического кода». Происхождение жизни и эволюция биосферы . 33 (4–5): 457–77. Бибкод : 2003OLEB...33..457F . дои : 10.1023/А:1025771327614 . ПМИД 14604186 . S2CID 18823745 .
- ^ Баранов П.В., Венин М., Прован Г (2009). Геммелл, Нью-Джерси (ред.). «Уменьшение размера кодонов как происхождение триплетного генетического кода» . ПЛОС ОДИН . 4 (5): е5708. Бибкод : 2009PLoSO...4.5708B . дои : 10.1371/journal.pone.0005708 . ПМЦ 2682656 . ПМИД 19479032 .
- ^ Тлусти Т. (ноябрь 2007 г.). «Модель возникновения генетического кода как переход в зашумленный информационный канал». Журнал теоретической биологии . 249 (2): 331–42. arXiv : 1007.4122 . Бибкод : 2007JThBi.249..331T . дои : 10.1016/j.jtbi.2007.07.029 . ПМИД 17826800 . S2CID 12206140 .
- ^ Зоннеборн ТМ (1965). Брайсон В., Фогель Х. (ред.). Эволюционирующие гены и белки . Нью-Йорк: Академическая пресса. стр. 377–397.
- ^ Тлусти Т. (февраль 2008 г.). «Сценарий искажения скорости возникновения и эволюции шумных молекулярных кодов». Письма о физических отзывах . 100 (4): 048101. arXiv : 1007.4149 . Бибкод : 2008PhRvL.100d8101T . doi : 10.1103/PhysRevLett.100.048101 . ПМИД 18352335 . S2CID 12246664 .
- ^ Селла Дж., Арделл Д.Х. (сентябрь 2006 г.). «Коэволюция генов и генетических кодов: новый взгляд на замороженную аварию Крика». Журнал молекулярной эволюции . 63 (3): 297–313. Бибкод : 2006JMolE..63..297S . дои : 10.1007/s00239-004-0176-7 . ПМИД 16838217 . S2CID 1260806 .
- ^ Тлусти Т. (сентябрь 2010 г.). «Красочное происхождение генетического кода: теория информации, статистическая механика и появление молекулярных кодов». Обзоры физики жизни . 7 (3): 362–76. arXiv : 1007.3906 . Бибкод : 2010PhLRv...7..362T . дои : 10.1016/j.plrev.2010.06.002 . ПМИД 20558115 . S2CID 1845965 .
- ^ Джи Дж., Сундстрем А., Мэсси С.Э., Мишра Б. (ноябрь 2013 г.). «Что информационно-асимметричные игры могут рассказать нам о контексте «замороженной аварии» Крика?» . Журнал Королевского общества, Интерфейс . 10 (88): 20130614. doi : 10.1098/rsif.2013.0614 . ПМЦ 3785830 . ПМИД 23985735 .
- ^ Ицковиц С., Алон У (2007). «Генетический код почти оптимален для предоставления дополнительной информации в последовательностях, кодирующих белок» . Геномные исследования . 17 (4): 405–412. дои : 10.1101/гр.5987307 . ПМЦ 1832087 . ПМИД 17293451 .
Дальнейшее чтение [ править ]
- Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гилберт В.М. (1999). Введение в генетический анализ (7-е изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-3771-1 .
- Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 .
- Лодиш Х.Ф., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж.Э. (2000). Молекулярно-клеточная биология (4-е изд.). Сан-Франциско: WH Freeman. ISBN 9780716737063 .
- Каски Коннектикут, Ледер П. (апрель 2014 г.). «Код РНК: природный Розеттский камень» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (16): 5758–9. Бибкод : 2014PNAS..111.5758C . дои : 10.1073/pnas.1404819111 . ПМЦ 4000803 . ПМИД 24756939 .
Внешние ссылки [ править ]
- Генетические коды: Таблицы генетических кодов
- База данных использования кодонов — таблицы частотности кодонов для многих организмов.
- История расшифровки генетического кода

биохимии Темы |
|---|