Канцерогенез

Канцерогенез , также вызываемый онкогенезом или онкогенез , является образованием рака чего нормальные клетки превращаются , в результате в раковые клетки . Процесс характеризуется изменениями на клеточных, генетических и эпигенетических уровнях и аномальным клеточным делением . Разделение клеток - это физиологический процесс, который происходит почти во всех тканях и при различных обстоятельствах. Обычно баланс между пролиферацией и запрограммированной гибелью клеток в форме апоптоза поддерживается для обеспечения целостности тканей и органов . Согласно преобладающей принятой теории канцерогенеза, теория соматических мутаций, мутации в ДНК и эпимутации , которые приводят к раку, нарушают эти упорядоченные процессы, вмешиваясь в программирование, регулирующее процессы, нарушая нормальный баланс между пролиферацией и гибелью клеток. [ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ] Это приводит к неконтролируемому делению клеток и эволюции этих клеток путем естественного отбора в организме. Только определенные мутации приводят к раку, тогда как большинство мутаций не делают. [ Цитация необходима ]
Варианты наследственных генов могут предрасполагать людей к раку. Кроме того, факторы окружающей среды, такие как канцерогены и радиация, вызывают мутации, которые могут способствовать развитию рака. Наконец, случайные ошибки при нормальной репликации ДНК могут привести к мутациям вызывающих рак. [ 6 ] Ряд нескольких мутаций к определенным классам генов обычно требуется до того, как нормальная клетка превратится в раковую клетку . [ 7 ] [ 8 ] [ 9 ] [ 10 ] [ 11 ] Недавняя комплексная классификация на уровне пациентов и количественная оценка событий водителя в когортах TCGA показала, что в среднем 12 событий драйвера на опухоль, из которых 0,6 являются точечными мутациями в онкогенах , 1,5 являются амплификациями онкогенов, 1,2 являются точечными мутациями в подавляющих опухолевых , 2,1- подавляющие опухолевые, 1, 1-й водитель, -водители , 2-х водителя, 2- х водителя, 2-х водителя , 2-х водителя, 2-х водителей , 2-х водителя, 2-х водителя , 2-х водителя, 2-х водителя , 2- х водителя. Потери , и 1,5 являются приростом рычага хромосомы водителя . [ 12 ] Мутации в генах, которые регулируют деление клеток, апоптоз (гибель клеток) и репарация ДНК , могут привести к неконтролируемой пролиферации клеток и раку.
Рак в основном является заболеванием регуляции роста тканей. Чтобы нормальная клетка превратилась в раковую клетку, должны быть изменены гены , которые регулируют рост и дифференцировку клеток. [ 13 ] Генетические и эпигенетические изменения могут происходить на многих уровнях, от усиления или потери целых хромосом до мутации, влияющей на один нуклеотид ДНК , или до молчания или активации микроРНК, которая контролирует экспрессию от 100 до 500 генов. [ 14 ] [ 15 ] Существуют две широкие категории генов, которые влияют на эти изменения. Онкогены могут быть нормальными генами, которые экспрессируются на ненадлежащих высоких уровнях, или измененные гены, которые обладают новыми свойствами. В любом случае экспрессия этих генов способствует злокачественному фенотипу раковых клеток. Гены супрессоров опухолей - это гены, которые ингибируют деление клеток, выживаемость или другие свойства раковых клеток. Гены-супрессоры опухолей часто инвалируют генетическими изменениями, способствующими раком. Наконец , oncovirinae , вирусы , содержащие онкоген , классифицируются как онкогенные, поскольку они вызывают рост опухолевых тканей у хозяина . Этот процесс также называется вирусной трансформацией . Также считается, что рак вызван хромосомными аномалиями, как объясняется в теории рака хромосом . [ 16 ]
Причины
[ редактировать ]Генетический и эпигенетический
[ редактировать ]Существует разнообразная схема классификации для различных геномных изменений, которые могут способствовать генерации раковых клеток . Многие из этих изменений являются мутациями или изменениями в нуклеотидной последовательности геномной ДНК. Есть также много эпигенетических изменений, которые изменяют, экспрессируются ли гены или не экспрессируются. Анеуплоидия , присутствие аномального числа хромосом, является одним из геномных изменений, которое не является мутацией и может включать либо усиление, либо потерю одной или нескольких хромосом посредством ошибок при митозе . Масштабные мутации включают либо делецию , либо дублирование части хромосомы. Геномная амплификация происходит, когда клетка получает много копий (часто 20 или более) небольшой хромосомной области, обычно содержащей один или несколько онкогенов и прилегающий генетический материал. Транслокация происходит, когда две отдельные хромосомные области становятся ненормально слитыми, часто в характерном месте. Хорошо известным примером этого является филадельфийская хромосома или транслокация хромосом 9 и 22, которые встречаются в Хроническая миелогенная лейкоза и приводит к выработке BCR - ABL слитого белка , онкогенной тирозинкиназы . Маленькие мутации включают точечные мутации , делеции и вставки , которые могут возникнуть в промоторе гена и влиять на его экспрессию гена или могут возникнуть в кодирующей последовательности и изменять функцию или стабильность его белкового продукта. Разрушение одного гена также может быть результатом интеграции геномного материала из ДНК -вируса или ретровируса , и такое событие может также привести к экспрессии вирусных онкогенов в пораженных клетке и ее потомках. [ Цитация необходима ]
Повреждение ДНК
[ редактировать ]
Повреждение ДНК считается основной причиной рака. [ 17 ] Более 60 000 новых естественных случаев повреждения ДНК в среднем возникают в среднем, на клетку человека в день из-за эндогенных клеточных процессов (см. Повреждение ДНК статьи (естественно встречается) ).
Дополнительное повреждение ДНК может возникнуть в результате воздействия экзогенных агентов. В качестве одного примера экзогенного канцерогенного агента, табачный дым вызывает увеличение повреждения ДНК, и это повреждение ДНК, вероятно, вызывает увеличение рака легких из -за курения. [ 18 ] В других примерах ультрафиолетовый свет от солнечного излучения вызывает повреждение ДНК, которое важно при меланоме , [ 19 ] Helicobacter pylori инфекция производит высокие уровни активных форм кислорода , которые повреждают ДНК и способствуют раку желудка , [ 20 ] А Aspergillus flavus метаболит афлатоксин является разрушительным агентом ДНК, который является причиной при раке печени. [ 21 ]
Повреждение ДНК также может быть вызвано веществами, полученными в организме . Макрофаги и нейтрофилы в воспаленном эпителии толстой кишки являются источником реактивных форм кислорода, вызывающих повреждение ДНК, которое инициирует онкогенез толстой кишки , [ 22 ] и желчные кислоты, на высоком уровне в колоне людей, придерживающихся диеты с высоким содержанием жиров, также вызывают повреждение ДНК и способствуют раку толстой кишки. [ 23 ]
Такие экзогенные и эндогенные источники повреждения ДНК указаны в коробках в верхней части рисунка в этом разделе. Центральная роль повреждения ДНК в прогрессировании рака указана на втором уровне рисунка. Центральные элементы повреждения ДНК, эпигенетических изменений и дефицита ДНК репарации при прогрессировании на рак показаны красными.
Дефицит восстановления ДНК приведет к большему повреждению ДНК для накопления и увеличит риск развития рака. Например, люди с унаследованным нарушением в любом из 34 генов репарации ДНК (см. Статья дефицита дефицита ДНК ) подвергаются повышенному риску рака, причем некоторые дефекты вызывают до 100% вероятность рака на 100% (например, p53 ). мутации [ 24 ] Такие мутации зародышевой линии показаны в ящике слева от рисунка, что указывает на их вклад в дефицит восстановления ДНК. Тем не менее, такие мутации зародышевой линии (которые вызывают синдромы с высоким содержанием пенетрантов рака) являются причиной всего около одного процента раковых заболеваний. [ 25 ]
Большинство раковых заболеваний называют негитарным или «спорадическим видом рака». Около 30% спорадических раковых заболеваний имеют какой -то наследственный компонент, который в настоящее время не определяется, в то время как большинство, или 70% спорадических раковых заболеваний, не имеют наследственного компонента. [ 26 ]
При спорадическом раке дефицит репарации ДНК иногда обусловлен мутацией в гене репарации ДНК; Гораздо чаще, сниженная или отсутствующая экспрессия генов репарации ДНК обусловлена эпигенетическими изменениями , которые уменьшают или молчат экспрессия генов . Это указано на рисунке на 3 -м уровне сверху. Например, для 113 колоректальных раковых заболеваний, изученных в последовательности, только четыре имели миссенс -мутацию в гене Mgmt , репаратирующей ДНК , в то время как большинство имели снижение экспрессии MGMT из -за метилирования MGMT промоторной области (эпигенетическое изменение). [ 27 ]
Когда экспрессия генов репарации ДНК снижается, это вызывает дефицит репарации ДНК. Это показано на рисунке на 4 -м уровне сверху. При дефиците восстановления ДНК повреждение ДНК сохраняется в клетках на более высоком, чем типичный уровень (5 -й уровень сверху на рисунке); Этот избыточный ущерб вызывает повышенную частоту мутации и/или эпимутации (6 -й уровень с вершины рисунка). Экспериментально, скорости мутаций существенно увеличиваются в клетках, дефектных при восстановлении несоответствия ДНК [ 28 ] [ 29 ] или в гомологичном рекомбинационном восстановлении (HRR). [ 30 ] Хромосомные перестройки и анеуплоидия также увеличиваются в клетках, дефективных HRR, [ 31 ] Во время восстановления двойных разрывов ДНК или восстановления других повреждений ДНК не полностью очищенные сайты восстановления могут вызвать эпигенетическое молчание генов. [ 32 ] [ 33 ]
Соматические мутации и эпигенетические изменения, вызванные повреждением ДНК и недостатками в репарации ДНК, накапливаются в полевых дефектах . Полевые дефекты представляют собой нормальные ткани с множественными изменениями (обсуждаются в разделе ниже), и являются распространенными предшественниками для развития неупорядоченного и чрезмерно распространенного клона ткани при раке. Такие дефекты поля (второй уровень от нижней части рисунка) могут иметь многочисленные мутации и эпигенетические изменения.
Невозможно определить первоначальную причину для большинства конкретных видов рака. В некоторых случаях существует только одна причина: например, вирус HHV-8 вызывает все сарком Капоси . Тем не менее, с помощью методов и информации эпидемиологии рака , можно получить оценку вероятной причины в гораздо большей ситуации. Например, рак легких имеет несколько причин, включая употребление табака и радоновый газ . У мужчин, которые в настоящее время курят табак, развивается рак легких со скоростью в 14 раз больше, чем у мужчин, которые никогда не курили табак: вероятность рака легких у нынешнего курильщика, вызванного курением, составляет около 93%; Существует 7% вероятность того, что рак легких курильщиков был вызван газом радона или какой-либо другой причиной некачественной. [ 34 ] Эти статистические корреляции позволили исследователям сделать вывод, что определенные вещества или поведение являются канцерогенными. Табачный дым вызывает увеличение экзогенного повреждения ДНК, и это повреждение ДНК является вероятной причиной рака легких из -за курения. Среди более 5000 соединений в табачном дыме, генотоксичные ДНК-разрушающие агенты, которые встречаются как в самых высоких концентрациях, и которые имеют наиболее сильные мутагенные эффекты,-это акролеин , формальдегид , акрилонитрил , 1,3-бутадиен , ацетальдегид , этиленоксид и изопрен . [ 18 ]
Используя молекулярные биологические методы, можно охарактеризовать мутации, эпимутации или хромосомные аберрации в опухоли, а в области прогнозирования прогнозирования определенных больных раком достигается быстрый прогресс на основе спектра мутаций. Например, до половины всех опухолей имеют дефектный ген p53. Эта мутация связана с плохим прогнозом, поскольку эти опухолевые клетки с меньшей вероятностью будут входить в апоптоз или запрограммированную гибель клеток при повреждении терапией. Мутации теломеразы Удаляют дополнительные барьеры, продлевая количество раз, когда ячейка может разделить. Другие мутации позволяют опухоли выращивать новые кровеносные сосуды , чтобы обеспечить больше питательных веществ или метастазировать , распространяясь на другие части тела. Однако, как только рак формируется, он продолжает развиваться и производить подколты. В 2012 году было сообщено, что один образец рака почек, отобранный в девяти различных областях, имел 40 «повсеместных» мутаций, обнаруженных во всех девяти областях, 59 мутаций, разделенных некоторыми, но не все девять областей и 29 «частных» мутаций, присутствующих только в одной области. [ 35 ]
Линии клеток, в которых накапливаются все эти изменения ДНК, трудно проследить, но две недавние данные свидетельствуют о том, что нормальные стволовые клетки могут быть клетками происхождения при раке. [ 36 ] [ 37 ] Во -первых, существует очень положительная корреляция (RHO Спирмена = 0,81; P <3,5 × 10–8) между риском развития рака в ткани и количеством нормальных делений стволовых клеток, происходящих в той же ткани. Корреляция применяется к 31 типам рака и распространялась на пять порядков . [ 38 ] Эта корреляция означает, что если нормальные стволовые клетки из ткани разделит один раз, риск рака в этой ткани составляет приблизительно 1x. Если они делят 1000 раз, риск рака составляет 1000x. И если нормальные стволовые клетки от ткани делят 100 000 раз, риск рака в этой ткани составляет приблизительно 100 000x. Это убедительно предполагает, что основным фактором инициации рака является тот факт, что «нормальные» стволовые клетки делятся, что подразумевает, что рак происходит в нормальных, здоровых стволовых клетках. [ 37 ]
Во -вторых, статистика показывает, что большинство рака человека диагностируются у пожилых людей. Возможное объяснение заключается в том, что раковые заболевания возникают, потому что клетки накапливают повреждение во времени. ДНК является единственным клеточным компонентом, который может накапливать повреждение в течение всего курса жизни, а стволовые клетки - единственные клетки, которые могут передавать ДНК от зиготы к клеткам в конце жизни. Другие клетки, полученные из стволовых клеток, не поддерживают ДНК с самого начала жизни, пока не произойдет возможный рак. Это подразумевает, что большинство видов рака возникают из нормальных стволовых клеток. [ 36 ] [ 37 ]
Вклад дефектов полевых
[ редактировать ]
Термин « полевой рак » был впервые использован в 1953 году для описания области или «поля» эпителия, который был предварительно кондиционирован (в то время) в значительной степени неизвестные процессы, чтобы предрасполагать его к развитию рака. [ 39 ] С тех пор термины «полевая рака» и «полевой дефект» были использованы для описания предслененной ткани, в которой могут возникнуть новые раковые заболевания. [ Цитация необходима ]
Полевые дефекты были идентифицированы в связи с раком и важны для прогрессирования рака. [ 40 ] [ 41 ] Тем не менее, это было указано Рубином [ 42 ] что «подавляющее большинство исследований в области исследований рака было проведено на четко определенных опухолях in vivo или на дискретных неопластических очагах in vitro. Тем не менее, есть доказательства того, что более 80% соматических мутаций, обнаруженных в мутаторном фенотипе колоректальных опухоли человека, возникают до начала терминальной клональной расширения…»… » [ 43 ] Более половины соматических мутаций, идентифицированных в опухолях, происходило в досшескованной фазе (в полевом дефекте) во время роста, по-видимому, нормальных клеток. Также можно ожидать, что многие из эпигенетических изменений, присутствующих в опухолях, могли происходить в досхеджных дефектах поля. [ 44 ]
В толстой кишке дефект поля, вероятно, возникает при естественном отборе мутантной или эпигенетически измененной клетки среди стволовых клеток у основания одной из кишечных склепок на внутренней поверхности толстой кишки. Мутант или эпигенетически измененная стволовая клетка может заменить другие близлежащие стволовые клетки на естественный отбор. Это может привести к возникновению участка аномальной ткани. Рисунок в этом разделе включает в себя фотографию свежеостадочного и открытого продольного сегмента толстой кишки, показывающий рак толстой кишки и четыре полипа. Ниже фотография существует схематическая схема того, как мог бы образоваться большой участок мутантных или эпигенетически измененных клеток, показанный большой площадью желтой на диаграмме. В рамках этого первого большого пятна в диаграмме может возникнуть большой клон клеток), может возникнуть вторая такая мутация или эпигенетическое изменение, так что данная стволовая клетка получает преимущество по сравнению с его соседями, и эта измененная стволовая клетка может расширяться клонально, образуя вторичное пятно или подклан, в рамках исходного пятно. Это обозначено на диаграмме четырьмя меньшими пятнами разных цветов в большой желтой исходной области. В этих новых пятнах (подколах) процесс можно повторить несколько раз, обозначаемые все еще меньшими пятнами в четырех вторичных пятнах (с все еще разными цветами на диаграмме), которые клонально расширяются, пока не возникнут стволовые клетки, которые генерируют либо небольшие полипы, либо иначе злокачественное новообразование (рак). На фотографии кажущийся дефект поля в этом сегменте толстой кишки генерировал четыре полипа (помеченные размером с полипов, 6 мм, 5 мм и двух из 3 мм и раком около 3 см в его самом длинном измерении). Эти новообразования также указаны (на диаграмме под фото) 4 небольшими загарными кругами (полипов) и большей красной областью (рак). Рак на фотографии произошел в области слежественности толстой кишки, где толстая кишка соединяет тонкую кишку (помеченную) и где происходит приложение (помечено). Жир на фото является внешним по отношению к внешней стене толстой кишки. В сегменте толстой кишки, показанном здесь, толстой кишке была разрезана вдоль, чтобы обнажить свою внутреннюю поверхность и демонстрировать рак и полипы, возникающие во внутренней эпителиальной слизистой оболочке толстой кишки. [ Цитация необходима ]
Если общий процесс, с помощью которого возникает спорадический рак толстой кишки,-это образование досшескованного клона, которое распространяется по естественному отбору, с последующим образованием внутренних субглонов внутри первоначального клона, и поднубковые клоны внутри, затем рак толстой кишки обычно должен быть связан и предыдует, поля растущей аномальности, отражающие претерпетиленные события. Наиболее обширная область аномалии (самая внешняя нерегулярная область в диаграмме) будет отражать самое раннее событие в формировании злокачественного новообразования.
В экспериментальной оценке специфических недостатков восстановления ДНК при раковых заболеваниях также было показано, что в полевых дефектах, окружающих эти раковые заболевания, также наблюдалось много специфических недостатков восстановления ДНК. В таблице ниже приведены примеры, для которых был показан дефицит восстановления ДНК при раке, вызван эпигенетическим изменением, и несколько более низкие частоты, с которыми был обнаружен тот же эпигенетически вызван дефицит восстановления ДНК в дефекте окружающего поля.
| Рак | Ген | Частота при раке | Частота в полевом дефекте | Ссылка |
|---|---|---|---|---|
| Колоректальный | Mgmt | 46% | 34% | [ 45 ] |
| Колоректальный | Mgmt | 47% | 11% | [ 46 ] |
| Колоректальный | Mgmt | 70% | 60% | [ 47 ] |
| Колоректальный | MSH2 | 13% | 5% | [ 46 ] |
| Колоректальный | ERC1 | 100% | 40% | [ 48 ] |
| Колоректальный | PMS2 | 88% | 50% | [ 48 ] |
| Колоректальный | XPF | 55% | 40% | [ 48 ] |
| Голова и шея | Mgmt | 54% | 38% | [ 49 ] |
| Голова и шея | MLH1 | 33% | 25% | [ 50 ] |
| Голова и шея | MLH1 | 31% | 20% | [ 51 ] |
| Желудок | Mgmt | 88% | 78% | [ 52 ] |
| Желудок | MLH1 | 73% | 20% | [ 53 ] |
| Пищевод | MLH1 | 77%–100% | 23%–79% | [ 54 ] |
Некоторые из небольших полипов в полевых дефекте, показанных на фотографии открытого сегмента толстой кишки, могут быть относительно доброкачественными новообразованиями. В исследовании полипов в 1996 году менее 10 мм размер, обнаруженного во время колоноскопии, и последовала повторная колоноскопия в течение 3 лет, 25% оставались неизменными в размерах, 35% регрессировали или сокращались по размеру, а 40% выросли в размере. [ 55 ]
Нестабильность генома
[ редактировать ]Известно, что раковые заболевания демонстрируют нестабильность генома или «мутаторский фенотип». [ 56 ] Кодирующая белок ДНК в ядре составляет около 1,5% от общей геномной ДНК. [ 57 ] В рамках этой ДНК, кодирующей белок ДНК (называемый экзом ), средний рак молочной железы или толстой кишки может иметь мутации, изменяющие белок от 60 до 70, из которых около 3 или 4 могут быть мутациями «водителя», а остальные могут быть мутациями «пассажир». [ 44 ] Однако среднее количество мутаций последовательности ДНК во всем геноме (включая непротеиновые области ) в образце ткани рака молочной железы составляет около 20 000. [ 58 ] В среднем образце ткани меланомы (меланомы имеют более высокую экзома ), частоту мутации [ 44 ] ) общее количество мутаций последовательности ДНК составляет около 80 000. [ 59 ] Эти высокие частоты мутаций в общих нуклеотидных последовательностях в раковых заболеваниях позволяют предположить, что часто раннее изменение в полевых дефектах, вызывающее рак (например, желтая область на диаграмме в предыдущем разделе) является дефицитом репарации ДНК. Большие полевые дефекты, окружающие рак толстой кишки (простирающиеся до 10 см на каждой стороне рака) обнаружены [ 48 ] Часто иметь эпигенетические дефекты в двух или трех белках репарации ДНК ( ERCC1 , ERCC4 (XPF) и/или PMS2 ) во всей области поля дефекта. Когда экспрессия генов репарации ДНК снижается, повреждение ДНК накапливается в клетках с более высокой, чем нормальной скоростью, и это избыточное повреждение вызывает повышенную частоту мутации и/или эпимутации. Скорость мутаций сильно увеличивается в клетках, дефектных при восстановлении несоответствия ДНК [ 28 ] [ 29 ] или в гомологичном рекомбинационном восстановлении (HRR). [ 30 ] Дефицит восстановления ДНК, сам по себе, может позволить повреждению ДНК накапливаться, и подверженное ошибкам синтез трансляции некоторых поврежденных областей может привести к мутациям. Кроме того, неисправное восстановление этого накопленного повреждения ДНК может привести к эпимутациям. Эти новые мутации и/или эпимутации могут обеспечить пролиферативное преимущество, генерируя полевой дефект. Хотя мутации/эпимутации в генах репарации ДНК не имеют, сами не дают избирательного преимущества, они могут выполняться в качестве пассажиров в клетках, когда клетка получает дополнительную мутацию/эпимутацию, которая обеспечивает пролиферативное преимущество. [ Цитация необходима ]
Теории без мазона
[ редактировать ]Существует ряд теорий канцерогенеза и лечения рака, которые выходят за рамки научного мнения из -за отсутствия научного обоснования, логики или доказательной базы. Эти теории могут быть использованы для оправдания различных альтернативных методов лечения рака. Их следует отличить от тех теорий канцерогенеза, которые имеют логическую основу в рамках биологии рака, и из которых могут быть сделаны традиционно проверенные гипотезы. [ Цитация необходима ]
Однако несколько альтернативных теорий канцерогенеза основаны на научных данных и все чаще признаются. Некоторые исследователи считают, что рак может быть вызван анеуплоидией (числовые и структурные аномалии в хромосомах) [ 60 ] а не мутациями или эпимутациями. Рак также рассматривался как метаболическое заболевание, при котором клеточный метаболизм кислорода перемещается от пути, который генерирует энергию ( окислительное фосфорилирование ) к пути, который генерирует активные формы кислорода . [ 61 ] Это вызывает переключение энергии от окислительного фосфорилирования в аэробный гликолиз ( эффект Варбурга ) и накопление активных форм кислорода, приводящих к окислительному стрессу («Теория окислительного стресса рака»). [ 61 ] Другая концепция развития рака основана на воздействии слабых магнитных и электромагнитных полей и их влияния на окислительный стресс , известный как магнитокарциногенез. [ 62 ]
Ряд авторов поставили под сомнение предположение, что раковые заболевания являются результатом последовательных случайных мутаций как упростит, что предполагает, что рак возникает в результате неспособности организма ингибировать врожденную, запрограммированную пролиферативную тенденцию. [ 63 ] Связанная теория предполагает, что рак - это атавизм , эволюционный возврат к более ранней форме многоклеточной жизни . [ 64 ] Гены, ответственные за неконтролируемый рост клеток и сотрудничество между раковыми клетками, очень похожи на г -н, которые позволили первым многоклеточным формам жизни группировать вместе и процветать. Эти гены все еще существуют в геномах более сложных метазой , таких как люди, хотя в последнее время развитые гены контролируют их. Когда новые управляющие гены терпят неудачу по какой -либо причине, ячейка может вернуться к более примитивному программированию и размножается из -под контроля. Теория является альтернативой представлению о том, что рак начинается с мошеннических ячеек, которые подвергаются эволюции в организме. Вместо этого они обладают фиксированным количеством примитивных генов, которые постепенно активируются, что дает им конечную изменчивость. [ 65 ] Другая эволюционная теория возвращает корни рака обратно к происхождению эукариоте ( зародышевой) клетки с помощью массивного горизонтального переноса генов , когда геномы инфицирования вирусов были расщеплены (и, таким образом, ослаблены), но их фрагменты, интегрированные в геном хозяина как иммунная защита. Таким образом, рак возникает, когда редкая соматическая мутация рекомбирует такие фрагменты в функциональный драйвер пролиферации клеток. [ 66 ]
Раковая клеточная биология
[ редактировать ]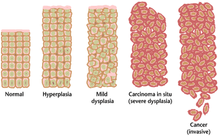
Часто множественные генетические изменения, которые приводят к раку, могут накапливаться много лет. В течение этого времени биологическое поведение презлокачественных клеток медленно изменяется от свойств нормальных клеток к ракоподобным свойствам. Предзнаменная ткань может иметь отличительный вид под микроскопом . Среди отличительных признаков презлоказного поражения-увеличение числа делящихся клеток , изменение размера и формы ядер , изменение размера и формы клеток , потеря специализированных клеточных признаков и потеря нормальной организации ткани. Дисплазия является аномальным типом чрезмерной пролиферации клеток, характеризующейся потерей нормального расположения ткани и структуры клеток в доловок клеток. Эти ранние неопластические изменения должны быть отличаются от гиперплазии , обратимого увеличения деления клеток, вызванного внешним стимулом, таким как гормональный дисбаланс или хроническое раздражение. [ Цитация необходима ]
Наиболее тяжелые случаи дисплазии называются карциномой in situ . На латыни термин in situ означает «на месте»; Карцинома in situ относится к неконтролируемому росту диспластических клеток, которые остаются в его первоначальном месте и не демонстрировали инвазию в другие ткани. Карцинома на месте может превратиться в инвазивную злокачественную опухоль и обычно удаляется хирургически при обнаружении.
Клональная эволюция
[ редактировать ]Подобно тому, как популяция животных подвергается эволюции , неконтролируемая популяция клеток также может подвергаться «эволюции». Этот нежелательный процесс называется соматической эволюцией , и как возникает рак и становится более злокачественным с течением времени. [ 67 ]
Большинство изменений в клеточном метаболизме, которые позволяют клеткам расти беспорядочной модой, приводят к гибели клеток. Однако, как только начинается рак, раковые клетки подвергаются процессу естественного отбора : немногие клетки с новыми генетическими изменениями, которые усиливают их выживание или размножение, и вскоре становятся все более быстрыми, и вскоре становятся все более широкими, чтобы доминировать в растущей опухоли, поскольку клетки с менее благоприятными генетическими изменениями выходят из строя. [ 68 ] Это тот же механизм, с помощью которого патогенные виды, такие как MRSA, могут стать устойчивыми к антибиотикам , и с помощью которого ВИЧ может стать устойчивым к лекарственным средствам , и с помощью которых заболевания растений и насекомые могут стать устойчивыми к пестицидам . рака Эта эволюция объясняет, почему рецидив часто включает в себя клетки, которые приобретали устойчивость к раке или устойчивость к лучевой терапии .
Биологические свойства раковых клеток
[ редактировать ]В статье 2000 года Ханахана и Вайнберга биологические свойства злокачественных опухолевых клеток были обобщены следующим образом: [ 69 ]
- Приобретение самодостаточности в сигналах роста , что приводит к неконтролируемому росту.
- Потеря чувствительности к сигналам против роста, что также приводит к неконтролируемому росту.
- Потеря способности при апоптозе , обеспечивая рост, несмотря на генетические ошибки и внешние сигналы против роста.
- Потеря способности к старению , что приводит к безграничному репликативному потенциалу (бессмертие)
- Приобретение устойчивого ангиогенеза , позволяя опухоли выходить за пределы ограничений пассивной диффузии питательных веществ.
- Приобретение способности вторгаться в соседние ткани , определяющая свойство инвазивной карциномы.
- Приобретение способности семян метастазов в отдаленных участках, поздних качества свойства некоторых злокачественных опухолей (карциномы или другие).
Завершение этих нескольких шагов было бы очень редким событием без:
- Потеря способности восстановить генетические ошибки, что приводит к увеличению скорости мутации (геномная нестабильность), что ускоряет все другие изменения.
Эти биологические изменения классические в карциномах ; Другие злокачественные опухоли, возможно, не потребуют их всех. Например, учитывая, что инвазия и смещение тканей в отдаленные участки являются нормальными свойствами лейкоцитов , эти шаги не нужны для развития лейкемии . Также различные шаги не обязательно представляют отдельные мутации. Например, инактивация одного гена, кодирующего белок p53 , вызовет геномную нестабильность, уклонение от апоптоза и увеличение ангиогенеза. Кроме того, не все раковые клетки делятся. Скорее, подмножество клеток в опухоли, называемой раковыми стволовыми клетками , воспроизводится, когда они генерируют дифференцированные клетки. [ 70 ]
Рак как дефект во взаимодействиях клеток
[ редактировать ]Обычно, как только ткань повреждена или инфицирована, поврежденные клетки вызывают воспаление, стимулируя специфические закономерности активности фермента и экспрессию генов цитокинов в окружающих клетках. [ 71 ] [ 72 ] Секретируются дискретные кластеры («кластеры цитокинов») молекул, которые действуют как посредники, вызывая активность последующих каскадов биохимических изменений. [ 73 ] Каждый цитокин связывается со специфическими рецепторами на различных типах клеток, и каждый тип клеток реагирует по очереди, изменяя активность внутриклеточных путей трансдукции сигнала, в зависимости от рецепторов, которые экспрессирует клетка, и сигнальные молекулы, присутствующие внутри клетки. [ 74 ] [ 75 ] В совокупности этот процесс перепрограммирования вызывает пошаговое изменение в клеточных фенотипах, что в конечном итоге приведет к восстановлению функции ткани и к восстановлению существенной структурной целостности. [ 76 ] [ 77 ] Таким образом, ткань может исцелить, в зависимости от продуктивной связи между клетками, присутствующими в месте повреждения и иммунной системой. [ 78 ] Одним из ключевых факторов в заживлении является регуляция экспрессии генов цитокинов, которая позволяет комплементарным группам клеток реагировать на воспалительные медиаторы таким образом, чтобы постепенно вызывает необходимые изменения в физиологии тканей. [ 79 ] [ 80 ] [ 81 ] Раковые клетки имеют либо постоянные (генетические) или обратимые (эпигенетические) изменения в их геноме, которые частично ингибируют их связь с окружающими клетками и с иммунной системой. [ 82 ] [ 83 ] Раковые клетки не общаются с их микроокружением тканей таким образом, чтобы защищать целостность тканей; Вместо этого движение и выживание раковых клеток становятся возможными в местах, где они могут нарушать функцию ткани. [ 84 ] [ 85 ] Раковые клетки выживают путем «перемешивания» сигнальных путей, которые обычно защищают ткань от иммунной системы. Это изменение иммунного ответа очевидно также на ранних стадиях злокачественности. [ 86 ] [ 87 ]
Одним из примеров повторного завода функции тканей при раке является активность транскрипционного фактора NF-κB . [ 88 ] NF-κB активирует экспрессию многочисленных генов, участвующих в переходе между воспалением и регенерацией, которые кодируют цитокины, коэффициенты адгезии и другие молекулы, которые могут изменить судьбу клеток. [ 89 ] Это перепрограммирование клеточных фенотипов обычно позволяет развивать полностью функциональную интактную ткань. [ 90 ] Активность NF-κB жестко контролируется несколькими белками, которые в совокупности гарантируют, что только дискретные кластеры генов индуцируются NF-κB в данной клетке и в данный момент времени. [ 91 ] Эта плотная регуляция обмена сигналами между клетками защищает ткани от чрезмерного воспаления и гарантирует, что различные типы клеток постепенно получают комплементарные функции и специфические положения. Неспособность этой взаимной регуляции между генетическим перепрограммированием и клеточным взаимодействием позволяет раковым клеткам вызывать метастазирование. Раковые клетки реагируют на цитокины и активируют сигнальные каскады, которые могут защитить их от иммунной системы. [ 88 ] [ 92 ]
В рыбе
[ редактировать ]Роль йода в морской рыбе (богатой йодом) и пресноводной рыбы (с дефицитом йода) не полностью понята, но сообщалось, что пресноводная рыба более восприимчива к инфекционным и, в частности, неопластическими и атеросклеротическими заболеваниями, чем морские рыбы. [ 93 ] [ 94 ] Морские рыбы Elasmobranch, такие как акулы, скаты и т. Д., Гораздо меньше влияют рак, чем пресноводные рыбы, и, следовательно, имеют стимулированные медицинские исследования, чтобы лучше понять канцерогенез. [ 95 ]
Механизмы
[ редактировать ]Чтобы клетки начинали неконтролируемо делить гены, которые регулируют рост клеток, должны быть нарушены. [ 96 ] Протоонкогены представляют собой гены, которые способствуют росту клеток и митозу , тогда как гены супрессоров опухолей препятствуют росту клеток или временно останавливают деление клеток для выполнения репарации ДНК . Как правило, серия нескольких мутаций в эти гены требуется перед нормальной клеточной трансформацией в раковую клетку . [ 10 ] Эта концепция иногда называется «Oncoevolution». Мутации в эти гены дают сигналы для опухолевых клеток, чтобы начать бесконтрольно делиться. Но неконтролируемое клеточное деление, которое характеризует рак, также требует, чтобы разделительная клетка дублировала все свои клеточные компоненты для создания двух дочерних клеток. Активация аэробного гликолиза ( эффект Варбурга ), которая не обязательно индуцируется мутациями в протоонкогенах и генах супрессора опухоли, [ 97 ] Обеспечивает большинство строительных блоков, необходимых для дублирования клеточных компонентов разделительной клетки, и, следовательно, также имеет важное значение для канцерогенеза. [ 61 ]
Онкогены
[ редактировать ]Онкогены способствуют росту клеток различными способами. Многие могут продуцировать гормоны , «химический посланник» между клетками, которые стимулируют митоз , эффект которого зависит от передачи сигнала приемной ткани или клеток. Другими словами, когда стимулируется гормональный рецептор на клетке реципиента, сигнал проводится с поверхности клетки в ядро клетки , чтобы влиять на некоторое изменение регуляции транскрипции генов на ядерном уровне. Некоторые онкогены являются частью самой системы передачи сигнала или рецепторов сигнала в клетках и самих тканях, что контролирует чувствительность к таким гормонам. Онкогены часто продуцируют митогены или участвуют в транскрипции ДНК в синтезе белка , что создает белки и ферменты, ответственные за продуцирование продуктов и биохимических клеток, использующих и взаимодействующие.
Мутации в протоонкогенах, которые являются нормально покоящимися аналогами онкогенов , могут модифицировать их экспрессию и функцию, увеличивая количество или активность белка продукта. Когда это происходит, протоонкогены становятся онкогенами , и этот переход расстраивает нормальный баланс регуляции клеточного цикла в клетке, что делает возможным неконтролируемый рост. Вероятность рака не может быть снижена путем удаления протоонкогенов из генома , даже если это было возможно, поскольку они имеют решающее значение для роста, восстановления и гомеостаза организма. Только когда они становятся мутированными, сигналы для роста становятся чрезмерными.
Одним из первых онкогенов , которые будут определены в исследованиях рака, является RAS Oncogene . Мутации в семействе протоонкогенов RAS (включающие H-Ras, N-Ras и K-Ras) очень распространены, обнаруживаются у 20-30% всех опухолей человека. [ 98 ] RAS был первоначально идентифицирован в геноме вируса саркомы Харви, и исследователи были удивлены тем, что этот ген присутствует не только в геноме человека, но и при лигировании стимулирующего контрольного элемента, но и индуцировать рак в культурах клеточных линий. [ 99 ] Недавно были предложены новые механизмы, что клеточная трансформация во время канцерогенеза была определена общим порогом онкогенных сетей (таких как передача сигналов RAS), но не статусом индивидуального онкогена. [ 100 ]
Протоонкогены
[ редактировать ]Протоонкогены способствуют росту клеток различными способами. Многие могут продуцировать гормоны , «химические посланники» между клетками, которые стимулируют митоз, эффект которого зависит от передачи сигнала приемной ткани или клеток. Некоторые из них ответственны за систему передачи сигнала и сигнальные рецепторы в самих клетках и самих тканях, что контролирует чувствительность к таким гормонам. Они часто продуцируют митогены или участвуют в транскрипции ДНК в синтезе белка , которые создают белки и ферменты, ответственные за продуцирование продуктов и биохимических клеток, которые используют и взаимодействуют.
Мутации в протоонкогенах могут модифицировать их экспрессию и функцию, увеличивая количество или активность продукта белка. Когда это происходит, они становятся онкогенами , и, таким образом, клетки имеют более высокую вероятность делиться чрезмерно и неудержимо. Вероятность рака не может быть снижена путем удаления протоонкогенов из генома , поскольку они имеют решающее значение для роста, восстановления и гомеостаза организма. Только когда они становятся мутированными, сигналы для роста становятся чрезмерными. Важно отметить, что ген, обладающий роли, способствующей росту, может увеличить канцерогенный потенциал клетки при условии, что все необходимые клеточные механизмы, которые позволяют росту, активируются. [ 101 ] Это состояние также включает в себя инактивацию специфических генов супрессоров опухоли (см. Ниже). Если условие не выполнено, ячейка может перестать расти и может приступить к смерти. Это делает выявление стадии и типа раковых клеток , которая растет под контролем заданного онкогена, имеющего решающее значение для разработки стратегий лечения.
Опухолевые гены
[ редактировать ]Гены-супрессоры опухолей кодируют сигналы антипролиферации и белки, которые подавляют митоз и рост клеток. Как правило, супрессоры опухолей являются факторами транскрипции , которые активируются клеточным стрессом или повреждением ДНК. Часто повреждение ДНК вызывает наличие свободно плавающего генетического материала, а также других признаков, и вызовет ферменты и пути, которые приводят к активации генов супрессоров опухолей . Функции таких генов заключаются в том, чтобы арестовать прогрессирование клеточного цикла для выполнения репарации ДНК, предотвращая передачу мутаций до дочерних клеток. Белок p53 , один из наиболее важных изученных генов -супрессоров опухолей, является фактором транскрипции, активируемым многими клеточными стрессорами, включая гипоксию и повреждение ультрафиолетового излучения .
Несмотря на почти половину всех раковых заболеваний, возможно, связанных с изменениями в p53, его функция подавления опухоли плохо изучена. P53 явно имеет две функции: одна ядерная роль в качестве фактора транскрипции, а другая цитоплазматическая роль в регуляции клеточного цикла, деления клеток и апоптоза.
Эффект Варбурга является преимущественным использованием гликолиза для энергии для поддержания роста рака. Было показано, что P53 регулирует переход от дыхания к гликолитическому пути. [ 102 ]
Тем не менее, мутация может повредить сам ген -супрессор опухоли, или сигнальный путь, который активирует его, «переключая его». Необходимым следствием этого является то, что репарация ДНК препятствует или ингибируется: повреждение ДНК накапливается без репарации, что неизбежно приводит к раку.
Мутации генов -супрессоров опухолей, которые встречаются в клетках зародышевой линии , передаются потомству и увеличивают вероятность диагностики рака в последующих поколениях. Члены этих семей имеют повышение частоты и снижение латентности множественных опухолей. Типы опухолей типичны для каждого типа мутации гена -супрессора опухоли, причем некоторые мутации вызывают определенные раковые заболевания, и другие мутации, вызывающие другие. Режим наследования мутантных опухолевых супрессоров заключается в том, что пораженный член наследует дефектную копию от одного родителя и обычную копию от другого. Например, у людей, которые наследуют один мутантный аллель p53 (и, следовательно, гетерозиготные по мутированному р53 ), могут развивать меланомы и рак поджелудочной железы , известный как синдром Лифамени . Другие унаследованные гены -синдромы опухоли включают в себя мутации Rb , связанные с ретинобластомой и мутациями гена APC , связанные с раком толстой кишки аденополипоза . Аденополипоз рак толстой кишки связан с тысячами полипов в Колоне, в то время как молодой, что приводит к Рак толстой кишки в относительно раннем возрасте. Наконец, унаследованные мутации в BRCA1 и BRCA2 приводят к раннему началу рака молочной железы .
Развитие рака было предложено в 1971 году, чтобы зависеть как минимум от двух мутационных событий. В том, что стало известно как Knudson гипотеза с двумя ударами , унаследованная мутация зародышевой линии в гене-супрессоре опухоли может вызвать рак только в том случае, если другое мутационное событие произошло позже в жизни организма, инактивируя другой аллель этого гена-супрессора опухоли . [ 103 ]
Обычно онкогены являются доминирующими , поскольку они содержат мутации усиления функции , в то время как мутированные супрессоры опухолей рецессив , поскольку они содержат мутации потери функции . Каждая ячейка имеет две копии одного и того же гена, по одной от каждого родителя, а в большинстве случаев усиление функциональных мутаций только в одной копии конкретного протоонкогена достаточно, чтобы сделать этот ген истинным онкогеном. С другой стороны, мутации потери функции должны произойти в обеих копиях гена-супрессора опухоли, чтобы сделать этот ген совершенно нефункциональным. Однако существуют случаи, в которых одна мутированная копия гена-супрессора опухоли может сделать другую, копию дикого типа не функциональными. Это явление называется доминирующим негативным эффектом и наблюдается во многих мутациях р53.
Две хитовые модели Кнудсона недавно была оспорена несколькими следователями. Инактивация одного аллеля некоторых генов -супрессоров опухолей достаточно, чтобы вызвать опухоли. Это явление называется гаплоинсуфанностью и было продемонстрировано рядом экспериментальных подходов. Опухоли, вызванные гаплоинсуфальностью, обычно имеют более поздний возраст начала по сравнению с опухолями с двумя ударами. [ 104 ]
Множественные мутации
[ редактировать ]
В целом, для возникновения рака необходимы мутации в обоих типах генов. Например, мутация, ограниченная одним онкогеном, будет подавлена нормальным контролем митоза и генами супрессора опухолей, сначала предполагаемой гипотезой Кнудсона . [ 8 ] Мутация только к одному гену опухолевого супрессора также не будет вызывать рак из -за наличия многих генов « резервного копирования », которые дублируют его функции. Только когда достаточное количество протоонкогенов мутировало в онкогены, и достаточно генов супрессоров опухолей дезактивировано или повреждено, сигналы для роста клеток подавляют сигналы для его регулирования, что рост клеток быстро выходит из-под контроля. [ 10 ] Часто, поскольку эти гены регулируют процессы, которые предотвращают большую часть повреждения самих генов, скорость мутаций увеличивается по мере того, как человек становится старше, потому что повреждение ДНК образует петлю обратной связи .
Мутация генов -супрессоров опухолей, которые передаются следующему поколению не только клеток, но их потомство , может вызвать повышенную вероятность того, что раковые заболевания будут унаследовать. Участники в этих семьях увеличили частоту и снижали латентность множественных опухолей. Режим наследования мутантных опухолевых супрессоров заключается в том, что пораженный член наследует дефектную копию от одного родителя и обычную копию от другого. Поскольку мутации в супрессорах опухолей действуют рецессивным образом (примечание, однако, есть исключения), потеря нормальной копии создает фенотип рака . Например, люди, которые гетерозиготны по мутациям р53, часто являются жертвами синдрома Лифаумени , и которые гетерозиготны по мутациям РБ развивают ретинобластому . Аналогичным образом, мутации в аденоматозном полипоз -гене Coli связаны с раком толстой кишки аденополипоза , с тысячами полипов в толстой кишке, в то время как мутации в BRCA1 и BRCA2 приводят к раннему началу рака молочной железы .
Новая идея, объявленная в 2011 году, представляет собой экстремальную версию множественных мутаций, называемую Chromothripsis по его сторонникам. Эта идея, затрагивающая только 2–3% случаев рака, хотя до 25% рака костей, включает катастрофическое разрушение хромосомы на десятки или сотни частей, а затем неправильно исправлено. Это разрушение, вероятно, происходит, когда хромосомы уплотнены во время нормального деления клеток , но триггер для разрушения неизвестен. В соответствии с этой моделью рак возникает в результате одного изолированного события, а не медленного накопления множественных мутаций. [ 105 ]
Не мутагенные канцерогены
[ редактировать ]Многие мутагены также являются канцерогенами , но некоторые канцерогены не являются мутагенами. Примеры канцерогенов, которые не являются мутагенами, включают алкоголь и эстроген . Считается, что они способствуют раку посредством их стимулирующего воздействия на скорость клеточного митоза . Более быстрые скорости митоза все больше оставляют меньше возможностей для восстановления ферментов для восстановления поврежденной ДНК во время репликации ДНК , увеличивая вероятность генетической ошибки. Ошибка, допущенная во время митоза, может привести к тому, что дочерние клетки получают неправильное количество хромосом , что приводит к анеуплоидии и может привести к раку.
Роль инфекций
[ редактировать ]Бактериальный
[ редактировать ]Helicobacter pylori может вызвать рак желудка . Хотя данные варьируются между разными странами, в целом от 1% до 3% людей, инфицированных Helicobacter pylori, развивается рак желудка в течение жизни по сравнению с 0,13% людей, у которых не было инфекции H. pylori . [ 106 ] [ 107 ] H. pylori инфекция очень распространена. Как оценивается в 2002 году, он присутствует в желудочных тканях 74% взрослых среднего возраста в развивающихся странах и 58% в развитых странах. [ 108 ] Поскольку от 1% до 3% инфицированных людей, вероятно, развивается рак желудка, [ 109 ] H. pylori -индуцированный рак желудка является третьей по величине причиной мировой смертности от рака с 2018 года. [ 110 ]
Инфекция H. pylori не вызывает никаких симптомов примерно у 80% инфицированных. [ 111 ] Около 75% людей, инфицированных H. pylori , развивают гастрит . [ 112 ] Таким образом, обычным следствием инфекции H. pylori является хронический бессимптомный гастрит. [ 113 ] Из -за обычного отсутствия симптомов, когда рак желудка, наконец, диагностируется, он часто довольно продвигается. Более половины пациентов с раком желудка имеют метастазирование лимфатических узлов, когда они первоначально диагностируются. [ 114 ]
Гастрит, вызванный H. pylori , сопровождается воспалением , характеризующимся инфильтрацией нейтрофилов и макрофагов в эпителий желудка, что способствует накоплению провоспалительных цитокинов и активных видов кислорода / реактивных видов азота (ROS / RNS). [ 115 ] Существенное присутствие ROS/RNS вызывает повреждение ДНК, включая 8-оксо-2'-дезоксигуанозин (8-OHDG). [ 115 ] Если инфицирование H. pylori несет ген цитотоксического CAGA (присутствует примерно в 60% западных изолятов и более высокий процент азиатских изолятов), они могут повысить уровень 8-OHDG в желудочных клетках на 8 раз, в то время как, если H. pylori не несет гена CAGA, увеличение в 8-ohDG составляет примерно 4-смех. [ 116 ] В дополнение к окислительному повреждению ДНК 8-OHDG, инфекция H. pylori вызывает другие характерные повреждения ДНК, включая разрывы ДНК двойной цепь. [ 117 ]
H. pylori также вызывает многие эпигенетические изменения, связанные с развитием рака. [ 118 ] [ 119 ] Эти эпигенетические изменения связаны с H. pylori -индуцированным метилированием сайтов CPG у промоторов генов [ 118 ] и H. pylori -индуцированная измененная экспрессия множественных микроРНК . [ 119 ]
Как рассмотрел Сантос и Рибейро [ 120 ] Инфекция H. pylori связана с эпигенетически сниженной эффективностью механизма восстановления ДНК, которая способствует накоплению мутаций и геномной нестабильности, а также канцерогенеза желудка. В частности, Raza et al. [ 121 ] показала, что экспрессия двух белков репарации ДНК, ERCC1 и PMS2 , была серьезно снижена после того, как H. pylori прогрессировала инфекция , вызванная диспепсией . Диспепсия встречается примерно у 20% инфицированных людей. [ 122 ] Кроме того, как рассмотрено Raza et al., [ 121 ] Человеческая желудочная инфекция H. pylori вызывает эпигенетически сниженную экспрессию белков белков, репаративных ДНК, MLH1 , MGMT и MRE11 . Снижение восстановления ДНК в присутствии повышенного повреждения ДНК увеличивает канцерогенные мутации и, вероятно, является значительной причиной канцерогенеза H. pylori .
Другие бактерии также могут играть роль в канцерогенезе. Контрольная точка контроля клеточного цикла и апоптоза с помощью p53 ингибируется микоплазменной бактерией, [ 123 ] позволяя клеткам с повреждением ДНК «запустить апоптотический красный свет» и пройти через клеточный цикл.
Популярный
[ редактировать ]Кроме того, многие раковые заболевания происходят из вирусной инфекции ; Это особенно верно для животных, таких как птицы , но меньше у людей . 12% рака человека можно отнести к вирусной инфекции. [ 124 ] Способ, индуцированного вирусом, можно разделить на два, остро, преобразуя или медленно трансформируя . При остро трансформирующихся вирусах вирусные частицы несут ген, который кодирует для гиперактивного онкогена, называемого вирусным онкогеном (V-ONC), и инфицированная клетка трансформируется, как только экспрессируется V-ONC. Напротив, в медленно трансформирующихся вирусах вставляется геном вируса, особенно в связи с тем, что вставка вирусного генома является обязательной частью ретровирусов , около протоонкогена в геноме хозяина. Вирусный промотор или другие элементы регуляции транскрипции, в свою очередь, вызывают чрезмерную экспрессию этого протоонкогена, который, в свою очередь, вызывает неконтролируемую клеточную пролиферацию. Поскольку вставка вирусного генома не специфична для протоонкогенов, и вероятность вставки вблизи этого протоонкогена является низкой, медленно трансформирующие вирусы имеют очень длинную латентность опухоли по сравнению с острым трансформирующим вирусом, который уже несет вирус-онкоген.
Вирусы, которые, как известно, вызывают рак, такой как ВПЧ ( рак шейки матки ), гепатит В ( рак печени ) и EBV (тип лимфомы ), являются вирусами ДНК. Считается, что когда вирус заражает клетку, он вводит часть своей собственной ДНК вблизи генов роста клеток, вызывая деление клеток. Группа измененных клеток, которые образуются из первого деления клеток, имеют одинаковую вирусную ДНК вблизи генов роста клеток. Группа измененных ячеек в настоящее время является особенной, потому что один из нормальных контролей роста был потерян.
В зависимости от их местоположения, клетки могут быть повреждены с помощью радиации, химикатов от сигаретного дыма и воспаления от бактериальной инфекции или других вирусов. У каждой ячейки есть шанс на повреждение. Клетки часто умирают, если они повреждены, путем неудачи жизненно важного процесса или иммунной системы, однако иногда повреждение выберет один ген рака. У старого человека есть тысячи, десятки тысяч или сотни тысяч нокаутированных клеток. Вероятность того, что любой из них сформирует рак, очень низкая. [ Цитация необходима ]
Когда повреждение происходит в любой области измененных ячеек, происходит что -то другое. Каждая из клеток имеет потенциал для роста. Измененные ячейки будут делиться быстрее, когда область повреждена физическими, химическими или вирусными агентами. Был установлен порочный круг : повреждение площади приведет к разделению измененных ячеек, вызывая большую вероятность того, что они будут испытывать нокауты.
Эта модель канцерогенеза популярна, потому что она объясняет, почему растут рак. Можно ожидать, что клетки, которые повреждены через радиацию, будут умереть или, по крайней мере, будут хуже, потому что у них меньше работы генов; Вирусы увеличивают количество работающих генов.
Одна мысль заключается в том, что мы можем получить тысячи вакцин, чтобы предотвратить каждый вирус, который может изменить наши клетки. Вирусы могут оказывать различное влияние на разные части тела. Возможно, можно было бы предотвратить ряд различных видов рака, иммунизируя против одного вирусного агента. Вполне вероятно, что ВПЧ, например, играет роль в раке слизистых оболочек рта.
Гельминтиаз
[ редактировать ]Определенные паразитические черви, как известно, являются канцерогенными. [ 125 ] К ним относятся:
- Clonorchis sinensis (организм, вызывающий клонорхиоз ) и opistorchis viverrini (вызывая опиорхиоз ) связаны с холангиокарциномой . [ 126 ]
- шистосомы Виды (организмы, вызывающие шистосомоз ) связаны с раком мочевого пузыря .
Эпигенетика
[ редактировать ]Эпигенетика -это изучение регуляции экспрессии генов посредством химических, не мутационных изменений в структуре ДНК. Теория эпигенетики в патогенезе рака заключается в том, что не мутационные изменения в ДНК могут привести к изменениям в экспрессии генов. Обычно онкогены молчат, например, из -за метилирования ДНК . Потеря этого метилирования может вызвать аберрантную экспрессию онкогенов , что приводит к патогенезу рака. Известные механизмы эпигенетического изменения включают метилирование ДНК и метилирование или ацетилирование гистоновых белков, связанных с хромосомной ДНК в специфических местах. Классы лекарств, известные как ингибиторы HDAC и ингибиторы ДНК метилтрансферазы , могут повторно регулировать эпигенетическую передачу сигналов в раковой клетке .
Эпимутации включают метилирование или деметилирование островов CPG промоторных областей генов , которые приводят к репрессии или депрессии, соответственно экспрессии генов. [ 127 ] [ 128 ] [ 129 ] Эпимутации также могут происходить путем ацетилирования, метилирования, фосфорилирования или других изменений в гистонах, создавая гистоновый код , который подавляет или активирует экспрессию генов, и такие эпимутации гистона могут быть важными эпигенетическими факторами при раке. [ 130 ] [ 131 ] Кроме того, канцерогенная эпимутация может происходить за счет изменений архитектуры хромосом, вызванных белками, такими как HMGA2 . [ 132 ] Дальнейший источник эпимутации обусловлено увеличением или снижением экспрессии микроРНК (miRNAs). Например, дополнительная экспрессия miR-137 может вызвать подавление экспрессии 491 генов, а miR-137 эпигенетически молчит при 32% колоректальных раков> [ 15 ]
Раковые стволовые клетки
[ редактировать ]Новый способ рассмотрения канцерогенеза происходит от интеграции идей биологии развития в онкологию . Гипотеза стволовых клеток рака предполагает , что различные виды клеток в гетерогенной опухоли возникают из одной клетки, называемой раковой стволовой клетки. Стволовые клетки рака могут возникать в результате трансформации взрослых стволовых клеток или дифференцированных клеток в организме. Эти клетки сохраняются в качестве подкомпонента опухоли и сохраняют ключевые свойства стволовых клеток. Они приводят к различным клеткам, способны к самообновлению и гомеостатическому контролю. [ 133 ] Кроме того, рецидив рака и появление метастазирования также объясняются этими клетками. Гипотеза рака стволовых клеток не противоречит более ранним концепциям канцерогенеза. Гипотеза раковых стволовых клеток была предложенным механизмом, который способствует неоднородности опухоли .
Клональная эволюция
[ редактировать ]В то время как генетические и эпигенетические изменения в генах супрессоров опухолей и онкогенах изменяют поведение клеток, эти изменения, в конце концов, приводят к раку через их влияние на популяцию опухолевых клеток и их микроокружение. [ 67 ] Мутантные клетки в новообразованиях конкурируют за пространство и ресурсы. Таким образом, клон с мутацией в гене -супрессоре опухоли или онкогена будет расширяться только в новообразованиях, если эта мутация дает клону конкурентное преимущество перед другими клонами и нормальными клетками в ее микроокружении. [ 134 ] Таким образом, процесс канцерогенеза является формально процессом дарвиновской эволюции , известной как соматическая или клональная эволюция . [ 68 ] Кроме того, в свете дарвинистических механизмов канцерогенеза, было теоретизировано, что различные формы рака можно классифицировать как половые и геронтологические. Антропологические исследования в настоящее время проводится на раке как естественный эволюционный процесс, посредством которого естественный отбор разрушает экологически неполноценные фенотипы, поддерживая других. Согласно этой теории, рак бывает в двух отдельных типах: от рождения до конца полового созревания (приблизительно 20 лет) телеологически склонен к поддерживающей групповой динамике и от середины жизни до смерти (приблизительно 40+) телеологически наклоненных от перенаселенной групповой динамики. [ Цитация необходима ]
Смотрите также
[ редактировать ]- Раковая клетка
- Armitage -Doll Многоступенчатая модель канцерогенеза
- Линейная модель безпорожирования
- Онкогенный белок ретровириды
Ссылки
[ редактировать ]- ^ Маджерус, Мари-Анжан (1 июля 2022 года). «Причина рака: объединяющая теория» . Достижения в области биологии рака - метастазирование . 4 : 100034. DOI : 10.1016/J.Adcanc.2022.100034 . ISSN 2667-3940 . S2CID 247145082 .
- ^ Новелл, Питер С. (1 октября 1976 г.). «Клональная эволюция популяций опухолевых клеток: приобретенная генетическая лабельность позволяет поэтапному выбору вариантов подсолин и лежит в основе прогрессирования опухоли» . Наука . 194 (4260): 23–28. Bibcode : 1976sci ... 194 ... 23n . doi : 10.1126/science.959840 . ISSN 0036-8075 . PMID 959840 . S2CID 38445059 .
- ^ Ханахан, Дуглас; Вайнберг, Роберт А (7 января 2000 г.). «Признаки рака» . Клетка . 100 (1): 57–70. doi : 10.1016/s0092-8674 (00) 81683-9 . ISSN 0092-8674 . PMID 10647931 . S2CID 1478778 .
- ^ Хан, Уильям С.; Вайнберг, Роберт А. (14 ноября 2002 г.). «Правила создания опухолевых клеток человека» . Новая Англия Журнал медицины . 347 (20): 1593–1603. doi : 10.1056/nejmra021902 . ISSN 0028-4793 . PMID 12432047 .
- ^ Калкинс, Гэри Н. (11 декабря 1914 г.). «О вопросе о появлении злокачественных опухолей. Бес. Бовери. Йена, Густав Фишер. 1914. 64 страницы» . Наука . 40 (1041): 857–859. Doi : 10.1126/science.40.1041.857 . ISSN 0036-8075 .
- ^ Tomasetti C, Li L, Vogelstein B (23 марта 2017 г.). «Разделители стволовых клеток, соматические мутации, этиология рака и профилактика рака» . Наука . 355 (6331): 1330–1334. BIBCODE : 2017SCI ... 355.1330T . doi : 10.1126/science.aaf9011 . PMC 5852673 . PMID 28336671 .
- ^ Вуд Л.Д., Парсонс Д.В., Джонс С., Лин Дж, Шеблом Т., Лири Р.Дж. и др. (Ноябрь 2007 г.). «Геномные ландшафты молочной железы человека и колоректального рака». Наука . 318 (5853): 1108–13. Bibcode : 2007sci ... 318.1108W . Citeseerx 10.1.1.218.5477 . doi : 10.1126/science.1145720 . PMID 17932254 . S2CID 7586573 .
- ^ Подпрыгнуть до: а беременный Knudson AG (ноябрь 2001 г.). «Два генетических хита (более или менее) для рака». Природные обзоры. Рак . 1 (2): 157–62. doi : 10.1038/35101030 . PMID 11905807 . S2CID 20201610 .
- ^ Fearon ER, Vogelstein B (июнь 1990 г.). «Генетическая модель для колоректального онкогенеза» . Клетка . 61 (5): 759–67. doi : 10.1016/0092-8674 (90) 90186-I . PMID 2188735 . S2CID 22975880 .
- ^ Подпрыгнуть до: а беременный в Belikov AV (сентябрь 2017 г.). «Количество ключевых канцерогенных событий может быть предсказано с частоты рака» . Научные отчеты . 7 (1): 12170. BIBCODE : 2017NATSR ... 712170B . doi : 10.1038/s41598-017-12448-7 . PMC 5610194 . PMID 28939880 .
- ^ Беликов А.В., Вьяткин А., Ленов С.В. (6 августа 2021 г.). «Распределение Erlang приближается к возрастному распределению заболеваемости в детстве и молодом раке взрослой жизни» . ПЕРЕЙ . 9 : E11976. doi : 10.7717/peerj.11976 . PMC 8351573 . PMID 34434669 .
- ^ Вьяткин, Алекси Д.; Otnyukov, Danila v.; Ленов, Сергей В.; Беликов, Алекси В. (14 января 2022 г.). «Комплексная классификация на уровне пациентов и количественная оценка событий водителя в когортах TCGA Pancanatlas» . PLOS Genetics . 18 (1): E1009996. doi : 10.1371/journal.pgen.1009996 . PMC 8759692 . PMID 35030162 .
- ^ Croce CM (январь 2008 г.). «Онкогены и рак». Новая Англия Журнал медицины . 358 (5): 502–11. doi : 10.1056/nejmra072367 . PMID 18234754 .
- ^ Лим Л.П., Лау Н.К., Гаррет-Энгеле П., Гримсон А., Шелтер Дж. М., Касл Дж., Бартел Д.П., Линсли П.С., Джонсон Дж. М. (февраль 2005 г.). «Анализ микрочипов показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа . 433 (7027): 769–73. Bibcode : 2005natur.433..769L . doi : 10.1038/nature03315 . PMID 15685193 . S2CID 4430576 .
- ^ Подпрыгнуть до: а беременный Balaguer F, Lozano JJ, Cuatreass M, Nagasaka T, Boland CR, Goel A (август 2010 г.). «Эпигенетическое молчание miR-137-это раннее событие в колоректальном канцерогенезе » РАНКА 70 (16): 6609–1 Doi : 10.1158/0008-5472.can- 10-0 PMC 2922409 PMID 20682795
- ^ «Как дисбаланс хромосом может стимулировать рак | Гарвардская медицинская школа» . Hms.harvard.edu . 6 июля 2023 года . Получено 2 апреля 2024 года .
- ^ Кастан М.Б. (апрель 2008 г.). «Ответы на повреждение ДНК: механизмы и роли в болезнях человека: 2007 г. Мемориальная премия GHA Clowes» . Молекулярное исследование рака . 6 (4): 517–24. doi : 10.1158/1541-7786.mcr-08-0020 . PMID 18403632 .
- ^ Подпрыгнуть до: а беременный Каннингем Ф.Х., Фибелкорн С., Джонсон М., Мередит С (ноябрь 2011). «Новое применение окраски подхода к воздействию: разделение токсикантов табачного дыма». Пищевая и химическая токсикология . 49 (11): 2921–33. doi : 10.1016/j.fct.2011.07.019 . PMID 21802474 .
- ^ Kanavy HE, Gerstenblith MR (декабрь 2011 г.). «Ультрафиолетовое излучение и меланома». Семинары в кожной медицине и хирургии . 30 (4): 222–8. doi : 10.1016/j.sder.2011.08.003 . PMID 22123420 .
- ^ Ханда О, Найто Y, Йошикава Т. (2011). «Окислительно -восстановительная биология и желудочный канцерогенез: роль Helicobacter pylori» . Окислительно -восстановительный отчет . 16 (1): 1–7. doi : 10.1179/174329211x12968219310756 . PMC 6837368 . PMID 21605492 .
- ^ Smela ME, Hamm ML, Henderson PT, Harris CM, Harris TM, Essigmann JM (май 2002). «Афлатоксин B (1) формамидопиримидиновый аддукт играет важную роль в выборе типов мутаций, наблюдаемых при гепатоцеллюлярной карциноме человека» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (10): 6655–60. Bibcode : 2002pnas ... 99,6655S . doi : 10.1073/pnas.102167699 . PMC 124458 . PMID 12011430 .
- ^ Кацурано М., Нива Т., Ясуи Ю., Шигемацу Ю., Ямашита С., Такешима Х., Ли М.С., Ким Ю.Дж., Танака Т., Ушиджима Т (январь 2012 г.). «Сформирование ранней стадии дефекта эпигенетического поля в модели колита мыши и несущественная роль Т- и В-клеток при индукции метилирования ДНК» . Онкоген . 31 (3): 342–51. doi : 10.1038/onc.2011.241 . PMID 21685942 .
- ^ Бернштейн С., Холубек Х., Бхаттачарья А.К., Нгуен Х., Пейн С.М., Зейтлин Б., Бернштейн Х (август 2011 г.). «Канцерогенность дезоксихолата, вторичная желчная кислота» . Архив токсикологии . 85 (8): 863–71. doi : 10.1007/s00204-011-0648-7 . PMC 3149672 . PMID 21267546 .
- ^ Малкин Д (апрель 2011 г.). «Синдром Ли-Фраумени» . Гены и рак . 2 (4): 475–84. doi : 10.1177/1947601911413466 . PMC 3135649 . PMID 21779515 .
- ^ Fearon ER (ноябрь 1997 г.). «Синдромы рака человека: подсказки о происхождении и природе рака». Наука . 278 (5340): 1043–50. Bibcode : 1997sci ... 278.1043f . doi : 10.1126/science.278.5340.1043 . PMID 9353177 .
- ^ Лихтенштейн П., Холм Н.В., Веркасало П.К., Илиаду А., Каприо Дж., Коскенвуо М., Пуккала Е., Скайт А., Хемминки К (июль 2000 г.). «Экологические и наследственные факторы в причинно-следственной связи-анализы когортов близнецов из Швеции, Дании и Финляндии» . Новая Англия Журнал медицины . 343 (2): 78–85. doi : 10.1056/nejm200007133430201 . PMID 10891514 .
- ^ Halford S, Rowan A, Sawyer E, Talbot I, Tomlinson I (июнь 2005 г.). «O (6) -метилгуаниновая метилтрансфераза при колоректальном раке: обнаружение мутаций, потеря экспрессии и слабая связь с переходами G: C> A: T» . Кишечник 54 (6): 797–802. doi : 10.1136/gut.2004.059535 . PMC 1774551 . PMID 15888787 .
- ^ Подпрыгнуть до: а беременный Нараянан Л., Фрицелл Дж.А., Бейкер С.М., Лискай Р.М., Глейзер П.М. (апрель 1997 г.). «Повышенные уровни мутации в нескольких тканях мышей с дефицитом в гене репарации несоответствия ДНК PMS2» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (7): 3122–7. Bibcode : 1997pnas ... 94.3122N . doi : 10.1073/pnas.94.7.3122 . PMC 20332 . PMID 9096356 .
- ^ Подпрыгнуть до: а беременный Hegan DC, Narayanan L, Jirik FR, Edelmann W, Liskay RM, Glazer PM (декабрь 2006 г.). «Разные паттерны генетической нестабильности у мышей с дефицитом в генах восстановления несоответствия PMS2, MLH1, MSH2, MSH3 и MSH6» . Канцерогенез . 27 (12): 2402–8. doi : 10.1093/carcin/bgl079 . PMC 2612936 . PMID 16728433 .
- ^ Подпрыгнуть до: а беременный Tutt AN, Van Oostrom CT, Ross GM, Van Steeg H, Ashworth A (март 2002 г.). «Разрушение BRCA2 увеличивает скорость спонтанной мутации in vivo: синергизм с ионизирующим радиацией» . Embo сообщает . 3 (3): 255–60. doi : 10.1093/embo-reports/kvf037 . PMC 1084010 . PMID 11850397 .
- ^ Немецкий J (март 1969 г.). «Синдром Блума. И. Генетические и клинические наблюдения у первых двадцати семи пациентов» . Американский журнал человеческой генетики . 21 (2): 196–227. PMC 1706430 . PMID 5770175 .
- ^ О'Хаган Х.М., Мохаммад Х.П., Бэйлин С.Б. (август 2008 г.). Ли JT (ред.). «Двойные разрывы цепей могут инициировать молчание генов и SIRT1-зависимое начало метилирования ДНК на экзогенном промоторе CPG Island» . PLOS Genetics . 4 (8): E1000155. doi : 10.1371/journal.pgen.1000155 . PMC 2491723 . PMID 18704159 .
- ^ Cuozzo C, Porcellini A, Angrisan T, Moranus, Lee B, No Pardo A, Messina S, Yulian R, Fusco A, Santillo MR, Muller MT, Chiartof L, Gottesman Me, Avvediment EV (Jony 2007). «Повреждение ДНК, репарация с гомологией и метилирование ДНК » PLOS Genetics 3 (7): E1 Doi : 10.1371/ journal.pgen.0 PMC 1913100 PMID 17616978
- ^ Villeneuve PJ, Mao Y (ноябрь 1994 г.). «Вероятность развития рака легких, статус курения, Канада». Канадский журнал общественного здравоохранения . 85 (6): 385–8. PMID 7895211 .
- ^ Gerlinger M, Rowan AJ, Horswell S, Larkin J, Endesfelder D, Gronroos E, et al. (Март 2012 г.). «Внутренняя неоднородность и разветвленная эволюция, выявленные с помощью множественного секвенирования» . Новая Англия Журнал медицины . 366 (10): 883–92. doi : 10.1056/nejmoa1113205 . PMC 4878653 . PMID 22397650 .
- ^ Подпрыгнуть до: а беременный Лопес-Лазаро М (август 2015 г.). Полем CLL CYM . 14 (16): 2547–8 doi : 10.1080/ 15384101.1015.1062330 PMC 5242319 . PMID 26090957 .
- ^ Подпрыгнуть до: а беременный в Лопес-Лазаро М (май 2015). «Миграционная способность стволовых клеток может объяснить существование рака неизвестного первичного сайта. Переосмысление метастазирования» . Онкосса . 2 (5): 467–75. doi : 10.18632/oncoscience.159 . PMC 4468332 . PMID 26097879 .
- ^ Tomasetti C, Vogelstein B (январь 2015 г.). «Этиология рака. Изменение риска рака среди тканей может быть объяснено количеством разделов стволовых клеток» . Наука . 347 (6217): 78–81. doi : 10.1126/science.1260825 . PMC 4446723 . PMID 25554788 .
- ^ Убэтер Д.П., Саутвик Х.В., Смейкал В. (сентябрь 1953 г.). «Полевая рака в пероральном стратифицированном плоскоклеточном эпителии; клинические последствия многоцентрического происхождения» . Рак . 6 (5): 963–8. doi : 10.1002/1097-0142 (195309) 6: 5 <963 :: AID-CNCR2820060515> 3.0.CO; 2-Q . PMID 13094644 . S2CID 6736946 .
- ^ Бернштейн С., Бернштейн Х., Пейн С.М., Дворак К., Гаревал Х (февраль 2008 г.). «Полевые дефекты в прогрессии к раке желудочно -кишечного тракта» . обзор. Раковые письма . 260 (1–2): 1–10. doi : 10.1016/j.canlet.2007.11.027 . PMC 2744582 . PMID 18164807 .
- ^ Ngyuyen H, Loustaunau C, Fasta A, Ramsey L, Hassounah N, Taylor H, Crouse R, Payne CM, Tsitis VL, Goldschmid S, Banerjee B, Perini RF, Bernstein C (2010). «Dficient PMS2, ERCC1, KU86, CCOI в полевых дефектах во время прогрессирования рака толстой кишки » Журнал визуализированных экспериментов (41): 1931. doi : 10.3791/ 1 3149991PMC PMID 20689513
- ^ Рубин Х (март 2011 г.). «Поля и полевые ракеты: преенепластическое происхождение рака: бессимптомные гиперпластические поля являются предшественниками неоплазии, а их прогрессирование до опухолей может отслеживать плотностью насыщения в культуре». Биологии . 33 (3): 224–31. doi : 10.1002/bies.201000067 . PMID 21254148 . S2CID 44981539 .
- ^ Цао Дж. Л., Ятабе Ю., Саловаара Р., Ярвинен Х.Дж., Меклин Дж.П., Аалтонен Л.А., Таваре С., Шибата Д. (февраль 2000 г.). «Генетическая реконструкция отдельных историй колоректальной опухоли» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (3): 1236–41. Bibcode : 2000pnas ... 97.1236t . doi : 10.1073/pnas.97.3.1236 . PMC 15581 . PMID 106555514 .
- ^ Подпрыгнуть до: а беременный в Vogelstein B, Papadopoulos N, Velculescu Ve, Zhou S, Diaz LA, Kinzler KW (март 2013 г.). «Ландшафты генома рака» . обзор. Наука . 339 (6127): 1546–58. BIBCODE : 2013SCI ... 339.1546V . doi : 10.1126/science.1235122 . PMC 3749880 . PMID 23539594 .
- ^ Шен Л., Кондо Y, Роснер Г.Л., Сяо Л., Эрнандес Н.С., Вилайтонг Дж., Хулихан П.С., Круз Р.С., Прасад А.Р., Эйнспар Дж. Г., Бакмейер Дж., Альбертс Д.С., Гамильтон С.Р., Исса Дж.П. (сентябрь 2005 г.). «Метилирование промотора MGMT и дефект поля при спорадическом колоректальном раке» . Журнал Национального института рака . 97 (18): 1330–8. doi : 10.1093/jnci/dji275 . PMID 16174854 .
- ^ Подпрыгнуть до: а беременный Ли К.Х., Ли Дж.С., Нам Дж. Х., Чой С., Ли М.К., Парк К.С., Юнг С.В., Ли Дж. Х. (октябрь 2011 г.). «Статус метилирования промотора генов HMLH1, HMSH2 и MGMT при колоректальном раке, связанных с последовательности аденомы-карциномы». Архив хирургии Лангенбека . 396 (7): 1017–26. doi : 10.1007/s00423-011-0812-9 . PMID 21706233 . S2CID 8069716 .
- ^ Svrcek M, Buhard O, Colas C, Coulet F, Dumont S, Massaoudi I, et al. (Ноябрь 2010). «Устойчивость к метилированию из-за дефекта поля O6-метилгуаниновой ДНК-метилтрансферазы (MGMT) в слизистой оболочке толстой кишки: инициирующая стадия развития несоответствующих раковых заболеваний с дефицитом восстановления несоответствия». Кишечник 59 (11): 1516–26. doi : 10.1136/gut.2009.194787 . PMID 20947886 . S2CID 206950452 .
- ^ Подпрыгнуть до: а беременный в дюймовый Facista A, Nguyen H, Lewis C, Prasad AR, Ramsey L, Zaitlin B, Nfonsam V, Krouse RS, Bernstein H, Payne CM, Stern S, Oatman N, Banerjee B, Bernstein C (апрель 2012 г.). «Недостаточная экспрессия ферментов репарации ДНК в раннем прогрессировании с спорадическим раком толстой кишки» . Целостность генома . 3 (1): 3. DOI : 10.1186/2041-9414-3-3 . PMC 3351028 . PMID 22494821 .
- ^ Paluszczak J, Misiak P, Wierzbicka M, Woźniak A, Baer-Dubska W (февраль 2011 г.). «Частое гиперметилирование DAPK, Rarbeta, Mgmt, Rass1a и FHIT в карциномах плоскоклеточных карт гортано Устная онкология 47 (2): 104–7 Doi : /j.oralocology.2010.11.0 10.1016 21147548PMID
- ^ Zuo C, Zhang H, Spencer HJ, Vural E, Suen JY, Schichman SA, Smoller BR, Kokoska MS, Fan Cy (октябрь 2009 г.). «Повышенная нестабильность микросателлита и эпигенетическая инактивация гена HMLH1 при плоскоклеточной карциноме головы и шеи». Отоларингология - голова и хирургия шеи . 141 (4): 484–90. doi : 10.1016/j.othns.2009.07.007 . PMID 19786217 . S2CID 8357370 .
- ^ Tawfik HM, El-Maqsoud NM, Hak BH, El-Sherbiny YM (2011). «Плоскоклеточная карцинома головы и шеи: репарация несоответствия иммуногистохимии и гиперметилирование гена HMLH1 HMLH1». Американский журнал отоларингологии . 32 (6): 528–36. doi : 10.1016/j.amjoto.2010.11.005 . PMID 21353335 .
- ^ Zou XP, Zhang B, Zhang XQ, Chen M, Cao J, Liu WJ (ноябрь 2009 г.). «Промоторное гиперметилирование нескольких генов при ранней желудочной аденокарциноме и предраковых поражениях». Человеческая патология . 40 (11): 1534–42. doi : 10.1016/j.humpath.2009.01.029 . PMID 19695681 .
- ^ Wani M, Afroze D, Makhdoomi M, Hamid I, Wani B, Bhat G, Wani R, Wani K (2012). «Статус метилирования промотора гена репарации ДНК (HMLH1) у пациентов с раком желудка в долине Кашмира» (PDF) . Азиатско -Тихоокеанский журнал профилактики рака . 13 (8): 4177–81. doi : 10.7314/apjcp.2012.13.8.4177 . PMID 23098428 .
- ^ Агарвал А., Полинени Р., Хусейн З, Вигода I, Бхагат Т.Д., Бхаттачарья С., Майтра А., Верма А. (2012). «Роль эпигенетических изменений в патогенезе пищевода Барретта и аденокарциномы пищевода» . Международный журнал клинической и экспериментальной патологии . 5 (5): 382–96. PMC 3396065 . PMID 22808291 . Обзор.
- ^ Hofstad B, Vatn MH, Andersen SN, Huitfeldt HS, Rognum T, Larsen S, Osnes M (сентябрь 1996 г.). «Рост колоректальных полипов: переопределение и оценка неразрываемых полипов в течение трех лет» . Кишечник 39 (3): 449–56. doi : 10.1136/gut.39.3.449 . PMC 1383355 . PMID 8949653 .
- ^ Schmitt MW, Prindle MJ, Loeb LA (сентябрь 2012 г.). «Последствия генетической гетерогенности при раке» . Анналы нью -йоркской академии наук . 1267 (1): 110–6. BIBCODE : 2012NASA1267..110S . doi : 10.1111/j.1749-6632.2012.06590.x . PMC 3674777 . PMID 22954224 .
- ^ Ландер Э.С., Линтон Л.М., Биррен Б., Нусбаум С., Зоди М.К., Болдуин Дж. И др. (Февраль 2001 г.). «Первоначальное секвенирование и анализ генома человека» . Природа . 409 (6822): 860–921. Bibcode : 2001natur.409..860L . doi : 10.1038/35057062 . HDL : 2027.42/62798 . PMID 11237011 .
- ^ Йост С.Е., Смит Э.Н., Шваб Р.Б., Бао Л., Юнг Х, Ван Х, Воэст Е., Пирс Дж.П., Мессер К., Паркер Б.А., Харизенди О., Фрейзер Ка (август 2012 г.). «Идентификация соматических мутаций с высокой достоверностью в последовательности всего генома фиксированных формалиновых образцов рака молочной железы» . Исследование нуклеиновых кислот . 40 (14): E107. doi : 10.1093/nar/gks299 . PMC 3413110 . PMID 22492626 .
- ^ Berger MF, Hodis E, Heffernan TP, Deribe YL, Lawrence MS, Protopopov A, et al. (Май 2012 г.). «Секвенирование генома меланомы выявляет частые мутации prex2» . Природа . 485 (7399): 502–6. Bibcode : 2012natur.485..502b . doi : 10.1038/nature11071 . PMC 3367798 . PMID 22622578 .
- ^ Rasnick D, Duesberg PH (июнь 1999 г.). «Как анеуплоидия влияет на метаболический контроль и вызывает рак» . Биохимический журнал . 340 (3): 621–30. doi : 10.1042/0264-6021: 3400621 . PMC 1220292 . PMID 10359645 .
- ^ Подпрыгнуть до: а беременный в Лопес-Лазаро М (март 2010 г.). «Новый взгляд на канцерогенез и альтернативный подход к терапии рака» . Молекулярная медицина . 16 (3–4): 144–53. doi : 10.2119/molmed.2009.00162 . PMC 2802554 . PMID 20062820 .
- ^ Juutilainen J, Herrala M, Luukkonen J, Naarala J, Hore PJ (май 2018). «Магнитокарциногенез: существует ли механизм канцерогенных эффектов слабых магнитных полей?» Полем Разбирательство. Биологические науки . 285 (1879): 20180590. DOI : 10.1098/rspb.2018.0590 . PMC 5998098 . PMID 29794049 .
- ^ Soto Am, Sonnenschein C (октябрь 2004 г.). «Теория рака соматических мутаций: растущие проблемы с парадигмой?». Биологии . 26 (10): 1097–107. doi : 10.1002/bies.20087 . PMID 15382143 .
- ^ Davies PC, LineWeaver CH (февраль 2011 г.). «Опухоли рака как метазоа 1.0: постукивание генов древних предков» . Физическая биология . 8 (1): 015001. BIBCODE : 2011 PHBIO ... 8A5001D . doi : 10.1088/1478-3975/8/1/015001 . PMC 3148211 . PMID 21301065 .
- ^ Дин, Тим. «Рак напоминает жизнь 1 миллиард лет назад, говорят астробиологи» , австралийский ученый , 8 февраля 2011 года. Получено 15 февраля 2011 года.
- ^ Sterrer, W (август 2016 г.). «Рак - мутационное воскресение прокариот эндофоссил» (PDF) . Рак гипотезы . 1 (1): 1–15. Архивировано из оригинала (PDF) 3 марта 2022 года . Получено 8 мая 2019 года .
- ^ Подпрыгнуть до: а беременный Nowell PC (октябрь 1976 г.). «Клональная эволюция популяций опухолевых клеток». Наука . 194 (4260): 23–8. Bibcode : 1976sci ... 194 ... 23n . doi : 10.1126/science.959840 . PMID 959840 . S2CID 38445059 .
- ^ Подпрыгнуть до: а беременный Merlo LM, Pepper JW, Reid BJ, Maley CC (декабрь 2006 г.). «Рак как эволюционный и экологический процесс». Природные обзоры. Рак . 6 (12): 924–35. doi : 10.1038/nrc2013 . PMID 17109012 . S2CID 8040576 .
- ^ Ханахан Д., Вайнберг Р.А. (январь 2000 г.). «Признаки рака» . Клетка . 100 (1): 57–70. doi : 10.1016/s0092-8674 (00) 81683-9 . PMID 10647931 .
- ^ CHO RW, Clarke MF (февраль 2008 г.). «Недавние достижения в стволовых клетках рака». Текущее мнение в области генетики и развития . 18 (1): 48–53. doi : 10.1016/j.gde.2008.01.017 . PMID 18356041 .
- ^ Танигучи К., Ву Л.В., Гривеников С.И., Де Йонг П.Р., Лиан И., Ю Ф.н., Ван К., Хо С.Б., Боланд Б.С., Чанг Дж.Т., Сандборн В.Дж., Хардиман Г., Раз Э, Маэхара Ю., Йошимура А., Зукман-Росси Дж., Гуан Кл, Карин М (март 2015). «Модуль GP130-SRC-YAP связывает воспаление с эпителиальной регенерацией» . Природа . 519 (7541): 57–62. Bibcode : 2015natur.519 ... 57t . doi : 10.1038/nature14228 . PMC 4447318 . PMID 25731159 .
- ^ You H, Lei P, Andreadis St (декабрь 2013 г.). «JNK - новый регулятор межклеточной адгезии» . Тканевые барьеры . 1 (5): E26845. doi : 10.4161/tisb.26845 . PMC 3942331 . PMID 24868495 .
- ^ Busillo JM, Azzam KM, Cidlowski JA (ноябрь 2011). «Глюкокортикоиды сенсибилизируют врожденную иммунную систему посредством регуляции воспаления NLRP3» . Журнал биологической химии . 286 (44): 38703–13. doi : 10.1074/jbc.m111.275370 . PMC 3207479 . PMID 21940629 .
- ^ Wang Y, Bugatti M, Ulland TK, Vermi W, Gilfillan S, Colonna M (март 2016 г.). «Недостативные роли IL-34, полученных из кератиноцитов и CSF1, полученного из нейтрофилов в обновлении клеток Лангерганса в устойчивом состоянии и во время воспаления» . Европейский журнал иммунологии . 46 (3): 552–9. doi : 10.1002/eji.201545917 . PMC 5658206 . PMID 26634935 .
- ^ Siqueira Mietto B, Kroner A, Girolami EI, Santos-Nogueira E, Zhang J, David S (декабрь 2015 г.). «Роль IL-10 в разрешении воспаления и функционального выздоровления после периферического нервного повреждения» . Журнал нейробиологии . 35 (50): 16431–42. Doi : 10.1523/jnerosci.2119-15.2015 . PMC 6605511 . PMID 26674868 .
- ^ Seifert AW, Maden M (2014). «Новое понимание регенерации кожи позвоночных». Международный обзор клеточной и молекулярной биологии . Тол. 310. С. 129–69. doi : 10.1016/b978-0-12-800180-6.00004-9 . ISBN 978-0-12-800180-6 Полем PMID 24725426 . S2CID 22554281 .
- ^ Kwon MJ, Shin HY, Cui Y, Kim H, Thi AH, Choi Jy, Kim EY, Hwang DH, Kim BG (декабрь 2015). «CCL2 опосредует нейрон-макрофаги взаимодействия для управления прооргенеративной активацией макрофагов после повреждения предварительного кондиционирования» . Журнал нейробиологии . 35 (48): 15934–47. doi : 10.1523/jneurosci.1924-15.2015 . PMC 6605453 . PMID 26631474 .
- ^ Hajishengallis G, Chavakis T (январь 2013 г.). «Эндогенные модуляторы рекрутирования воспалительных клеток» . Тенденции в иммунологии . 34 (1): 1–6. doi : 10.1016/j.it.2012.08.003 . PMC 3703146 . PMID 22951309 .
- ^ Нельсон А.М., Кацефф А.С., Ратлифф Т.С., Гарза Л.А. (февраль 2016 г.). «Интерлейкин 6 и STAT3 регулируют экспрессию изоформы p63 в кератиноцитах во время регенерации» . Экспериментальная дерматология . 25 (2): 155–7. doi : 10.1111/exd.12896 . PMC 4724264 . PMID 26566817 .
- ^ Vidal PM, Lemmens E, Dooley D, Hendrix S (февраль 2013 г.). «Роль« противовоспалительных »цитокинов в регенерации аксонов». Обзоры цитокинов и факторов роста . 24 (1): 1–12. doi : 10.1016/j.cytogfr.2012.08.008 . PMID 22985997 .
- ^ Hsueh Yy, Chang YJ, Huang CW, Handayani F, Chiang YL, Fan SC, Ho CJ, Kuo YM, Yang SH, Chen YL, Lin SC, Huang CC, Wu CC (октябрь 2015). «Синергия эндотелиальных и нервных клеток-предшественников из жировых стволовых клеток, полученных из жировых стволовых клеток, для сохранения нейросулярных структур в гипоксическом повреждении головного мозга крысы» . Научные отчеты . 5 : 14985. Bibcode : 2015natsr ... 514985h . doi : 10.1038/srep14985 . PMC 4597209 . PMID 26447335 .
- ^ Янив М (сентябрь 2014 г.). «Реконструирование хроматина: от транскрипции к раку». Рак генетика . 207 (9): 352–7. doi : 10.1016/j.cancergen.2014.03.006 . PMID 24825771 .
- ^ Zhang X, He N, Gu D, Wickliffe J, Salazar J, Boldogh I, Xie J (октябрь 2015 г.). «Генетические доказательства взаимодействия XPC-KRAS во время развития рака легких» . Журнал генетики и геномики = Yi Chuan Xue Bao . 42 (10): 589–96. doi : 10.1016/j.jgg.2015.09.006 . PMC 4643398 . PMID 26554912 .
- ^ Dubois-Pot-Schneider H, Fekir K, Coulouarn C, Glaise D, Aninat C, Jarnouen K, Le Guével R, Kubo T, Ishida S, Morel F, Corlu A (декабрь 2014 г.). «Воспалительные цитокины способствуют ретродифференцировке гепатоцит-подобных клеток, полученных из опухоли, в клетки-предшественники». Гепатология . 60 (6): 2077–90. doi : 10.1002/hep.27353 . PMID 25098666 . S2CID 11182192 .
- ^ Finkin S, Yuan D, Stein I, Taniguchi K, Weber A, Unger K, et al. (Декабрь 2015). «Эктопические лимфоидные структуры функционируют как микрониши для клеток -предшественников опухоли при гепатоцеллюлярной карциноме» . Природа иммунология . 16 (12): 1235–44. doi : 10.1038/ni.3290 . PMC 4653079 . PMID 26502405 .
- ^ Beane JE, Mazzilli SA, Campbell JD, Duclos G, Krysan K, Moy C, et al. (Апрель 2019). «Молекулярное подтипирование выявляет иммунные изменения, связанные с прогрессированием бронхиальных предварительных поражений» . Природная связь . 10 (1): 1856. Bibcode : 2019natco..10.1856b . doi : 10.1038/s41467-019-09834-2 . PMC 6478943 . PMID 31015447 .
- ^ Маус А., Меренштейн С., Кога Ю., Поттер А., Гауэр А.К., Лю Г. и др. (Сентябрь 2021 г.). «Повышенное разнообразие репертуара Т -клеток связано с прогрессированием плоскоклеточных поражений легких» . Журнал для иммунотерапии рака . 9 (9): E002647. doi : 10.1136/jitc-2021-002647 . PMC 8477334 . PMID 34580161 .
- ^ Подпрыгнуть до: а беременный Vlahopoulos SA, Cen O, Hengen N, Agan J, Moschovi M, Critselis E, Adamaki M, Bacopoulou F, Copland JA, Boldogh I, Karin M, Chrousos GP (август 2015). «Динамический аберрантный NF-κB-термогенез: новая модель, охватывающая микроокружение» . Обзоры цитокинов и факторов роста . 26 (4): 389–403. doi : 10.1016/j.cytogfr.2015.06.001 . PMC 4526340 . PMID 26119834 .
- ^ Гривеников С.И., Карин М (февраль 2010 г.). «Опасные связи: сотрудничество и перекрестные помехи STAT3 и NF-Kappab в раке» . Обзоры цитокинов и факторов роста . 21 (1): 11–9. doi : 10.1016/j.cytogfr.2009.11.005 . PMC 2834864 . PMID 20018552 .
- ^ Ригер С., Чжао Х, Мартин П., Абэ К, Лисс Т.С. (январь 2015). «Роль рецепторов ядерного гормона в восстановлении кожной раны» . Клеточная биохимия и функция . 33 (1): 1–13. doi : 10.1002/cbf.3086 . PMC 4357276 . PMID 25529612 .
- ^ Lu X, Yarbrough WG (февраль 2015 г.). «Негативная регуляция фосфорилирования RELA: новых игроков и их роль в раке». Обзоры цитокинов и факторов роста . 26 (1): 7–13. doi : 10.1016/j.cytogfr.2014.09.003 . PMID 25438737 .
- ^ Сион Р.В., Фридлендер З.Г., Гранот Z (декабрь 2015). «Многогранные роли нейтрофилы играют в микроокружении опухоли» . Раковая микроокружение . 8 (3): 125–58. doi : 10.1007/s12307-014-0147-5 . PMC 4714999 . PMID 24895166 .
- ^ Вентури, Себастьяно (2011). «Эволюционная значимость йода». Текущая химическая биология . 5 (3): 155–162. doi : 10.2174/187231311796765012 . ISSN 1872-3136 .
- ^ Вентури С. (2014). «Йод, PUFA и йодолипиды в отношении здоровья и болезней: эволюционная перспектива». Человеческая эволюция . 29 (1–3): 185–205. ISSN 0393-9375 .
- ^ Уолш К.Дж., Люер К.А., Бодин А.Б., Смит К.А., Кокс Х.Л., Нойес Д.Р., Маура Г. (декабрь 2006 г.). «Иммунные клетки Elasmobranch как источник новых ингибиторов опухолевых клеток: последствия для общественного здравоохранения» . Интегративная и сравнительная биология . 46 (6): 1072–1081. doi : 10.1093/ICB/ICL041 . PMC 2664222 . PMID 19343108 .
- ^ Vogelstein B, Kinzler KW (август 2004 г.). «Гены рака и пути, которые они контролируют». Природная медицина . 10 (8): 789–99. doi : 10.1038/nm1087 . PMID 15286780 . S2CID 205383514 .
- ^ Бренд К.А., Гермфисс У (апрель 1997 г.). «Аэробный гликолиз путем пролиферирующих клеток: защитная стратегия против активных форм кислорода» . FASEB Journal . 11 (5): 388–95. doi : 10.1096/fasebj.11.5.9141507 . PMID 9141507 . S2CID 16745745 .
- ^ Bos JL (сентябрь 1989 г.). «Ras Oncogenes при раке человека: обзор» . РАНКА . 49 (17): 4682–9. PMID 2547513 .
- ^ Чанг Э.Х., Форт я, Сколник Эм, Лоуи Д.Р. (июнь 1982 г.). «Онкогенная трансформация клеток млекопитающих, индуцированных нормальным геном человека, гомологичным онкогена вируса саркомы Харви мыши». Природа . 297 (5866): 479–83. Bibcode : 1982natur.297..479c . doi : 10.1038/297479a0 . PMID 6283358 . S2CID 4234621 .
- ^ Zhan H, Bhattacharya S, Cai H, Iglesias PA, Huang CH, Devreotes PN (сентябрь 2020 г.). «Вовлеченная сигнальная сеть RAS/PI3K/ERK контролирует миграцию и онкогенную трансформацию в эпителиальных клетках» . Ячейка развития . 54 (5): 608–623. doi : 10.1016/j.devcel.2020.08.001 . PMC 7505206 . PMID 32877650 .
- ^ Vlahopolos SA, Logotheti S, Mikas D, Girika A, Gorgoulis V, Zoumpourlis V (апрель 2008 г.). «Роль ATF-2 в онкогенезе» Биссайс 30 (4): 314–2 Doi : 10.1002/Бис . PMID 18348191 678541S2CID
- ^ Matoba S, Kang JG, Patino WD, Wragg A, Boehm M, Gavrilova O, Hurley PJ, Bunz F, Hwang PM (июнь 2006 г.). «P53 регулирует митохондриальное дыхание». Наука . 312 (5780): 1650–3. Bibcode : 2006sci ... 312.1650M . doi : 10.1126/science.1126863 . PMID 16728594 . S2CID 36668814 .
- ^ Knudson AG (апрель 1971 г.). «Мутация и рак: статистическое исследование ретинобластомы» . Труды Национальной академии наук Соединенных Штатов Америки . 68 (4): 820–3. Bibcode : 1971pnas ... 68..820K . doi : 10.1073/pnas.68.4.820 . PMC 389051 . PMID 5279523 .
- ^ Fodde R, Smits R (октябрь 2002 г.). «Биология рака. Вопрос дозировки». Наука . 298 (5594): 761–3. doi : 10.1126/science.1077707 . PMID 12399571 . S2CID 50724 .
- ^ Stephens PJ, Greenman CD, Fu B, Yang F, Bignell GR, Mudie LJ, et al. (Январь 2011). «Массовая геномная перестройка, приобретенная в одном катастрофическом событии во время развития рака» . Клетка . 144 (1): 27–40. doi : 10.1016/j.cell.2010.11.055 . PMC 3065307 . PMID 21215367 .
- Сводка на: Уэйд N (10 января 2011 г.). «Рак может развиваться в катастрофическом взрыве» . New York Times .
- ^ Kuipers EJ (март 1999 г.). «Обзорная статья: Изучение связи между Helicobacter pylori и раком желудка» . Пищевая фармакология и терапия . 13 (Suppl 1): 3–11. doi : 10.1046/j.1365-2036.1999.00002.x . PMID 10209681 .
- ^ Kusters JG, Van Vliet AH, Kuipers EJ (июль 2006 г.). «Патогенез инфекции Helicobacter pylori» . Клин Микробиол. Преподобный 19 (3): 449–90. doi : 10.1128/cmr.00054-05 . PMC 1539101 . PMID 16847081 .
- ^ Паркин Д.М. (июнь 2006 г.). «Глобальное бремя здравоохранения рака, связанных с инфекцией, в 2002 году» . Инт. Дж. Рак . 118 (12): 3030–44. doi : 10.1002/ijc.21731 . PMID 16404738 .
- ^ Wroblewski LE, Peek RM, Wilson KT (октябрь 2010). «Helicobacter pylori и рак желудка: факторы, которые модулируют риск заболевания» . Клин Микробиол. Преподобный 23 (4): 713–39. doi : 10.1128/cmr.00011-10 . PMC 2952980 . PMID 20930071 .
- ^ Ferlay J, Colombet M, Soerjomataram I, Mathers C, Parkin DM, Piñeros M, Znaor A, Bray F (апрель 2019). «Оценка глобальной заболеваемости и смертности в 2018 году: источники и методы глобокана» . Инт. Дж. Рак . 144 (8): 1941–1953. doi : 10.1002/ijc.31937 . PMID 30350310 .
- ^ Meurer Ln, Bower DJ (апрель 2002 г.). «Управление инфекцией Helicobacter pylori». Am Fam Врач . 65 (7): 1327–36. PMID 11996414 .
- ^ Прабху С.Р., Ранганатан С., Амарапуркар Д.Н. (ноябрь 1994 г.). «Helicobacter pylori в нормальной слизистой оболочке желудка». J Assoc Physicians India . 42 (11): 863–4. PMID 7868485 .
- ^ White Jr, Winter JA, Robinson K (2015). «Дифференциальный воспалительный ответ на инфекцию Helicobacter pylori: этиология и клинические результаты» . J Plantamm Res . 8 : 137–47. doi : 10.2147/jir.s64888 . PMC 4540215 . PMID 26316793 .
- ^ Денг Джи, Лян Х (апрель 2014 г.). «Клиническая значимость метастазирования лимфатических узлов при раке желудка» . World J. Gastroenterol . 20 (14): 3967–75. doi : 10.3748/wjg.v20.i14.3967 . PMC 3983452 . PMID 24744586 .
- ^ Подпрыгнуть до: а беременный Валенсуэла М.А., Каналес Дж., Корвалан А.Х., Quest AF (декабрь 2015 г.). «Хеликобактер-индуцированное воспаление и эпигенетические изменения во время канцерогенеза желудка» . World J. Gastroenterol . 21 (45): 12742–56. doi : 10.3748/wjg.v21.i45.12742 . PMC 4671030 . PMID 26668499 .
- ^ Раза Й., Хан А., Фаруки А., Мубарак М., Фасиста А., Ахтар С.С., Хан С., Кази Джи, Бернштейн С., Казми Су (2014). «Окислительное повреждение ДНК как потенциальный ранний биомаркер геликобактерного пилори, связанный с канцерогенезом». Патоль Онкол. Резерв 20 (4): 839–46. doi : 10.1007/s12253-014-9762-1 . PMID 24664859 . S2CID 18727504 .
- ^ Koeppel M, Garcia-Alcalde F, Glowinski F, Schlaermann P, Meyer TF (июнь 2015 г.). «Helicobacter pylori инфекция вызывает характерные паттерны повреждения ДНК в клетках человека» . Камерный реп . 11 (11): 1703–13. doi : 10.1016/j.celrep.2015.05.030 . PMID 26074077 .
- ^ Подпрыгнуть до: а беременный Muhammad JS, Eladl MA, Ходер G (февраль 2019 г.). «Хеликобактер-индуцированное пилори-метилированием ДНК в качестве эпигенетического модулятора рака желудка: последние результаты и будущее направление» . Патогены . 8 (1): 23. doi : 10.3390/pathogens8010023 . PMC 6471032 . PMID 30781778 .
- ^ Подпрыгнуть до: а беременный Noto JM, Peek RM (2011). «Роль микроРНК в патогенезе Helicobacter pylori и канцерогенез желудка» . Микробиол передних клеток . 1 : 21. doi : 10.3389/fcimb.2011.00021 . PMC 3417373 . PMID 22919587 .
- ^ Santos JC, Ribeiro ML (август 2015 г.). «Эпигенетическая регуляция механизма восстановления ДНК в хилеобактерном пило-индуцированном желудочном канцерогенезе» . World J. Gastroenterol . 21 (30): 9021–37. doi : 10.3748/wjg.v21.i30.9021 . PMC 4533035 . PMID 26290630 .
- ^ Подпрыгнуть до: а беременный Раза Й., Ахмед А., Хан А., Чишти А.А., Ахтер С.С., Мубарак М., Бернштейн С., Зейтлин Б., Казми Су (февраль 2020 г.). «Helicobacter pylori строго снижает экспрессию белков, резидентных ДНК, PMS2 и ERCC1 при гастрите и раке желудка» . Репарация ДНК (Amst.) . 89 : 102836. DOI : 10.1016/j.dnarep.2020.102836 . PMID 32143126 .
- ^ Dore MP, PES GM, Bassotti G, USAI-Satta P (2016). «Диспепсия: когда и как проверить на инфекцию Helicobacter pylori» . Гастроэнтерол Res Pract . 2016 : 8463614. DOI : 10.1155/2016/8463614 . PMC 4864555 . PMID 27239194 .
- ^ Borchsenius, Sn; Дакс, а; Федорова, о; Чернова, о; Barlev, NA (2018). «Влияние микоплазмы на реакцию организма хозяина посредством передачи сигналов p53/nf-κB». Журнал клеточной физиологии . 234 (1): 171–180. doi : 10.1002/jcp.26781 . PMID 30146800 .
- ^ Carrillo-Infante C, Abbadessa G, Bagella L, Giordano A (июнь 2007 г.). «Вирусные инфекции как причина рака (обзор)» . Международный журнал онкологии . 30 (6): 1521–8. doi : 10.3892/ijo.30.6.1521 . PMID 17487374 .
- ^ Сафдар А (2011). Лечение инфекций у больных раком . Спрингер. п. 478. ISBN 978-1-60761-643-6 .
- ^ Samaras V, Rafailidis Pi, Mourtzoukou EG, Peppas G, Falagas ME (июнь 2010 г.). «Хронические бактериальные и паразитические инфекции и рак: обзор» . Журнал инфекции в развивающихся странах . 4 (5): 267–81. doi : 10.3855/jidc.819 . PMID 20539059 .
- ^ Даниэль Ф.И., Черубини К., Юргель Л.С., де Фигейредо М.А., Салум Ф.Г. (февраль 2011 г.). «Роль эпигенетической репрессии транскрипции и метилтрансферазы ДНК при раке» . Рак . 117 (4): 677–87. doi : 10.1002/cncr.25482 . PMID 20945317 . Обзор.
- ^ Канвал Р., Гупта С (апрель 2012 г.). «Эпигенетические модификации при раке» . Клиническая генетика . 81 (4): 303–11. doi : 10.1111/j.1399-0004.2011.01809.x . PMC 3590802 . PMID 22082348 .
- ^ Паттани К.М., Саудри Э., Глейзер К.А., Охс М.Ф., Ван Х., Шуссель Дж., Сун В., Хеннесси П., Мидларз В., Лойо М., Демокан С., Смит И.М., Калифорния Д.А. (2012). Дао Q (ред.). «MAGEB2 активируется промотором деметилирования в плоскоклеточной карциноме головы и шеи» . Plos один . 7 (9): E45534. Bibcode : 2012ploso ... 745534p . doi : 10.1371/journal.pone.0045534 . PMC 3454438 . PMID 23029077 .
- ^ Sampath D, Liu C, Vasan K, Sulda M, Puduvalli VK, Wierda WG, Keating MJ (февраль 2012 г.). «Гистоновые деацетилазы опосредуют молчание miR-15A, miR-16 и miR-29b при хронической лимфоцитарной лейкозе» . Кровь . 119 (5): 1162–72. doi : 10.1182/blood-2011-05-351510 . PMC 3277352 . PMID 22096249 .
- ^ Хитчлер М.Дж., Оберли Л.В., Доманн Фе (декабрь 2008 г.). «Эпигенетическое молчание SOD2 путем модификаций гистонов в клетках рака молочной железы человека» . Свободная радикальная биология и медицина . 45 (11): 1573–80. doi : 10.1016/j.freeradbiomed.2008.09.005 . PMC 2633123 . PMID 18845242 .
- ^ Baldassarre G, Battista S, Belletti B, Thakur S, Pentimalli F, Trapasso F, Fedele M, Pierantoni G, Croce CM, Fusco A (апрель 2003 г.). «Негативная регуляция экспрессии гена BRCA1 белками HMGA1 объясняет сниженные уровни белка BRCA1 при спорадической карциноме молочной железы» . Молекулярная и клеточная биология . 23 (7): 2225–38. doi : 10.1128/mcb.23.7.2225-2238.2003 . PMC 150734 . PMID 12640109 .
- ^ Далерба П., Чо Р.В., Кларк М.Ф. (2007). «Стволовые клетки рака: модели и концепции». Ежегодный обзор медицины . 58 : 267–84. doi : 10.1146/annurev.med.58.062105.204854 . PMID 17002552 .
- ^ Zhang W, Hanks AN, Boucher K, Florell SR, Allen SM, Александр A, Brash De, Grossman D (январь 2005 г.). «Апоптоз, индуцированный UVB, стимулирует клональную экспансию во время развития опухоли кожи» . Канцерогенез . 26 (1): 249–57. doi : 10.1093/carcin/bgh300 . PMC 2292404 . PMID 15498793 .
Дальнейшее чтение
[ редактировать ]- Tokar EJ, Benbrahim-Tallaa L, Waalkes MP (2011). «Чептер 14. ионы металлов в развитии рака человека». В Sigel A, Sigel H, Sigel Rk (Eds.). Ионы металлов в токсикологии: эффекты, взаимодействия, взаимозависимости . Металлические ионы в науках о жизни. Тол. 8. RSC Publishing. С. 375–401. doi : 10.1039/9781849732116-00375 . ISBN 978-1-84973-091-4 .
- Диксон К, Копрас Е (декабрь 2004 г.). «Генетические изменения и репарация ДНК в канцерогенезе человека». Семинары по биологии рака . 14 (6): 441–8. doi : 10.1016/j.semcancer.2004.06.007 . PMID 15489137 .
- Клейнсмит Л.Дж. (2006). Принципы биологии рака . Сан -Франциско: Пирсон Бенджамин Каммингс. ISBN 978-0-8053-4003-7 .
- Сарасин А (ноябрь 2003 г.). «Обзор механизмов мутагенеза и канцерогенеза». Мутационные исследования . 544 (2–3): 99–106. Bibcode : 2003mrrmr.544 ... 99S . doi : 10.1016/j.mrrev.2003.06.024 . PMID 14644312 .
- Schottenfeld D, Beebe-Dimmer JL (2005). «Достижения в области эпидемиологии рака: понимание причинных механизмов и доказательства для реализации вмешательств» . Ежегодный обзор общественного здравоохранения . 26 : 37–60. doi : 10.1146/annurev.publhealth.26.021304.144402 . PMID 15760280 .
- Tannock I, Hill R, Bristow R, Harrington L (2005). Основная наука онкологии (4 -е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-07-138774-3 .
- Wicha MS, Liu S, Dontu G (февраль 2006 г.). «Стволовые клетки рака: старая идея-сдвиг парадигмы» . РАНКА . 66 (4): 1883–90, обсуждение 1895–6. doi : 10.1158/0008-5472.can-05-3153 . PMID 16488983 .