Некодирующая РНК

Некодирующая РНК ( NCRNA ) представляет собой функциональную молекулу РНК , которая не транслируется в белок . Последовательность ДНК , из которой транскрибируется функциональная некодирующая РНК, часто называют геном РНК . Обильные и функционально важные типы некодирующих РНК, включающие передаточные РНК (ТРНК) и рибосомальные РНК а также небольшие РНК, такие как микроРНК , мирнасы , пирнасы , snornas , snrnas , exrnas , scrnas и длинные Ncrnas, такие (РРНК) , и Hotair .
Количество некодирующих РНК в геноме человека неизвестно; Тем не менее, недавние транскриптомные и биоинформационные исследования показывают, что существуют тысячи некодирующих транскриптов. [ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] Многие из недавно идентифицированных NCRNAs имеют неизвестные функции, если таковые имеются. [ 8 ] Нет единого мнения о том, сколько не кодирующей транскрипции является функциональной: некоторые считают, что большинство NCRNAs являются нефункциональной «нежелательной РНК», ложными транскрипциями, [ 9 ] [ 10 ] В то время как другие ожидают, что многие не кодирующие транскрипты имеют функции, которые должны быть обнаружены. [ 11 ] [ 12 ]
История и открытие
[ редактировать ]Нуклеиновые кислоты были впервые обнаружены в 1868 году Фридрихом Мишером , [ 13 ] и к 1939 году РНК была вовлечена в синтез белка . [ 14 ] Два десятилетия спустя Фрэнсис Крик предсказал функциональный компонент РНК, который опосредовал трансляцию ; Он рассуждал, что РНК лучше подходит для базовой пары с транскриптом мРНК, чем чистый полипептид . [ 15 ]

Первой некодирующей РНК, которая была охарактеризована аланиновая тРНК, найденная в дрожжах пекаря , ее структура была опубликована в 1965 году. [ 16 ] Чтобы получить очищенный образец аланиновой тРНК, Роберт В. Холли и соавт. использовал 140 кг коммерческих дрожжей пекаря, чтобы дать всего 1 г очищенной тРНК Земля Для анализа. [ 17 ] 80 нуклеотидной тРНК секвенировали сначала расщепляемой с помощью рибонуклеазы поджелудочной железы (продуцируя фрагменты, заканчивающиеся цитозином или уридина ), а затем с такадиастазой рибонуклеазой TL (продуцируя фрагменты, которые закончились гуанозином ). Хроматография и идентификация концов 5 'и 3', а затем помогли организовать фрагменты для установления последовательности РНК. [ 17 ] Из трех структур, первоначально предложенных для этой тРНК, [ 16 ] Структура «Cloverleaf» была независимо предложена в нескольких следующих публикациях. [ 18 ] [ 19 ] [ 20 ] [ 21 ] Cloverleaf Вторичная структура была завершена после анализа рентгеновской кристаллографии, выполненного двумя независимыми исследовательскими группами в 1974 году. [ 22 ] [ 23 ]
Рибосомальная РНК была обнаружена, после чего в начале 1980 -х годов следовала урна. С тех пор открытие новых некодирующих РНК продолжалось с Snornas , Xist , CRISPR и многими другими. [ 24 ] Недавние примечательные дополнения включают рибосвитчи и miRNA ; Открытие механизма РНКи , связанного с последним, заработало Крейг С. Мелло и Эндрю Нобелевскую премию 2006 года в области физиологии или медицины . [ 25 ]
Недавние открытия NCRNAS были достигнуты как с помощью экспериментальных, так и биоинформационных методов .
Биологические роли
[ редактировать ]Некодирующие РНК относятся к нескольким группам и участвуют во многих клеточных процессах. [ 26 ] Они варьируются от NCRNAS, имеющих центральное значение, которые сохраняются во всех или большинстве клеточных сроков до более переходных NCRNAS, специфичных для одного или нескольких близкородственных видов. Считается, что более консервативные NCRNAs являются молекулярными окаменелостями или реликвиями из последнего универсального общего предка и мира РНК , и их текущие роли остаются в основном в регуляции потока информации от ДНК к белке. [ 27 ] [ 28 ] [ 29 ]
В переводе
[ редактировать ]
Многие из консервативных, важных и обильных NCRNAs участвуют в переводе . рибонуклеопротеина Частицы (RNP), называемые рибосомами, представляют собой «фабрики», где трансляция происходит в ячейке. Рибосома состоит из более чем 60% рибосомной РНК ; Они состоят из 3 NCRNAs в прокариотах и 4 NCRNAs у эукариот . Рибосомальные РНК катализируют трансляцию нуклеотидных последовательностей в белок. Другой набор NCRNAS, переносит RNA , образует «молекулу адаптера» между мРНК и белком. H /ACA Box и C/D Box Snornas - NCRNAs, найденные в Archaea и Eukaryotes. RNASE MRP ограничен эукариотами. Обе группы NCRNA участвуют в созревании рРНК. Snornas направляют ковалентные модификации рРНК, тРНК и SNRNAS ; RNASE MRP расщепляет внутреннюю транскрибируемую проставку 1 между 18S до 5,8 с рРНК. Вездесущая NCRNA, RNASE P , является эволюционным родственником RNASE MRP. [ 31 ] РНКаза P составляет последовательности тРНК, генерируя зрелые 5'-конец тРНК, путем расщепления 5'-лидерных элементов предшественников-трнас. Другой вездесущий RNP, называемый SRP , распознает и транспортирует специфические зарождающиеся белки в эндоплазматический ретикулум у эукариот и плазматическую мембрану у прокариот . У бактерий РНК переноса-мссенджера (TMRNA) представляет собой RNP, участвующий в спасении застопорившихся рибосомов, помечая неполные полипептиды и способствуя деградации аберрантной мРНК. [ Цитация необходима ]
В РНК сплайсинг
[ редактировать ]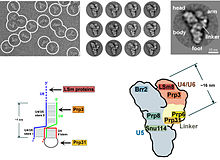
У эукариот сплайсосома выполняет реакции сплайсинга , необходимые для удаления последовательностей интрона , этот процесс необходим для образования зрелой мРНК . Сплисосома -это еще одна RNP , часто известная как SNRNP или Tri-Snrnp. Есть две разные формы сплайсосомы, основные и незначительные формы. Компонентами NCRNA основной сплайсосомы являются U1 , U2 , U4 , U5 и U6 . Компонентами NCRNA незначительной сплайсосомы являются U11 , U12 , U5 , U4ATAC и U6ATAC . [ Цитация необходима ]
Другая группа интронов может катализировать свое собственное удаление из транскриптов хозяина; Они называются самолетами РНК. Существуют две основные группы самолетов RNA: каталитический интрон I и каталитический интрон группы II . Эти NCRNAs катализируют свое собственное удаление из предшественников мРНК, тРНК и рРНК в широком диапазоне организмов. [ Цитация необходима ]
У млекопитающих было обнаружено, что Snornas также могут регулировать альтернативный сплайсинг мРНК, например, Snorna HBII-52 регулирует сплайсинг рецептора серотонина 2C . [ 32 ]
В нематодах SMY NCRNA, по-видимому, участвует в транс-распылении мРНК . [ 33 ]
В репликации ДНК
[ редактировать ]
Y RNA являются петлями STEM, необходимыми для репликации ДНК посредством взаимодействия с хроматином и белками инициации (включая комплекс распознавания происхождения ). [ 35 ] [ 36 ] Они также являются компонентами частицы рибонуклеопротеина RO60 [ 37 ] который является мишенью аутоиммунных антител у пациентов с системной волчанкой эритематозу . [ 38 ]
В регуляции генов
[ редактировать ]Экспрессия . многих тысяч генов регулируется NCRNAS Это регулирование может происходить в транс или в цис . Появляется все больше доказательств того, что особый тип NCRNAS, называемые Enhancer RNA , транскрибируется из области энхансера гена, действует для стимулирования экспрессии генов. [ Цитация необходима ]
Транс-действия
[ редактировать ]У более высоких эукариот микроРНК регулируют экспрессию генов. Одна miRNA может снизить уровни экспрессии сотен генов. Механизм, с помощью которого действуют зрелые молекулы miRNA, заключается в частичной комплементарности к одному или нескольким молекулам РНК (мРНК) мессенджера (мРНК), как правило, в 3 'UTR . Основная функция miRNAS-это подавлять экспрессию генов.
NCRNA RNase P Также было показано, что влияет на экспрессию генов. В ядре человека РНКаза P необходима для нормальной и эффективной транскрипции различных NCRNAS, транскрибируемых РНК -полимеразой III . К ним относятся тРНК, 5S рРНК , SRP РНК и гены snRNA U6 . РНКаза P оказывает свою роль в транскрипции посредством ассоциации с Pol III и хроматином активных генов тРНК и 5S рРНК. [ 39 ]
Было показано, что РНК 7SK , метазоанная NCRNA, действует как негативный регулятор РНК -полимеразы II- фактора P-TEFB , и что на эту активность влияет пути реакции на стресс. [ Цитация необходима ]
Бактериальная NCRNA, 6S РНК , в частности, ассоциируется с РНК -полимеразной голоюферментом, содержащим фактор специфичности Sigma70 . Это взаимодействие подавляет экспрессию от Sigma70-зависимого промотора во время стационарной фазы . [ Цитация необходима ]
Другая бактериальная NCRNA, Oxys РНК репрессирует трансляцию путем связывания с последовательностями Shine-Dalgarno, тем самым окклюзируя связывание рибосом. РНК Oxys индуцируется в ответ на окислительный стресс в Escherichia coli. [ Цитация необходима ]
РНК B2 представляет собой небольшую некодирующую транскрипт РНК -полимеразы III, которая подавляет транскрипцию мРНК в ответ на тепловой удар у мыши ячейки B2 РНК ингибирует транскрипцию путем связывания с ядра Pol II. Благодаря этому взаимодействию РНК B2 собирается в предварительную проживание Комплексы на промоторе и блокируют синтез РНК. [ 40 ]
Недавнее исследование показало, что только акт транскрипции последовательности NCRNA может повлиять на экспрессию генов. Транскрипция РНК -полимеразы II NCRNAS требуется для ремоделирования хроматина в Pombe Schizosaccharomyces . Хроматин постепенно преобразуется в открытую конфигурацию, так как транскрибируется несколько видов NCRNAS. [ 41 ]
Цис-действие
[ редактировать ]Ряд NCRNAs встроен в 5 ' UTR (неперечисленные области) генов, кодирующих белок , и влияют на их экспрессию различными способами. Например, рибосвитч может напрямую связывать небольшую целевую молекулу ; Связывание цели влияет на активность гена. [ Цитация необходима ]
Последовательности лидеров РНК находятся выше первого гена аминокислотных биосинтетических оперонов. Эти РНК -элементы образуют одну из двух возможных структур в областях, кодирующих очень короткие пептидные последовательности, которые богаты аминокислотой конечного продукта оперона. Структура терминатора образуется, когда существует избыток регуляторного движения аминокислоты и рибосомы над транскриптом лидера, не затрудняется. Когда существует дефицит заряженной тРНК регуляторной аминокислоты, рибосома, транслирующая лидерские пептидные киоски и образуются противотритерскую структуру. Это позволяет РНК -полимеразу транскрибировать оперон. Известными лидерами РНК являются лидер гистидинового оперона , лидер оперона лейцина , лидер треонинового оперона и лидер триптофана . [ Цитация необходима ]
Элементы ответа железа (IRE) связаны белками ответа железа (IRP). IRE встречается в UTRS различных мРНК , продукты которых участвуют в метаболизме железа . Когда концентрация железа низкая, IRP связывают гнев мРНК ферритина, что приводит к репрессии трансляции. [ Цитация необходима ]
Внутренние сайты ввода рибосом (IRES) представляют собой РНК -структуры , которые позволяют инициировать трансляцию в середине последовательности мРНК как часть процесса синтеза белка . [ Цитация необходима ]
В защите генома
[ редактировать ]Piwi-Interacting RNAS (PIRNAS), выраженные в млекопитающих Яички и соматические клетки образуют РНК-белковые комплексы с Piwi белками . Эти комплексы PIRNA (PIRC) были связаны с молчанием транскрипционных генов ретротранспозонов и других генетических элементов в клетках зародышевой линии , особенно в сперматогенезе .
Регулярно кластеризованные короткие палиндромные повторения (CRISPR) - это повторения, обнаруженные в ДНК многих бактерий и археи . Повторения разделены проставками одинаковой длины. Было продемонстрировано, что эти проставки могут быть получены из фага и впоследствии помочь защитить клетку от инфекции.
Хромосомная структура
[ редактировать ]Теломераза RNP представляет собой фермент , который добавляет специфические повторения последовательности ДНК («ttaggg» у позвоночных) к теломерным областям, которые обнаружены на концах эукариотических хромосом . Теломеры содержат конденсированный материал ДНК, обеспечивающий стабильность хромосомах. Фермент представляет собой обратную транскриптазу , которая несет РНК теломеразы , которая используется в качестве шаблона, когда она удлиняет теломеры, которые сокращаются после каждого цикла репликации .
XIST (x-неактивная транскрипция) является длинным геном NCRNA на X-хромосоме плацентарных млекопитающих , который действует как основной эффектор процесса инактивации X-хромосомы, образующих тела BARR . Антисмысловая РНК , TSIX , является отрицательным регулятором XIST. Х -хромосомы, отсутствующие экспрессии TSIX (и, следовательно, высокие уровни транскрипции XIST) инактивируются чаще, чем нормальные хромосомы. У Drosophilids , которые также используют систему определения пола XY , ROX (РНК на x) РНК участвуют в компенсации дозировки. [ 42 ] Как XIST, так и ROX работают путем эпигенетической регуляции транскрипции посредством рекрутирования гистон-модифицирующих ферментов .
Бифункциональная РНК
[ редактировать ]Бифункциональные РНК , или РНК с двойным функциями , представляют собой РНК, которые имеют две различные функции. [ 43 ] [ 44 ] Большинство известных бифункциональных РНК представляют собой мРНК, которые кодируют как белок, так и NCRNAS. Тем не менее, растущее число NCRNAS делится на две разные категории NCRNA; Например, H/ACA Box Snorna и miRNA . [ 45 ] [ 46 ]
Два известных примеров бифункциональных РНК являются SGRS RNA и RNAIII . Тем не менее, известно, что несколько других бифункциональных РНК (например, активатор стероидного рецептора/SRA, [ 47 ] Овощная РНК, [ 48 ] [ 49 ] Оскар РНК, [ 50 ] Enod40 , [ 51 ] P53 РНК [ 52 ] SR1 РНК , [ 53 ] и Spot 42 РНК . [ 54 ] ) Бифункциональные РНК были предметом специального выпуска биохими в 2011 году . [ 55 ]
Как гормон
[ редактировать ]Существует важная связь между определенными некодирующими РНК и контролем гормоновых путей. У Drosophila такие гормоны, как экдизон и ювенильный гормон, могут способствовать экспрессии определенных miRNAs. Кроме того, эта регуляция происходит в различных временных точках в рамках Caenorhabditis Elegans . развития [ 56 ] У млекопитающих miR-206 является важным регулятором эстрогена -рецептор-альфа. [ 57 ]
Некодирующие РНК имеют решающее значение в развитии нескольких эндокринных органов, а также при эндокринных заболеваниях, таких как сахарный диабет . [ 58 ] В частности, в клеточной линии MCF-7 добавление 17β- эстрадиола увеличивало глобальную транскрипцию некодирующих РНК, называемых LNCRNAs вблизи активируемых эстроген генов. [ 59 ]
В патогенном избегании
[ редактировать ]C. elegans было показано, что он учится и наследует патогенное избегание после воздействия одной некодирующей РНК бактериального патогена . [ 60 ] [ 61 ]
Роли в болезнях
[ редактировать ]Как и в случае белков , мутации или дисбаланс в репертуаре NCRNA в организме могут вызвать различные заболевания.
Рак
[ редактировать ]Многие NCRNAs показывают аномальные паттерны экспрессии в раковых тканях. [ 6 ] К ним относятся miRNAs , длинные мРНК-подобные NCRNAs , [ 62 ] [ 63 ] Газ5 , [ 64 ] Snord50 , [ 65 ] Теломераза РНК и y rnas . [ 66 ] MiRNAs участвуют в крупномасштабной регуляции многих кодирующих белок генов, [ 67 ] [ 68 ] Y RNA важны для инициации репликации ДНК, [ 35 ] РНК теломеразы, которая служит праймером для теломеразы, RNP, который расширяет теломерные области на концах хромосомы (см. Теломеры и заболевание для получения дополнительной информации). Прямая функция длинных мРНК-подобных NCRNAS менее ясна.
мутации зародышевой линии в первичных предшественниках miR-16-1 и miR-15 Было показано, что являются гораздо более частыми у пациентов с хроническим лимфоцитарным лейкозом по сравнению с контрольными популяциями. [ 69 ] [ 70 ]
Было высказано предположение, что редкий SNP ( RS11614913 ), который перекрывает HSA-MIR-196A-2, было обнаружено, связан с немелкоклеточным раком легких . [ 71 ] Аналогичным образом, экран из 17 miRNAs, которые, как было предсказано, регулирует ряд генов, связанных с раком молочной железы, обнаружил вариации у с микроРНК MIR-17 и miR-30C-1F пациентов ; Эти пациенты были не носителями мутаций BRCA1 или BRCA2 , предоставляя возможность того, что семейный рак молочной железы может быть вызван изменением в этих miRNAs. [ 72 ] Супрессор опухоли P53 , возможно, является наиболее важным агентом в предотвращении образования и прогрессирования опухоли. Белок p53 функционирует как фактор транскрипции с решающей ролью в организации реакции клеточного стресса. В дополнение к своей решающей роли в раке, p53 участвовал в других заболеваниях, включая диабет, гибель клеток после ишемии и различные нейродегенеративные заболевания, такие как Хантингтон, Паркинсон и Альцгеймер. Исследования показали, что экспрессия p53 подвержена регуляции некодирующей РНК. [ 5 ]
Другим примером некодирующей РНК, нерегулируемой в раковых клетках, является длинная некодирующая РНК LINC00707. LINC00707 активируется и губки miRNAs в мезенхимальных стволовых клетках, полученных из костного мозга человека, [ 73 ] в гепатоцеллюлярной карциноме, [ 74 ] рак желудка [ 75 ] или рак молочной железы, [ 76 ] [ 77 ] и, таким образом, способствует остеогенезу, способствует прогрессированию гепатоцеллюлярной карциномы, способствует пролиферации и метастазированию, или косвенно регулирует экспрессию белков, участвующих в агрессивности рака, соответственно.
Синдром Прадера - Уилли
[ редактировать ]Было показано , что удаление 48 копий C/D Snorna Snord116 является основной причиной синдрома Прадера -Уилли . [ 78 ] [ 79 ] [ 80 ] [ 81 ] Prader-Willi-это расстройство развития, связанное с чрезмерными трудностями и обучением. Snord116 имеет потенциальные сайты-мишени в ряде генов, кодирующих белок, и может играть роль в регуляции альтернативного сплайсинга. [ 82 ]
Аутизм
[ редактировать ]Хромосомный локус, содержащий малый кластер генов генов нуклеолярной РНК, был дублируется примерно у 5% людей с аутистическими признаками . [ 83 ] [ 84 ] Модель мыши, разработанную для того, чтобы иметь дублирование кластера SNORD115, отображает аутистическое поведение. [ 85 ] Недавнее небольшое исследование посмертной ткани головного мозга продемонстрировало измененную экспрессию длинных некодирующих РНК в префронтальной коре и мозжечке аутичного мозга по сравнению с контрольной группой. [ 86 ]
Хрящ -гипоплазия
[ редактировать ]мутации в RNASE MRP Было показано, что вызывают гипоплазию хряща и хаира , заболевание, связанное с множеством симптомов, таких как короткий рост, редкие волосы, скелетные аномалии и подавленную иммунную систему, которая часто бывает среди амишей и финских . [ 87 ] [ 88 ] [ 89 ] A-to-G Лучше всего охарактеризованный вариант-это переход в нуклеотиде 70, который находится в области петли двух оснований 5 ' консервативного псевдокнота . Тем не менее, многие другие мутации в RNASE MRP также вызывают CHH.
Болезнь Альцгеймера
[ редактировать ]Антисмысловая РНК, BACE1-AS, транскрибируется от противоположной нити к BACE1 и активируется у пациентов с болезнью Альцгеймера . [ 90 ] BACE1-регулирует экспрессию BACE1, увеличивая стабильность мРНК BACE1 и генерируя дополнительную BACE1 с помощью механизма подачи после транскрипции. По тому же механизму он также повышает концентрации бета -амилоида , основной компонент старших бляшек. Концентрации BACE1-AS повышены у субъектов с болезнью Альцгеймера и у трансгенных мышей-предшественников амилоида.
miR-96 и потеря слуха
[ редактировать ]Различия в области семян зрелого miR-96 были связаны с аутосомной доминантной , прогрессирующей потерей слуха у людей и мышей. Гомозиготные мутанты мышей были глубоко глухи, не показывая кохлеарных ответов. Гетерозиготные мыши и люди постепенно теряют способность слышать. [ 91 ] [ 92 ] [ 93 ]
Митохондриальная перенос RNAS
[ редактировать ]Ряд мутаций в митохондриальных тРНК был связан с такими заболеваниями, как синдром Меласа , синдром MERRF и хроническая прогрессивная внешняя офтальмоплегия . [ 94 ] [ 95 ] [ 96 ] [ 97 ]
Различие между функциональной РНК (FRNA) и NCRNA
[ редактировать ]Ученые начали различать функциональную РНК ( FRNA ) от NCRNA, чтобы описать области функциональных на уровне РНК, которые могут или не могут быть автономными транскриптами РНК. [ 98 ] [ 99 ] [ 100 ] Это подразумевает, что FRNA (такая как рибосвитчи, элементы SECIS и другие цис-регуляторные области) не является NCRNA. Тем не менее, FRNA может также включать мРНК , так как это РНК -кодирующая белок, и, следовательно, функциональная. Кроме того, искусственно эволюционированные РНК также подпадают под термин зонтика FRNA. Некоторые публикации [ 24 ] Укажите, что NCRNA и FRNA являются почти синонимичными, однако другие указали, что большая часть аннотированных NCRNAS, вероятно, не имеет функции. [ 9 ] [ 10 ] Также было предложено просто использовать термин РНК , поскольку различие от белка, кодирующей РНК ( Мессенджер РНК ), уже дано мРНК квалификатора . [ 101 ] Это устраняет двусмысленность при обращении к геном «кодирующей некодирующей» РНК. Кроме того, может быть ряд NCRNAS, которые неверны в опубликованной литературе и наборах данных. [ 102 ] [ 103 ] [ 104 ]
Смотрите также
[ редактировать ]- Внеклеточный РНК
- Список РНК
- Структура нуклеиновой кислоты
- RFAM
- Рибосвитч
- Рибозим
- РНК, присутствующие в образцах окружающей среды
- VA (вирусная) РНК
Ссылки
[ редактировать ]- ^ Cheng J, Kapranov P, Drenkow J, Dike S, Brubaker S, Patel S, et al. (Май 2005 г.). «Транскрипционные карты 10 хромосом человека при разрешении 5-нуклеотидов». Наука . 308 (5725): 1149–1154. Bibcode : 2005sci ... 308.1149c . doi : 10.1126/science.1108625 . PMID 15790807 . S2CID 13047538 .
- ^ Birney E, Stamatoyannopoulos JA, Dutta A , Guigó R, Gingeras TR, Margulies EH, et al. (Кодирование консорциума проекта) (июнь 2007 г.). «Идентификация и анализ функциональных элементов в 1% человеческого генома с помощью пилотного проекта Encode» . Природа . 447 (7146): 799–816. Bibcode : 2007natur.447..799b . doi : 10.1038/nature05874 . PMC 2212820 . PMID 17571346 .
- ^ Thind AS, Monga I, Thakur PK, Kumari P, Dindhoria K, Krzak M, et al. (Ноябрь 2021 г.). «Демистификация новых приложений RNA-seq: применение и полезность биоинформационной методологии». Брифинги в биоинформатике . 22 (6). doi : 10.1093/bib/bbab259 . PMID 34329375 .
- ^ Washietl S, Pedersen JS, Korbel JO, Stocsits C, Gruber AR, Hackermüller J, et al. (Июнь 2007 г.). «Структурированные РНК в выбранных областях генома человека» . Исследование генома . 17 (6): 852–864. doi : 10.1101/gr.5650707 . PMC 1891344 . PMID 17568003 .
- ^ Jump up to: а беременный Моррис К.В., изд. (2012). Некодирующие РНК и эпигенетическая регуляция экспрессии генов: драйверы естественного отбора . Caister Academic Press . ISBN 978-1-904455-94-3 .
- ^ Jump up to: а беременный Shahrouki P, Larsson E (2012). "Некодирующий онкоген: случай отсутствующих доказательств ДНК?" Полем Границы в генетике . 3 : 170. DOI : 10.3389/fgene.2012.00170 . PMC 3439828 . PMID 22988449 .
- ^ Ван Бакель Х., Нислоу С., Бленкоу Б.Дж., Хьюз Т.Р. (май 2010). Эдди С.Р. (ред.). «Большинство" темной материи "связаны с известными генами» . PLOS Биология . 8 (5): E1000371. doi : 10.1371/journal.pbio.1000371 . PMC 2872640 . PMID 20502517 .
- ^ Hüttenhofer A, Schattner P, Polacek N (май 2005 г.). «Некодирующие РНК: надежда или ажиотаж?». Тенденции в генетике . 21 (5): 289–297. doi : 10.1016/j.tig.2005.03.007 . PMID 15851066 .
- ^ Jump up to: а беременный Брозиус Дж (май 2005 г.). «Не хочу, не хочу-перевозить избыток в многоклеточных эукариотах». Тенденции в генетике . 21 (5): 287–288. doi : 10.1016/j.tig.2005.02.014 . PMID 15851065 .
- ^ Jump up to: а беременный Palazzo AF, Lee ES (2015). "Некодирующая РНК: что такое функционально и что такое мусор?" Полем Границы в генетике . 6 : 2. DOI : 10.3389/fgene.2015.00002 . PMC 4306305 . PMID 25674102 .
- ^ Mattick J, Amaral P (2022). РНК, эпицентр генетической информации: новое понимание молекулярной биологии . CRC Press. ISBN 9780367623920 .
- ^ Lee H, Zhang Z, Krause HM (декабрь 2019 г.). «Длинные некодирующие РНК и повторяющиеся элементы: мусор или интимные эволюционные партнеры?» Полем Тенденции в генетике . 35 (12): 892–902. doi : 10.1016/j.tig.2019.09.006 . PMID 31662190 . S2CID 204975291 .
- ^ Дам Р (февраль 2005 г.). «Фридрих Мишер и открытие ДНК». Биология развития . 278 (2): 274–288. doi : 10.1016/j.ydbio.2004.11.028 . PMID 15680349 .
- ^ Caspersson T, Schultz J (1939). «Пентозные нуклеотиды в цитоплазме растущих тканей». Природа . 143 (3623): 602–3. Bibcode : 1939natur.143..602c . doi : 10.1038/143602c0 . S2CID 4140563 .
- ^ Крик Ф.Х. (1958). «О синтезе белка». Симпозии Общества экспериментальной биологии . 12 : 138–163. PMID 13580867 .
- ^ Jump up to: а беременный Холли Р.В., Апгар Дж., Эверет Г.А., Мэдисон Дж. Т., Маркизи М., Меррилл Ш. С. и др. (Март 1965 г.). «Структура рибонуклеиновой кислоты». Наука . 147 (3664): 1462–1465. Bibcode : 1965sci ... 147.1462H . doi : 10.1126/science.147.3664.1462 . PMID 14263761 . S2CID 40989800 .
- ^ Jump up to: а беременный «Нобелевская премия по физиологии или медицине 1968» . Нобелевский фонд . Получено 2007-07-28 .
- ^ Мэдисон Дж. Т., Эверет Дж., Кунг Х (июль 1966 г.). «Нуклеотидная последовательность дрожжевой РНК -переноса тирозина». Наука . 153 (3735): 531–534. Bibcode : 1966sci ... 153..531m . Citeseerx 10.1.1.1001.2662 . doi : 10.1126/science.153.3735.531 . PMID 5938777 . S2CID 9265016 .
- ^ Захау Х.Г., Дюттинг Д., Фельдманн Х, Мелчерс Ф., Карау В. (1966). «Серин специфический перенос рибонуклеиновые кислоты. XIV. Сравнение нуклеотидных последовательностей и моделей вторичной структуры». Симпозии Cold Spring Harbor по количественной биологии . 31 : 417–424. doi : 10.1101/sqb.1966.031.01.054 . PMID 5237198 .
- ^ Дудок Б.С., Кац Г., Тейлор Эк, Холли Р.В. (март 1969 г.). «Первичная структура РНК переноса фенилаланина зародышевой фенилаланина» . Труды Национальной академии наук Соединенных Штатов Америки . 62 (3): 941–945. Bibcode : 1969pnas ... 62..941d . doi : 10.1073/pnas.62.3.941 . PMC 223689 . PMID 5257014 .
- ^ Cramer F, Doepner H, Haar F VD, Schlimme E, Seidel H (декабрь 1968 г.). «О конформации передачи РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 61 (4): 1384–1391. Bibcode : 1968pnas ... 61.1384c . doi : 10.1073/pnas.61.4.1384 . PMC 225267 . PMID 4884685 .
- ^ Ladner JE, Jack A, Robertus JD, Brown RS, Rhodes D, Clark BF, Klug A (ноябрь 1975 г.). «Структура дрожжевой фенилаланиновой передачи РНК при 2,5 резолюции» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (11): 4414–4418. Bibcode : 1975pnas ... 72.4414L . doi : 10.1073/pnas.72.11.4414 . PMC 388732 . PMID 1105583 .
- ^ Ким Ш., Куигли Г.Дж., Суддат Ф.Л., Макферсон А., Снеден Д., Ким Дж.Дж. и др. (Январь 1973). «Трехмерная структура дрожжевой РНК переноса фенилаланина: складывание полинуклеотидной цепи». Наука . 179 (4070): 285–288. Bibcode : 1973sci ... 179..285k . doi : 10.1126/science.179.4070.285 . PMID 4566654 . S2CID 28916938 .
- ^ Jump up to: а беременный Эдди С.Р. (декабрь 2001 г.). «Некодирующие гены РНК и современный мир РНК». Природные обзоры. Генетика . 2 (12): 919–929. doi : 10.1038/35103511 . PMID 11733745 . S2CID 18347629 .
- ^ Данехольт Б. "Расширенная информация: РНК -интерференция" . Нобелевская премия по физиологии или медицине 2006 . Архивировано из оригинала 2007-01-20 . Получено 2007-01-25 .
- ^ Монга I, Банерджи I (ноябрь 2019). «Вычислительная идентификация PIRNAS с использованием признаков, основанных на последовательности РНК, структуры, термодинамических и физико -химических свойств» . Текущая геномика . 20 (7): 508–518. doi : 10.2174/1389202920666191129112705 . PMC 7327968 . PMID 32655289 .
- ^ Джеффрес, округ Колумбия, Пул А.М., Пенни Д. (январь 1998 г.). «Реликвии из мира РНК». Журнал молекулярной эволюции . 46 (1): 18–36. Bibcode : 1998jmole..46 ... 18J . doi : 10.1007/pl00006280 . PMID 9419222 . S2CID 2029318 .
- ^ Пул А.М., Джеффрес, округ Колумбия, Пенни Д (январь 1998 г.). «Путь от мира РНК». Журнал молекулярной эволюции . 46 (1): 1–17. Bibcode : 1998jmole..46 .... 1p . doi : 10.1007/pl00006275 . PMID 9419221 . S2CID 17968659 .
- ^ Пул А., Джеффрес Д., Пенни Д (октябрь 1999 г.). «Ранняя эволюция: прокариоты, новые дети на блоке». Биологии . 21 (10): 880–889. doi : 10.1002/(SICI) 1521-1878 (199910) 21:10 <880 :: AID-bies11> 3.0.co; 2-P . PMID 10497339 . S2CID 45607498 .
- ^ Бан Н., Ниссен П., Хансен Дж., Мур П.Б., Стейц Та (август 2000). «Полная атомная структура большой рибосомальной субъединицы при 2,4 в разрешении». Наука . 289 (5481): 905–920. Bibcode : 2000sci ... 289..905b . Citeseerx 10.1.1.58.2271 . doi : 10.1126/science.289.5481.905 . PMID 10937989 .
- ^ Zhu Y, Serbinskis V, Ramos KS, L Y (май 2006 г.). «Аннализ продолжения РНК RNP выявляет . РНК 12 (5): 699–7 doi : 10.1261/РНК . 1440897PMC PMID 16540690 .
- ^ Кишор С., Стамм С (январь 2006 г.). «Snorna HBII-52 регулирует альтернативный сплайсинг рецептора серотонина 2C» . Наука . 311 (5758): 230–232. Bibcode : 2006sci ... 311..230K . doi : 10.1126/science.1118265 . PMID 16357227 . S2CID 44527461 .
- ^ Джонс, Томас А.; Отто, Вольфганг; Марз, Манджа; Эдди, Шон Р.; Stadler, Peter F. (2009). «Обзор нематоде SMY RNAS» . РНК -биология . 6 (1): 5–8. doi : 10.4161/rna.6.1.7634 . ISSN 1555-8584 . PMID 19106623 .
- ^ Stein AJ, Fuchs G, Fu C, Wolin SL, Reinisch KM (май 2005 г.). «Структурное понимание контроля качества РНК: RO Autoantigen связывает неправильно свернутые РНК через его центральную полость» . Клетка . 121 (4): 529–539. doi : 10.1016/j.cell.2005.03.009 . PMC 1769319 . PMID 15907467 .
- ^ Jump up to: а беременный Кристова К.П., Гардинер Т.Дж., Шютс Д., Круд Т (сентябрь 2006 г.). «Функциональные потребности некодирующих Y RNA для репликации хромосомной ДНК человека» . Молекулярная и клеточная биология . 26 (18): 6993–7004. doi : 10.1128/mcb.01060-06 . PMC 1592862 . PMID 16943439 .
- ^ Zhang AT, Langley AR, Christov CP, Kheir E, Shafee T, Gardiner TJ, Crude T (июнь 2011 г.). «Динамическое взаимодействие Y RNA с хроматином и белками инициации во время репликации ДНК человека» . Журнал сотовой науки . 124 (Pt 12): 2058–2069. doi : 10.1242/jcs.086561 . PMC 3104036 . PMID 21610089 .
- ^ Hall AE, Turnbull C, Dalmay T (апрель 2013 г.). «Y RNAS: последние события» . Биомолекулярные концепции . 4 (2): 103–110. doi : 10.1515/bmc-2012-0050 . PMID 25436569 . S2CID 12575326 .
- ^ Лернер М.Р., Бойл Дж.А., Хардин Дж.А., Стейц Дж.А. (январь 1981). «Два новых класса небольших рибонуклеопротеинов, обнаруженных антителами, связанными с волчанкой эритематозу». Наука . 211 (4480): 400–402. Bibcode : 1981sci ... 211..400L . doi : 10.1126/science.6164096 . PMID 6164096 .
- ^ Рейнер Р., Бен-Асули Y, Криловецки I, Jarrous N (июнь 2006 г.). «Роль каталитической рибонуклеопротеин РНКазы P в транскрипции РНК -полимеразы III» . Гены и развитие . 20 (12): 1621–1635. doi : 10.1101/gad.386706 . PMC 1482482 . PMID 16778078 .
- ^ Espinoza CA, Allen TA, Hieb AR, Kugel JF, Goodrich JA (сентябрь 2004 г.). «B2 РНК связывается непосредственно с РНК -полимеразой II, чтобы репрессировать синтез транскрипта». Природа структурная и молекулярная биология . 11 (9): 822–829. doi : 10.1038/nsmb812 . PMID 15300239 . S2CID 22199826 .
- ^ Хирота К., Миёси Т., Кугу К., Хоффман К.С., Шибата Т., Охта К (ноябрь 2008 г.). «Пошаговое ремоделирование хроматина путем каскада транскрипционной инициации некодирующих РНК». Природа . 456 (7218): 130–134. Bibcode : 2008natur.456..130H . doi : 10.1038/nature07348 . PMID 18820678 . S2CID 4416402 .
- ^ Park Y, Kelley RL, OH H, Kuroda Mi, Meller VH (ноябрь 2002 г.). «Степень распространения хроматина, определяемая рекрутированием ROX РНК белков MSL». Наука . 298 (5598): 1620–1623. Bibcode : 2002sci ... 298.1620p . doi : 10.1126/science.1076686 . PMID 12446910 . S2CID 27167367 .
- ^ Wadler CS, Vanderpool CK (декабрь 2007 г.). «Двойная функция для бактериальной малой РНК: SGR выполняет зависимую от основания регуляцию и кодирует функциональный полипептид» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (51): 20454–20459. Bibcode : 2007pnas..10420454W . doi : 10.1073/pnas.0708102104 . PMC 2154452 . PMID 18042713 .
- ^ Dinger Me, Pang KC, Mercer TR, Mattick JS (ноябрь 2008 г.). Mcentyre J (ред.). «Дифференцирование кодирования белка и некодирующая РНК: проблемы и неоднозначности» . PLOS Computational Biology . 4 (11): E1000176. BIBCODE : 2008PLSCB ... 4E0176D . doi : 10.1371/journal.pcbi.1000176 . PMC 2518207 . PMID 19043537 .
- ^ Saraiya AA, Wang CC (ноябрь 2008 г.). Голдберг де (ред.). «Снорна, новый предшественник микроРНК в Giardia Lamblia» . PLO -патогены . 4 (11): E1000224. doi : 10.1371/journal.ppat.1000224 . PMC 2583053 . PMID 19043559 .
- ^ Ender C, Krek A, Friedländer MR, Beitzinger M, Weinmann L, Chen W, et al. (Ноябрь 2008 г.). «Человеческая Snorna с микроРНК, подобными функциям» . Молекулярная клетка . 32 (4): 519–528. doi : 10.1016/j.molcel.2008.10.017 . PMID 19026782 .
- ^ Лейга Е (август 2007 г.). «РНК -активатор стероидных рецепторов (SRA1): необычные продукты генов бифаса с подозрением на рак молочной железы» . Передача сигналов ядерного рецептора . 5 : E006. doi : 10.1621/nrs.05006 . PMC 1948073 . PMID 17710122 .
- ^ Zhang J, King ML (декабрь 1996 г.). «РНК Xenopus Vegt локализуется в растительном коре во время оогенеза и кодирует новый фактор транскрипции T-BOX, участвующий в мезодермальном паттерне». Разработка . 122 (12): 4119–4129. doi : 10.1242/dev.122.12.4119 . PMID 9012531 . S2CID 28462527 .
- ^ Kloc M, Wilk K, Vargas D, Shirato Y, Bilinski S, Etkin LD (август 2005 г.). «Потенциальная структурная роль некодирующих и кодирующих РНК в организации цитоскелета в растительной коре ооцитов Xenopus» . Разработка . 132 (15): 3445–3457. doi : 10.1242/dev.01919 . PMID 16000384 .
- ^ Дженни А., Хахет О., Заворски П., Сирклафф А., Уэстон М.Д., Джонстон Д.С. и др. (Август 2006 г.). «Независимая от трансляции роль Оскар РНК в раннем дрозофиле оогенеза» . Разработка . 133 (15): 2827–2833. doi : 10.1242/dev.02456 . PMID 16835436 .
- ^ Gultyaev AP, Roussis A (2007). «Идентификация консервативных вторичных структур и сегментов расширения в РНК enod40 выявляет новые гомологи enod40 в растениях» . Исследование нуклеиновых кислот . 35 (9): 3144–3152. doi : 10.1093/nar/gkm173 . PMC 1888808 . PMID 17452360 .
- ^ Кандейайс М.М., Малберт-Колас Л., Пауэлл Д.Дж., Даскалогянни С., Маслон М.М., Наски Н. и др. (Сентябрь 2008 г.). «МРНК P53 контролирует активность p53 путем управления функциями MDM2». Природная клеточная биология . 10 (9): 1098–1105. doi : 10.1038/ncb1770 . PMID 19160491 . S2CID 5122088 .
- ^ Gimpel M, Preis H, Barth E, Gramzow L, Brantl S (декабрь 2012 г.). «SR1-небольшая РНК с двумя удивительно консервативными функциями» . Исследование нуклеиновых кислот . 40 (22): 11659–11672. doi : 10.1093/nar/gks895 . PMC 3526287 . PMID 23034808 .
- ^ Aoyama JJ, Raina M, Zhong A, Storz G (март 2022 г.). «Двойная функция Пятно 42 РНК кодирует белок с 15 аминокислотой, который регулирует фактор транскрипции СРБ» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (10): E21198661119. Bibcode : 2022pnas..11919866a . doi : 10.1073/pnas.21198661119 . PMC 8916003 . PMID 35239441 .
- ^ Francastel C, Hubé F (ноябрь 2011 г.). "Кодирование или некодирование: нужно ли они быть эксклюзивными?" Полем Биохими . 93 (11): vi - vii. doi : 10.1016/s0300-9084 (11) 00322-1 . PMID 21963143 .
- ^ Sempere LF, Sokol NS, Dubrovsky EB, Berger EM, Ambros V (июль 2003 г.). «Временная регуляция экспрессии микроРНК в Drosophila melanogaster, опосредованную гормональными сигналами и активностью гена широкого комплекса» . Биология развития . 259 (1): 9–18. doi : 10.1016/s0012-1606 (03) 00208-2 . PMID 12812784 . S2CID 17249847 .
- ^ Адамс Б.Д., Ферно Х, Белый Ба (май 2007 г.). «Микрорибонуклеиновая кислота (miRNA) miR-206 нацелен на рецептор эстрогена человека (Eralpha) и подавляет РНК Eralpha Messenger и экспрессию белка в клеточных линиях рака молочной железы» . Молекулярная эндокринология . 21 (5): 1132–1147. doi : 10.1210/me.2007-0022 . PMID 17312270 .
- ^ Knoll M, Lodish HF, Sun L (март 2015 г.). «Длинные некодирующие РНК как регуляторы эндокринной системы» . Природные обзоры. Эндокринология . 11 (3): 151–160. doi : 10.1038/nrendo.2014.229 . HDL : 1721.1/116703 . PMC 4376378 . PMID 25560704 .
- ^ Li W, Notani D, Ma Q, Tanasa B, Nunez E, Chen AY, et al. (Июнь 2013 г.). «Функциональная роль энхансерных РНК для эстрогензависимой активации транскрипции» . Природа . 498 (7455): 516–520. Bibcode : 2013natur.498..516L . doi : 10.1038/nature12210 . PMC 3718886 . PMID 23728302 .
- ^ «Исследователи обнаруживают, как черви передают знание патогена потомству» . Phys.org . Получено 11 октября 2020 года .
- ^ Калетский Р., Мур Р.С., Врла Г.Д., Парсонс Л.Р., Гитай З., Мерфи К.Т. (октябрь 2020 г.). «C. elegans интерпретирует бактериальные некодирующие РНК для изучения патогенного избегания» . Природа . 586 (7829): 445–451. Bibcode : 2020nater.586..445K . doi : 10.1038/s41586-020-2699-5 . PMC 8547118 . PMID 32908307 . S2CID 221626129 .
- ^ Pibouin L, Villaudy J, Ferbus D, Muleris M, Propéri MT, Remvikos Y, Goubin G (февраль 2002 г.). «Клонирование мРНК сверхэкспрессии в раке толстой кишки-1: последовательность сверхэкспрессирована в подмножестве карциномы толстой кишки». Генетика рака и цитогенетика . 133 (1): 55–60. doi : 10.1016/s0165-4608 (01) 00634-3 . PMID 11890990 .
- ^ Fu X, Ravindranath L, Tran N, Petrovics G, Srivastava S (март 2006 г.). «Регуляция апоптоза с помощью специфического для простаты и некодируемого гена, ассоциированного с раком простаты, PCGEM1». ДНК и клеточная биология . 25 (3): 135–141. doi : 10.1089/dna.2006.25.135 . PMID 16569192 .
- ^ Mourtada-Maarabouni M, Pickard MR, Hedge VL, Farzaneh F, Williams GT (январь 2009 г.). «GAS5, не белковой РНК, контролирует апоптоз и подавляется при раке молочной железы» . Онкоген . 28 (2): 195–208. doi : 10.1038/onc.2008.373 . PMID 18836484 .
- ^ Dong Xy, Guo P, Boyd J, Sun X, Li Q, Zhou W, Dong JT (август 2009 г.). «Смысл Snorna U50 при раке молочной железы человека» . Журнал генетики и геномики = Yi Chuan Xue Bao . 36 (8): 447–454. doi : 10.1016/s1673-8527 (08) 60134-4 . PMC 2854654 . PMID 19683667 .
- ^ Кристова К.П., Тривье Э., Круд Т (март 2008 г.). «Некодирующие человеческие RNAs сверхэкспрессируются в опухолях и требуются для пролиферации клеток» . Британский журнал рака . 98 (5): 981–988. doi : 10.1038/sj.bjc.6604254 . PMC 2266855 . PMID 18283318 .
- ^ Farh KK, Grimson A, Jan C, Lewis BP, Johnston WK, Lim LP, et al. (Декабрь 2005 г.). «Широко распространенное влияние микроРНК млекопитающих на репрессию и эволюцию мРНК». Наука . 310 (5755): 1817–1821. Bibcode : 2005sci ... 310.1817f . doi : 10.1126/science.1121158 . PMID 16308420 . S2CID 1849875 .
- ^ Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, et al. (Февраль 2005 г.). «Анализ микрочипов показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа . 433 (7027): 769–773. Bibcode : 2005natur.433..769L . doi : 10.1038/nature03315 . PMID 15685193 . S2CID 4430576 .
- ^ Калин Г.А., Феррацин М., Cimmino A, Di Leva G, Shimizu M, Wojcik SE, et al. (Октябрь 2005 г.). «Подпись микроРНК, связанная с прогнозом и прогрессированием при хронической лимфоцитарной лейкозе» . Новая Англия Журнал медицины . 353 (17): 1793–1801. doi : 10.1056/nejmoa050995 . PMID 16251535 .
- ^ Калин Г.А., Думитру К.Д., Шимизу М., Бичи Р., Зупо С., Ноч Е. и др. (Ноябрь 2002). «Частые делеции и подавляемая регуляция микромеровских генов miR15 и miR16 при 13q14 при хронической лимфоцитарной лейкозе» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (24): 15524–15529. Bibcode : 2002pnas ... 9915524c . doi : 10.1073/pnas.242606799 . PMC 137750 . PMID 12434020 .
- ^ Hu Z, Chen J, Tian T, Zhou X, Gu H, Xu L, et al. (Июль 2008 г.). «Генетические варианты последовательностей miRNA и немелкоклеточного рака легких» . Журнал клинических исследований . 118 (7): 2600–2608. doi : 10.1172/jci34934 . PMC 2402113 . PMID 18521189 .
- ^ Shen J, Ambrosone CB, Zhao H (март 2009 г.). «Новые генетические варианты в генах микроРНК и семейный рак молочной железы» . Международный журнал рака . 124 (5): 1178–1182. doi : 10.1002/ijc.24008 . PMID 19048628 . S2CID 20960029 .
- ^ Jia B, Wang Z, Sun X, Chen J, Zhao J, Qiu X (февраль 2019). «Длинные некодирующие РНК LINC00707 SPONGES MIR-370-3P, чтобы способствовать остеогенезу мезенхимальных стволовых клеток, полученных из костного мозга человека, посредством активации WNT2B» . Исследование стволовых клеток и терапия . 10 (1): 67. DOI : 10.1186/S13287-019-1161-9 . PMC 6387535 . PMID 30795799 .
- ^ Tu J, Zhao Z, Xu M, Chen M, Weng Q, Wang J, Ji J (июль 2019). «LINC00707 способствует прогрессированию гепатоцеллюлярной карциномы посредством губчатого miR-206, чтобы увеличить CDK14». Журнал клеточной физиологии . 234 (7): 10615–10624. doi : 10.1002/jcp.27737 . PMID 30488589 . S2CID 54119752 . (Втянут, см Два : 10.1002/jcp.30712 , PMID 35297045 )
- ^ Xie M, Ma T, Xue J, Ma H, Sun M, Zhang Z, et al. (Февраль 2019 г.). «Длинная межгенная небетральная кодирующая РНК 707 способствует пролиферации и метастазированию рака желудка путем взаимодействия со стабилизирующим мРНК белком Hur». Раковые письма . 443 : 67–79. doi : 10.1016/j.canlet.2018.11.032 . PMID 30502359 . S2CID 54611497 .
- ^ Li T, Li Y, Sun H (2019-06-06). «Microrna-876 обливается длинной некодирующей РНК LINC00707 и непосредственно нацелен на метадгерин, чтобы ингибировать злокачественную опухоль рака молочной железы» . Управление раком и исследования . 11 : 5255–5269. doi : 10.2147/cmar.s210845 . PMC 6559252 . PMID 31239777 .
- ^ Yuan RX, Bao D, Zhang Y (май 2020). «LINC00707 способствует пролиферации, инвазии и миграции клеток через регуляторную петлю miR-30C/CTHRC1 при раке молочной железы». Европейский обзор медицинских и фармакологических наук . 24 (9): 4863–4872. doi : 10.26355/eurrev_202005_21175 . PMID 32432749 . S2CID 218759508 .
- ^ Sahoo T, Del Gaudio D, German Jr, Shinawi M, Peters Su, Person Re, et al. (Июнь 2008 г.). «Фенотип Prader-Willi, вызванный отцовским дефицитом для HBII-85 C/D Box мелкого кластера нуклеолярной РНК» . Природа генетика . 40 (6): 719–721. doi : 10.1038/ng.158 . PMC 2705197 . PMID 18500341 .
- ^ Skryabin BV, Gubar LV, Seeger B, Pfeiffer J, Handel S, Robeck T, et al. (Декабрь 2007 г.). «Удаление кластера генов Snorna MBII-85 у мышей приводит к последующему задержке роста» . PLOS Genetics . 3 (12): E235. doi : 10.1371/journal.pgen.0030235 . PMC 2323313 . PMID 18166085 .
- ^ Ding F, Li HH, Zhang S, Solomon NM, Camper SA, Cohen P, Francke U (март 2008 г.). Акбарян С (ред.). «Делуция Snorna Snord116 (PWCR1/MBII-85) вызывает дефицит роста и гиперфагию у мышей» . Plos один . 3 (3): E1709. Bibcode : 2008ploso ... 3.1709d . doi : 10.1371/journal.pone.0001709 . PMC 2248623 . PMID 18320030 .
- ^ Ding F, Prints Y, Dhar MS, Johnson DK, Garnacho-Montero C, Nicholls Rd, Francke U (июнь 2005 г.). «Отсутствие Snorna PWCR1/MBII-85 имеет решающее значение для летальности новорожденных в моделях мыши синдрома Прадер-Вилли». Геном млекопитающих . 16 (6): 424–431. doi : 10.1007/s00335-005-2460-2 . PMID 16075369 . S2CID 12256515 .
- ^ Bazeley PS, Shepelev V, Talebizadeh Z, Butler MG, Fedorova L, Filatov V, Fedorov A (январь 2008 г.). «Сонеаржет показывает, что мишени для Snorna человека, расположенные близко к альтернативным сплайсинским соединениям» . Ген . 408 (1–2): 172–179. doi : 10.1016/j.gene.2007.10.037 . PMC 6800007 . PMID 18160232 .
- ^ Bolton PF, Veltman MW, Weisblatt E, Holmes JR, Thomas NS, Youings SA, et al. (Сентябрь 2004 г.). «Хромосома 15q11-13 Аномалии и другие заболевания у людей с расстройствами аутистического спектра». Психиатрическая генетика . 14 (3): 131–137. doi : 10.1097/00041444-200409000-00002 . PMID 15318025 . S2CID 37344935 .
- ^ Кук Эх, Шерер С.В. (октябрь 2008 г.). «Вариации копий-номера, связанные с нейропсихиатрическими условиями». Природа . 455 (7215): 919–923. Bibcode : 2008natur.455..919c . doi : 10.1038/nature07458 . PMID 18923514 . S2CID 4377899 .
- ^ Накатани Дж., Тамада К., Хатанака Ф., Исе С., Охта Х., Иноуэ К. и др. (Июнь 2009 г.). «Аномальное поведение в модели мыши, оборудованной хромосомой для дублирования человека 15Q11-13, наблюдаемое при аутизме» . Клетка . 137 (7): 1235–1246. doi : 10.1016/j.cell.2009.04.024 . PMC 3710970 . PMID 19563756 .
- ^ Ziats MN, Rennert OM (март 2013 г.). «Аберрантная экспрессия длинных некодирующих РНК в аутичном мозге» . Журнал молекулярной нейробиологии . 49 (3): 589–593. doi : 10.1007/s12031-012-9880-8 . PMC 3566384 . PMID 22949041 .
- ^ Ridanpää M, Van Eenennaam H, Pelin K, Chadwick R, Johnson C, Yuan B, et al. (Январь 2001). «Мутации в РНК-компоненте RNASE MRP вызывают плейотропное заболевание человека, гипоплазию хряща» . Клетка . 104 (2): 195–203. doi : 10.1016/s0092-8674 (01) 00205-7 . HDL : 2066/185709 . PMID 11207361 . S2CID 13977736 .
- ^ Мартин Ан, Ли Y (март 2007 г.). «РНК МРП РНК и генетические заболевания человека» . Клеточные исследования . 17 (3): 219–226. doi : 10.1038/sj.cr.7310120 . PMID 17189938 .
- ^ Kavadas FD, Giliani S, Gu Y, Mazzolari E, Bates A, Pegoiani E, et al. (Декабрь 2008 г.). «Изменчивость клинических и лабораторных особенностей у пациентов с рибонуклеазой митохондриальной РНК -обработки мутаций гена эндорибонуклеазы» . Журнал аллергии и клинической иммунологии . 122 (6): 1178–1184. doi : 10.1016/j.jaci.2008.07.036 . PMID 18804272 .
- ^ Faghihi MA, Modarresi F, Khalil AM, Wood DE, Sahagan BG, Morgan TE и др. (Июль 2008 г.). «Экспрессия некодирующей РНК повышается при болезни Альцгеймера и приводит к быстрой регуляции бета-секретазы быстрого приема пищи» . Природная медицина . 14 (7): 723–730. doi : 10.1038/nm1784 . PMC 2826895 . PMID 18587408 .
- ^ Менсия А., Менсия-Хиккибьер С. (Там 2009). Это семян. Природа генетика 41 (5): 609–613. doi : 10.1038/g.355 . PMID 19363479 . 11113852S2CID
- ^ Льюис М.А., Квинт Е., Глейзер А.М., Фукс Х., Де Анжелис М.Х., Лэнгфорд С. и др. (Май 2009 г.). «ENU-индуцированная мутация miR-96, связанная с прогрессирующей потерей слуха у мышей» . Природа генетика . 41 (5): 614–618. doi : 10.1038/ng.369 . PMC 2705913 . PMID 19363478 .
- ^ Soukup GA (июнь 2009 г.). «Маленькие, но громкие: маленькие РНК оказывают громкое влияние на развитие ушей» . Исследование мозга . 1277 : 104–114. doi : 10.1016/j.brainres.2009.02.027 . PMC 2700218 . PMID 19245798 .
- ^ Тейлор Р.В., Тернбулл Д.М. (май 2005 г.). «Митохондриальные ДНК -мутации при болезни человека» . Природные обзоры. Генетика . 6 (5): 389–402. doi : 10.1038/nrg1606 . PMC 1762815 . PMID 15861210 .
- ^ Yarham JW, Elson JL, Blakely EL, McFarland R, Taylor RW (сентябрь 2010 г.). «Митохондриальные мутации тРНК и болезнь». Wiley междисциплинарные обзоры. РНК . 1 (2): 304–324. doi : 10.1002/wrna.27 . PMID 21935892 . S2CID 43123827 .
- ^ Zifa E, Jiannouli S, Theotokis P, Stamatis C, Mamuris Z, Stathopoulos C (январь 2007 г.). «Митохондриальные мутации тРНК: клинические и функциональные возмущения» . РНК -биология . 4 (1): 38–66. doi : 10.4161/rna.4.1.4548 . PMID 17617745 . S2CID 11965790 .
- ^ Эбботт Дж.А., Франклин К.С., Роби-Бонд С.М. (2014). «Передача РНК и болезнь человека» . Границы в генетике . 5 : 158. doi : 10.3389/fgene.2014.00158 . PMC 4042891 . PMID 24917879 .
- ^ Картер Р.Дж., Дубчак I, Холбрук С.Р. (октябрь 2001 г.). «Вычислительный подход для идентификации генов для функциональных РНК в геномных последовательностях» . Исследование нуклеиновых кислот . 29 (19): 3928–3938. doi : 10.1093/nar/29.19.3928 . PMC 60242 . PMID 11574674 .
- ^ Pedersen JS, Bejerano G, Siepel A, Rosenbloom K, Lindblad-Toh K, Lander ES, et al. (Апрель 2006 г.). «Идентификация и классификация консервативных РНК вторичных структур в геноме человека» . PLOS Computational Biology . 2 (4): E33. Bibcode : 2006plscb ... 2 ... 33p . doi : 10.1371/journal.pcbi.0020033 . PMC 1440920 . PMID 16628248 .
- ^ Томас Дж. М., Хорспул Д., Браун Г., Черепанов В., Аптон С (январь 2007 г.). «ГрафДНК: Java -программа для графического отображения анализа композиции ДНК» . BMC Bioinformatics . 8 : 21. doi : 10.1186/1471-2105-8-21 . PMC 1783863 . PMID 17244370 .
- ^ Brosius J, Raabe CA (февраль 2015 г.). «Что такое РНК? Верхний слой для классификации РНК» . РНК -биология . 13 (2): 140–144. doi : 10.1080/15476286.2015.1128064 . PMC 4829331 . PMID 26818079 .
- ^ Ji Z, Song R, Regev A, Struhl K (декабрь 2015 г.). «Многие lncrnas, 5'UTR и псевдогены переводятся, а некоторые могут экспрессировать функциональные белки» . элиф . 4 : E08890. doi : 10.7554/elife.08890 . PMC 4739776 . PMID 26687005 .
- ^ Tosar JP, Rovira C, Cayota A (2018-01-22). «Некодирующие фрагменты РНК составляют большинство аннотированных PIRNAS, экспрессируемых в соматических не-гонадальных тканях» . Биология связи . 1 (1): 2. DOI : 10.1038/S42003-017-0001-7 . PMC 6052916 . PMID 30271890 .
- ^ Housman G, Ulitsky I (январь 2016 г.). «Методы различения кодирования белка и длинных некодирующих РНК и неуловимой биологической цели трансляции длинных некодирующих РНК». Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов . 1859 (1): 31–40. doi : 10.1016/j.bbagrm.2015.07.017 . PMID 26265145 .
Внешние ссылки
[ редактировать ]- "Rnadb" . Архивировано из оригинала 2007-08-29.
Эта база данных представляет собой комплексную базу данных РНК млекопитающих (RNADB)
- База данных RFAM - курируемый список сотен семей связанных NCRNAS
- Noncode.org - бесплатная база данных всех видов некодирующих РНК (кроме тРНК и рРНК)
- RNACON Прогнозирование и классификация NCRNA BMC Genomics 2014, 15: 127
- Кодировать потоки исследователя некодирующей характеристики РНК. Природа (журнал)
- Ресурс баз данных некодирующих баз данных РНК (NRDR) -курированный источник данных, связанных с некодирующими базами данных РНК, доступными в Интернете
- DASHR - база данных небольших некодирующих RNAS Bioinformatics 2018
Типы нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Избиратели | |||||||
| Рибонуклеиновые кислоты (Кодирование, не кодирование ) |
| ||||||
| Дезоксирибонуклеический кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||