Сукцинилкоэнзим А-синтетаза
| Сукцинат - КоА-лигаза (формирующая ВВП) | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 6.2.1.4 | ||
| Номер CAS. | 9014-36-2 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| Сукцинат — КоА-лигаза (АДФ-образующая) | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 6.2.1.5 | ||
| Номер CAS. | 9080-33-5 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Сукцинил-Коэнзим А-синтетаза ( SCS , также известная как сукцинил-КоА-синтетаза или сукцинат-тиокиназа или сукцинат-КоА-лигаза ) представляет собой фермент, который катализирует обратимую реакцию сукцинил-КоА в сукцинат . [3] Фермент облегчает соединение этой реакции с образованием молекулы нуклеозидтрифосфата ( GTP или АТФ ) из молекулы неорганического фосфата и молекулы нуклеозиддифосфата ( GDP или ADP ). Он играет ключевую роль в качестве одного из катализаторов, участвующих в цикле лимонной кислоты , центральном пути клеточного метаболизма , и расположен в митохондриальном матриксе клетки. [4]
Химическая реакция и ферментативный механизм
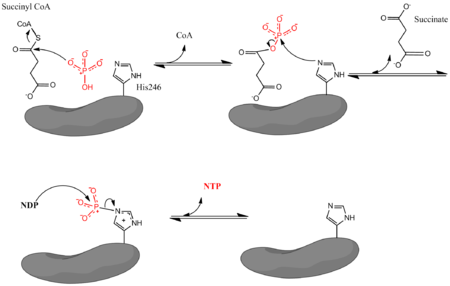
[ редактировать ]Сукцинил-КоА-синтетаза катализирует следующую обратимую реакцию :
- Сукцинил-КоА + Пи + НДП ↔ Сукцинат + КоА + NTP
где Pi обозначает неорганический фосфат, NDP обозначает нуклеотиддифосфат (GDP или ADP), а NTP обозначает нуклеотидтрифосфат (GTP или АТФ). Как уже упоминалось, фермент способствует сочетанию превращения сукцинил-КоА в сукцинат с образованием NTP из NDP и Pi. Реакция имеет биохимическое стандартное изменение свободной энергии в состоянии -3,4 кДж/моль. [4] Реакция протекает по трехстадийному механизму. [3] что изображено на изображении ниже. Первый этап включает замещение КоА из сукцинил-КоА нуклеофильной молекулой неорганического фосфата с образованием сукцинилфосфата. Затем фермент использует остаток гистидина для удаления фосфатной группы из сукцинилфосфата и образования сукцината. Наконец, фосфорилированный гистидин переносит фосфатную группу на нуклеозиддифосфат, который генерирует высокоэнергетический нуклеозидтрифосфат.
Структура
[ редактировать ]Субъединицы
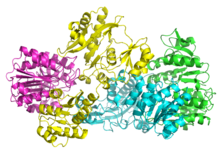
[ редактировать ]SCS бактерий и млекопитающих состоят из субъединиц α и β . [5] В E. coli αβ два гетеродимера соединяются вместе, образуя структуру α 2 β 2 гетеротетрамерную . Однако митохондриальные SCS млекопитающих активны как αβ-димеры и не образуют гетеротетрамер. [6] Гететереттрамер SCS E. coli был кристаллизован и очень подробно охарактеризован. [6] [7] Как видно на изображении 2, две субъединицы α (розовая и зеленая) расположены на противоположных сторонах структуры, а две субъединицы β (желтая и синяя) взаимодействуют в средней области белка. Две субъединицы α взаимодействуют только с одной единицей β, тогда как единицы β взаимодействуют с одной единицей α (с образованием димера αβ) и субъединицей β другого димера αβ. [6] Короткая аминокислотная цепь соединяет две β-субъединицы, образуя тетрамерную структуру.

Кристаллическая структура альфа-субъединицы сукцинил-КоА-синтетазы (сукцинил-КоА-связывающая изоформа) была определена Joyce et al. с разрешением 2,10 А, с кодом PDB 1CQJ. [1] . [8]
Каталитические остатки
[ редактировать ]Кристаллические структуры SCS E. coli доказывают, что кофермент А связывается внутри каждой α-субъединицы (внутри складки Россмана ) в непосредственной близости от остатка гистидина (His246α). [7] Этот остаток гистидина фосфорилируется на стадии образования сукцината в механизме реакции. Точное место связывания сукцината четко не определено. [9] Образование нуклеотидтрифосфата происходит в домене захвата АТФ, который расположен вблизи N-конца каждой β-субъединицы. Однако этот домен захвата расположен примерно в 35 Å от фосфорилированного остатка гистидина. [8] Это заставляет исследователей полагать, что фермент должен претерпеть серьезные изменения в конформации , чтобы перенести гистидин в домен захвата и облегчить образование нуклеозидтрифосфата. Эксперименты по мутагенезу определили, что два остатка глутамата (один рядом с каталитическим гистидином, Glu208α, и один рядом с доменом захвата АТФ, Glu197β) играют роль в фосфорилировании и дефосфорилировании гистидина, но точный механизм, с помощью которого фермент меняет конформацию, неизвестен. полностью понял. [9]
Изоформы
[ редактировать ]Джонсон и др. описывают две изоформы сукцинил-КоА-синтетазы у амниот : одна, которая отвечает за синтез АТФ, и другая, которая синтезирует ГТФ. [10]
У амниот фермент представляет собой гетеродимер α- и β-субъединицы. Специфичность аденозин- или гуанозинфосфатов определяется β-субъединицей, [10] который кодируется двумя генами. SUCLG2 специфичен для GTP, а SUCLA2 специфичен для АТФ, тогда как SUCLG1 кодирует общую α-субъединицу. β-варианты производятся в разных количествах в разных тканях. [10] вызывая потребность в субстратах GTP или АТФ .
Ткани, потребляющие в основном вещества, такие как сердце и мозг, содержат больше АТФ-специфической сукцинил-КоА-синтетазы (ATPSCS), тогда как синтетические ткани, такие как почки и печень, имеют более специфичную для GTP форму (GTPCSS). [11] Анализ кинетики ATPSCS из грудной мышцы голубей и GTPSCS из печени голубей показал, что их кажущиеся константы Михаэлиса были схожи для КоА, но различались для нуклеотидов, фосфата и сукцината. Наибольшая разница наблюдалась для сукцината: K m app ATPSCS = 5 мМ по сравнению с GTPSCS = 0,5 мМ. [10]
Функция
[ редактировать ]Генерация нуклеотидтрифосфатов
[ редактировать ]SCS — единственный фермент цикла лимонной кислоты, который катализирует реакцию, в которой нуклеотидтрифосфат (GTP или АТФ) образуется путем фосфорилирования на уровне субстрата . [4] Исследования показали, что SCS E. coli могут катализировать образование GTP или АТФ. [7] Однако млекопитающие обладают разными типами SCS, которые специфичны либо для GTP (G-SCS), либо для АТФ (A-SCS) и являются нативными для разных типов тканей организма. Интересное исследование с использованием голубиных клеток показало, что SCS, специфичные для GTP, расположены в клетках печени голубя, а SCS, специфичные для АТФ, расположены в мышечных клетках молочной железы голубя. [12] Дальнейшие исследования выявили аналогичный феномен ГТФ и АТФ-специфических SCS в тканях крыс, мышей и человека. Похоже, что ткани, обычно участвующие в анаболическом метаболизме (например, печень и почки), экспрессируют G-SCS, тогда как ткани, участвующие в катаболическом метаболизме (например, мозг, сердце и мышечная ткань), экспрессируют A-SCS. [11]
Образование промежуточных продуктов метаболизма
[ редактировать ]SCS облегчает поток молекул в другие метаболические пути , контролируя взаимное превращение сукцинил-КоА и сукцината. [13] Это важно, поскольку сукцинил-КоА является промежуточным продуктом, необходимым для образования порфирина , гема , [14] и кетоновых тел биосинтез . [15]
Регулирование и ингибирование
[ редактировать ]У некоторых бактерий фермент регулируется на уровне транскрипции. [16] Было продемонстрировано, что ген SCS (sucCD) транскрибируется вместе с геном α-кетоглутаратдегидрогеназы (sucAB) под контролем промотора sdhC , который является частью сукцинатдегидрогеназы оперона . Этот оперон активируется присутствием кислорода и реагирует на различные источники углерода. Антибактериальные препараты , предотвращающие фосфорилирование гистидина, такие как молекула LY26650, являются мощными ингибиторами бактериальных SCS. [17]
Оптимальная активность
[ редактировать ]Измерения (выполненные с использованием SCS соевых бобов) указывают на оптимальную температуру 37 °C и оптимальный pH 7,0–8,0. [18]
Роль в болезни
[ редактировать ]Фатальный младенческий лактоацидоз. Дефектный СХП считается причиной фатального детского лактоацидоза , заболевания у младенцев, характеризующегося накоплением токсических уровней молочной кислоты. Состояние (в наиболее тяжелом случае) приводит к смерти обычно в течение 2–4 дней после рождения. [19] Было установлено, что у пациентов с этим заболеванием наблюдается двух пар оснований делеция в гене, известном как SUCLG1 , который кодирует субъединицу α SCS. [19] В результате функциональный SCS отсутствует в метаболизме, что приводит к серьезному дисбалансу потоков между гликолизом и циклом лимонной кислоты. Поскольку в клетках нет функционального цикла лимонной кислоты, возникает ацидоз, поскольку клетки вынуждены выбирать выработку молочной кислоты в качестве основного средства производства АТФ.
См. также
[ редактировать ]- Цикл лимонной кислоты
- Сукцинатдегидрогеназа
- Сукцинат — КоА-лигаза (АДФ-образующая)
- Сукцинат - КоА-лигаза (формирующая ВВП)
Ссылки
[ редактировать ]- ^ Фрейзер М.Э., Хаякава К., Хьюм М.С., Райан Д.Г., Брауни Э.Р. (апрель 2006 г.). «Взаимодействие GTP с доменом АТФ-захвата GTP-специфической сукцинил-КоА-синтетазы» . Журнал биологической химии . 281 (16): 11058–65. дои : 10.1074/jbc.M511785200 . ПМИД 16481318 .
- ^ Фрейзер М.Э., Джеймс М.Н., Бриджер В.А., Володко В.Т. (январь 1999 г.). «Подробное структурное описание сукцинил-КоА-синтетазы Escherichia coli». Журнал молекулярной биологии . 285 (4): 1633–53. дои : 10.1006/jmbi.1998.2324 . ПМИД 9917402 .
- ^ Перейти обратно: а б Воэт, Дональд Дж. (2011). Биохимия / Дональд Дж. Воэт; Джудит Г. Воэт . Нью-Йорк, штат Нью-Йорк: Wiley, J. ISBN 978-0-470-57095-1 .
- ^ Перейти обратно: а б с Берг, Джереми М. (Джереми М.); Тимочко, Джон Л.; Страйер, Люберт; Страйер, Люберт. Биохимия. (2002). Биохимик . Нью-Йорк: WH Freeman. стр. 475–477 . ISBN 0-7167-3051-0 .
- ^ Нисимура Дж.С. (1986). «Структурно-функциональные взаимоотношения сукцинил-КоА-синтетазы и другие соображения». Достижения энзимологии и смежных областей молекулярной биологии . Достижения в энзимологии и смежных областях молекулярной биологии. Том. 58. стр. 141–72. дои : 10.1002/9780470123041.ch4 . ISBN 9780470123041 . ПМИД 3521216 .
- ^ Перейти обратно: а б с Володько В.Т., Кей К.М., Бриджер В.А. (сентябрь 1986 г.). «Активная седиментация ферментов, скорость седиментации и исследования седиментационного равновесия сукцинил-КоА-синтетаз сердца свиньи и Escherichia coli». Биохимия . 25 (19): 5420–5. дои : 10.1021/bi00367a012 . ПМИД 3535876 .
- ^ Перейти обратно: а б с Фрейзер М.Э., Джеймс М.Н., Бриджер В.А., Володко Дж. (май 1999 г.). «Подробное структурное описание сукцинли-КоА-синтетазы Escherichia coli» . Журнал молекулярной биологии . 288 (3): 501. doi : 10.1006/jmbi.1999.2773 . ПМИД 10329157 .
- ^ Перейти обратно: а б Джойс М.А., Фрейзер М.Е., Джеймс М.Н., Бриджер В.А., Володко В.Т. (январь 2000 г.). «АДФ-связывающий сайт сукцинил-КоА-синтетазы Escherichia coli, выявленный методом рентгеновской кристаллографии». Биохимия . 39 (1): 17–25. дои : 10.1021/bi991696f . ПМИД 10625475 .
- ^ Перейти обратно: а б Фрейзер М.Э., Джойс М.А., Райан Д.Г., Володко В.Т. (январь 2002 г.). «Два остатка глутамата, Glu 208 альфа и Glu 197 бета, имеют решающее значение для фосфорилирования и дефосфорилирования остатка гистидина в активном центре сукцинил-КоА-синтетазы». Биохимия . 41 (2): 537–46. дои : 10.1021/bi011518y . ПМИД 11781092 .
- ^ Перейти обратно: а б с д Джонсон Дж.Д., Мехус Дж.Г., Тьюс К., Милавец Б.И., Ламбет Д.О. (октябрь 1998 г.). «Генетические доказательства экспрессии АТФ- и ГТФ-специфических сукцинил-КоА-синтетаз у многоклеточных эукариот» . Журнал биологической химии . 273 (42): 27580–6. дои : 10.1074/jbc.273.42.27580 . ПМИД 9765291 .
- ^ Перейти обратно: а б Ламбет Д.О., Тьюс К.Н., Адкинс С., Фрелих Д., Милавец Б.И. (август 2004 г.). «Экспрессия двух сукцинил-КоА-синтетаз с различной нуклеотидной специфичностью в тканях млекопитающих» . Журнал биологической химии . 279 (35): 36621–4. дои : 10.1074/jbc.M406884200 . ПМИД 15234968 .
- ^ Джонсон Дж.Д., Мухонен В.В., Ламбет Д.О. (октябрь 1998 г.). «Характеристика АТФ- и ГТФ-специфичных сукцинил-КоА-синтетаз у голубей. Ферменты включают одну и ту же альфа-субъединицу» . Журнал биологической химии . 273 (42): 27573–9. дои : 10.1074/jbc.273.42.27573 . ПМИД 9765290 .
- ^ Лаббе Р.Ф., Курумада Т., Онисава Дж. (декабрь 1965 г.). «Роль сукцинил-КоА-синтетазы в контроле биосинтеза гема». Biochimica et Biophysical Acta (BBA) – Общие предметы . 111 (2): 403–15. дои : 10.1016/0304-4165(65)90050-4 . ПМИД 5879477 .
- ^ Оттауэй Дж. Х., Макклеллан Дж. А., Сондерсон К. Л. (1981). «Сукциновая тиокиназа и метаболический контроль». Международный журнал биохимии . 13 (4): 401–10. дои : 10.1016/0020-711x(81)90111-7 . ПМИД 6263728 .
- ^ Дженкинс Т.М., Вейцман П.Д. (сентябрь 1986 г.). «Особые физиологические роли сукцинаттиокиназ животных. Связь сукцинаттиокиназы, связанной с гуаниновым нуклеотидом, с утилизацией кетоновых тел» . Письма ФЭБС . 205 (2): 215–8. дои : 10.1016/0014-5793(86)80900-0 . ПМИД 2943604 . S2CID 23667115 .
- ^ Круспль В., Штрайтманн Б. (февраль 1975 г.). «[Узловатый ретикулез с келоидными образованиями]». Журнал кожных заболеваний . 50 (3): 117–25. ПМИД 179232 .
- ^ Хангер-Глейзер И., Брун Р., Линдер М., Зеебек Т. (май 1999 г.). «Ингибирование фосфорилирования гистидина сукцинил-КоА-синтетазы у Trypanosoma brucei ингибитором бактериальных двухкомпонентных систем». Молекулярная и биохимическая паразитология . 100 (1): 53–9. дои : 10.1016/s0166-6851(99)00032-8 . ПМИД 10376993 .
- ^ Wider de Xifra E, del C Batlle AM (март 1978 г.). «Биосинтез порфиринов: иммобилизованные ферменты и лиганды. VI. Исследования сукцинил-КоА-синтетазы из культивируемых клеток сои». Биохимика и биофизика Acta . 523 (1): 245–9. дои : 10.1016/0005-2744(78)90027-x . ПМИД 564714 .
- ^ Перейти обратно: а б Остергаард Э., Кристенсен Э., Кристенсен Э., Могенсен Б., Дуно М., Шубридж Э.А., Вибранд Ф. (август 2007 г.). «Дефицит альфа-субъединицы сукцинат-коэнзима А-лигазы вызывает фатальный детский лактоацидоз с истощением митохондриальной ДНК» . Американский журнал генетики человека . 81 (2): 383–7. дои : 10.1086/519222 . ПМК 1950792 . ПМИД 17668387 .
Внешние ссылки
[ редактировать ]- Сукцинил + коэнзим + А + синтетазы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)