Подокезавр
| Подокезавр Временной диапазон: ранняя юра ,
| |
|---|---|

| |
| Образец голотипа с хвостом и неопределенными костями слева и телом справа. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Суперсемейство: | † Целофизоиды |
| Род: | † Подокезавр Талбот , 1911 год. |
| Разновидность: | † П. Holyokensis
|
| Биномиальное имя | |
| † Подокезавр холокенсис Талбот, 1911 год.
| |
| Синонимы | |
| |
Подокезавр — это род целофизоидных раннем динозавров , который жил на территории нынешней восточной части США в юрском периоде. Первая окаменелость была обнаружена геологом Миньон Тэлбот недалеко от горы Холиок , штат Массачусетс , в 1910 году. Образец был фрагментарным, сохранив большую часть тела, конечностей и хвоста. новый род и вид Podokesaurus Holyokensis В 1911 году Талбот описал и назвал на его основе . Полное название можно перевести как «Быстроногая ящерица Холиока». Это открытие сделало Талбот первой женщиной, которая нашла и описала динозавра, не являющегося птицей. была Окаменелость голотипа признана значимой и изучалась другими исследователями, но была утеряна, когда в 1917 году сгорело здание, в котором он хранился; однозначных экземпляров подокезавра с тех пор не было обнаружено . В 2022 году он был объявлен динозавром штата Массачусетс.
имел длину около 1 м (3 фута) и вес 1–40 кг (2–90 фунтов), По оценкам, подокезавр имел легкое телосложение с полыми костями и был похож на целофиза : стройный, с длинной шеей, и с острыми загнутыми зубами. Позвонки были очень легкими и полыми, а некоторые слегка вогнутыми на каждом конце. Шейные ( задними (шейные) позвонки были относительно большими по длине и диаметру по сравнению со спинными ) позвонками, а хвостовые (хвостовые) позвонки были длинными и тонкими. ( Плечевая кость кость плеча) была маленькой и нежной, ее длина составляла менее половины длины бедренной кости (бедренной кости). ( Лобковая кость лобковая кость) была очень длинной, расширяющейся как на переднем, так и на заднем концах. Бедренная кость стройная, почти прямая, с тонкими стенками и расширенная на задней стороне нижнего конца. Три плюсневые кости голени были плотно прижаты друг к другу, образуя компактную структуру.
Поскольку это был один из немногих мелких теропод, известных на момент его описания, родство подокезавра долгое время оставалось неясным. Он был отнесен к семейству Podokesauridae вместе с другими мелкими тероподами и, как предполагалось, был похож на протоптицу . Было высказано предположение, что это синоним целофиза , и к нему был отнесен природный слепок, но в настоящее время эти идеи не приняты. Семейство Podokesauridae больше не используется, его заменили Coelophysidae, а Podokesaurus , как полагают, был целофизоидом. Таким образом, Подокезавр был быстроногим хищником с мощными передними конечностями и хватательными руками. По оценкам, он мог двигаться со скоростью 15–20 км/ч (9–12 миль в час). Подокезавр , как полагают, был собран из портлендской формации , возраст которой долгое время был неясен, но в настоящее время считается, что он относится к геттангско - синемурскому этапам ранней юры, между 201 и 190 миллионами лет назад.
Открытие и история
[ редактировать ]

В 1910 году американский геолог Миньон Талбот шла со своей сестрой Эллен в Холиок , штат Массачусетс , на востоке США, когда они проходили мимо фермы и заметили неподалеку небольшой холм. С одной стороны у него был гравийный карьер, образованный скоплением песка, гравия и валунов, оставленных отступающим ледником . Талбот заметила белую полосу на валуне из песчаника на дне гравийного карьера и, обнаружив, что это кости, сказала сестре, что нашла «настоящее живое ископаемое». множество ископаемых следов Это произошло потому, что ранее было обнаружено в долине Коннектикута (на которые она часто возила своих учеников), но мало настоящих скелетов динозавров и ни одного на горе Холиок . Владелец земли предоставил ей разрешение собрать образец для колледжа Маунт-Холиок (женский колледж в нескольких милях оттуда), где она возглавляла геологический факультет. [ 1 ] [ 2 ] [ 3 ] [ 4 ]
На следующий день она привела группу рабочих, чтобы собрать образец, и нашла еще один кусок песчаника, который содержал остальную часть окаменелости, а также отпечатки тех, что были в первой плите. Образец, судя по всему, лежал под открытым небом в течение многих лет, и никто его не замечал, поскольку валун был взломан людьми или морозом. Окаменелость была доставлена в лабораторию, где ее подготовили и сфотографировали. [ 1 ] На неполном экземпляре сохранились 5 шейных (шейных), 11 спинных (спинных) и 24 хвостовых (хвостовых) позвонков, фрагмент левой лопатки (лопатки) и правого клювовидного отростка (часть плечевого пояса грудного пояса ), частичная левая плечевая кость (верхняя часть ). кость руки), кости фаланг 3 пальцев, в том числе 2 когтей (когтевые кости), ребра, лобковая и седалищная кости (кости, образующие соответственно нижняя передняя и нижняя задняя часть таза), бедренная кость (кость бедра), левая большеберцовая кость (кость голени), фрагмент правой таранной кости (кость голеностопного сустава), сочлененные плюсневые кости (кости стопы) левая стопа, за ней следуют 3 педальные фаланги (кости пальцев ног), частичные плюсневые кости и, возможно, фаланги правой стопы, а также фрагменты неопределенной идентичности. [ 5 ] [ 6 ] Легкие и хрупкие кости находились в своем естественном или почти естественном положении внутри камня, за исключением хвоста и неопределенных фрагментов, которые находились в нескольких сантиметрах от скелета, после длинного разрыва в скелете за тазом. Передняя половина шеи отсутствовала. [ 1 ] [ 7 ]


Значение окаменелости было подтверждено на межвузовском совещании факультетов геологии, и когда американский палеонтолог Ричард Суонн Лулл впоследствии предложил Тэлботу описать образец, она ответила, что ничего не знает о динозаврах, но Лулл предложил ей изучить их, а затем опишите это. В декабре 1910 года Талбот прочитала предварительное описание окаменелости на заседании Палеонтологического общества в Питтсбурге , а в июне 1911 года опубликовала краткое научное описание , в котором сделала экземпляр голотипом нового рода и вида Podokesaurus Holyokensis . [ 1 ] [ 8 ] Родовое название происходит от древнегреческих слов podōkēs ( ποδώκης ), что означает «быстроногий» ( эпитет , обычно используемый в отношении греческого героя Ахилла) , и saura ( σαύρα ), означающего «ящерица», в то время как конкретное название относится к Холиоку. Полностью название можно перевести как «Быстроногая ящерица Холиока». [ 8 ] [ 2 ] Открытие и наименование подокезавра сделало Талбот первой женщиной, которая нашла и описала динозавра, не являющегося птицей. [ 9 ] Американский палеонтолог Роберт Т. Баккер заявил в 2014 году, что, хотя старые профессора во время его учебы в университете ворчали, что женщины непригодны для работы с окаменелостями, открытие Талботом подокезавра было контраргументом этому. [ 10 ]
К тому времени, когда описание было опубликовано, Талбот отправил окаменелость в Музей естественной истории Пибоди в Йельском университете для дальнейшей подготовки и изучения, где также были изготовлены слепки из костей, лежавших в скале. Там Лулл нарисовал реконструкцию скелета, взяв за основу части, отсутствующие в окаменелости, на эквивалентах компсогната , и создал модель живого животного, которое Талбот позже описал как обладающее «сардонической улыбкой». Лулл подробно остановился на статье Талбота в публикации 1915 года, в которой он также предложил другие названия некоторых костей, отличные от того, что она первоначально предлагала, в том числе частичный клювовидный отросток вместо лопатки и часть большеберцовой кости вместо подвздошной кости (часть бедра) . ). Лулл предположил, что валун, содержащий окаменелость, должно быть, прибыл с уступа, который лежал на южной стороне хребта Холиок , примерно в двух или трех милях к северу от места, где он был найден, и уточнил, что он был из песчаника Лонгмидоу . [ 6 ] [ 2 ] [ 11 ] [ 7 ]

Лулл отправил свою рукопись датскому орнитологу Герхарду Хейльману Перед публикацией для критики. Хейльманн опубликовал свой ответ в статье 1913 года, в которую он включил ранее неопубликованные фотографии окаменелости, полученной от Талбота, а также свои собственные реставрации. Он не согласился с некоторыми анатомическими интерпретациями Луллия и переписывался по поводу окаменелости с американским зоологом Робертом Уилсоном Шуфельдтом по поводу своих утверждений. Статья Хейльмана была одной из серии о происхождении птиц , в которой он исследовал скелеты доисторических рептилий, чтобы найти черты, которые могли быть предками птиц. [ 12 ] На заседании Вашингтонской академии наук в 1916 году Шуфельдт представил отчет о своей переписке с Луллем, Талботом и Хейльманном и согласился с последним в некоторых критических замечаниях по поводу восстановления Луллем подокезавра . [ 13 ]
Тэлбот хотел, чтобы ископаемое оставалось в Йельском университете или Вашингтоне на постоянной выставке, где оно могло бы «быть среди себе подобных», но оно хранилось на горе Холиок в старом научном здании Уиллистон-Холл как местный образец, где оно стало « Любопытство» для студентов. Во время рождественских каникул 1917 года Уиллистон-холл сгорел, и никаких останков окаменелости подокезавра среди развалин обнаружено не было. Американский писатель Кристофер Бенфей отметил в 2002 году, что подокезавр имел особую особенность: он был динозавром, который исчез дважды. [ 2 ] [ 1 ] Немецкий палеонтолог Фридрих фон Хюне был последним, кто изучил и описал образец голотипа в публикации 1914 года, прежде чем он был уничтожен (он изучал его в Йельском университете до того, как было опубликовано описание Талбота). [ 7 ] [ 14 ] Хотя коллекции окаменелостей колледжа были почти полностью уничтожены пожаром, его помещения и коллекции впоследствии продолжали расти и улучшаться благодаря усилиям Талбота. [ 15 ]
Никаких других однозначных экземпляров подокезавра с тех пор обнаружено не было, но отлитые копии типового экземпляра остались в Музее естественной истории Пибоди и Американском музее естественной истории в Нью-Йорке . Подокезаврам не уделялось особого внимания, пока в конце 1940-х годов не были обнаружены новые и многочисленные окаменелости целофиза , и анатомия мелких тероподных динозавров не стала более ясной; Американские палеонтологи Эдвин Харрис Колберт и Дональд Бэрд сравнили подокезавра с целофизом в 1958 году, используя слепки первого. Наряду с зауроподоморфом анчизавром , подокезавр был одной из первых значительных коллекций скелетного материала динозавров, известных на восточном побережье Северной Америки. За исключением зуба (приписанного Coelophys в 1976 году), все остальные находки раннеюрских теропод на Восточном побережье представляют собой следы. [ 16 ] [ 17 ] [ 18 ]
В 2021 году Подокезавра и Анчизавра предложил в качестве претендентов на звание динозавра штата Массачусетс член Палаты представителей Джек Патрик Льюис , при этом Подокезавр выиграл онлайн-опрос. [ 19 ] [ 20 ] [ 21 ] 2 мая 2022 года губернатор Чарли Бейкер подписал закон, делающий его официальным динозавром штата. [ 22 ]
Описание
[ редактировать ]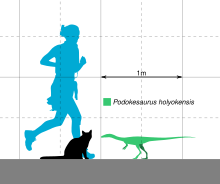
Сохранившееся тело единственной известной окаменелости подокезавра имело длину 18 см (7 дюймов). [ 8 ] По оценкам Лулла, полная длина подокезавра составляла 1150 мм (3,77 фута), шея - 130 мм (5,1 дюйма), спина - 170 мм (6,7 дюйма), крестец ( позвонки тазовой области) - 45. мм (1,8 дюйма), а хвост — 730 мм (2,40 фута). [ 6 ] По оценкам фон Хюне, длина животного составляла 1100 мм (3,6 фута), а длина хвоста составляла около 70 см (2,3 фута), что более чем в 1,5 раза превышало остальную часть скелета вместе взятого. [ 7 ] Кольбер оценил длину животного примерно в 1 м (3,3 фута). [ 17 ] В 1995 году писатель Ян Печкис оценил, что Подокезавр весил 10–40 кг (22–88 фунтов), путем определения высоты таза (на основе измерений фон Хюне). [ 23 ] Палеонтолог Грегори С. Пол подсчитал, что в 2016 году он весил 1 кг (2 фунта) (на основе оценки длины в 1 м). [ 24 ] В 1937 году Талбот заявил, что те ученые, которые видели этот образец, не думали, что это был молодой человек, поскольку не было никаких признаков хряща , который с возрастом превратился бы в кость. [ 1 ] в то время как Пол думал, что это, возможно, было юношеским. [ 24 ] В целом животное было легкого телосложения, с полыми костями . [ 8 ] Как целофизоид , он, вероятно, был бы похож по телосложению, например, на целофизиса : стройный, с длинной шеей и острыми, загнутыми назад зубами. [ 16 ]
Поскольку на момент описания Podokesaurus было известно немного других мелких теропод , его в основном сравнивали с более поздними родами Compsognathus , Ornitholestes и Ornithomimus , которые считались родственными. Из-за фрагментарности экземпляра подокезавра и того факта, что он утерян, идентичность и форма некоторых его костей сегодня неясны. Поскольку сейчас остались только слепки экземпляра, можно получить лишь общее представление о сохранившемся скелете и его пропорциях. Талбот идентифицировал костные фрагменты рядом с хвостом как кости черепа, потому что две из них были двусторонне симметричными , а один был широко выпуклым с бороздой (или бороздой) на средней линии и, по мнению Лулла, возможно, находился на верхней стенке черепа. , недалеко от лобных костей . Лулл предположил, что другие из этих элементов могли быть частью носовых костей . [ 6 ] [ 17 ] Вместо этого фон Хюне интерпретировал эти кости как хвостовые части середины хвоста. [ 7 ]
Посткраниальный скелет
[ редактировать ]




Позвонки были очень легкими и полыми, а некоторые слегка вогнутыми на каждом конце. Шейные позвонки были относительно большими по длине и диаметру по сравнению со спинными и имели шиловидные (по форме напоминающие ручку или щетину) шейные ребра, длинные, прямые и узкие, как у Anchisaurus . Передние спинные кости были значительно короче шейных, затем снова увеличивались в размерах к тазу. Их длина составляла 12–13 мм (0,47–0,51 дюйма). Нейральные отростки спинных костей были узкими, высокими и изогнутыми назад, но не такими расширенными спереди назад, как у Compsognathus . Спинные ребра были сильно изогнуты, очень тонкие и полые, напоминая ребра Saltopus и Hallopus , самое большое из которых имело длину 52 мм (2,0 дюйма) и ширину 2 мм (0,1 дюйма). Поперечные отростки (которые соединялись с ребрами на каждой стороне позвонка) образовывали круговую дугу вниз с каждой стороны спинного позвонка, простираясь примерно на такое же расстояние от центра («тела» позвонка), как и остистый отросток. был высоким. Зигапофизы (отростки , соединяющие соседние позвонки) были хорошо развиты. Центры были относительно длиннее, чем у Орнитомима , а у Компсогната больше напоминали таковые у Подокезавра , но были короче. [ 8 ] [ 6 ] [ 7 ]
Хотя задняя часть хвоста располагалась на некотором расстоянии от остального скелета, Лулл считал, что она находилась в своем естественном положении и что она должна была быть очень длинной, поскольку промежуточные позвонки были снесены. Хвостовые позвонки были очень длинными и тонкими и вместе с шевронами (костями на нижней стороне хвостовых позвонков) напоминали таковые у Ornitholestes . Типичная хвостовая часть имела длину 17 мм (0,67 дюйма) и диаметр 4 мм (0,16 дюйма). Хвостовые отростки были очень похожи по длине и не уменьшались в длину до последних нескольких в последовательности. Прямо перед лобком располагалось скопление тонких гастралий (брюшных ребер), самое длинное из которых имело длину 18 мм (0,71 дюйма). Передние гастралии имели расширенные концы, что часто характерно для грудинных ребер . [ 8 ] [ 6 ] [ 7 ]
Плечевая кость была маленькой и очень нежной, длиной 42 мм (1,7 дюйма), что составляло менее половины длины бедренной кости. У него был сильный латеральный отросток ( отросток, направленный в сторону) на 30 мм (1,2 дюйма) выше его нижнего конца. Лулл обнаружил, что он похож на орнитолестес , за исключением того, что радиальный гребень (который соединялся с лучевой костью предплечья) не был таким высоким и демонстрировал небольшую мышечную силу. Фон Хюне оценил длину радиуса в 30 мм (1,2 дюйма). Лулль обнаружил, что рука больше похожа на руку Компсогната, чем на Орнитолеста . [ 8 ] [ 6 ] [ 7 ] Сохранились только тонкие кости фаланг трех пальцев, включая острые изогнутые когти. [ 7 ]

Лобковая кость была очень длинной, расширяющейся как на переднем, так и на заднем концах. Он имел ширину 6 мм (0,24 дюйма) и длину 95 мм (3,7 дюйма), а диаметр нижнего расширения составлял 10 мм (0,39 дюйма). Талбот обнаружил, что лобок похож на лобок неописанного тогда Procompsognathus . Лулл утверждал, что лобковая кость у окаменелости лежала примерно в том же положении, направленная вперед. [ 8 ] [ 6 ] Вместо этого Хейльманн считал, что кость перед захоронением сместилась с места и должна была быть направлена назад. [ 12 ] Кольбер и Бэрд предположили, что лобковая кость была направлена вперед, но слегка изогнута вверх, а не вниз из-за естественного коробления и растрескивания кости. Седалищная кость имела длину 55 мм (2,2 дюйма), ширину 15,5 мм (0,61 дюйма) на конце, ближайшем к телу, а ее тонкий стержень имел диаметр 4 мм (0,16 дюйма) и несколько утолщался на нижнем конце. Лулль нашел его сходным с таковым у Компсогната и Орнитолеста . Ниже вертлужной впадины (там, где бедренная кость соединяется с тазом) имелась широкая пластинка , идущая к лобковой кости. [ 6 ] [ 7 ]
Бедренная кость стройная, почти прямая, с тонкими стенками и расширенная на задней стороне нижнего конца. Он был 86 мм (3,4 дюйма) в длину и 6,5 мм (0,26 дюйма) в диаметре чуть ниже четвертого вертела (фланца, расположенного посередине длины в задней части бедренной кости). Четвертый вертел был сильным, длиной 18 мм (0,71 дюйма) и высотой около 2 мм (0,079 дюйма) и располагался за серединой стержня, ближе к нижнему концу. Мыщелки (круглые выступы ) нижнего конца бедренной кости сильно выступали назад. Большеберцовая кость представляла собой тонкий узкий стержень длиной 104 мм (4,1 дюйма) и диаметром около 7 мм (0,28 дюйма). Тальбот заявил, что очень тонкая малоберцовая кость лежала близко к большеберцовой кости и имела почти одинаковую длину, в то время как фон Хюне считал, что это была часть раздавленной большеберцовой кости. Три плюсневые кости голени были тесно прижаты друг к другу, образуя компактную структуру, похожую на ту, что наблюдалась у Ornithomimus , но не сросшуюся в предплюсне-плюсневую кость , как это наблюдается у птиц. Длина плюсневых костей составляла 75 мм (3,0 дюйма), а их ширина - 9 мм (0,35 дюйма). Самая полная педальная фаланга была очень тонкой и длиной 12 мм (0,47 дюйма). [ 6 ] [ 8 ] [ 7 ]
Классификация
[ редактировать ]Ранние интерпретации
[ редактировать ]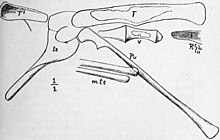
Читая свое предварительное описание в 1910 году, Талбот предположила, что Подокезавр был «травоядным динозавром», но дальнейшие работы в Йельском университете показали, что некоторые кости были идентифицированы неправильно, а седалищная кость таза с хорошо развитым гребнем оказалась напоминают Компсогната . Она воздержалась от однозначной классификации экземпляра в своем описании 1911 года из-за отсутствия челюсти и костей стопы, которые могли бы помочь в этом, но пришла к выводу, что он принадлежал «плотоядному динозавру», основываясь на форме и положении лобковой кости, а также отсутствие постлобковой кости . Она считала, что окаменелость относится к триасовому периоду (первый из трех мезозойских периодов). [ 8 ] В 1914 году фон Хюэне назвал новое семейство Podokesauridae, куда он, помимо Podokesaurus , включил Saltopus , Procompsognathus , Coelophys и Tanystropheus . Он поместил эту группу в категорию Coelurosauria , которая в то время использовалась для обозначения мелких теропод в целом. [ 25 ] [ 7 ] [ 26 ]
Хейльманн считал Подокезавра очень похожим на раннего археоптерикса в 1913 году и задавался вопросом, почему Талбот не провел с ним никаких сравнений. В частности, он обнаружил ноги и таз, когда лобковая кость интерпретировалась как направленная назад (как у птиц), подобно тому, как она могла выглядеть у предков птиц, но при этом считала реконструкцию Луллия направленной вперед лобковой кости маловероятной. Он также нашел части лопаток, передних конечностей и даже костей копчика, похожие на птичьи. Кроме того, он указал на наличие брюшных ребер и на то, что кости были полыми, и пришел к выводу, что ничто не мешает Подокезавру быть очень примитивной протоптицей с еще не сросшимися костями средней части стопы. Единственной особенностью, которая, по его мнению, не соответствовала этой интерпретации, был короткий палец с когтем, если только он не был частью пальца ноги. череп, грудина и ключица — кости, которые могли бы стать важным ключом к классификации. Он предположил, что проблему можно было бы решить, если бы в скале были найдены [ 12 ]


Хейльманн хотел получить более четкое объяснение расположения костей средней части стопы по отношению друг к другу, и нашел удивительным, что реконструкция Подокезавра, проведенная Луллием , показала еще более ущемленную среднюю часть стопы, чем она присутствовала у более позднего предполагаемого потомка Орнитомима , которого он Мысль к тому времени развила бы полностью птичью среднюю ногу. Хейльманн нашел подокезавра очень необычным для динозавра триаса и считал его одним из самых ранних двуногих позвоночных. Он предположил, что, поскольку он настолько отличался от текодонтозавра и платеозавра , возможно, это не были динозавры, или же подокезавр мог быть связан с Saltopus или паразухиями , но обнаружил, что его окаменелостей слишком недостаточно, чтобы сказать что-то определенное по этому вопросу. [ 12 ]
Шуфельдт подробно остановился на своей и Хейльманной интерпретации лобка в 1915 году и заявил, что кость, вероятно, сместилась во время окаменения. Он указывал, что если бы оно было обращено вперед, то имело бы силовой контакт с брюшными ребрами, что было бы опасно для внутренних органов во время движения. [ 11 ] Лулл счел критику Хейльмана и Шуфельдта по поводу реконструкции его таза вероятной в 1915 году. Он включил Podokesauridae в надсемейство Compsognatha и предположил, что Podokesaurus, возможно, был предком более поздних североамериканских родов Ornitholestes и Ornithimimus . [ 6 ] В 1916 году Хейльманн отметил, что ранние динозавры, паразухи и птерозавры имели сходство с птицами, а также друг с другом, и что триасовых рептилий, таких как Scleromochlus , Saltopus и Podokesaurus , было трудно разделить. Он предположил, что подокезавр и другие двуногие рептилии триаса произошли от псевдозухий многие типы архозавров ( к которым в то время относились ), которые, возможно, развили двуногость в пермском периоде . Он пришел к выводу, что птицы произошли от орнитозухов- псевдозухов, а не от динозавров, из-за их двуногости и птичьих ног. [ 27 ]
Палеонтолог Генри Фэйрфилд Осборн в 1917 году использовал подокезавра как пример «плотоядного динозавра птичьего типа», заявляя при этом, что сходство между птицами и динозаврами обусловлено параллельной эволюцией . [ 28 ] Палеонтолог Франц Нопца фон Фельсо-Сильвас в 1928 году создал подсемейство Podokesaurinae, в которое он включил Podokesaurus , Procerosaurus , Saltopus и Tanystropeus . [ 29 ] Фон Хюэн сгруппировал Podokesaurus , Coelophys , Spinosurus , Halticosaurus , Saltopus , Avipes и Velocipes в Podokesauridae в 1932 году. [ 30 ]
Отношение к целофизу
[ редактировать ]
В 1958 году Кольбер и Бэрд описали образец теропода, состоящий из естественных слепков костей в песчанике (вероятно образовавшихся в результате растворения костей в кислой воде, оставив после себя слепки костей), включая лобковую кость, большую берцовую кость и некоторые ребра. Они нашли экземпляр, похожий на Coelophys и Podokesaurus , назвав его C. sp. (неизвестного вида). Они считали, что естественный вид и подокезавр происходят из Портленда Аркозы группы Ньюарк в долине Коннектикута. Хотя подокезавр был меньше остальных (по размеру его можно было сравнить с самыми маленькими известными экземплярами целофиза ), исследователи предположили, что, поскольку он был так похож на них, это вызвало вопросы относительно его достоверности. [ 31 ] [ 32 ] В 1964 году Кольбер синонимизировал Podokesaurus с Coelophys (поскольку последнее название было старше), создав новую комбинацию C. Holyokensis . Он также предположил, что естественный слепок принадлежал C. Holyokensis . Кольбер заявил, что Подокезавра можно отличить от Целофиза только по тому, что нервные отростки его позвонков не такие длинные спереди назад, а также потому, что седалищная кость различалась по форме. Он обнаружил, что эти различия аналогичны тем, которые наблюдаются между современными видами рептилий одного и того же рода, и что они представляют восточные ( C. Holyokensis ) и западные ( C. bauri ) виды одного и того же рода. Хотя он признал, что эти выводы не были железными и что Подокезавр действительно мог отличаться от других, он сказал, что бремя доказывания должно лежать на сторонниках такой точки зрения. [ 17 ]
В 1977 году палеонтологи Пол Э. Олсен и Питер Гальтон отнесли супергруппу Ньюарк (к которой принадлежит портлендская формация) к раннему юрскому периоду вместо предшествующего триаса, как считалось ранее. Это было частью исследования, в котором Олсен сравнил фауну различных формаций и пришел к выводу, что на границе триаса и юры не произошло внезапного и широкомасштабного вымирания , а было постепенное. [ 33 ] Олсен заявил в 1980 году, что, хотя точное происхождение и систематическое положение подокезавра, вероятно, останется неопределенным, портлендская формация, из которой он, как предполагалось, произошел, вероятно, имела раннеюрский возраст и, следовательно, на 15 миллионов лет моложе целофиза из триаса. Поскольку черты, общие для Podokesaurus и Coelophys , также известны и у других мелких теропод, таких как Syntarsus , Олсен предположил, что Podokesaurus следует сохранить как отдельный род неопределенных теропод, чтобы избежать чрезмерного расширения временного диапазона целофизов . Он также применил этот аргумент к естественному составу, рассматривая его как incertae sedis (с неопределенными отношениями) среди теропод, а не как Podokesaurus или Coelophys . [ 34 ]
Палеонтолог Сэмюэл П. Уэллс заявил в 1984 году, что семейство Podokesauridae стало «универсальным» для большинства триасовых теропод. Хотя он нашел целофиза наиболее похожим на подокезавра среди теропод, он считал, что они сильно различаются тем, что четвертый вертел последнего находился ниже середины бедра (в отличие от более высокого положения у большинства других теропод), а его плюсневые кости были одинаковой длины. . [ 14 ] Палеонтолог Кевин Падиан заявил в 1986 году, что, хотя предложение Кольбера о синонимии было возможным, заметные сходства между подокезавром и целофизом заключались в примитивных чертах теропод, и эти два существа не были так близки во времени, как считалось раньше. [ 35 ] В 1988 году Пол сказал, что, хотя семейство Podokesauridae все еще использовалось для целофизов и его родственников, Podokesaurus изначально не был основан на хороших останках, и, поскольку остались только плохие копии, он думал, что всегда будут разногласия по поводу таксона. Он счел, что это слишком много двусмысленности, чтобы с этим мириться, и предложил вместо этого использовать название Coelophysidae (Halticosauridae, еще один претендент, также был основан на слишком фрагментарных останках). [ 36 ] В 1989 году Кольбер также усомнился в синонимии подокезавра с целофизом и в том, что естественные слепки принадлежали последнему из-за их пересмотренного раннеюрского возраста, и он не предпринял попыток определить статус подокезавра из-за потери единственного экземпляра. Он сохранил за семейством название Podokesauridae. [ 37 ]

В 1990 году палеонтологи Тимоти Роу и Жак Готье считали Podokesauridae таксономической корзиной для мусора , в которой таксоны были сгруппированы на основе фенетического сходства и стратиграфического разделения и, следовательно, находились под постоянным пересмотром и нестабильностью. Они считали возможным, что Подокезавр и естественный слепок были целофизами , но обнаружили, что их сходства не были присущи исключительно им, а были наследственными чертами среди теропод. Поэтому они согласились, что название Podokesaurus должно быть ограничено голотипом и что его и естественный состав следует считать Theropoda incertae sedis . [ 38 ] В том же томе палеонтолог Дэвид Б. Норман согласился с этой оценкой и заявил, что черты, используемые для объединения подокезавра с целофизом, просто подтверждают, что они были динозаврами, а не устанавливают между ними определенную связь. [ 5 ] Палеонтолог Томас Р. Хольц в 1994 году согласился с тем, что название семейства Podokesauridae следует заменить на Coelophysidae. К этому времени идея о том, что мелкие тероподы должны быть сгруппированы в Coelurosauria, а крупные тероподы в Carnosauria, вышла из моды, и Coelophysoidea считалась отдельной группой изящных ранних теропод. [ 39 ] Палеонтологи Дэвид Б. Вейшампель и Лютер Янг предположили подокезавра сходство с цератозаврами в 1996 году. [ 40 ]
Палеонтолог Рональд С. Тыкоски и Роу отметили в 2004 году, что, хотя у подокезавра были черты целофизоида (такие как небольшое шишковидное расширение в нижней части лобковой кости), у него не было каких-либо производных черт , которые могли бы объединить его с целофизом . Они согласились с более ранними исследователями в том, что название Podokesaurus следует ограничить голотипом, и пришли к выводу, что естественный вид может быть целофизоидом, но не может быть идентифицирован, кроме как теропод. [ 41 ] В 2004 году палеонтологи Мэтью Т. Каррано и Скотт Д. Сэмпсон заявили, что Подокезавр почти наверняка был целофизоидом, похожим на Целофизис . У него были целофизоидные черты, такие как длинная, изогнутая вниз лобковая кость, которая была длиннее седалищной кости, и дополнительное отверстие (отверстие) на лобковой кости, ниже запирательного отверстия можно было сказать немного больше , но о его родстве . [ 42 ] Каррано и его коллеги заявили в 2004 году, что Сегизавр и Подокезавр были одними из последних выживших целофизоидов и что эволюционное излучение этой группы, возможно, закончилось в конце ранней юры. [ 43 ] Также в 2004 году палеонтологи Антея Бристоу и Майкл А. Раат без каких-либо подробностей перечислили подокезавра как синоним целофиза . [ 44 ] Тыкоски отверг синонимичность этих двух видов в своей диссертации 2005 года , но исключил Подокезавра из своего анализа. [ 45 ] Пол предположил в 2016 году, что Podokesaurus , возможно, образовал семейство Coelophysidae с Coelophys , Panguraptor и Procompsognathus . [ 24 ]
Палеобиология
[ редактировать ]
Талбот предположил, что короткая, тонкая плечевая кость, длинные, прямые кости задних конечностей и хорошо развитый четвертый вертел бедренной кости указывают на то, что подокезавр был двуногим. Она обнаружила, что тот факт, что большеберцовая кость была намного длиннее бедренной, плюсневые кости были очень длинными, более половины длины голени, а легкая конструкция скелета свидетельствовала о быстром передвижении. Талбот заявил, что Лулл считал, что это адаптация к климатическим условиям, поскольку животное, должно быть, могло быстро и далеко путешествовать в поисках воды в своем полузасушливом регионе. Талбот также сообщил о небольшом кусочке гладкого полированного кварца среди ребер и предположил, что это мог быть гастролит (желудочный камень), и поэтому впервые они были обнаружены вместе с плотоядным динозавром. [ 8 ] Фон Хюэне согласился, что камень представлял собой гастролит, отличный от окружающего его песчаника, и добавил, что его длина составляет 11 мм (0,43 дюйма). [ 7 ]
Лулл обнаружил, что это животное было по сути стройным, беглым (приспособленным к бегу) животным с плотоядными привычками, но стройность, которая делала его быстрым, также ограничивала его мелкой добычей. [ 6 ] В 1932 году фон Хюне предположил, что маленькие целурозавры имели прыгающую походку, поскольку их нижние конечности длиннее верхних, что контрастирует с попеременными шагами карнозавров. Он подумал, что с его большим четвертым трокантером подокезавр , вероятно, отказался от этой прыжковой походки, вместо этого двигаясь быстрыми, попеременными шагами, как у бескилевых птиц. [ 30 ]
В 1982 году палеонтолог Ричард А. Талборн оценил скорость различных динозавров, основываясь на взаимосвязи между скоростью, походкой и размером тела современных животных (в основном млекопитающих). Экстраполируя длину шага и частоту шагов подокезавра , он подсчитал, что он мог бегать со скоростью около 15–20 км/ч (9–12 миль в час). Он пришел к выводу, что, если динозавр мог сравниться с млекопитающим в скорости, он должен был иметь аналогичный уровень эффективности двигательных способностей и физиологии. Он предупредил, что сравнение затруднено, поскольку анатомия динозавров значительно отличается от анатомии млекопитающих и птиц. Различия включали массивные хвосты динозавров и мышцы-ретракторы задних конечностей, которые возникли позади бедренной кости, тогда как у млекопитающих и современных птиц эти мышцы берут свое начало более вперед. [ 46 ] В 2006 году Вейшампель заявил, что, будучи целофизоидом, подокезавр был бы быстроногим хищником с мощными передними конечностями и хватательными руками. [ 16 ]
Лулл обнаружил, что ископаемые следы под названием Grallator курсориус совпадают с ступнями и шагами подокезавра . Их было много в карьере в Южном Хэдли (где они были найдены рядом с большими следами Отозума настроения ), что, по его мнению, подтверждало эту идентичность. [ 6 ] В 1926 году Хейльманн предположил, что стопа Procompsognathus лучше соответствует следам G. курсориуса из-за пропорций костей пальцев ног. [ 47 ] Палеонтолог Вильгельм Бок заявил в 1952 году, что, хотя очень похожие следы Граллятора приписывались различным мелким динозаврам (таким как подокезавры ), он считал такие корреляции слишком специфичными, и что лучшее, что можно было сказать, это то, что такие следы представляли мелких целурозавров. [ 48 ] Писатель Дональд Ф. Глут предположил в 1997 году, что плита со следами из формации Брансуик в Нью-Йорке, ранее приписывавшаяся целофизу, могла быть произведена подокезавром , исходя из его юрского возраста. [ 49 ] Эта формация теперь известна как формация Пассаик, а возраст ее считается позднетриасовым. [ 50 ]
Палеосреда
[ редактировать ]
Считается, что единственный известный экземпляр подокезавра был собран в формации Портленд (ранее известной как Портленд Аркоз) в бассейне Хартфорд в штате Массачусетс. Возраст этой формации долгое время был неясен (до 1977 года считалось, что она относится к триасовому периоду), но в настоящее время считается, что она относится к геттан - синемюрскому этапам ранней юры, между 201 и 190 миллионами лет назад (ранее считалось, что это плинсбах - ярус тоарский ). [ 51 ] [ 16 ] [ 34 ] В 2016 году палеонтолог Роберт Э. Уимс и его коллеги предложили возвести портлендскую формацию в геологическую группу в составе супергруппы Ньюарка (как Портлендскую группу), заменив тем самым прежнее название «группа Агавам». Они также восстановили песчаник Лонгмидоу, где был найден подокезавр , как образование (в пределах самой верхней Портлендской группы); ранее он считался идентичным портлендской формации. [ 51 ] [ 6 ]
Портлендская группа представляет собой самую верхнюю часть Ньюаркской супергруппы и образовалась после того, Центральноатлантическая магматическая провинция как в конце триаса и начале юры образовалась . Песчаник Лонгмидоу состоит из речных (отложенных реками и ручьями) красных аргиллитов , песчаников, конгломератов , второстепенных красных эоловых (отложенных ветром) песчаников и алевролитов . [ 51 ] Другие животные, известные из этой формации, включают динозавра-зауроподоморфа Anchisaurus , крокодиломорфа Stegomosochus и рыб, таких как Acentrophorus и Semionotus . [ 34 ] Следы динозавров включают ихнородов Anchisauripus , Anomoepus , Eubrontes и Grallator . [ 52 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Уорнер, Флорида (1937). «XII. Пропавший динозавр». В кампусе Новой Англии . Кембридж: Компания Houghton Mifflin. п. 279. АСИН Б00085ТО0О .
- ^ Jump up to: а б с д Бенфей, К. (2002). «Предисловие: «Путь исчезновения» » . Меняющиеся перспективы: вид с горы Холиок . Итака: Издательство Корнельского университета. п. 9. ISBN 978-0801441196 .
- ^ «Смело копать: Миньон Талбот (1869–1950)» . www.museumoftheearth.org . Архивировано из оригинала 11 февраля 2021 года . Проверено 14 августа 2020 г.
- ^ Росситер, М.В. (1981). «Геология в женском образовании девятнадцатого века в Соединенных Штатах» . Журнал геологического образования . 29 (5): 228–232. Бибкод : 1981JGeoE..29..228R . дои : 10.5408/0022-1368-29.5.228 .
- ^ Jump up to: а б Норман, Д.Б. (1990). «Проблематическая Теропода». В Вейшампеле, Д.Б.; Осмольская, Х.; Додсон, П. (ред.). Динозаврия (1-е изд.). Беркли: Издательство Калифорнийского университета. п. 298. ИСБН 978-0-520-06727-1 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Затишье, RS (1915). «Триасовая жизнь долины Коннектикута» . Государственная служба геологии и естествознания Коннектикута . 24 : 155–169. дои : 10.5962/bhl.title.70405 . ISBN 978-1167291937 . Архивировано из оригинала 6 октября 2020 года . Проверено 21 августа 2020 г.
- ^ Jump up to: а б с д и ж г час я дж к л м фон Хюне, Ф. (1914). «Вклад в историю архозавров» . Геологические и палеонтологические трактаты (на немецком языке). 13 :1–53. Архивировано из оригинала 9 октября 2020 года . Проверено 7 октября 2020 г.
- ^ Jump up to: а б с д и ж г час я дж к Талбот, М. (1911). « Podokesaurus Holyokensis , новый динозавр из триаса долины Коннектикута» . Американский научный журнал . с4-31 (186): 469–479. Бибкод : 1911AmJS...31..469T . дои : 10.2475/ajs.s4-31.186.469 . Архивировано из оригинала 18 августа 2020 года . Проверено 16 августа 2020 г.
- ^ Тернер, С.; Бурек, резюме; Муди, RTJ (2010). «Забытые женщины в вымершем ящеричьем (мужском) мире». Геологическое общество, Лондон, специальные публикации . 343 (1): 111–153. Бибкод : 2010GSLSP.343..111T . дои : 10.1144/SP343.7 . S2CID 130338204 .
- ^ Баккер, RT (2014). «Повесть о двух компьютерах: что в Парке Юрского периода было правильным и неправильным в отношении анатомии динозавров» . Хьюстонский музей естественных наук. Архивировано из оригинала 21 сентября 2020 года . Проверено 18 августа 2020 г.
- ^ Jump up to: а б Муди, Р.Л. (1913). «Некоторые последние достижения в палеонтологии позвоночных. II». Американский натуралист . 47 (556): 248–256. дои : 10.1086/279347 . ISSN 0003-0147 . JSTOR 2455799 .
- ^ Jump up to: а б с д Хейльманн, Г. (1913–1914). «Наши современные знания о происхождении птиц. Второй раздел: Птичье сходство древних ящериц» . Журнал Датской орнитологической ассоциации (на датском языке). 8 : 56–65.
- ^ Вегеманн, Швейцария; Лион, МВт (1916). «Труды Академии и дочерних обществ». Журнал Вашингтонской академии наук . 6 (9): 251–259. ISSN 0043-0439 . JSTOR 24521263 .
- ^ Jump up to: а б Уэллс, СП (1984). « Dilophosaurus wetherilli (Dinosauria, Theropoda), остеология и сравнения». Палеонтографика Абтейлунг А. 185 : 172.
- ^ Старейшина, Э.С. (1982). «Женщины в ранней геологии» . Журнал геологического образования . 30 (5): 287–293. Бибкод : 1982JGeoE..30..287E . дои : 10.5408/0022-1368-30.5.287 .
- ^ Jump up to: а б с д Вейшампель, Д.Б. (2006). Еще один взгляд на динозавров восточного побережья Северной Америки . III Международная конференция по палеонтологии динозавров и окружающей среде, Салас-де-лос-Инфантес, Бургос, Испания. Протоколы археолого-палеонтологического коллектива Саленсе. стр. 129–168.
- ^ Jump up to: а б с д Кольбер, Э.Х. (1964). «Триасовые динозавры родов Podokesaurus и Coelophys ». Американский музей Novitates (2168): 1–12. HDL : 2246/3350 .
- ^ Гальтон, премьер-министр (1976). «Динозавры-прозауроподы (Reptilia, Saurischia) Северной Америки». Постилла, Музей Пибоди, Йельский университет (169): 6.
- ^ Моррисон, Х. (4 февраля 2021 г.). «Динозавр штата Массачусетс: Podokesaurus Holyokensis побеждает в голосовании фанатов, законодатель подает законопроект, официально объявляющий динозавра штатом» . Масслайв . Архивировано из оригинала 21 марта 2021 года.
- ^ Кент, Л. (2021). «Массачусетс скоро предложит официального государственного динозавра» . CNN . Архивировано из оригинала 18 января 2021 года . Проверено 18 января 2021 г.
- ^ Кьянка, П. (2021). «Читатели Boston.com хотят, чтобы это был официальный динозавр Массачусетса» . www.бостон.com . Архивировано из оригинала 17 января 2021 года . Проверено 18 января 2021 г.
- ^ Морриссон, Х. (3 мая 2022 г.). «В Массачусетсе официально есть динозавр штата» . МассЛайв . Проверено 4 мая 2022 г.
- ^ Печкис, Дж. (1995). «Последствия оценок массы тела динозавров» . Журнал палеонтологии позвоночных . 14 (4): 530–531. дои : 10.1080/02724634.1995.10011575 . ISSN 0272-4634 .
- ^ Jump up to: а б с Пол, GS (2016). «Тероподы». Принстонский полевой справочник по динозаврам (2-е изд.). Принстон: Издательство Принстонского университета . стр. 76, 78. ISBN. 978-0691167664 .
- ^ фон Хюне, Ф. (1914). «Природная система ящериц» . Centralblatt по минералогии, геологии и палеонтологии (на немецком языке). 5 : 154-158. Архивировано из оригинала 27 октября 2020 года . Проверено 24 октября 2020 г.
- ^ Падиан, К. (2012). «Проблема происхождения динозавров: объединение трех подходов к возникновению динозавров». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 103 (3–4): 423–442. дои : 10.1017/S1755691013000431 . S2CID 128420276 .
- ^ Хейльманн, Г. (1916). «Наши современные знания о происхождении птиц. Пятый раздел: Ранняя птица Проавис» . Журнал Датской орнитологической ассоциации (на датском языке). 11:90 , 113, 137.
- ^ Осборн, HF (1917). Происхождение и эволюция жизни, по теории действия, реакции и взаимодействия энергии . Нью-Йорк: Сыновья К. Скрибнера. стр. 210–215. дои : 10.5962/bhl.title.7397 . Архивировано из оригинала 16 июня 2021 года . Проверено 21 сентября 2020 г.
- ^ Нопца, Ф. (1928). «Роды пресмыкающихся». Палеобиология (1): 163–188.
- ^ Jump up to: а б фон Хюне, Ф. (1932). «Ископаемый отряд рептилий Saurischia, его эволюция и история» (PDF) . Монографии по геологии и палеонтологии (на немецком языке). 4 (1): 24, 35, 332. Архивировано (PDF) из оригинала 25 февраля 2021 года . Проверено 20 декабря 2020 г.
- ^ Кольбер, Э.Х.; Бэрд, Д. (1958). «Слепки костей целурозавра из триаса долины Коннектикута» . Американский музей Novitates (1901): 1–11. hdl : 2246/2486 . Архивировано из оригинала 14 ноября 2020 года . Проверено 14 ноября 2020 г.
- ^ Гетти, PR; Буш, AM (2011). «Песчаные псевдоморфозы костей динозавров: последствия для (не) сохранения скелетного материала четвероногих в бассейне Хартфорд, США». Палеогеография, Палеоклиматология, Палеоэкология . 302 (3–4): 407. Бибкод : 2011PPP...302..407G . дои : 10.1016/j.palaeo.2011.01.029 .
- ^ Олсен, ЧП; Гальтон, премьер-министр (1977). «Триасово-юрское вымирание четвероногих: реальны ли они?». Наука . 197 (4307): 983–986. Бибкод : 1977Sci...197..983O . дои : 10.1126/science.197.4307.983 . ПМИД 17784133 . S2CID 44654197 .
- ^ Jump up to: а б с Олсен, ЧП (1980). «Сравнение комплексов позвоночных из бассейнов Ньюарка и Хартфорда (ранний мезозой, супергруппа Ньюарк) восточной части Северной Америки» (PDF) . В Джейкобсе, LL (ред.). Аспекты истории позвоночных: очерки в честь Эдвина Харриса Кольберта . Флагстафф: Музей прессы Северной Аризоны. стр. 35–53. ISBN 978-0897340526 . Архивировано (PDF) из оригинала 16 июня 2021 г. Проверено 16 ноября 2020 г.
- ^ Падиан, К. (1986). «О типовом материале Coelophys Cope (Saurischia: Theropoda) и новом экземпляре из Окаменевшего леса Аризоны (поздний триас: формация Чинл)». В Падиане, К. (ред.). Начало эпохи динозавров: изменения фауны на границе триаса и юры . Кембридж: Издательство Кембриджского университета. стр. 57–60. ISBN 978-0521367790 .
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 258–259. ISBN 978-0-671-61946-6 .
- ^ Кольбер, Э.Х. (1989). «Триасовый динозавр Целофиз». Бюллетень Музея Северной Аризоны . 57 : 24, 28–29. ISBN 978-0897340977 .
- ^ Роу, Т.; Готье, Дж. (1990). «Цератозаврия». В Вейшампеле, Д.Б.; Осмольская, Х.; Додсон, П. (ред.). Динозаврия (1-е изд.). Беркли: Издательство Калифорнийского университета. стр. 153, 166. ISBN. 978-0-520-06727-1 .
- ^ Хольц, Т.Р. (1994). «Филогенетическое положение тираннозаврид: значение для систематики теропод» . Журнал палеонтологии . 68 (5): 1100–1117. дои : 10.1017/S0022336000026706 . ISSN 0022-3360 . JSTOR 1306180 . Архивировано из оригинала 3 октября 2020 года . Проверено 3 октября 2020 г.
- ^ Вейшампель, Д.Б.; Янг, Л. (1996). Динозавры Восточного побережья . Балтимор: Издательство Университета Джонса Хопкинса. п. 275. ИСБН 978-0801852176 .
- ^ Тыкоски, РС; Роу, Т. (2004). «Цератозаврия». В Вейшампеле, Д.Б.; Додсон, П.; Осмольская, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. п. 68. ИСБН 978-0-520-24209-8 .
- ^ Каррано, Монтана; Сэмпсон, Скотт Д. (2004). «Обзор целофизоидов (Dinosauria: Theropoda) ранней юры Европы с комментариями к поздней истории Coelophysoidea». Neues Jahrbuch für Geologie und Paläontologie - Monatshefte . 2004 (9): 537–558. дои : 10.1127/njgpm/2004/2004/537 .
- ^ Каррано, Монтана; Хатчинсон-младший; Сэмпсон, С.Д. (2005). «Новая информация о Segisaurus Halli , небольшом динозавре-тероподе из ранней юры Аризоны». Журнал палеонтологии позвоночных . 25 (4): 835–849. doi : 10.1671/0272-4634(2005)025[0835:NIOSHA]2.0.CO;2 .
- ^ Бристоу, А.; Раат, Массачусетс (2004). «Молодой целофизоидный череп из ранней юры Зимбабве и синонимы целофиза и синтарсуса » . Палеонтология Африканская . ISSN 0078-8554 . Архивировано из оригинала 9 августа 2020 года . Проверено 20 октября 2020 г.
- ^ Тыкоски, Р.С. (2005). Анатомия, онтогенез и филогения целофизоидных теропод . Техасский университет в Остине (Диссертация). п. 69. Архивировано из оригинала 29 июля 2020 года . Проверено 20 октября 2020 г.
- ^ Талборн, РА (1982). «Скорость и походка динозавров». Палеогеография, Палеоклиматология, Палеоэкология . 38 (3–4): 227–256. Бибкод : 1982PPP....38..227T . дои : 10.1016/0031-0182(82)90005-0 .
- ^ Хейльманн, Г. (1926). Происхождение птиц . Нью-Йорк: Dover Publications. п. 181. ИСБН 0-486-22784-7 .
- ^ Бок, В. (1952). «Триасовые следы рептилий и тенденции локомотивной эволюции: с замечаниями о корреляции» . Журнал палеонтологии . 26 (3): 395–433. ISSN 0022-3360 . JSTOR 1299951 . Архивировано из оригинала 16 июня 2021 года . Проверено 30 сентября 2020 г.
- ^ Перенасыщение, Д.Ф. (1997). Динозавры: Энциклопедия . Джефферсон: McFarland & Company, Inc., стр. 718–719. ISBN 978-0786472222 .
- ^ Мец, Р. (2000). «Окаменелости триасового периода из озерных береговых отложений формации Пассаик, Дугласвилл, Пенсильвания». Ихнос . 7 (4): 253–266. дои : 10.1080/10420940009380165 . S2CID 128688223 .
- ^ Jump up to: а б с Уимс, RE; Таннер, Л.Х.; Лукас, СГ (2016). «Синтез и пересмотр литостратиграфических групп и формаций в верхней перми? – нижней юрской супергруппе Ньюарка на востоке Северной Америки» . Стратиграфия . 13 (2): 111–153. Архивировано из оригинала 5 августа 2020 года . Проверено 20 октября 2020 г.
- ^ Коллетт, Дж. Х.; Гетти, PR; Хагадорн, JW (2011). «Взгляд на среду обитания динозавров ранней юры: ихнофации и загадочные структуры из портлендской формации, карьер Гувера, Массачусетс, США» Atlantic Geology . 47 : 81–98. дои : 10.4138/atlgeol.2011.003 .
Внешние ссылки
[ редактировать ]- Музей науки, Бостон - «Прославьте Podokesaurus Holyokensis как динозавра штата Массачусетс». 24-минутное видео
- Музей науки, Бостон – 49-минутное видео «Выбор динозавра штата Массачусетс».














