Микробное сотрудничество
Эта статья нуждается в дополнительных цитатах для проверки . ( апрель 2019 г. ) |
Микроорганизмы участвуют в самых разных социальных взаимодействиях, включая сотрудничество . Кооперативное поведение – это поведение, приносящее пользу отдельному лицу (получателю), а не тому, кто выполняет такое поведение (актору). [ 1 ] В этой статье описываются различные формы кооперативного взаимодействия ( мутуализм и альтруизм ), наблюдаемые в микробных системах, а также преимущества, которые могли стимулировать эволюцию этого сложного поведения.
Введение
[ редактировать ]Микроорганизмы или микробы охватывают все три области жизни : бактерии , археи и многие одноклеточные эукариоты , включая некоторые грибы и простейшие . Обычно определяемые как одноклеточные формы жизни, которые можно наблюдать только с помощью микроскопа, микроорганизмы были первыми клеточными формами жизни и сыграли решающую роль в создании условий для эволюции более сложных многоклеточных форм.
Хотя микробы слишком малы, чтобы их можно было увидеть невооруженным глазом, они представляют подавляющее большинство биологического разнообразия и, таким образом, служат отличной системой для изучения вопросов эволюции. Одной из таких тем, которую ученые исследовали на микробах, является эволюция социального поведения, включая сотрудничество. Кооперативное взаимодействие приносит пользу получателю и выбирается на этом основании. Было документально подтверждено, что в микробных системах клетки, принадлежащие к одним и тем же таксонам, участвуют в кооперативных взаимодействиях, выполняя широкий спектр сложных многоклеточных действий, таких как расселение, поиск пищи, построение биопленок , размножение, химическая война и передача сигналов. В этой статье будут описаны различные формы кооперативного взаимодействия, наблюдаемые в микробных системах, а также преимущества, которые могли способствовать эволюции этого сложного поведения.
История
[ редактировать ]| Влияние на получателя | |||
|---|---|---|---|
| + | − | ||
| Влияние на актера | + | Взаимная выгода | Эгоизм |
| − | Альтруизм | Злость | |
С эволюционной точки зрения поведение является социальным, если оно имеет последствия для приспособленности как для человека, который выполняет это поведение (актера), так и для другого человека (реципиента). Гамильтон впервые классифицировал социальное поведение в зависимости от того, являются ли последствия, которые оно влечет за собой для действующего лица и получателя, полезными (увеличение прямой приспособленности) или дорогостоящими (уменьшение прямой приспособленности). [ 2 ] Согласно определению Гамильтона, существует четыре уникальных типа социальных взаимодействий: мутуализм (+/+), эгоизм (+/-), альтруизм (-/+) и злоба (-/-) (табл. 1). Мутуализм и альтруизм считаются кооперативными взаимодействиями, поскольку они приносят пользу получателю, и именно им будет посвящена эта статья.
Объяснение сотрудничества остается одной из величайших задач эволюционной биологии, независимо от того, считается ли поведение взаимовыгодным или альтруистическим. Согласно классической эволюционной теории, организм будет вести себя только так, чтобы максимизировать свою приспособленность . Таким образом, происхождение кооперативных взаимодействий или действий отдельных лиц, в результате которых другие люди получают преимущества в фитнесе, кажется нелогичным.
Теоретические объяснения эволюции сотрудничества можно разделить на две категории: прямые выгоды от приспособленности и косвенные выгоды от приспособленности. Это следует из предположения Гамильтона 1964 года о том, что особи приобретают инклюзивную приспособленность непосредственно через влияние на собственное воспроизводство (прямые эффекты приспособленности), а также через влияние на воспроизводство особей с родственными генами (косвенные эффекты приспособленности). [ 2 ]
Виды сотрудничества
[ редактировать ]мутуализм
[ редактировать ]Возможно, наиболее распространенные кооперативные взаимодействия, наблюдаемые в микробных системах, являются взаимовыгодными . Взаимовыгодные социальные взаимодействия обеспечивают прямую выгоду для фитнеса обоих участников, перевешивая при этом любые затраты на выполнение такого поведения. [ 3 ] В среде с отдельными микробами мутуализм чаще всего применяется с целью увеличения индивидуальной пользы для фитнеса. Однако в сообществе микроорганизмы будут взаимодействовать в больших масштабах, обеспечивая сохранение популяции, что тем самым повысит их собственную приспособленность. [ 4 ]
В большинстве случаев организмы, участвующие в таком поведении, имеют общий интерес к сотрудничеству. В микробных системах это часто наблюдается при производстве метаболически дорогих молекул, известных как общественные блага. Многие микробы, особенно бактерии, производят многочисленные общественные блага, которые высвобождаются во внеклеточную среду. Происходящая диффузия позволяет им использоваться соседними организмами, несмотря на то, что они производятся для человека.
Одним из очень популярных примеров взаимовыгодного микробного взаимодействия является производство сидерофоров . Сидерофоры — это молекулы, поглощающие железо, продуцируемые многими таксонами микробов , включая бактерии и грибы. Эти молекулы известны как хелатирующие агенты и играют важную роль в облегчении поглощения и метаболизма железа в окружающей среде, поскольку оно обычно существует в нерастворимой форме. [ 5 ] Чтобы бактерии получили доступ к этому ограничивающему фактору, клетки будут производить эти молекулы, а затем секретировать их во внеклеточное пространство. [ 6 ] После высвобождения сидерофоры будут связывать железо и образовывать комплекс, который распознается рецепторами бактериальных клеток. Затем его можно транспортировать в клетку и восстанавливать, делая железо метаболически доступным для бактерий. Производство сидерофоров часто используется как пример мутуализма, поскольку соединения не ограничены индивидуальным использованием. Пока в организме есть рецептор комплекса сидерофор-Fe (III), их можно воспринимать и использовать. [ 7 ]
Существует множество объяснений, оправдывающих эволюцию взаимовыгодного взаимодействия. Самое главное, чтобы производство общественных благ было эволюционно выгодным, поведение должно приносить прямую выгоду репродуктивным характеристикам субъекта, которая перевешивает затраты на реализацию такого поведения. [ 5 ] Чаще всего это наблюдается в случае прямой пользы для фитнеса. Поскольку бактерии чаще всего встречаются колониями, соседние бактерии, вероятно, будут иметь генетическую общность. Таким образом, увеличивая шансы на рост и деление близлежащей бактерии, хозяин увеличивает собственный проход генетического материала. В случае сидерофоров была обнаружена положительная корреляция между родством бактериальных линий и производством сидерофоров. [ 6 ]
Однако микробные сообщества заинтересованы не только в выживании и продуктивности своего вида. Было обнаружено, что в смешанном сообществе разные виды бактерий адаптируются к разным источникам пищи, включая продукты жизнедеятельности других видов, чтобы предотвратить ненужную конкуренцию. [ 8 ] Это позволяет повысить эффективность работы сообщества в целом.
Наличие сбалансированного сообщества очень важно для микробного успеха. В случае производства сидерофоров должно существовать равновесие между микробами, которые тратят свою энергию на производство хелатирующих агентов, и теми, которые могут использовать ксеносидерофоры. В противном случае эксплуататорские микробы в конечном итоге вытеснили бы производителей, оставив сообщество без организмов, способных производить сидерофоры, и, следовательно, неспособных выжить в условиях с низким содержанием железа. Эта способность балансировать между двумя популяциями в настоящее время исследуется. Считается, что это связано с наличием рецепторов с низким сродством у непродуцентов или продуцентов, генерирующих механизм интерференции, опосредованный токсином. [ 9 ]

Хотя производство общественных благ направлено на благо всех людей, оно также приводит к появлению мошенников, то есть людей, которые не оплачивают затраты на производство блага, но все равно получают выгоду (рис. 1). Чтобы минимизировать затраты на приспособленность, естественный отбор будет отдавать предпочтение особям, которые не выделяют секреты, используя при этом секреты своих соседей. В популяции клеток, секретирующих сидерофоры, несекретирующие мутантные клетки не оплачивают затраты на секрецию, но все же получают ту же выгоду, что и соседи дикого типа. Недавно Гриффин и др. (2004) исследовали социальную природу продукции сидерофоров у Pseudomonas aeruginosa . [ 10 ] Когда клетки, выращенные в чистой культуре, были помещены в среду с ограничением железа, популяции клеток, секретирующих сидерофоры ( дикий тип ), вытесняли популяцию мутантных несекреторных клеток. Следовательно, производство сидерофоров полезно при ограничении железа. Однако, когда те же популяции были помещены в среду, богатую железом, мутантная популяция вытеснила популяцию дикого типа, продемонстрировав, что производство сидерофоров является метаболически дорогостоящим. Наконец, когда бактерии дикого типа и мутантные бактерии были помещены в одну и ту же смешанную популяцию, мутанты могут получить выгоду от производства сидерофоров без оплаты затрат и, следовательно, увеличить частоту появления. Эту концепцию обычно называют трагедией общего достояния .
Игра «Дилемма заключенного» — это еще один способ, с помощью которого биологи-эволюционисты объясняют наличие мошенничества в кооперативных микробных системах. Первоначально сформулированная Меррилом Флудом и Мелвином Дрешером в 1950 году, «Дилемма узника» представляет собой фундаментальную проблему теории игр и демонстрирует, что два человека могут не сотрудничать, даже если это отвечает их интересам обоих. В этой дилемме каждый из двух человек выбирает, сотрудничать ли с другим человеком или обманывать. Сотрудничество обоих индивидов дает наибольшее среднее преимущество. Однако если один человек решит обмануть, он получит большее индивидуальное преимущество. Если игра проводится только один раз, мошенничество является превосходной стратегией, поскольку это превосходная стратегия. Однако в биологически реалистичных ситуациях с повторяющимися взаимодействиями (играми), мутациями и гетерогенной средой часто не существует единого стабильного решения, и успех отдельных стратегий может варьироваться в бесконечных периодических или хаотических циклах. Конкретное решение игры будет в решающей степени зависеть от способа реализации итераций и от того, как выигрыши преобразуются в динамику населения и сообщества.
В бактериях Escherichia coli ситуация «дилеммы заключенного» может наблюдаться, когда мутанты демонстрируют фенотип преимущества роста в стационарной фазе (GASP). [ 11 ] конкурировать со штаммом дикого типа (WT) в периодической культуре. [ 12 ] В таких условиях периодического культивирования, где среда роста гомогенизируется путем встряхивания культур, клетки WT взаимодействуют, останавливая рост бактерий , чтобы предотвратить экологический коллапс , в то время как мутанты GASP продолжают расти, переходя к регуляторному механизму дикого типа. В результате такого перехода к саморегуляции роста клетками GASP, хотя в краткосрочной перспективе достигается более высокая плотность клеток, в долгосрочной перспективе достигается популяционный коллапс из-за трагедии общего пользования (рис. 1). Напротив, хотя клетки ДТ не достигают такой высокой плотности населения, их популяции устойчивы при той же плотности в долгосрочной перспективе. Как предсказывает теория, [ 13 ] в пространственной обстановке, например, реализованной экспериментально с помощью микрофлюидных чипов, сосуществование двух штаммов возможно из-за локализации взаимодействий и пространственной сегрегации мошенников. [ 14 ] При наличии такой пространственной среды бактерии могут самоорганизовываться в динамические модели агрегации и десегрегации клеток, которые гарантируют, что сотрудничающие клетки WT могут пожинать плоды сотрудничества (рис. 2).


Greig & Travisano (2004) рассмотрели эти идеи в экспериментальном исследовании дрожжей Saccharomyces cerevisiae . [ 15 ] S. cerevisiae обладает множеством генов, каждый из которых продуцирует инвертазу — фермент, секретируемый для переваривания сахарозы вне клетки. Как обсуждалось выше, это производство общественного блага создает для отдельных клеток возможность обмана, крадя сахар, переваренный их соседями, без внесения самого фермента. Greig & Travisano (2004) измерили приспособленность типа мошенника (который обладал уменьшенным количеством генов инвертазы) по сравнению с типом кооператора (который содержал все возможные гены инвертазы). [ 15 ] Манипулируя уровнем социального взаимодействия внутри сообщества путем изменения плотности населения, они обнаружили, что мошенник менее приспособлен, чем сотрудничающий, на низких уровнях социальности, но более приспособлен к плотным сообществам. Поэтому они предполагают, что отбор на «обман» вызывает естественные вариации количества генов инвертазы, которыми может обладать индивид, и что вариации генов инвертазы отражают постоянную адаптацию к постоянно меняющейся биотической среде, что является следствием нестабильности кооперативных взаимодействий. .
Альтруизм
[ редактировать ]Второй тип кооперативных взаимодействий — альтруистический , или взаимодействия, которые выгодны получателю, но дорого обходятся действующему лицу (-/+). Обоснование эволюционной выгоды альтруистического поведения является широко обсуждаемой темой. Распространенным оправданием наличия альтруистического поведения является то, что оно приносит косвенную выгоду, поскольку поведение направлено на других людей, несущих ген сотрудничества. [ 2 ] Самая простая и наиболее распространенная причина, по которой два человека имеют общие гены, заключается в том, что они являются генеалогическими родственниками (родственниками), и поэтому это часто называют родственным отбором . [ 16 ] По мнению Гамильтона, альтруистический акт эволюционно выгоден, если родство индивида, получающего прибыль от альтруистического действия, выше, чем соотношение затрат и выгод, которое налагает этот акт. Это обоснование называется правилом Гамильтона .
Естественный отбор обычно благоприятствует гену, если он увеличивает воспроизводство, потому что потомство разделяет копии этого гена. Однако ген также может оказаться предпочтительным, если он помогает другим родственникам, у которых также есть общие копии. Таким образом, помогая близкому родственнику размножаться, особь все равно передает свои гены следующему поколению, хотя и косвенно. Гамильтон отметил, что родственный отбор может происходить посредством двух механизмов: (а) родственной дискриминации , когда сотрудничество преимущественно направлено на родственников, и (б) ограниченного расселения (вязкости популяции), которое удерживает родственников в пространственной близости друг к другу, позволяя сотрудничеству быть направлено без разбора на всех соседей (которые, как правило, являются родственниками). [ 2 ] В микробных системах эти два механизма одинаково важны. Например, большинство микробных популяций часто начинаются с небольшого числа колонизаторов. Поскольку большинство микробов размножаются бесполым путем , по мере роста популяции клетки будут окружать близкие генетические родственники. Эти клональные популяции часто приводят к чрезвычайно высокой плотности, особенно в наземных системах. Поэтому вероятность того, что альтруистическое поведение клеток пойдет на пользу близкому родственнику, чрезвычайно высока.
Хотя альтруистическое поведение наиболее распространено среди людей с высоким генетическим родством, оно не является абсолютно необходимым. Альтруистическое поведение также может быть эволюционно полезным, если сотрудничество направлено на людей, которые разделяют интересующий ген, независимо от того, связано ли это с общим происхождением или каким-либо другим механизмом. [ 17 ] Пример этого известен как механизм « зеленой бороды » и требует одного гена (или ряда тесно связанных генов), который одновременно вызывает кооперативное поведение и может быть распознан другими людьми благодаря отличительному фенотипическому маркеру, такому как зеленая борода. [ 2 ]
Наиболее изученной слизевиком с этой точки зрения является Dictyostelium discoideum , хищник бактерий, распространенный в почве. При голодании обычно одиночные одноклеточные амебы объединяются и образуют многоклеточный слизень, который может содержать 10 4 –10 6 клетки. Этот слизень мигрирует на поверхность почвы, где превращается в плодовое тело, состоящее из сферической верхушки спор и стебля, состоящего из нежизнеспособных клеток стебля, которые удерживают споры наверху (рис. 2). Примерно 20% клеток развиваются в нерепродуктивный стебель, поднимая споры и способствуя их распространению. [ 18 ]
Запрограммированная гибель клеток (ПКС) — еще одна предполагаемая форма микробного альтруистического поведения. Хотя запрограммированная гибель клеток (также известная как апоптоз или аутолиз ), очевидно, не обеспечивает прямой пользы для приспособленности, она может быть эволюционно-адаптивной, если она обеспечивает косвенные выгоды для людей с высоким генетическим родством ( родственный отбор ). Для PCD было предложено несколько альтруистических возможностей, таких как предоставление ресурсов, которые могли бы использоваться другими клетками для роста и выживания Saccharomyces cerevisiae . [ 19 ] [ 20 ] Хотя использование родственного отбора для объяснения эволюционных преимуществ PCD является обычным явлением, такое рассуждение содержит некоторые присущие ему проблемы. Чарльзворт (1978) отметил, что гену, вызывающему самоубийство, чрезвычайно трудно распространиться, поскольку в конечном итоге от этого выиграют только родственники, НЕ разделяющие этот ген. [ 21 ] Следовательно, возможное решение этой проблемы у микробов состоит в том, что отбор может способствовать низкой вероятности ПКС среди большой популяции клеток, возможно, в зависимости от индивидуальных условий, условий окружающей среды или передачи сигналов.
Другие микробные взаимодействия
[ редактировать ]Определение кворума
[ редактировать ]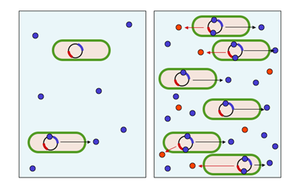
Интеграция кооперативных и коммуникативных взаимодействий представляется чрезвычайно важной для микробов; например, 6–10% всех генов бактерии Pseudomonas aeruginosa контролируются межклеточными сигнальными системами. [ 22 ] Одним из способов, с помощью которого микробы общаются и организуются друг с другом, чтобы участвовать в более продвинутых совместных взаимодействиях, является чувство кворума . Чувство кворума описывает явление, при котором накопление сигнальных молекул в окружающей среде позволяет одной клетке оценить количество особей (плотность клеток), чтобы популяция в целом могла дать скоординированный ответ. Это взаимодействие довольно распространено среди бактериальных таксонов и включает секрецию отдельными клетками «сигнальных» молекул, называемых аутоиндукторами или феромонами . У этих бактерий также есть рецептор , который может специфически обнаруживать сигнальную молекулу. Когда индуктор связывается с рецептором, он активирует транскрипцию определенных генов, в том числе генов синтеза индуктора. Вероятность того, что бактерия обнаружит собственный секретируемый индуктор, мала. Таким образом, чтобы активировать транскрипцию гена, клетка должна столкнуться с сигнальными молекулами, секретируемыми другими клетками в ее окружении. Когда поблизости находится всего несколько других бактерий того же вида, диффузия снижает концентрацию индуктора в окружающей среде практически до нуля, поэтому бактерии производят мало индуктора. Однако по мере роста популяции концентрация индуктора превышает порог, в результате чего синтезируется больше индуктора. Это формирует петлю положительной обратной связи , и рецептор полностью активируется. Активация рецептора вызывает активацию других специфических генов, в результате чего все клетки начинают транскрипцию примерно в одно и то же время. Другими словами, когда локальная концентрация этих молекул достигает порога, клетки реагируют включением определенных генов. Таким образом, отдельные клетки могут чувствовать локальную плотность бактерий, так что популяция в целом может дать скоординированный ответ. [ 23 ]
Во многих ситуациях затраты, которые бактериальные клетки платят за координацию поведения, перевешивают выгоды, если только нет достаточного количества сотрудников. Например, биолюминесцентная люцифераза, продуцируемая Vibrio fischeri, не была бы видна, если бы она вырабатывалась одной клеткой. Используя определение кворума для ограничения выработки люциферазы ситуациями, когда популяции клеток велики, клетки V. fischeri могут избежать траты энергии на производство бесполезного продукта. Во многих ситуациях бактериальная деятельность, такая как производство упомянутых общественных благ, имеет смысл только в случае совместной деятельности достаточного числа сотрудников. Регулирование посредством определения кворума позволит клеткам проявлять соответствующее поведение только тогда, когда оно эффективно, тем самым экономя ресурсы в условиях низкой плотности. Таким образом, ощущение кворума интерпретируется как бактериальная система коммуникации для координации поведения на уровне популяции.
Условно-патогенные бактерии Pseudomonas aeruginosa также используют чувство кворума для координации образования биопленок , роевой подвижности , производства экзополисахаридов и агрегации клеток. [ 24 ] Эти бактерии могут расти внутри хозяина, не причиняя ему вреда, пока не достигнут определенной концентрации. Затем они становятся агрессивными, их количество достаточно, чтобы преодолеть иммунную систему хозяина, и образуют биопленку, приводящую к заболеванию внутри хозяина. Другая форма регуляции генов, которая позволяет бактериям быстро адаптироваться к изменениям окружающей среды, — это передача сигналов окружающей среды. Недавние исследования показали, что анаэробиоз может существенно повлиять на основной регуляторный контур восприятия кворума. Эта важная связь между ощущением кворума и анаэробиозом оказывает существенное влияние на выработку факторов вирулентности этого организма. [ 25 ] Есть надежда, что терапевтическая ферментативная деградация сигнальных молекул предотвратит образование таких биопленок и, возможно, ослабит существующие биопленки. Нарушение сигнального процесса таким образом называется ингибированием кворума.
Бактериальное спаривание
[ редактировать ]S. pneumoniae развил сложную кооперативную систему определения кворума, которая регулирует выработку бактериоцинов , а также переход в компетентное состояние, необходимое для естественной генетической трансформации . [ 26 ] У естественно компетентных S. pneumoniae компетентное состояние, а не конститутивное свойство, индуцируется пептидным феромоном , включающим механизм определения кворума. [ 27 ] Индукция компетентности вызывает высвобождение ДНК из части популяции S. pneumoniae , скорее всего, за счет лизиса клеток. Впоследствии большинство клеток S. pneumoniae , которым индуцируется компетентность, действуют как реципиенты и поглощают ДНК, высвобождаемую донорами. [ 27 ] Естественная трансформация S. pneumoniae , вероятно, является естественным кооперативным механизмом, способствующим генетической рекомбинации , аналогичной половой у высших организмов. [ 27 ] Несмотря на наличие эффективных вакцин, S. pneumoniae является причиной смерти более миллиона человек ежегодно. [ 28 ]
V. cholerae обладает способностью эффективно взаимодействовать на клеточном уровне, и этот процесс признан формой совместного чувства кворума. [ 29 ] [ 30 ] Два различных стимула, которые встречаются в тонком кишечнике, отсутствие кислорода и присутствие солей желчных кислот , вырабатываемых хозяином , влияют на функцию восприятия V. cholerae quorum и, следовательно, на его патогенность. [ 31 ] Совместное ощущение кворума, вероятно, способствует естественной генетической трансформации , процессу, который включает поглощение V. cholerae внеклеточной ДНК ( компетентными ) клетками V. cholerae . [ 32 ] V. cholerae — бактериальный патоген, вызывающий холеру — заболевание, вызывающее тяжелую заразную диарею, от которой страдают миллионы людей во всем мире.
Бактериальная трансформация
[ редактировать ]По крайней мере, 80 видов бактерий, по-видимому, способны к трансформации, причем примерно поровну между грамположительными и грамотрицательными бактериями. [ 33 ]
Подразумеваемое
[ редактировать ]Хотя эволюция кооперативных взаимодействий позволила микробным таксонам повысить свою приспособленность, предполагается, что кооперация стала непосредственной причиной других крупных эволюционных переходов , включая эволюцию многоклеточности . [ 34 ] Эта идея, часто называемая колониальной теорией, была впервые предложена Геккелем в 1874 году и утверждает, что симбиоз многих организмов одного и того же вида (в отличие от теории симбиоза, предполагающей симбиоз разных видов) привел к появлению многоклеточного организма. . В некоторых случаях многоклеточность возникает в результате разделения клеток, а затем воссоединения (например, клеточные слизевики), тогда как для большинства многоклеточных типов многоклеточность возникает вследствие того, что клетки не могут разделиться после деления. [ 35 ] Механизм формирования последней колонии может быть таким же простым, как неполный цитокинез, хотя обычно считается, что многоклеточность включает клеточную дифференцировку. [ 36 ]
Преимущество гипотезы колониальной теории состоит в том, что она неоднократно наблюдалась независимо (в 16 различных типах протоктистана). Например, во время нехватки пищи клетки Dictyostelium discoideum группируются в колонию, которая как одно перемещается на новое место. Некоторые из этих клеток затем слегка дифференцируются друг от друга. Другими примерами колониальной организации простейших являются Volvocaceae , такие как Eudorina и Volvox . Однако часто бывает трудно отличить колониальные простейшие от настоящих многоклеточных организмов, поскольку эти две концепции не различны. Эта проблема мешает большинству гипотез о том, как могла произойти многоклеточность.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Вест С.А., Гриффин А.С., Гарднер А. 2007. Социальная семантика: альтруизм, сотрудничество, мутуализм, сильная взаимность и групповой отбор. Евро. Соц. для Эвола. Биол. 20: 415–432.
- ^ Перейти обратно: а б с д и Гамильтон ВД. 1964. Генетическая эволюция социального поведения, I&II. Дж. Теория. Биол. 7:1–52.
- ^ Сакс Дж.Л. и др. 2004. Эволюция сотрудничества. Ежеквартальный обзор биологии 79:135-160. doi: 10.1128/mBio.00099-12
- ^ Гимарайнш, П.Р., Пирес, М.М., Маркитти, Ф.М. и Раймундо, Р.Л. 2016. Экология мутуализмов. ЭЛС . 1–9. doi:10.1002/9780470015902.a0026295
- ^ Перейти обратно: а б Ниландс Дж.Б. Сидерофоры 1995. Структура и функции микробных соединений железа, переносящих железо. Ж. Биол. хим. 270:26723–6. 7. дои: 10.1074/jbc.270.45.26723
- ^ Перейти обратно: а б Вест С.А., Баклинг А. 2003. Сотрудничество, вирулентность и производство сидерофоров у бактериальных паразитов. Учеб. Р. Сок. Лон. Сер. Б 270:37–44. дои: 10.1098/rspb.2002.2209
- ^ Митке, М., Марахил М.А., 2007. Приобретение железа на основе сидерофоров и контроль патогенов. Микробиол. Мол. Биол. Откровение 71:413-451. doi: 10.1128/MMBR.00012-07
- ^ Лоуренс, Д. и др. 2010. Взаимодействие видов меняет эволюционные реакции на новую окружающую среду. ПЛОС. Био. дои: https://doi.org/10.1371/journal.pbio.1001330
- ^ Бутайте, Э. и др. 2017. Обман сидерофоров и устойчивость к мошенничеству формируют конкуренцию за железо в почвенных и пресноводных сообществах Pseudomonas. Нат. Коммун. 8. дои: 10.1038/s41467-017-00509-4
- ^ Гриффин А.С., Вест С.А., Баклинг А. 2004. Сотрудничество и конкуренция среди патогенных бактерий. Природа 430:1024–27.
- ^ Зинсер Э., Колтер Р. 2004. Эволюция Escherichia coli во время стационарной фазы. Рез. Микробиол . 155: 328–336
- ^ Вулич М., Колтер Р. 2001. Эволюционный обман в культурах стационарной фазы Escherichia coli. Генетика 158: 519–526.
- ^ Новак М., Бонхефффер С., Мэй Р. 1994. Пространственные игры и поддержание сотрудничества. ПНАС 91:4877-4881
- ^ Хол Ф., Галайда П., Надь К., Вултуис Р., Деккер К., Кеймер Дж. Э. 2013. Пространственная структура облегчает сотрудничество в социальной дилемме: эмпирические данные бактериального сообщества PLoS One 8 (10): e77042.
- ^ Перейти обратно: а б Грейг Д., Травизано М. 2004. Дилемма заключенного и полиморфизм генов SUC дрожжей. Биол. Летт. 271:С25–26.
- ^ Мейнард Смит Дж. 1964. Групповой отбор и родственный отбор. Природа 201: 1145–1147.
- ^ Вест SA и др. 2006. Теория социальной эволюции микробов. Нат. Преподобный Микробиол. 4:597–607.
- ^ Вест SA и др. 2007. Социальная жизнь микробов. Анну. Преподобный Экол. Эвол. Сист. 38:53-77.
- ^ Фабрицио П. и др. 2006. Супероксид является медиатором альтруистической программы старения у S. cerevisiae . Дж. Клеточная Биол. 166:1055–67.
- ^ Гурли CW, Ду В, Эйскоу КР. 2006. Апоптоз дрожжей – механизмы и преимущества для одноклеточного организма. Мол. Микробиол. 62:1515–21.
- ^ Чарльзуорт Б. 1978. Некоторые модели эволюции альтруистического поведения между братьями и сестрами. Дж. Теория. Биол. 72:297–319.
- ^ Шустер М. и др. 2003. Идентификация, время и специфичность сигнала генов, контролируемых кворумом Pseudomonas aeruginosa: анализ транскриптома. Дж. Бактериол. 185:2066–79.
- ^ Царан Т., Хоекстра РФ. 2009. Микробная коммуникация, сотрудничество и обман: ощущение кворума стимулирует эволюцию сотрудничества у бактерий. PLoS One 4:6655.
- ^ Льюис Зауэр К., Кампер А., Эрлих Г., Костертон Дж., Дэвис Д. 2002. Pseudomonas aeruginosa проявляет множественные фенотипы во время развития в виде биопленки. Журнал бактериологии 184: 1140–1154.
- ^ Корнелис П. 2008. Pseudomonas: геномика и молекулярная биология (1-е изд.). Кайстер Академик Пресс.
- ^ Шанкер Э., Федерле МЮ (январь 2017 г.). «Регуляция компетентности в отношении кворума и бактериоцины у Streptococcus pneumoniae и mutans». Гены (Базель) . 8 (1). дои : 10.3390/genes8010015 . ПМИД 28067778 .
{{cite journal}}: CS1 maint: неотмеченный бесплатный DOI ( ссылка ) - ^ Перейти обратно: а б с Стейнмоен Х., Кнутсен Э., Ховарштайн Л.С. (май 2002 г.). «Индукция естественной компетентности Streptococcus pneumoniae запускает лизис и высвобождение ДНК из части клеточной популяции». Proc Natl Acad Sci США . 99 (11): 7681–6. дои : 10.1073/pnas.112464599 . ПМИД 12032343 .
- ^ Юнгес Р., Сальвадори Г., Шекхар С., Амдал Х.А., Перисельнерис Дж.Н., Чен Т., Браун Дж.С., Петерсен ФК (2017). «Система определения кворума, которая регулирует образование биопленки Streptococcus pneumoniae и выработку поверхностных полисахаридов» . мСфера . 2 (5). дои : 10.1128/mSphere.00324-17 . ПМК 5597970 . ПМИД 28932816 .
- ^ Садживан А., Рамамурти Т., Соломон А.П. (март 2024 г.). «Вирулентность холерного вибриона и его подавление с помощью системы кворума». Крит. Рев. Микробиол : 1–22. дои : 10.1080/1040841X.2024.2320823 . ПМИД 38441045 .
- ^ Ли Ю, Ян Дж, Ли Дж, Сюэ Икс, Ван Ю, Цао Б (декабрь 2023 г.). «Новый регулятор восприятия кворума LuxT способствует вирулентности холерного вибриона» . Вирулентность . 14 (1): 2274640. doi : 10.1080/21505594.2023.2274640 . ПМЦ 10621291 . ПМИД 37908129 .
- ^ Машрувала А.А., Басслер Б.Л. (июль 2020 г.). «Кворум-чувствительный белок Vibrio cholerae VqmA объединяет сигналы плотности клеток, окружающей среды и хозяина в контроль вирулентности» . мБио . 11 (4). дои : 10.1128/mBio.01572-20 . ПМЦ 7387800 . ПМИД 32723922 .
- ^ Блокеш М. (сентябрь 2012 г.). «Переключение, опосредованное ощущением кворума, способствует естественной трансформации холерного вибриона». Моб Генет Элементы . 2 (5): 224–227. дои : 10.4161/mge.22284 . ПМИД 23446800 .
- ^ Джонстон С., Мартин Б., Фичант Г., Полард П., Клаверис Дж. П. (март 2014 г.). «Бактериальная трансформация: распространение, общие механизмы и дивергентный контроль». Nat Rev Микробиол . 12 (3): 181–96. дои : 10.1038/nrmicro3199 . ПМИД 24509783 .
- ^ Мейнард Смит Дж., Сатмари Э. 1995. Основные переходы в эволюции. Фриман, Оксфорд.
- ^ Вулперт Л., Сатмари Э. 2002. Многоклеточность: эволюция и яйцо. Природа 420:747-748.
- ^ Кирк Д.Л. 2005. Программа двенадцати шагов развития многоклеточности и разделения труда. Биоэссе 27: 299–310.