Некодирующая РНК

( Некодирующая РНК нкРНК ) — это функциональная молекула РНК , которая не транслируется в белок . Последовательность ДНК , из которой транскрибируется функциональная некодирующая РНК, часто называют геном РНК . Обильные и функционально важные типы некодирующих РНК включают транспортные РНК (тРНК) и рибосомальные РНК (рРНК), а также малые РНК, такие как микроРНК , миРНК , пиРНК , мяРНК , мяРНК , эксРНК , скаРНК и длинные нкРНК, такие как Xist. и ХОТЭР .
Количество некодирующих РНК в геноме человека неизвестно; однако недавние транскриптомные и биоинформатические исследования показывают, что существуют тысячи некодирующих транскриптов. [1] [2] [3] [4] [5] [6] [7] Многие из недавно идентифицированных нкРНК имеют неизвестные функции, если таковые имеются. [8] Нет единого мнения относительно того, какая часть некодирующей транскрипции является функциональной: некоторые полагают, что большинство нкРНК являются нефункциональными «мусорными РНК», ложными транскрипциями, [9] [10] в то время как другие ожидают, что многие некодирующие транскрипты имеют функции, которые еще предстоит открыть. [11] [12]
История и открытия
[ редактировать ]Нуклеиновые кислоты были впервые открыты в 1868 году Фридрихом Мишером . [13] а к 1939 году РНК была вовлечена в синтез белка . [14] Два десятилетия спустя Фрэнсис Крик предсказал функциональный компонент РНК, который опосредует трансляцию ; он пришел к выводу, что РНК лучше подходит для соединения пар оснований с транскриптом мРНК, чем чистый полипептид . [15]

Первой охарактеризованной некодирующей РНК была аланиновая тРНК, обнаруженная в пекарских дрожжах , ее структура была опубликована в 1965 году. [16] Чтобы получить очищенный образец аланиновой тРНК, Robert W. Holley et al. использовали 140 кг коммерческих пекарских дрожжей, чтобы получить всего 1 г очищенной тРНК. Земля для анализа. [17] 80- нуклеотидную тРНК секвенировали, сначала расщепляя рибонуклеазой поджелудочной железы (с получением фрагментов, оканчивающихся на цитозин или уридин ), а затем с помощью такадиастазы рибонуклеазы T1 (с получением фрагментов, заканчивающихся гуанозином ). Затем хроматография и идентификация 5'- и 3'-концов помогли расположить фрагменты для установления последовательности РНК. [17] Из трех структур, первоначально предложенных для этой тРНК, [16] Структура «клеверного листа» была независимо предложена в нескольких последующих публикациях. [18] [19] [20] [21] клеверного листа Вторичная структура была окончательно определена после рентгеновского кристаллографического анализа, проведенного двумя независимыми исследовательскими группами в 1974 году. [22] [23]
рибосомальная РНК Следующей была открыта , а в начале 1980-х годов — УрНА. С тех пор открытие новых некодирующих РНК продолжилось с помощью мякРНК , Xist , CRISPR и многих других. [24] Недавние заметные дополнения включают рибопереключатели и микроРНК ; открытие механизма РНКи , связанного с последним, принесло Крейгу К. Мелло и Эндрю Файру 2006 года Нобелевскую премию по физиологии и медицине . [25]
Недавние открытия нкРНК были достигнуты как экспериментальными, так и биоинформационными методами .
Биологические роли
[ редактировать ]Некодирующие РНК относятся к нескольким группам и участвуют во многих клеточных процессах. [26] Они варьируются от нкРНК центральной важности, которые сохраняются на протяжении всей или большей части клеточной жизни, до более временных нкРНК, специфичных для одного или нескольких близкородственных видов. Считается, что более консервативные нкРНК представляют собой молекулярные окаменелости или реликты от последнего универсального общего предка и мира РНК , и их нынешняя роль остается в основном в регуляции потока информации от ДНК к белку. [27] [28] [29]
В переводе
[ редактировать ]
Многие из консервативных, незаменимых и распространенных нкРНК участвуют в трансляции . рибонуклеопротеина Частицы (РНП), называемые рибосомами, являются «фабриками», на которых в клетке происходит трансляция. Рибосома состоит более чем на 60% из рибосомальной РНК ; они состоят из 3 нкРНК у прокариот и 4 нкРНК у эукариот . Рибосомальные РНК катализируют трансляцию нуклеотидных последовательностей в белок. Другой набор нкРНК, транспортные РНК , образуют «молекулу-адаптер» между мРНК и белком. МпРНК H /ACA-бокса и C/D-бокса представляют собой нкРНК, обнаруженные у архей и эукариот. РНКаза MRP доступна только эукариотам. Обе группы нкРНК участвуют в созревании рРНК. мякРНК осуществляют ковалентные модификации рРНК, тРНК и мяРНК ; РНКаза MRP расщепляет внутренний транскрибируемый спейсер 1 между 18S и 5,8S рРНК. Вездесущая нкРНК, РНКаза P , является эволюционным родственником РНКазы MRP. [31] РНКаза P созревает последовательности тРНК путем образования зрелых 5'-концов тРНК путем расщепления 5'-лидерных элементов тРНК-предшественников. Другой вездесущий RNP, называемый SRP, распознает и транспортирует специфические возникающие белки в эндоплазматический ретикулум у эукариот и на плазматическую мембрану у прокариот . У бактерий транспортная РНК (тмРНК) представляет собой РНП, участвующий в спасении застрявших рибосом, маркировании неполных полипептидов и содействии деградации аберрантной мРНК. [ нужна ссылка ]
При сплайсинге РНК
[ редактировать ]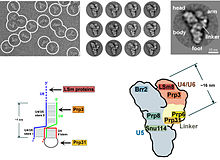
У эукариот сплайсосома осуществляет реакции сплайсинга, необходимые для удаления последовательностей интронов ; этот процесс необходим для образования зрелой мРНК . Сплайсосома — это еще один РНП , часто известный как мяРНП или три-мяРНП. Существуют две разные формы сплайсосомы: большая и второстепенная. Компонентами нкРНК основной сплайсосомы являются U1 , U2 , U4 , U5 и U6 . Компонентами нкРНК минорной сплайсосомы являются U11 , U12 , U5 , U4atac и U6atac . [ нужна ссылка ]
Другая группа интронов может катализировать собственное удаление из транскриптов хозяина; они называются самосплайсинговыми РНК. Существуют две основные группы самосплайсинговых РНК: каталитический интрон группы I и каталитический интрон группы II . Эти нкРНК катализируют собственное удаление из мРНК, тРНК и предшественников рРНК у широкого круга организмов. [ нужна ссылка ]
У млекопитающих было обнаружено, что мякРНК также могут регулировать альтернативный сплайсинг мРНК, например мякРНК HBII-52 регулирует сплайсинг рецептора серотонина 2C . [32]
У нематод нкРНК SmY , по-видимому, участвует в транссплайсинге мРНК . [33]
При репликации ДНК
[ редактировать ]
Y-РНК представляют собой петли ствола, необходимые для репликации ДНК посредством взаимодействия с хроматином и белками инициации (включая комплекс распознавания происхождения ). [35] [36] Они также являются компонентами рибонуклеопротеиновой частицы Ro60. [37] который является мишенью аутоиммунных антител у больных системной красной волчанкой . [38]
В регуляции генов
[ редактировать ]Экспрессия генов многих тысяч . регулируется нкРНК Эта регуляция может происходить в транс или в цис . Появляется все больше доказательств того, что особый тип нкРНК, называемый энхансерными РНК , транскрибируемый из энхансерной области гена, способствует экспрессии генов. [ нужна ссылка ]
Транзакции
[ редактировать ]У высших эукариот микроРНК регулируют экспрессию генов. Одна микроРНК может снизить уровень экспрессии сотен генов. Механизм действия зрелых молекул микроРНК заключается в частичной комплементарности одной или нескольким молекулам информационной РНК (мРНК), обычно в 3'-UTR . Основная функция микроРНК — подавление экспрессии генов.
что нкРНК РНКаза P Также было показано, влияет на экспрессию генов. В ядре человека РНКаза P необходима для нормальной и эффективной транскрипции различных нкРНК, транскрибируемых РНК-полимеразой III . К ним относятся гены тРНК, 5S рРНК , SRP РНК и гены мяРНК U6 . РНКаза P участвует в транскрипции посредством ассоциации с Pol III и хроматином активных генов тРНК и 5S рРНК. [39]
Было показано, что 7SK РНК , нкРНК многоклеточных животных , действует как негативный регулятор фактора элонгации РНК-полимеразы II P-TEFb , и что на эту активность влияют пути реакции на стресс. [ нужна ссылка ]
Бактериальная нкРНК, 6S РНК , специфически связывается с голоферментом РНК-полимеразы, содержащим фактор специфичности сигма70 . Это взаимодействие подавляет экспрессию сигма70-зависимого промотора во время стационарной фазы . [ нужна ссылка ]
Другая бактериальная нкРНК, OxyS РНК, подавляет трансляцию путем связывания с последовательностями Шайна-Дальгарно, тем самым блокируя связывание рибосом. РНК OxyS индуцируется в ответ на окислительный стресс у Escherichia coli. [ нужна ссылка ]
РНК B2 представляет собой небольшой некодирующий транскрипт РНК-полимеразы III, который подавляет транскрипцию мРНК в ответ на тепловой шок у мышей.клетки. РНК B2 ингибирует транскрипцию путем связывания с ядром Pol II. Благодаря этому взаимодействию РНК B2 собирается в преинициацию. комплексы на промоторе и блокируют синтез РНК. [40]
Недавнее исследование показало, что сам акт транскрипции последовательности нкРНК может влиять на экспрессию генов. Транскрипция нкРНК РНК-полимеразой II необходима для ремоделирования хроматина у Schizosaccharomyces pombe . Хроматин постепенно преобразуется в открытую конфигурацию по мере транскрипции нескольких видов нкРНК. [41]
Цис-действующий
[ редактировать ]Ряд нкРНК встроены в 5'- UTR (нетранслируемые области) генов, кодирующих белки , и различными способами влияют на их экспрессию. Например, рибопереключатель может напрямую связывать небольшую молекулу-мишень ; связывание мишени влияет на активность гена. [ нужна ссылка ]
Лидерные последовательности РНК находятся выше первого гена оперонов биосинтеза аминокислот. Эти элементы РНК образуют одну из двух возможных структур в участках, кодирующих очень короткие пептидные последовательности, богатые аминокислотой конечного продукта оперона. Терминаторная структура формируется, когда имеется избыток регуляторной аминокислоты и движение рибосомы над лидерным транскриптом не затруднено. При дефиците заряженной тРНК регуляторной аминокислоты рибосома, транслирующая лидерный пептид, останавливается и формируется антитерминаторная структура. Это позволяет РНК-полимеразе транскрибировать оперон. Известными лидерами РНК являются лидер гистидинового оперона , лидер лейцинового оперона , лидер треонинового оперона и лидер триптофанового оперона . [ нужна ссылка ]
Элементы реакции железа (IRE) связываются белками реакции железа (IRP). IRE обнаружен в UTR различных мРНК , продукты которых участвуют в метаболизме железа . Когда концентрация железа низкая, IRP связывают мРНК ферритина IRE, что приводит к репрессии трансляции. [ нужна ссылка ]
Внутренние сайты входа в рибосому (IRES) представляют собой структуры РНК , которые позволяют инициировать трансляцию в середине последовательности мРНК как часть процесса синтеза белка . [ нужна ссылка ]
В защиту генома
[ редактировать ]Piwi-взаимодействующие РНК (piRNA), экспрессируемые в млекопитающих Семенники и соматические клетки образуют РНК-белковые комплексы с Piwi белками . Эти комплексы piRNA (piRCs) связаны с подавлением транскрипционного гена ретротранспозонов и других генетических элементов в клетках зародышевой линии , особенно в клетках сперматогенеза .
Кластерные регулярно расположенные короткие палиндромные повторы (CRISPR) — это повторы, обнаруженные в ДНК многих бактерий и архей . Повторы разделены спейсерами одинаковой длины. Было продемонстрировано, что эти спейсеры могут быть получены из фага и впоследствии помочь защитить клетку от инфекции.
Хромосомная структура
[ редактировать ]Теломераза RNP — это фермент , который добавляет специфические повторы последовательности ДНК («TTAGGG» у позвоночных) к теломерным областям, которые находятся на концах эукариотических хромосом . Теломеры содержат конденсированный материал ДНК, придающий хромосомам стабильность. Фермент представляет собой обратную транскриптазу , которая несет теломеразную РНК , которая используется в качестве матрицы при удлинении теломер, которые укорачиваются после каждого цикла репликации .
Xist (X-неактивный транскрипт) представляет собой длинный ген нкРНК на Х-хромосоме плацентарных млекопитающих , который действует как главный эффектор процесса инактивации Х-хромосомы, образуя тельца Барра . Антисмысловая РНК Tsix является негативным регулятором Xist. Х-хромосомы, лишенные экспрессии Tsix (и, следовательно, имеющие высокий уровень транскрипции Xist), инактивируются чаще, чем нормальные хромосомы. У дрозофилид , которые также используют систему определения пола XY , РНК roX (РНК на X) участвуют в компенсации дозы. [42] И Xist, и roX действуют посредством эпигенетической регуляции транскрипции посредством рекрутирования ферментов, модифицирующих гистоны .
Бифункциональная РНК
[ редактировать ]Бифункциональные РНК или РНК с двойной функцией — это РНК, которые имеют две различные функции. [43] [44] Большинство известных бифункциональных РНК представляют собой мРНК, которые кодируют как белок, так и нкРНК. Однако все большее число нкРНК делятся на две разные категории нкРНК; например, H/ACA-бокса мякРНК и микроРНК . [45] [46]
Двумя хорошо известными примерами бифункциональных РНК являются SgrS РНК и РНКIII . Однако известно, что существует несколько других бифункциональных РНК (например, активатор стероидных рецепторов/SRA, [47] ВегТ РНК, [48] [49] Оскар РНК, [50] ЭНОД40 , [51] р53 РНК [52] SR1 РНК , [53] и РНК Spot 42 . [54] ) Бифункциональные РНК были темой специального выпуска журнала Biochimie за 2011 год . [55]
В качестве гормона
[ редактировать ]Существует важная связь между некоторыми некодирующими РНК и контролем путей, регулируемых гормонами. У дрозофилы такие гормоны, как экдизон и ювенильный гормон, могут способствовать экспрессии определенных микроРНК. Более того, эта регуляция происходит в различные временные моменты развития Caenorhabditis elegans . [56] У млекопитающих миР-206 является важнейшим регулятором альфа-рецептора эстрогена . [57]
Некодирующие РНК имеют решающее значение в развитии ряда эндокринных органов, а также при эндокринных заболеваниях, таких как сахарный диабет . [58] В частности, в клеточной линии MCF-7 добавление 17β- эстрадиола увеличивало глобальную транскрипцию некодирующих РНК, называемых днРНК, рядом с кодирующими генами, активируемыми эстрогеном. [59]
При патогенном избегании
[ редактировать ]Было показано, что C. elegans обучается и наследует патогенов избегание после воздействия одной некодирующей РНК бактериального патогена . [60] [61]
Роль в болезни
[ редактировать ]Как и в случае с белками , мутации или дисбаланс в репертуаре нкРНК в организме могут вызывать различные заболевания.
Рак
[ редактировать ]Многие нкРНК демонстрируют аномальные паттерны экспрессии в раковых тканях. [6] К ним относятся микроРНК , длинные мРНК-подобные нкРНК , [62] [63] ГАЗ5 , [64] СНОРД50 , [65] теломеразные РНК и Y-РНК . [66] МикроРНК участвуют в крупномасштабной регуляции многих генов, кодирующих белки. [67] [68] Y-РНК важны для инициации репликации ДНК, [35] теломеразная РНК, которая служит праймером для теломеразы, РНП, который расширяет теломерные области на концах хромосом ( см. в разделе «Теломеры и болезни дополнительную информацию »). Прямая функция длинных мРНК-подобных нкРНК менее ясна.
мутации зародышевой линии в первичных предшественниках миР-16-1 и миР-15 Было показано, что встречаются гораздо чаще у пациентов с хроническим лимфоцитарным лейкозом по сравнению с контрольной популяцией. [69] [70]
Было высказано предположение, что редкий SNP ( rs11614913 ), который перекрывается с hsa-mir-196a-2, связан с немелкоклеточной карциномой легкого . [71] Аналогичным образом, анализ 17 микроРНК, которые, как было предсказано, регулируют ряд генов, связанных с раком молочной железы, обнаружил вариации в микроРНК miR-17 и miR-30c-1 пациентов; эти пациенты не были носителями мутаций BRCA1 или BRCA2 , что дает возможность того, что семейный рак молочной железы может быть вызван вариациями этих микроРНК. [72] Супрессор опухоли р53 , возможно, является наиболее важным агентом в предотвращении образования и прогрессирования опухоли. Белок p53 действует как транскрипционный фактор, играющий решающую роль в организации клеточной реакции на стресс. Помимо своей решающей роли в развитии рака, р53 участвует в других заболеваниях, включая диабет, гибель клеток после ишемии и различные нейродегенеративные заболевания, такие как болезни Хантингтона, Паркинсона и Альцгеймера. Исследования показали, что экспрессия р53 регулируется некодирующей РНК. [5]
Другим примером нарушения регуляции некодирующей РНК в раковых клетках является длинная некодирующая РНК Linc00707. Linc00707 активируется и губчат микроРНК в мезенхимальных стволовых клетках, полученных из костного мозга человека, [73] при гепатоцеллюлярной карциноме, [74] рак желудка [75] или рак молочной железы, [76] [77] и, таким образом, способствует остеогенезу, способствует прогрессированию гепатоцеллюлярной карциномы, способствует пролиферации и метастазированию или косвенно регулирует экспрессию белков, участвующих в агрессивности рака, соответственно.
Синдром Прадера-Вилли
[ редактировать ]делеция 48 копий C/D-бокса мякРНК SNORD116 Было показано, что является основной причиной синдрома Прадера-Вилли . [78] [79] [80] [81] Прадера-Вилли — нарушение развития, связанное с перееданием и трудностями в обучении. SNORD116 имеет потенциальные сайты-мишени в ряде генов, кодирующих белок, и может играть роль в регуляции альтернативного сплайсинга. [82]
Аутизм
[ редактировать ]Хромосомный локус, содержащий кластер генов малой ядрышковой РНК SNORD115, дублируется примерно у 5% людей с аутистическими чертами . [83] [84] Модель мыши, созданная с дублированием кластера SNORD115, демонстрирует поведение, подобное аутизму. [85] Недавнее небольшое исследование посмертной ткани головного мозга продемонстрировало измененную экспрессию длинных некодирующих РНК в префронтальной коре и мозжечке аутичного мозга по сравнению с контрольной группой. [86]
Хрящево-волосовая гипоплазия
[ редактировать ]мутации в РНКазе MRP Было показано, что вызывают гипоплазию хряща и волос — заболевание, связанное с рядом симптомов, таких как низкий рост, редкие волосы, аномалии скелета и подавленная иммунная система, которая часто встречается среди амишей и финнов . [87] [88] [89] Наиболее охарактеризованным вариантом является переход A-to-G в нуклеотиде 70, который находится в области петли, состоящей из двух оснований 5' консервативного псевдоузла . Однако многие другие мутации в MRP РНКазы также вызывают CHH.
болезнь Альцгеймера
[ редактировать ]Антисмысловая РНК BACE1-AS транскрибируется с цепи, противоположной BACE1 , и ее активность повышается у пациентов с болезнью Альцгеймера . [90] BACE1-AS регулирует экспрессию BACE1 путем повышения стабильности мРНК BACE1 и генерации дополнительного BACE1 посредством посттранскрипционного механизма прямой связи. По тому же механизму он также повышает концентрацию бета-амилоида , основного компонента старческих бляшек. Концентрации BACE1-AS повышены у субъектов с болезнью Альцгеймера и у трансгенных мышей, содержащих белок-предшественник амилоида.
миР-96 и потеря слуха
[ редактировать ]Вариации в зародышевой области зрелой миР-96 связаны с аутосомно-доминантной прогрессирующей потерей слуха у людей и мышей. Гомозиготные мутантные мыши были глубоко глухими и не проявляли кохлеарной реакции. Гетерозиготные мыши и люди постепенно теряют способность слышать. [91] [92] [93]
Митохондриальные транспортные РНК
[ редактировать ]Ряд мутаций в митохондриальных тРНК связан с такими заболеваниями, как синдром MELAS , синдром MERRF и хроническая прогрессирующая наружная офтальмоплегия . [94] [95] [96] [97]
Различие между функциональной РНК (фРНК) и нкРНК
[ редактировать ]Ученые начали отличать функциональную РНК ( фРНК ) от нкРНК, чтобы описать функциональные области на уровне РНК, которые могут быть или не быть отдельными транскриптами РНК. [98] [99] [100] Это означает, что фРНК (например, рибопереключатели, элементы SECIS и другие цис-регуляторные области) не является нкРНК. Однако фРНК может также включать мРНК , поскольку она кодирует белок и, следовательно, функциональна. Кроме того, искусственно созданные РНК также подпадают под общий термин фРНК. Некоторые публикации [24] заявляют, что нкРНК и фРНК почти синонимы, однако другие отмечают, что большая часть аннотированных нкРНК, вероятно, не имеет никакой функции. [9] [10] Также было предложено просто использовать термин РНК , поскольку отличие от белка, кодирующего РНК ( информационная РНК ), уже определяется квалификатором мРНК . [101] Это устраняет двусмысленность при обращении к гену, «кодирующему некодирующую» РНК. Кроме того, может существовать ряд нкРНК, которые неправильно указаны в опубликованной литературе и наборах данных. [102] [103] [104]
См. также
[ редактировать ]- Внеклеточная РНК
- Список РНК
- Структура нуклеиновой кислоты
- Рфам
- Рибопереключатель
- Рибозим
- РНК присутствуют в образцах окружающей среды
- ВА (вирусная ассоциированная) РНК
Ссылки
[ редактировать ]- ^ Ченг Дж., Капранов П., Дренков Дж., Дике С., Брубейкер С., Патель С. и др. (май 2005 г.). «Транскрипционные карты 10 хромосом человека с разрешением 5 нуклеотидов». Наука . 308 (5725): 1149–1154. Бибкод : 2005Sci...308.1149C . дои : 10.1126/science.1108625 . ПМИД 15790807 . S2CID 13047538 .
- ^ Бирни Э., Стаматояннопулос Х.А., Дутта А. , Гиго Р., Гингерас Т.Р., Маргулис Э.Х. и др. (Консорциум проекта ENCODE) (июнь 2007 г.). «Идентификация и анализ функциональных элементов в 1% генома человека в рамках пилотного проекта ENCODE» . Природа . 447 (7146): 799–816. Бибкод : 2007Natur.447..799B . дои : 10.1038/nature05874 . ПМК 2212820 . ПМИД 17571346 .
- ^ Тинд А.С., Монга И., Тхакур П.К., Кумари П., Диндория К., Крзак М. и др. (ноябрь 2021 г.). «Демистификация новых приложений массового секвенирования РНК: применение и полезность биоинформатической методологии». Брифинги по биоинформатике . 22 (6). дои : 10.1093/нагрудник/bbab259 . ПМИД 34329375 .
- ^ Вашитл С., Педерсен Дж.С., Корбель Дж.О., Стокситс С., Грубер А.Р., Хакермюллер Дж. и др. (июнь 2007 г.). «Структурированные РНК в ENCODE выбранных регионах генома человека» . Геномные исследования . 17 (6): 852–864. дои : 10.1101/гр.5650707 . ЧВК 1891344 . ПМИД 17568003 .
- ^ Jump up to: а б Моррис К.В., изд. (2012). Некодирующие РНК и эпигенетическая регуляция экспрессии генов: движущие силы естественного отбора . Кайстер Академик Пресс . ISBN 978-1-904455-94-3 .
- ^ Jump up to: а б Шахруки П., Ларссон Э (2012). «Некодирующий онкоген: случай отсутствия доказательств ДНК?» . Границы генетики . 3 : 170. дои : 10.3389/fgene.2012.00170 . ПМЦ 3439828 . ПМИД 22988449 .
- ^ ван Бакель Х., Нислоу С., Бленкоу Б.Дж., Хьюз Т.Р. (май 2010 г.). Эдди С.Р. (ред.). «Большинство транскриптов «темной материи» связаны с известными генами» . ПЛОС Биология . 8 (5): e1000371. дои : 10.1371/journal.pbio.1000371 . ПМЦ 2872640 . ПМИД 20502517 .
- ^ Хюттенхофер А., Шаттнер П., Полачек Н. (май 2005 г.). «Некодирующие РНК: надежда или обман?». Тенденции в генетике . 21 (5): 289–297. дои : 10.1016/j.tig.2005.03.007 . ПМИД 15851066 .
- ^ Jump up to: а б Брозиус Дж. (май 2005 г.). «Не тратьте зря, не хотите — избыток транскриптов у многоклеточных эукариот». Тенденции в генетике . 21 (5): 287–288. дои : 10.1016/j.tig.2005.02.014 . ПМИД 15851065 .
- ^ Jump up to: а б Палаццо AF, Ли ES (2015). «Некодирующая РНК: что функционально, а что мусор?» . Границы генетики . 6 :2. дои : 10.3389/fgene.2015.00002 . ПМК 4306305 . ПМИД 25674102 .
- ^ Мэттик Дж., Амарал П. (2022). РНК, эпицентр генетической информации: новое понимание молекулярной биологии . ЦРК Пресс. ISBN 9780367623920 .
- ^ Ли Х., Чжан З., Краузе Х.М. (декабрь 2019 г.). «Длинные некодирующие РНК и повторяющиеся элементы: мусор или близкие эволюционные партнеры?» . Тенденции в генетике . 35 (12): 892–902. дои : 10.1016/j.tig.2019.09.006 . ПМИД 31662190 . S2CID 204975291 .
- ^ Дам Р. (февраль 2005 г.). «Фридрих Мишер и открытие ДНК». Биология развития . 278 (2): 274–288. дои : 10.1016/j.ydbio.2004.11.028 . ПМИД 15680349 .
- ^ Касперссон Т., Шульц Дж (1939). «Пентозные нуклеотиды в цитоплазме растущих тканей». Природа . 143 (3623): 602–3. Бибкод : 1939Natur.143..602C . дои : 10.1038/143602c0 . S2CID 4140563 .
- ^ Крик Ф.Х. (1958). «О синтезе белка». Симпозиумы Общества экспериментальной биологии . 12 : 138–163. ПМИД 13580867 .
- ^ Jump up to: а б Холли Р.В., Апгар Дж., Эверетт Г.А., Мэдисон Дж.Т., Маркизи М., Меррилл С.Х. и др. (март 1965 г.). «Структура рибонуклеиновой кислоты». Наука . 147 (3664): 1462–1465. Бибкод : 1965Sci...147.1462H . дои : 10.1126/science.147.3664.1462 . ПМИД 14263761 . S2CID 40989800 .
- ^ Jump up to: а б «Нобелевская премия по физиологии и медицине 1968 года» . Нобелевский фонд . Проверено 28 июля 2007 г.
- ^ Мэдисон Дж.Т., Эверетт Г.А., Кунг Х. (июль 1966 г.). «Нуклеотидная последовательность дрожжевой тирозиновой РНК». Наука . 153 (3735): 531–534. Бибкод : 1966Sci...153..531M . CiteSeerX 10.1.1.1001.2662 . дои : 10.1126/science.153.3735.531 . ПМИД 5938777 . S2CID 9265016 .
- ^ Захау Х.Г., Дюттинг Д., Фельдманн Х., Мельчерс Ф., Карау В. (1966). «Серинспецифические переносящие рибонуклеиновые кислоты. XIV. Сравнение нуклеотидных последовательностей и моделей вторичной структуры». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 31 : 417–424. дои : 10.1101/SQB.1966.031.01.054 . ПМИД 5237198 .
- ^ Дадок Б.С., Кац Г., Тейлор Э.К., Холли Р.В. (март 1969 г.). «Первичная структура РНК-переносчика фенилаланина зародышей пшеницы» . Труды Национальной академии наук Соединенных Штатов Америки . 62 (3): 941–945. Бибкод : 1969ПНАС...62..941Д . дои : 10.1073/pnas.62.3.941 . ПМК 223689 . ПМИД 5257014 .
- ^ Крамер Ф., Допнер Х., Хаар Ф. В.Д., Шлимме Э., Зайдель Х. (декабрь 1968 г.). «О конформации транспортных РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 61 (4): 1384–1391. Бибкод : 1968PNAS...61.1384C . дои : 10.1073/pnas.61.4.1384 . ПМК 225267 . ПМИД 4884685 .
- ^ Ладнер Дж. Э., Джек А., Робертус Дж. Д., Браун Р. С., Роудс Д., Кларк Б. Ф., Клуг А. (ноябрь 1975 г.). «Структура дрожжевой РНК-переносчика фенилаланина с разрешением 2,5 А» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (11): 4414–4418. Бибкод : 1975PNAS...72.4414L . дои : 10.1073/pnas.72.11.4414 . ПМЦ 388732 . ПМИД 1105583 .
- ^ Ким Ш., Куигли Г.Дж., Суддат Ф.Л., Макферсон А., Снеден Д., Ким Дж.Дж. и др. (январь 1973 г.). «Трехмерная структура дрожжевой РНК-переносчика фенилаланина: сворачивание полинуклеотидной цепи». Наука . 179 (4070): 285–288. Бибкод : 1973Sci...179..285K . дои : 10.1126/science.179.4070.285 . ПМИД 4566654 . S2CID 28916938 .
- ^ Jump up to: а б Эдди С.Р. (декабрь 2001 г.). «Некодирующие РНК-гены и современный мир РНК». Обзоры природы. Генетика . 2 (12): 919–929. дои : 10.1038/35103511 . ПМИД 11733745 . S2CID 18347629 .
- ^ Дэнехольт Б. «Дополнительная информация: РНК-интерференция» . Нобелевская премия по физиологии и медицине 2006 года . Архивировано из оригинала 20 января 2007 г. Проверено 25 января 2007 г.
- ^ Монга I, Банерджи I (ноябрь 2019 г.). «Вычислительная идентификация piRNA с использованием признаков, основанных на последовательности, структуре, термодинамических и физико-химических свойствах РНК» . Современная геномика . 20 (7): 508–518. дои : 10.2174/1389202920666191129112705 . ПМЦ 7327968 . PMID 32655289 .
- ^ Джеффарес, округ Колумбия, Пул А.М., Пенни Д. (январь 1998 г.). «Реликвии из мира РНК». Журнал молекулярной эволюции . 46 (1): 18–36. Бибкод : 1998JMolE..46...18J . дои : 10.1007/PL00006280 . ПМИД 9419222 . S2CID 2029318 .
- ^ Пул А.М., Джеффарес округ Колумбия, Пенни Д. (январь 1998 г.). «Путь из мира РНК». Журнал молекулярной эволюции . 46 (1): 1–17. Бибкод : 1998JMolE..46....1P . дои : 10.1007/PL00006275 . ПМИД 9419221 . S2CID 17968659 .
- ^ Пул А., Джеффарес Д., Пенни Д. (октябрь 1999 г.). «Ранняя эволюция: прокариоты, новые дети на районе». Биоэссе . 21 (10): 880–889. doi : 10.1002/(SICI)1521-1878(199910)21:10<880::AID-BIES11>3.0.CO;2-P . ПМИД 10497339 . S2CID 45607498 .
- ^ Бан Н., Ниссен П., Хансен Дж., Мур П.Б., Стейц Т.А. (август 2000 г.). «Полная атомная структура большой субъединицы рибосомы с разрешением 2,4 А». Наука . 289 (5481): 905–920. Бибкод : 2000Sci...289..905B . CiteSeerX 10.1.1.58.2271 . дои : 10.1126/science.289.5481.905 . ПМИД 10937989 .
- ^ Чжу Ю, Стрибинскис В, Рамос К.С., Ли Ю (май 2006 г.). «Анализ последовательности РНКазы MRP РНК показывает ее происхождение из РНК эукариотической РНКазы P» . РНК . 12 (5): 699–706. дои : 10.1261/rna.2284906 . ПМЦ 1440897 . ПМИД 16540690 .
- ^ Кишор С., Стамм С. (январь 2006 г.). «МякоРНК HBII-52 регулирует альтернативный сплайсинг рецептора серотонина 2C» . Наука . 311 (5758): 230–232. Бибкод : 2006Sci...311..230K . дои : 10.1126/science.1118265 . ПМИД 16357227 . S2CID 44527461 .
- ^ Джонс, Томас А.; Отто, Вольфганг; Марз, Манья; Эдди, Шон Р.; Стадлер, Питер Ф. (2009). «Обзор SmY-РНК нематод» . Биология РНК . 6 (1): 5–8. дои : 10.4161/rna.6.1.7634 . ISSN 1555-8584 . ПМИД 19106623 .
- ^ Штейн А.Дж., Фукс Г., Фу С., Волин С.Л., Райниш К.М. (май 2005 г.). «Структурные данные о контроле качества РНК: аутоантиген Ro связывает неправильно свернутые РНК через свою центральную полость» . Клетка . 121 (4): 529–539. дои : 10.1016/j.cell.2005.03.009 . ПМЦ 1769319 . ПМИД 15907467 .
- ^ Jump up to: а б Христов К.П., Гардинер Т.Дж., Сюц Д., Круде Т. (сентябрь 2006 г.). «Функциональные требования некодирующих Y-РНК для репликации хромосомной ДНК человека» . Молекулярная и клеточная биология . 26 (18): 6993–7004. дои : 10.1128/MCB.01060-06 . ПМК 1592862 . ПМИД 16943439 .
- ^ Чжан А.Т., Лэнгли А.Р., Христов К.П., Хейр Э., Шафи Т., Гардинер Т.Дж., Круд Т. (июнь 2011 г.). «Динамическое взаимодействие Y-РНК с хроматином и белками инициации во время репликации ДНК человека» . Журнал клеточной науки . 124 (Часть 12): 2058–2069. дои : 10.1242/jcs.086561 . ПМК 3104036 . ПМИД 21610089 .
- ^ Холл А.Е., Тернбулл С., Далмей Т. (апрель 2013 г.). «Y-РНК: последние разработки» . Биомолекулярные концепции . 4 (2): 103–110. дои : 10.1515/bmc-2012-0050 . ПМИД 25436569 . S2CID 12575326 .
- ^ Лернер М.Р., Бойл Дж.А., Хардин Дж.А., Стейц Дж.А. (январь 1981 г.). «Два новых класса малых рибонуклеопротеинов, обнаруженных с помощью антител, связанных с красной волчанкой». Наука . 211 (4480): 400–402. Бибкод : 1981Sci...211..400L . дои : 10.1126/science.6164096 . ПМИД 6164096 .
- ^ Райнер Р., Бен-Асули Ю., Криловецкий И., Джаррус Н. (июнь 2006 г.). «Роль каталитического рибонуклеопротеина РНКазы P в транскрипции РНК-полимеразы III» . Гены и развитие . 20 (12): 1621–1635. дои : 10.1101/gad.386706 . ПМЦ 1482482 . ПМИД 16778078 .
- ^ Эспиноза К.А., Аллен Т.А., Хиб А.Р., Кугель Дж.Ф., Гудрич Дж.А. (сентябрь 2004 г.). «РНК B2 напрямую связывается с РНК-полимеразой II, подавляя синтез транскрипта». Структурная и молекулярная биология природы . 11 (9): 822–829. дои : 10.1038/nsmb812 . ПМИД 15300239 . S2CID 22199826 .
- ^ Хирота К., Миёси Т., Куго К., Хоффман К.С., Сибата Т., Охта К. (ноябрь 2008 г.). «Поэтапное ремоделирование хроматина путем каскадной инициации транскрипции некодирующих РНК». Природа . 456 (7218): 130–134. Бибкод : 2008Natur.456..130H . дои : 10.1038/nature07348 . ПМИД 18820678 . S2CID 4416402 .
- ^ Пак Ю, Келли Р.Л., О Х, Курода М.И., Меллер В.Х. (ноябрь 2002 г.). «Степень распространения хроматина определяется рекрутированием roX РНК белков MSL». Наука . 298 (5598): 1620–1623. Бибкод : 2002Sci...298.1620P . дои : 10.1126/science.1076686 . ПМИД 12446910 . S2CID 27167367 .
- ^ Уодлер CS, Вандерпул CK (декабрь 2007 г.). «Двойная функция бактериальной малой РНК: SgrS осуществляет регуляцию, зависящую от спаривания оснований, и кодирует функциональный полипептид» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (51): 20454–20459. Бибкод : 2007PNAS..10420454W . дои : 10.1073/pnas.0708102104 . ПМК 2154452 . ПМИД 18042713 .
- ^ Дингер М.Э., Панг К.К., Мерсер Т.Р., Мэттик Дж.С. (ноябрь 2008 г.). Макинтайр Дж. (ред.). «Дифференциация белок-кодирующей и некодирующей РНК: проблемы и двусмысленности» . PLOS Вычислительная биология . 4 (11): e1000176. Бибкод : 2008PLSCB...4E0176D . дои : 10.1371/journal.pcbi.1000176 . ПМК 2518207 . ПМИД 19043537 .
- ^ Сарайя А.А., Ван CC (ноябрь 2008 г.). Гольдберг Д.Е. (ред.). «мякРНК, новый предшественник микроРНК в лямблиях лямблий» . ПЛОС Патогены . 4 (11): e1000224. дои : 10.1371/journal.ppat.1000224 . ПМЦ 2583053 . ПМИД 19043559 .
- ^ Эндер С., Крек А., Фридлендер М.Р., Бейтцингер М., Вайнманн Л., Чен В. и др. (ноябрь 2008 г.). «Человеческая мяРНК с функциями, подобными микроРНК» . Молекулярная клетка . 32 (4): 519–528. doi : 10.1016/j.molcel.2008.10.017 . ПМИД 19026782 .
- ^ Лига E (август 2007 г.). «Активатор РНК стероидного рецептора (SRA1): необычные двугранные генные продукты, предположительно имеющие отношение к раку молочной железы» . Передача сигналов ядерных рецепторов . 5 : e006. дои : 10.1621/nrs.05006 . ПМК 1948073 . ПМИД 17710122 .
- ^ Чжан Дж, Кинг ML (декабрь 1996 г.). «РНК Xenopus VegT локализуется в вегетативной коре во время оогенеза и кодирует новый фактор транскрипции Т-бокса, участвующий в формировании мезодермального паттерна». Разработка . 122 (12): 4119–4129. дои : 10.1242/dev.122.12.4119 . ПМИД 9012531 . S2CID 28462527 .
- ^ Клок М., Уилк К., Варгас Д., Ширато Ю., Билински С., Эткин Л.Д. (август 2005 г.). «Потенциальная структурная роль некодирующих и кодирующих РНК в организации цитоскелета вегетативной коры ооцитов Xenopus» . Разработка . 132 (15): 3445–3457. дои : 10.1242/dev.01919 . ПМИД 16000384 .
- ^ Дженни А., Хачет О., Заворски П., Цирклафф А., Уэстон М.Д., Джонстон Д.С. и др. (август 2006 г.). «Независимая от трансляции роль РНК oskar в раннем оогенезе дрозофилы» . Разработка . 133 (15): 2827–2833. дои : 10.1242/dev.02456 . ПМИД 16835436 .
- ^ Гультяев А.П., Руссис А (2007). «Идентификация консервативных вторичных структур и сегментов расширения в РНК enod40 выявляет новые гомологи enod40 у растений» . Исследования нуклеиновых кислот . 35 (9): 3144–3152. дои : 10.1093/нар/gkm173 . ПМЦ 1888808 . ПМИД 17452360 .
- ^ Кандейас М.М., Мальберт-Колас Л., Пауэлл Д.Д., Даскалоянни С., Маслон М.М., Наски Н. и др. (сентябрь 2008 г.). «МРНК P53 контролирует активность p53, управляя функциями Mdm2». Природная клеточная биология . 10 (9): 1098–1105. дои : 10.1038/ncb1770 . ПМИД 19160491 . S2CID 5122088 .
- ^ Гимпел М., Прейс Х., Барт Э., Грамцов Л., Брантл С. (декабрь 2012 г.). «SR1 — небольшая РНК с двумя удивительно консервативными функциями» . Исследования нуклеиновых кислот . 40 (22): 11659–11672. дои : 10.1093/nar/gks895 . ПМЦ 3526287 . ПМИД 23034808 .
- ^ Аояма Дж.Дж., Райна М., Чжун А., Сторц Г. (март 2022 г.). «РНК Spot 42 с двойной функцией кодирует белок из 15 аминокислот, который регулирует фактор транскрипции CRP» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (10): e2119866119. Бибкод : 2022PNAS..11919866A . дои : 10.1073/pnas.2119866119 . ПМЦ 8916003 . ПМИД 35239441 .
- ^ Франкастель К., Хубе Ф (ноябрь 2011 г.). «Кодирование или некодирование: должны ли они быть эксклюзивными?» . Биохимия . 93 (11): vi – vii. дои : 10.1016/S0300-9084(11)00322-1 . ПМИД 21963143 .
- ^ Семпере Л.Ф., Сокол Н.С., Дубровский Е.Б., Бергер Э.М., Амброс В. (июль 2003 г.). «Временная регуляция экспрессии микроРНК у Drosophila melanogaster, опосредованная гормональными сигналами и активностью генов широкого комплекса» . Биология развития . 259 (1): 9–18. дои : 10.1016/S0012-1606(03)00208-2 . ПМИД 12812784 . S2CID 17249847 .
- ^ Адамс Б.Д., Фюрно Х., Уайт Б.А. (май 2007 г.). «Микрорибонуклеиновая кислота (миРНК) миР-206 нацелена на альфа-рецептор эстрогена человека (ERalpha) и подавляет экспрессию информационной РНК и белка ERalpha в клеточных линиях рака молочной железы» . Молекулярная эндокринология . 21 (5): 1132–1147. дои : 10.1210/me.2007-0022 . ПМИД 17312270 .
- ^ Нолл М., Лодиш Х.Ф., Сан Л. (март 2015 г.). «Длинные некодирующие РНК как регуляторы эндокринной системы» . Обзоры природы. Эндокринология . 11 (3): 151–160. дои : 10.1038/nrendo.2014.229 . hdl : 1721.1/116703 . ПМЦ 4376378 . ПМИД 25560704 .
- ^ Ли В., Нотани Д., Ма К., Танаса Б., Нуньес Э., Чен А.Ю. и др. (июнь 2013 г.). «Функциональная роль энхансерных РНК в эстроген-зависимой активации транскрипции» . Природа . 498 (7455): 516–520. Бибкод : 2013Natur.498..516L . дои : 10.1038/nature12210 . ПМЦ 3718886 . ПМИД 23728302 .
- ^ «Исследователи выяснили, как черви передают знания о патогене потомству» . физ.орг . Проверено 11 октября 2020 г.
- ^ Калецкий Р., Мур Р.С., Врла Г.Д., Парсонс Л.Р., Гитай З., Мерфи К.Т. (октябрь 2020 г.). «C. elegans интерпретирует бактериальные некодирующие РНК, чтобы научиться избегать патогенов» . Природа . 586 (7829): 445–451. Бибкод : 2020Natur.586..445K . дои : 10.1038/s41586-020-2699-5 . ПМЦ 8547118 . ПМИД 32908307 . S2CID 221626129 .
- ^ Пибуэн Л., Виллауди Ж., Фербус Д., Мулерис М., Проспери М.Т., Ремвикос Ю., Губен Г. (февраль 2002 г.). «Клонирование мРНК со сверхэкспрессией при карциноме толстой кишки-1: последовательность, сверхэкспрессируемая в подмножестве карцином толстой кишки». Генетика рака и цитогенетика . 133 (1): 55–60. дои : 10.1016/S0165-4608(01)00634-3 . ПМИД 11890990 .
- ^ Фу X, Равиндранат Л., Тран Н., Петрович Г., Шривастава С. (март 2006 г.). «Регуляция апоптоза с помощью некодирующего гена, специфичного для простаты и связанного с раком простаты, PCGEM1». ДНК и клеточная биология . 25 (3): 135–141. дои : 10.1089/dna.2006.25.135 . ПМИД 16569192 .
- ^ Муртада-Маарабуни М., Пикард М.Р., Хедж В.Л., Фарзане Ф., Уильямс Г.Т. (январь 2009 г.). «GAS5, некодирующая белок РНК, контролирует апоптоз, и его активность снижается при раке молочной железы» . Онкоген . 28 (2): 195–208. дои : 10.1038/onc.2008.373 . ПМИД 18836484 .
- ^ Донг XY, Го П, Бойд Дж, Сунь X, Ли Q, Чжоу В, Донг JT (август 2009 г.). «Влияние мякРНК U50 на рак молочной железы человека» . Журнал генетики и геномики = И Чуань Сюэ Бао . 36 (8): 447–454. дои : 10.1016/S1673-8527(08)60134-4 . ПМЦ 2854654 . ПМИД 19683667 .
- ^ Христов К.П., Тривье Э., Круде Т. (март 2008 г.). «Некодирующие человеческие Y-РНК сверхэкспрессируются в опухолях и необходимы для пролиферации клеток» . Британский журнал рака . 98 (5): 981–988. дои : 10.1038/sj.bjc.6604254 . ПМК 2266855 . ПМИД 18283318 .
- ^ Фарх К.К., Гримсон А., Ян С., Льюис Б.П., Джонстон В.К., Лим Л.П. и др. (декабрь 2005 г.). «Широко распространенное влияние микроРНК млекопитающих на репрессию и эволюцию мРНК». Наука . 310 (5755): 1817–1821. Бибкод : 2005Sci...310.1817F . дои : 10.1126/science.1121158 . ПМИД 16308420 . S2CID 1849875 .
- ^ Лим Л.П., Лау Н.К., Гарретт-Энгеле П., Гримсон А., Шелтер Дж.М., Касл Дж. и др. (февраль 2005 г.). «Анализ микрочипов показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа . 433 (7027): 769–773. Бибкод : 2005Natur.433..769L . дои : 10.1038/nature03315 . ПМИД 15685193 . S2CID 4430576 .
- ^ Калин Г.А., Феррацин М., Чиммино А., Ди Лева Г., Симидзу М., Войчик С.Е. и др. (октябрь 2005 г.). «Сигнатура микроРНК, связанная с прогнозом и прогрессированием хронического лимфоцитарного лейкоза» . Медицинский журнал Новой Англии . 353 (17): 1793–1801. doi : 10.1056/NEJMoa050995 . ПМИД 16251535 .
- ^ Калин Г.А., Думитру С.Д., Симидзу М., Бичи Р., Зупо С., Ноч Э. и др. (ноябрь 2002 г.). «Частые делеции и подавление генов микроРНК miR15 и miR16 в 13q14 при хроническом лимфоцитарном лейкозе» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (24): 15524–15529. Бибкод : 2002PNAS...9915524C . дои : 10.1073/pnas.242606799 . ПМК 137750 . ПМИД 12434020 .
- ^ Ху З, Чен Дж, Тянь Т, Чжоу Х, Гу Х, Сюй Л и др. (июль 2008 г.). «Генетические варианты последовательностей микроРНК и выживаемость при немелкоклеточном раке легкого» . Журнал клинических исследований . 118 (7): 2600–2608. дои : 10.1172/JCI34934 . ПМК 2402113 . ПМИД 18521189 .
- ^ Шен Дж., Амброзон CB, Чжао Х. (март 2009 г.). «Новые генетические варианты генов микроРНК и семейный рак молочной железы» . Международный журнал рака . 124 (5): 1178–1182. дои : 10.1002/ijc.24008 . ПМИД 19048628 . S2CID 20960029 .
- ^ Цзя Б, Ван Цзы, Сунь Х, Чэнь Дж, Чжао Дж, Цю Икс (февраль 2019 г.). «Длинная некодирующая РНК LINC00707 губчат миР-370-3p, чтобы способствовать остеогенезу мезенхимальных стволовых клеток, полученных из костного мозга человека, посредством активации WNT2B» . Исследования и терапия стволовыми клетками . 10 (1): 67. дои : 10.1186/s13287-019-1161-9 . ПМК 6387535 . ПМИД 30795799 .
- ^ Ту Дж, Чжао З, Сюй М, Чен М, Венг Ц, Ван Дж, Цзи Дж (июль 2019 г.). «LINC00707 способствует прогрессированию гепатоцеллюлярной карциномы посредством губчатой миР-206 для увеличения CDK14». Журнал клеточной физиологии . 234 (7): 10615–10624. дои : 10.1002/jcp.27737 . ПМИД 30488589 . S2CID 54119752 . (Отозвано, см. дои : 10.1002/jcp.30712 , ПМИД 35297045 )
- ^ Се М, Ма Т, Сюэ Дж, Ма Х, Сунь М, Чжан Цз и др. (февраль 2019 г.). «Длинная межгенная небелковая кодирующая РНК 707 способствует пролиферации и метастазированию рака желудка путем взаимодействия со стабилизирующим мРНК белком HuR». Письма о раке . 443 : 67–79. дои : 10.1016/j.canlet.2018.11.032 . ПМИД 30502359 . S2CID 54611497 .
- ^ Ли Т, Ли Ю, Сунь Х (06 июня 2019 г.). «МикроРНК-876 состоит из длинной некодирующей РНК LINC00707 и напрямую нацелена на метадгерин, подавляя злокачественное развитие рака молочной железы» . Управление раком и исследования . 11 : 5255–5269. дои : 10.2147/cmar.s210845 . ПМК 6559252 . ПМИД 31239777 .
- ^ Юань Р.С., Бао Д., Чжан Ю. (май 2020 г.). «Linc00707 способствует пролиферации, инвазии и миграции клеток через регуляторную петлю miR-30c/CTHRC1 при раке молочной железы». Европейский обзор медицинских и фармакологических наук . 24 (9): 4863–4872. дои : 10.26355/eurrev_202005_21175 . ПМИД 32432749 . S2CID 218759508 .
- ^ Саху Т., дель Гаудио Д., Герман Дж.Р., Шинави М., Петерс С.У., Персон Р.Э. и др. (июнь 2008 г.). «Фенотип Прадера-Вилли, вызванный отцовским дефицитом малого ядрышкового РНК-кластера HBII-85 C/D» . Природная генетика . 40 (6): 719–721. дои : 10.1038/ng.158 . ПМК 2705197 . ПМИД 18500341 .
- ^ Скрябин Б.В., Губар Л.В., Сигер Б., Пфайффер Дж., Гендель С., Робек Т. и др. (декабрь 2007 г.). «Удаление кластера генов мякРНК MBII-85 у мышей приводит к постнатальной задержке роста» . ПЛОС Генетика . 3 (12): е235. дои : 10.1371/journal.pgen.0030235 . ПМЦ 2323313 . ПМИД 18166085 .
- ^ Дин Ф., Ли Х.Х., Чжан С., Соломон Н.М., Кампер С.А., Коэн П., Франк У (март 2008 г.). Акбарян С. (ред.). «Делегация SnoRNA Snord116 (Pwcr1/MBII-85) вызывает задержку роста и гиперфагию у мышей» . ПЛОС ОДИН . 3 (3): e1709. Бибкод : 2008PLoSO...3.1709D . дои : 10.1371/journal.pone.0001709 . ПМК 2248623 . ПМИД 18320030 .
- ^ Дин Ф., Принс Ю, Дхар М.С., Джонсон Д.К., Гарначо-Монтеро С., Николлс Р.Д., Франк У (июнь 2005 г.). «Отсутствие мякРНК Pwcr1/MBII-85 имеет решающее значение для неонатальной смертности в мышиных моделях синдрома Прадера-Вилли». Геном млекопитающих . 16 (6): 424–431. дои : 10.1007/s00335-005-2460-2 . ПМИД 16075369 . S2CID 12256515 .
- ^ Базелей П.С., Шепелев В., Талебизаде З., Батлер М.Г., Федорова Л., Филатов В., Федоров А. (январь 2008 г.). «snoTARGET показывает, что мишени мякРНК-сироты человека располагаются вблизи альтернативных соединений сплайсинга» . Джин . 408 (1–2): 172–179. дои : 10.1016/j.gene.2007.10.037 . ПМК 6800007 . ПМИД 18160232 .
- ^ Болтон П.Ф., Вельтман М.В., Вейсблатт Э., Холмс Дж.Р., Томас Н.С., Юингс С.А. и др. (сентябрь 2004 г.). «Хромосомные нарушения 15q11-13 и другие заболевания у людей с расстройствами аутистического спектра». Психиатрическая генетика . 14 (3): 131–137. дои : 10.1097/00041444-200409000-00002 . ПМИД 15318025 . S2CID 37344935 .
- ^ Кук Э.Х., Шерер С.В. (октябрь 2008 г.). «Вариации числа копий, связанные с нейропсихиатрическими состояниями». Природа . 455 (7215): 919–923. Бибкод : 2008Natur.455..919C . дои : 10.1038/nature07458 . ПМИД 18923514 . S2CID 4377899 .
- ^ Накатани Дж., Тамада К., Хатанака Ф., Исе С., Охта Х., Иноуэ К. и др. (июнь 2009 г.). «Аномальное поведение на мышиной модели, созданной с помощью хромосомной инженерии, для дупликации человеческого 15q11-13, наблюдаемой при аутизме» . Клетка . 137 (7): 1235–1246. дои : 10.1016/j.cell.2009.04.024 . ПМК 3710970 . ПМИД 19563756 .
- ^ Зиац М.Н., Реннерт О.М. (март 2013 г.). «Аберрантная экспрессия длинных некодирующих РНК в мозге аутистов» . Журнал молекулярной нейронауки . 49 (3): 589–593. дои : 10.1007/s12031-012-9880-8 . ПМЦ 3566384 . ПМИД 22949041 .
- ^ Риданпаа М., ван Эененнаам Х., Пелин К., Чедвик Р., Джонсон С., Юань Б. и др. (январь 2001 г.). «Мутации в РНК-компоненте РНКазы MRP вызывают плейотропное заболевание человека — хрящево-волосяную гипоплазию» . Клетка . 104 (2): 195–203. дои : 10.1016/S0092-8674(01)00205-7 . hdl : 2066/185709 . ПМИД 11207361 . S2CID 13977736 .
- ^ Мартин А.Н., Ли Ю. (март 2007 г.). «РНКаза MRP РНК и генетические заболевания человека» . Клеточные исследования . 17 (3): 219–226. дои : 10.1038/sj.cr.7310120 . ПМИД 17189938 .
- ^ Кавадас Ф.Д., Джилиани С., Гу Й., Маццолари Е., Бейтс А., Пегояни Е. и др. (декабрь 2008 г.). «Вариабельность клинических и лабораторных особенностей у пациентов с мутациями гена эндорибонуклеазы, процессирующей рибонуклеазную митохондриальную РНК» . Журнал аллергии и клинической иммунологии . 122 (6): 1178–1184. дои : 10.1016/j.jaci.2008.07.036 . ПМИД 18804272 .
- ^ Фагихи М.А., Модарреси Ф., Халил А.М., Вуд Д.Э., Сааган Б.Г., Морган Т.Е. и др. (июль 2008 г.). «Экспрессия некодирующей РНК повышается при болезни Альцгеймера и приводит к быстрой прямой регуляции бета-секретазы» . Природная медицина . 14 (7): 723–730. дои : 10.1038/nm1784 . ПМЦ 2826895 . ПМИД 18587408 .
- ^ Менсия А., Модамио-Хойбьор С., Редшоу Н., Морин М., Майо-Мерино Ф., Олаварриета Л. и др. (май 2009 г.). «Мутации в семенной области человеческой миР-96 ответственны за несиндромальную прогрессирующую потерю слуха». Природная генетика . 41 (5): 609–613. дои : 10.1038/ng.355 . ПМИД 19363479 . S2CID 11113852 .
- ^ Льюис М.А., Квинт Э., Глейзер А.М., Фукс Х., Де Анджелис М.Х., Лэнгфорд С. и др. (май 2009 г.). «Вызванная ENU мутация миР-96, связанная с прогрессирующей потерей слуха у мышей» . Природная генетика . 41 (5): 614–618. дои : 10.1038/ng.369 . ПМК 2705913 . ПМИД 19363478 .
- ^ Соукуп Г.А. (июнь 2009 г.). «Мало, но громко: малые РНК оказывают сильное влияние на развитие ушей» . Исследования мозга . 1277 : 104–114. дои : 10.1016/j.brainres.2009.02.027 . ПМК 2700218 . ПМИД 19245798 .
- ^ Тейлор Р.В., Тернбулл DM (май 2005 г.). «Мутации митохондриальной ДНК при заболеваниях человека» . Обзоры природы. Генетика . 6 (5): 389–402. дои : 10.1038/nrg1606 . ПМК 1762815 . ПМИД 15861210 .
- ^ Ярхэм Дж.В., Элсон Дж.Л., Блейкли Э.Л., МакФарланд Р., Тейлор Р.В. (сентябрь 2010 г.). «Мутации и болезни митохондриальной тРНК». Междисциплинарные обзоры Wiley. РНК . 1 (2): 304–324. дои : 10.1002/wrna.27 . ПМИД 21935892 . S2CID 43123827 .
- ^ Зифа Э., Джаннули С., Теотокис П., Стаматис С., Мамурис З., Статопулос С. (январь 2007 г.). «Мутации митохондриальной тРНК: клинические и функциональные нарушения» . Биология РНК . 4 (1): 38–66. дои : 10.4161/rna.4.1.4548 . ПМИД 17617745 . S2CID 11965790 .
- ^ Эбботт Дж.А., Франклин К.С., Роби-Бонд С.М. (2014). «Переносная РНК и болезни человека» . Границы генетики . 5 : 158. дои : 10.3389/fgene.2014.00158 . ПМК 4042891 . ПМИД 24917879 .
- ^ Картер Р.Дж., Дубчак И., Холбрук С.Р. (октябрь 2001 г.). «Вычислительный подход к идентификации генов функциональных РНК в геномных последовательностях» . Исследования нуклеиновых кислот . 29 (19): 3928–3938. дои : 10.1093/нар/29.19.3928 . ПМК 60242 . ПМИД 11574674 .
- ^ Педерсен Дж.С., Беджерано Г., Сипель А., Розенблум К., Линдблад-Тох К., Ландер Э.С. и др. (апрель 2006 г.). «Идентификация и классификация консервативных вторичных структур РНК в геноме человека» . PLOS Вычислительная биология . 2 (4): е33. Бибкод : 2006PLSCB...2...33P . дои : 10.1371/journal.pcbi.0020033 . ПМЦ 1440920 . ПМИД 16628248 .
- ^ Томас Дж.М., Хорспул Д., Браун Дж., Черепанов В., Аптон С. (январь 2007 г.). «GraphDNA: программа Java для графического отображения результатов анализа состава ДНК» . БМК Биоинформатика . 8:21 . дои : 10.1186/1471-2105-8-21 . ПМЦ 1783863 . ПМИД 17244370 .
- ^ Брозиус Дж., Раабе, Калифорния (февраль 2015 г.). «Что такое РНК? Верхний уровень классификации РНК» . Биология РНК . 13 (2): 140–144. дои : 10.1080/15476286.2015.1128064 . ПМЦ 4829331 . ПМИД 26818079 .
- ^ Джи З, Сонг Р, Регев А, Струл К (декабрь 2015 г.). «Многие днРНК, 5'UTR и псевдогены транслируются, и некоторые из них, вероятно, экспрессируют функциональные белки» . электронная жизнь . 4 : e08890. doi : 10.7554/eLife.08890 . ПМЦ 4739776 . ПМИД 26687005 .
- ^ Тосар Дж. П., Ровира С., Кайота А (22 января 2018 г.). «Некодирующие фрагменты РНК составляют большинство аннотированных piRNA, экспрессируемых в соматических негонадных тканях» . Коммуникационная биология . 1 (1): 2. дои : 10.1038/s42003-017-0001-7 . ПМК 6052916 . ПМИД 30271890 .
- ^ Хаусман Г., Улицкий И. (январь 2016 г.). «Методы различия между кодирующими белок и длинными некодирующими РНК и неуловимая биологическая цель трансляции длинных некодирующих РНК». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1859 (1): 31–40. дои : 10.1016/j.bbagrm.2015.07.017 . ПМИД 26265145 .
Внешние ссылки
[ редактировать ]- «РНКдб» . Архивировано из оригинала 29 августа 2007 г.
Эта база данных представляет собой комплексную базу данных некодирующих РНК млекопитающих (RNAdb).
- База данных Rfam — тщательно подобранный список сотен семейств родственных нкРНК.
- NONCODE.org — бесплатная база данных всех видов некодирующих РНК (кроме тРНК и рРНК).
- RNAcon Прогнозирование и классификация нкРНК BMC Genomics 2014, 15:127
- Обозреватель потоков ENCODE. Характеристика некодирующих РНК. Природа (журнал)
- Ресурс баз данных некодирующих РНК (NRDR) — тщательно подобранный источник данных, связанных с базами данных по некодирующим РНК, доступными в Интернете.
- DASHR - база данных малых некодирующих РНК Биоинформатика 2018
Виды нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Составляющие | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновая кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||