Белок-носитель митохондрий мозга 1
| SLC25A14 | ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | SLC25A14 , семейство 25 переносчиков растворенных веществ (митохондриальный носитель, мозг), член 14, BMCP1, UCP5, семейство 25 переносчиков растворенных веществ, член 14 | |||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 300242 ; МГИ : 1330823 ; Гомологен : 2937 ; Генные карты : SLC25A14 ; OMA : SLC25A14 — ортологи | |||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||






Белок-носитель 1 митохондрий мозга — это белок , который у человека кодируется SLC25A14 геном . [ 5 ] [ 6 ]
Митохондриальные разобщающие белки (UCP) являются членами более крупного семейства митохондриальных белков-переносчиков анионов (MACP). UCP отделяют окислительное фосфорилирование от синтеза АТФ, при этом энергия рассеивается в виде тепла, что также называется утечкой митохондриальных протонов. UCP облегчают перенос анионов с внутренней на внешнюю митохондриальную мембрану и обратный перенос протонов с внешней на внутреннюю митохондриальную мембрану. Они также снижают мембранный потенциал митохондрий в клетках млекопитающих. Тканевая специфичность наблюдается для различных UCP, и точные методы переноса UCP H+/OH- неизвестны. UCP содержат три гомологичных белковых домена MACP. Этот ген широко экспрессируется во многих тканях, наибольшее его количество — в головном мозге и семенниках. Генный продукт имеет N-концевой гидрофобный домен, которого нет в других UCP. Для этого гена были обнаружены два варианта сплайсинга. [ 6 ]
Митохондриальные разобщающие белки играют значительную роль в геноме человека, а также в геномах растений, прокариот, грибов и других эукариот, включая всех млекопитающих. Люди содержат пять различных изоформ разобщающих белков, и каждый UCP выполняет свою собственную функцию в организме человека; однако все они жизненно важны для генома. Предыдущие исследования продемонстрировали, как снижение окислительно-восстановительной передачи сигналов и специфические UCP играют нейропротекторную роль, способную защитить от ряда нейродегенеративных заболеваний, включая гипоксию, ишемию и такие заболевания, как болезнь Альцгеймера, болезнь Паркинсона и шизофрения.
УКП1
UCP1 был обнаружен у некоторых представителей Rodentia, но в основном в бурой жировой ткани, обнаруженной у младенцев, и играет роль в энергетическом гомеостазе. Местоположение UCP1 находится во внутренней мембране митохондрий. Бурая жировая ткань организма предотвращает гипотермию, высвобождая движущую силу протонов в результате окислительного фосфорилирования. Таким образом, UCP1 контролирует как независимый, так и термогенный метаболизм. Заболевания, связанные с UCP1, включают диабет 2 типа и кардиометаболические заболевания.
ОГП 2
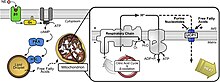
UCP2 выполняет различные функции в организме, например, в мозге, легких, почках, селезенке, сердце и скелетных мышцах. Кроме того, в нескольких областях центральной нервной системы наблюдается стабильно высокий уровень экспрессии мРНК UCP2. Непосредственные нейроны, вторгающиеся или спящие моноциты, клетки хориоидального сплетения и эндотелиальные клетки связаны с повышенными уровнями мРНК UCP2. Митохондриально-ассоциированные заболевания оказывают нейропротекторное действие при некоторых нейродегенеративных заболеваниях, включая смерть нейронов, вызванную СД2, ожирение SCZ, воспалительные заболевания, рак и церебральную ишемию (Kumar, 2022). Из-за неисправности митохондрий и изменений синаптической пластичности с UCP2 связаны многочисленные заболевания. В поврежденном участке мозга после черепно-мозговой травмы экспрессия UP2 повышается. В мозге основная роль UCP2 заключается в пластичности и выживании нейронов. В частности, исследования показывают, что при ишемии головного мозга UCP2 регулирует выработку АФК, нейровоспаление, митохондриальный потенциал, энергетический баланс, нейроэндокринную функцию и гибель клеток. Другая важная функция UCP2 — контроль температуры микроокружения пресинаптического терминала. Согласно исследованиям, UCP2 имеет решающее значение для биоэнергетической адаптации нейронов к увеличению активности нейронов и синаптической пластичности. В результате данные различных исследований позволяют предположить, что UCP2 является вероятной терапевтической мишенью для лечения неврологических заболеваний. УКП3
UCP3 в основном экспрессируется в жировой ткани, легких и скелетных мышцах; однако он также обнаруживается в мозге, почках, селезенке и сердце. Функции этого UCP заключаются в регулировании скорости метаболизма в состоянии покоя, регулировании метаболизма жирных кислот, предотвращении диабетической нейропатии путем предотвращения потери нервов, ограничении уровней свободных радикалов в клетках и ограничении уровня АФК. UCP3 также снижает риск диабета 1 типа, указывая на то, что UCP предотвращают митохондриально-опосредованное повреждение нейронов (Kumar, 2022). UCP 1, 2 и 3 обладают способностью контролировать элементы, на которые влияет энергетический баланс, такие как агонисты 3-адренорецепторов, прием пищи и воздействие холода. Наконец, с UCP3 связаны сердечно-сосудистые заболевания.
ОГП 4 и 5
Центральная нервная система является основным местом экспрессии UCP 4 и 5, и каждый UCP имеет отдельную функцию.
UCP4 поддерживает гомеостаз за счет повышения активности митохондриального комплекса II в клетках нейробластомы SH-SY5Y, усиления окисления жирных кислот и чувствительности к инсулину, а также снижения потенциала митохондриальной мембраны в астроцитах, тем самым поддерживая выживаемость нейронов. UCP4 связан с нейродегенеративными заболеваниями, такими как болезнь Альцмира, Паркинсона и шиперферия, СД2, а также с индуцированной гибелью нейронов (Kumar, 2022). UCP4 расположен в таких областях мозга, как гиппокамп, черная субстанция и полосатое тело.
UCP5 контролирует гомеостаз путем увеличения синтеза АТФ, поляризации митохондрий и предотвращения апоптоза. Он также поддерживает потенциал митохондриальной мембраны и уменьшает окислительный стресс против токсичности 1-метил-4-фенилпиридиния MPP+ и дофамина в клетках SH-SY5Y. Экспрессия UCP5 происходит в нейронах и клетках астроцитов, и это BMCP1, митохондриальный белок-переносчик головного мозга. UCP5 связан со старением и теми же нейродегенеративными заболеваниями, что и UCP4.
Структура
Как упоминалось выше, функции и распределение пяти изоформ UCP различаются; некоторые из них расположены в одной хромосоме у людей, крыс и мышей. UCP1 расположен на хромосомах 4, 19 и 8. UCP2 обнаружен на хромосомах 11, 1 и 7. Кроме того, UCP2 имеет сходство с UCP 1, 3, 4 и 5.
Нейродегенеративные заболевания
Во всем мире нейродегенеративные расстройства представляют серьезную угрозу для общественного здравоохранения. Наряду с окислительным стрессом, нейровоспалением и дисбалансом энергетического гомеостаза ключевым фактором является митохондриальная дисфункция. Одним из наиболее примитивных и решающих событий в патогенезе болезни Альцгеймера является окислительный стресс. Активные формы кислорода и индуцибельная синтаза оксида азота производятся преимущественно в митохондриях, что при болезни Альцгеймера вызывает образование активных форм кислорода и в конечном итоге приводит к гибели нейронов (Kumar 2022). Факторы созревания глии, обнаруженные в глиальных клетках, связаны с подавлением UCP 2 и 4. Провоспалительный белок, известный как фактор созревания глии, был обнаружен рядом с амилоидными бляшками и нейрофибриллярными клубками при болезни Альцгеймера. Это связано с экспрессией индуцибельной синтазы оксида азота и NF-kB.
В различных областях мозга при болезни Альцгеймера наблюдается повышенная экспрессия GMF. Было показано, что нейровоспаление является фактором, способствующим нейродегенеративным заболеваниям, а UCP2 был идентифицирован как основной регулятор пластичности M1 и M2 в отношении защитного фенотипа при заболеваниях головного мозга. Повышенное производство активных форм кислорода в UCP 2 и 4 приводит к развитию болезни Альцгеймера из-за увеличения вероятности апоптоза.
Болезнь Паркинсона обычно поражает взрослых старше 60 лет; однако исследования показывают, что травма головного мозга у людей в возрасте от 18 до 30 лет может вызвать это состояние. Более 90% случаев болезни Паркинсона являются изолированными, однако менее 10% являются генетическими. Симптомы болезни Паркинсона включают тремор, замедление движений, нарушение равновесия и другие расстройства, сопровождающие болезнь Паркинсона. Этот симптом возникает из-за потери дофаминергических нейронов во время выведения дофаминовых нейротрансмиттеров. Поскольку UCP4 является потенциальным регулятором гомеостаза кальция, блокирование кальциевых каналов, которые снижают окислительный стресс и останавливают гибель нейрональных клеток, предотвращает экскрецию кальция в митохондрии. Утечка протонов и потребление кислорода увеличиваются из-за увеличения производства UCP5, что приводит к снижению митохондриального потенциала и синтеза АТФ.
Старение, гипоксия (повреждение головного мозга) и диабет 2 типа соответствуют UCP 2 и 4. Старение мозга является фактором риска нейродегенеративных заболеваний, а сверхэкспрессия UCP2 может остановить процесс старения и остановить потерю нейронов. Гипоксия возникает, когда в тканях снижается содержание кислорода, что вызывает когнитивные нарушения. Уровни UCP4 повышаются в ответ на воздействие окружающей среды у людей, защищая мозг от повреждений. Апопостис способствует ишемии головного мозга и необходим для гомеостаза кальция, высвобождения свободных радикалов, образования активных форм кислорода (АФК) и энергетического метаболизма. Поскольку дальнейшие исследования все еще проводятся, исследования показывают, что UCP1 и UCP2 связаны с диабетом 2 типа.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000102078 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031105 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Санчис Д., Флери С., Чомики Н., Губерн М., Хуанг К., Неверова М. и др. (декабрь 1998 г.). «BMCP1, новый митохондриальный носитель с высокой экспрессией в центральной нервной системе человека и грызунов, а также активностью разобщения дыхания у рекомбинантных дрожжей» . Журнал биологической химии . 273 (51): 34611–34615. дои : 10.1074/jbc.273.51.34611 . ПМИД 9852133 .
- ^ Jump up to: а б «Ген Энтрез: семейство 25 растворенных носителей SLC25A14 (митохондриальный носитель, мозг), член 14» .
Дальнейшее чтение
[ редактировать ]- Рикье Д., Буйо Ф. (январь 2000 г.). «Гомологи разобщающих белков: UCP1, UCP2, UCP3, StUCP и AtUCP» . Биохимический журнал . 345 Ч. 2 (Часть 2): 161–179. дои : 10.1042/0264-6021:3450161 . ПМК 1220743 . ПМИД 10620491 .
- Езек П., Урбанкова Э. (январь 2000 г.). «Конкретная последовательность мотивов митохондриальных разобщающих белков» . ИУБМБ Жизнь . 49 (1): 63–70. дои : 10.1080/713803586 . ПМИД 10772343 .
- Ю XX, Мао В., Чжун А., Шоу П., Браш Дж., Шервуд С.В. и др. (август 2000 г.). «Характеристика новых изоформ UCP5/BMCP1 и дифференциальная регуляция экспрессии UCP4 и UCP5 посредством диетических или температурных манипуляций» . Журнал ФАСЭБ . 14 (11): 1611–1618. дои : 10.1096/fj.14.11.1611 . ПМИД 10928996 .
- Ким-Хан Дж.С., Райхерт С.А., Квик К.Л., Дуган Л.Л. (ноябрь 2001 г.). «BMCP1: митохондриальный разобщающий белок в нейронах, который регулирует функцию митохондрий и выработку оксидантов» . Журнал нейрохимии . 79 (3): 658–668. дои : 10.1046/j.1471-4159.2001.00604.x . ПМИД 11701769 .
- Ян X, Пратли Р.Э., Токракс С., Татаранни П.А., Пермана П.А. (апрель 2002 г.). «Изоформы транскрипта UCP5/BMCP1 в скелетных мышцах человека: связь изоформ с короткой вставкой с окислением липидов и скоростью метаболизма в состоянии покоя» . Молекулярная генетика и обмен веществ . 75 (4): 369–373. дои : 10.1016/S1096-7192(02)00008-2 . ПМИД 12051969 .
- Хо П.В., Чу А.С., Квок К.Х., Кунг М.Х., Рамсден Д.Б., Хо С.Л. (ноябрь 2006 г.). «Нокдаун разобщающего белка-5 в нейрональных клетках SH-SY5Y: влияние на MPP+-индуцированную деполяризацию митохондриальной мембраны, дефицит АТФ и окислительную цитотоксичность». Журнал нейробиологических исследований . 84 (6): 1358–1366. дои : 10.1002/jnr.21034 . ПМИД 16941493 . S2CID 24763520 .
- Ясуно К., Андо С., Мисуми С., Макино С., Кульски Дж.К., Муратаке Т. и др. (март 2007 г.). «Синергическая ассоциация генов митохондриального разобщающего белка (UCP) с шизофренией». Американский журнал медицинской генетики. Часть B. Нейропсихиатрическая генетика . 144Б (2): 250–253. дои : 10.1002/ajmg.b.30443 . ПМИД 17066476 . S2CID 30129988 .
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .
Эта мембранных белках статья о незавершена . Вы можете помочь Википедии, расширив ее . |