Рецептор, похожий на Toll
| Рецептор, похожий на Toll | |
|---|---|
 Изогнутая богатая лейцином повторная область рецепторов, подобных платежам, представленная здесь TLR3 | |
| Идентификаторы | |
| Символ | Рецептор, похожий на Toll |
| Мембраном | 7 |
| PIRSF037595 | |
Toll-подобные рецепторы ( TLR ) представляют собой класс белков , которые играют ключевую роль в врожденной иммунной системе . Они представляют собой однопространенные рецепторы, обычно экспрессируемые на дозорных клетках, таких как макрофаги и дендритные клетки , которые распознают структурно консервативные молекулы, полученные из микробов . Как только эти микробы достигли физических барьеров, таких как кожи или кишечного тракта слизистая оболочка , они распознаются TLR, которые активируют иммунных клеток реакции . TLR включают TLR1 , TLR2 , TLR3 , TLR4 , TLR5 , TLR6 , TLR7 , TLR8 , TLR9 , TLR10 , TLR11 , TLR12 и TLR13 . У людей не хватает генов для TLR11, TLR12 и TLR13 [ 1 ] и мышам не хватает функционального гена для TLR10. [ 2 ] Рецепторы TLR1, TLR2, TLR4, TLR5, TLR6 и TLR10 расположены на клеточной мембране , тогда как TLR3, TLR7, TLR8 и TLR9 расположены во внутриклеточных везикулах (поскольку они являются датчиками ядерных кислот ). [ 3 ]
TLRS получила свое имя от их сходства с белком, кодируемом геном Toll . [ 4 ]
Функция
[ редактировать ]Способность иммунной системы распознавать молекулы , которые широко распространены патогенами , частично, из-за присутствия иммунных рецепторов , называемых платных рецепторов (TLR), которые экспрессируются на мембранах лейкоцитов , включая дендритные клетки , макрофаги , натуральные Клетки-киллер , клетки адаптивных иммунитет Т-клеток и В-клетки , а также неиммунные клетки ( эпителиальные и эндотелиальные клетки и фибробласты ). [ 5 ]
Связывание лигандов - либо в форме адъюванта, используемого при вакцинациях , либо в форме инвазивных фрагментов во время естественной инфекции - к TLR отмечает ключевые молекулярные события, которые в конечном итоге приводят к врожденным иммунным реакциям и развитию антиген -специфических приобретенных приобретенных иммунитет. [ 6 ] [ 7 ]
После активации TLR рекатации адаптерных белков (белки, которые опосредуют другие белковые взаимодействия) в цитозоле иммунной клетки, чтобы распространять антиген-индуцированный путь передачи сигнала . Эти рекрутированные белки затем ответственны за последующую активацию других нижестоящих белков, включая протеинкиназы (IKKI, IRAK1 , IRAK4 и TBK1 ), которые дополнительно усиливают сигнал и в конечном итоге приводят к повышению регуляции или подавления генов , которые организуют воспаление и другие транскрипцию и другие события. Некоторые из этих событий приводят к производству цитокинов , пролиферации и выживанию, в то время как другие приводят к большему адаптивному иммунитету. [ 7 ] Если лиганд является бактериальным фактором, патоген может быть фагоцитозирован и расщеплен, а его антигены представлены в CD4+ T -клетках . В случае вирусного фактора инфицированная клетка может отключить синтез своего белка и может подвергаться запрограммированной гибели клеток ( апоптоз ). Иммунные клетки, которые обнаружили вирус, также могут выделять антивирусные факторы, такие как интерфероны .
Также было показано, что рецепторы, подобные платежам, являются важной связью между врожденным и адаптивным иммунитетом посредством их присутствия в дендритных клетках . [ 8 ] Флагеллин , лиганд TLR5, индуцирует секрецию цитокинов при взаимодействии с TLR5 на Т -клетках человека. [ 8 ]
Суперсемейство
[ редактировать ]
TLR представляют собой тип рецептора распознавания паттернов (PRR) и распознают молекулы, которые широко распространены патогенами , но различимы от молекул хозяина, в совокупности называемых патогенными молекулярными паттернами (PAMPS). В дополнение к распознаванию экзогенных PAMP, TLR также могут связываться с эндогенными молекулярными паттернами, связанными с эндогенными повреждениями (DAMP), такими как белки теплового шока (HSP) или компоненты плазматической мембраны. [ 9 ] TLR вместе с рецепторами интерлейкина-1 образуют рецепторную суперсемейство , известное как «рецептор-рецептор интерлейкина-1 / суперсемейство рецептора»; У всех членов этой семьи общий так называемый домен TIR (рецептор Toll-IL-1).
Существуют три подгруппы доменов TIR. Белки с доменами TIR подгруппа 1 представляют собой рецепторы для интерлейкинов , которые продуцируются макрофагами , моноцитами и дендритными клетками и все имеют внеклеточные домены иммуноглобулина (IG). Белки с доменами TIR подгруппы 2 являются классическими TLR и прямо или косвенно связываются с молекулами микробного происхождения. Третья подгруппа белков, содержащих домены TIR, состоит из адаптерных белков , которые являются исключительно цитозольными и опосредуют передачу сигналов из белков подгрупп 1 и 2.
Большая семья
[ редактировать ]В этом разделе отсутствует информация о TLR choanoflagellate (PMID29848444). ( Декабрь 2021 г. ) |
TLR присутствуют как у позвоночных, так и в беспозвоночных . Молекулярные строительные блоки TLR представлены в бактериях и у растений, а рецепторы распознавания растителей, как известно, требуются для защиты хозяина от инфекции. Таким образом, TLR, по -видимому, являются одним из самых древних, консервативных компонентов иммунной системы .
В последние годы TLR были идентифицированы также в нервной системе млекопитающих. Члены семейства TLR были обнаружены на глиа, нейронах и на клетках нейронных предшественников, в которых они регулируют решение клеток. [ 10 ]
Было подсчитано, что большинство видов млекопитающих имеют от десяти до пятнадцати типов рецепторов, похожих на платные,. Тринадцать TLR (названные просто TLR1 до TLR13) были идентифицированы у людей и мышей вместе, и эквивалентные формы многих из них были обнаружены у других видов млекопитающих. [ 11 ] [ 12 ] [ 13 ] Однако эквиваленты определенного TLR, обнаруженного у людей, не присутствуют во всех млекопитающих. Например, ген, кодирующий белок, аналогичный TLR10 у людей, присутствует у мышей , но, по -видимому, был поврежден в какой -то момент в прошлом ретровирусом . С другой стороны, мыши экспресс 11, 12 и 13, ни один из которых не представлен у людей. Другие млекопитающие могут выразить TLR, которые не встречаются у людей. Другие виды, не являющиеся млекопитающими, могут отличать TLR от млекопитающих, как продемонстрировано антиклеточной стенкой TLR14 , которая встречается в Takifugu Pufferfish. [ 14 ] Это может усложнить процесс использования экспериментальных животных в качестве моделей человеческого врожденного иммунитета.
TLR позвоночных делятся на сходство на семейства TLR 1/2/6/10/14/15, TLR 3, TLR 4, TLR 5, TLR 7/8/9 и TLR 11/12/13/16/21 /22/23. [ 14 ]
TLRS в Drosophila Immunity
[ редактировать ]
Участие передачи сигналов в иммунитете впервые было продемонстрировано в фруктовой мух, Drosophila melanogaster . [ 19 ] Фруктовые мухи имеют только врожденные иммунные ответы, позволяющие исследованиям избежать вмешательства адаптивных иммунных механизмов при передаче сигнала. Реакция мухи на грибковую или бактериальную инфекцию происходит через два отдельных сигнальных каскада, одним из которых является платный путь, а другой - путь иммунного дефицита . Платный путь похож на передачу сигналов TLR млекопитающих, но в отличие от TLR млекопитающих, платы не активируются непосредственно с помощью патогенов, ассоциированных с молекулярными паттернами ( PAMP ). Его рецепторный эктодомен распознает расщепленную форму цитокинового спени, которая секретируется в гемолимфе как неактивный димерный предшественник. Рецептор TOLL разделяет цитоплазматический домен TIR с TLR млекопитающих, но эктодомен и внутрицитоплазматический хвост разные. Это различие может отражать функцию этих рецепторов в виде цитокиновых рецепторов, а не PRR .
Платный путь активируется различными стимулами, такими как грамположительные бактерии , грибы и факторы вирулентности . [ 17 ] [ 20 ] Во -первых, фермент обработки Spätzle (SPE) активируется в ответ на инфекцию и расщепляет Spätzle ( SPZ ). Затем расщепленный Spätzle связывается с рецептором Toll и сшивает его эктодомены. Это запускает конформационные изменения в рецепторе, что приводит к передаче сигналов с помощью потери. С этого момента сигнальный каскад очень похож на передачу сигналов млекопитающих через TLR. Сигнальный комплекс, индуцированный Toll (TICS), состоит из MyD88 , Tube и Pelle (ортолог IRAK млекопитающих). Сигнал из тиков затем трансдуцируется кактусу (гомолог IκB DIF (дорсальный фактор иммунитета; гомолог NF-κB млекопитающего), фосфорилированный кактус является полиубиквиквизированным и разлагается, что позволяет ядерная транслокация млекопитаю Пептиды (ампер), такие как дрозомицин . [ 21 ]
Drosophila сложности 9 имеют в общей генов семейства SPZ , которые взаимодействуют друг с другом в разной степени. [ 22 ]
TLR2
[ редактировать ]TLR2 также был обозначен как CD282 (кластер дифференцировки 282).
TLR3
[ редактировать ]TLR3 не использует зависимый путь MyD88. Его лигандом представляет собой ретровирусную двухцепочечную РНК ( дцРНК ), которая активирует сигнальный путь, зависящий от TRIF . Чтобы исследовать роль этого пути в ретровирусном перепрограммировании, были получены методы спускания TLR3 или TRIF, и результаты показали, что только путь TLR3 необходим для полной индукции экспрессии гена -мишеней вектором ретровирусного экспрессии. Эта ретровирусная экспрессия четырех транскрипционных факторов ( OCT4 , SOX2 , KLF4 и C-MYC ; OSKM) индуцирует плюрипотентность в соматических клетках. Это подтверждается исследованием, которое показывает, что эффективность и количество генерации IPSC человека с использованием ретровирусных векторов снижаются путем нокдауна пути с ингибиторами пептидов или ShRNA нокдауном TLR3 или его адаптерным белком TRIF. Взятые вместе, стимуляция TLR3 вызывает серьезные изменения в ремоделировании хроматина и ядерном перепрограммировании, и для этих изменений требуется активация воспалительных путей, индукция генов плюрипотентности и генерации колоний плюрипотентных стволовых клеток (IPSC). [ 23 ]
TLR11
[ редактировать ]Как отмечалось выше, клетки человека не экспрессируют TLR11 , но клетки мышей делают. Специфичный для мыши TLR11 распознает уропатогенный E.Coli и апикомпетксан-паразит токсоплазма Gondii . С токсоплазмой его лиганд является белковым профилин, а лигандом для кишечной палочки является флагеллин . Флагеллин из энтеропатогенной сальмонеллы также распознается TLR11. [ 24 ]
Поскольку мыши TLR11 способны эффективно распознавать сальмонеллу , нормальные мыши не заражаются пероральной сальмонеллой Typhi , что вызывает пищевую и водяную гастроэнтерит и брюшную лихорадку у людей. дефицитом TLR11 Мыши с , с другой стороны, эффективно заражены. В результате эта нокаутная мышь может действовать как модель заболевания человеческой брюшной лихорадки. [ 25 ]
Резюме известных TLR млекопитающих
[ редактировать ]Toll-подобные рецепторы связывают и активируются различными лигандами, которые, в свою очередь, расположены на разных типах организмов или структур. У них также есть разные адаптеры, чтобы реагировать на активацию и иногда расположены на клеточной поверхности, а иногда и во внутренних ячеек . [ 26 ] Кроме того, они экспрессируются различными типами лейкоцитов или другими типами клеток :
| Рецептор | Лиганд (ы) [ 27 ] | Расположение лиганда [ 27 ] | Адаптер (ы) | Расположение | Типы клеток [ 27 ] |
|---|---|---|---|---|---|
| TLR 1 | Множественные триацилпопептиды | Бактериальный липопротеин | Myd88 /mal | клеточная поверхность |
|
| TLR 2 | Множественные гликолипиды | Бактериальные пептидогликаны | Myd88/mal | клеточная поверхность |
|
| множественные липопептиды и протеолипиды | Бактериальные пептидогликаны | ||||
| липотеховая кислота | Грамположительные бактерии | ||||
| HSP70 | Клетки -хозяина | ||||
| зимозан ( бета-глюкан ) | Грибы | ||||
| Многочисленные другие | |||||
| TLR 3 | двухцепочечная РНК , Poly I: C | вирусы | Триф | клеточный отсек |
|
| TLR 4 | липополисахарид | Грамотрицательные бактерии | Myd88/mal/ trif /tram | клеточная поверхность |
|
| Несколько белков теплового шока | Бактерии и клетки -хозяев | ||||
| Фибриноген | клетки -хозяина | ||||
| гепаран сульфата фрагменты | клетки -хозяина | ||||
| гиалуроновой кислоты Фрагменты | клетки -хозяина | ||||
| никель [ 32 ] | |||||
| Различные опиоидные препараты | |||||
| TLR 5 | Бактериальный флагеллин | Бактерии | Myd88 | клеточная поверхность |
|
| Профиль [ 33 ] | Toxoplasma gondii | ||||
| TLR 6 | множественные диацилпопептиды | Микоплазма | Myd88/mal | клеточная поверхность |
|
| TLR 7 | Имидазохинолин | Маленькие синтетические соединения | Myd88 | клеточный отсек |
|
| локсорибин ( аналог гуанозина ) | |||||
| Бропиримин | |||||
| Resiquimod | |||||
| одноцепочечная РНК | РНК -вирусы | ||||
| TLR 8 | небольшие синтетические соединения; одноцепочечная вирусная РНК, фагоцитизированная бактериальная РНК (24) | Myd88 | клеточный отсек |
| |
| TLR 9 | неметилированная CPG олигодезоксинуклеотидная ДНК | Бактерии, ДНК -вирусы | Myd88 | клеточный отсек |
|
| TLR 10 | трицилированные липопептиды [ 35 ] | неизвестный | клеточная поверхность | ||
| TLR 11 | Профиль | Toxoplasma gondii [ 39 ] | Myd88 | клеточный отсек [ 40 ] |
|
| Флагеллин | Бактерии ( E. coli , сальмонелла ) [ 24 ] | ||||
| TLR 12 | Профиль | Toxoplasma gondii [ 41 ] | Myd88 | клеточный отсек |
|
| TLR 13 [ 43 ] [ 44 ] | Бактериальная рибосомальная РНК -последовательность "CGGAAAGACC" (но не метилированная версия) [ 45 ] | Вирус, бактерии | MyD88, TAK-1 | клеточный отсек |
|
Лиганды
[ редактировать ]
Из-за специфичности рецепторов, похожих на Toll (и других врожденных иммунных рецепторов), они не могут быть легко изменены в ходе эволюции, эти рецепторы распознают молекулы, которые постоянно связаны с угрозами (то есть патогеном или клеточным стрессом) и очень специфичны к Эти угрозы (то есть не могут быть приняты за самостоятельные молекулы, которые обычно выражаются в физиологических условиях). Считается, что молекулы, связанные с патогеном, которые отвечают этим требованиям, имеют решающее значение для функции патогена и трудно изменить с помощью мутации; Говорят, что они эволюционно сохраняются. Несколько консервативных признаков в патогенах включают бактериальные клеточные липополисахариды (LPS), липопротеины , липопептиды и липоарабиноманнан ; белки, такие как флагеллин из бактериальной жгутики ; двухцепочечная РНК вирусов; или неметилированные острова CPG бактериальной и вирусной ДНК ; а также островов CPG, найденных в промоторах эукариотической ДНК; а также некоторые другие молекулы РНК и ДНК. Поскольку лиганды TLR присутствуют в большинстве патогенных микроорганизмов, они также могут присутствовать в вакцинах, полученных из патогенов (например, MMR, гриппа, вакцины полиомиелита), большинство коммерчески доступных вакцин были оценены для их неотъемлемых лигандов TLR активировать отдельные подгруппы иммунных клеток. [ 46 ] [ 47 ] Для большинства TLR специфичность распознавания лигандов в настоящее время была установлена с помощью гена (также известного как «нокаут гена»): метод, с помощью которого отдельные гены могут быть избирательно удалены у мышей. [ 48 ] [ 49 ] См. Таблицу выше для краткого изложения известных лигандов TLR.
Эндогенные лиганды
[ редактировать ]Стереотипный воспалительный ответ, спровоцированный активацией рецепторов Toll-подобного, вызвал предположение, что эндогенные активаторы рецепторов, подобных Toll, могут участвовать в аутоиммунных заболеваниях. TLR были подозреваны в связывании с молекулами хозяина, включая фибриноген (вовлеченные в свертывание крови ), белки теплового шока (HSP), HMGB1 , компоненты внеклеточного матрикса и самостоятельный Комплекс с эндогенными белками становятся устойчивыми к этим нуклеазам и получают доступ к эндосомальным TLR как TLR7 или TLR9). Эти эндогенные лиганды обычно продуцируются в результате нефизиологической гибели клеток. [ 50 ]
Сигнализация
[ редактировать ]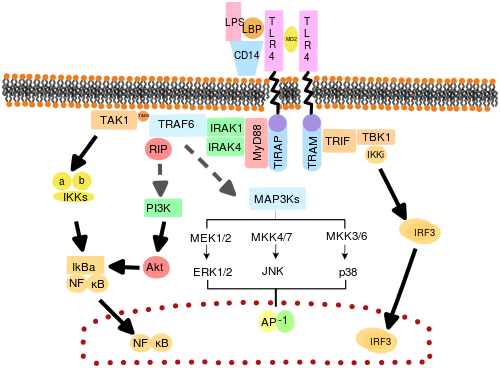
Считается, что TLR функционируют как димеры . Хотя большинство TLR, по -видимому, функционируют как гомодимеры , TLR2 образует гетеродимеры с TLR1 или TLR6, каждый димер имеет различную специфичность лиганда. TLR также могут зависеть от других со-рецепторов для полной чувствительности к лиганду, например, в случае распознавания TLR4 LPS , который требует MD-2. Известно, что CD14 и LPS-связывающий белок ( LBP ) облегчают представление LPS для MD-2.
Набор эндосомных TLR, содержащих TLR3, TLR7, TLR8 и TLR9, распознают нуклеиновую кислоту, полученную из вирусов, а также эндогенные нуклеиновые кислоты в контексте патогенных событий. Активация этих рецепторов приводит к выработке воспалительных цитокинов , а также интерферонам I типа ( интерферон тип I ), чтобы помочь бороться с вирусной инфекцией.
Адаптерные белки и киназы, которые опосредуют передачу сигналов TLR, также были нацелены. Кроме того, случайный мутагенез зародышевой линии с ENU был использован для расшифровки сигнальных путей TLR. При активации TLRS рекрутирует молекулы адаптера в цитоплазме клеток, чтобы распространять сигнал. Известно, что четыре молекулы адаптера участвуют в передаче сигналов. Эти белки известны как MyD88 , TIRAP (также называемый MAL), TRIF и TRAM (молекула адаптера, связанную с TRIF). [ 51 ] [ 52 ] [ 53 ]
Передача сигналов TLR делится на два различных сигнальных путей: MyD88-зависимый и TRIF-зависимый путь.
MyD88-зависимый путь
[ редактировать ]Зависимый от MyD88 отклик возникает при димеризации TLR и используется каждым TLR, кроме TLR3. Его первичным эффектом является активация NFκB и митоген-активируемой протеинкиназы . Связывание лиганда и конформационные изменения, которые возникают в рецепторе, рекрутирует адаптерный белок MyD88, член семейства TIR . MyD88 затем набирает Irak4 , Irak1 и Irak2 . Затем киназы irak затем фосфорилируют и активируют белок TRAF6 , который, в свою очередь, полиубиквинает белок TAK1, а также сам по себе, чтобы облегчить связывание с IKK-β . При связывании TAK1 фосфорилирует IKK-β, который затем фосфорилирует IκB, вызывая его деградацию, и позволяет NFκB диффундировать в ядро клеток и активировать транскрипцию и, как следствие, индукция воспалительных цитокинов. [ 50 ]
Триф-зависимый путь
[ редактировать ]Как TLR3, так и TLR4 используют TRIF-зависимый путь, который запускается дцРНК и LPS соответственно. Для TLR3 дцРНК приводит к активации рецептора, рекрутируя адаптер TRIF . TRIF активирует киназы TBK1 и RIPK1 , которые создают ветвь в сигнальном пути. Сигнальный комплекс TRIF/TBK1 фосфорилирует IRF3, позволяющий его транслокацию в ядро и производство интерферона типа I. Между тем, активация RIPK1 вызывает полиубиквитинирование и активацию транскрипции TAK1 и NFκB таким же образом, как и MyD88-зависимый путь. [ 50 ]
Передача сигналов TLR в конечном итоге приводит к индукции или подавлению генов, которые организуют воспалительный ответ. В целом, тысячи генов активируются передачей сигналов TLR, и в совокупности TLR представляют собой один из самых плейотропных , но строгого регулируемых шлюзов для модуляции генов.
TLR4 - единственный TLR, который использует все четыре адаптера. Комплекс, состоящий из TLR4, MD2 и LPS, рекрутирует TIR-домен, содержащие адаптеры TIRAP и MYD88 и, таким образом, инициирует активацию NFκB (ранняя фаза) и MAPK. Комплекс TLR4-MD2-LPS затем подвергается эндоцитозу, а в эндосоме он образует сигнальный комплекс с адаптерами трамвая и TRIF. Этот TRIF-зависимый путь снова приводит к активации IRF3 и производству интерферонов типа I, но он также активирует активацию NFκB поздней фазы. Как поздняя, так и ранняя фаза активация NFκB необходима для выработки воспалительных цитокинов. [ 50 ]
Медицинская значимость
[ редактировать ]Имикимод (кардинально используется в дерматологии ) является агонистом TLR7, а его преемник -восстановитель - агонист TLR7 и TLR8. [ 54 ] Недавно Resiquimod был изучен в качестве агента иммунотерапии рака, [ 55 ] действуя через стимуляцию макрофагов, связанных с опухолью.
Несколько лигандов TLR находятся в клинической разработке или тестируются на моделях на животных в качестве адъювантов вакцины , [ 56 ] С первым клиническим применением у людей в рекомбинантной вакцине зостера герпеса в 2017 году, которая содержит монофосфорильный липид компонент.
Сообщалось о уровнях экспрессии РНК мессенджера TLR7 у молочных животных при естественной вспышке заболеваний с ногами. [ 57 ]
Было показано, что TLR4 важен для долгосрочных побочных эффектов опиоидов . Считается, что его активация приводит к снижению воспалительных модуляторов, включая TNF-α и IL-1β , и, как полагают, постоянное высвобождение этих модуляторов низкого уровня снижает эффективность лечения опиоидных лекарств со временем и участвует в толерантности к опиоидам, устойчивости к опиоидным [ 58 ] [ 59 ] гипералгезия и аллодиния . [ 60 ] [ 61 ] Активация TLR4, вызванная морфином, ослабляет обезболивания подавление опиоидами и усиливает развитие опиоидной толерантности и зависимости , злоупотребления наркотиками и других негативных побочных эффектов, таких как депрессия дыхания и гипералгезия. [ 62 ] Было показано, что лекарства, которые блокируют действие TNF-α или IL-1β, увеличивают анальгетические эффекты опиоидов и снижают развитие толерантности и других побочных эффектов, [ 63 ] [ 64 ] И это также было продемонстрировано с помощью лекарств, которые блокируют сам TLR4.
«Неестественные» энантиомеры опиоидных препаратов, таких как (+)-морфин и (+)-налоксон, не имеет сродства к опиоидным рецепторам, все еще вызывают ту же активность в TLR4, что и их «нормальные» энантиомеры. [ 65 ] [ 66 ] Итак, «неестественные» энтианомеры опиоидов, таких как (+)-налоксон, могут использоваться для блокировки активности TLR4 опиоидных анальгетических препаратов без какого-либо сродства к μ-опиоидному рецептору [ 67 ] [ 66 ] [ 68 ]
Открытие
[ редактировать ]Когда микробы сначала были признаны в качестве причины инфекционных заболеваний, было сразу ясно, что многоклеточные организмы должны быть способны распознавать их при заражении и, следовательно, способны распознавать молекулы, уникальные для микробов. Большая литература, охватывающая большую часть прошлого века, свидетельствует о поиске ключевых молекул и их рецепторов. Более 100 лет назад Ричард Пфайффер , ученик Роберта Коха , придумал термин « эндотоксин », чтобы описать вещество, создаваемое грамотрицательными бактериями , который может вызвать лихорадку и шок у экспериментальных животных . В последующие десятилетия эндотоксин был химически охарактеризован и идентифицирован как липополисахарид (LPS), продуцируемый большинством грамотрицательных бактерий. Этот липополисахарид является неотъемлемой частью грамотрицательной мембраны и высвобождается при разрушении бактерии. Другие молекулы (бактериальные липопептиды , флагеллин и неметилированная ДНК ), в свою очередь, показали, как обычно защищают ответы хозяина. Тем не менее, эти ответы могут быть вредными, если они чрезмерно продолжительны или интенсивны. Следует логически, что должны быть рецепторы для таких молекул, способные предупредить хозяина о наличии инфекции, но они оставались неуловимыми в течение многих лет. Toll-подобные рецепторы теперь учитываются среди ключевых молекул, которые предупреждают иммунная система к присутствию микробных инфекций.
Прототипический член семьи, рецептор Toll ( P08953 ; TL) в фруктовой мух Drosophila Melanogaster , был обнаружен в 1985 году нобелевскими лауреатами Кристианом Нюслейн-Вольхард и Эриком Висчаусом и коллегами. Он был известен своей функцией развития в эмбриогенезе путем установления дорсальной вентральной . оси Он был назван в честь восклицания Кристиана Нюслейна-Вольхарда 1985 года « Das Ist Ja Toll ! » («Это удивительно!») В связи с недоразвитой вентральной частью личинки фруктовой мухи. [ 4 ] Он был клонирован лабораторией Кэтрин Андерсон в 1988 году. [ 69 ] В 1996 году что в результате и его коллеги обнаружили, Жюль А. Хоффманн выяснилось важную роль в иммунитете мухи в грибковой инфекции , которую он достиг путем активации синтеза антимикробных пептидов. [ 19 ]
Первый зарегистрированный рецептор, похожий на человека, был описан Nomura и коллегами в 1994 году, [ 70 ] Намечен на хромосому Тагучи и коллегами в 1996 году. [ 71 ] Поскольку иммунная функция платы за Drosophila не была тогда известна, предполагалось, что TIL (теперь известный как TLR1) может участвовать в развитии млекопитающих. Однако в 1991 году (до обнаружения TIL) было отмечено, что молекула с четкой ролью в иммунной функции у млекопитающих, рецептора интерлейкина-1 (IL-1), также имела гомологию с Drosophila Toll; Цитоплазматические части обеих молекул были одинаковыми. [ 72 ]
В 1997 году Чарльз Дженуэй и Руслан Меджитов показали, что рецептор, похожий на Toll, теперь известный как TLR4, когда при искусственном лигировании с использованием антител индуцирует активацию определенных генов, необходимых для инициирования адаптивного иммунного ответа . [ 7 ] TLR 4 функционирует как рецептор LPS, который был обнаружен Брюсом А. Битлером и его коллегами. [ 73 ] Эти работники использовали позиционное клонирование , чтобы доказать, что мыши, которые не могли реагировать на LPS, имели мутации, которые отменили функцию TLR4. Это идентифицировало TLR4 как один из ключевых компонентов рецептора для LPS.

В свою очередь, другие гены TLR были удалены у мышей с помощью генов, в основном в лаборатории Шизуо Акиры и его коллег. В настоящее время считается, что каждый TLR обнаруживает отдельную коллекцию молекул - некоторые из микробного происхождения, а также некоторые продукты повреждения клеток - и сигнализировать о наличии инфекций. [ 74 ]
Гомологи завода были обнаружены Памелой Рональдом в 1995 году (Rice XA21) [ 75 ] и Томас Боллер в 2000 году ( Arabidopsis FLS2). [ 76 ]
В 2011 году Бутлер и Хоффманн были удостоены Нобелевской премии по медицине или физиологии за их работу. [ 77 ] Хоффманн и Акира получили Международную награду Канады Гарднер в 2011 году. [ 78 ]
Примечания и ссылки
[ редактировать ]- ^ Махла Р.С., Редди М.К., Прасад Д.В., Кумар Х (сентябрь 2013 г.). «Подслащивают Pamps: роль сахарных комплексных PAMPS в врожденном иммунитете и биологии вакцины» . Границы в иммунологии . 4 : 248. DOI : 10.3389/fimmu.2013.00248 . PMC 3759294 . PMID 24032031 .
- ^ Передняя вера; Indriputri, разрезать; Мамутс, Джанет; Nugraha, Jusak (2020). «TLR10 и его уникальные противовоспалительные свойства и потенциальное использование в качестве цели в терапии» . Иммунная сеть . 20 (3): E21. doi : 10.4110/in.2020.20.e21 . ISSN 1598-2629 . PMC 7327153 . PMID 32655969 .
- ^ Kemball CC, Alirezaei M, Whitton JL (2010). «Тип B Coxsackieviruses и их взаимодействие с врожденной и адаптивной иммунной системой» . Будущая микробиология . 5 (9): 1329–1347. doi : 10.2217/fmb.10.101 . PMC 3045535 . PMID 20860480 .
- ^ Jump up to: а беременный Hansson GK, Edfeldt K (июнь 2005 г.). «Проезд, чтобы быть заплаченным у ворот к стене суда» . Артериосклероз, тромбоз и сосудистая биология . 25 (6): 1085–7. doi : 10.1161/01.atv.0000168894.43759.47 . PMID 15923538 .
- ^ Delneste Y, Beauvillain C, Jeannin P (январь 2007 г.). «[Врожденный иммунитет: структура и функция TLRS]» . МЕДЕЦИНА/НАУКА . 23 (1): 67–73. doi : 10.1051/medsci/200723167 . PMID 17212934 .
- ^ Такеда К, Акира С. (январь 2005 г.). «Рецепторы, похожие на платы, в врожденном иммунитете». Международная иммунология . 17 (1): 1–14. doi : 10.1093/intimm/dxh186 . PMID 15585605 .
- ^ Jump up to: а беременный в Medzhitov R, Preston-Hurlburt P, Janeway CA (июль 1997 г.). «Гомолог человека Drosophila Toll Signals активации адаптивного иммунитета» . Природа . 388 (6640): 394–7. Bibcode : 1997natur.388..394M . doi : 10.1038/41131 . PMID 9237759 .
- ^ Jump up to: а беременный Шарма Н., Ахаде А.С., Кадри А (апрель 2013 г.). «Сфингозин-1-фосфат подавляет TLR-индуцированную секрецию CXCL8 из Т-клеток человека». Журнал биологии лейкоцитов . 93 (4): 521–8. doi : 10.1189/jlb.0712328 . PMID 23345392 .
- ^ Someer AS, Nissar S. Toll-подобные рецепторы (TLR): структура, функции, передача сигналов и роль их полиморфизмов в чувствительности к колоректальному раку. Biomed Res Int. 2021 сентябрь 12; 2021: 1157023. doi: 10.1155/2021/1157023. PMID 34552981; PMCID: PMC8452412.
- ^ Rolls A, Shechter R, London A, Ziv Y, Ronen A, Levy R, Schwartz M (сентябрь 2007 г.). «Рецепторы, похожие на Toll, модулируют нейрогенез гиппокампа взрослых». Природная клеточная биология . 9 (9): 1081–8. doi : 10.1038/ncb1629 . PMID 17704767 . S2CID 12517461 .
- ^ Du X, Poltorak A, Wei Y, Beutler B (сентябрь 2000 г.). «Три новых рецептора, похожие на млекопитающих: структура гена, экспрессия и эволюция» . Европейская сеть цитокинов . 11 (3): 362–71. PMID 11022119 .
- ^ Chuang Th, Ulevitch RJ (сентябрь 2000 г.). «Клонирование и характеристика подсемейства человеческих рецепторов: HTLR7, HTLR8 и HTLR9» . Европейская сеть цитокинов . 11 (3): 372–8. PMID 11022120 .
- ^ Табета К., Джорджел П., Янссен Е., Дю Х, Хобе К., Крозат К. и др. (Март 2004 г.). «Рецепторы, похожие на Toll, 9 и 3 как важные компоненты врожденной иммунной защиты от цитомегаловирусной инфекции мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (10): 3516–21. Bibcode : 2004pnas..101.3516t . doi : 10.1073/pnas.0400525101 . PMC 373494 . PMID 14993594 .
- ^ Jump up to: а беременный Roach JC, Glusman G, Rowen L, Kaur A, Purcell MK, Smith KD и др. (Июль 2005 г.). «Эволюция рецепторов, подобных позвоночным» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (27): 9577–82. Bibcode : 2005pnas..102.9577r . doi : 10.1073/pnas.0502272102 . PMC 1172252 . PMID 15976025 .
- ^ Lemaitre B, Hoffmann J (2007). «Защита хозяина Drosophila melanogaster» . Ежегодный обзор иммунологии . 25 : 697–743. doi : 10.1146/annurev.immunol.25.022106.141615 . PMID 17201680 .
- ^ Valanne S, Wang JH, Rämet M (январь 2011 г.). «Сигнальный путь Drosophila Toll» . Журнал иммунологии . 186 (2): 649–56. doi : 10.4049/jimmunol.1002302 . PMID 21209287 .
- ^ Jump up to: а беременный Dudzic JP, Hanson MA, Iatsenko I, Kondo S, Lemaitre B (апрель 2019 г.). «Больше, чем черный или белый: меланизация и регуляторные сериновые протеазы в Drosophila» . Сотовые отчеты . 27 (4): 1050–1061.e3. doi : 10.1016/j.celrep.2019.03.101 . PMID 31018123 .
- ^ Хансон М.А., Гамильтон П.Т., Перлман С.Дж. (октябрь 2016 г.). «Иммунные гены и дивергентные антимикробные пептиды в мухах подрода дрозофила» . BMC Эволюционная биология . 16 (1): 228. doi : 10.1186/s12862-016-0805-y . PMC 5078906 . PMID 27776480 .
- ^ Jump up to: а беременный Лемитр Б., Николас Э., Михаут Л., Рейххарт Дж. М., Хоффманн Дж.А. (сентябрь 1996 г.). «Дорсовентральная регуляторная гена кассета Spätzle/Toll/тактус контролирует мощный противогрибковой ответ у взрослых дрозофилы» . Клетка . 86 (6): 973–83. doi : 10.1016/s0092-8674 (00) 80172-5 . PMID 8808632 . S2CID 10736743 .
- ^ Issa N, Guillaumot N, Lauret E, Matt N, Schaeffer-Reiss C, Van Dorsselaer A, et al. (Февраль 2018 г.). «Циркулирующая протеаза персефона является иммунным датчиком для микробной протеолитической активности выше по течению от пути Drosophila» . Молекулярная клетка . 69 (4): 539–550.e6. doi : 10.1016/j.molcel.2018.01.029 . PMC 5823974 . PMID 29452635 .
- ^ Ferrandon D, Imler JL, Hetru C, Hoffmann JA (ноябрь 2007 г.). «Системный иммунный ответ Drosophila: восприятие и передача сигналов во время бактериальных и грибковых инфекций». Природные обзоры. Иммунология . 7 (11): 862–74. doi : 10.1038/nri2194 . PMID 17948019 . S2CID 11104900 .
- ^ Chowdhury M, Li CF, He Z, Lu Y, Liu XS, Wang YF, et al. (Июнь 2019). «Дрозофила» . Журнал биологической химии . 294 (26): 10172–10181. doi : 10.1074/jbc.ra118.006804 . PMC 6664172 . PMID 31088910 .
- ^ Ли Дж., Сайед Н., Хантер А., Ау К.Ф., Вонг В.Х., Мокарски Е.С. и др. (Октябрь 2012 г.). «Активация врожденного иммунитета необходима для эффективного ядерного перепрограммирования» . Клетка . 151 (3): 547–58. doi : 10.1016/j.cell.2012.09.034 . PMC 3506423 . PMID 23101625 .
- ^ Jump up to: а беременный Хатай, Хироцугу; Лепелли, Алиса; Зенг, Вангёнг; Хейден, Мэтью С.; Гош, Санкар (2016). «Toll-подобный рецептор 11 (TLR11) взаимодействует с флагеллином и профилин с помощью разрозненных механизмов» . Plos один . 11 (2): E0148987. BIBCODE : 2016PLOSO..1148987H . doi : 10.1371/journal.pone.0148987 . ISSN 1932-6203 . PMC 4747465 . PMID 26859749 .
- ^ Mathur R, Oh H, Zhang D, Park SG, Seo J, Koblansky A, et al. (Октябрь 2012 г.). «Мышиная модель инфекции Salmonella Typhi» . Клетка . 151 (3): 590–602. doi : 10.1016/j.cell.2012.08.042 . PMC 3500584 . PMID 23101627 .
- ^ Флайник, Мартин Ф.; Сингх, Невил Дж.; Голландия, Стивен М., ред. (2023). «Глава 12 Рецепторы распознавания образцов и семейство IL-1». Фундаментальная иммунология Павла (8 -е изд.). Филадельфия Балтимор Нью -Йорк Лондон Буэнос -Айрес Гонконг Сидней Токио: Wolters Kluwer/Lippincott Williams & Wilkins. ISBN 978-1-9751-4251-3 Полем
В целом, TLR 1, 2, 4-6 и 10 выражены на плазматической мембране, а TLRS 3, 7–9 и от 11 до 13 ограничены эндосомами.
- ^ Jump up to: а беременный в Если еще не указано в коробках, то ссылка: Waltenbaugh C, Doan T, Melvold R, Viselli S (2008). Иммунология . Иллюстрированные отзывы Липпинкотта. Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins. п. 17. ISBN 978-0-7817-9543-2 .
- ^ Jump up to: а беременный Сабро I, Дауэр С.К., Уайт М.К. (ноябрь 2005 г.). «Роль рецепторов, подобных Toll, в регуляции миграции, активации и апоптоза нейтрофилов» . Клинические инфекционные заболевания . 41 (Suppl 7): S421-6. doi : 10.1086/431992 . PMID 16237641 .
- ^ Jump up to: а беременный в дюймовый Sallusto F, Lanzavecchia A (2002). «Поучительная роль дендритных клеток в Т-клетках» . Исследование артрита . 4 (Suppl 3): S127-32. doi : 10.1186/ar567 . PMC 3240143 . PMID 12110131 .
- ^ Gerondakis S, Grumont RJ, Banerjee A (2007). «Регулирование активации B-клеток и выживаемости в ответ на сигналы TLR» . Иммунология и клеточная биология . 85 (6): 471–5. doi : 10.1038/sj.icb.7100097 . PMID 17637697 . S2CID 30443009 .
- ^ Cario E, Rosenberg IM, Brandwein SL, Beck PL, Reinecker HC, Podolsky DK (январь 2000 г.). «Липополисахарид активирует различные сигнальные пути в эпителиальных клеточных линиях кишечника, экспрессирующих рецепторы, подобные плате» » . Журнал иммунологии . 164 (2): 966–72. doi : 10.4049/jimmunol.164.2.966 . PMID 10623846 .
- ^ Peana M, Zdyb K, Medici S, Pelucelli A, Simula G, Gumienna-Kontecka E, Zoroddu MA (декабрь 2017 г.). «Ni (II) взаимодействие с пептидной моделью человеческого эктодомена TLR4». Журнал следовых элементов в медицине и биологии . 44 : 151–160. doi : 10.1016/j.jtemb.2017.07.006 . PMID 28965571 .
- ^ Салазар Гонсалес Р.М., Шехата Х., О'Коннелл М.Дж., Ян Ю., Морено-Фернандес М.Е., Чугнет К.А., Алиберти Дж. (2014). «Toxoplasma Gondii-полученное профилин запускает 5-зависимую 5-зависимую выработку цитокинов человека» . Журнал врожденного иммунитета . 6 (5): 685–94. doi : 10.1159/000362367 . PMC 4141014 . PMID 24861338 .
- ^ SEIZER L, Rahimi S, Santos-Sierra S, Drexel M (2022) Экспрессия платы, подобно рецептору 8 (TLR8) в определенных группах мышиных интернейрон гиппокампа. PLO One 17 (5): E0267860. https://doi.org/10.1371/journal.pone.0267860
- ^ Guan Y, Ranoa DR, Jiang S, Mutha SK, Li X, Baudry J, Tapping RI (май 2010 г.). «Человеческие TLRS 10 и 1 имеют общие механизмы врожденного иммунного зондирования, но не передачи сигналов» . Журнал иммунологии . 184 (9): 5094–103. doi : 10.4049/jimmunol.0901888 . PMID 20348427 .
- ^ Chuang T, Ulevitch RJ (март 2001 г.). «Идентификация HTLR10: новый рецептор, похожий на человека, предпочтительно экспрессируется в иммунных клетках». Biochimica et Biophysica Acta (BBA) - структура и экспрессия гена . 1518 (1–2): 157–61. doi : 10.1016/s0167-4781 (00) 00289-x . PMID 11267672 .
- ^ Hornung V, Rothenfusser S, Britsch S, Krug A, Jahrsdörfer B, Giese T, Endres S, Hartmann G (май 2002). «Количественная экспрессия мРНК 1-10 рецепторов рецептора 1-10 в клеточных подмножествах мононуклеарных клеток периферической крови человека и чувствительность к олигодезоксинуклеотидам CPG» . Журнал иммунологии . 168 (9). Балтимор, штат Мэриленд: 1950: 4531–7. doi : 10.4049/jimmunol.168.9.4531 . PMID 11970999 .
{{cite journal}}: CS1 Maint: местоположение ( ссылка ) - ^ Jump up to: а беременный Риган Т., Нолли К, Кармоди Р., Хьюстон А., Шанахан Ф., Макшарри Дж., Брит Е (декабрь 2013 г.). «Идентификация TLR10 в качестве ключевого медиатора воспалительного ответа на Listeria Monocytogenes в эпителиальных клетках кишечника и макрофагах» . Журнал иммунологии . 191 (12): 6084–92. doi : 10.4049/jimmunol.1203245 . PMID 24198280 .
- ^ Yarovinsky F, Zhang D, Andersen JF, Bannenberg GL, Serhan CN, Hayden MS, et al. (Июнь 2005 г.). «TLR11 активация дендритных клеток протозойским профилиноподобным белком» . Наука . 308 (5728): 1626–9. Bibcode : 2005sci ... 308.1626y . doi : 10.1126/science.1109893 . PMID 15860593 . S2CID 34165967 .
- ^ Pifer R, Benson A, Sturge CR, Yarovinsky F (февраль 2011 г.). «UNC93B1 необходим для активации TLR11 и IL-12-зависимой устойчивости хозяина к токсоплазме Gondii» . Журнал биологической химии . 286 (5): 3307–14. doi : 10.1074/jbc.m110.171025 . PMC 3030336 . PMID 21097503 .
- ^ Koblansky AA, Jankovic D, Oh H, Hieny S, Sungnak W, Mathur R, et al. (Январь 2013). «Распознавание профилина с помощью Toll-подобного рецептора 12 имеет решающее значение для устойчивости хозяина Gondii Toxoplasma» . Иммунитет . 38 (1): 119–30. doi : 10.1016/j.immuni.2012.09.016 . PMC 3601573 . PMID 23246311 .
- ^ Мишра Б.Б., Гундра Ум, Тил Дж. М. (декабрь 2008 г.). «Экспрессия и распределение платежных рецепторов 11-13 в мозге во время мышиного нейроцистицероза» . Журнал нейровоспаления . 5 : 53. DOI : 10.1186/1742-2094-5-53 . PMC 2631477 . PMID 19077284 .
- ^ Shi Z, Cai Z, Sanchez A, Zhang T, Wen S, Wang J, et al. (Февраль 2011 г.). «Новый платный рецептор, который распознает вирус везикулярного стоматита» . Журнал биологической химии . 286 (6): 4517–24. doi : 10.1074/jbc.m110.159590 . PMC 3039399 . PMID 21131352 .
- ^ Олденбург М., Крюгер А., Ферстл Р., Кауфманн А., Ниес Г., Сигмунд А. и др. (Август 2012 г.). «TLR13 распознает бактериальную 23S рРНК, лишенную модификации резистентности к эритромицину». Наука . 337 (6098): 1111–5. Bibcode : 2012Sci ... 337.11111o . doi : 10.1126/science.1220363 . PMID 22821982 . S2CID 206540638 .
- ^ Hochrein H, Kirschning CJ (март 2013 г.). «Бактерии уклоняются от иммунного распознавания через TLR13 и связывание их 23S рРНК с помощью антибиотиков MLS с помощью тех же механизмов» . Онкоиммунология . 2 (3): E23141. doi : 10.4161/onci.23141 . PMC 3661153 . PMID 23802068 .
- ^ Шреббет, Герти; Бенитес-Рибас, Даниэль; Хуже того, данные; Lambetck, annelchin you; Хаут-Куджир, Маверкс; Шфт, Нильс; Point, Cornelis JA; Figdor, Carl G.; Ademe, Gest J.; Freeze, I. Jolanda M. (29 июля 2010 г.). «Обычно сваренные профилактические вакцины в качестве альтернативы для синтетических продуцируемых к клеткам с моноцитами-дендерными клетками матоцитов» . Кровь . 116 (4): 564–574. doi : 10 1182/кровь-2009-11-251884 . HDL : 2066/89493 . ISSN 1528-0020 . PMID 20424184 .
- ^ Элейник, Марк; Свенссон-Арвелунд, Джудит; Pantsulaia, Gvantsa; Ким, Кристи; Роуз, Самуил А.; Упадхьяй, Ранджан; Йеллин, Майкл; Марш, Генри; Орепер, Даниэль; Jhunjhunwala, Сучит; Мусс, Кристина Карин; Мерад, Мириам; Браун, Брайан Д.; Броди, Джошуа Д. (июль 2023 г.). «Агонисты рецепторов распознавания паттернов в вакцинах против патогенов опосредуют противоопухолевое перекрестное примирование Т-клеток» . Журнал для иммунотерапии рака . 11 (7): E007198. doi : 10.1136/jitc-2023-007198 . ISSN 2051-1426 . PMC 10373699 . PMID 37487664 .
- ^ Hoebe K, Du X, Georgel P, Janssen E, Tabeta K, Kim SO, et al. (Август 2003 г.). «Идентификация LPS2 в качестве ключевого преобразователя MyD88-независимой передачи сигналов TIR». Природа . 424 (6950): 743–8. Bibcode : 2003natur.424..743H . doi : 10.1038/nature01889 . PMID 12872135 . S2CID 15608748 .
- ^ Хемми Х., Такеучи О., Каваи Т., Кайшо Т., Сато С., Санджо Х. и др. (Декабрь 2000 г.). «Рецептор, похожий на Toll, распознает бактериальную ДНК» Природа 408 (6813): 740–5 Bibcode : 2000natur.408..740h . Doi : 10.1038/ 350471 PMID 1130078 S2CID 4405163
- ^ Jump up to: а беременный в дюймовый Kawai T, Akira S (май 2010 г.). «Роль рецепторов распознавания паттернов в врожденном иммунитете: обновление о платных рецепторах» . Природа иммунология . 11 (5): 373–84. doi : 10.1038/ni.1863 . PMID 20404851 . S2CID 39414949 .
- ^ Shigeoka AA, Holscher TD, King AJ, Hall FW, Kiosses WB, Tobias PS, et al. (Май 2007). «TLR2 конститутивно экспрессируется в почках и участвует в ишемическом повреждении почек как через MyD88, так и по -зависимые пути» . Журнал иммунологии . 178 (10): 6252–8. doi : 10.4049/jimmunol.178.10.6252 . PMID 17475853 .
- ^ Yamamoto M, Sato S, Hemmi H, Uematsu S, Hoshino K, Kaisho T, et al. (Ноябрь 2003). «Трамвай специально участвует в 4-опосредованном рецепторе 4-опосредованного рецептора MyD88». Природа иммунология . 4 (11): 1144–50. doi : 10.1038/ni986 . PMID 14556004 . S2CID 13016860 .
- ^ Yamamoto M, Sato S, Hemmi H, Sanjo H, Uematsu S, Kaisho T, et al. (Ноябрь 2002). «Основная роль TIRAP в активации сигнального каскада, разделяемого TLR2 и TLR4». Природа . 420 (6913): 324–9. Bibcode : 2002natur.420..324y . doi : 10.1038/nature01182 . PMID 12447441 . S2CID 16163262 .
- ^ Питер Фрич (2004). Дерматология Венерология: Основы. Клиника. Атлас (на немецком языке). Берлин: Спрингер. ISBN 3-540-00332-0 .
- ^ Rodell CB, Arlauckas SP, Cuccarese MF, Garris CS, Li R, Ahmed MS, et al. (Август 2018). «Наночастицы, нагруженные TLR7/8-агонистом, способствуют поляризации макрофагов, ассоциированных с опухолом, для усиления иммунотерапии раком» . Природная биомедицинская инженерия . 2 (8): 578–588. doi : 10.1038/s41551-018-0236-8 . PMC 6192054 . PMID 31015631 .
- ^ Toussi DN, Massari P (апрель 2014 г.). «Иммунный адъювантный эффект молекулярно определенных платтных рецепторных лигандов» . Вакцина . 2 (2): 323–53. doi : 10.3390/vaccines2020323 . PMC 4494261 . PMID 26344622 .
- ^ Audarya, SD; Pattnaik, B.; Саньял, А.; Мохапатра, JK (2017). «Подобные уровни экспрессии экспрессии рибонуклеиновой кислоты рецептора 7 у молочных животных при вспышке заболеваний с ногами» (PDF) . Бюллетень Буффало . 36 (3). Архивировано из оригинала (PDF) 28 апреля 2021 года.
- ^ Shavit Y, Wolf G, Goshen I, Livshits D, Yirmiya R (май 2005). «Интерлейкин-1 противодействует анальгезии морфина и лежит в основе толерантности к морфину». Боль . 115 (1–2): 50–9. doi : 10.1016/j.pain.2005.02.003 . PMID 15836969 . S2CID 7286123 .
- ^ Мохан С., Дэвис Р.Л., Десилва У, Стивенс К.В. (октябрь 2010 г.). «Двойная регуляция рецепторов опиоидов MU в клетках нейробластомы SK-N-SH с помощью морфина и интерлейкина-1β: доказательства опиоид-иммунных перекрестных помех» . Журнал нейроиммунологии . 227 (1–2): 26–34. doi : 10.1016/j.jneuroim.2010.06.007 . PMC 2942958 . PMID 20615556 .
- ^ Komatsu T, Sakurada S, Katsuyama S, Sanai K, Sakurada T (2009). Механизм аллодинии, вызванный интратекальным морфином-3-глюкуронидом у мышей . Международный обзор нейробиологии. Тол. 85. С. 207–19. doi : 10.1016/s0074-7742 (09) 85016-2 . ISBN 9780123748935 Полем PMID 19607972 .
- ^ Льюис С.С., Хатчинсон М.Р., Резвани Н., Лорам Л.К., Чжан Ю., Майер С.Ф., Райс К.С., Уоткинс Л.Р. (январь 2010 г.). «Свидетельство о том, что интратекальный морфин-3-глюкуронид может вызвать усиление боли с помощью Toll-подобного рецептора 4/MD-2 и интерлейкина-1бета» . Нейробиология . 165 (2): 569–83. doi : 10.1016/j.neuroscience.2009.10.011 . PMC 2795035 . PMID 19833175 .
- ^ Драль С (22 августа 2012 г.). «Маленькие молекулы нацелены на рецепторы» . Химические и инженерные новости .
- ^ Shen CH, Tsai Ry, Shih MS, Lin SL, Tai YH, Chien CC, Wong CS (февраль 2011 г.). «Etanercept восстанавливает антиноцицептивное действие морфина и подавляет нейровоспаление позвоночника у крыс, устойчивых к морфину» . Анестезия и анальгезия . 112 (2): 454–9. doi : 10.1213/ane.0b013e3182025b15 . PMID 21081778 . S2CID 12295407 .
- ^ Хук М.А., Уошберн С.Н., Морено Г., Воллер С.А., Пуга Д., Ли К.Х., Грау Дж.В. (февраль 2011 г.). «Антагонист рецептора IL-1 блокирует индуцированное морфином ослабление локомоторного восстановления после повреждения спинного мозга» . Мозг, поведение и иммунитет . 25 (2): 349–59. doi : 10.1016/j.bbi.2010.10.018 . PMC 3025088 . PMID 20974246 .
- ^ Уоткинс Л.Р., Хатчинсон М.Р., Райс К.С., Майер С.Ф. (ноябрь 2009 г.). «« Плата »индуцированной опиоидами глиальной активации: улучшение клинической эффективности опиоидов путем нацеливания на глию» . Тенденции в фармакологических науках . 30 (11): 581–91. doi : 10.1016/j.tips.2009.08.002 . PMC 2783351 . PMID 19762094 .
- ^ Jump up to: а беременный Hutchinson MR, Zhang Y, Brown K, Coats BD, Shridhar M, Sholar PW, Patel SJ, Crysdale NY, Harrison JA, Maier SF, Rice KC, Watkins LR (июль 2008 г.). «Нестеоселективное изменение невропатической боли с помощью налоксона и налтрексона: вовлечение платежного рецептора 4 (TLR4)» . Европейский журнал нейробиологии . 28 (1): 20–9. doi : 10.1111/j.1460-9568.2008.06321.x . PMC 2588470 . PMID 18662331 .
- ^ Hutchinson MR, Coats BD, Lewis SS, Zhang Y, Sprunger DB, Rezvani N, Baker EM, Jekich BM, Wieseler JL, Somogyi AA, Martin D, Poole S, Judd CM, Maier SF, Watkins LR (ноябрь 2008). «Провоспалительные цитокины противодействуют индуцированной опиоидам острого и хронической анальгезии» . Мозг, поведение и иммунитет . 22 (8): 1178–89. doi : 10.1016/j.bbi.2008.05.004 . PMC 2783238 . PMID 18599265 .
- ^ Hutchinson MR, Lewis SS, Coats BD, Rezvani N, Zhang Y, Wieseler JL, Somogyi AA, Yin H, Maier SF, Rice KC, Watkins LR (май 2010 г.). «Возможное вовлечение Toll-подобного рецептора 4/миелоидного фактора дифференцировки-2 опиоидных неактивных изомеров вызывает провоспаление позвоночника и связанные с ними поведенческие последствия» . Нейробиология . 167 (3): 880–93. doi : 10.1016/j.neuroscience.2010.02.011 . PMC 2854318 . PMID 20178837 .
- ^ Hashimoto C, Hudson KL, Anderson KV (январь 1988 г.). «Ген drosophila, необходимый для дорсально-вентральной эмбриональной полярности, по-видимому, кодирует трансмембранное белок». Клетка . 52 (2): 269–79. doi : 10.1016/0092-8674 (88) 90516-8 . PMID 2449285 . S2CID 19439405 .
- ^ Nomura N, Miyajima N, Sazuka T, Tanaka A, Kawarabayasi Y, Sato S, et al. (1994). «Прогнозирование кодирующих последовательностей неопознанных генов человека. I. кодирующие последовательности 40 новых генов (KIAA0001-KIAA0040), выведенные путем анализа случайно отобранных клонов кДНК из линии миелоидных клеток человека KG-1» . ДНК -исследования . 1 (1): 27–35. doi : 10.1093/dnares/1.1.27 . PMID 7584026 .
- ^ Тагучи Т., Митчам Дж.Л., Дауэр С.К., Симс Дж., Теста -младший (март 1996 г.). «Хромосомная локализация TIL, гена, кодирующего белок, связанный с Drosophila трансмембранным рецептором, с хромосомой человека 4p14». Геномика . 32 (3): 486–8. doi : 10.1006/geno.1996.0150 . PMID 8838819 .
- ^ Гей Нью -Джерси, Кит Ф.Дж. (май 1991). «Drosophila Toll и рецептор IL-1». Природа . 351 (6325): 355–6. Bibcode : 1991natur.351..355G . doi : 10.1038/3513555B0 . PMID 1851964 . S2CID 1700458 .
- ^ Poltorak A, He X, Smirnova I, Liu My, Van Huffel C, Du X, et al. (Декабрь 1998). «Дефектная передача сигналов LPS у мышей C3H/HEJ и C57BL/10SCCR: мутации в гене TLR4». Наука . 282 (5396): 2085–8. Bibcode : 1998sci ... 282.2085p . doi : 10.1126/science.282.5396.2085 . PMID 9851930 .
- ^ Хошино К., Такеучи О., Каваи Т., Санджо Х, Огава Т., Такеда Ю. и др. (Апрель 1999). «Передовая кромка: мыши с дефицитом Toll-подобного рецептора 4 (TLR4)-гипореактивны липополисахариду: доказательства TLR4 как продукта гена LPS» . Журнал иммунологии . 162 (7): 3749–52. doi : 10.4049/jimmunol.162.7.3749 . PMID 10201887 . S2CID 7419784 .
- ^ Song Wy, Wang GL, Chen LL, Kim HS, Pi Ly, Holsten T, et al. (Декабрь 1995). «Рецепторный киназоподобный белок, кодируемый геном устойчивости к болезням риса, Xa21» . Наука . 270 (5243): 1804–6. Bibcode : 1995sci ... 270.1804s . doi : 10.1126/science.270.5243.1804 . PMID 8525370 . S2CID 10548988 .
- ^ Gómez-Gómez L, Boller T (июнь 2000 г.). «FLS2: рецепторная рецепторная киназа LRR, участвующая в восприятии бактериального элиситора флагеллина у арабидопсиса» . Молекулярная клетка . 5 (6): 1003–11. doi : 10.1016/s1097-2765 (00) 80265-8 . PMID 10911994 .
- ^ «Нобелевская премия по физиологии или медицине 2011» . Нобелевские СМИ AB . 3 октября 2011 года.
- ^ Митчелл Б (23 марта 2011 г.). «Доктор Британской Колумбии выигрывает престижный медицинский приз» . Звезда .
Смотрите также
[ редактировать ]Внешние ссылки
[ редактировать ]- Toll-подобные+рецепторы в Национальной медицинской библиотеке Медицинской библиотеки США (Mesh)
- Toll+белок,+Drosophila в Национальной библиотеке Медицинской библиотеки Медицинской библиотеки (Mesh)
- Tollml: база данных с рецепторами и лигандами в Мюнхене , в Мюнхенском университете
- Семейство врожденных иммунных рецепторов рецепторов, подобных рецепторам (PDF)
- Toll-подобный рецепторный путь
- Биологические анимации