Вирус Эпштейна-Барра
| Гаммагерпесвирус человека 4 | |
|---|---|
 | |
| Электронная микрофотография двух вирионов Эпштейна-Барра (вирусных частиц), показывающая круглые капсиды, свободно окруженные мембранной оболочкой. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Дуплоднавирус |
| Королевство: | Хынггунвирэ |
| Тип: | Пепловирикота |
| Сорт: | Гервивирицеты |
| Заказ: | Герпесвирусные инфекции |
| Семья: | Ортогерпесвирусиды |
| Род: | Лимфокриптовирус |
| Разновидность: | Гаммагерпесвирус человека 4 |
| Синонимы [1] | |
| |
Вирус Эпштейна-Барра ( ВЭБ ), формально называемый гаммагерпесвирусом человека 4 , является одним из девяти известных типов вируса герпеса человека семейства герпеса и одним из наиболее распространенных вирусов у человека. ВЭБ представляет собой вирус с двухцепочечной ДНК . [2] Вирус Эпштейна-Барра (ЭБВ) — первый выявленный онкогенный вирус , вызывающий стойкую инфекцию у человека. ВЭБ вызывает инфекционный мононуклеоз , а также тесно связан со многими злокачественными заболеваниями. Различные составы вакцин прошли испытания на разных животных или на людях. Однако ни один из них не смог предотвратить заражение ВЭБ, и на сегодняшний день ни одна вакцина не одобрена. [3]
Вирус вызывает инфекционный мононуклеоз («моно» или «железистая лихорадка»). Он также связан с различными доброкачественными, предраковыми и злокачественными лимфопролиферативными заболеваниями, связанными с вирусом Эпштейна-Барра, такими как лимфома Беркитта , гемофагоцитарный лимфогистиоцитоз , [4] и лимфома Ходжкина ; нелимфоидные злокачественные новообразования, такие как рак желудка и карцинома носоглотки ; и состояния, связанные с вирусом иммунодефицита человека , такие как волосатая лейкоплакия и центральной нервной системы лимфомы . [5] [6] Вирус также связан с детскими расстройствами, такими как синдром Алисы в стране чудес. [7] и острая мозжечковая атаксия [8] и, по некоторым данным, более высокий риск развития некоторых аутоиммунных заболеваний , [9] особенно дерматомиозит , системная красная волчанка , ревматоидный артрит и синдром Шегрена . [10] [11] Считается, что около 200 000 случаев рака во всем мире в год связаны с ВЭБ. [12] [13] В 2022 году крупное исследование (10 миллионов человек за 20 лет) показало, что ВЭБ является основной причиной рассеянного склероза , при этом недавняя инфекция ВЭБ привела к 32-кратному увеличению риска развития рассеянного склероза. [14] [15] [16] [17] [18]
Заражение ВЭБ происходит при пероральном переносе слюны. [19] и половых выделений.Большинство людей заражаются ВЭБ и приобретают адаптивный иммунитет . В США около половины всех пятилетних детей и около 90% взрослых имеют признаки предшествующего заражения. [20] Младенцы становятся восприимчивыми к ВЭБ, как только исчезает защита материнских антител . У многих детей, инфицированных ВЭБ, симптомы не проявляются или симптомы неотличимы от других легких и кратковременных заболеваний детского возраста. [21] Когда заражение происходит в подростковом или молодом возрасте, оно вызывает инфекционный мононуклеоз в 35–50% случаев. [22]
ВЭБ инфицирует В-клетки иммунной системы и эпителиальные клетки . инфекция EBV Как только первоначальная литическая взята под контроль, латентный период человека EBV сохраняется в В-клетках памяти на всю оставшуюся жизнь. [19] [23] [24]
Вирусология
[ редактировать ]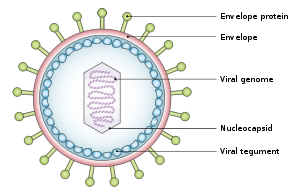
Структура и геном
[ редактировать ]Вирус кодирующих имеет диаметр около 122–180 нм и состоит из двойной спирали дезоксирибонуклеиновой кислоты (ДНК), которая содержит около 172 000 пар оснований, 85 генов . [19] ДНК окружена белковым нуклеокапсидом , который окружен тегументом , состоящим из белка, который, в свою очередь, окружен оболочкой, содержащей как липиды , так и поверхностные проекции гликопротеинов , которые необходимы для заражения клетки-хозяина . [25] В июле 2020 года группа исследователей сообщила о первой полной атомной модели нуклеокапсида вируса. Эта «первая полная атомная модель [включает] икосаэдрический капсид, капсид-ассоциированный тегументный комплекс (CATC) и додекамерный портал — аппарат транслокации вирусного генома». [26] [27]
Тропизм
[ редактировать ]Термин вирусный тропизм относится к тому, какие типы клеток инфицирует ВЭБ. ВЭБ может инфицировать различные типы клеток, включая В-клетки и эпителиальные клетки . [28]
Вирусные трехкомпонентные гликопротеиновые комплексы gHgL gp42 опосредуют слияние мембран В-клеток; хотя двухкомпонентные комплексы gHgL опосредуют слияние мембран эпителиальных клеток. ВЭБ, образующийся в В-клетках, имеет небольшое количество комплексов gHgLgp42, поскольку эти трехчастные комплексы взаимодействуют с молекулами человеческого лейкоцитарного антигена класса II, присутствующими в В-клетках эндоплазматического ретикулума, и разрушаются. Напротив, EBV из эпителиальных клеток богат трехкомпонентными комплексами, поскольку эти клетки обычно не содержат молекул HLA класса II . Как следствие, ВЭБ, полученный из В-клеток, более инфекционен для эпителиальных клеток, а ВЭБ, полученный из эпителиальных клеток, более инфекционен для В-клеток. Вирусы, у которых отсутствует часть gp42 , способны связываться с В-клетками человека, но не способны инфицировать. [29]
Цикл репликации
[ редактировать ]
Вход в камеру
[ редактировать ]ВЭБ может инфицировать как В-клетки, так и эпителиальные клетки. Механизмы проникновения в эти две клетки различны.
Чтобы проникнуть в В-клетки, вирусный гликопротеин gp350 связывается с клеточным рецептором CD21 (также известным как CR2). [30] Затем вирусный гликопротеин gp42 взаимодействует с клеточными молекулами MHC класса II . Это вызывает слияние вирусной оболочки с клеточной мембраной, позволяя ВЭБ проникнуть в В-клетку. [25] CD35 человека, также известный как рецептор комплемента 1 (CR1), является дополнительным фактором прикрепления gp350/220 и может обеспечивать путь проникновения EBV в CD21-негативные клетки, включая незрелые B-клетки. Инфекция EBV снижает экспрессию CD35. [31]
Для проникновения в эпителиальные клетки вирусный белок BMRF-2 взаимодействует с клеточными интегринами β1 . Затем вирусный белок gH/gL взаимодействует с клеточными интегринами αvβ6 / αvβ8 . Это вызывает слияние вирусной оболочки с мембраной эпителиальной клетки, позволяя ВЭБ проникнуть в эпителиальную клетку. [25] В отличие от проникновения B-клеток, проникновение эпителиальных клеток фактически затруднено вирусным гликопротеином gp42. [30]
Как только EBV проникает в клетку, вирусный капсид растворяется, и вирусный геном транспортируется в ядро клетки . [32]
Литическая репликация
[ редактировать ]Литический цикл , или продуктивная инфекция, приводит к образованию инфекционных вирионов . ВЭБ может подвергаться литической репликации как в В-клетках, так и в эпителиальных клетках. В В-клетках литическая репликация обычно происходит только после реактивации из латентного периода . В эпителиальных клетках литическая репликация часто следует непосредственно за проникновением вируса . [25]
Для того чтобы произошла литическая репликация, вирусный геном должен быть линейным. Латентный геном ВЭБ имеет кольцевую форму, поэтому он должен линеаризоваться в процессе литической реактивации. Во время литической репликации вирусная ДНК-полимераза отвечает за копирование вирусного генома. Это контрастирует с латентным периодом, при котором ДНК-полимераза клетки-хозяина копирует вирусный геном. [25]
Продукты литического гена производятся в три последовательные стадии: немедленную-раннюю, раннюю и позднюю. [25] Продукты немедленных и ранних литических генов действуют как трансактиваторы , усиливая экспрессию более поздних литических генов. Продукты немедленно-раннего литического гена включают BZLF1 (также известный как Zta, EB1, связанный с его геном-продуктом ZEBRA ) и BRLF1 (связанный с его геном-продуктом Rta ). [25] Ранние литические генные продукты выполняют гораздо больше функций, таких как репликация, метаболизм и блокада процессинга антигена . Ранние литические генные продукты включают BNLF2 . [25] Наконец, поздние литические генные продукты обычно представляют собой белки со структурной ролью, такие как VCA , который образует вирусный капсид . Другие продукты позднего литического гена, такие как BCRF1, помогают ВЭБ уклоняться от иммунной системы. [25]
EGCG , полифенол, содержащийся в зеленом чае , в исследовании показал, что он ингибирует спонтанную литическую инфекцию EBV на уровне ДНК, транскрипции генов и белка в зависимости от времени и дозы ; экспрессия литических генов EBV Zta, Rta и раннего антигенного комплекса EA-D (индуцированная Rta ), однако высокостабильный ген EBNA-1, обнаруженный на всех стадиях инфекции EBV, не затрагивается. [33] Специфические ингибиторы (путей) предполагают, что путь Ras/MEK/MAPK способствует литической инфекции EBV через путь BZLF1 и PI3-K через BRLF1, причем последний полностью устраняет способность аденовирусного вектора BRLF1 индуцировать литическую форму инфекции EBV. [33] Кроме того, активация некоторых генов, но не других, изучается, чтобы определить, как вызвать иммунное разрушение латентно инфицированных В-клеток с помощью ТРА или бутирата натрия . [33]
Задержка
[ редактировать ]
В отличие от литической репликации, латентный период не приводит к образованию вирионов. [25] Вместо этого кольцевая ДНК генома EBV находится в ядре клетки в виде эписомы и копируется ДНК-полимеразой клетки-хозяина . [25] человека Он сохраняется в В-клетках памяти . [19] [24] Эпигенетические изменения, такие как метилирование ДНК и состав клеточного хроматина , подавляют большинство вирусных генов в латентно инфицированных клетках. [34] только часть генов ВЭБ Экспрессируется , что поддерживает латентное состояние вируса. [34] [19] [35] Латентный ВЭБ экспрессирует свои гены по одному из трех паттернов, известных как латентные программы. ВЭБ может латентно персистировать внутри В-клеток и эпителиальных клеток , но в этих двух типах клеток возможны разные латентные программы. [36] [37]
EBV может иметь одну из трех программ задержки: Latency I, Latency II или Latency III. Каждая латентная программа приводит к выработке ограниченного, отдельного набора вирусных белков и вирусных РНК . [38] [39]
| Ген выражен | ЭБНА-1 | ЭБНА-2 | ЕБНА -3А | ЭБНА-3 Б | ЭБНА -3С | ЭБНА-ЛП | ЛМП1 | ЛМП- 2А | ЛМП-2 Б | ЭБЕР |
|---|---|---|---|---|---|---|---|---|---|---|
| Продукт | Белок | Белок | Белок | Белок | Белок | Белок | Белок | Белок | Белок | нкРНК |
| Задержка I | + | – | – | – | – | – | – | – | – | + |
| Задержка II | + | – | – | – | – | + | + | + | + | + |
| Задержка III | + | + | + | + | + | + | + | + | + | + |
Кроме того, постулируется программа, в которой отключается вся экспрессия вирусных белков (латентность 0). [40]
Внутри В-клеток возможны все три латентные программы. [19] Латентный период EBV внутри B-клеток обычно прогрессирует от латентного периода III до латентного периода II, а затем до латентного периода I. Каждая стадия латентного периода уникальным образом влияет на поведение B-клеток. [19] При инфицировании покоящейся наивной В-клетки ВЭБ переходит в латентный период III. Набор белков и РНК, вырабатываемых в латентном периоде III, превращает В-клетку в пролиферирующий бласт (также известный как активация В-клеток). [19] [25] Позже вирус ограничивает экспрессию своих генов и переходит в латентный период II. Более ограниченный набор белков и РНК, вырабатываемых в Латентном периоде II, побуждает В-клетку дифференцироваться в В-клетку памяти . [19] [25] Наконец, EBV еще больше ограничивает экспрессию генов и переходит в латентный период I. Экспрессия EBNA-1 позволяет геному EBV реплицироваться при делении B-клеток памяти. [19] [25]
Внутри эпителиальных клеток возможна только латентность II. [41]
При первичной инфекции ВЭБ реплицируется в эпителиальных клетках ротоглотки и создает латентную стадию III, II и I инфекции в В-лимфоцитах. Латентная инфекция В-лимфоцитов ВЭБ необходима для персистенции вируса, последующей репликации в эпителиальных клетках и выхода инфекционного вируса в слюну. Латентные инфекции III и II ВЭБ В-лимфоцитов, латентная инфекция II эпителиальных клеток полости рта и латентная II инфекция NK- или Т-клеток могут привести к злокачественным новообразованиям, характеризующимся однородным присутствием генома EBV и экспрессией генов. [42]
Реактивация
[ редактировать ]Латентный EBV в B-клетках может быть реактивирован для переключения на литическую репликацию . Известно, что это происходит in vivo , но что именно вызывает это, точно неизвестно. In vitro латентный EBV в B-клетках может быть реактивирован путем стимуляции рецептора B-клеток, поэтому вполне вероятно, что реактивация in vivo происходит после того, как латентно инфицированные B-клетки отвечают на несвязанные инфекции. [25]
Трансформация В-лимфоцитов
[ редактировать ]Когда EBV инфицирует B-клетки in vitro , в конечном итоге возникают линии лимфобластоидных клеток, способные к неопределенному росту. Ростовая трансформация этих клеточных линий является следствием экспрессии вирусного белка. [43]
EBNA-2, EBNA-3C и LMP-1 необходимы для трансформации, тогда как EBNA-LP и EBER — нет. [44]
Считается, что после естественного заражения EBV вирус выполняет некоторые или все свои программы экспрессии генов, создавая стойкую инфекцию. Учитывая первоначальное отсутствие иммунитета хозяина , литический цикл производит большое количество вирионов для заражения других (предположительно) B-лимфоцитов внутри хозяина.
Латентные программы перепрограммируют и разрушают инфицированные В-лимфоциты, заставляя их пролиферировать и доставлять инфицированные клетки в места, где предположительно сохраняется вирус. В конце концов, когда иммунитет хозяина развивается, вирус сохраняется, отключая большую часть (или, возможно, все) своих генов, и лишь изредка реактивируется и производит вирионы-потомки. В конечном итоге достигается баланс между случайной реактивацией вируса и иммунным надзором хозяина, удаляющим клетки, которые активируют экспрессию вирусных генов. Манипулирование эпигенетикой человеческого организма с помощью EBV может изменить геном клетки, оставив онкогенные фенотипы. [45] В результате модификация EBV увеличивает вероятность развития рака, связанного с EBV. [46] Рак, связанный с ВЭБ, уникален тем, что он часто вызывает эпигенетические изменения, но с меньшей вероятностью мутирует. [47]
Местом персистенции ВЭБ может быть костный мозг . оказываются EBV-отрицательными EBV-положительные пациенты, у которых был заменен костный мозг костного мозга от EBV-отрицательного донора, после трансплантации . [48]
Латентные антигены
[ редактировать ]Все ядерные белки EBV производятся путем альтернативного сплайсинга транскрипта, начинающегося с промоторов Cp или Wp на левом конце генома ( в общепринятой номенклатуре). Гены упорядочены EBNA-LP/EBNA-2/EBNA-3A/EBNA-3B/EBNA-3C/EBNA-1 в геноме .
Инициирующий кодон кодирующей области EBNA-LP создается путем альтернативного сплайсинга транскрипта ядерного белка. В отсутствие этого инициирующего кодона EBNA-2/EBNA-3A/EBNA-3B/EBNA-3C/EBNA-1 будут экспрессироваться в зависимости от того, какой из этих генов альтернативно сплайсирован в транскрипт.
Белок/гены
[ редактировать ]| Белок/ген/антиген | Этап | Описание |
|---|---|---|
| ЭБНА-1 | латентный+литический | Белок EBNA-1 связывается с точкой начала репликации (oriP) внутри вирусного генома и опосредует репликацию и разделение эписомы во время деления клетки-хозяина. Это единственный вирусный белок, экспрессируемый во время латентного периода группы I. |
| ЭБНА-2 | латентный+литический | EBNA-2 является основным вирусным трансактиватором . |
| ЭБНА-3 | латентный+литический | хозяина Эти гены также связывают белок RBP-Jκ . |
| ЛМП-1 | скрытый | LMP-1 представляет собой шестизвенный трансмембранный белок, который также необходим для трансформации роста, опосредованной EBV. |
| ЛМП-2 | скрытый | LMP-2A/LMP-2B представляют собой трансмембранные белки, которые блокируют передачу сигналов тирозинкиназы . |
| ЭБЕР | скрытый | EBER-1/EBER-2 представляют собой небольшие ядерные РНК, которые связываются с определенными нуклеопротеиновыми частицами, обеспечивая связывание с PKR (дцРНК-зависимой серин/треониновой протеинкиназой), тем самым ингибируя ее функцию. EBER на сегодняшний день являются наиболее распространенными продуктами EBV, транскрибируемыми в инфицированных EBV клетках. Их обычно используют в качестве мишеней для обнаружения EBV в гистологических тканях. [49] ЭР-частицы также индуцируют выработку IL-10, который усиливает рост и ингибирует цитотоксические Т-клетки. |
| v-snoRNA1 | скрытый | SnoRNA1 вируса Эпштейна-Барра представляет собой коробчатую CD-snoRNA, генерируемую вирусом в латентном периоде. V-snoRNA1 может действовать как микроРНК-подобный предшественник, который процессируется во фрагменты РНК размером 24 нуклеотида, нацеленные на 3'UTR мРНК вирусной ДНК-полимеразы. [39] |
| ebv-sisРНК | скрытый | Ebv-sisRNA-1 представляет собой стабильную интронную последовательность РНК, генерируемую во время латентной программы III. После EBER это третья по распространенности малая РНК, вырабатываемая вирусом в ходе этой программы. [50] |
| микроРНК | скрытый | МикроРНК EBV кодируются двумя транскриптами: один расположен в гене BART , а другой - рядом с кластером BHRF1 . Три pri-микроРНК BHRF1 (генерирующие четыре микроРНК) экспрессируются во время латентного периода типа III, тогда как большой кластер микроРНК BART (до 20 микроРНК) экспрессируется на высоком уровне во время латентного периода типа II и лишь умеренно во время латентного периода типа I и II. [51] Предыдущая ссылка также дает отчет об известных функциях этих микроРНК. |
| ЭБВ-ЕА | литический | ранний антиген |
| ЭБВ-МА | литический | мембранный антиген |
| EBV-VCA | литический | вирусный капсидный антиген |
| ЭБВ-АН | литический | щелочная нуклеаза [52] |
Подтипы ВЭБ
[ редактировать ]ВЭБ можно разделить на два основных типа: ВЭБ типа 1 и ВЭБ типа 2. Эти два подтипа имеют разные гены EBNA-3 . В результате эти два подтипа различаются по своим преобразующим способностям и способности к реактивации. Тип 1 преобладает на большей части мира, но оба типа одинаково распространены в Африке . Отличить ВЭБ типа 1 от ВЭБ типа 2 можно, разрезав вирусный геном с помощью фермента рестрикции и сравнив полученные закономерности переваривания с помощью гель-электрофореза . [25]
Обнаружение
[ редактировать ]| Типичная интерпретация | VCA-IgG | VCA-IgM | советник | ЭБНА |
|---|---|---|---|---|
| Никогда не заражался | - | - | - | - |
| Острая инфекция (ИМ) | + | + | +/- | - |
| Острая или недавняя инфекция | + | +/- | + | +/- |
| Недавнее перенесенное инфицирование | + | - | +/- | + |
| Заражение в далеком прошлом | + | - | - | + |
| Хроническая инфекция/реактивация | + | - | + | +/- |
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( март 2022 г. ) |
Малые РНК, кодируемые вирусом Эпштейна-Барр (EBER), на сегодняшний день являются наиболее распространенными продуктами EBV, транскрибируемыми в клетках, инфицированных EBV. Их обычно используют в качестве мишеней для обнаружения ВЭБ в гистологических тканях. [49]
Клинически наиболее распространенным способом выявления наличия ВЭБ является иммуноферментный анализ (ИФА). антитела (IgM и IgG) к белкам, кодируемым ДНК ВЭБ. Выявляются [53] Прямое выявление присутствия генома ВЭБ с помощью полимеразной цепной реакции (ПЦР) проводится редко, поскольку этот метод ничего не говорит о реакции иммунной системы на вирус. Вирусная нагрузка ВЭБ плохо коррелирует с клиническими симптомами инфекции. [54]
Роль в болезни
[ редактировать ]- См. также Инфекционный мононуклеоз и другие заболевания, перечисленные в этом разделе.
ВЭБ вызывает инфекционный мононуклеоз. [55] Дети, инфицированные ВЭБ, имеют мало симптомов или могут протекать бессимптомно, но когда заражение происходит в подростковом или взрослом возрасте, оно может вызвать усталость , лихорадку , воспаление горла , увеличение лимфатических узлов на шее, увеличение селезенки , опухание печени или сыпь. [20] Постинфекционный синдром хронической усталости также связан с инфекцией ВЭБ. [56] [57]
ВЭБ также участвует в ряде других заболеваний, включая лимфому Беркитта . [58] гемофагоцитарный лимфогистиоцитоз , [59] лимфома Ходжкина , [60] рак желудка , [12] [61] рак носоглотки , [62] рассеянный склероз , [15] [16] [63] [17] и лимфоматоидный гранулематоз . [64]
В частности, было показано, что В-клетки, инфицированные EBV, находятся в очагах поражения головного мозга пациентов с рассеянным склерозом . [17] а исследование 10 миллионов исторических образцов крови солдат, проведенное в 2022 году, показало, что «Люди, которые не были инфицированы вирусом Эпштейна-Барра, практически никогда не заболевают рассеянным склерозом. Только после заражения вирусом Эпштейна-Барра риск рассеянного склероза возрастает более чем в 30 раз», и что только ВЭБ из многих инфекций имел такую четкую связь с заболеванием. [65]
Дополнительные заболевания, связанные с ВЭБ, включают синдром Джанотти-Крости , мультиформную эритему , острые генитальные язвы и волосатую лейкоплакию полости рта . [66] Вирусная инфекция также связана и часто способствует развитию широкого спектра доброкачественных лимфопролиферативных заболеваний, таких как тяжелые аллергические реакции гиперчувствительности на укусы комаров , [67] слизисто-кожные язвы, положительные на вирус Эпштейна-Барра , и вакциниформная водянка, а также злокачественные лимфопролиферативные заболевания, такие как положительная на вирус Эпштейна-Барра лимфома Беркитта , [68] вирус-положительная лимфома Ходжкина Эпштейна-Барра , [69] и первичная выпотная лимфома . [70]
Вирус Эпштейна-Барр вовлечен в нарушения, связанные с агрегацией альфа-синуклеина (например, болезнь Паркинсона , деменция с тельцами Леви и множественная системная атрофия ). [71]
Было обнаружено, что EBNA1 может вызывать хромосомные разрывы в 11-й хромосоме , особенно в области 11q23 между геном FAM55D и FAM55B, к которой EBNA-1, по-видимому, имеет высокое сродство из-за его ДНК-связывающего домена, проявляющего интерес к специфический палиндромный повтор в этом участке генома. [72] Хотя причина и точный механизм этого неизвестны, побочный продукт приводит к ошибкам и нарушению хромосомной структуры, поскольку клетки, происходящие из линии испорченного генома, подвергаются митозу . Поскольку гены в этой области вовлечены в развитие лейкемии и являются домом для гена-супрессора опухоли , который модифицирован или отсутствует в экспрессии большинства опухолевых генов, была выдвинута гипотеза, что нарушение в этой области является основным виновником рака, что ВЭБ увеличивает вероятность из. Поломка также зависит от дозы: у человека с латентной инфекцией поломка будет меньше, чем у человека с новой или реактивированной инфекцией, поскольку уровни EBNA1 в ядре и ядрышке выше во время активной атаки организма из-за постоянной репликации и поглощения. переизбыток клеток в организме.
История
[ редактировать ]Вирус Эпштейна-Барра был назван в честь М.А. Эпштейна и Ивонн Барр , открывших вирус вместе с Бертом Ачонгом . [73] [74] В 1961 году Эпштейн, патолог и эксперт по электронной микроскопии , посетил лекцию Д. П. Беркитта , хирурга, практикующего в Уганде , на тему «Самый распространенный детский рак в тропической Африке — до сих пор нераспознанный синдром», в которой Беркитт описал «эндемичный вариант» ( педиатрическая форма) заболевания, которое теперь носит его имя . В 1963 году образец был отправлен из Уганды в больницу Миддлсекса для культивирования. Вирусные частицы были идентифицированы в культивируемых клетках, и результаты были опубликованы в журнале The Lancet в 1964 году Эпштейном, Ачонгом и Барром. [74] [75] Клеточные линии были отправлены Вернеру и Гертруде Хенле в Детскую больницу Филадельфии, которые разработали серологические маркеры. [76] В 1967 году у техника в их лаборатории развился мононуклеоз, и они смогли сравнить сохраненный образец сыворотки, показав, что выработались антитела к вирусу. [75] [77] [78] В 1968 году они обнаружили, что ВЭБ может непосредственно иммортализовать В-клетки после заражения, имитируя некоторые формы инфекций, связанных с ВЭБ. [76] и подтвердили связь между вирусом и инфекционным мононуклеозом. [79]
Исследовать
[ редактировать ]Будучи относительно сложным вирусом, ВЭБ еще не до конца изучен. Лаборатории по всему миру продолжают изучать вирус и разрабатывать новые способы лечения заболеваний, которые он вызывает. Одним из популярных способов изучения ВЭБ in vitro является использование искусственных бактериальных хромосом . [80] Вирус Эпштейна-Барра можно поддерживать и манипулировать им в лаборатории в непрерывном латентном периоде (свойство, общее с герпесвирусом, ассоциированным с саркомой Капоши , еще одним из восьми вирусов герпеса человека). Хотя предполагается, что многие вирусы обладают этим свойством во время заражения своих естественных хозяев, не существует легко управляемой системы для изучения этой части жизненного цикла вируса. Геномные исследования EBV позволили изучить литическую реактивацию и регуляцию латентной вирусной эписомы. [81]
Несмотря на активные исследования, вакцина против вируса Эпштейна-Барра еще не доступна. Разработка эффективной вакцины могла бы предотвратить до 200 000 случаев рака во всем мире в год. [12] [13] Отсутствие эффективных моделей на животных является препятствием для разработки профилактических и терапевтических вакцин против ВЭБ. [24]
Как и другие вирусы герпеса человека, вирус Эпштейна-Барра может позволить уничтожить его самостоятельно с помощью курса препарата валацикловир , но необходимы дальнейшие исследования, чтобы определить, действительно ли эрадикация достижима. [45] Противовирусные агенты действуют путем ингибирования репликации вирусной ДНК, но имеется мало доказательств того, что они эффективны против вируса Эпштейна-Барр. Более того, они дороги, рискуют вызвать устойчивость к противовирусным препаратам и (в 1–10% случаев) могут вызывать неприятные побочные эффекты . [46]
См. также
[ редактировать ]- Инфекция вирусом Эпштейна-Барра
- Лимфопролиферативные заболевания, ассоциированные с вирусом Эпштейна-Барра
- Джеймс Корсон Нидерман , врач, доказавший, как передается вирус Эпштейна-Барра при инфекционном мононуклеозе.
Ссылки
[ редактировать ]- ^ «История таксономии ICTV: гаммагерпесвирус человека 4 » . Международный комитет по таксономии вирусов (ICTV) . Проверено 10 января 2019 г.
- ^ Занелла М., Корди С., Кайзер Л. (2020). «Помимо цитомегаловируса и вируса Эпштейна-Барра: обзор вирусов, составляющих виром крови реципиентов трансплантатов твердых органов и гемопоэтических стволовых клеток» . Обзоры клинической микробиологии . 33 (4): e00027-20. дои : 10.1128/CMR.00027-20 . ПМЦ 7462738 . ПМИД 32847820 .
- ^ Актуальность и необходимость профилактических вакцин против вируса Эпштейна-Барр. 2022. НПЖ Вакцины. 7/1. Л. Чжун, К. Крумменачер, В. Чжан, Дж. Хонг, К. Фэн, Ю. Чен и др. дои: 10.1038/s41541-022-00587-6.
- ^ Резк С.А., Чжао X, Вайс Л.М. (сентябрь 2018 г.). «Лимфоидная пролиферация, связанная с вирусом Эпштейна-Барра (ВЭБ), обновленная информация за 2018 год». Патология человека . 79 : 18–41. дои : 10.1016/j.humpath.2018.05.020 . ПМИД 29885408 . S2CID 47010934 .
- ^ Маэда Э., Акахане М., Кирю С., Като Н., Ёсикава Т., Хаяши Н. и др. (январь 2009 г.). «Спектр заболеваний, связанных с вирусом Эпштейна-Барра: иллюстрированный обзор». Японский журнал радиологии . 27 (1): 4–19. дои : 10.1007/s11604-008-0291-2 . ПМИД 19373526 . S2CID 6970917 .
- ^ Черри-Пепперс Дж., Дэниелс К.О., Микс В., Сандерс К.Ф., Резник Д. (февраль 2003 г.). «Оральные проявления в эпоху ВААРТ» . Журнал Национальной медицинской ассоциации . 95 (2, дополнение 2): 21S–32S. ПМЦ 2568277 . ПМИД 12656429 .
- ^ Мастрия Г, Манчини В, Вигано А, ди Пьеро В (2016). «Синдром Алисы в стране чудес: клинический и патофизиологический обзор» . БиоМед Исследования Интернэшнл . 2016 : 8243145. doi : 10.1155/2016/8243145 . ПМК 5223006 . ПМИД 28116304 .
- ^ Нусинович М., Прайс Д., Воловиц Б., Шапиро Р., Амир Дж. (сентябрь 2003 г.). «Постинфекционная острая мозжечковая атаксия у детей». Клиническая педиатрия . 42 (7): 581–584. дои : 10.1177/000992280304200702 . ПМИД 14552515 . S2CID 22942874 .
- ^ Туссиро Э., Рудье Ж (октябрь 2008 г.). «Вирус Эпштейна-Барра при аутоиммунных заболеваниях». Передовая практика и исследования: Клиническая ревматология . 22 (5): 883–896. дои : 10.1016/j.berh.2008.09.007 . ПМИД 19028369 .
- ^ Дрейфус Д.Х. (декабрь 2011 г.). «Аутоиммунные заболевания: роль новых противовирусных методов лечения?». Обзоры аутоиммунитета . 11 (2): 88–97. дои : 10.1016/j.autrev.2011.08.005 . ПМИД 21871974 .
- ^ Пендер, депутат парламента (2012). «Дефицит CD8+ Т-клеток, инфекция вируса Эпштейна-Барр, дефицит витамина D и шаги к аутоиммунитету: объединяющая гипотеза» . Аутоиммунные заболевания . 2012 : 189096. дои : 10.1155/2012/189096 . ПМК 3270541 . ПМИД 22312480 .
- ^ Перейти обратно: а б с «Разработка вакцины против вируса Эпштейна-Барра может предотвратить до 200 000 случаев рака во всем мире, говорят эксперты» . Cancer Research UK (пресс-релиз). 24 марта 2014 г. Архивировано из оригинала 19 марта 2017 г.
- ^ Перейти обратно: а б Хан Дж., Фицморис С., Нагави М., Ахмед Л.А. (август 2020 г.). «Глобальная и региональная заболеваемость, смертность и продолжительность жизни с поправкой на инвалидность для злокачественных новообразований, связанных с вирусом Эпштейна-Барра, 1990–2017 гг.» . БМЖ Опен . 10 (8): e037505. doi : 10.1136/bmjopen-2020-037505 . ПМЦ 7462312 . ПМИД 32868361 .
- ^ Робинсон У.Х., Штейнман Л. (январь 2022 г.). «Вирус Эпштейна-Барра и рассеянный склероз» . Наука . 375 (6578): 264–265. Бибкод : 2022Sci...375..264R . дои : 10.1126/science.abm7930 . ПМИД 35025606 . S2CID 245978874 .
- ^ Перейти обратно: а б Бьёрневик К., Кортезе М., Хили Б.С., Куле Дж., Мина М.Дж., Ленг Ю. и др. (январь 2022 г.). «Продольный анализ показывает высокую распространенность вируса Эпштейна-Барра, связанного с рассеянным склерозом». Наука . 375 (6578). Американская ассоциация содействия развитию науки (AAAS): 296–301. Бибкод : 2022Sci...375..296B . дои : 10.1126/science.abj8222 . ПМИД 35025605 . S2CID 245983763 . Связанная нетехническая статья: Кокс Д. (20 марта 2022 г.). «Можем ли мы сделать прививку от вируса Эпштейна-Барра, о котором вы не знали?» . Наблюдатель .
- ^ Перейти обратно: а б Ашерио А., Мангер К.Л. (сентябрь 2010 г.). «Вирусная инфекция Эпштейна-Барра и рассеянный склероз: обзор». Журнал нейроиммунной фармакологии . 5 (3): 271–277. дои : 10.1007/s11481-010-9201-3 . ПМИД 20369303 . S2CID 24409610 .
- ^ Перейти обратно: а б с Морено М.А., Ор-Гева Н., Афтаб Б.Т., Ханна Р., Кроз Э., Стейнман Л. и др. (июль 2018 г.). «Молекулярная подпись вирусной инфекции Эпштейна-Барра при поражениях головного мозга при рассеянном склерозе» . Неврология . 5 (4): е466. дои : 10.1212/NXI.0000000000000466 . ПМК 5994704 . ПМИД 29892607 .
- ^ Хассани А., Корбой-младший, Аль-Салам С., Хан Г. (2018). «Вирус Эпштейна-Барра присутствует в мозге в большинстве случаев рассеянного склероза и может поражать не только В-клетки» . ПЛОС Один . 13 (2): e0192109. Бибкод : 2018PLoSO..1392109H . дои : 10.1371/journal.pone.0192109 . ПМЦ 5796799 . ПМИД 29394264 .
- ^ Перейти обратно: а б с д и ж г час я дж Амон В., Фаррелл П.Дж. (ноябрь 2004 г.). «Реактивация вируса Эпштейна-Барра из латентного состояния». Обзоры по медицинской вирусологии . 15 (3): 149–156. дои : 10.1002/rmv.456 . ПМИД 15546128 . S2CID 19433994 .
- ^ Перейти обратно: а б «Около 90% взрослых имеют антитела, которые указывают на то, что они в настоящее время или в прошлом перенесли ВЭБ-инфекцию» . Национальный центр инфекционных заболеваний . США Центр по контролю и профилактике заболеваний . 28 сентября 2020 г. Архивировано из оригинала 8 августа 2016 г.
- ^ Штраус С.Е., Коэн Дж.И., Тосато Дж., Мейер Дж. (январь 1993 г.). «Конференция НИЗ. Вирусные инфекции Эпштейна-Барра: биология, патогенез и лечение». Анналы внутренней медицины . 118 (1): 45–58. дои : 10.7326/0003-4819-118-1-199301010-00009 . ПМИД 8380053 . S2CID 53090545 .
- ^ «Вирус Эпштейна-Барра и инфекционный мононуклеоз» . Центры США по контролю и профилактике заболеваний (CDC). Архивировано из оригинала 20 апреля 2012 года . Проверено 29 декабря 2011 г.
- ^ Хан Дж., Мияшита Э.М., Ян Б., Бэбкок Г.Дж., Торли-Лоусон Д.А. (август 1996 г.). «Является ли персистенция EBV in vivo моделью гомеостаза B-клеток?» . Иммунитет . 5 (2): 173–179. дои : 10.1016/s1074-7613(00)80493-8 . ПМИД 8769480 .
- ^ Перейти обратно: а б с Жан-Пьер В., Лупо Дж., Жерми Р. (2021 г.). «Основные цели разработки профилактической или терапевтической вакцины против вируса Эпштейна-Барр» . Границы микробиологии . 12 : 701611. дои : 10.3389/fmicb.2021.701611 . ПМЦ 8258399 . ПМИД 34239514 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Одумаде О.А., Хогквист К.А., Бальфур Х.Х. (январь 2011 г.). «Прогресс и проблемы в понимании и лечении первичных инфекций, вызванных вирусом Эпштейна-Барра» . Обзоры клинической микробиологии . 24 (1): 193–209. дои : 10.1128/CMR.00044-10 . ПМК 3021204 . ПМИД 21233512 .
- ^ Цзя Л (17 июля 2020 г.). «Ученые обнаружили первую атомную структуру нуклеокапсида вируса Эпштейна-Бар» . phys.org (пресс-релиз) . Проверено 4 октября 2020 г.
- ^ Ли З, Чжан Х, Донг Л, Пан Дж, Сюй М, Чжун Q и др. (октябрь 2020 г.). «КриоЭМ-структура тегументированного капсида вируса Эпштейна-Барра» . Клеточные исследования . 30 (10): 873–884. дои : 10.1038/s41422-020-0363-0 . ПМК 7608217 . ПМИД 32620850 . S2CID 220309464 .
- ^ Шеннон-Лоу С., Роу М. (февраль 2014 г.). «Вход вируса Эпштейна-Барра; поцелуи и спряжение» . Современное мнение в вирусологии . 4 : 78–84. дои : 10.1016/j.coviro.2013.12.001 . ПМИД 24553068 .
- ^ Ван X, Хатт-Флетчер LM (январь 1998 г.). «Вирус Эпштейна-Барра, лишенный гликопротеина gp42, может связываться с В-клетками, но не способен инфицировать» . Журнал вирусологии . 72 (1): 158–163. doi : 10.1128/jvi.72.1.158-163.1998 . ПМК 109360 . ПМИД 9420211 .
- ^ Перейти обратно: а б «Ген Энтреза: рецептор 2 компонента комплемента CR2 (3d/вирус Эпштейна-Барра)» . ncbi.nlm.nih.gov . Архивировано из оригинала 5 декабря 2010 года.
- ^ Огембо Дж.Г., Каннан Л., Гиран И., Николсон-Веллер А., Финберг Р.В., Цокос Г.К. и др. (февраль 2013 г.). «Рецептор человеческого комплемента типа 1 / CD35 представляет собой рецептор вируса Эпштейна-Барра» . Отчеты по ячейкам . 3 (2): 371–85. дои : 10.1016/j.celrep.2013.01.023 . ПМЦ 3633082 . ПМИД 23416052 .
- ^ Бушле А., Хаммершмидт В. (апрель 2020 г.). «Эпигенетический образ жизни вируса Эпштейна-Барра» . Семинары по иммунопатологии . 42 (2): 131–142. дои : 10.1007/s00281-020-00792-2 . ПМЦ 7174264 . ПМИД 32232535 .
- ^ Перейти обратно: а б с Лю С., Ли Х., Чен Л., Ян Л., Ли Л., Тао Ю. и др. (март 2013 г.). «(-)-Эпигаллокатехин-3-галлат ингибирование спонтанной литической инфекции вирусом Эпштейна-Барра включает передачу сигналов ERK1/2 и PI3-K/Akt в EBV-положительных клетках». Канцерогенез . 34 (3): 627–637. дои : 10.1093/carcin/bgs364 . ПМИД 23180656 .
- ^ Перейти обратно: а б Бушле А., Хаммершмидт В. (апрель 2020 г.). «Эпигенетический образ жизни вируса Эпштейна-Барра» . Семинары по иммунопатологии . 42 (2): 131–142. дои : 10.1007/s00281-020-00792-2 . ПМЦ 7174264 . ПМИД 32232535 .
- ^ Торли-Лоусон Д.А., Мияшита Э.М., Хан Г. (май 1996 г.). «Вирус Эпштейна-Барра и В-клетка: это все, что нужно». Тенденции в микробиологии . 4 (5): 204–208. дои : 10.1016/s0966-842x(96)90020-7 . ПМИД 8727601 .
- ^ Мерчант М., Сварт Р., Кацман Р.Б., Икеда М., Икеда А., Лонгнекер Р. и др. (2001). «Влияние латентного мембранного белка 2А вируса Эпштейна-Барр на функцию В-клеток». Международные обзоры иммунологии . 20 (6): 805–835. дои : 10.3109/08830180109045591 . ISSN 0883-0185 . ПМИД 11913951 .
- ^ Кикучи К., Иноуэ Х., Миядзаки Ю., Иде Ф., Кодзима М., Кусама К. (2017). «Эпителиальные и неэпителиальные поражения полости рта, ассоциированные с вирусом Эпштейна-Барра (ЭБВ) . Японский обзор стоматологической науки . 53 (3): 95–109. дои : 10.1016/j.jdsr.2017.01.002 . ISSN 1882-7616 . ПМК 5501733 . ПМИД 28725300 .
- ^ Колдервуд М.А., Венкатесан К., Син Л., Чейз М.Р., Васкес А., Холтхаус А.М. и др. (май 2007 г.). «Карты взаимодействия вируса Эпштейна-Барра и вирусных белков человека» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (18): 7606–11. Бибкод : 2007PNAS..104.7606C . дои : 10.1073/pnas.0702332104 . ПМК 1863443 . ПМИД 17446270 . Здесь используется номенклатура Киева. Другие лаборатории используют другие номенклатуры.
- ^ Перейти обратно: а б Хутцингер Р., Фидерле Р., Мразек Дж., Шифермайер Н., Балвирц П.Дж., Заволан М. и др. (август 2009 г.). «Экспрессия и процессинг малой ядрышковой РНК из генома вируса Эпштейна-Барр» . ПЛОС Патогены . 5 (8): e1000547. дои : 10.1371/journal.ppat.1000547 . ПМЦ 2718842 . ПМИД 19680535 .
- ^ Plottel CS, Blaser MJ (октябрь 2011 г.). «Микробиом и злокачественные новообразования» . Клетка-хозяин и микроб . 10 (4): 324–335. дои : 10.1016/j.chom.2011.10.003 . ПМК 3264051 . ПМИД 22018233 .
- ^ Ансари М.А., Сингх В.В., Дутта С., Веттил М.В., Дутта Д., Чикоти Л. и др. (август 2013 г.). «Конститутивный интерферон-индуцируемый белок 16-воспалительная активация во время латентного периода I, II и III вируса Эпштейна-Барра в B и эпителиальных клетках» . Журнал вирусологии . 87 (15): 8606–8623. дои : 10.1128/JVI.00805-13 . ПМК 3719826 . ПМИД 23720728 .
- ^ Робертсон Э.С., изд. (2010). Вирус Эпштейна-Барра: латентный период и трансформация . Кайстер Академик Пресс . ISBN 978-1-904455-62-2 .
- ^ Мюнц С (ноябрь 2019 г.). «Латентность и литическая репликация при онкогенезе, связанном с вирусом Эпштейна-Барра» (PDF) . Обзоры природы Микробиология . 17 (11): 691–700. дои : 10.1038/s41579-019-0249-7 . PMID 31477887 . S2CID 201713873 .
- ^ Йейтс Дж.Л., Уоррен Н., Сагден Б. (1985). «Стабильная репликация плазмид, полученных из вируса Эпштейна-Барр, в различных клетках млекопитающих». Природа . 313 (6005): 812–815. Бибкод : 1985Natur.313..812Y . дои : 10.1038/313812a0 . ПМИД 2983224 . S2CID 4334367 .
- ^ Перейти обратно: а б Хосино Ю., Катано Х., Зоу П., Хохман П., Маркес А., Тайринг С.К. и др. (ноябрь 2009 г.). «Длительное применение валацикловира снижает количество В-клеток, инфицированных вирусом Эпштейна-Барра (ЭБВ), но не количество копий ДНК ВЭБ на В-клетку у здоровых добровольцев» . Журнал вирусологии . 83 (22): 11857–11861. дои : 10.1128/JVI.01005-09 . ПМЦ 2772668 . ПМИД 19740997 .
- ^ Перейти обратно: а б де Паор М., О'Брайен К., Смит С.М. (2016). «Противовирусные средства при инфекционном мононуклеозе (железистой лихорадке)» . Кокрановская база данных систематических обзоров . 2016 (12): CD011487. дои : 10.1002/14651858.CD011487.pub2 . ПМК 6463965 . ПМИД 27933614 .
- ^ Скотт Р.С. (октябрь 2017 г.). «Вирус Эпштейна-Барра: мастер эпигенетического манипулятора» . Современное мнение в вирусологии . 26 : 74–80. дои : 10.1016/j.coviro.2017.07.017 . ПМЦ 5742862 . ПМИД 28780440 .
- ^ Гратама Дж.В., Остервир М.А., Цваан Ф.Е., Лепутр Дж., Кляйн Г., Эрнберг И. (ноябрь 1988 г.). «Ликвидация вируса Эпштейна-Барра путем аллогенной трансплантации костного мозга: последствия для участков латентного вируса» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (22): 8693–8696. Бибкод : 1988PNAS...85.8693G . дои : 10.1073/pnas.85.22.8693 . ПМК 282526 . ПМИД 2847171 .
- ^ Перейти обратно: а б Хан Дж., Коутс П.Дж., Кангро Х.О., Славин Дж. (июль 1992 г.). «Вирус Эпштейна-Барра (ВЭБ), кодирующий малые РНК: мишени для обнаружения путем гибридизации in situ с олигонуклеотидными зондами» . Журнал клинической патологии . 45 (7): 616–620. дои : 10.1136/jcp.45.7.616 . ПМК 495191 . ПМИД 1325480 .
- ^ Мосс В.Н., Стейц Дж.А. (август 2013 г.). «Полногеномный анализ вируса Эпштейна-Барр выявил консервативные структуры РНК и новую стабильную интронную последовательность РНК» . БМК Геномика . 14 :543. дои : 10.1186/1471-2164-14-543 . ПМЦ 3751371 . ПМИД 23937650 .
- ^ Форте Э, Люфтиг Массачусетс (декабрь 2011 г.). «Роль микроРНК в латентном периоде и литической реактивации вируса Эпштейна-Барр» . Микробы заражают . 13 (14, 15): 1156–1167. дои : 10.1016/j.micinf.2011.07.007 . ПМЦ 4911174 . ПМИД 21835261 .
- ^ Бюиссон М., Жеуи Т., Флот Д., Тарбурьех Н., Рессинг М.Е., Вирц Э.Дж. и др. (август 2009 г.). «Мост пересекает каньон активного центра нуклеазы вируса Эпштейна-Барра с активностью ДНКазы и РНКазы». Журнал молекулярной биологии . 391 (4): 717–728. дои : 10.1016/j.jmb.2009.06.034 . ПМИД 19538972 .
- ^ Мэтисон Б.А., Чисхолм С.М., Хо-Йен Д.О. (август 1990 г.). «Оценка экспресс-теста ИФА для выявления вирусной инфекции Эпштейна-Барра» . Журнал клинической патологии . 43 (8): 691–693. дои : 10.1136/jcp.43.8.691 . ISSN 0021-9746 . ПМК 502656 . ПМИД 2169485 .
- ^ Кимура Х., Квонг Ю.Л. (12 февраля 2019 г.). «Вирусная нагрузка EBV в диагностике, мониторинге и оценке ответа» . Границы онкологии . 9:62 . doi : 10.3389/fonc.2019.00062 . ISSN 2234-943X . ПМК 6379266 . ПМИД 30809508 .
- ^ Вайс Л.М., О'Мэлли Д. (январь 2013 г.). «Доброкачественные лимфаденопатии» . Современная патология . 26 (Дополнение 1): С88–С96. дои : 10.1038/modpathol.2012.176 . ПМИД 23281438 .
- ^ Скули RT (1988). «Синдром хронической усталости: проявление вирусной инфекции Эпштейна-Барр?». Актуальные клинические темы инфекционных болезней . 9 : 126–146. ПМИД 2855828 .
- ^ Проал АД, ВанЭльзаккер МБ (23 июня 2021 г.). «Длительный COVID или пост-острые последствия COVID-19 (PASC): обзор биологических факторов, которые могут способствовать стойким симптомам» . Границы микробиологии . 12 : 698169. doi : 10.3389/fmicb.2021.698169 . ПМК 8260991 . ПМИД 34248921 .
- ^ Панноне Дж., Зампарезе Р., Пейс М., Педичилло М.С., Каджано С., Сомма П. и др. (2014). «Роль EBV в патогенезе лимфомы Беркитта: исследование, проведенное в итальянской больнице» . Инфекционные агенты и рак . 9 (1): 34. дои : 10.1186/1750-9378-9-34 . ПМК 4216353 . ПМИД 25364378 .
- ^ Марш РА (2017). «Вирус Эпштейна-Барра и гемофагоцитарный лимфогистиоцитоз» . Границы в иммунологии . 8 : 1902. дои : 10.3389/fimmu.2017.01902 . ПМК 5766650 . ПМИД 29358936 .
- ^ Ганди М.К., Теллам Дж.Т., Ханна Р. (май 2004 г.). «Лиммома Ходжкина, связанная с вирусом Эпштейна-Барра» . Британский журнал гематологии . 125 (3): 267–281. дои : 10.1111/j.1365-2141.2004.04902.x . ПМИД 15086409 . S2CID 2355660 .
- ^ Яу ТО, Тан СМ, Ю Дж (июнь 2014 г.). «Эпигенетическая дисрегуляция при карциноме желудка, связанной с вирусом Эпштейна-Барра: заболевание и лечение» . Всемирный журнал гастроэнтерологии . 20 (21): 6448–6456. дои : 10.3748/wjg.v20.i21.6448 . ПМК 4047330 . ПМИД 24914366 .
- ^ Доган С., Хедберг М.Л., Феррис Р.Л., Рат Т.Дж., Ассаад А.М., Чиосеа С.И. (апрель 2014 г.). «Вирус папилломы человека и вирус Эпштейна-Барра при раке носоглотки в популяции с низкой заболеваемостью» . Голова и шея . 36 (4): 511–516. дои : 10.1002/hed.23318 . ПМЦ 4656191 . ПМИД 23780921 .
- ^ Мечелли Р., Манзари С., Поликано С., Аннесе А., Пикарди Е., Уметон Р. и др. (март 2015 г.). «Генетические варианты вируса Эпштейна-Барра связаны с рассеянным склерозом» . Неврология . 84 (13): 1362–1368. дои : 10.1212/WNL.0000000000001420 . ПМЦ 4388746 . ПМИД 25740864 .
- ^ Тальявини Э., Росси Г., Валли Р., Занелли М., Кадиоли А., Менголи М.К. и др. (август 2013 г.). «Лимфоматоидный гранулематоз: практический обзор для патологов, занимающихся этим редким легочным лимфопролиферативным процессом». Патологика . 105 (4): 111–116. ПМИД 24466760 .
- ^ «Вызывает ли вирус, который есть у всех нас, рассеянный склероз?» . Новости Би-би-си . 13 апреля 2022 г.
- ^ ди Лерния В., Мансури Ю. (октябрь 2013 г.). «Вирус Эпштейна-Барра и кожные проявления в детском возрасте» . Международный журнал дерматологии . 52 (10): 1177–1184. дои : 10.1111/j.1365-4632.2012.05855.x . ПМИД 24073903 . S2CID 30557756 .
- ^ Кириакидис И, Василиу Э, Карастрати С, Трагианнидис А, Гомпакис Н, Хацистилиану М (декабрь 2016 г.). «Первичная инфекция EBV и повышенная чувствительность к укусам комаров: описание случая» . Вирусология Синика . 31 (6): 517–520. дои : 10.1007/s12250-016-3868-4 . ПМК 8193400 . ПМИД 27900557 . S2CID 7996104 .
- ^ Навари М., Этебари М., де Фалько Г., Амбросио М.Р., Джибеллини Д., Леончини Л. и др. (2015). «Присутствие вируса Эпштейна-Барра существенно влияет на профиль транскрипции при лимфоме Беркитта, связанной с иммунодефицитом» . Границы микробиологии . 6 : 556. дои : 10.3389/fmicb.2015.00556 . ПМЦ 4462103 . ПМИД 26113842 .
- ^ Шеннон-Лоу С., Рикинсон А.Б., Белл А.И. (октябрь 2017 г.). «Лимфомы, ассоциированные с вирусом Эпштейна-Барр» . Философские труды Лондонского королевского общества, серия B, Биологические науки . 372 (1732): 20160271. doi : 10.1098/rstb.2016.0271 . ПМЦ 5597738 . ПМИД 28893938 .
- ^ Арора Н., Гупта А., Садеги Н. (июль 2017 г.). «Первичная выпотная лимфома: современные концепции и лечение». Современное мнение в области легочной медицины . 23 (4): 365–370. дои : 10.1097/MCP.0000000000000384 . ПМИД 28399009 . S2CID 4514140 .
- ^ Вульф Дж., Хугендорн Х., Тарнопольски М., Муньос Д.Г. (ноябрь 2000 г.). «Моноклональные антитела против вируса Эпштейна-Барра перекрестно реагируют с альфа-синуклеином в мозге человека». Неврология . 55 (9): 1398–1401. дои : 10.1212/WNL.55.9.1398 . ПМИД 11087792 . S2CID 84387269 .
- ^ Фраппье Л (апрель 2023 г.). «Вирус Эпштейна-Барра — агент геномной нестабильности» . Природа . 616 (7957): 441–442. Бибкод : 2023Natur.616..441F . дои : 10.1038/d41586-023-00936-y . ПМИД 37045952 . S2CID 258110880 .
- ^ МакГрат П. (6 апреля 2014 г.). «Открытию вируса рака способствовала задержка рейса» . Здоровье. Новости Би-би-си . Архивировано из оригинала 8 октября 2015 года . Проверено 4 ноября 2015 г.
- ^ Перейти обратно: а б Эпштейн М.А., Ачонг Б.Г., Барр Ю.М. (март 1964 г.). «Вирусные частицы в культивируемых лимфобластах лимфомы Беркитта». Ланцет . 1 (7335): 702–703. дои : 10.1016/s0140-6736(64)91524-7 . ПМИД 14107961 .
- ^ Перейти обратно: а б Эпштейн М.А. (2005). «1. Истоки исследований ВЭБ: открытие и характеристика вируса» . В Робертсоне ES (ред.). Вирус Эпштейна-Барра . Уаймондхэм, Великобритания: Caister Academic Press. стр. 1–14. ISBN 978-1-904455-03-5 . Проверено 18 сентября 2010 г.
- ^ Перейти обратно: а б Хенле В., Хенле Г. (1980). «Эпидемиологические аспекты заболеваний, связанных с вирусом Эпштейна-Барра (ВЭБ)». Анналы Нью-Йоркской академии наук . 354 : 326–331. дои : 10.1111/j.1749-6632.1980.tb27975.x . ПМИД 6261650 . S2CID 30025994 .
- ^ Робертсон Э.С. (2005). Вирус Эпштейна-Барр . Горизонт Научная Пресса. п. 18. ISBN 978-1-904455-03-5 . Проверено 3 июня 2012 г.
- ^ Миллер Дж. (21 декабря 2006 г.). « Вирус Эпштейна-Барра ». Медицинский журнал Новой Англии (рецензия на книгу). 355 (25): 2708–2709. дои : 10.1056/NEJMbkrev39523 .
- ^ Молодой Л.С. (2009). Настольная энциклопедия человеческой и медицинской вирусологии . Бостон, Массачусетс: Академическая пресса. стр. 532–533.
- ^ Делеклюз Х.Дж., Фидерле Р., Берендс У., Маутнер Дж. (декабрь 2008 г.). «Вклад вирусных рекомбинантов в изучение иммунного ответа против вируса Эпштейна-Барра». Семинары по биологии рака . 18 (6): 409–415. doi : 10.1016/j.semcancer.2008.09.001 . ПМИД 18938248 .
- ^ Арви А., Темпера И., Цай К., Чен Х.С., Тихмянова Н., Кличинский М. и др. (август 2012 г.). «Атлас транскриптома и эпигенома вируса Эпштейна-Барра раскрывает регуляторные взаимодействия вируса-хозяина» . Клетка-хозяин и микроб . 12 (2): 233–245. дои : 10.1016/j.chom.2012.06.008 . ПМЦ 3424516 . ПМИД 22901543 .
Дальнейшее чтение
[ редактировать ]- Чжан С. (3 марта 2022 г.). «Загадочный вирус, который заражает почти всех» . Атлантика .
Внешние ссылки
[ редактировать ]- «Транскриптом и эпигеном ВЭБ» . Пенсильванский университет . Архивировано из оригинала 18 января 2019 года.