Митоз




а. неделящиеся клетки
б. ядра готовятся к делению (стадия спиремы)
в. делящиеся клетки, показывающие фигуры митоза
е. пара дочерних клеток вскоре после деления
Митоз ( / m aɪ ˈ t oʊ s ɪ s / ) — часть клеточного цикла , в ходе которой реплицированные хромосомы разделяются на два новых ядра . Деление клеток митозом — это эквационное деление, в результате которого образуются генетически идентичные клетки, в которых сохраняется общее число хромосом. [ 1 ] Митозу предшествует фаза интерфазы S - (во время которой происходит репликация ДНК ), за ней следуют телофаза и цитокинез , которые делят цитоплазму , органеллы и клеточную мембрану одной клетки на две новые клетки, содержащие примерно равные доли этих клеточных компонентов. [ 2 ] Различные стадии митоза в целом определяют митотическую фазу (М-фазу) клеточного цикла — деление материнской клетки на две дочерние клетки, генетически идентичные друг другу. [ 3 ]
Процесс митоза делится на этапы, соответствующие завершению одного комплекса действий и началу следующего. Этими стадиями являются препрофаза (специфичная для растительных клеток), профаза , прометафаза , метафаза , анафаза и телофаза . Во время митоза хромосомы, уже продублированные во время интерфазы, конденсируются и прикрепляются к волокнам веретена , которые тянут по одной копии каждой хромосомы к противоположным сторонам клетки. [ 4 ] В результате появляются два генетически идентичных дочерних ядра. Остальная часть клетки может затем продолжить деление посредством цитокинеза с образованием двух дочерних клеток. [ 5 ] Различные фазы митоза можно визуализировать в режиме реального времени с помощью визуализации живых клеток . [ 6 ]
Ошибка в митозе может привести к образованию трех или более дочерних клеток вместо обычных двух. Это называется триполярным митозом и мультиполярным митозом соответственно. Эти ошибки могут быть причиной нежизнеспособных эмбрионов, которые не могут имплантироваться . [ 7 ] Другие ошибки во время митоза могут вызвать митотическую катастрофу , апоптоз (запрограммированную гибель клеток) или вызвать мутации . Некоторые виды рака могут возникнуть в результате таких мутаций. [ 8 ]
Митоз происходит только в эукариотических клетках и варьируется у разных организмов. [ 9 ] Например, клетки животных обычно подвергаются открытому митозу, при котором ядерная оболочка разрушается до разделения хромосом, тогда как клетки грибов обычно подвергаются закрытому митозу, при котором хромосомы делятся внутри интактного ядра клетки. [ 10 ] [ 11 ] Большинство клеток животных претерпевают изменение формы, известное как округление митотических клеток , приобретая почти сферическую морфологию в начале митоза. Большинство клеток человека образуются путем митотического деления клеток. Важными исключениями являются гаметы – сперматозоиды и яйцеклетки , которые образуются в результате мейоза . Прокариоты , бактерии и археи, у которых нет настоящего ядра, делятся другим процессом, называемым бинарным делением . [ 12 ]
Открытие
[ редактировать ]В XVIII и XIX веках были сделаны многочисленные описания клеточного деления с разной степенью точности. [ 13 ] В 1835 году немецкий ботаник Гуго фон Моль описал деление клеток зеленых водорослей Cladophora glomerata , заявив, что размножение клеток происходит посредством клеточного деления. [ 14 ] [ 15 ] [ 16 ] В 1838 году Матиас Якоб Шлейден подтвердил, что «образование новых клеток внутри них было общим правилом размножения клеток у растений», точка зрения, позже отвергнутая в пользу модели Моля, благодаря вкладу Роберта Ремака и других. [ 17 ]
В клетках животных деление клеток митозом было обнаружено в клетках роговицы лягушки, кролика и кошки в 1873 году и впервые описано польским гистологом Вацлавом Майзелем в 1875 году. [ 18 ] [ 19 ]
Бючли, Шнайдер и Фоль могли также заявить об открытии процесса, известного сейчас как «митоз». [ 13 ] В 1873 году немецкий зоолог Отто Бючли опубликовал данные наблюдений за нематодами . Несколько лет спустя он открыл и описал митоз на основе этих наблюдений. [ 20 ] [ 21 ] [ 22 ]
Термин «митоз», введенный Вальтером Флеммингом в 1882 году. [ 23 ] происходит от греческого слова μίτος ( митос , «нить основы»). [ 24 ] [ 25 ] Есть несколько альтернативных названий этого процесса. [ 26 ] например, «кариокинез» (деление ядра), термин, введенный Шлейхером в 1878 году, [ 27 ] [ 28 ] или «эквационное деление», предложенное Августом Вейсманом в 1887 году. [ 29 ] Однако термин «митоз» также используется некоторыми авторами в широком смысле для обозначения одновременно кариокинеза и цитокинеза. [ 30 ] В настоящее время «уравненное деление» чаще используется для обозначения мейоза II , части мейоза, наиболее похожей на митоз. [ 31 ]
Фазы
[ редактировать ]Обзор
[ редактировать ]родительской клетки Первичным результатом митоза и цитокинеза является передача генома в две дочерние клетки. Геном состоит из ряда хромосом — комплексов плотно скрученной ДНК , которые содержат генетическую информацию, жизненно важную для правильного функционирования клеток. [ 32 ] Поскольку каждая полученная дочерняя клетка должна быть генетически идентична родительской клетке, родительская клетка должна сделать копию каждой хромосомы перед митозом. Это происходит во время S-фазы интерфазы. [ 33 ] Дупликация хромосом приводит к образованию двух идентичных сестринских хроматид, связанных вместе белками когезина в центромере .
Когда начинается митоз, хромосомы конденсируются и становятся видимыми. У некоторых эукариот, например животных, ядерная оболочка , отделяющая ДНК от цитоплазмы, распадается на мелкие пузырьки. Исчезает и ядрышко , образующее в клетке рибосомы. Микротрубочки выступают из противоположных концов клетки, прикрепляются к центромерам и выравнивают хромосомы по центру клетки. Затем микротрубочки сокращаются, разрывая сестринские хроматиды каждой хромосомы. [ 34 ] Сестринские хроматиды в этот момент называются дочерними хромосомами . По мере удлинения клетки соответствующие дочерние хромосомы притягиваются к противоположным концам клетки и максимально конденсируются в поздней анафазе. Вокруг каждого набора дочерних хромосом формируется новая ядерная оболочка, которая деконденсируется с образованием интерфазных ядер.
Во время митотической прогрессии, обычно после начала анафазы, клетка может подвергаться цитокинезу. В клетках животных клеточная мембрана сжимается внутрь между двумя развивающимися ядрами, образуя две новые клетки. В растительных клетках . клеточная пластинка между двумя ядрами образуется Цитокинез происходит не всегда; ценоцитарные (тип многоядерных клеток) подвергаются митозу без цитокинеза.

Интерфаза
[ редактировать ]Интерфаза — гораздо более длинная фаза клеточного цикла , чем относительно короткая М-фаза. Во время интерфазы клетка готовится к процессу клеточного деления. Интерфаза делится на три подфазы: G 1 (первый разрыв) , S (синтез) и G 2 (второй разрыв) . В течение всех трех частей интерфазы клетка растет, производя белки и цитоплазматические органеллы. Однако хромосомы реплицируются только во время S-фазы . Таким образом, клетка растет (G 1 ), продолжает расти, дублируя свои хромосомы (S), растет больше и готовится к митозу (G 2 ) и, наконец, делится (М) перед возобновлением цикла. [ 33 ] Все эти фазы клеточного цикла в высокой степени регулируются циклинами , циклин-зависимыми киназами и другими белками клеточного цикла. Фазы следуют одна за другой в строгом порядке, и существуют контрольные точки клеточного цикла , которые дают клеточным сигналам переходить или нет от одной фазы к другой. [ 35 ] Клетки также могут временно или навсегда выйти из клеточного цикла и войти в G0 , фазу чтобы прекратить деление. Это может произойти, когда клетки становятся перенаселенными ( зависимое от плотности торможение ) или когда они дифференцируются для выполнения определенных функций организма, как в случае с клетками сердечной мышцы и нейронами человека . Некоторые клетки G0 обладают способностью повторно вступать в клеточный цикл.
Двухцепочечные разрывы ДНК могут быть восстановлены во время интерфазы с помощью двух основных процессов. [ 36 ] Первый процесс, негомологическое соединение концов (NHEJ), может соединять два разорванных конца ДНК в G1 , S и G2 фазах интерфазы . Второй процесс, гомологичная рекомбинационная репарация (HRR), более точен, чем NHEJ, при восстановлении двухцепочечных разрывов. HRR активен во время фаз интерфазы S и G2, когда репликация ДНК либо частично завершена, либо после ее завершения, поскольку HRR требует двух соседних гомологов .
Интерфаза помогает подготовить клетку к митотическому делению. От этого зависит, произойдет ли митотическое деление клеток. Он тщательно останавливает клетку, если ДНК клетки повреждена или не завершила важную фазу. Интерфаза очень важна, поскольку она определяет успешность завершения митоза. Это уменьшит количество поврежденных клеток и производство раковых клеток. Просчет ключевых белков интерфазы может иметь решающее значение, поскольку последние потенциально могут создать раковые клетки. [ 37 ]
Митоз
[ редактировать ]
Препрофаза (растительные клетки)
[ редактировать ]Только в растительных клетках профазе предшествует стадия препрофазы . В сильно вакуолизированных растительных клетках ядро должно мигрировать в центр клетки, прежде чем сможет начаться митоз. Это достигается за счет образования фрагмосомы — поперечного листка цитоплазмы, который делит клетку пополам вдоль будущей плоскости клеточного деления. Помимо образования фрагмосом, препрофаза характеризуется образованием кольца микротрубочек и актиновых нитей (называемого препрофазной полосой ) под плазматической мембраной вокруг экваториальной плоскости будущего митотического веретена . Эта полоса отмечает место, где клетка в конечном итоге будет делиться. В клетках высших растений (например, цветковых ) центриоли отсутствуют ; вместо этого микротрубочки образуют веретено на поверхности ядра, а затем сами хромосомы организуют его в веретено после разрушения ядерной оболочки. [ 38 ] Полоса препрофазы исчезает во время разрушения ядерной оболочки и образования веретена в прометафазе. [ 39 ] : 58–67
Профаза
[ редактировать ]

Во время профазы, которая наступает после интерфазы G2 , клетка готовится к делению путем плотной конденсации своих хромосом и инициирования формирования митотического веретена. Во время интерфазы генетический материал в ядре состоит из рыхло упакованного хроматина . В начале профазы волокна хроматина конденсируются в отдельные хромосомы, которые обычно видны при большом увеличении в световой микроскоп . На этой стадии хромосомы длинные, тонкие и нитевидные. Каждая хромосома имеет две хроматиды. Две хроматиды соединяются в центромере.
Транскрипция гена прекращается во время профазы и не возобновляется до поздней анафазы и начала фазы G1 . [ 40 ] [ 41 ] [ 42 ] Ядрышко также исчезает во время ранней профазы. [ 43 ]
Близко к ядру животной клетки расположены структуры, называемые центросомами , состоящие из пары центриолей, окруженных рыхлым набором белков . клетки Центросома является координирующим центром микротрубочек . Клетка наследует одну центросому при делении клетки, которая дублируется клеткой перед началом нового раунда митоза, образуя пару центросом. Две центросомы полимеризуют тубулин , помогая сформировать веретенообразный аппарат микротрубочек . Затем моторные белки толкают центросомы вдоль этих микротрубочек к противоположным сторонам клетки. Центросомы хотя и помогают организовать сборку микротрубочек, но они не необходимы для формирования веретенообразного аппарата, поскольку отсутствуют у растений. [ 38 ] и не являются абсолютно необходимыми для митоза животных клеток. [ 44 ]
Прометафаза
[ редактировать ]В начале прометафазы в животных клетках фосфорилирование ядерных ламинов приводит к распаду ядерной оболочки на мелкие мембранные пузырьки . Когда это происходит, микротрубочки вторгаются в ядерное пространство. Это называется открытым митозом и встречается у некоторых многоклеточных организмов. Грибы и некоторые протисты , такие как водоросли или трихомонады , подвергаются варианту, называемому закрытым митозом , когда веретено формируется внутри ядра или микротрубочки проникают в неповрежденную ядерную оболочку. [ 45 ] [ 46 ]
В поздней прометафазе кинетохорные микротрубочки начинают искать хромосомные кинетохоры и прикрепляться к ним . [ 47 ] Кинетохор структуру, связывающую микротрубочки, которая образуется на хромосомной представляет собой белковую центромере во время поздней профазы. [ 47 ] [ 48 ] Ряд полярных микротрубочек находят и взаимодействуют с соответствующими полярными микротрубочками противоположной центросомы, образуя митотическое веретено. [ 49 ] Хотя структура и функция кинетохора до конца не изучены, известно, что он содержит некоторую форму молекулярного мотора . [ 50 ] Когда микротрубочка соединяется с кинетохорой, мотор активируется, используя энергию АТФ , чтобы «ползти» вверх по трубке к возникшей центросоме. Эта двигательная активность в сочетании с полимеризацией и деполимеризацией микротрубочек обеспечивает силу тяги, необходимую для последующего разделения двух хроматид хромосомы. [ 50 ]
Метафаза
[ редактировать ]

После того, как микротрубочки расположены и прикреплены к кинетохорам в прометафазе, две центросомы начинают тянуть хромосомы к противоположным концам клетки. Возникающее в результате напряжение заставляет хромосомы выравниваться вдоль метафазной пластинки в экваториальной плоскости, воображаемой линии, расположенной в центре между двумя центросомами (приблизительно на средней линии клетки). [ 49 ] Чтобы гарантировать справедливое распределение хромосом в конце митоза, метафазная контрольная точка гарантирует, что кинетохоры правильно прикреплены к митотическому веретену и что хромосомы выровнены вдоль метафазной пластинки. [ 51 ] Если клетка успешно проходит через контрольную точку метафазы, она переходит в анафазу.
Анафаза
[ редактировать ]
Во время анафазы А когезины, связывающие сестринские хроматиды, расщепляются, образуя две идентичные дочерние хромосомы. [ 52 ] Укорочение микротрубочек кинетохор тянет вновь образовавшиеся дочерние хромосомы к противоположным концам клетки. Во время анафазы B полярные микротрубочки прижимаются друг к другу, заставляя клетку удлиняться. [ 53 ] В поздней анафазе хромосомы также достигают общего максимального уровня конденсации, что способствует сегрегации хромосом и повторному формированию ядра. [ 54 ] В большинстве клеток животных анафаза А предшествует анафазе В, но в некоторых яйцеклетках позвоночных наблюдается противоположный порядок событий. [ 52 ]
Телофаза
[ редактировать ]
Телофаза (от греческого слова τελος, означающего «конец») — это изменение места событий профазы и прометафазы. В телофазе полярные микротрубочки продолжают удлиняться, удлиняя клетку еще больше. Если ядерная оболочка разрушена, новая ядерная оболочка формируется с использованием мембранных везикул старой ядерной оболочки родительской клетки. Новая оболочка формируется вокруг каждого набора разделенных дочерних хромосом (хотя мембрана не окружает центросомы), и снова появляется ядрышко. Оба набора хромосом, теперь окруженные новой ядерной мембраной, начинают «расслабляться» или деконденсироваться. Митоз завершен. Каждое дочернее ядро имеет идентичный набор хромосом. Деление клеток может происходить или не происходить в это время в зависимости от организма.
Цитокинез
[ редактировать ]

Цитокинез — это не фаза митоза, а отдельный процесс, необходимый для завершения деления клетки. В клетках животных на месте метафазной пластинки развивается борозда расщепления (защемление), содержащая сократительное кольцо , отщемляющее отделившиеся ядра. [ 55 ] Как в животных, так и в растительных клетках деление клеток также осуществляется за счет пузырьков, происходящих из аппарата Гольджи , которые перемещаются по микротрубочкам к середине клетки. [ 56 ] У растений эта структура объединяется в клеточную пластинку в центре фрагмопласта и развивается в клеточную стенку, разделяющую два ядра. Фрагмопласт представляет собой структуру микротрубочек, типичную для высших растений, тогда как некоторые зеленые водоросли используют массив микротрубочек фикопласта во время цитокинеза. [ 39 ] : 64–7, 328–9 Каждая дочерняя клетка имеет полную копию генома родительской клетки. Окончание цитокинеза знаменует окончание М-фазы.
Существует множество клеток, в которых митоз и цитокинез происходят раздельно, образуя отдельные клетки с множеством ядер. Наиболее заметно это явление наблюдается среди грибов , слизевиков и ценоцитарных водорослей, но это явление встречается и у различных других организмов. Даже у животных цитокинез и митоз могут происходить независимо, например, на определенных стадиях эмбрионального развития плодовых мух . [ 57 ]
Функция
[ редактировать ]Функция или значение митоза заключается в поддержании хромосомного набора; каждая образовавшаяся клетка получает хромосомы, сходные по составу и равные по количеству хромосомам родительской клетки.
Митоз возникает в следующих случаях:
- Развитие и рост: количество клеток в организме увеличивается за счет митоза. Это основа развития многоклеточного тела из одной клетки, т. е. зиготы , а также основа роста многоклеточного тела.
- Замена клеток. В некоторых частях тела, например, в коже и пищеварительном тракте, клетки постоянно отслаиваются и заменяются новыми. [ 58 ] Новые клетки образуются в результате митоза, а также являются точными копиями заменяемых клеток. Аналогичным образом, эритроциты имеют короткую продолжительность жизни (всего около 3 месяцев), и новые эритроциты образуются в результате митоза. [ 59 ]
- Регенерация: Некоторые организмы могут регенерировать части тела. Производство новых клеток в таких случаях достигается путем митоза. Например, морские звезды регенерируют утраченные руки посредством митоза.
- Бесполое размножение. Некоторые организмы производят генетически сходное потомство посредством бесполого размножения . Например, гидра размножается бесполым путем почкованием. Клетки на поверхности гидры подвергаются митозу и образуют массу, называемую почкой. Митоз продолжается в клетках почки, и из нее вырастает новая особь. при бесполом или вегетативном размножении . Такое же деление происходит у растений
Вариации
[ редактировать ]Формы митоза
[ редактировать ]Процесс митоза в клетках эукариотических организмов следует аналогичной схеме, но с вариациями в трех основных деталях. «Закрытый» и «открытый» митоз можно отличить по тому, остается ли ядерная оболочка неповрежденной или разрушается. Промежуточная форма с частичной деградацией ядерной оболочки называется «полуоткрытым» митозом. Что касается симметрии веретенообразного аппарата во время метафазы, то приблизительно аксиально-симметричная (центрированная) форма называется «ортомитозом» и отличается от эксцентрических веретен «плевромитоза», при котором митотический аппарат имеет двустороннюю симметрию. Наконец, третий критерий — расположение центрального веретена при закрытом плевромитозе: «внеядерное» (веретено расположено в цитоплазме) или «внутриядерное» (в ядре). [ 9 ]
-
 закрыто
закрыто
внутриядерный
плевромитоз -
 закрыто
закрыто
внеядерный
плевромитоз -
 закрыто
закрыто
ортомитоз -
 полуоткрытый
полуоткрытый
плевромитоз -
 полуоткрытый
полуоткрытый
ортомитоз -
 открыть
открыть
ортомитоз






Деление ядра происходит только в клетках организмов эукариотического домена, так как у бактерий и архей ядро отсутствует. Бактерии и археи подвергаются разному типу деления. [ 60 ] [ 61 ] Внутри каждой из супергрупп эукариот можно встретить митоз открытой формы, а также закрытый митоз, за исключением одноклеточных Excavata , у которых наблюдается исключительно закрытый митоз. [ 62 ] Ниже приводится возникновение форм митоза у эукариот: [ 9 ] [ 63 ]
- Закрытый внутриядерный плевромитоз характерен для Foraminifera , некоторых Prasinomonadida , некоторых Kinetoplastida , Oxymonadida , Haplosporidia , многих грибов ( хитриды , оомицеты , зигомицеты , аскомицеты ), некоторых радиолярий ( Spumellaria и Acantharia ); кажется, это самый примитивный тип.
- Закрытый внеядерный плевромитоз встречается у Trichomonadida и Dinoflagellata .
- Закрытый ортомитоз встречается у диатомей , инфузорий , некоторых микроспоридий , одноклеточных дрожжей и некоторых многоклеточных грибов .
- полуоткрытый плевромитоз характерен Для большинства Apicomplexa .
- Полуоткрытый ортомитоз встречается с различными вариантами у некоторых амеб ( Lobosa ) и некоторых зеленых жгутиконосцев (например, Raphidophyta или Volvox ).
- Открытый ортомитоз типичен для млекопитающих и др. многоклеточных животных , а также для наземных растений ; но это также встречается у некоторых простейших.
Ошибки и другие варианты
[ редактировать ]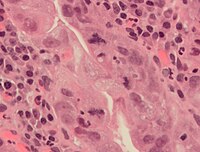
Ошибки могут возникать во время митоза, особенно на ранних стадиях эмбрионального развития человека. [ 64 ] На каждом этапе митоза обычно существуют контрольные точки, которые контролируют нормальный исход митоза. [ 65 ] Но иногда (почти редко) ошибки случаются. Митотические ошибки могут привести к образованию анеуплоидных клеток, в которых содержится слишком мало или слишком много одной или нескольких хромосом, что приводит к раку . [ 66 ] [ 67 ] Ранние человеческие эмбрионы, раковые клетки, инфицированные или интоксицированные клетки также могут страдать от патологического деления на три или более дочерних клеток (триполярный или мультиполярный митоз), что приводит к серьезным ошибкам в их хромосомном наборе. [ 7 ]
При нерасхождении сестринские хроматиды не могут разделиться во время анафазы. [ 68 ] Одна дочерняя клетка получает обе сестринские хроматиды из нерасщепляющейся хромосомы, а другая — ни одной. В результате первая клетка получает три копии хромосомы — состояние, известное как трисомия , а вторая — только одну копию — состояние, известное как моносомия . Иногда, когда клетки испытывают нерасхождение, они не могут завершить цитокинез и сохраняют оба ядра в одной клетке, что приводит к образованию двуядерных клеток . [ 69 ]
Задержка анафазы возникает, когда движение одной хроматиды затруднено во время анафазы. [ 68 ] Это может быть вызвано неспособностью митотического веретена правильно прикрепиться к хромосоме. Отстающая хроматида исключается из обоих ядер и теряется. Следовательно, одна из дочерних клеток будет моносомной по этой хромосоме.
Эндоредупликация (или эндорепликация) происходит, когда хромосомы удваиваются, но клетка впоследствии не делится. В результате образуются полиплоидные клетки или, если хромосомы многократно удваиваются, политенные хромосомы . [ 68 ] [ 70 ] Эндоредупликация обнаружена у многих видов и, по-видимому, является нормальной частью развития . [ 70 ] Эндомитоз — это вариант эндоредупликации, при котором клетки реплицируют свои хромосомы во время S-фазы и вступают в митоз, но преждевременно завершают его. Вместо разделения на два новых дочерних ядра реплицированные хромосомы остаются внутри исходного ядра. [ 57 ] [ 71 ] Затем клетки повторно входят в фазы G1 и S и снова реплицируют свои хромосомы. [ 71 ] Это может происходить несколько раз, увеличивая количество хромосом с каждым раундом репликации и эндомитозом. тромбоциты, , продуцирующие Мегакариоциты во время дифференцировки клеток проходят эндомитоз. [ 72 ] [ 73 ]
Амитоз у инфузорий и в тканях плаценты животных приводит к случайному распределению родительских аллелей.
Кариокинез без цитокинеза приводит к образованию многоядерных клеток, называемых ценоцитами .
Диагностический маркер
[ редактировать ]
В гистопатологии скорость митоза (митотическое количество или митотический индекс) является важным параметром в различных типах образцов тканей для диагностики, а также для дальнейшего уточнения агрессивности опухолей. Например, при классификации рака молочной железы обычно проводится количественная оценка количества митозов . [ 74 ] Митозы необходимо подсчитывать в зоне наибольшей митотической активности. Визуально идентифицировать эти области сложно в опухолях с очень высокой митотической активностью. [ 75 ] Также выявление атипичных форм митоза может использоваться как диагностический, так и прогностический маркер. [ нужна ссылка ] Например, митоз лаг-типа (неприкрепленный конденсированный хроматин в области митотической фигуры) указывает на высокий риск папилломавирусной инфекцией , связанного с рака шейки матки . [ нужна ссылка ] Чтобы улучшить воспроизводимость и точность подсчета митозов, был предложен автоматизированный анализ изображений с использованием алгоритмов глубокого обучения. [ 76 ] Однако необходимы дальнейшие исследования, прежде чем эти алгоритмы можно будет использовать для рутинной диагностики.
-
 Нормальные и атипичные формы митоза раковых клеток. А — нормальный митоз; Б, хроматиновый мостик ; С — мультиполярный митоз; D, кольцевой митоз; Е — рассеянный митоз; F — асимметричный митоз; G — митоз лагового типа; и H, микроядра. Пятно H&E.
Нормальные и атипичные формы митоза раковых клеток. А — нормальный митоз; Б, хроматиновый мостик ; С — мультиполярный митоз; D, кольцевой митоз; Е — рассеянный митоз; F — асимметричный митоз; G — митоз лагового типа; и H, микроядра. Пятно H&E.

Связанные клеточные процессы
[ редактировать ]Округление ячеек
[ редактировать ]
В тканях животных большинство клеток во время митоза округляются до почти сферической формы. [ 77 ] [ 78 ] [ 79 ] В эпителии и эпидермисе эффективный процесс округления коррелирует с правильным выравниванием митотического веретена и последующим правильным расположением дочерних клеток. [ 78 ] [ 79 ] [ 80 ] [ 81 ] Более того, исследователи обнаружили, что если сильно подавлять округление, это может привести к дефектам веретена, в первую очередь к расщеплению полюсов и неспособности эффективно захватывать хромосомы . [ 82 ] Следовательно, округление митотических клеток играет защитную роль в обеспечении точного митоза. считается, что [ 81 ] [ 83 ]
Силы округления обусловлены реорганизацией F-актина и миозина (актомиозина) в сократительную гомогенную клеточную кору , которая: 1) укрепляет периферию клетки. [ 83 ] [ 84 ] [ 85 ] 2) способствует созданию внутриклеточного гидростатического давления (до 10 раз выше, чем в интерфазе ). [ 86 ] [ 87 ] [ 88 ] Генерация внутриклеточного давления особенно важна в условиях ограниченного пространства, что может быть важно в сценарии с тканями, где должны создаваться внешние силы для воздействия на окружающие клетки и/или внеклеточный матрикс . Генерация давления зависит от формин -опосредованной F-актина. нуклеации [ 88 ] и Rho-киназа (ROCK)-опосредованное сокращение миозина II , [ 84 ] [ 86 ] [ 88 ] оба из которых регулируются сигнальными путями RhoA и ECT2. [ 84 ] [ 85 ] посредством активности Cdk1 . [ 88 ] Из-за своей важности в митозе молекулярные компоненты и динамика митотической коры актомиозина являются областью активных исследований.
Митотическая рекомбинация
[ редактировать ]Митотические клетки, облученные рентгеновскими лучами в фазе G1 клеточного цикла, восстанавливают рекомбиногенные повреждения ДНК преимущественно за счет рекомбинации между гомологичными хромосомами . [ 89 ] Митотические клетки, облученные в фазе G2 , восстанавливают такие повреждения преимущественно путем рекомбинации сестринских хроматид . [ 89 ] Мутации в генах , кодирующих ферменты, используемые в рекомбинации, приводят к тому, что клетки становятся более чувствительными к уничтожению различными агентами, повреждающими ДНК. [ 90 ] [ 91 ] [ 92 ] Эти данные позволяют предположить, что митотическая рекомбинация является адаптацией для восстановления повреждений ДНК, в том числе потенциально смертельных.
Эволюция
[ редактировать ]

Существуют прокариотические гомологи всех ключевых молекул эукариотического митоза (например, актинов, тубулинов). Будучи универсальным свойством эукариот, митоз, вероятно, возник у основания эукариотического дерева. Поскольку митоз менее сложен, чем мейоз , мейоз мог возникнуть после митоза. [ 93 ] Однако половое размножение с участием мейоза также является примитивной характеристикой эукариот. [ 94 ] Таким образом, мейоз и митоз могли развиваться параллельно из предковых прокариотических процессов.
В то время как при делении бактериальной клетки после дупликации ДНК две кольцевые хромосомы прикрепляются к особому участку клеточной мембраны, для эукариотического митоза обычно характерно наличие множества линейных хромосом, кинетохоры которых прикрепляются к микротрубочкам веретена. По формам митоза закрытый внутриядерный плевромитоз представляется наиболее примитивным типом, так как он больше похож на деление бактерий. [ 9 ]
Галерея
[ редактировать ]Митотические клетки можно визуализировать микроскопически, окрашивая их флуоресцентными антителами и красителями .
-
 Ранняя профаза : полярные микротрубочки, показанные в виде зеленых нитей, сформировали матрикс вокруг ныне неповрежденного ядра, причем конденсирующиеся хромосомы обозначены синим цветом. Красные узелки — это центромеры.
Ранняя профаза : полярные микротрубочки, показанные в виде зеленых нитей, сформировали матрикс вокруг ныне неповрежденного ядра, причем конденсирующиеся хромосомы обозначены синим цветом. Красные узелки — это центромеры. -
 Ранняя прометафаза : ядерная мембрана только что разобралась, что позволяет микротрубочкам быстро взаимодействовать с кинетохорами, которые собираются на центромерах конденсирующихся хромосом.
Ранняя прометафаза : ядерная мембрана только что разобралась, что позволяет микротрубочкам быстро взаимодействовать с кинетохорами, которые собираются на центромерах конденсирующихся хромосом. -
 Метафаза : Центросомы переместились к полюсам клетки и установили митотическое веретено. Хромосомы съехали в метафазной пластинке.
Метафаза : Центросомы переместились к полюсам клетки и установили митотическое веретено. Хромосомы съехали в метафазной пластинке. -
 Анафаза : микротрубочки кинетохора раздвигают два набора хромосом, а удлинение полярных микротрубочек раздвигает половинки делящейся клетки дальше друг от друга, в то время как хромосомы максимально конденсируются.
Анафаза : микротрубочки кинетохора раздвигают два набора хромосом, а удлинение полярных микротрубочек раздвигает половинки делящейся клетки дальше друг от друга, в то время как хромосомы максимально конденсируются. -
 Телофаза : Реверс событий профазы и прометафазы и, таким образом, завершение клеточного цикла .
Телофаза : Реверс событий профазы и прометафазы и, таким образом, завершение клеточного цикла .





См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Деление и рост клеток» . britannica.com . БРИТАНСКАЯ ЭНЦИКЛОПЕДИЯ. Архивировано из оригинала 28 октября 2018 г. Проверено 4 ноября 2018 г.
- ^ Картер Дж.С. (14 января 2014 г.). «Митоз» . biology.clc.uc.edu . Архивировано из оригинала 27 октября 2012 г. Проверено 12 ноября 2019 г.
- ^ «Митоз — обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 24 ноября 2020 г.
- ^ «Деление клеток: этапы митоза | Изучайте науку в Scitable» . www.nature.com . Архивировано из оригинала 14 ноября 2015 г. Проверено 16 ноября 2015 г.
- ^ Матон А., Хопкинс Дж. Дж., ЛаХарт С., Куон Уорнер Д., Райт М., Джилл Д. (1997). Клетки: строительные блоки жизни . Нью-Джерси: Прентис Холл. стр. 70–4 . ISBN 978-0-13-423476-2 .
- ^ ПО «Сандоз» (декабрь 2019 г.). «Анализ живых клеток млекопитающих на основе изображений с использованием трехмерных карт показателя преломления без меток выявляет новую динамику органелл и поток сухой массы» . ПЛОС Биология . 17 (12): e3000553. doi : 10.1371/journal.pbio.3000553 . ПМЦ 6922317 . ПМИД 31856161 .
- ^ Перейти обратно: а б Калатова Б, Есенска Р, Глинка Д, Дудас М (январь 2015 г.). «Триполярный митоз в клетках и эмбрионах человека: возникновение, патофизиология и медицинские значения» . Акта гистохимика . 117 (1): 111–25. дои : 10.1016/j.acthis.2014.11.009 . ПМИД 25554607 .
- ^ Копс Г.Дж., Уивер Б.А., Кливленд Д.В. (октябрь 2005 г.). «На пути к раку: анеуплоидия и митотическая контрольная точка». Обзоры природы. Рак . 5 (10): 773–85. дои : 10.1038/nrc1714 . ПМИД 16195750 . S2CID 2515388 .
- ^ Перейти обратно: а б с д Райков И.Б. (1994). «Разнообразие форм митоза у простейших: сравнительный обзор». Европейский журнал протистологии . 30 (3): 253–69. дои : 10.1016/S0932-4739(11)80072-6 .
- ^ Де Соуза CP, Османи С.А. (сентябрь 2007 г.). «Митоз, а не только открытый или закрытый» . Эукариотическая клетка . 6 (9): 1521–7. дои : 10.1128/EC.00178-07 . ПМК 2043359 . ПМИД 17660363 .
- ^ Бетчер Б., Баррал Ю. (2013). «Клеточная биология открытого и закрытого митоза» . Ядро . 4 (3): 160–5. дои : 10.4161/nucl.24676 . ПМЦ 3720745 . ПМИД 23644379 .
- ^ Патил, К.с. Клеточная биология . Издательство АПХ. ISBN 978-81-313-0416-7 .
- ^ Перейти обратно: а б Росс, Анна Э. «Анатомия и физиология человека I: Хронология описания митоза». Университет христианских братьев . Проверено 2 мая 2018 г. ссылка. Архивировано 12 мая 2016 г. в Wayback Machine .
- ^ Моля Х. (1835 г.). О размножении растительных клеток путем деления . Вступительная диссертация (диссертация). Тюбинген.
- ^ Карл Мэгдефрау (1994), «Моль, Хьюго фон» , Новая немецкая биография (на немецком языке), том. 17, Берлин: Duncker & Humblot, стр. 690–691 ; ( полный текст онлайн )
- ^ «Заметки и меморандумы: покойный профессор фон Моль». Ежеквартальный журнал микроскопической науки , т. XV, новая серия, с. 178-181, 1875. ссылка .
- ^ Вейерс, Вольфганг (2002). 150 лет деления клеток. Дерматопатология: практическая и концептуальная , Vol. 8, № 2. ссылка. Архивировано 2 апреля 2019 г. на Wayback Machine.
- ^ Комендер Дж (2008). «Несколько слов о докторе Вацлаве Майзеле и его открытии» [О Вацлаве Майзеле и его наблюдениях за митотическим делением] (PDF) . Достижения клеточной биологии (на польском языке). 35 (3): 405–407. Архивировано (PDF) из оригинала 27 октября 2012 г.
- ^ Иловецкий М (1981). История польской науки . Варшава: Издательство Интерпресс. стр. 187. ISBN. 978-83-223-1876-8 .
- ^ Бючли, О. (1873). Вклад в знание свободноживущих нематод. Nova Acta Императорской Леопольдинско-Каролинской немецкой академии естествознания 36, 1-144. ссылка. Архивировано 11 августа 2018 г. на Wayback Machine .
- ^ Бючли, О. (1876). Исследования первых процессов развития яйцеклетки, деления клеток и конъюгации инфузорий. Отд.д. Сенкенб. Натурф. Гес Франкфурт А. М. 10, 213-452. ссылка. Архивировано 9 августа 2018 г. на Wayback Machine .
- ^ Фокин С.И. (2013). «Отто Бючли (1848–1920). Где мы преклоним колени?» (PDF) . Протистология . 8 (1): 22–35. Архивировано (PDF) из оригинала 8 августа 2014 г. Проверено 6 августа 2014 г.
- ^ Шарп Л.В. (1921). Введение в цитологию . Нью-Йорк: McGraw Hill Book Company Inc., с. 143.
- ^ «митоз» . Интернет-словарь этимологии . Архивировано из оригинала 28 сентября 2017 г. Проверено 12 ноября 2019 г.
- ^ μίτος . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ Батталья Э (2009). «Карионема, альтернатива хромосоме, и новая кариологическая номенклатура» (PDF) . Кариология . 62 (4): 1–80. Архивировано из оригинала (PDF) 4 марта 2016 г.
- ^ Шлейхер В. (1878 г.). «Die Knorpelzelltheilung» . Арх. Миркроскоп. Анат . 16 : 248–300. дои : 10.1007/BF02956384 . S2CID 163374324 . Архивировано из оригинала 11 августа 2018 г.
- ^ Топфер Г. «Кариокинез» . Биоконцепции . Архивировано из оригинала 3 мая 2018 г. Проверено 2 мая 2018 г.
- ^ Батталья Э (1987). «Эмбриологические вопросы: 12. Polygonum и Allium Правильно ли установлены типы ?». Энн Бот . 45 . Рим: 81–117.
п. 85: Уже в 1887 году Вейсман дал названия Aequationstheilung обычному делению клеток и Reduktionstheilungen двум делениям, участвующим в процессе деления пополам числа Kernsegmente.
- ^ Маусет Дж.Д. (1991). Ботаника: введение в биологию растений . Филадельфия: Издательство Saunders College. ISBN 9780030302220 .
п. 102: Деление клеток – это цитокинез, а деление ядра – это кариокинез. Слова «митоз» и «мейоз» технически относятся только к кариокинезу, но часто используются и для описания цитокинеза.
- ^ Купер, Джеффри М. (2000). «Мейоз и оплодотворение» . Клетка: молекулярный подход. 2-е издание .
- ^ Браун, Теренс А. (2002). Геном человека . Вили-Лисс.
- ^ Перейти обратно: а б Блоу Джей-Джей, Танака ТУ (ноябрь 2005 г.). «Хромосомный цикл: координация репликации и сегрегации. Второй в серии обзоров циклов» . Отчеты ЭМБО . 6 (11): 1028–34. дои : 10.1038/sj.embor.7400557 . ПМК 1371039 . ПМИД 16264427 .
- ^ Чжоу Дж., Яо Дж., Джоши Х.К. (сентябрь 2002 г.). «Крепление и натяжение в КПП шпиндельного узла» . Журнал клеточной науки . 115 (Часть 18): 3547–55. дои : 10.1242/jcs.00029 . ПМИД 12186941 .
- ^ Биология онлайн (28 апреля 2020 г.). «Митоз» . Биология онлайн .
- ^ Сибата А (2017). «Регуляция выбора пути репарации при двухконцевых разрывах двухцепочечной ДНК». Мутат Рес . 803–805: 51–55. дои : 10.1016/j.mrfmmm.2017.07.011 . ПМИД 28781144 .
- ^ Бернат, РЛ; Борисы, Г.Г.; Ротфилд, Северная Каролина; Эрншоу, WC (1 октября 1990 г.). «Инъекция антицентромерных антител в интерфазе нарушает события, необходимые для движения хромосом при митозе» . Журнал клеточной биологии . 111 (4): 1519–1533. дои : 10.1083/jcb.111.4.1519 . ISSN 0021-9525 . ПМК 2116233 . ПМИД 2211824 .
- ^ Перейти обратно: а б Ллойд С., Чан Дж. (февраль 2006 г.). «Не так разделено: общая основа деления клеток растений и животных». Обзоры природы. Молекулярно-клеточная биология . 7 (2): 147–52. дои : 10.1038/nrm1831 . ПМИД 16493420 . S2CID 7895964 .
- ^ Перейти обратно: а б Рэйвен П.Х., Эверт Р.Ф., Эйххорн С.Е. (2005). Биология растений (7-е изд.). Нью-Йорк: WH Freeman and Co. ISBN 978-0716710073 .
- ^ Прасант К.В., Сакко-Бубуля П.А., Прасант С.Г., Спектор Д.Л. (март 2003 г.). «Последовательное проникновение компонентов механизма экспрессии генов в дочерние ядра» . Молекулярная биология клетки . 14 (3): 1043–57. дои : 10.1091/mbc.E02-10-0669 . ПМК 151578 . ПМИД 12631722 .
- ^ Кадауке С., Блобель Г.А. (апрель 2013 г.). «Митотическая закладка факторами транскрипции» . Эпигенетика и хроматин . 6 (1): 6. дои : 10.1186/1756-8935-6-6 . ПМК 3621617 . ПМИД 23547918 .
- ^ Прескотт Д.М., Бендер М.А. (март 1962 г.). «Синтез РНК и белка во время митоза в клетках культуры тканей млекопитающих». Экспериментальные исследования клеток . 26 (2): 260–8. дои : 10.1016/0014-4827(62)90176-3 . ПМИД 14488623 .
- ^ Олсон М.О. (2011). Ядрышко . Том. 15 обзоров протеинов. Берлин: Springer Science & Business Media. п. 15. ISBN 9781461405146 .
- ^ Басто Р., Лау Дж., Виноградова Т., Гардиол А., Вудс К.Г., Ходжаков А., Рафф Дж.В. (июнь 2006 г.). «Мухи без центриолей» . Клетка . 125 (7): 1375–86. дои : 10.1016/j.cell.2006.05.025 . ПМИД 16814722 . S2CID 2080684 .
- ^ Хейвуд П. (июнь 1978 г.). «Ультраструктура митоза хлоромонадофицеевой водоросли Vacuolaria virescens». Журнал клеточной науки . 31 : 37–51. дои : 10.1242/jcs.31.1.37 . ПМИД 670329 .
- ^ Рибейро К.К., Перейра-Невес А., Бенчимол М. (июнь 2002 г.). «Митотическое веретено и связанные с ним мембраны в закрытом митозе трихомонад». Биология клетки . 94 (3): 157–72. дои : 10.1016/S0248-4900(02)01191-7 . ПМИД 12206655 . S2CID 29081466 .
- ^ Перейти обратно: а б Чан Г.К., Лю С.Т., Йен ТиДжей (ноябрь 2005 г.). «Структура и функции кинетохора». Тенденции в клеточной биологии . 15 (11): 589–98. дои : 10.1016/j.tcb.2005.09.010 . ПМИД 16214339 .
- ^ Чизмен И.М., Десаи А. (январь 2008 г.). «Молекулярная архитектура интерфейса кинетохор-микротрубочки». Обзоры природы. Молекулярно-клеточная биология . 9 (1): 33–46. дои : 10.1038/nrm2310 . ПМИД 18097444 . S2CID 34121605 .
- ^ Перейти обратно: а б Вайни М., Мамай К.Л., О'Тул Э.Т., Мастронард Д.Н., Гиддингс Т.Х., Макдональд К.Л., Макинтош-младший (июнь 1995 г.). «Трехмерный ультраструктурный анализ митотического веретена Saccharomyces cerevisiae» . Журнал клеточной биологии . 129 (6): 1601–15. дои : 10.1083/jcb.129.6.1601 . ПМК 2291174 . ПМИД 7790357 .
- ^ Перейти обратно: а б Майато Х., ДеЛука Дж., Салмон Э.Д., Эрншоу У.К. (ноябрь 2004 г.). «Динамический интерфейс кинетохор-микротрубочки» (PDF) . Журнал клеточной науки . 117 (Часть 23): 5461–77. дои : 10.1242/jcs.01536 . ПМИД 15509863 . S2CID 13939431 . Архивировано (PDF) из оригинала 18 августа 2017 г. Проверено 20 апреля 2018 г.
- ^ Чан Г.К., Йен ТиДжей (2003). «Митотическая контрольная точка: сигнальный путь, который позволяет одному неприкрепленному кинетохору ингибировать выход из митоза». Прогресс в исследованиях клеточного цикла . 5 : 431–9. ПМИД 14593737 .
- ^ Перейти обратно: а б ФитцХаррис Дж. (март 2012 г.). «Анафаза B предшествует анафазе A в яйце мыши» (PDF) . Современная биология . 22 (5): 437–44. Бибкод : 2012CBio...22..437F . дои : 10.1016/j.cub.2012.01.041 . ПМИД 22342753 . Архивировано (PDF) из оригинала 24 июля 2018 г. Проверено 17 сентября 2019 г.
- ^ Миллер К.Р., Левин Дж. (2000). «Анафаза». Биология (5-е изд.). Пирсон Прентис Холл. стр. 169–70 . ISBN 978-0-13-436265-6 .
- ^ Европейская лаборатория молекулярной биологии (12 июня 2007 г.). «Конденсация хромосом посредством митоза» . Наука Дейли . Архивировано из оригинала 13 июня 2007 года . Проверено 4 октября 2020 г.
- ^ Глотцер М. (март 2005 г.). «Молекулярные требования для цитокинеза». Наука . 307 (5716): 1735–9. Бибкод : 2005Sci...307.1735G . дои : 10.1126/science.1096896 . ПМИД 15774750 . S2CID 34537906 .
- ^ Альбертсон Р., Риггс Б., Салливан В. (февраль 2005 г.). «Мембранный трафик: движущая сила цитокинеза». Тенденции в клеточной биологии . 15 (2): 92–101. дои : 10.1016/j.tcb.2004.12.008 . ПМИД 15695096 .
- ^ Перейти обратно: а б Лилли М.А., Дуронио Р.Дж. (апрель 2005 г.). «Новое понимание контроля клеточного цикла эндоцикла дрозофилы» . Онкоген . 24 (17): 2765–75. дои : 10.1038/sj.onc.1208610 . ПМИД 15838513 .
- ^ Сандерленд (2000). Клетка: молекулярный подход. 2-е издание (2-е изд.). Синауэр Ассошиэйтс.
- ^ Франко, Роберт (27 августа 2012 г.). «Измерение продолжительности жизни эритроцитов и старения» . Трансфузиология и гемотерапия . 39 (5): 302–307. дои : 10.1159/000342232 . ПМЦ 3678251 . ПМИД 23801920 .
- ^ Хоган (23 августа 2011 г.). «Архея» . Энциклопедия жизни .
- ^ «Бинарное деление и другие формы размножения бактерий» . Корнеллский колледж сельского хозяйства и наук о жизни .
- ^ Бетчер Б., Баррал Ю. (2013). «Клеточная биология открытого и закрытого митоза» . Ядро . 4 (3): 160–5. дои : 10.4161/nucl.24676 . ПМЦ 3720745 . ПМИД 23644379 .
- ^ Р. Десаль, Б. Шируотер: Ключевые переходы в эволюции животных. CRC Press, 2010, с. 12, ссылка. Архивировано 2 января 2019 г. на Wayback Machine .
- ^ Мантику Э., Вонг К.М., Реппинг С., Мастенбридж С. (декабрь 2012 г.). «Молекулярное происхождение митотических анеуплоидий у предимплантационных эмбрионов» . Биохимия и биофизика Acta (BBA) - Молекулярные основы болезней . 1822 (12): 1921–30. дои : 10.1016/j.bbadis.2012.06.013 . ПМИД 22771499 .
- ^ Вассманн, Катя; Бенезра, Роберт (1 февраля 2001 г.). «Митотические контрольные точки: от дрожжей до рака» . Текущее мнение в области генетики и развития . 11 (1): 83–90. дои : 10.1016/S0959-437X(00)00161-1 . ISSN 0959-437X . ПМИД 11163156 .
- ^ Дравиам В.М., Се С., Зоргер П.К. (апрель 2004 г.). «Сегрегация хромосом и стабильность генома». Текущее мнение в области генетики и развития . 14 (2): 120–5. дои : 10.1016/j.где.2004.02.007 . ПМИД 15196457 .
- ^ Сантагуида С., Амон А. (август 2015 г.). «Кратко- и долгосрочные последствия неправильной сегрегации хромосом и анеуплоидии». Обзоры природы. Молекулярно-клеточная биология . 16 (8): 473–85. дои : 10.1038/nrm4025 . hdl : 1721.1/117201 . ПМИД 26204159 . S2CID 205495880 .
- ^ Перейти обратно: а б с Юров И.Ю., Ворсанова С.Г., Юров Ю.Б. (2006). «Хромосомные вариации в нейрональных клетках млекопитающих: известные факты и привлекательные гипотезы». В Чон К.Дж. (ред.). Международный обзор цитологии: обзор клеточной биологии . Том. 249. Уолтем, Массачусетс: Академическая пресса. п. 146. ИСБН 9780080463506 .
- ^ Ши Q, King RW (октябрь 2005 г.). «Нерасхождение хромосом приводит к образованию тетраплоидных, а не анеуплоидных клеток в клеточных линиях человека». Природа . 437 (7061): 1038–42. Бибкод : 2005Natur.437.1038S . дои : 10.1038/nature03958 . ПМИД 16222248 . S2CID 1093265 .
- ^ Перейти обратно: а б Эдгар Б.А., Орр-Уивер Т.Л. (май 2001 г.). «Клеточные циклы эндорепликации: больше за меньшие деньги» . Клетка . 105 (3): 297–306. дои : 10.1016/S0092-8674(01)00334-8 . ПМИД 11348589 . S2CID 14368177 .
- ^ Перейти обратно: а б Ли Х.О., Дэвидсон Дж.М., Дуронио Р.Дж. (ноябрь 2009 г.). «Эндорепликация: целенаправленная полиплоидия» . Гены и развитие . 23 (21): 2461–77. дои : 10.1101/gad.1829209 . ПМЦ 2779750 . ПМИД 19884253 .
- ^ Итальяно Дж. Э., Шивдасани Р. А. (июнь 2003 г.). «Мегакариоциты и не только: рождение тромбоцитов». Журнал тромбозов и гемостаза . 1 (6): 1174–82. дои : 10.1046/j.1538-7836.2003.00290.x . ПМИД 12871316 . S2CID 24325966 .
- ^ Витрат Н., Коэн-Солал К., Пике С., Ле Куэдик Ж.П., Норол Ф., Ларсен А.К., Кац А., Вайнченкер В., Дебили Н. (май 1998 г.). «Эндомитоз мегакариоцитов человека возникает в результате абортивного митоза» . Кровь . 91 (10): 3711–23. дои : 10.1182/blood.V91.10.3711 . ПМИД 9573008 .
- ^ «Инфильтрирующая протоковая карцинома молочной железы (карцинома особого типа)» . Медицинский факультет Стэнфордского университета . Архивировано из оригинала 11 сентября 2019 г. Проверено 2 октября 2019 г.
- ^ Бертрам К.А., Обревиль М., Гуртнер С., Бартель А., Корнер С.М., Деттвайлер М. и др. (март 2020 г.). «Компьютерный расчет распределения числа митозов в срезах тучных клеток кожи собак: количество митозов зависит от площади» (PDF) . Ветеринарная патология . 57 (2): 214–226. дои : 10.1177/0300985819890686 . ПМИД 31808382 . S2CID 208767801 .
- ^ Бертрам, Кристоф А; Обревиль, Марк; Донован, Тэрин А; Бартель, Александр; Вильм, Фрауке; Марзал, Кристиан; Ассенмахер, Шарль-Антуан; Беккер, Кэтрин; Беннетт, Марк; Корнер, Сара; Коссик, Бриё; Денк, Даниэла; Деттвайлер, Мартина; Гонсалес, Беатрис Гарсия; Гуртнер, Коринн; Хаверкамп, Анн-Катрин; Хейер, Аннабель; Лембекер, Анника; Мерц, Софи; Ноланд, Эрика Л; Плог, Стефани; Шмидт, Аня; Себастьян, Франциска; Сани, Додд Джи; Смедли, Ребекка С; Течилла, Марко; Тайвонг, Таддоу; Фукс-Баумгартингер, Андреа; Меутен, Дональд Дж; Брейнингер, Катарина; Киупель, Матти; Майер, Андреас; Клопфляйш, Роберт (2021). «Компьютерный подсчет митозов с использованием алгоритма, основанного на глубоком обучении, улучшает воспроизводимость и точность результатов между наблюдателями» . Ветеринарная патология . 59 (2): 211–226. дои : 10.1177/03009858211067478 . ПМЦ 8928234 . ПМИД 34965805 . S2CID 245567911 .
- ^ Зауэр (1935). «Митоз в нервной трубке». Журнал сравнительной неврологии . 62 (2): 377–405. дои : 10.1002/cne.900620207 . S2CID 84960254 .
- ^ Перейти обратно: а б Мейер Э.Дж., Икми А., Гибсон MC (март 2011 г.). «Интеркинетическая ядерная миграция является широко консервативной особенностью деления клеток в псевдомногослойном эпителии» . Современная биология . 21 (6): 485–91. Бибкод : 2011CBio...21..485M . дои : 10.1016/j.cub.2011.02.002 . ПМИД 21376598 .
- ^ Перейти обратно: а б Люксенбург С., Пасолли Х.А., Уильямс С.Е., Фукс Э. (март 2011 г.). «Роль Srf в развитии, кортикального цитоскелета и формы клеток в ориентации эпидермального веретена» . Природная клеточная биология . 13 (3): 203–14. дои : 10.1038/Ncb2163 . ПМЦ 3278337 . ПМИД 21336301 .
- ^ Накадзима Ю., Мейер Э.Дж., Кроесен А., МакКинни С.А., Гибсон MC (август 2013 г.). «Эпителиальные соединения поддерживают архитектуру ткани, направляя плоскую ориентацию веретена». Природа . 500 (7462): 359–62. Бибкод : 2013Natur.500..359N . дои : 10.1038/nature12335 . ПМИД 23873041 . S2CID 4418619 .
- ^ Перейти обратно: а б Кадар С., Злотек-Злоткевич Е., Ле Берр М., Пиль М., Мэтьюз Х.К. (апрель 2014 г.). «Изучение функции формы и размера клеток во время митоза» . Развивающая клетка . 29 (2): 159–69. дои : 10.1016/j.devcel.2014.04.009 . ПМИД 24780736 .
- ^ Ланкастер О.М., Ле Берр М., Димитракопулос А., Бонацци Д., Злотек-Злоткевич Е., Пиконе Р., Дьюк Т., Пиль М., Баум Б. (май 2013 г.). «Митотическое округление изменяет геометрию клеток, обеспечивая эффективное формирование биполярного веретена» . Развивающая клетка . 25 (3): 270–83. дои : 10.1016/j.devcel.2013.03.014 . ПМИД 23623611 .
- ^ Перейти обратно: а б Ланкастер О.М., Баум Б. (октябрь 2014 г.). «Формирование для деления: координация ремоделирования цитоскелета актина и микротрубочек во время митоза». Семинары по клеточной биологии и биологии развития . 34 : 109–15. дои : 10.1016/j.semcdb.2014.02.015 . ПМИД 24607328 .
- ^ Перейти обратно: а б с Мэддокс А.С., Берридж К. (январь 2003 г.). «RhoA необходим для ретракции коры и ригидности во время округления митотических клеток» . Журнал клеточной биологии . 160 (2): 255–65. дои : 10.1083/jcb.200207130 . ПМК 2172639 . ПМИД 12538643 .
- ^ Перейти обратно: а б Мэтьюз Х.К., Делабр Ю., Рон Дж.Л., Гак Дж., Кунда П., Баум Б. (август 2012 г.). «Изменения в локализации Ect2 сочетают изменения формы актомиозин-зависимых клеток с митотической прогрессией» . Развивающая клетка . 23 (2): 371–83. дои : 10.1016/j.devcel.2012.06.003 . ПМЦ 3763371 . ПМИД 22898780 .
- ^ Перейти обратно: а б Стюарт М.П., Хелениус Дж., Тойода Ю., Раманатан С.П., Мюллер Д.Д., Хайман А.А. (январь 2011 г.). «Гидростатическое давление и кора актомиозина приводят к округлению митотических клеток». Природа . 469 (7329): 226–30. Бибкод : 2011Natur.469..226S . дои : 10.1038/nature09642 . ПМИД 21196934 . S2CID 4425308 .
- ^ Фишер-Фридрих Э., Хайман А.А., Юлихер Ф., Мюллер Д.Д., Хелениус Дж. (август 2014 г.). «Количественная оценка поверхностного натяжения и внутреннего давления, создаваемого одиночными митотическими клетками» . Научные отчеты . 4 (6213): 6213. Бибкод : 2014NatSR...4E6213F . дои : 10.1038/srep06213 . ПМК 4148660 . ПМИД 25169063 .
- ^ Перейти обратно: а б с д Раманатан С.П., Хелениус Дж., Стюарт М.П., Кэттин С.Дж., Хайман А.А., Мюллер DJ (февраль 2015 г.). «Cdk1-зависимое митотическое обогащение коркового миозина II способствует округлению клеток против удержания». Природная клеточная биология . 17 (2): 148–59. дои : 10.1038/ncb3098 . ПМИД 25621953 . S2CID 5208968 .
- ^ Перейти обратно: а б Кадык Л.К., Хартвелл Л.Х. (октябрь 1992 г.). «Сестринские хроматиды предпочтительнее гомологов в качестве субстратов для рекомбинационной репарации у Saccharomyces cerevisiae» . Генетика . 132 (2): 387–402. дои : 10.1093/генетика/132.2.387 . ПМК 1205144 . ПМИД 1427035 .
- ^ Ботхоф Ю.Г., Бельчик-Мачиньска Е., Феррейра Л., Цвейич А. (май 2017 г.). «rad51 приводит к симптомам, подобным анемии Фанкони, у рыбок данио» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (22): Е4452–Е4461. дои : 10.1073/pnas.1620631114 . ПМЦ 5465903 . ПМИД 28512217 .
Здесь мы приводим in vivo доказательства того, что снижение количества HSPC у взрослых рыб действительно происходит из-за сочетания снижения пролиферации и увеличения апоптоза во время эмбрионального развития. Этот дефект, по-видимому, опосредован через p53(10), поскольку наши двойные мутанты p53/rad51 не обнаруживали каких-либо заметных гематологических дефектов у эмбрионов или взрослых особей.
- ^ Штюрцбехер Х.В., Донзельманн Б., Хеннинг В., Книппшильд У., Буххоп С. (апрель 1996 г.). «p53 напрямую связан с процессами гомологичной рекомбинации посредством взаимодействия белков RAD51/RecA» . Журнал ЭМБО . 15 (8): 1992–2002. дои : 10.1002/j.1460-2075.1996.tb00550.x . ПМК 450118 . ПМИД 8617246 .
- ^ Сонода Э., Сасаки М.С., Бюрстедде Дж.М., Беззубова О., Шинохара А., Огава Х. и др. (январь 1998 г.). «Клетки позвоночных с дефицитом Rad51 накапливают хромосомные разрывы перед смертью клеток» . Журнал ЭМБО . 17 (2): 598–608. дои : 10.1093/emboj/17.2.598 . ПМК 1170409 . ПМИД 9430650 .
- ^ Уилкинс А.С., Холлидей Р. (январь 2009 г.). «Эволюция мейоза из митоза» . Генетика . 181 (1): 3–12. doi : 10.1534/genetics.108.099762 . ПМК 2621177 . ПМИД 19139151 .
- ^ Бернштейн, Х., Бернштейн, К. Эволюционное происхождение и адаптивная функция мейоза. В «Мейозе», Intech Publ (редакторы Кэрол Бернштейн и Харрис Бернштейн), глава 3: 41-75 (2013).
Дальнейшее чтение
[ редактировать ]- Морган Д.Л. (2007). Клеточный цикл: принципы управления . Лондон: Опубликовано New Science Press совместно с Oxford University Press. ISBN 978-0-9539181-2-6 .
- Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Митоз» . Молекулярная биология клетки (4-е изд.). Гирляндная наука . Проверено 22 января 2006 г.
- Кэмпбелл Н., Рис Дж. (декабрь 2001 г.). «Клеточный цикл» . Биология (6-е изд.). Сан-Франциско: Бенджамин Каммингс/Эддисон-Уэсли. стр. 217–224 . ISBN 978-0-8053-6624-2 .
- Купер Дж. (2000). «События фазы М» . Клетка: молекулярный подход (2-е изд.). Синаур Ассошиэйтс, Инк . Проверено 22 января 2006 г.
- Фриман С. (2002). «Деление клеток». Биологическая наука . Река Аппер-Седл, Нью-Джерси: Прентис-Холл. стр. 155–174 . ISBN 978-0-13-081923-9 .
- Лодиш Х., Берк А., Зипурски Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «Обзор клеточного цикла и его контроля» . Молекулярно-клеточная биология (4-е изд.). У. Х. Фриман . Проверено 22 января 2006 г.
Внешние ссылки
[ редактировать ]- Флеш-анимация, сравнивающая митоз и мейоз.
- Академия Хана, лекция
- Изучение митоза в культивируемых клетках млекопитающих
- Общие классные ресурсы K-12 для митоза
- Онтология клеточного цикла
- WormWeb.org: Интерактивная визуализация C. elegans клеточного происхождения — визуализируйте все клеточное дерево происхождения и все клеточные деления нематоды C. elegans.