EZH2
| EZH2 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | EZH2 , ENX-1, ENX1, EZH1, EZH2B, KMT6, KMT6A, WVS, WVS2, энхансер Zeste 2 Polycomb Represse Complex 2 субъединицы 2 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Омим : 601573 ; MGI : 107940 ; Гомологен : 37926 ; GeneCards : EZH2 ; OMA : EZH2 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викидид | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Энхансер Zeste Homolog 2 (EZH2) представляет собой гистон-лизин N-метилтрансфераза фермент ( EC 2.1.1.43 ), кодируемый EZH2 геном , который участвует в метилировании гистонов и, в конечном счете, транскрипционной репрессии . [ 5 ] EZH2 катализирует добавление метильных групп в гистон H3 в лизине 27, [ 6 ] используя кофактор S-аденозил-L-метионин . Активность метилирования EZH2 облегчает образование гетерохроматина , тем самым замолчало функцию гена. [ 5 ] Реконструирование хромосомного гетерохроматина с помощью EZH2 также требуется во время клеточного митоза.
EZH2 является функциональным ферментативным компонентом поликомб -репрессивного комплекса 2 ( PRC2 ), который отвечает за здоровое эмбриональное развитие посредством эпигенетического поддержания генов, ответственных за регулирование развития и дифференцировки . [ 7 ] EZH2 отвечает за метилирование активности PRC2, и комплекс также содержит белки, необходимые для оптимальной функции ( EED , SUZ12 , JARID2 , AEBP2 , RBAP46/48 и PCL ). [ 8 ]
Мутация или чрезмерная экспрессия EZH2 были связаны со многими формами рака. [ 9 ] EZH2 ингибирует гены, ответственные за подавление развития опухоли, и блокирование активности EZH2 может замедлить рост опухоли. EZH2 был нацелен на ингибирование, потому что он активируется при нескольких видах рака, включая, но не ограничиваясь, грудью, [ 10 ] простата , [ 11 ] меланома , [ 12 ] и рак мочевого пузыря. [ 13 ] Мутации в гене EZH2 также связаны с синдромом ткача , редким врожденным расстройством, [ 14 ] и EZH2 участвует в выборе нейродегенеративных симптомов при заболевании нервной системы, атаксии телеангиэктазии . [ 15 ]
Функция
[ редактировать ]| Гистон-лизин N-метилтрансфераза | |||
|---|---|---|---|
| Идентификаторы | |||
| ЕС №. | 2.1.1.43 | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| |||
EZH2 является каталитической субъединицей поликомбного репрессивного комплекса 2 (PRC2). [ 16 ] Каталитическая активность EZH2 зависит от его образования комплекса, по крайней мере, с двумя другими компонентами PRC2, SUZ12 и EED . [ 17 ]
В качестве гистон-метилтрансферазы (HMTase) первичная функция EZH2-метилат Lys -27 на гистоне 3 (H3K27ME) путем переноса метильной группы из кофактора S-аденозил-L-метионина (SAM). EZH2 способен к моно-, ди- и триметилированию H3K27 и был связан с различными биологическими функциями, включая регуляцию транскрипции при гематопоэзе , развитие и дифференцировку клеток . [ 17 ] [ 18 ] [ 19 ] [ 20 ]
EZH2 также был идентифицирован как способный к метилирующим не гистоновым белкам. [ 17 ] [ 18 ]
Репрессия транскрипции
[ редактировать ]EZH2, как часть PRC2, катализирует триметилирование H3K27 ( H3K27ME3 ), которая является модификацией гистонов, которая была охарактеризована как часть кода гистона . [ 16 ] [ 20 ] [ 21 ] [ 22 ] Гистоновой код - это теория, что химические модификации, такие как метилирование , ацетилирование и убиквитинирование , гистоновых белков играют отличительную роль в эпигенетической генов регуляции транскрипции . EZH2-опосредованный катализ H3K27ME3 связан с долгосрочной репрессией транскрипции. [ 16 ] [ 20 ] [ 21 ]
EZH2, а также другие белки группы поликомб, участвуют в создании и поддержании репрессии генов через деление клеток . [ 17 ] [ 20 ] Считается, что это транскрипционно репрессивное состояние обусловлено PRC2/EZH2-EED-опосредованным метилированием H3K27 и последующим рекрутированием PRC1 , что облегчает конденсацию хроматина и образование гетерохроматина . [ 20 ] Гетерохроматин является плотно упакованным хроматином, который ограничивает доступность транскрипционной машины до базовой ДНК, тем самым подавляя транскрипцию. [ 23 ]
Во время деления клеток образование гетерохроматина необходимо для правильной сегрегации хромосом . [ 24 ] Комплекс PRC2/EED-EZH2 также может быть вовлечен в рекрутирование ДНК-метилтрансфераз ( DNMT ), что приводит к увеличению метилирования ДНК , еще одного эпигенетического слоя репрессии транскрипции. [ 16 ] [ 17 ] Специфические гены, которые были идентифицированы как мишени EZH2-опосредованной транскрипционной репрессии, включают HOXA9 , HOXC8 , MYT1 , CDKN2A и ретиноевой кислоты . гены мишеней [ 16 ]
Активация транскрипции
[ редактировать ]При раке EZH2 может играть роль в активации транскрипции, независимо от PRC2 . [ 17 ] Было продемонстрировано, что в клетках рака молочной железы EZH2 активирует гены-мишени NF-κB , которые участвуют в ответах на стимулы. [ 17 ] Функциональная роль этой активности и ее механизм до сих пор неизвестны.
Развитие и дифференциация клеток
[ редактировать ]EZH2 играет важную роль в разработке. В частности, это помогает контролировать транскрипционную репрессию генов, которые регулируют дифференцировку клеток. [ 17 ] [ 18 ] [ 20 ] [ 21 ] В эмбриональных стволовых клетках EZH2-опосредованное триметилирование H3K27ME3 в областях, содержащих гены развития, по-видимому, важно для поддержания нормальной дифференцировки клеток. [ 20 ] H3K27ME3 также важен для управления инактивацией X , молчания одной X-хромосомы у женщин во время развития. [ 22 ] Во время Инактивации X считается, что EZH2 участвует в инициировании образования гетерохроматина путем триметилизирующего H3K27 и что другие гистон-метилтрансферазы и знаки гистонов могут быть вовлечены в поддержание молчаливого состояния. [ 25 ]
Кроме того, EZH2 был идентифицирован как важный белок, участвующий в развитии и дифференцировке B-клеток и Т-клеток . [ 18 ] H3K27ME3 участвует в подавлении генов, которые способствуют дифференцировке, тем самым поддерживая недифференцированное состояние B- и Т-клеток и играют важную роль в регуляции гематопоэза . [ 18 ] [ 26 ] [ 27 ]
Регуляция активности EZH2
[ редактировать ]Активность EZH2 регулируется посттрансляционным фосфорилированием треониновых сериновых и остатков на EZH2. [ 28 ] В частности, фосфорилирование T350 было связано с увеличением активности EZH2, в то время как фосфорилирование T492 и S21 было связано с снижением активности EZH2. [ 21 ] [ 28 ] Предполагается, что фосфорилирование T492 нарушает контакты между человеком EZH2 и его партнерами по связыванию в комплексе PRC2, что препятствует его каталитической активности. [ 21 ]
В дополнение к фосфорилированию также было показано, что активность PRC2/EZH2-EED противоречит транскрипции, активирующих марок гистонов, такими как ацетилирование H3K27 ( H3K27AC ) и метилирование H3K36 ( H3K36ME ). [ 21 ] [ 29 ]
Экспрессия EZH2 регулируется передачей сигналов эстрогена при нормальном эпителии молочной железы человека и рака молочной железы человека. [ 30 ]
Ферментативная активность
[ редактировать ]Функция EZH2 сильно зависит от его найма комплексом PRC2. В частности, эмбриональное развитие белка-эмбриональной эмбриональной эктодермы (EED) и супрессор белка цинкового белка Zeste 12 (SUZ12). для стабилизации взаимодействия EZH2 необходимы [ 31 ] [ 32 ] две изоформы EZH2, полученные из альтернативного сплайсинга : EZH2α и EZH2β. Недавно у людей были идентифицированы [ 33 ] Обе изоформы содержат элементы, которые были идентифицированы как важные для функции EZH2, включая сигнал ядерной локализации , сайты связывания EED и SUZ12, а также консервативный набор. [ 33 ] В большинстве исследований до сих пор было сосредоточено на более длинной изоформе EZH2α, но было показано, что EZH2β, в котором отсутствуют экзоны 4 и 8. [ 33 ] Кроме того, комплексы PRC2/EZH2β действуют на различные гены от его аналогов PRC2/EZH2α, что позволяет предположить, что каждая изоформа может действовать для регулирования специфической подмножества генов. [ 33 ] Дополнительные данные свидетельствуют о том, что EZH2 также может быть способен к метилированию лизина, независимо от ассоциации с PRC2, когда EZH2 сильно активируется. [ 17 ]
Метилирование лизина
[ редактировать ]
Метилирование -это добавление hCh 3 или метильной группы к другой молекуле. В биологии метилирование обычно катализируется ферментами, а метильные группы обычно добавляются либо к белкам, либо к нуклеиновой кислоте. При катализируемом EZH2 метилирование аминокислотный лизин в гистоне H3 метилирован. Этот аминокислотный остаток может быть метилирован до трех раз в ее терминальной группе аммония . Эти метилированные лизины важны для контроля экспрессии генов млекопитающих и играют функциональную роль в образовании гетерохроматина , инактивации X-хромосомы и регуляции транскрипции. [ 34 ] У хромосомов млекопитающих метилирование гистона лизина может либо активировать, либо репрессировать гены в зависимости от сайта метилирования. Недавняя работа показала, что, по крайней мере, часть функции молчания комплекса EZH2 является метилированием гистона H3 на лизине 27. [ 35 ] Метилирование и другие модификации происходят на гистонах. Модификации метила могут влиять на связывание белков с этими гистонами и активировать или ингибировать транскрипцию . [ 24 ]
Механизм катализа
[ редактировать ]
EZH2 является членом семейства SET доменов лизина метилтрансфераз , которые функционируют для добавления метильных групп к боковым цепям лизина субстратных белков. [ 36 ] Настройки метилтрансферазы зависят от кофактора S-аденозилметионина (SAM), чтобы действовать как донор метила для их каталитической активности. Установленные доменные белки отличаются от других SAM-зависимых метилтрансфераз тем, что они связывают свой субстрат и кофактор SAM на противоположных сторонах активного сайта фермента. Эта ориентация субстрата и кофактора позволяет SAM диссоциации без нарушения связывания субстрата и может привести к нескольким раундам метилирования лизина без диссоциации субстрата. [ 36 ]
Хотя не была определена ни подложка, ни связанная с SAM-структура для EZH2, не было определено, выравнивание марки с помощью человека SET7/9 метилтрансфераза показывает консервативные остатки тирозина в почти идентичных положениях в предполагаемом активном участке EZH2.

Ранее было высказано предположение, что тирозин 726 в активном участке EZH2 выступал в качестве общего основания для протонирования лизина субстрата, но эффекты кинетических изотопов показали, что остатки активного сайта не участвуют непосредственно в химии реакции метилтрансферазы. [ 37 ] Вместо этого эти эксперименты подтверждают механизм, в котором остатки опускают PKA остатка лизина субстрата, одновременно предоставляя канал для воды для доступа к боковой цепи лизина во внутренней части активного участка. Тогда вода объемного растворителя может легко депротонировать боковую цепь лизина, активируя ее для нуклеофильной атаки кофактора SAM в S N 2 -подобной реакции, приводящей к переносу метильной группы из SAM к боковой цепи лизина. [ 37 ]

EZH2 в первую очередь катализирует моно- и диметилирование H3K27, но клинически значимая мутация остатков тирозина 641 к фенилаланину (Y641F) приводит к более высокой активности триметилирования H3K27. [ 37 ] [ 38 ] Предполагается, что удаление гидроксильной группы на Y641 устраняет стерические помехи и позволяет размещать третью метильную группу на субстрате лизин.
Клиническое значение
[ редактировать ]Рак
[ редактировать ]EZH2 является привлекательной мишенью для противораковой терапии, потому что он помогает раковым клеткам разделить и пролиферировать. Он обнаруживается в больших количествах, чем в здоровых клетках в широком диапазоне рака, включая рак молочной железы, простату, мочевого пузыря, матки и рака почек , а также меланома и лимфому . EZH2 является супрессором генов , поэтому, когда он становится сверхэкспрессией, многие гены -супрессоры опухолей, которые обычно включаются, выключены. Ингибирование функции EZH2 уменьшает злокачественные опухоли в некоторых случаях, потому что эти гены -супрессоры опухолей не замолчали EZH2. [ 39 ] EZH2 обычно не выражается у здоровых взрослых; Он обнаруживается только в активно делящихся клетках, таких как те, которые активны во время развития плода. [ 40 ] Из -за этой характеристики сверхэкспрессия EZH2 может использоваться в качестве диагностического маркера рака и некоторых нейродегенеративных расстройств. [ 15 ] Тем не менее, есть случаи, когда трудно сказать, является ли сверхэкспрессия EZH2 причиной заболевания или просто следствием. Если это только следствие, нацеливание на EZH2 на ингибирование может не вылечить заболевание. Одним из примеров пути рака, в котором EZH2 играет роль, является путь PRB-E2F. Он находится ниже по течению от пути PRB-E2F , и сигналы от этого пути приводят к сверхэкспрессии EZH2. [ 41 ] Другая важная характеристика EZH2 заключается в том, что когда EZH2 сверхэкспрессирован, он может активировать гены без формирования PRC2 . Это проблема, потому что это означает, что метилирование активности фермента не опосредована комплексным образованием. В клетках рака молочной железы EZH2 активирует гены, которые способствуют пролиферации и выживанию клеток. [ 17 ] Он также может активировать регуляторные гены, такие как C-Myc и Cyclin D1, взаимодействуя с сигнальными факторами Wnt . [ 42 ] Важно отметить, что мутация тирозина 641 в домене активного набора к ряду различных аминокислот является общей чертой некоторых B-клеточных лимфом. [ 43 ]

Ингибиторы
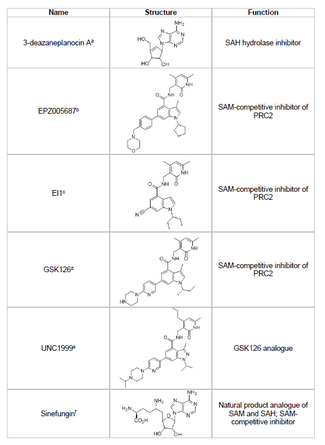
[ редактировать ]Разработка ингибитора EZH2 и предотвращение нежелательного метилирования гистонов генов супрессоров опухолей является жизнеспособной областью исследований рака. Развитие ингибитора EZH2 была сосредоточена на нацеливании на активное место сбора домена белка. Несколько ингибиторов EZH2 были разработаны по состоянию на 2015 год, включая 3-деазанепланоцин A (DZNEP), EPZ005687, EI1, GSK126 и UNC1999.
- DZNEP
- DZNEP обладает потенциальными противовирусными и противораковыми свойствами, поскольку он снижает уровни EZH2 и вызывает апоптоз в клетках рака молочной железы и толстой кишки. [ 44 ] DZNEP ингибирует гидролиз S-аденозил-L-гомоцистеин (SAH), который является ингибитором всех метилтрансфераз белка на основе продукта, что приводит к повышенным клеточным концентрациям SAH, что, в свою очередь, ингибирует EZH2. Тем не менее, DZNEP не специфичен для EZH2, а также ингибирует другие ДНК -метилтрансферазы.
- EPZ005687
- В 2012 году компания под названием Epizyme выявила EPZ005687, S-аденозилметиониновый ( SAM ) конкурентный ингибитор, который более селективный, чем DZNEP; Он имеет 50-кратное увеличение селективности для EZH2 по сравнению с EZH1 . Препарат блокирует активность EZH2 путем связывания с активным сайтом сет -домена фермента. EPZ005687 также может ингибировать мутанты Y641 и A677 EZH2, которые могут быть применимы для лечения неходжкинской лимфомы. [ 45 ]
- Tazemetostat
- В 2013 году эпизим начал клинические испытания фазы I с другим ингибитором EZH2 Tazemetostat (EPZ-6438), для пациентов с B-клеточной лимфомой. [ 49 ] В 2020 году Tazemetostat , с Tradename Tazverik, получил ускоренное одобрение FDA для лечения метастатической или местной эпителиоидной саркомы [ Цитация необходима ] и ускоренное одобрение для лечения пациентов с рецидивирующей фолликулярной лимфомой позже в том же году. [ 50 ]
- Синефунгин
- SINEFUNGIN является еще одним ингибитором SAM-совместимости, однако, как и DZNEP, он не специфичен для EZH2. [ 48 ] Он работает путем связывания в кофакторном связывающем кармане ДНК метилтрансферазы для блокировки переноса метила. EI1 является еще одним ингибитором, разработанным Novartis , который показал ингибирующую активность EZH2 в опухолевых клетках лимфомы, включая клетки с мутацией Y641. [ 46 ] Механизм этого ингибитора также включает в себя конкуренцию с кофактором SAM за связывание с EZH2. [ 46 ]
- GSK126
- GSK126 является мощным, конкурентным SAM-конкурентоспособным ингибитором EZH2, разработанным GlaxoSmithKline , который имеет 150-кратную селективность по сравнению с EZH1 и A K I 0,5-3 нм. [ 47 ] UNC1999 был разработан в качестве аналога GSK126 и был первым перорально биодоступным ингибитором EZH2, чтобы показать активность. Тем не менее, он менее селективен, чем его аналог GSK126, и также связывается с EZH1, увеличивая потенциал для эффектов вне цели.
Комбинированная терапия изучается в качестве возможных методов лечения, когда первичные методы лечения начинают терпеть неудачу. Этопозид , ингибитор топоизомеразы , в сочетании с ингибитором EZH2, становится более эффективным для немелкоклеточных раков легких с мутациями BRG1 и EGFR . [ 39 ] Однако метилирование EZH2 и лизина может обладать активностью подавления опухоли, например, при миелодиспластическом синдроме , [ 51 ] указывая на то, что ингибирование EZH2 не может быть полезным во всех случаях.
Развитие скелета
[ редактировать ]EZH2 имеет решающее значение для эпигенетической регуляции специфического паттерна во время остеохондрогенеза, [ 52 ] или развитие костей и хряща, черепно -лицевых скелетных элементов. Подавляя ингибиторы, EZH2 способствует образованию костей и хряща в скелетных особенностях лица, возникающих в результате нейронного гребня. Выше среднего экспрессия EZH2 стала биологическим маркером для наиболее агрессивной формы рака молочной железы, известной как воспалительный рак молочной железы (IBC). Но в 2013 году исследование, проведенное Zhaomei MU и его партнерами, пришло к выводу, что нокдаунный ген для EZH2 ингибирует как миграцию, так и инвазию в клеток IBC. Также in vivo его нокдаун гена подавлял рост опухоли, скорее всего, из -за наличия меньшего количества кровеносных сосудов или снижения ангиогенеза в опухоли нокдауна EZH2 по сравнению с опухолями EZH2. [ 53 ]
Синдром ткача
[ редактировать ]Мутации в гене EZH2 были связаны с синдромом ткача , редким расстройством, характеризующимся прогрессирующим возрастом кости, макроцефалией и гипертелоризмом . [ 14 ] Остаток гистидина в активном сайте EZH2 дикого типа мутировали к тирозину у пациентов с диагнозом синдрома ткача. [ 14 ] Мутация, вероятно, мешает связыванию кофакторов и вызывает нарушение естественной функции белка. [ 14 ]
Таксономическое распределение
[ редактировать ]
Энхансер Zeste (E (z)) был первоначально идентифицирован у Drosophila melanogaster , а его гомологи млекопитающих были впоследствии идентифицированы и назвали EZH1 (энхансер Zeste Homolog 1) и EZH2 (энхансер Zeste Homolog 2). [ 55 ] EZH2 высоко консервативен благодаря эволюции . Это и его гомологи играют важную роль в развитии, дифференцировке клеток и делении клеток у растений, насекомых, рыб и млекопитающих. [ 17 ] [ 21 ] [ 56 ] [ 57 ] Следующее таксономическое дерево является изображением распределения EZH2 по широкому разнообразию видов. [ 58 ] [ 59 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в GRCH38: Ensembl Release 89: ENSG00000106462 - Ensembl , май 2017 г.
- ^ Jump up to: а беременный в GRCM38: Ensembl Release 89: Ensmusg00000029687 - Ensembl , май 2017 г.
- ^ «Человеческая PubMed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
- ^ Jump up to: а беременный Viré E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, et al. (Февраль 2006 г.). «Поликомбский групповой белок EZH2 непосредственно контролирует метилирование ДНК». Природа . 439 (7078): 871–874. Bibcode : 2006natur.439..871v . doi : 10.1038/nature04431 . PMID 16357870 . S2CID 4409726 .
- ^ Cao R, Wang L, Wang H, Xia L, Erdjument-Bromage H, Tempst P, et al. (Ноябрь 2002). «Роль гистона H3 лизина 27 Метилирование в молчане поликомб-группа». Наука . 298 (5595): 1039–1043. Bibcode : 2002sci ... 298.1039c . doi : 10.1126/science.1076997 . PMID 12351676 . S2CID 6265267 .
- ^ Morey L, Helin K (июнь 2010 г.). «Поликомбинская группа, опосредованная белком, репрессия транскрипции». Тенденции в биохимических науках . 35 (6): 323–332. doi : 10.1016/j.tibs.2010.02.009 . PMID 20346678 .
- ^ Margueron R, Reinberg D (январь 2011 г.). «Комплекс Polycomb PRC2 и его знак в жизни» . Природа . 469 (7330): 343–349. Bibcode : 2011natur.469..343m . doi : 10.1038/nature09784 . PMC 3760771 . PMID 21248841 .
- ^ Ким К.Х., Робертс CW (февраль 2016 г.). «Нацеливание на EZH2 в раке» . Природная медицина . 22 (2): 128–134. doi : 10.1038/nm.4036 . PMC 4918227 . PMID 26845405 .
- ^ Yoo Kh, Hennighausen L (2012). «EZH2 метилтрансфераза и метилирование H3K27 при раке молочной железы» . Международный журнал биологических наук . 8 (1): 59–65. doi : 10.7150/ijbs.8.59 . PMC 3226033 . PMID 22211105 .
- ^ Varambally S, Dhanasekaran SM, Zhou M, Barrette TR, Kumar-Sinha C, Sanda MG, et al. (Октябрь 2002). «Polycomb Group Protein EZH2 участвует в прогрессировании рака простаты». Природа . 419 (6907): 624–629. Bibcode : 2002natur.419..624V . doi : 10.1038/nature01075 . HDL : 2027.42/62896 . PMID 12374981 . S2CID 4414767 .
- Грэм С (10 октября 2002 г.). «Ученые идентифицируют ген, который отмечает самую смертельную форму рака простаты» . Scientific American . Архивировано из оригинала 2015-02-23 . Получено 2015-02-23 .
- ^ Zingg D, Debbache J, Schaefer SM, Tuncer E, Frommel SC, Cheng P, et al. (Январь 2015). «Эпигенетический модификатор EZH2 контролирует рост меланомы и метастазирование посредством молчания отдельных опухолевых супрессоров» . Природная связь . 6 : 6051. Bibcode : 2015natco ... 6.6051z . doi : 10.1038/ncomms7051 . PMID 25609585 .
- «25 января эпигенетический контрольный белок позволяет меланоме клетки метатазировать» . Medicalresearch . 25 января 2015 года. Архивировано с оригинала 23 февраля 2015 года . Получено 23 февраля 2015 года .
- ^ Arrisan S, Buyuktuceer ED, Palavan-Nnsal N, Cakir Да. Полем Internationalis 75 (3): 252–2 doi : 10.1159/00088804 . PMID 16215315 . S2CID 26843362 .
- ^ Jump up to: а беременный в дюймовый Gibson WT, Hood RL, Zhan SH, Bulman DE, Fejes AP, Moore R, et al. (Январь 2012 г.). «Мутации в EZH2 вызывают синдром ткача» . Американский журнал человеческой генетики . 90 (1): 110–118. doi : 10.1016/j.ajhg.2011.11.018 . PMC 3257956 . PMID 22177091 .
- ^ Jump up to: а беременный Li J, Hart RP, Mallimo EM, Swerdel MR, Kusnecov AW, Herrup K (декабрь 2013 г.). «Опосредованное EZH2 триметилирование H3K27 опосредует нейродегенерацию при атаксии-телагиэктазии» . Nature Neuroscience . 16 (12): 1745–1753. doi : 10.1038/nn.3564 . PMC 3965909 . PMID 24162653 .
- ^ Jump up to: а беременный в дюймовый и Универсальный белковый ресурс. Вступление в ресурс Q15910 в Uniprot .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Тан Дж. Г., Ян Й., Ван XX, Цзян Й., Сюй Хе (февраль 2014 г.). «EZH2: биология, болезнь и структура, основанные на открытии лекарств» . Acta Pharmacologica Sinica . 35 (2): 161–174. doi : 10.1038/aps.2013.161 . PMC 3914023 . PMID 24362326 .
- ^ Jump up to: а беременный в дюймовый и Лунд К, Адамс П.Д., Копленд М (январь 2014 г.). «EZH2 в нормальном и злокачественном крове» . Лейкемия . 28 (1): 44–49. doi : 10.1038/leu.2013.288 . PMID 24097338 . S2CID 736796 .
- ^ "Refseq" . Refseq ген EZH2 . Архивировано из оригинала 23 февраля 2015 года . Получено 1 февраля 2015 года .
- ^ Jump up to: а беременный в дюймовый и фон глин Ding X, Wang X, Sontag S, Qin J, Wanek P, Lin Q, et al. (Май 2014). «Поликомб белок EZH2 влияет на индуцированную плюрипотентную генерацию стволовых клеток» . Стволовые клетки и развитие . 23 (9): 931–940. doi : 10.1089/scd.2013.0267 . PMC 3996971 . PMID 24325319 .
- ^ Jump up to: а беременный в дюймовый и фон глин O'Meara MM, Simon Ja (июнь 2012 г.). «Внутренние работы и регулирующие входы, которые контролируют поликомб -репрессивный комплекс 2» . Хромосома . 121 (3): 221–234. doi : 10.1007/s00412-012-0361-1 . PMC 3351537 . PMID 22349693 .
- ^ Jump up to: а беременный «Гистон H3K27» . Эпигени . Архивировано из оригинала 2015-02-23 . Получено 2015-02-23 .
- ^ Grewal Si, Jia S (январь 2007 г.). «Гетерохроматин повторно» . Природные обзоры. Генетика . 8 (1): 35–46. doi : 10.1038/nrg2008 . PMID 17173056 . S2CID 31811880 . Архивировано из оригинала 2021-06-27 . Получено 2018-11-04 .
- ^ Jump up to: а беременный Стюарт MD, Li J, Wong J (апрель 2005 г.). «Связь между гистоном H3 лизин 9 метилирования, репрессии транскрипции и рекрутирования гетерохроматина белка 1» . Молекулярная и клеточная биология . 25 (7): 2525–2538. doi : 10.1128/mcb.25.7.2525-2538.2005 . PMC 1061631 . PMID 15767660 .
- ^ Jentorur P (2008). Эпигенетика и хроматин . Пружины. ISBN 9783540852360 Полем Архивировано из оригинала 2024-04-22 . Получено 2020-10-12 .
- ^ Neo WH, Booth CA, Azzoni E, Chi L, Delgado-olguín P, de Bruijn Mf, et al. (Май 2018). «Клеточно-экстразическое гематопоэтическое влияние инактивации EZH2 в эндотелиальных клетках печени плода» . Кровь . 131 (20): 2223–2234. doi : 10.1182/blood-2017-10-811455 . PMC 5960588 . PMID 29555646 .
- ^ Neo WH, Meng Y, Rodriguez-Meira A, Fadlullah MZ, Booth CA, Azzoni E, et al. (Декабрь 2021 г.). «EZH2 необходим для генерации функционального желточного мешка, полученного эритромелоидными предшественниками» . Природная связь . 12 (1): 7019. Bibcode : 2021Natco..12.7019n . doi : 10.1038/s41467-021-27140-8 . HDL : 10281/339795 . PMC 8640066 . PMID 34857757 .
- ^ Jump up to: а беременный Kaneko S, Li G, Son J, Xu CF, Margueron R, Neubert TA, et al. (Декабрь 2010). «Фосфорилирование компонента PRC2 EZH2 является регулируемым клеточным циклом и усиливает его связывание с NCRNA» . Гены и развитие . 24 (23): 2615–2620. doi : 10.1101/gad.1983810 . PMC 2994035 . PMID 21123648 .
- ^ Tie F, Banerjee R, Stratton CA, Prasad-Sinha J, Stepeanik V, Zlobin A, et al. (Сентябрь 2009 г.). «CBP-опосредованное ацетилирование гистона H3 лизин 27 противодействует молчанию поликомба Drosophila» . Разработка . 136 (18): 3131–3141. doi : 10.1242/dev.037127 . PMC 2730368 . PMID 19700617 .
- ^ Осако Т., Ли Х, Турашвили Г., Чиу Д., МакКинни С., Джустен С.Е. и др. (Май 2020). «Возрастные коррелированные белок и экспрессия транскрипта при раке молочной железы и нормальной ткани молочной железы преобладают эндокринные эффекты хозяина». Природная рак . 1 (5): 518–532. doi : 10.1038/s43018-020-0060-4 . PMID 35121983 . S2CID 218955089 .
- ^ Cao R, Zhang Y (июль 2004 г.). «SUZ12 необходим как для активности гистонметилтрансферазы, так и для функции молчания комплекса EED-EZH2» . Молекулярная клетка . 15 (1): 57–67. doi : 10.1016/j.molcel.2004.06.020 . PMID 15225548 .
- ^ Денизенко О., Шнирева М., Сузуки Х, Бомзтик К (октябрь 1998). «Точечные мутации в домене wd40 eed блокируют его взаимодействие с EZH2» . Молекулярная и клеточная биология . 18 (10): 5634–5642. doi : 10.1128/mcb.18.10.5634 . PMC 109149 . PMID 9742080 .
- ^ Jump up to: а беременный в дюймовый Grzenda A, Lomberk G, Svving P, Mathison A, Calvo E, Iovanna J, et al. (Февраль 2013 г.). «Функциональная характеристика EZH2β выявляет повышенную сложность изоформ EZH2, участвующих в регуляции экспрессии генов млекопитающих» . Эпигенетика и хроматин . 6 (1): 3. DOI : 10.1186/1756-8935-6-3 . PMC 3606351 . PMID 23448518 .
- ^ Мартин С, Чжан Y (ноябрь 2005 г.). «Разнообразные функции метилирования гистонового лизина». Природные обзоры. Молекулярная клеточная биология . 6 (11): 838–849. doi : 10.1038/nrm1761 . PMID 16261189 . S2CID 31300025 .
- ^ Brien GL, Gambero G, O'Connell DJ, Jerman E, Turner SA, Egan CM, et al. (Декабрь 2012 г.). «PolyComb PHF19 связывает H3K36ME3 и рекрутирует PRC2 и деметилазу №66 с генами эмбриональных стволовых клеток во время дифференцировки». Природа структурная и молекулярная биология . 19 (12): 1273–1281. doi : 10.1038/nsmb.2449 . HDL : 2262/97536 . PMID 23160351 . S2CID 1017805 .
- ^ Jump up to: а беременный Dillon SC, Zhang X, Trievel RC, Cheng X (2005). «Суперсемейство белка сет-домен: метилтрансферазы белка» . Биология генома . 6 (8): 227. DOI : 10.1186/GB-2005-6-8-227 . PMC 1273623 . PMID 16086857 .
- ^ Jump up to: а беременный в Kipp DR, Quinn CM, Fortin PD (октябрь 2013 г.). «Фермент-зависимый депротонирование лизина в катализе EZH2». Биохимия . 52 (39): 6866–6878. doi : 10.1021/bi400805w . PMID 24000826 .
- ^ Yap DB, Chu J, Berg T, Schapira M, Cheng SW, Moradian A, et al. (Февраль 2011 г.). «Соматические мутации в EZH2 Y641 активно действуют через механизм селективно измененной каталитической активности PRC2, чтобы увеличить триметилирование H3K27» . Кровь . 117 (8): 2451–2459. doi : 10.1182/blood-2010-11-321208 . PMC 3062411 . PMID 21190999 .
- ^ Jump up to: а беременный «Окно уязвимости» . Гарвардская медицинская школа . 28 января 2015 года. Архивировано с оригинала 23 февраля 2015 года . Получено 23 февраля 2015 года .
- ^ Jump up to: а беременный Konze KD, Ma A, Li F, Barsyte-Lovejoy D, Parton T, Macnevin CJ, et al. (2013). «Перорально биодоступный химический зонд лизина метилтрансферазы EZH2 и EZH1» . ACS Химическая биология . 8 (6): 1324–1334. doi : 10.1021/cb400133j . PMC 3773059 . PMID 23614352 .
- ^ Bracken AP, Pasini D, Capra M, Prosperini E, Colli E, Helin K (октябрь 2003 г.). «EZH2 находится ниже по течению от пути PRB-E2F, необходимо для пролиферации и усиливается при раке» . Embo Journal . 22 (20): 5323–5335. doi : 10.1093/emboj/cdg542 . PMC 213796 . PMID 14532106 .
- ^ Shi B, Liang J, Yang X, Wang Y, Zhao Y, Wu H, et al. (Июль 2007 г.). «Интеграция сигнальных схем эстрогена и Wnt с помощью белка группы Polycomb Group EZH2 в клетках рака молочной железы» . Молекулярная и клеточная биология . 27 (14): 5105–5119. doi : 10.1128/mcb.00162-07 . PMC 1951944 . PMID 17502350 .
- ^ Морин Р.Д., Джонсон Н.А., Северсон Т.М., Мунгалл А.Дж., Дж., Гоя Р. и др. (Февраль 2010 г.). «Соматические мутации, изменяющие EZH2 (TYR641) в фолликулярных и диффузных крупных В-клеточных лимфомах происхождения зародышевого центра» . Природа генетика . 42 (2): 181–185. doi : 10.1038/ng.518 . PMC 2850970 . PMID 20081860 .
- ^ Jump up to: а беременный Tan J, Yang X, Zhuang L, Jiang X, Chen W, Lee PL, et al. (Май 2007). «Фармакологическое нарушение поликомб-репрессивной комплексной 2-опосредованной репрессии генов селективно индуцирует апоптоз в раковых клетках» . Гены и развитие . 21 (9): 1050–1063. doi : 10.1101/gad.1524107 . PMC 1855231 . PMID 17437993 .
- ^ Jump up to: а беременный Knutson SK, Wigle TJ, Warholic NM, Sneeringer CJ, Allain CJ, Klaus CR, et al. (Ноябрь 2012). «Селективный ингибитор EZH2 блокирует метилирование H3K27 и убивает клетки мутантной лимфомы». Природная химическая биология . 8 (11): 890–896. doi : 10.1038/nchembio.1084 . PMID 23023262 .
- ^ Jump up to: а беременный в Qi W, Chan H, Teng L, Li L, Chuai S, Zhang R, et al. (Декабрь 2012 г.). «Селективное ингибирование EZH2 небольшой молекул -ингибитором блокирует пролиферацию опухолевых клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (52): 21360–21365. BIBCODE : 2012PNAS..10921360Q . doi : 10.1073/pnas.1210371110 . PMC 3535655 . PMID 23236167 .
- ^ Jump up to: а беременный McCabe MT, Ott HM, Ganji G, Korenchuk S, Thompson C, Van Aller GS, et al. (Декабрь 2012 г.). «Ингибирование EZH2 как терапевтическая стратегия для лимфомы с активирующими EZH2 мутациями». Природа . 492 (7427): 108–112. Bibcode : 2012natur.492..108m . doi : 10.1038/nature11606 . PMID 23051747 . S2CID 4385729 .
- ^ Jump up to: а беременный Couture JF, Hauk G, Thompson MJ, Blackburn GM, Trievel RC (июль 2006 г.). «Каталитические роли для водородных связей углерода-кислорода в метилтрансферазах лизина в установлении» . Журнал биологической химии . 281 (28): 19280–19287. doi : 10.1074/jbc.m602257200 . PMID 16682405 .
- ^ Эпизим объявил клинические данные из фазы 1 исследования ингибитора EZH2 EPZ-6438 (E7438), которые будут представлены на симпозиуме EORTC-NCI-AACR. (2014, 1 октября).
- ^ «FDA предоставило ускоренное одобрение Tazemetostat для фолликулярной лимфомы» . FDA . 18 июня 2020 года. Архивировано с оригинала 1 августа 2020 года . Получено 27 июля 2020 года .
- ^ Николоски Г., Лангемейер С.М., Куйпер Р.П., Кнопс Р., Массоп М., Тённиссен Э.Р. и др. (Август 2010 г.). «Соматические мутации гена гистонметилтрансферазы EZH2 при миелодиспластических синдромах». Природа генетика . 42 (8): 665–667. doi : 10.1038/ng.620 . PMID 20601954 . S2CID 5814891 .
- ^ Дудакович А., Камиллери И.Т., Сюй Ф., Ристер С.М., МакГи-Лоуренс М.Е., Брэдли Э.В. и др. (Ноябрь 2015). «Эпигенетический контроль развития скелета с помощью гистонметилтрансферазы EZH2» . Журнал биологической химии . 290 (46): 27604–27617. doi : 10.1074/jbc.m115.672345 . PMC 4646012 . PMID 26424790 .
- ^ Mu Z, Li H, Fernandez SV, Alpaugh KR, Zhang R, Cristofanilli M (сентябрь 2013 г.). «Нокдаун EZH2 подавляет рост и инвазию воспалительных клеток рака молочной железы человека» . Журнал экспериментальных и клинических исследований рака . 32 (1): 70. DOI : 10.1186/1756-9966-32-70 . PMC 3850122 . PMID 24294976 .
- ^ "Ensembl" . Геновое дерево ezh2 . Архивировано из оригинала 23 февраля 2015 года . Получено 19 февраля 2015 года .
- ^ Laible G, Wolf A, Dorn R, Reuter G, Nislow C, Lebersorger A, et al. (Июнь 1997 г.). «Гомологи млекопитающих энхансера гена поликомб-группы Zeste опосредуют молчание генов в гетерохроматине дрозофилы и в Tolomeres S. cerevisiae» . Embo Journal . 16 (11): 3219–3232. doi : 10.1093/emboj/16.11.3219 . PMC 1169939 . PMID 9214638 .
- ^ "NCBI Unigene" . Энхансер Zeste Homolog 2 (Drosophila) (EZH2) . Получено 1 февраля 2015 года .
- ^ "GeneCards" . Энхансер Zeste Homolog 2 (Drosophila) . Архивировано из оригинала 23 февраля 2015 года . Получено 1 февраля 2015 года .
- ^ "Ensembl" . Геновое дерево ezh2 . Архивировано из оригинала 22 апреля 2024 года . Получено 1 февраля 2015 года .
- ^ Flicek P, Amode MR, Barrell D, Beal K, Billis K, Brent S, et al. (Январь 2014). "Ensembl 2014" . Исследование нуклеиновых кислот . 42 (выпуск базы данных): D749 - D755. doi : 10.1093/nar/gkt1196 . PMC 3964975 . PMID 24316576 .
Дальнейшее чтение
[ редактировать ]- Zeidler M, Kleer CG (сентябрь 2006 г.). «Поликомбский групповый белок энхансер Zeste 2: его связь с репарацией ДНК и раком молочной железы». Журнал молекулярной гистологии . 37 (5–7): 219–223. doi : 10.1007/s10735-006-9042-9 . PMID 16855786 . S2CID 2332105 .
- De Haan G, Gerrits A (июнь 2007 г.). «Эпигенетический контроль гематопоэтических стволовых клеток. Старение EZH2». Анналы нью -йоркской академии наук . 1106 (1): 233–239. Bibcode : 2007nyasa1106..233d . doi : 10.1196/annals.1392.008 . PMID 17332078 . S2CID 25177748 .
- Hobert O, Jallal B, Ullrich A (июнь 1996 г.). «Взаимодействие VAV с ENX-1, предполагаемым регулятором транскрипции экспрессии гена гомеобокс» . Молекулярная и клеточная биология . 16 (6): 3066–3073. doi : 10.1128/mcb.16.6.3066 . PMC 231301 . PMID 8649418 .
- Бональдо М.Ф., Леннон Г., Соарс М.Б. (сентябрь 1996 г.). «Нормализация и вычитание: два подхода для облегчения открытия генов» . Исследование генома . 6 (9): 791–806. doi : 10.1101/gr.6.9.791 . PMID 8889548 .
- Абель К.Дж., Броди Л.К., Вальдес Дж.М., Эрдос М.Р., МакКинли Д.Р., Кастилья Л.Х. и др. (Октябрь 1996). «Характеристика EZH1, человеческого гомолога Drosophila Enhancer Zeste вблизи BRCA1» . Геномика . 37 (2): 161–171. doi : 10.1006/geno.1996.0537 . PMID 8921387 .
- Laible G, Wolf A, Dorn R, Reuter G, Nislow C, Lebersorger A, et al. (Июнь 1997 г.). «Гомологи млекопитающих энхансера гена поликомб-группы Zeste опосредуют молчание генов в гетерохроматине дрозофилы и в Tolomeres S. cerevisiae» . Embo Journal . 16 (11): 3219–3232. doi : 10.1093/emboj/16.11.3219 . PMC 1169939 . PMID 9214638 .
- Cardoso C, Timsit S, Villard L, Khrestchatisky M, Fontès M, Collequaux L (апрель 1998 г.). «Специфическое взаимодействие между продуктом гена XNP/ATR-X и установленным доменом белка EZH2 человека» . Молекулярная генетика человека . 7 (4): 679–684. doi : 10.1093/hmg/7.4.679 . PMID 9499421 .
- Van Lohuizen M, Tijms M, Voncken JW, Schumacher A, Magnuson T, Wientjens E (июнь 1998 г.). «Взаимодействие белков мышиной поликомб-группы (PC-G) ENX1 и ENX2 с EED: индикация для отдельных комплексов PC-G» . Молекулярная и клеточная биология . 18 (6): 3572–3579. doi : 10.1128/mcb.18.6.3572 . PMC 108938 . PMID 9584197 .
- Sewalt RG, Van der Vlag J, Gunster MJ, Hamer KM, Den Blaauwen JL, Satijn DP, et al. (Июнь 1998 г.). «Характеристика взаимодействий между белками поликомб-группы млекопитающих ENX1/EZH2 и EED предполагает существование различных белковых комплексов поликомби-группы млекопитающих» . Молекулярная и клеточная биология . 18 (6): 3586–3595. doi : 10.1128/mcb.18.6.3586 . PMC 108940 . PMID 9584199 .
- Денизенко О., Шнирева М., Сузуки Х, Бомзтик К (октябрь 1998). «Точечные мутации в домене wd40 eed блокируют его взаимодействие с EZH2» . Молекулярная и клеточная биология . 18 (10): 5634–5642. doi : 10.1128/mcb.18.10.5634 . PMC 109149 . PMID 9742080 .
- van der Vlag J, Otte AP (декабрь 1999 г.). «Транскрипционная репрессия, опосредованная белком поликомб-группы человека, включает в себя деацетилирование гистона». Природа генетика . 23 (4): 474–478. doi : 10.1038/70602 . PMID 10581039 . S2CID 6748531 .
- Cardoso C, Mignon C, Hetet G, Grandchamps B, Fontes M, Colleaux L (март 2000 г.). «Ген EZH2 человека: геномная организация и пересмотренное картирование в 7Q35 в критической области для злокачественных миелоидных расстройств» . Европейский журнал человеческой генетики . 8 (3): 174–180. doi : 10.1038/sj.ejhg.5200439 . PMID 10780782 .
- Raaphorst FM, Otte AP, Van Kemenade FJ, Blokzijl T, Fieret E, Hamer KM, et al. (Май 2001). «Отличительные паттерны экспрессии BMI-1 и EZH2 в тимоцитах и зрелых Т-клетках предполагают роль для поликомбных генов в дифференцировке Т-клеток человека» . Журнал иммунологии . 166 (10): 5925–5934. doi : 10.4049/jimmunol.166.10.5925 . PMID 11342607 .
- О'Коннелл С., Ван Л., Роберт С., Джонс К.А., Сент Р., Джонс Р.С. (ноябрь 2001 г.). «Поликомблейковые пальцы PhD опосредуют консервативное взаимодействие с энхансером белка цеста» . Журнал биологической химии . 276 (46): 43065–43073. doi : 10.1074/jbc.m104294200 . PMID 11571280 .
- Varambally S, Dhanasekaran SM, Zhou M, Barrette TR, Kumar-Sinha C, Sanda MG, et al. (Октябрь 2002). «Polycomb Group Protein EZH2 участвует в прогрессировании рака простаты». Природа . 419 (6907): 624–629. Bibcode : 2002natur.419..624V . doi : 10.1038/nature01075 . HDL : 2027.42/62896 . PMID 12374981 . S2CID 4414767 .
- Kleer CG, Cao Q, Varambally S, Shen R, Ota I, Tomlins SA, et al. (Сентябрь 2003 г.). «EZH2 является маркером агрессивного рака молочной железы и способствует опухолевой трансформации эпителиальных клеток молочной железы» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (20): 11606–11611. Bibcode : 2003pnas..10011606K . doi : 10.1073/pnas.1933744100 . PMC 208805 . PMID 14500907 .