ДНК-вакцина

ДНК -вакцина — это тип вакцины , которая трансфицирует специфическую антиген, , кодирующую последовательность ДНК в клетки организма в качестве механизма индукции иммунного ответа. [1] [2]
ДНК-вакцины действуют путем инъекции генно-инженерной плазмиды, содержащей последовательность ДНК, кодирующую антиген (ы), против которых требуется иммунный ответ, поэтому клетки непосредственно продуцируют антиген, вызывая тем самым защитный иммунологический ответ . [3] ДНК-вакцины имеют теоретические преимущества перед обычными вакцинами, включая «способность вызывать более широкий спектр типов иммунного ответа». [4] Несколько ДНК-вакцин были протестированы для использования в ветеринарии . [3] В одних случаях удалось добиться защиты животных от болезней, в других — нет. [3] Продолжаются исследования в отношении вирусных, бактериальных и паразитарных заболеваний человека, а также рака. [4] В августе 2021 года власти Индии дали экстренное разрешение на использование ZyCoV-D . Разработанная Cadila Healthcare , это первая ДНК-вакцина, одобренная для людей. [5]
История
[ редактировать ]Обычные вакцины содержат либо специфические антигены возбудителя, либо ослабленные вирусы, которые стимулируют иммунный ответ в привитом организме. ДНК-вакцины являются членами генетических вакцин , поскольку они содержат генетическую информацию (ДНК или РНК), которая кодирует клеточное производство ( биосинтез белка ) антигена . ДНК-вакцины содержат ДНК, которая кодирует специфические антигены возбудителя. ДНК вводится в организм и поглощается клетками, нормальные метаболические процессы которых синтезируют белки на основе генетического кода плазмиды, которую они поглотили. Поскольку эти белки содержат области аминокислотных последовательностей, характерных для бактерий или вирусов, они распознаются как чужеродные, и когда они обрабатываются клетками-хозяевами и отображаются на их поверхности, иммунная система предупреждается, что затем запускает иммунные реакции. [6] [7] Альтернативно, ДНК может быть инкапсулирована в белок для облегчения проникновения в клетку. Если этот капсидный белок будет включен в ДНК, полученная вакцина сможет сочетать в себе эффективность живой вакцины без риска реверсии. [ нужна ссылка ]
В 1983 году Энцо Паолетти и Деннис Паникали из Министерства здравоохранения Нью-Йорка разработали стратегию производства вакцин на основе рекомбинантной ДНК с помощью генной инженерии для преобразования обычной вакцины против оспы в вакцины, которые могут предотвращать другие заболевания. [8] Они изменили ДНК вируса коровьей оспы , вставив ген других вирусов (а именно вируса простого герпеса , гепатита В и гриппа ). [9] [10] В 1993 году Джеффри Улмер и его коллеги из исследовательской лаборатории Merck продемонстрировали, что прямая инъекция мышам плазмидной ДНК, кодирующей антиген гриппа, защищает животных от последующего экспериментального заражения вирусом гриппа. [11] В 2016 году ДНК-вакцина против вируса Зика начала испытания на людях в Национальных институтах здравоохранения . В исследовании планировалось принять участие до 120 субъектов в возрасте от 18 до 35 лет. Отдельно Inovio Pharmaceuticals и GeneOne Life Science начали испытания другой ДНК-вакцины против вируса Зика в Майами. Вакцина NIH вводится в плечо под высоким давлением. По состоянию на август 2016 года массовое производство вакцин оставалось нерешенным. [12] Клинические испытания ДНК-вакцин для профилактики ВИЧ продолжаются. [13]
В августе 2021 года власти Индии дали экстренное разрешение на использование ZyCoV-D. , разработанная Cadila Healthcare Это первая ДНК-вакцина против COVID-19 . [5]
Приложения
[ редактировать ]По состоянию на 2021 год [update] ни одна ДНК-вакцина не была одобрена для использования человеком в Соединенных Штатах. Лишь немногие экспериментальные испытания вызвали достаточно сильную реакцию, чтобы защитить от болезней, и полезность этого метода еще предстоит доказать на людях.
Одобрена ветеринарная ДНК-вакцина для защиты лошадей от вируса Западного Нила. [14] Еще одна вакцина против вируса Западного Нила была успешно испытана на американских малиновках. [15]
ДНК-иммунизация также исследуется как средство разработки противоядных сывороток. [1] ДНК-иммунизация может использоваться в качестве технологической платформы для индукции моноклональных антител. [2]
Преимущества
[ редактировать ]- Нет риска заражения [7]
- Презентация антигена MHC класса I и класса II молекулами [7]
- Поляризовать реакцию Т-клеток в сторону типа 1 или типа 2. [7]
- Иммунный ответ сосредоточен на интересующем антигене.
- Простота разработки и производства [7]
- Стабильность при хранении и транспортировке
- Экономическая эффективность
- Устраняет необходимость синтеза пептидов, экспрессии и очистки рекомбинантных белков и использования токсичных адъювантов. [16]
- Длительная персистенция иммуногена [6]
- Экспрессия in vivo обеспечивает более близкое сходство белка с нормальной эукариотической структурой с сопутствующими посттрансляционными модификациями. [6]
Недостатки
[ редактировать ]- Ограничено белковыми иммуногенами (не применимо для небелковых антигенов, таких как бактериальные полисахариды)
- Возможность атипичной обработки белков бактерий и паразитов. [7]
- Потенциал при использовании назального спрея введения наночастиц плазмидной ДНК для трансфекции нецелевых клеток, таких как клетки головного мозга. [17]
- Перекрестное загрязнение при производстве разных типов живых вакцин на одном предприятии.
Плазмидные векторы
[ редактировать ]Векторный дизайн
[ редактировать ]ДНК-вакцины вызывают лучший иммунный ответ при использовании векторов с высокой экспрессией. Это плазмиды , которые обычно состоят из сильного вирусного промотора , который управляет транскрипцией и трансляцией гена (или комплементарной ДНК ). in vivo интересующего [18] интрон А Иногда можно включать для улучшения стабильности мРНК и, следовательно, увеличения экспрессии белка. [19] Плазмиды также содержат сильный полиаденилирования /транскрипции сигнал терминации , такой как последовательности полиаденилирования бычьего гормона роста или кроличьего бета-глобулина . [6] [7] [20] Полицистронные векторы (с несколькими представляющими интерес генами) иногда конструируют для экспрессии более чем одного иммуногена или для экспрессии иммуногена и иммуностимулирующего белка. [21]
Поскольку плазмида, несущая относительно небольшой генетический код размером примерно до 200 тыс. пар оснований, является «транспортным средством», из которого экспрессируется иммуноген, необходима оптимизация дизайна вектора для максимальной экспрессии белка. [21] Одним из способов усиления экспрессии белка является оптимизация использования кодонов патогенных мРНК в эукариотических клетках. Патогены часто имеют другое содержание АТ, чем целевые виды, поэтому изменение последовательности гена иммуногена, чтобы отразить кодоны, более часто используемые у целевых видов, может улучшить его экспрессию. [22]
Еще одним соображением является выбор промоутера . Промотор SV40 традиционно использовался до тех пор, пока исследования не показали, что векторы, управляемые промотором вируса саркомы Рауса (RSV), имеют гораздо более высокую скорость экспрессии. [6] Совсем недавно экспрессия и иммуногенность были дополнительно увеличены в модельных системах за счет использования непосредственно раннего промотора цитомегаловируса (CMV) и ретровирусного цис-действующего транскрипционного элемента . [23] Дополнительные модификации для улучшения скорости экспрессии включают вставку энхансерных последовательностей, синтетических интронов , трехсторонних лидерных последовательностей аденовируса (TPL) и модификации последовательностей полиаденилирования и терминации транскрипции. [6] Примером ДНК-вакцинной плазмиды является pVAC, в которой используется промотор SV40 .
Явления структурной нестабильности вызывают особую озабоченность при производстве плазмид, ДНК-вакцинации и генной терапии. [24] Дополнительные области, относящиеся к основной цепи плазмиды, могут участвовать в широком спектре явлений структурной нестабильности. Хорошо известные катализаторы генетической нестабильности включают прямые, инвертированные и тандемные повторы, которые заметны во многих коммерчески доступных векторах клонирования и экспрессии. Следовательно, уменьшение или полное устранение посторонних некодирующих последовательностей основной цепи могло бы заметно снизить склонность к таким событиям и, следовательно, общий рекомбиногенный потенциал плазмиды. [25]
Механизм плазмид
[ редактировать ]Как только плазмида внедряется в ядро трансфицированной клетки, она кодирует пептидную цепочку чужеродного антигена. На своей поверхности клетка отображает чужеродный антиген с молекулами комплекса гистосовместимости (MHC) классов I и II. Затем антигенпрезентирующая клетка перемещается в лимфатические узлы и представляет антигенный пептид и костимулирующую молекулу, передающую сигнал Т-клетке, инициируя иммунный ответ. [26]
Дизайн вкладыша для вакцины
[ редактировать ]Иммуногены могут быть направлены на различные клеточные компартменты для улучшения антител или цитотоксических Т-клеточных ответов. Секретируемые или связанные с плазматической мембраной антигены более эффективны в индукции гуморального ответа, чем цитозольные антигены, в то время как ответы цитотоксических Т-клеток могут быть улучшены путем нацеливания антигенов на цитоплазматическую деградацию и последующее вступление в путь главного комплекса гистосовместимости (MHC) класса I. [7] Обычно это достигается добавлением сигналов N-концевого убиквитина . [27] [28] [29]
Конформация . белка также может влиять на реакцию антител «Упорядоченные» структуры (например, вирусные частицы) более эффективны, чем неупорядоченные. [30] Цепочки минигенов (или эпитопов MHC класса I ) от различных патогенов вызывают цитотоксические реакции Т-клеток на некоторые патогены, особенно если также включен эпитоп TH. [7]
Доставка
[ редактировать ]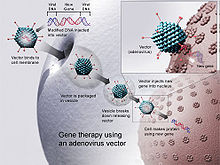
ДНК-вакцины вводили в ткани животных несколькими методами. В 1999 году двумя наиболее популярными подходами были инъекции ДНК в физиологическом растворе : с помощью стандартной иглы для подкожных инъекций или с помощью доставки генной пушки . [31] За прошедшие годы было зарегистрировано несколько других методов.
Инъекция физиологического раствора
[ редактировать ]Инъекцию физиологического раствора обычно проводят внутримышечно (в/м) в скелетные мышцы или внутрикожно (внутрикожно) с доставкой ДНК во внеклеточные пространства. Этому можно способствовать либо 1) электропорацией ; [32] 2) путем временного повреждения мышечных волокон миотоксинами , такими как бупивакаин ; или 3) с помощью гипертонических растворов физиологического раствора или сахарозы . [6] На иммунную реакцию на этот метод могут влиять такие факторы, как тип иглы, [16] выравнивание иглы, скорость инъекции, объем инъекции, тип мышц, а также возраст, пол и физиологическое состояние реципиента. [6]
Генный пистолет
[ редактировать ]Доставка генной пушки баллистически ускоряет плазмидную ДНК (пДНК), которая была поглощена микрочастицами золота или вольфрама , в клетки-мишени, используя сжатый гелий в качестве ускорителя. [6] [21]
Доставка на поверхность слизистой оболочки
[ редактировать ]Альтернативы включали аэрозольную инстилляцию обнаженной ДНК на поверхности слизистых оболочек , таких как носа и легких слизистая оболочка . [21] и местное введение пДНК в глаз [33] и слизистой влагалища. [21] Доставка на поверхность слизистой оболочки также была достигнута с использованием катионных липосомальных препаратов ДНК. [7] биоразлагаемые микросферы, [34] [21] ослабленная сальмоналла , [35] Векторы шигелл или листерий для перорального введения в слизистую оболочку кишечника. [36] и рекомбинантные аденовирусные векторы. [21]
Полимерный автомобиль
[ редактировать ]гибридный носитель, состоящий из бактериальных клеток и синтетических полимеров Для доставки ДНК-вакцины был использован . Внутреннее ядро E. coli и внешняя оболочка из поли(бета-аминоэфира) действуют синергически, повышая эффективность путем устранения барьеров, связанных с доставкой генов в антиген-представляющие клетки , которые включают клеточное поглощение и интернализацию, выход из фагосом и внутриклеточную концентрацию груза. [ жаргон ] При испытаниях на мышах было обнаружено, что гибридный вектор вызывает иммунный ответ. [37] [38]
иммунизация ЭЛИ
[ редактировать ]Другим подходом к ДНК-вакцинации является иммунизация экспрессионной библиотекой (ELI). Используя этот метод, потенциально все гены патогена могут быть доставлены одновременно, что может быть полезно для патогенов, которые трудно ослабить или культивировать. [6] ELI можно использовать для определения того, какие гены вызывают защитную реакцию. Это было проверено на Mycoplasma pulmonis , мышином легочном патогене с относительно небольшим геномом . Даже библиотеки частичной экспрессии могут обеспечить защиту от последующего заражения. [39]
Полезное табличное сравнение
[ редактировать ]| Способ доставки | Формулировка ДНК | Целевая ткань | Количество ДНК | |
|---|---|---|---|---|
| Парентеральный | Инъекция (игла для подкожных инъекций) | Водный раствор в физиологическом растворе | ИМ (скелетный); ИДЕНТИФИКАТОР ; ( ВВ , подкожно и внутрибрюшинно с переменным успехом) | Большие количества (приблизительно 100-200 мкг) |
| Генный пистолет | Золотые бусины, покрытые ДНК | ЭД (кожа живота); слизистая влагалища; хирургически обнаженные мышцы и другие органы | Небольшие количества (всего 16 нг) | |
| Пневматический (струйный) впрыск | Водный раствор | ЭД | Очень высокая (до 300 мкг) | |
| Местное применение | Водный раствор | окуляр; интравагинальный | Небольшие количества (до 100 мкг) | |
| Цитофектин-опосредованный [ жаргон ] | Липосомы (катионные); микросферы; рекомбинантные аденовирусные векторы; аттенуированный вектор шигелл ; аэрозольные катионных липидов составы | Я; IV (для системной трансфекции тканей); внутрибрюшинный; пероральная иммунизация слизистой оболочки кишечника; слизистые оболочки носа/легких | переменная | |
| Способ доставки | Преимущество | Недостаток |
|---|---|---|
| Внутримышечная или внутрикожная инъекция |
|
|
| Генный пистолет |
|
|
| Струйный впрыск |
|
|
| Липосомальная доставка |
|
|
Дозировка
[ редактировать ]Способ доставки определяет дозу, необходимую для повышения эффективного иммунного ответа. Для инъекций физиологического раствора требуется разное количество ДНК: от 10 мкг до 1 мг, тогда как для доставки генной пушкой требуется в 100–1000 раз меньше. [40] Обычно требуется 0,2–20 мкг, хотя сообщалось о таких низких количествах, как 16 нг. [6] Эти количества варьируются в зависимости от вида. Например, мышам требуется примерно в 10 раз меньше ДНК, чем приматам . [7] Для инъекций физиологического раствора требуется больше ДНК, поскольку ДНК доставляется во внеклеточные пространства ткани-мишени (обычно мышц), где ей приходится преодолевать физические барьеры (такие как базальная пластинка и большое количество соединительной ткани ), прежде чем она будет поглощена клетки, в то время как доставка генной пушки направляет/нагнетает ДНК непосредственно в клетки, что приводит к меньшим «потери». [6] [7]
Иммунный ответ
[ редактировать ]Ответы Т-хелперов
[ редактировать ]
Иммунизация ДНК может вызывать множественные реакции Т H , включая лимфопролиферацию и образование различных профилей цитокинов . Основным преимуществом ДНК-вакцин является легкость, с которой ими можно манипулировать, чтобы сместить тип помощи Т-клеток в сторону ответа TH1 или TH2. [41] Каждый тип имеет отличительные закономерности экспрессии лимфокинов и хемокинов, специфические типы иммуноглобулинов , закономерности перемещения лимфоцитов и типы врожденных иммунных ответов .
Другие виды помощи Т-клеток
[ редактировать ]На тип получаемой помощи Т-клеток влияет способ доставки и тип экспрессируемого иммуногена, а также нацеливание на различные лимфоидные компартменты. [6] [42] Как правило, инъекции физиологического раствора иглой (в/м или внутримышечно) имеют тенденцию индуцировать реакцию TH1, тогда как доставка генной пушки повышает реакцию TH2. [41] [42] Это справедливо для внутриклеточных и связанных с плазматической мембраной антигенов, но не для секретируемых антигенов, которые, по-видимому, вызывают ответы TH2 независимо от метода доставки. [43]
Как правило, тип вырабатываемой помощи Т-клеток стабилен с течением времени и не меняется при заражении или после последующих иммунизаций, которые обычно вызывали бы противоположный тип ответа у наивного образца. [41] [42] Однако Мор и др. . (1995) [18] иммунизировали и иммунизировали мышей пДНК, кодирующей циркумспорозоитный белок мышиного малярийного паразита Plasmodium yoelii (PyCSP), и обнаружили, что первоначальный ответ TH2 после ревакцинации изменился на ответ TH1.
Основа для различных типов помощи Т-клеток
[ редактировать ]Как действуют эти различные методы, формы экспрессируемого антигена и различные профили помощи Т-клеток, непонятно. Считалось, что относительно большие количества ДНК, использованные при внутримышечной инъекции, ответственны за индукцию ответов TH1. Однако данные не показывают дозозависимых различий в типе ТГ. [41] Тип вырабатываемой Т-клеточной помощи определяется дифференцированным состоянием антигенпредставляющих клеток . Дендритные клетки могут дифференцироваться и секретировать IL-12 (который поддерживает развитие клеток TH1) или IL-4 (который поддерживает ответы TH2). [44] пДНК, инъецированная иглой, эндоцитируется в дендритную клетку, которая затем стимулируется к дифференцировке для производства цитокина TH1 (IL-12). [45] в то время как генная пушка бомбардирует ДНК непосредственно в клетку, минуя стимуляцию TH1.
Практическое использование поляризованных Т-клеток.
[ редактировать ]Поляризация в Т-клетках помогает влиять на аллергические реакции и аутоиммунные заболевания . При аутоиммунных заболеваниях цель состоит в том, чтобы сместить саморазрушительный ответ TH1 (с связанной с ним цитотоксической активностью Т-клеток) на неразрушающий ответ TH2. Это было успешно применено при предзаболевании для желаемого типа ответа в доклинических моделях. [7] и в некоторой степени успешно меняет реакцию на уже установленное заболевание. [46]
Цитотоксические Т-клеточные реакции
[ редактировать ]Одним из преимуществ ДНК-вакцин является то, что они способны индуцировать цитотоксические Т-лимфоциты (CTL) без риска, присущего живым вакцинам. Реакция CTL может быть вызвана против иммунодоминантных и иммунорецессивных эпитопов CTL. [47] а также субдоминантные эпитопы CTL , [34] [ жаргон ] таким образом, который имитирует естественную инфекцию . Это может оказаться полезным инструментом для оценки эпитопов CTL и их роли в обеспечении иммунитета.
Цитотоксические Т-клетки распознают небольшие пептиды (8-10 аминокислот ), образующие комплекс с MHC класса I. молекулами [48] Эти пептиды происходят из цитозольных белков, которые расщепляются и доставляются к зарождающейся молекуле MHC класса I в эндоплазматическом ретикулуме (ЭР). [48] Таким образом, нацеливание генных продуктов непосредственно на ER (путем добавления сигнальной последовательности вставки ER на N-конце ) должно усиливать ответы CTL. Это было успешно продемонстрировано с использованием рекомбинантных коровьей оспы вирусов гриппа . , экспрессирующих белки [48] однако этот принцип должен быть применим и к ДНК-вакцинам. Было показано, что нацеливание антигенов на внутриклеточную деградацию (и, таким образом, на вход в путь MHC класса I) путем добавления убиквитина сигнальных последовательностей или мутации других сигнальных последовательностей эффективно увеличивает ответы CTL. [28]
Ответы CTL можно усилить путем совместной инокуляции костимулирующими молекулами, такими как B7-1 или B7-2, для ДНК-вакцин против нуклеопротеина гриппа, [47] [49] или GM-CSF для ДНК-вакцин против мышиной модели малярии P. yoelii . [50] Было показано, что совместная инокуляция плазмидами, кодирующими костимулирующие молекулы IL-12 и TCA3, увеличивает активность CTL против антигенов нуклеопротеина ВИЧ-1 и гриппа. [49] [51]
Гуморальный (антитетельный) ответ
[ редактировать ]
На реакцию антител, вызванную вакцинацией ДНК, влияют множество переменных, включая тип антигена; расположение антигена (т.е. внутриклеточное или секретируемое); количество, частота и доза иммунизации; место и способ доставки антигена.
Кинетика реакции антител
[ редактировать ]Гуморальные реакции после однократной инъекции ДНК могут быть гораздо более продолжительными, чем после однократной инъекции рекомбинантного белка. Реакция антител против оболочечного белка вируса гепатита B (HBV) (HBsAg) сохранялась до 74 недель без ревакцинации, в то время как пожизненное сохранение защитного ответа на гемагглютинин гриппа было продемонстрировано у мышей после доставки генной пушки. [52] Клетки, секретирующие антитела (ASC), мигрируют в костный мозг и селезенку для длительного производства антител и обычно локализуются там через год. [52]
Сравнение ответов антител, вызванных естественной (вирусной) инфекцией, иммунизацией рекомбинантным белком и иммунизацией пДНК, суммировано в таблице 4. Ответы антител, вызванные ДНК, растут гораздо медленнее, чем при естественной инфекции или иммунизации рекомбинантным белком. Для достижения пиковых титров у мышей может потребоваться до 12 недель, хотя ревакцинация может сократить этот интервал. Этот ответ, вероятно, обусловлен низкими уровнями антигена, экспрессируемого в течение нескольких недель, что поддерживает как первичную, так и вторичную фазы гуморального ответа. [ нужны разъяснения ] ДНК-вакцина, экспрессирующая белок малой и средней оболочки HBV, вводилась взрослым с хроническим гепатитом. Вакцина привела к выработке специфического интерферона гамма-клетками. Также были разработаны специфические Т-клетки для антигенов белков средней оболочки. Иммунный ответ пациентов был недостаточно сильным, чтобы контролировать инфекцию HBV. [53]
| Способ иммунизации | |||
|---|---|---|---|
| ДНК-вакцина | Рекомбинантный белок | Естественная инфекция | |
| Количество индуцирующего антигена | из | мкг | ? (в мкг) |
| Продолжительность презентации антигена | несколько недель | < 1 неделя | несколько недель |
| Кинетика реакции антител | медленный подъем | быстрый рост | быстрый рост |
| Количество прививок для получения высокой авидности IgG и миграции АСК в костный мозг | один | два | один |
| Изотип Ab (мышиные модели) | С'-зависимый или С'-независимый | C'-зависимый | C'-независимый |
Кроме того, титры специфических антител, вырабатываемые при вакцинации ДНК, ниже, чем титры, полученные после вакцинации рекомбинантным белком. Однако антитела, индуцированные ДНК-иммунизацией, проявляют большее сродство к нативным эпитопам, чем антитела, индуцированные рекомбинантным белком. Другими словами, иммунизация ДНК вызывает качественно лучший ответ. Антитела могут быть индуцированы после одной вакцинации ДНК, тогда как вакцинация рекомбинантными белками обычно требует повторной вакцинации. Иммунизация ДНК может использоваться для смещения ТН-профиля иммунного ответа и, следовательно, изотипа антитела, что невозможно ни при естественной инфекции, ни при иммунизации рекомбинантным белком. Ответы антител, генерируемые ДНК, полезны в качестве препаративного инструмента. Например, можно получить поликлональные и моноклональные антитела для использования в качестве реагентов. [ нужна ссылка ]
Механистическая основа иммунных реакций, вызванных ДНК
[ редактировать ]Механизм поглощения ДНК
[ редактировать ]Когда поглощение и последующая экспрессия ДНК была впервые продемонстрирована in vivo в мышечных клетках, [54] эти клетки считались уникальными из-за их обширной сети Т-трубочек. С помощью электронной микроскопии было высказано предположение, что поглощению ДНК способствуют кавеолы (или ямки, не покрытые клатрином). [55] Однако последующие исследования показали, что другие клетки (такие как кератиноциты , фибробласты и эпителиальные клетки Лангерганса ) также могут усваивать ДНК. [46] [56] Механизм поглощения ДНК неизвестен.
Доминируют две теории: поглощение ДНК in vivo происходит неспецифически, методом, подобным фаго- или пиноцитозу . [21] или через специфические рецепторы. [57] Они могут включать поверхностные рецепторы массой 30 кДа или рецепторы-поглотители макрофагов . [ нужны разъяснения ] Поверхностный рецептор массой 30 кДа специфически связывается с фрагментами ДНК длиной 4500 пар оснований (которые затем интернализуются) и обнаруживается на профессиональных АПК и Т-клетках. Макрофаговые скэвенджер-рецепторы связываются с различными макромолекулами, включая полирибонуклеотиды , и, таким образом, являются кандидатами на поглощение ДНК. [57] [58] Рецепторно-опосредованное поглощение ДНК может быть облегчено наличием полигуанилатных последовательностей . [ нужны разъяснения ] [ нужна ссылка ] генных пушек Системы доставки , упаковка катионных липосом и другие методы доставки обходят этот метод проникновения, но понимание его может быть полезно для снижения затрат (например, за счет снижения потребности в цитофектинах), что может быть важно в животноводстве.
Презентация антигена клетками, происходящими из костного мозга.
[ редактировать ]
Исследования с использованием химерных мышей показали, что антиген представлен клетками, происходящими из костного мозга, которые включают дендритные клетки, макрофаги и специализированные B-клетки, называемые профессиональными антигенпрезентирующими клетками (APC). [49] [59] После инокуляции генной пушкой на кожу трансфицированные клетки Лангерганса мигрируют в дренирующий лимфатический узел и представляют антигены. [7] После внутримышечных и внутрикожных инъекций дендритные клетки представляют антиген в дренирующем лимфатическом узле. [56] трансфицированные макрофаги были обнаружены в периферической крови. [60]
Помимо прямой трансфекции дендритных клеток или макрофагов, перекрестное праймирование происходит после доставки ДНК IM, ID и генной пушки. Перекрестное праймирование происходит, когда клетка, происходящая из костного мозга, представляет пептиды из белков, синтезированных в другой клетке в контексте MHC класса 1. Это может стимулировать цитотоксические Т-клеточные ответы и, по-видимому, важно для полного первичного иммунного ответа. [7] [61]
Роль целевого сайта
[ редактировать ]Доставка ДНК IM и ID по-разному инициирует иммунные ответы. В коже кератиноциты, фибробласты и клетки Лангерганса поглощают и экспрессируют антигены и отвечают за индукцию первичного гуморального ответа. Трансфицированные клетки Лангерганса мигрируют из кожи (в течение 12 часов) в дренирующий лимфатический узел, где они запускают вторичные В- и Т-клеточные реакции. В скелетных мышцах чаще всего трансфицируются поперечно-полосатые мышечные клетки, но они, по-видимому, не играют важной роли в иммунном ответе. Вместо этого ДНК, инокулированная IM, в течение нескольких минут «вымывается» в дренирующий лимфатический узел, где дистальные дендритные клетки трансфицируются, а затем инициируют иммунный ответ. Трансфицированные миоциты, по-видимому, действуют как «резервуар» антигена для торговли профессиональными АПК. [21] [54] [61]
Поддержание иммунного ответа
[ редактировать ]ДНК-вакцинация создает эффективную иммунную память посредством отображения комплексов антиген-антитело на фолликулярных дендритных клетках (ФДК), которые являются мощными стимуляторами В-клеток. Т-клетки могут стимулироваться аналогичными дендритными клетками зародышевого центра. FDC способны генерировать иммунную память, поскольку выработка антител «перекрывает» долговременную экспрессию антигена, позволяя формироваться и проявляться иммунокомплексам антиген-антитело с помощью FDC. [7]
Интерфероны
[ редактировать ]И хелперные, и цитотоксические Т-клетки могут контролировать вирусные инфекции, секретируя интерфероны. Цитотоксические Т-клетки обычно убивают инфицированные вирусом клетки. Однако их также можно стимулировать к секреции противовирусных цитокинов, таких как IFN-γ и TNF-α , которые не убивают клетку, но ограничивают вирусную инфекцию за счет снижения экспрессии вирусных компонентов. [62] ДНК-вакцинации могут использоваться для сдерживания вирусных инфекций путем неразрушающего контроля, опосредованного IFN. Это было продемонстрировано для гепатита B. [63] IFN-γ имеет решающее значение в борьбе с малярийными инфекциями. [64] и это необходимо учитывать при разработке противомалярийных ДНК-вакцин.
Модуляция иммунного ответа
[ редактировать ]Цитокиновая модуляция
[ редактировать ]Эффективная вакцина должна вызывать соответствующий иммунный ответ на данный патоген. ДНК-вакцины могут поляризовать помощь Т-клеток в сторону профилей TH1 или TH2 и при необходимости генерировать CTL и/или антитела. Этого можно достичь путем модификации формы экспрессируемого антигена (т.е. внутриклеточного или секретируемого), способа и пути доставки или дозы. [41] [42] [65] [66] [67] Это также может быть достигнуто путем совместного введения плазмидной ДНК, кодирующей иммунорегуляторные молекулы, т.е. цитокины, лимфокины или костимулирующие молекулы. Эти «генетические адъюванты » можно применять в виде:
- смесь двух плазмид, одна из которых кодирует иммуноген, а другая - цитокин.
- одиночный би- или полицистронный вектор, разделенный спейсерными областями
- , кодируемая плазмидой химера , или гибридный белок
В целом, одновременное введение провоспалительных агентов (таких как различные интерлейкины , фактор некроза опухоли и GM-CSF) и TH2-индуцирующих цитокинов увеличивает реакцию антител, тогда как провоспалительные агенты и TH1-индуцирующие цитокины снижают гуморальные реакции и повышают цитотоксические реакции (более важные для защиты от вирусов). костимулирующие молекулы, такие как B7-1 , B7-2 и CD40L Иногда используются .
Эта концепция была применена при местном введении пДНК, кодирующей IL-10 . [33] Плазмида, кодирующая B7-1 (лиганд APC), успешно усиливала иммунный ответ на моделях опухолей. Смешение плазмид, кодирующих GM-CSF и циркумспорозоитный белок P. yoelii (PyCSP), усиливало защиту от последующего заражения (тогда как только PyCSP, кодируемый плазмидой, этого не делало). Было высказано предположение, что GM-CSF заставляет дендритные клетки более эффективно представлять антиген и усиливать выработку IL-2 и активацию TH-клеток, тем самым вызывая усиление иммунного ответа. [50] Это можно еще больше усилить путем первого праймирования смесью pPyCSP и pGM-CSF с последующей бустерной стимуляцией рекомбинантным поксвирусом, экспрессирующим PyCSP. [68] Однако совместная инъекция плазмид, кодирующих GM-CSF (или IFN-γ, или IL-2) и слитого белка P. chabaudi поверхностного белка 1 мерозоита (C-конец)-поверхностного белка вируса гепатита B (PcMSP1-HBs) упразднена защита от заражения по сравнению с защитой, полученной путем доставки только pPcMSP1-HB. [30]
Преимуществами генетических адъювантов являются их низкая стоимость и простота применения, а также отсутствие нестабильных рекомбинантных цитокинов и потенциально токсичных «обычных» адъювантов (таких как квасцы , фосфат кальция , монофосфориллипид А, холерный токсин, катионные липосомы и липосомы, покрытые маннаном). , QS21 , карбоксиметилцеллюлоза и убенимекс ). [7] [21] Однако потенциальная токсичность продолжительной экспрессии цитокинов не установлена. У многих коммерчески важных видов животных гены цитокинов не идентифицированы и не изолированы. Кроме того, различные цитокины, кодируемые плазмидами, по-разному модулируют иммунную систему в зависимости от времени доставки. Например, некоторые цитокиновые плазмидные ДНК лучше всего доставляются после иммуногенной пДНК, поскольку предварительная или совместная доставка может снизить специфические ответы и повысить неспецифические ответы. [69]
Иммуностимулирующие мотивы CpG
[ редактировать ]Плазмидная ДНК сама по себе оказывает адъювантное действие на иммунную систему. [6] [7] ДНК бактериального происхождения может запускать механизмы врожденной иммунной защиты, активацию дендритных клеток и выработку цитокинов TH1. [45] [70] Это связано с распознаванием определенных динуклеотидных последовательностей CpG, обладающих иммуностимулирующим действием. [66] [71] Стимулирующие CpG (CpG-S) последовательности встречаются в двадцать раз чаще в ДНК бактериального происхождения, чем у эукариот. Это связано с тем, что эукариоты демонстрируют «супрессию CpG», то есть пары динуклеотидов CpG встречаются гораздо реже, чем ожидалось. Кроме того, последовательности CpG-S гипометилированы. Это часто происходит в бактериальной ДНК, тогда как мотивы CpG, встречающиеся у эукариот, метилированы по цитозиновому нуклеотиду. Напротив, нуклеотидные последовательности, которые ингибируют активацию иммунного ответа (так называемые нейтрализующие CpG или CpG-N), чрезмерно представлены в геномах эукариот. [72] Оптимальной иммуностимулирующей последовательностью является неметилированный динуклеотид CpG, фланкированный двумя 5'- пуринами и двумя 3'- пиримидинами . [66] [70] Кроме того, фланкирующие области за пределами этого иммуностимулирующего гексамера должны быть богаты гуанином , чтобы обеспечить связывание и поглощение клетками-мишенями.
Врожденная система работает вместе с адаптивной иммунной системой, чтобы вызвать реакцию против белка, кодируемого ДНК. Последовательности CpG-S индуцируют поликлональную активацию В-клеток и усиление экспрессии и секреции цитокинов. [73] Стимулированные макрофаги секретируют IL-12, IL-18 , TNF-α, IFN-α, IFN-β и IFN-γ, тогда как стимулированные B-клетки секретируют IL-6 и некоторое количество IL-12. [21] [73] [74]
Манипулирование последовательностями CpG-S и CpG-N в плазмидном остове ДНК-вакцин может обеспечить успех иммунного ответа на кодируемый антиген и привести иммунный ответ к фенотипу TH1. Это полезно, если патогену требуется реакция TH для защиты. Последовательности CpG-S также использовались в качестве внешних адъювантов для вакцинации как ДНК, так и рекомбинантным белком с различной степенью успеха. Другие организмы с гипометилированными мотивами CpG продемонстрировали стимуляцию поликлональной экспансии B-клеток. [75] Механизм этого может быть более сложным, чем простое метилирование: не обнаружено, что гипометилированная мышиная ДНК вызывает иммунный ответ.
Большая часть доказательств наличия иммуностимулирующих последовательностей CpG получена в исследованиях на мышах. Экстраполяция этих данных на другие виды требует осторожности: отдельным видам могут потребоваться разные фланкирующие последовательности, поскольку специфичность связывания рецепторов-мусорщиков варьируется у разных видов. Кроме того, такие виды, как жвачные животные, могут быть нечувствительны к иммуностимулирующим последовательностям из-за большой нагрузки на желудочно-кишечный тракт.
Альтернативные усиления
[ редактировать ]Иммунные реакции, основанные на ДНК, можно усилить введением рекомбинантного белка или рекомбинантных поксвирусов. Стратегии «прайм-буст» с использованием рекомбинантного белка успешно увеличили как титр нейтрализующих антител, так и авидность и устойчивость антител к слабым иммуногенам, таким как белок оболочки ВИЧ-1. [7] [76] Было показано, что стимуляция рекомбинантными вирусами очень эффективна для усиления реакций CTL, основанных на ДНК. Праймирование ДНК фокусирует иммунный ответ на необходимом иммуногене, в то время как бустирование рекомбинантным вирусом обеспечивает большее количество экспрессируемого антигена, что приводит к значительному увеличению специфических ответов CTL.
В ряде исследований стратегии Prime-boost оказались успешными в обеспечении защиты от малярии. Мыши, которым примировали плазмидную ДНК, кодирующую поверхностный белок циркумспорозоита Plasmodium yoelii (PyCSP), а затем бустерировали рекомбинантным вирусом коровьей оспы, экспрессирующим тот же белок, имели значительно более высокие уровни антител, активности CTL и IFN-γ и, следовательно, более высокие уровни защиты, чем иммунизированные мыши. и бустировали только плазмидной ДНК. [77] Это можно дополнительно усилить путем праймирования смесью плазмид, кодирующих PyCSP и мышиный GM-CSF, перед бустерной стимуляцией рекомбинантным вирусом коровьей оспы. [68] эффективная стратегия первичной стимуляции для обезьяньей модели малярии P. Knowlesi . Также была продемонстрирована [78] Макакам-резусам вводили многокомпонентную многоэтапную ДНК-вакцину, кодирующую два антигена на стадии печени – поверхностный белок циркумспорозоитов (PkCSP) и поверхностный белок 2 спорозоитов (PkSSP2) – и два антигена на стадии крови – поверхностный белок 1 апикального мерозоита (PkAMA1) и Поверхностный белок 1 мерозоита (PkMSP1p42). Затем им вводили рекомбинантный вирус оспы канареек, кодирующий все четыре антигена (ALVAC-4). У иммунизированных обезьян вырабатывались антитела против спорозоитов и инфицированных эритроцитов, а также Т-клеточные ответы, секретирующие IFN-γ, против пептидов PkCSP. Была достигнута частичная защита от заражения спорозоитами, а средняя паразитемия значительно снизилась по сравнению с контрольными обезьянами. Эти модели, хотя и не идеальны для экстраполяции на P. falciparum у людей, будут важны в доклинических испытаниях.
Усиление иммунных реакций
[ редактировать ]ДНК
[ редактировать ]Эффективность иммунизации ДНК можно повысить за счет стабилизации ДНК от деградации и повышения эффективности доставки ДНК в антигенпрезентирующие клетки . [7] Это было продемонстрировано путем покрытия биоразлагаемых катионных микрочастиц (таких как поли(лактид-со-гликолид), приготовленных с бромидом цетилтриметиламмония ) ДНК. Такие микрочастицы, покрытые ДНК, могут быть столь же эффективны в повышении CTL, как и рекомбинантные вирусы, особенно при смешивании с квасцами. Частицы диаметром 300 нм, по-видимому, наиболее эффективны для поглощения антигенпрезентирующими клетками. [7]
Альфавирусные векторы
[ редактировать ]на основе рекомбинантных альфавирусов Векторы использовались для повышения эффективности вакцинации ДНК. [7] Ген, кодирующий интересующий антиген, вставляется в репликон альфавируса, заменяя структурные гены, но оставляя неструктурные гены репликазы нетронутыми. Вирус Синдбис и вирус леса Семлики использовались для создания рекомбинантного альфавируса репликонов . В отличие от обычных ДНК-вакцинаций, альфавирусные векторы убивают трансфицированные клетки и экспрессируются лишь временно. Гены репликазы альфавируса экспрессируются помимо вакцинной вставки. Неясно, как репликоны альфавируса повышают иммунный ответ, но это может быть связано с высокими уровнями белка, экспрессируемыми этим вектором, репликон-индуцированными цитокиновыми реакциями или репликон-индуцированным апоптозом, приводящим к усиленному поглощению антигена дендритными клетками.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Энрике Роман Рамос и Пауло Ли Хо. «Разработка сыворотки против змеиного яда путем генетической иммунизации: обзор» . Клиническая токсинология в Азиатско-Тихоокеанском регионе и Африке . 2 : 401–414.
- ^ Перейти обратно: а б Лю С., Ван С., Лу С. (апрель 2016 г.). «ДНК-иммунизация как технологическая платформа для индукции моноклональных антител» . Новые микробы и инфекции . 5 (4): е33. дои : 10.1038/emi.2016.27 . ПМЦ 4855071 . ПМИД 27048742 .
- ^ Перейти обратно: а б Хан К.Х. (март 2013 г.). «ДНК-вакцины: роль в борьбе с болезнями» . Микробы . 3 (1): 26–35. дои : 10.11599/germs.2013.1034 . ПМЦ 3882840 . ПМИД 24432284 .
- ^ Перейти обратно: а б «Индия дает экстренное одобрение на первую в мире ДНК-вакцину против COVID-19» . Рейтер . 20 августа 2021 г. Проверено 22 августа 2021 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Аларкон Дж.Б., Уэйн Г.В., Макманус Д.П. (1999). «ДНК-вакцины: технология и применение в качестве противопаразитарных и противомикробных средств». Достижения паразитологии Том 42 . Том. 42. С. 343–410. дои : 10.1016/S0065-308X(08)60152-9 . ISBN 9780120317424 . ПМИД 10050276 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v Робинсон Х.Л., Пертмер Т.М. (2000). ДНК-вакцины против вирусных инфекций: фундаментальные исследования и применение . Достижения в области исследования вирусов. Том. 55. стр. 1–74. дои : 10.1016/S0065-3527(00)55001-5 . ISBN 9780120398553 . ПМИД 11050940 .
- ^ Уайт Л.О., Гибб Э., Ньюхэм Х.К., Ричардсон М.Д., Уоррен Р.К. (июль 1979 г.). «Сравнение роста вирулентных и ослабленных штаммов Candida albicans в почках нормальных и обработанных кортизоном мышей с помощью анализа хитина». Микопатология . 67 (3): 173–177. дои : 10.1007/bf00470753 . ПМИД 384256 . S2CID 31914107 .
- ^ Паолетти Э., Липинскас Б.Р., Самсонофф С., Мерсер С., Паникали Д. (январь 1984 г.). «Создание живых вакцин с использованием генно-инженерных поксвирусов: биологическая активность рекомбинантов вируса коровьей оспы, экспрессирующих поверхностный антиген вируса гепатита В и гликопротеин D вируса простого герпеса» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (1): 193–197. Бибкод : 1984PNAS...81..193P . дои : 10.1073/pnas.81.1.193 . ПМЦ 344637 . ПМИД 6320164 .
- ^ Патент США 4722848 - Способ иммунизации животных синтетически модифицированным вирусом коровьей оспы.
- ^ Улмер Дж.Б., Доннелли Дж.Дж., Паркер С.Е., Роудс Г.Х., Фельгнер П.Л., Дварки В.Дж. и др. (март 1993 г.). «Гетерологичная защита от гриппа путем инъекции ДНК, кодирующей вирусный белок». Наука . 259 (5102): 1745–1749. Бибкод : 1993Sci...259.1745U . дои : 10.1126/science.8456302 . ПМИД 8456302 .
- ^ Регаладо А (2 августа 2016 г.). «Правительство США начало испытания своей первой вакцины против Зика на людях» . Журнал MIT Technology Review . Проверено 06 августа 2016 г.
- ^ Чен Ю, Ван С, Лу С (февраль 2014 г.). «ДНК-иммунизация для разработки вакцины против ВИЧ» . Вакцина . 2 (1): 138–159. doi : 10.3390/vaccines2010138 . ПМК 4494200 . ПМИД 26344472 .
- ^ Леджервуд Дж.Э. , Пирсон Т.К., Хубка С.А., Десаи Н., Ракер С., Гордон И.Дж. и др. (май 2011 г.). «ДНК-вакцина против вируса Западного Нила, использующая модифицированный промотор, индуцирует нейтрализующие антитела у молодых и пожилых здоровых взрослых в ходе I фазы клинических испытаний» . Журнал инфекционных болезней . 203 (10): 1396–1404. дои : 10.1093/infdis/jir054 . ПМК 3080891 . ПМИД 21398392 .
- ^ Килпатрик А.М., Дюпюи А.П., Чанг Г.Дж., Крамер Л.Д. (май 2010 г.). «ДНК-вакцинация американской малиновки (Turdus migratorius) против вируса Западного Нила» . Трансмиссивные и зоонозные болезни . 10 (4): 377–380. дои : 10.1089/vbz.2009.0029 . ПМЦ 2883478 . ПМИД 19874192 .
- ^ Перейти обратно: а б Седега М., Хедстром Р., Хобарт П., Хоффман С.Л. (октябрь 1994 г.). «Защита от малярии путем иммунизации плазмидной ДНК, кодирующей белок циркумспорозоита» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (21): 9866–9870. Бибкод : 1994PNAS...91.9866S . дои : 10.1073/pnas.91.21.9866 . JSTOR 2365723 . ПМК 44918 . ПМИД 7937907 .
- ^ Хармон Б.Т., Али А.Е., Падегимас Л., Сесеноглу-Лэрд О., Купер М.Дж., Ващак Б.Л. (май 2014 г.). «Интраназальное введение наночастиц плазмидной ДНК приводит к успешной трансфекции и экспрессии репортерного белка в мозге крысы» . Генная терапия . 21 (5): 514–521. дои : 10.1038/gt.2014.28 . ПМИД 24670994 . S2CID 5560134 .
- ^ Перейти обратно: а б Мор Г., Клинман Д.М., Шапиро С., Хагивара Э., Седега М., Норман Дж.А. и др. (август 1995 г.). «Сложность реакции цитокинов и антител, вызванная иммунизацией мышей плазмидной ДНК белка циркумспорозоита Plasmodium yoelii» . Журнал иммунологии . 155 (4): 2039–2046. doi : 10.4049/jimmunol.155.4.2039 . ПМИД 7636255 . S2CID 37290980 .
- ^ Лейтнер В.В., Сеген М.К., Баллоу В.Р., Зейтц Дж.П., Шульц А.М., Шихи М.Дж., Лион Дж.А. (декабрь 1997 г.). «Иммунные реакции, вызванные внутримышечной инъекцией или инъекцией генной пушки защитных вакцин дезоксирибонуклеиновой кислоты, которые экспрессируют белок циркумспорозоита малярийных паразитов Plasmodium berghei» . Журнал иммунологии . 159 (12): 6112–6119. дои : 10.4049/jimmunol.159.12.6112 . ПМИД 9550412 . S2CID 37685499 .
- ^ Бём В., Кухрёбер А., Пайер Т., Мертенс Т., Рейманн Дж., Ширмбек Р. (июнь 1996 г.). «Векторные конструкции ДНК, которые стимулируют реакцию цитотоксических Т-лимфоцитов, специфичных для поверхностного антигена гепатита B, и антител у мышей после внутримышечной инъекции». Журнал иммунологических методов . 193 (1): 29–40. дои : 10.1016/0022-1759(96)00035-X . ПМИД 8690928 .
- ^ Перейти обратно: а б с д и ж г час я дж к Льюис П.Дж., Бабюк Л.А. (1999). ДНК-вакцины: обзор . Том. 54. Академическая пресса. стр. 129–88. дои : 10.1016/S0065-3527(08)60367-X . ISBN 978-0-12-039854-6 . ПМИД 10547676 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Андре С., Сид Б, Эберле Дж., Шраут В., Бюльтманн А., Хаас Дж. (февраль 1998 г.). «Усиление иммунного ответа, вызванное вакцинацией ДНК синтетической последовательностью gp120 с оптимизированным использованием кодонов» . Журнал вирусологии . 72 (2): 1497–1503. doi : 10.1128/JVI.72.2.1497-1503.1998 . ПМЦ 124631 . ПМИД 9445053 .
- ^ Мутхумани К., Чжан Д., Дайес Н.С., Хван Д.С., Каларота С.А., Чу А.Ю. и др. (сентябрь 2003 г.). «Новая сконструированная плазмидная конструкция gp160 восточноафриканской клады-A ВИЧ-1 индуцирует сильные гуморальные и клеточно-опосредованные иммунные реакции in vivo» . Вирусология . 314 (1): 134–146. дои : 10.1016/S0042-6822(03)00459-8 . ПМИД 14517067 .
- ^ Оливейра П.Х., Пратер К.Дж., Празерес Д.М., Монтейро Г.А. (сентябрь 2009 г.). «Структурная нестабильность плазмидных биофармацевтических препаратов: проблемы и последствия». Тенденции в биотехнологии . 27 (9): 503–511. дои : 10.1016/j.tibtech.2009.06.004 . ПМИД 19656584 .
- ^ Оливейра П.Х., Майрхофер Дж. (сентябрь 2013 г.). «Безмаркерные плазмиды для биотехнологических применений - значение и перспективы». Тенденции в биотехнологии . 31 (9): 539–547. дои : 10.1016/j.tibtech.2013.06.001 . ПМИД 23830144 .
- ^ Куцлер М.А., Вайнер Д.Б. (октябрь 2008 г.). «ДНК-вакцины: готовы к прайм-тайму?» . Обзоры природы. Генетика . 9 (10): 776–788. дои : 10.1038/nrg2432 . ПМЦ 4317294 . ПМИД 18781156 .
- ^ Родригес Ф., Чжан Дж., Уиттон Дж.Л. (ноябрь 1997 г.). «ДНК-иммунизация: убиквитинирование вирусного белка усиливает индукцию цитотоксических Т-лимфоцитов и противовирусную защиту, но отменяет индукцию антител» . Журнал вирусологии . 71 (11): 8497–8503. doi : 10.1128/JVI.71.11.8497-8503.1997 . ЧВК 192313 . ПМИД 9343207 .
- ^ Перейти обратно: а б Тобери Т.В., Силициано РФ (март 1997 г.). «Нацеливание антигенов ВИЧ-1 на быструю внутриклеточную деградацию усиливает распознавание цитотоксических Т-лимфоцитов (CTL) и индукцию ответов de novo CTL in vivo после иммунизации» . Журнал экспериментальной медицины . 185 (5): 909–920. дои : 10.1084/jem.185.5.909 . ПМК 2196169 . ПМИД 9120397 .
- ^ Хюбенер Н., Фест С., Страндсби А., Михальски Е., Прейсснер Р., Цзэн Ю. и др. (июль 2008 г.). «Рационально разработанная ДНК-вакцина тирозингидроксилазы индуцирует специфический иммунитет против нейробластомы». Молекулярная терапия рака . 7 (7): 2241–2251. дои : 10.1158/1535-7163.MCT-08-0109 . ПМИД 18645033 . S2CID 35652424 .
- ^ Перейти обратно: а б Вундерлих Г., Моура И.С., дель Портильо Х.А. (октябрь 2000 г.). «Генетическая иммунизация мышей BALB/c плазмидой, несущей ген, кодирующий гибридный поверхностный белок мерозоитов и слитый поверхностный белок вируса гепатита B, защищает мышей от летальной инфекции Plasmodium chabaudi chabaudi PC1» . Инфекция и иммунитет . 68 (10): 5839–5845. дои : 10.1128/IAI.68.10.5839-5845.2000 . ПМЦ 101545 . ПМИД 10992493 .
- ^ Вайнер Д.Б., Кеннеди Р.К. (июль 1999 г.). «Генетические вакцины» . Научный американец . 281 (1): 50–57. Бибкод : 1999SciAm.281a..50W . doi : 10.1038/scientificamerican0799-50 . ПМИД 10396782 . Архивировано из оригинала 25 марта 2009 г. Проверено 21 ноября 2007 г.
- ^ Видера Г., Остин М., Рабуссей Д., Голдбек С., Барнетт С.В., Чен М. и др. (май 2000 г.). «Увеличение доставки ДНК-вакцины и иммуногенности с помощью электропорации in vivo» . Журнал иммунологии . 164 (9): 4635–4640. дои : 10.4049/jimmunol.164.9.4635 . ПМИД 10779767 .
- ^ Перейти обратно: а б Дахешиа М., Куклин Н., Канангат С., Маникан Э., Роуз Б.Т. (август 1997 г.). «Подавление продолжающегося воспалительного заболевания глаз путем местного введения плазмидной ДНК, кодирующей IL-10» . Журнал иммунологии . 159 (4): 1945–1952. doi : 10.4049/jimmunol.159.4.1945 . ПМИД 9257860 . S2CID 43203331 .
- ^ Перейти обратно: а б Чен Ю, Вебстер Р.Г., Вудленд Д.Л. (март 1998 г.). «Индукция CD8+ Т-клеточных ответов на доминантные и субдоминантные эпитопы и защитный иммунитет к инфекции вирусом Сендай с помощью ДНК-вакцинации» . Журнал иммунологии . 160 (5): 2425–2432. дои : 10.4049/jimmunol.160.5.2425 . ПМИД 9498786 . S2CID 2250871 .
- ^ Лоде Х.Н., Хюбенер Н., Зенг Ю., Фест С., Вейкслер С., Гаедике Г. (декабрь 2004 г.). «ДНК-минигенная вакцинация для адъювантной терапии нейробластомы». Анналы Нью-Йоркской академии наук . 1028 (1): 113–121. Бибкод : 2004NYASA1028..113L . дои : 10.1196/анналы.1322.012 . ПМИД 15650237 . S2CID 27240738 .
- ^ Сайзмор Д.Р., Бранстром А.А., Садофф Дж.К. (октябрь 1995 г.). «Аттенуированные шигеллы как средство доставки ДНК для ДНК-опосредованной иммунизации» . Наука . 270 (5234): 299–302. Бибкод : 1995Sci...270..299S . дои : 10.1126/science.270.5234.299 . ПМИД 7569980 . S2CID 12532901 .
- ^ Нилон, Кори (25 ноября 2014 г.). «Гибридный автомобиль, доставляющий ДНК» . Государственный университет Нью-Йорка в Буффало . Проверено 16 декабря 2014 г.
- ^ Джонс Ч., Равикришнан А., Чен М., Реддингер Р., Камаль Ахмади М., Рэйн С. и др. (август 2014 г.). «Разработка вектора гибридной биосинтетической генной терапии и двойной инженерный потенциал» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (34): 12360–12365. Бибкод : 2014PNAS..11112360J . дои : 10.1073/pnas.1411355111 . ПМК 4151754 . ПМИД 25114239 .
- ^ Барри М.А., Лай У.К., Джонстон С.А. (октябрь 1995 г.). «Защита от микоплазменной инфекции с использованием экспрессионной библиотечной иммунизации» . Природа . 377 (6550): 632–635. Бибкод : 1995Natur.377..632B . дои : 10.1038/377632a0 . ПМИД 7566175 . S2CID 4306972 .
- ^ Финан Э.Ф., Вебстер Р.Г., Фуллер Д.Х., Хейнс-младший, Санторо Дж.К., Робинсон Х.Л. (декабрь 1993 г.). «ДНК-вакцины: защитная иммунизация путем парентеральных прививок, прививок через слизистую оболочку и генной пушки» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (24): 11478–11482. Бибкод : 1993PNAS...9011478F . дои : 10.1073/pnas.90.24.11478 . ПМК 48007 . ПМИД 8265577 .
- ^ Перейти обратно: а б с д и Фелткват Д.М., Хини С., Вебстер Р.Г., Робинсон Х.Л. (март 1997 г.). «Различные типы Т-хелперных клеток и изотипы антител, генерируемые иммунизацией ДНК физиологическим раствором и генной пушкой» . Журнал иммунологии . 158 (5): 2278–2284. дои : 10.4049/jimmunol.158.5.2278 . ПМИД 9036975 . S2CID 41368723 .
- ^ Перейти обратно: а б с д Бойл С.М., Морин М., Вебстер Р.Г., Робинсон Х.Л. (декабрь 1996 г.). «Роль различных лимфоидных тканей в инициировании и поддержании реакций антител, вызванных ДНК, к гликопротеину H1 вируса гриппа» . Журнал вирусологии . 70 (12): 9074–9078. doi : 10.1128/JVI.70.12.9074-9078.1996 . ПМК 191015 . ПМИД 8971047 .
- ^ Селлберг М., Таунсенд К., Чен М., О'Ди Дж., Бэнкс Т., Джолли DJ и др. (июль 1997 г.). «Характеристика гуморальных и CD4+ клеточных ответов после генетической иммунизации ретровирусными векторами, экспрессирующими различные формы ядра вируса гепатита В и е-антигенов» . Журнал вирусологии . 71 (7): 5295–5303. doi : 10.1128/JVI.71.7.5295-5303.1997 . ЧВК 191766 . ПМИД 9188598 .
- ^ Баншеро Дж. , Стейнман Р.М. (март 1998 г.). «Дендритные клетки и контроль иммунитета». Природа . 392 (6673): 245–252. Бибкод : 1998Natur.392..245B . дои : 10.1038/32588 . ПМИД 9521319 . S2CID 4388748 .
- ^ Перейти обратно: а б Джейкоб Т., Уокер П.С., Криг А.М., Удей М.К., Фогель Дж.К. (сентябрь 1998 г.). «Активация дендритных клеток кожи CpG-содержащими олигодезоксинуклеотидами: роль дендритных клеток в усилении ответов Th1 иммуностимулирующей ДНК» . Журнал иммунологии . 161 (6): 3042–3049. дои : 10.4049/jimmunol.161.6.3042 . ПМИД 9743369 . S2CID 35107733 .
- ^ Перейти обратно: а б Раз Э., Тайге Х., Сато Ю., Корр М., Дадлер Дж.А., Роман М. и др. (май 1996 г.). «Преимущественная индукция иммунного ответа Th1 и ингибирование образования специфических антител IgE путем иммунизации плазмидной ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (10): 5141–5145. Бибкод : 1996PNAS...93.5141R . дои : 10.1073/pnas.93.10.5141 . ПМК 39421 . ПМИД 8643542 .
- ^ Перейти обратно: а б Фу Т.М., Фридман А., Улмер Дж.Б., Лю М.А., Доннелли Дж.Дж. (апрель 1997 г.). «Защитный клеточный иммунитет: ответы цитотоксических Т-лимфоцитов против доминантных и рецессивных эпитопов нуклеопротеина вируса гриппа, индуцированные иммунизацией ДНК» . Журнал вирусологии . 71 (4): 2715–2721. doi : 10.1128/JVI.71.4.2715-2721.1997 . ЧВК 191393 . ПМИД 9060624 .
- ^ Перейти обратно: а б с Рестифо Н.П., Бачик И., Ирвин К.Р., Юделл Дж.В., Маккейб Б.Дж., Андерсон Р.В. и др. (май 1995 г.). «Обработка антигена in vivo и выявление первичных ответов CTL» . Журнал иммунологии . 154 (9): 4414–4422. дои : 10.4049/jimmunol.154.9.4414 . ЧВК 1952186 . ПМИД 7722298 .
- ^ Перейти обратно: а б с Ивасаки А., Штирнхольм Б.Дж., Чан А.К., Беринштейн Н.Л., Барбер Б.Х. (май 1997 г.). «Усиление ответов CTL, опосредованных иммуногенами плазмидной ДНК, кодирующими костимулирующие молекулы и цитокины» . Журнал иммунологии . 158 (10): 4591–4601. дои : 10.4049/jimmunol.158.10.4591 . ПМИД 9144471 . S2CID 41779568 .
- ^ Перейти обратно: а б Вайс В.Р., Исии К.Дж., Хедстром Р.К., Седега М., Ичино М., Барнхарт К. и др. (сентябрь 1998 г.). «Плазмида, кодирующая мышиный гранулоцитарно-макрофагальный колониестимулирующий фактор, увеличивает защиту, обеспечиваемую ДНК-вакциной против малярии» . Журнал иммунологии . 161 (5): 2325–2332. дои : 10.4049/jimmunol.161.5.2325 . ПМИД 9725227 . S2CID 21038927 .
- ^ Цудзи Т., Хамадзима К., Фукусима Дж., Синь К.К., Исии Н., Аоки И. и др. (апрель 1997 г.). «Усиление клеточного иммунитета против ВИЧ-1, индуцированное совместной инокуляцией кодируемого плазмидой антигена ВИЧ-1 с плазмидой, экспрессирующей IL-12» . Журнал иммунологии . 158 (8): 4008–4013. дои : 10.4049/гиммунол.158.8.4008 . ПМИД 9103472 . S2CID 38099483 .
- ^ Перейти обратно: а б Юстевич Д.М., Вебстер Р.Г. (октябрь 1996 г.). «Долгосрочное поддержание В-клеточного иммунитета к гемагглютинину вируса гриппа у мышей после иммунизации на основе ДНК» . Вирусология . 224 (1): 10–17. дои : 10.1006/виро.1996.0501 . ПМИД 8862394 .
- ^ Манчини-Буржин М., Фонтен Х., Брешо С., Поль С., Мишель М.Л. (май 2006 г.). «Иммуногенность ДНК-вакцины против гепатита В, вводимой хроническим носителям ВГВ». Вакцина . 24 (21): 4482–4489. doi : 10.1016/j.vaccine.2005.08.013 . ПМИД 16310901 .
- ^ Перейти обратно: а б Вольф Дж.А., Даути М.Э., Цзяо С., Репетто Дж., Берг Р.К., Лудтке Дж.Дж. и др. (декабрь 1992 г.). «Экспрессия голых плазмид культивируемыми мышечными трубками и проникновение плазмид в Т-трубочки и кавеолы скелетных мышц млекопитающих». Журнал клеточной науки . 103. 103 (4): 1249–1259. дои : 10.1242/jcs.103.4.1249 . ПМИД 1487500 .
- ^ Андерсон Р.Г., Камен Б.А., Ротберг К.Г., Лейси С.В. (январь 1992 г.). «Потоцитоз: секвестрация и транспорт малых молекул кавеолами». Наука . 255 (5043): 410–411. Бибкод : 1992Sci...255..410A . дои : 10.1126/science.1310359 . ПМИД 1310359 .
- ^ Перейти обратно: а б Касарес С., Инаба К., Брумяну Т.Д., Стейнман Р.М., Бона К.А. (ноябрь 1997 г.). «Презентация антигена дендритными клетками после иммунизации ДНК, кодирующей вирусный эпитоп, ограниченный классом II главного комплекса гистосовместимости» . Журнал экспериментальной медицины . 186 (9): 1481–1486. дои : 10.1084/jem.186.9.1481 . ПМК 2199124 . ПМИД 9348305 .
- ^ Перейти обратно: а б Беннетт Р.М., Габор Г.Т., Мерритт М.М. (декабрь 1985 г.). «Связывание ДНК с лейкоцитами человека. Доказательства рецептор-опосредованной ассоциации, интернализации и деградации ДНК» . Журнал клинических исследований . 76 (6): 2182–2190. дои : 10.1172/JCI112226 . ПМК 424340 . ПМИД 3001145 .
- ^ Беннет Р.М., Хефенейдер С.Х., Бакке А., Мерритт М., Смит К.А., Моурих Д., Генрих М.К. (май 1988 г.). «Производство и характеристика мышиных моноклональных антител к рецептору ДНК на лейкоцитах человека» . Журнал иммунологии . 140 (9): 2937–2942. doi : 10.4049/jimmunol.140.9.2937 . ПМИД 2452195 . S2CID 22923379 .
- ^ Корр М., Ли DJ, Карсон Д.А., Тай Х. (октябрь 1996 г.). «Генная вакцинация обнаженной плазмидной ДНК: механизм праймирования CTL» . Журнал экспериментальной медицины . 184 (4): 1555–1560. дои : 10.1084/jem.184.4.1555 . ПМК 2192808 . ПМИД 8879229 .
- ^ Чаттергун М.А., Робинсон Т.М., Бойер Дж.Д., Вайнер Д.Б. (июнь 1998 г.). «Специфическая иммунная индукция после иммунизации на основе ДНК посредством трансфекции in vivo и активации макрофагов/антигенпрезентирующих клеток» . Журнал иммунологии . 160 (12): 5707–5718. дои : 10.4049/jimmunol.160.12.5707 . ПМИД 9637479 . S2CID 33499198 .
- ^ Перейти обратно: а б Торрес К.А., Ивасаки А., Барбер Б.Х., Робинсон Х.Л. (май 1997 г.). «Дифференциальная зависимость от ткани-мишени для иммунизации генной пушкой и внутримышечной ДНК» . Журнал иммунологии . 158 (10): 4529–4532. дои : 10.4049/jimmunol.158.10.4529 . ПМИД 9144463 . S2CID 45069087 .
- ^ Франко А., Гуидотти Л.Г., Хоббс М.В., Паскетто В., Чисари Ф.В. (август 1997 г.). «Патогенетическая эффекторная функция CD4-положительных Т-хелперов 1 у трансгенных мышей вируса гепатита В» . Журнал иммунологии . 159 (4): 2001–2008. doi : 10.4049/jimmunol.159.4.2001 . ПМИД 9257867 . S2CID 20528634 .
- ^ Манчини М., Хашуэль М., Дэвис Х.Л., Уэлен Р.Г., Тиолле П., Мишель М.Л. (октябрь 1996 г.). «ДНК-опосредованная иммунизация на модели трансгенных мышей с хроническим носительством поверхностного антигена гепатита В» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (22): 12496–12501. Бибкод : 1996PNAS...9312496M . дои : 10.1073/pnas.93.22.12496 . ПМК 38020 . ПМИД 8901610 .
- ^ Дулан Д.Л., Хоффман С.Л. (июль 1999 г.). «IL-12 и NK-клетки необходимы для антигенспецифического адаптивного иммунитета против малярии, инициируемого CD8+ Т-клетками в модели Plasmodium yoelii» . Журнал иммунологии . 163 (2): 884–892. дои : 10.4049/jimmunol.163.2.884 . ПМИД 10395683 . S2CID 41651105 .
- ^ Кардозо А.И., Бликсенкроне-Моллер М., Файоль Дж., Лю М., Бакленд Р., Уайлд Т.Ф. (ноябрь 1996 г.). «Иммунизация плазмидной ДНК, кодирующей гемагглютинин и нуклеопротеин вируса кори, приводит к гуморальному и клеточному иммунитету» . Вирусология . 225 (2): 293–299. дои : 10.1006/виро.1996.0603 . ПМИД 8918915 .
- ^ Перейти обратно: а б с Сато Ю., Роман М., Тиге Х., Ли Д., Корр М., Нгуен М.Д. и др. (июль 1996 г.). «Иммуностимулирующие последовательности ДНК, необходимые для эффективной внутрикожной иммунизации генов». Наука . 273 (5273): 352–354. Бибкод : 1996Sci...273..352S . дои : 10.1126/science.273.5273.352 . ПМИД 8662521 . S2CID 9333197 .
- ^ Вайс Р., Лейтнер В.В., Шайблхофер С., Чен Д., Бернхаупт А., Мостбёк С. и др. (октябрь 2000 г.). «Генетическая вакцинация против малярийной инфекции путем внутрикожных и эпидермальных инъекций плазмиды, содержащей ген, кодирующий белок циркумспорозоита Plasmodium berghei» . Инфекция и иммунитет . 68 (10): 5914–5919. дои : 10.1128/IAI.68.10.5914-5919.2000 . ПМК 101554 . ПМИД 10992502 .
- ^ Перейти обратно: а б Седега М., Вайс В., Саччи Дж.Б., Чароенвит Ю., Хедстром Р., Гауда К. и др. (июнь 2000 г.). «Улучшение защитного иммунитета, индуцированного иммунизацией на основе ДНК: праймирование антигеном и плазмидной ДНК, кодирующей GM-CSF, и бустирование рекомбинантным поксвирусом, экспрессирующим антиген» . Журнал иммунологии . 164 (11): 5905–5912. дои : 10.4049/jimmunol.164.11.5905 . ПМИД 10820272 .
- ^ Баруш Д.Х., Сантра С., Стенбеке Т.Д., Чжэн XX, Перри Х.К., Дэвис М.Э. и др. (август 1998 г.). «Усиление и подавление иммунного ответа на ДНК-вакцину ВИЧ-1 путем введения плазмидных цитокинов/Ig» . Журнал иммунологии . 161 (4): 1875–1882. doi : 10.4049/jimmunol.161.4.1875 . ПМИД 9712056 . S2CID 36488254 .
- ^ Перейти обратно: а б Криг А.М., Йи А.К., Мэтсон С., Вальдшмидт Т.Дж., Бишоп Г.А., Тисдейл Р. и др. (апрель 1995 г.). «Мотивы CpG в бактериальной ДНК вызывают прямую активацию B-клеток». Природа . 374 (6522): 546–549. Бибкод : 1995Natur.374..546K . дои : 10.1038/374546a0 . ПМИД 7700380 . S2CID 4261304 .
- ^ Клинман Д.М., Ямщиков Г., Исигацубо Ю. (апрель 1997 г.). «Вклад мотивов CpG в иммуногенность ДНК-вакцин» . Журнал иммунологии . 158 (8): 3635–3639. дои : 10.4049/jimmunol.158.8.3635 . ПМИД 9103425 . S2CID 41861994 .
- ^ Криг А.М., Ву Т., Виратна Р., Эфлер С.М., Лав-Хоман Л., Ян Л. и др. (октябрь 1998 г.). «Последовательные мотивы в аденовирусной ДНК блокируют иммунную активацию посредством стимулирующих мотивов CpG» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (21): 12631–12636. Бибкод : 1998PNAS...9512631K . дои : 10.1073/pnas.95.21.12631 . ПМК 22882 . ПМИД 9770537 .
- ^ Перейти обратно: а б Клинман Д.М., Йи А.К., Бокаж С.Л., Коновер Дж., Криг А.М. (апрель 1996 г.). «Мотивы CpG, присутствующие в ДНК бактерий, быстро побуждают лимфоциты секретировать интерлейкин 6, интерлейкин 12 и гамма-интерферон» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (7): 2879–2883. Бибкод : 1996PNAS...93.2879K . дои : 10.1073/pnas.93.7.2879 . ПМК 39727 . ПМИД 8610135 .
- ^ Йи А.К., Чейс Дж.Х., Каудери Дж.С., Криг А.М. (январь 1996 г.). «ИФН-гамма способствует секреции IL-6 и IgM в ответ на мотивы CpG в бактериальной ДНК и олигодезоксинуклеотидах» . Журнал иммунологии . 156 (2): 558–564. дои : 10.4049/jimmunol.156.2.558 . ПМИД 8543806 . S2CID 42145608 .
- ^ Барвик Б.Г., Шарер К.Д., Мартинес Р.Дж., Прайс М.Дж., Вейн А.Н., Хейнс Р.Р. и др. (май 2018 г.). «Активация В-клеток и дифференцировка плазматических клеток ингибируются метилированием ДНК de novo» . Природные коммуникации . 9 (1): 1900. Бибкод : 2018NatCo...9.1900B . дои : 10.1038/s41467-018-04234-4 . ПМЦ 5953949 . ПМИД 29765016 .
- ^ Летвин Н.Л., Монтефиори Д.С., Ясутоми Ю., Перри Х.К., Дэвис М.Э., Лекутис С. и др. (август 1997 г.). «Мощные защитные иммунные реакции против ВИЧ, генерируемые бимодальной ДНК оболочки ВИЧ плюс белковая вакцинация» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (17): 9378–9383. Бибкод : 1997PNAS...94.9378L . дои : 10.1073/pnas.94.17.9378 . ПМК 23198 . ПМИД 9256490 .
- ^ Седега М., Джонс Т.Р., Каур М., Хедстром Р., Хобарт П., Тайн Дж.А., Хоффман С.Л. (июнь 1998 г.). «Бункционирование рекомбинантной вакциной повышает иммуногенность и защитную эффективность ДНК-вакцины против малярии» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (13): 7648–7653. Бибкод : 1998PNAS...95.7648S . дои : 10.1073/pnas.95.13.7648 . ПМК 22711 . ПМИД 9636204 .
- ^ Роджерс В.О., Бэрд Дж.К., Кумар А., Тайн Дж.А., Вайс В., Агиар Дж.К. и др. (сентябрь 2001 г.). «Многоступенчатая мультиантигенная гетерологичная первичная буст-вакцина от малярии Plasmodium Knowlesi обеспечивает частичную защиту макак-резус» . Инфекция и иммунитет . 69 (9): 5565–5572. дои : 10.1128/IAI.69.9.5565-5572.2001 . ПМК 98670 . ПМИД 11500430 .
Дальнейшее чтение
[ редактировать ]- Хупер Дж.В., Томпсон Э., Вильгельмсен С., Циммерман М., Ичоу М.А., Штеффен С.Е. и др. (май 2004 г.). «ДНК-вакцина против оспы защищает приматов от смертельной оспы обезьян» . Журнал вирусологии . 78 (9): 4433–4443. doi : 10.1128/JVI.78.9.4433-4443.2004 . ПМЦ 387704 . ПМИД 15078924 .