Секс
| Часть серии о |
| Секс |
|---|
 |
| Биологические термины |
| Половое размножение |
| Сексуальность |
| Сексуальная система |
Пол — это биологический признак , определяющий, , размножающийся половым путем, производит ли организм мужские или женские гаметы . [ 1 ] [ 2 ] [ 3 ] [ 4 ] Во время полового размножения мужская и женская гаметы сливаются, образуя зиготу , из которой развивается потомство , унаследовавшее черты каждого родителя. Условно организмы, производящие более мелкие, более подвижные гаметы ( сперматозоиды , сперматозоиды ), называются мужскими , а организмы, производящие более крупные, неподвижные гаметы ( яйцеклетки , часто называемые яйцеклетками), называются женскими . [ 5 ] Организм, производящий оба типа гамет, является гермафродитом . [ 3 ] [ 6 ]
У негермафродитных видов пол особи определяется с помощью одной из нескольких биологических систем определения пола . Большинство видов млекопитающих имеют систему определения пола XY , при которой самец обычно несет X и Y-хромосому (XY), а самка обычно несет две Х-хромосомы (XX). Другие хромосомные системы определения пола у животных включают систему ZW у птиц и систему XO у некоторых насекомых. [ 7 ] Различные системы окружающей среды включают определение пола в зависимости от температуры у рептилий и ракообразных. [ 8 ]
Самец и самка вида могут быть физически похожими (половой мономорфизм) или иметь физические различия ( половой диморфизм ). У видов с половым диморфизмом, включая большинство птиц и млекопитающих, пол особи обычно определяется этой особи путем наблюдения за половыми признаками . Половой отбор или выбор партнера могут ускорить эволюцию различий между полами.
Термины «самец» и «самка» обычно не применяются к недифференцированным по половому признаку видам, у которых особи изоморфны (выглядят одинаково), а гаметы изогамны ( неразличимы по размеру и форме), например, к зеленой водоросли Ulvalactuca . Некоторые виды функциональных различий между особями, например, у грибов , [ 9 ] можно назвать типами спаривания . [ 10 ]
Половое размножение
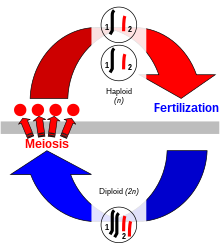
Половое размножение, при котором две особи производят потомство, обладающее набором генетических признаков каждого родителя, свойственно исключительно эукариотам . Генетические признаки закодированы в дезоксирибонуклеиновой кислоте (ДНК) хромосом . Клетка эукариота имеет набор парных гомологичных хромосом , по одной от каждого родителя, и эта двуххромосомная стадия называется « диплоидной ». Во время полового размножения диплоидный организм производит специализированные гаплоидные половые клетки, называемые гаметами посредством мейоза . [ 11 ] каждый из которых имеет единственный набор хромосом. Мейоз включает стадию генетической рекомбинации посредством хромосомного кроссовера , при котором участки ДНК обмениваются между совпавшими парами хромосом с образованием новых хромосом, каждая из которых содержит новую комбинацию генов родителей. Затем хромосомы разделяются на отдельные наборы в гаметах. Когда гаметы сливаются во время оплодотворения, образующаяся зигота содержит половину генетического материала матери и половину отца. [ 12 ] Комбинация хромосомного скрещивания и оплодотворения , объединяющая два отдельных набора хромосом для создания новой диплоидной зиготы , приводит к образованию нового организма, содержащего различный набор генетических признаков каждого родителя.
У животных гаплоидная стадия возникает только в гаметах, половых клетках, которые сливаются, образуя зиготу, которая развивается непосредственно в новый диплоидный организм. У видов растений диплоидный организм в результате мейоза производит тип гаплоидных спор , которые способны подвергаться повторному делению клеток с образованием многоклеточного гаплоидного организма. В любом случае гаметы могут быть внешне похожи ( изогамия ), как у зеленой водоросли Ulva , или могут отличаться по размеру и другим аспектам ( анизогамия ). [ 13 ] Наибольшая разница в размерах наблюдается при оогамии — типе анизогамии, при котором маленькая подвижная гамета соединяется с гораздо более крупной неподвижной гаметой. [ 14 ]
У анизогамных организмов по соглашению большая гамета (называемая яйцеклеткой или яйцеклеткой) считается женской, а меньшая гамета (называемая сперматозоидом или сперматозоидом) считается мужской. Особь, производящая крупные гаметы, — женщина, а особь, производящая мелкие гаметы, — мужчина. [ 15 ] Особь, у которой образуются оба типа гамет, является гермафродитом . У некоторых видов гермафродиты могут самооплодотворяться и производить потомство самостоятельно.
Животные

Большинство животных, размножающихся половым путем, проводят свою жизнь как диплоидные, при этом гаплоидная стадия редуцируется до одноклеточных гамет. [ 16 ] Гаметы животных имеют мужскую и женскую формы — сперматозоиды и яйцеклетки соответственно. Эти гаметы объединяются, образуя эмбрионы , из которых развиваются новые организмы.
Мужская гамета, сперматозоид (продуцируемый у позвоночных в семенниках ), представляет собой небольшую клетку, содержащую один длинный жгутик , который приводит ее в движение. [ 17 ] Сперматозоиды представляют собой чрезвычайно уменьшенные клетки, в которых отсутствуют многие клеточные компоненты, необходимые для эмбрионального развития. Они специализируются на подвижности, ищут яйцеклетку и сливаются с ней в процессе, называемом оплодотворением .
Женские гаметы представляют собой яйцеклетки. У позвоночных они вырабатываются в яичниках . Это большие неподвижные клетки, содержащие питательные вещества и клеточные компоненты, необходимые развивающемуся эмбриону. [ 18 ] Яйцеклетки часто связаны с другими клетками, которые поддерживают развитие эмбриона, образуя яйцеклетку . У млекопитающих оплодотворенный эмбрион развивается внутри самки, получая питание непосредственно от матери.
ищут партнера противоположного пола Животные обычно подвижны и для спаривания . Животные, живущие в воде, могут спариваться с помощью внешнего оплодотворения , при котором яйцеклетки и сперма высвобождаются в окружающую воду и соединяются с ней. [ 19 ] Однако большинство животных, живущих вне воды, используют внутреннее оплодотворение , передавая сперму непосредственно самке, чтобы предотвратить высыхание гамет.
У большинства птиц и выделение, и размножение осуществляются через одно заднее отверстие, называемое клоакой : самцы и самки птиц прикасаются к клоаке для передачи спермы - процесс, называемый «клоакальным поцелуем». [ 20 ] У многих других наземных животных самцы используют специальные половые органы для транспортировки спермы — эти мужские половые органы называются интромиттантными органами . У людей и других млекопитающих этот мужской орган известен как пенис , который входит в репродуктивный тракт самки (называемый влагалищем ) для достижения оплодотворения — процесса, называемого половым актом . Пенис содержит трубку, по которой проходит сперма (жидкость, содержащая сперматозоиды). У самок млекопитающих влагалище соединяется с маткой , органом, который непосредственно поддерживает развитие оплодотворенного эмбриона внутри (процесс, называемый беременностью ).
Из-за их подвижности сексуальное поведение животных может включать принудительный секс. Травматическое оплодотворение , например, используется некоторыми видами насекомых для оплодотворения самок через рану в брюшной полости — процесс, вредный для здоровья самки.
Растения
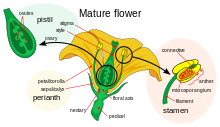
Как и животные, наземные растения имеют специализированные мужские и женские гаметы. [ 21 ] [ 22 ] У семенных растений мужские гаметы производятся редуцированными мужскими гаметофитами , которые содержатся в пыльце и имеют твердую оболочку, защищающую клетки, образующие мужские гаметы, во время транспорта от пыльников к рыльцу . Женские гаметы семенных растений содержатся в семязачатках . После оплодотворения они образуют семена , которые, как и яйца, содержат питательные вещества, необходимые для начального развития эмбрионального растения.

Цветки . цветковых растений содержат половые органы Большинство цветковых растений являются гермафродитами: как мужские, так и женские части находятся в одном цветке или на одном растении в однополых цветках, около 5% видов растений имеют отдельные растения одного или другого пола. [ 23 ] Женские части, расположенные в центре гермафродитного или женского цветка, представляют собой пестики , каждая часть которых состоит из плодолистика , столбика и рыльца . Две или более из этих репродуктивных единиц могут быть объединены в один сложный пестик , при этом сросшиеся плодолистики образуют завязь . Внутри плодолистиков находятся семязачатки , из которых после оплодотворения развиваются семена. Мужские части цветка — тычинки : они состоят из длинных нитей, расположенных между пестиком и лепестками, которые производят пыльцу в пыльниках на их кончиках. Когда пыльцевое зерно попадает на рыльце пестика на вершине плодолистика, оно прорастает, образуя пыльцевую трубку , которая прорастает через ткани столбика в плодолистик, где оно доставляет ядра мужских гамет для оплодотворения семяпочки, которая в конечном итоге развивается в плодолистик. семя.
Некоторые гермафродитные растения самоплодны, но растения выработали множество различных механизмов самонесовместимости , чтобы избежать самооплодотворения, включая последовательный гермафродитизм , системы молекулярного распознавания и морфологические механизмы, такие как гетеростилия . [ 24 ] : 73, 74
У сосен и других хвойных половые органы образуются внутри шишек , имеющих мужскую и женскую формы. Мужские шишки меньше женских и производят пыльцу, которая переносится ветром на землю в женских шишках. Более крупные и долгоживущие женские шишки, как правило, более долговечны и содержат внутри себя семяпочки, из которых после оплодотворения развиваются семена.
Поскольку семенные растения неподвижны, они зависят от пассивных методов транспортировки пыльцевых зерен к другим растениям. Многие из них, в том числе хвойные и травы, производят легкую пыльцу, которая переносится ветром на соседние растения. У некоторых цветковых растений более тяжелая и липкая пыльца, предназначенная для транспортировки насекомыми или более крупными животными, такими как колибри и летучие мыши , которых могут привлечь цветы, содержащие нектар и пыльцу. Эти животные переносят пыльцу по мере перемещения к другим цветкам, которые также содержат женские репродуктивные органы, что приводит к опылению .
Грибы

Большинство видов грибов могут размножаться половым путем и имеют жизненный цикл как с гаплоидными, так и с диплоидными фазами. Эти виды грибов обычно изогамны , то есть лишены мужской и женской специализации. Один гаплоидный гриб соприкасается с другим, а затем они сливают свои клетки. В некоторых случаях слияние асимметрично, и клетку, которая отдает только ядро (без сопутствующего клеточного материала), можно считать мужской. [ 25 ] Грибы также могут иметь более сложные аллельные системы спаривания, при этом представители других полов неточно описываются как мужские, женские или гермафродитные. [ 26 ]
Некоторые грибы, в том числе пекарские дрожжи , имеют типы спаривания , определяющие совместимость. Дрожжи с одинаковыми типами спаривания не сливаются друг с другом с образованием диплоидных клеток, а только с дрожжами, несущими другой тип спаривания. [ 27 ]
Многие виды высших грибов образуют грибы в процессе полового размножения . Внутри гриба образуются диплоидные клетки, позже делящиеся на гаплоидные споры .
Сексуальные системы
Половая система — это распределение мужских и женских функций между организмами одного вида. [ 28 ]
Животные
Примерно 95% видов животных имеют отдельных самцов и самок и считаются гонохорическими . Около 5% видов животных являются гермафродитами. [ 28 ] Этот низкий процент частично объясняется очень большим количеством видов насекомых , у которых гермафродитизм отсутствует. [ 29 ] Около 99% позвоночных — гонохоры, а оставшийся 1% — гермафродиты — почти все рыбы. [ 30 ]
Растения
Большинство растений двуполые . [ 31 ] : 212 либо гермафродит (с тычинками и пестиком в одном цветке), либо однодомный . [ 32 ] [ 33 ] У двудомных видов мужской и женский пол находятся на отдельных растениях. [ 34 ] Около 5% цветковых растений двудомные и имеют до 5000 независимых источников. [ 35 ] Двудомность распространена у голосеменных , у которых около 65% видов двудомны, но большинство хвойных однодомны. [ 36 ]
Эволюция секса
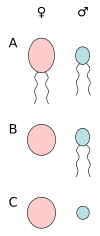
А) анизогамия подвижных клеток, Б) оогамия (яйцеклетка и сперматозоид), В) анизогамия неподвижных клеток (яйцеклетка и сперматозоид).
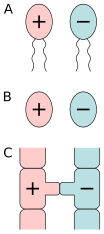
А) изогамия подвижных клеток , Б) изогамия неподвижных клеток, В) конъюгация.
Принято считать, что изогамия была предком анизогамии . [ 37 ] и что анизогамия развивалась несколько раз независимо в разных группах эукариот, включая простейших, водоросли, растения и животные. [ 29 ] Эволюция анизогамии является синонимом происхождения мужчины и происхождения женщины . [ 38 ] Это также первый шаг к половому диморфизму. [ 39 ] и повлияли на эволюцию различных половых различий. [ 40 ]
Неясно, привела ли анизогамия сначала к эволюции гермафродитизма или к эволюции гонохоризма . [ 31 ] : 213 а эволюция сперматозоидов и яйцеклеток не оставила никаких ископаемых свидетельств. [ 41 ]
Окаменелость Bangiomorpha pubescens возрастом 1,2 миллиарда лет предоставила старейшую летопись окаменелостей, позволяющую дифференцировать мужские и женские репродуктивные типы, и показала, что у эукариотов полы эволюционировали на ранней стадии. [ 42 ] Исследования зеленых водорослей предоставили генетические доказательства эволюционной связи между полами и типами спаривания . [ 43 ]
Первоначальной формой секса было наружное оплодотворение . Внутреннее оплодотворение , или пол, как мы его знаем, возникло позже. [ 44 ] и стал доминирующим для позвоночных после их выхода на сушу . [ 45 ]
Адаптивная функция пола
Самая основная роль мейоза , по-видимому, заключается в сохранении целостности генома , который передается потомству родителями. [ 46 ] [ 47 ] Два наиболее фундаментальных аспекта полового размножения , мейотическая рекомбинация и ауткроссинг , вероятно, поддерживаются соответственно адаптивными преимуществами рекомбинационного восстановления повреждений геномной ДНК и генетической комплементации , которая маскирует проявление вредных рецессивных мутаций . [ 48 ] Генетическая изменчивость , часто возникающая как побочный продукт этих процессов, может обеспечить долгосрочные преимущества в тех половых линиях, которые благоприятствуют ауткроссингу . [ 48 ]
Системы определения пола

Биологическая причина развития организма в тот или иной пол называется детерминацией пола . Причиной может быть генетика, окружающая среда, гаплодиплоидия или множество факторов. [ 29 ] У животных и других организмов, имеющих генетическую систему определения пола, определяющим фактором может быть наличие половой хромосомы . У растений с половым диморфизмом, например у гинкго двулопастного , [ 24 ] : 203 у печеночницы Marchantia polymorpha или двудомного вида цветковых растений рода Silene пол также может определяться половыми хромосомами. [ 49 ] Негенетические системы могут использовать сигналы окружающей среды, такие как температура на ранних этапах развития крокодилов , для определения пола потомства. [ 50 ]
Определение пола часто отличается от половой дифференциации . Определение пола — это обозначение стадии развития по направлению к мужскому или женскому полу, тогда как дифференциация пола — это путь к развитию фенотипа . [ 51 ]
Генетический
Определение пола XY

У людей и большинства других млекопитающих есть система определения пола XY : Y-хромосома несет в себе факторы, ответственные за запуск мужского развития, поэтому определение пола XY в основном основано на наличии или отсутствии Y-хромосомы . Именно мужская гамета определяет пол потомства. [ 52 ] В этой системе XX млекопитающие обычно являются самками, а XY — самцами. [ 29 ] Однако люди с XXY или XYY — мужчины, а люди с X и XXX — женщины. [ 8 ] Необычно то, что утконос , однопроходное млекопитающее, имеет десять половых хромосом; у женщин есть десять Х-хромосом, а у мужчин — пять Х-хромосом и пять Y-хромосом. Все яйцеклетки утконоса имеют пять Х-хромосом, тогда как сперматозоиды могут иметь либо пять Х-хромосом, либо пять Y-хромосом. [ 53 ]
Определение пола XY встречается и у других организмов, в том числе у насекомых, таких как обыкновенная плодовая мушка , [ 54 ] и некоторые растения. [ 55 ] В некоторых случаях пол определяет количество Х-хромосом, а не наличие Y-хромосомы. [ 8 ] У плодовой мухи особи с XY — самцы, а особи с XX — самки; однако люди с XXY или XXX также могут быть женщинами, а люди с X могут быть мужчинами. [ 56 ]
ZW определение пола
У птиц, имеющих систему определения пола ZW , W-хромосома несет факторы, ответственные за развитие самок, а развитие по умолчанию – самцов. [ 57 ] В этом случае особи ZZ — мужчины, а ZW — женщины. Именно женская гамета определяет пол потомства. Эту систему используют птицы, некоторые рыбы и некоторые ракообразные . [ 8 ]
У большинства бабочек и мотыльков также есть система определения пола ZW. У женщин могут быть Z, ZZW и даже ZZWW. [ 58 ]
XO определение пола
В системе определения пола XO мужчины имеют одну Х-хромосому (XO), а женщины - две (XX). Все остальные хромосомы у этих диплоидных организмов парные, но организмы могут наследовать одну или две Х-хромосомы. Эта система встречается у большинства паукообразных , насекомых, таких как чешуйница ( Apterygota ), стрекоз ( Paleoptera ) и кузнечиков ( Exopterygota ), а также у некоторых нематод, ракообразных и брюхоногих моллюсков. [ 59 ] [ 60 ]
у полевых сверчков насекомые с одной Х-хромосомой развиваются как самцы, а с двумя — как самки. Например, [ 61 ]
У нематоды Caenorhabditis elegans большинство червей являются самооплодотворяющимися гермафродитами с кариотипом ХХ, но случайные аномалии в наследовании хромосом могут привести к появлению особей только с одной Х-хромосомой — эти особи ХО являются плодовитыми самцами (и половина их потомства — самцы). [ 62 ]
ЗО определение пола
В системе определения пола ZO у мужчин есть две Z-хромосомы, а у женщин - одна. Эта система встречается у нескольких видов бабочек. [ 63 ]
Относящийся к окружающей среде
Для многих видов пол определяется не наследственными признаками, а факторами окружающей среды, такими как температура, испытываемая во время развития или на более позднем этапе жизни. [ 64 ]
У папоротника Ceratopteris и других гомоспоровых видов папоротников пол по умолчанию - гермафродитный, но особи, растущие в почве, которая ранее содержала гермафродитов, под влиянием феромона антеридиогена развиваются как мужские. [ 49 ] Личинки bonelliidae могут развиваться как самцы только при встрече с самкой. [ 29 ]
Последовательный гермафродитизм

Некоторые виды могут менять пол в течение своей жизни — явление, называемое последовательным гермафродитизмом . [ 65 ]
Костистые рыбы — единственная линия позвоночных , у которой встречается последовательный гермафродитизм. У рыбы-клоуна более мелкими рыбами являются самцы, а доминирующей и самой крупной рыбой в группе становится самка; когда доминирующая самка отсутствует, ее партнер меняет пол с мужского на женский. У многих губанов все наоборот: рыбы изначально являются самками и становятся самцами, когда достигают определенного размера. [ 66 ]
Последовательный гермафродитизм также встречается у таких растений, как Arisaema triphyllum .
Определение пола в зависимости от температуры

У многих рептилий , включая всех крокодилов и большинство черепах , определение пола зависит от температуры . У этих видов температура, которую испытывают эмбрионы во время развития, определяет их пол. [ 29 ]
Например, у некоторых черепах самцы производятся при более низких температурах, чем самки; но самки Macroclemys производятся при температуре ниже 22 ° C или выше 28 ° C, а самцы - в промежутке между этими температурами. [ 68 ]
Гаплодиплоиды
Некоторые насекомые, такие как медоносные пчелы и муравьи , используют гаплодиплоидную систему определения пола . [ 69 ] Диплоидные пчелы и муравьи — это, как правило, самки, а гаплоидные особи (которые развиваются из неоплодотворенных яиц) — самцы. Эта система определения пола приводит к сильно смещенному соотношению полов , поскольку пол потомства определяется оплодотворением ( арренотокия или псевдоарренотокия, приводящая к образованию самцов), а не набором хромосом во время мейоза. [ 70 ]
Соотношение полов
Соотношение полов — это соотношение мужчин и женщин в популяции . Как объясняется принципом Фишера , по эволюционным причинам это соотношение обычно составляет примерно 1:1 у видов, размножающихся половым путем . [ 71 ] [ 72 ] Однако многие виды отклоняются от равномерного соотношения полов периодически или постоянно. Примеры включают партеногенные виды, периодически спаривающиеся организмы, такие как тля, некоторые эусоциальные осы , пчелы , муравьи и термиты . [ 73 ]
Соотношение полов человека представляет особый интерес для антропологов и демографов. В человеческих обществах соотношение полов при рождении может быть значительно искажено такими факторами, как возраст матери при рождении. [ 74 ] а также аборты по признаку пола и детоубийство. Воздействие пестицидов и других загрязнителей окружающей среды также может быть существенным фактором, способствующим этому. [ 75 ] По состоянию на 2024 год глобальное соотношение полов при рождении оценивается в 107 мальчиков на 100 девочек (1000 мальчиков на 934 девочки). [ 76 ] К старости соотношение полов меняется: на каждые 100 пожилых женщин приходится 81 пожилой мужчина; Для всех возрастов население мира почти сбалансировано: на каждые 100 женщин приходится 101 мужчина. [ 76 ]Половые различия
Анизогамия – фундаментальное различие между мужчиной и женщиной. [ 77 ] [ 78 ] Ричард Докинз заявил, что все различия между полами можно интерпретировать как вытекающие из этого. [ 79 ]
Сексуальные характеристики
Половой диморфизм


У многих животных и некоторых растений особи мужского и женского пола различаются по размеру и внешнему виду — явление, называемое половым диморфизмом . [ 81 ] Половой диморфизм у животных часто связан с половым отбором : спариванием особей одного пола с противоположным полом. [ 82 ] Другие примеры демонстрируют, что половой диморфизм обусловлен предпочтением самок, как, например, в случае со стебельчатой мухой . [ 83 ]
Половые различия у людей включают, как правило, больший размер и больше волос на теле у мужчин, в то время как у женщин большая грудь, более широкие бедра и более высокий процент жира в организме. У других видов могут быть различия в окраске или других особенностях, и они могут быть настолько выраженными, что представители разных полов могут быть ошибочно приняты за два совершенно разных таксона. [ 82 ]
Самки составляют более крупный пол у большинства животных. [ 81 ] Например, самки южных пауков черной вдовы обычно в два раза длиннее самцов. [ 84 ] Эта разница в размерах может быть связана со стоимостью производства яйцеклеток, для чего требуется больше питания, чем для производства спермы: более крупные самки способны производить больше яйцеклеток. [ 85 ] [ 81 ] Во многих других случаях самец вида крупнее самки. Виды млекопитающих с крайним половым диморфизмом размеров, такие как морские слоны , как правило, имеют весьма полигинную систему спаривания, предположительно из-за отбора на успех в конкуренции с другими самцами.
Половой диморфизм может быть крайним: самцы, такие как некоторые удильщики , паразитируют на самке. Некоторые виды растений также демонстрируют диморфизм, при котором самки значительно крупнее самцов, например, у рода мхов Dicranum. [ 86 ] и род печеночников Sphaerocarpos . [ 87 ] Есть некоторые свидетельства того, что у этих родов диморфизм может быть связан с половой хромосомой. [ 87 ] [ 88 ] или химической передаче сигналов от самок. [ 89 ]
У птиц самцы часто имеют более яркую внешность и могут иметь особенности (например, длинный хвост у самцов павлинов), которые, по-видимому, ставят их в невыгодное положение (например, яркие цвета делают птицу более заметной для хищников). Одним из предлагаемых объяснений этого является принцип гандикапа . [ 90 ] Эта гипотеза утверждает, что, демонстрируя, что он может выжить с такими недостатками, мужчина рекламирует свою генетическую приспособленность женщинам - черты, которые принесут пользу и дочерям, которые не будут обременены такими недостатками.
Половые различия в поведении
Полы гонохорных видов обычно различаются по поведению. У большинства видов животных самки больше вкладывают в родительскую заботу. [ 91 ] хотя у некоторых видов, таких как некоторые кукулы , самцы уделяют больше родительской заботы . [ 92 ] Самки также, как правило, более разборчивы в выборе того, с кем они спариваются. [ 93 ] как и большинство видов птиц. [ 94 ] Самцы, как правило, более конкурентоспособны при спаривании, чем самки. [ 38 ]
Отличие от пола
См. также
Ссылки
- ^ Стивенсон А., Уэйт М. (2011). «Секс» . Краткий Оксфордский словарь английского языка: книга и набор компакт-дисков . ОУП Оксфорд . п. 1302. ИСБН 978-0-19-960110-3 . Проверено 23 марта 2018 г.
Пол: одна из двух основных категорий (мужская и женская), на которые делятся люди и большинство других живых существ на основе их репродуктивных функций. Факт принадлежности к одной из этих категорий. Группа всех представителей обоего пола.
- ^ Миллс А (1 января 2018 г.). «Секс и репродукция» . Биология секса . Университет Торонто Пресс. стр. 43–45. ISBN 978-1-4875-9337-7 . Проверено 3 октября 2023 г.
- ^ Перейти обратно: а б Первс В.К., Садава Д.Е., Орианс Г.Х. , Хеллер Х.К. (2000). Жизнь: наука биология . Макмиллан . п. 736. ИСБН 978-0-7167-3873-2 . Проверено 23 марта 2018 г.
Одно и то же тело может функционировать как мужское, так и женское. Для полового размножения необходимы как мужские, так и женские гаплоидные гаметы. У большинства видов эти гаметы производятся особями мужского или женского пола. Виды, у которых есть мужские и женские особи, называются раздельнополыми (от греческого «два дома»). У некоторых видов одна особь может обладать как женской, так и мужской репродуктивной системой. Такие виды называются однодомными («один дом») или гермафродитами.
- ^ Де Луф А (31 января 2018 г.). «Только две половые формы, но несколько гендерных вариантов: как объяснить?» . Коммуникативная и интегративная биология . 11 (1): e1427399. дои : 10.1080/19420889.2018.1427399 . ISSN 1942-0889 . ПМЦ 5824932 . ПМИД 29497472 .
- ^ Кокко Х. , Дженнионс М. (2012). «Половые различия в родительской заботе» . В Ройле, штат Нью-Джерси, Смизет П.Т., Кёлликер М. (ред.). Эволюция родительской заботы . Издательство Оксфордского университета. п. 103. ИСБН 978-0-19-969257-6 .
Ответ заключается в том, что существует условное соглашение: особи, производящие меньший из двух типов гамет (сперматозоиды или пыльца), являются самцами, а те, которые производят более крупные гаметы (яйцеклетки или семязачатки), — самки.
- ^ Авизе Дж.К. (2011). «Два пола в одном» . Гермафродитизм: учебник по биологии, экологии и эволюции двойной сексуальности . Издательство Колумбийского университета. стр. 1–7. ISBN 978-0-231-52715-6 . Проверено 18 сентября 2020 г.
- ^ Блэкмон Х., Росс Л., Бахтрог Д. (январь 2017 г.). «Определение пола, половые хромосомы и эволюция кариотипа у насекомых» . Журнал наследственности . 108 (1): 78–93. doi : 10.1093/jhered/esw047 . ISSN 0022-1503 . ПМК 6281344 . ПМИД 27543823 .
- ^ Перейти обратно: а б с д Хейк Л., О'Коннор С. (2008). «Генетические механизмы определения пола | Изучайте науку в Scitable» . Природное образование . 1 (1): 25. Архивировано из оригинала 19 августа 2017 года . Проверено 13 апреля 2021 г.
- ^ Мур Д., Робсон Дж. Д., Тринчи А. П. (2020). Путеводитель по грибам 21 века (2-е изд.). Издательство Кембриджского университета. стр. 211–228. ISBN 978-1-108-74568-0 .
- ^ Кумар Р., Мина М., Свапнил П. (2019). "Анизогамия". В Вонк Дж, Шакелфорд Т. (ред.). Энциклопедия познания и поведения животных . Чам: Международное издательство Springer. стр. 1–5. дои : 10.1007/978-3-319-47829-6_340-1 . ISBN 978-3-319-47829-6 .
Анизогамию можно определить как способ полового размножения, при котором сливающиеся гаметы, образованные участвующими родителями, различаются по размеру.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Мейоз» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 25 января 2017 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Польза секса» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 .
- ^ Гилберт С.Ф. (2000). «Многоклеточность: эволюция дифференцировки» . Биология развития (6-е изд.). Сандерленд (Массачусетс): Sinauer Associates. ISBN 978-0-87893-243-6 . Архивировано из оригинала 8 марта 2021 года . Проверено 17 апреля 2021 г.
- ^ Аллаби М (2012). Словарь наук о растениях . ОУП Оксфорд. п. 350. ИСБН 978-0-19-960057-1 .
- ^ Ну и дела Х (22 ноября 1999 г.). «Размер и единственная половая клетка» . Природа . Архивировано из оригинала 11 октября 2017 года . Проверено 4 июня 2018 г.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Менделевская генетика в жизненных циклах эукариот» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 2 апреля 2017 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Сперма» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 29 июня 2009 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Яйца» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 29 июня 2009 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Оплодотворение» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 19 декабря 2008 года.
- ^ Ритчисон Г. «Размножение птиц» . Университет Восточного Кентукки. Архивировано из оригинала 12 апреля 2008 года . Проверено 3 апреля 2008 г.
- ^ Гилберт С.Ф. (2000). «Производство гамет у покрытосеменных» . Биология развития (6-е изд.). Сандерленд (Массачусетс): Sinauer Associates. ISBN 978-0-87893-243-6 . Архивировано из оригинала 21 апреля 2021 года . Проверено 17 апреля 2021 г.
- ^ Дюзенбери Д.Б. (2009). Жизнь в микромасштабе: неожиданная физика маленького размера . Издательство Гарвардского университета. стр. 308–326. ISBN 978-0-674-03116-6 .
- ^ Уилтон П. (12 марта 2009 г.). «Растения, секс и Дарвин» . OxSciБлог . Оксфордский университет . Проверено 10 января 2024 г.
- ^ Перейти обратно: а б Джадд В.С. , Кэмпбелл К.С., Келлог Э.А. , Стивенс П.Ф. , Донохью М.Дж. (2002). Систематика растений, филогенетический подход (2-е изд.). Sinauer Associates Inc. Сандерленд, Массачусетс: ISBN 0-87893-403-0 .
- ^ Ник Лейн (2005). Власть, секс, самоубийство: митохондрии и смысл жизни . Издательство Оксфордского университета. стр. 236–237 . ISBN 978-0-19-280481-5 .
- ^ Уоткинсон СК, Бодди Л. , Деньги Н. (2015). Грибы . Эльзевир Наука. п. 115. ИСБН 978-0-12-382035-8 . Проверено 18 февраля 2018 г.
- ^ Лодиш Х., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «Спецификация типа клетки и преобразование типа спаривания в дрожжах» . Молекулярно-клеточная биология (Четвертое изд.). WH Freeman and Co. ISBN 978-0-7167-4366-8 . Архивировано из оригинала 1 июля 2009 года.
- ^ Перейти обратно: а б Леонард Дж.Л. (22 августа 2013 г.). «Парадокс Уильямса и роль фенотипической пластичности в сексуальных системах» . Интегративная и сравнительная биология . 53 (4): 671–688. дои : 10.1093/icb/ict088 . ISSN 1540-7063 . ПМИД 23970358 .
- ^ Перейти обратно: а б с д и ж Бахтрог Д., Манк Дж.Э. , Пайчел К.Л., Киркпатрик М. , Отто С.П. , Эшман Т.Л. и др. (июль 2014 г.). «Определение пола: почему так много способов сделать это?» . ПЛОС Биология . 12 (7): e1001899. дои : 10.1371/journal.pbio.1001899 . ПМК 4077654 . ПМИД 24983465 .
- ^ Кувамура Т., Сунобе Т., Сакаи Ю., Кадота Т., Савада К. (1 июля 2020 г.). «Гермафродитизм у рыб: аннотированный список видов, филогения и система спаривания» . Ихтиологические исследования . 67 (3): 341–360. Бибкод : 2020IchtR..67..341K . дои : 10.1007/s10228-020-00754-6 . ISSN 1616-3915 . S2CID 218527927 .
- ^ Перейти обратно: а б Климан Р.М. (2016). Энциклопедия эволюционной биологии . Том. 2. Академическая пресса. стр. 212–224. ISBN 978-0-12-800426-5 . Архивировано из оригинала 6 мая 2021 года . Проверено 14 апреля 2021 г.
- ^ Сабат Н., Голдберг Э.Э., Глик Л., Эйнхорн М., Эшман Т.Л., Минг Р. и др. (февраль 2016 г.). «Двудомность не ускоряет и не замедляет диверсификацию линий нескольких родов покрытосеменных растений» . Новый фитолог . 209 (3): 1290–1300. дои : 10.1111/nph.13696 . ПМИД 26467174 .
- ^ Бентдже Х (2016). Глоссарий завода в Кью (2-е изд.). Королевский ботанический сад, Кью: Издательство Кью. ISBN 978-1-84246-604-9 .
- ^ Лейте Монтальвао А.П., Керстен Б., Фладунг М., Мюллер Н.А. (2021). «Разнообразие и динамика определения пола у двудомных растений» . Границы в науке о растениях . 11 : 580488. doi : 10.3389/fpls.2020.580488 . ПМЦ 7843427 . ПМИД 33519840 .
- ^ Реннер СС (октябрь 2014 г.). «Относительные и абсолютные частоты половых систем покрытосеменных: двудомность, моноэция, гинодиэция и обновленная онлайн-база данных» . Американский журнал ботаники . 101 (10): 1588–1596. дои : 10.3732/ajb.1400196 . ПМИД 25326608 .
- ^ Валас Л., Мандрик В., Томас П.А., Тырала-Веруцка Ж., Ишкуло Г. (2018). «Половая система голосеменных растений: обзор» (PDF) . Фундаментальная и прикладная экология . 31 : 1–9. Бибкод : 2018BApEc..31....1W . дои : 10.1016/j.baae.2018.05.009 . S2CID 90740232 . Архивировано (PDF) из оригинала 27 января 2022 года . Проверено 7 июня 2021 г.
- ^ Авасти АК. Учебник Водорослей . Издательство Викас. п. 363. ИСБН 978-93-259-9022-7 .
- ^ Перейти обратно: а б Лехтонен Дж., Кокко Х. , Паркер Г.А. (октябрь 2016 г.). «Что изогамные организмы говорят нам о сексе и двух полах?» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1706). дои : 10.1098/rstb.2015.0532 . ПМК 5031617 . ПМИД 27619696 .
- ^ Тогаши Т., Бартелт Дж.Л., Йошимура Дж., Тайнака К., Кокс П.А. (август 2012 г.). «Эволюционные траектории объясняют разнообразную эволюцию изогамии и анизогамии морских зеленых водорослей» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (34): 13692–13697. Бибкод : 2012PNAS..10913692T . дои : 10.1073/pnas.1203495109 . ПМЦ 3427103 . ПМИД 22869736 .
- ^ Секели Т. , DJ Fairbairn, Blanckenhorn WU (2007). Пол, размер и гендерные роли: эволюционные исследования диморфизма полового размера . ОУП Оксфорд. стр. 167–169, 176, 185. ISBN. 978-0-19-920878-4 .
- ^ Питник С.С., Хоскен DJ, Биркхед Т.Р. (2008). Биология спермы: эволюционная перспектива . Академическая пресса. стр. 43–44. ISBN 978-0-08-091987-4 .
- ^ Хёрандл Э., Хадачек Ф (август 2020 г.). «Кислород, формы жизни и эволюция полов у многоклеточных эукариот» . Наследственность . 125 (1–2): 1–14. дои : 10.1038/s41437-020-0317-9 . ПМЦ 7413252 . ПМИД 32415185 .
- ^ Савада Х., Иноуэ Н., Ивано М., ред. (2014). Половое размножение животных и растений . Спрингер. стр. 215–227. дои : 10.1007/978-4-431-54589-7 . ISBN 978-4-431-54589-7 .
- ^ Черный R (19 октября 2014 г.). «Бронированная рыба стала пионером секса, каким вы его знаете» . Животные . Нэшнл Географик. Архивировано из оригинала 2 марта 2021 года . Проверено 10 июля 2023 г.
- ^ «43.2А: Внешнее и внутреннее оплодотворение» . Свободные тексты по биологии . 17 июля 2018 года. Архивировано из оригинала 24 мая 2022 года . Проверено 9 ноября 2020 г.
- ^ Брандейс М. (май 2018 г.). «Новые идеи о древнем сексе: отделение мейоза от спаривания может решить загадку вековой давности». Биологические обзоры Кембриджского философского общества . 93 (2): 801–810. дои : 10.1111/brv.12367 . ПМИД 28913952 .
- ^ Хёрандль Э (июнь 2024 г.). «Апомиксис и парадокс пола у растений». Анналы ботаники . 134 (1): 1–18. дои : 10.1093/aob/mcae044 . PMC 11161571. PMID 38497809 .
- ^ Перейти обратно: а б Бернштейн Х., Байерли Х.К., Хопф Ф.А., Мишод Р.Э. (сентябрь 1985 г.). «Генетические повреждения, мутации и эволюция пола». Наука . 229 (4719). Нью-Йорк, штат Нью-Йорк: 1277–81. дои : 10.1126/science.3898363 . ПМИД 3898363 .
- ^ Перейти обратно: а б Танурдзич М., Бэнкс Дж.А. (2004). «Механизмы определения пола у наземных растений» . Растительная клетка . 16 (Дополнение): S61–S71. дои : 10.1105/tpc.016667 . ПМЦ 2643385 . ПМИД 15084718 .
- ^ Уорнер Д.А., Шайн Р. (январь 2008 г.). «Адаптационное значение температурно-зависимого определения пола у рептилий». Природа . 451 (7178): 566–568. Бибкод : 2008Natur.451..566W . дои : 10.1038/nature06519 . ПМИД 18204437 . S2CID 967516 .
- ^ Бекебум Л.В., Перрин Н. (2014). Эволюция определения пола . Издательство Оксфордского университета. п. 16. ISBN 978-0-19-965714-8 .
- ^ Уоллис MC, Уотерс PD, Грейвс Дж. А. (октябрь 2008 г.). «Определение пола у млекопитающих – до и после эволюции SRY» . Клеточные и молекулярные науки о жизни . 65 (20): 3182–95. дои : 10.1007/s00018-008-8109-z . ПМЦ 11131626 . ПМИД 18581056 . S2CID 31675679 .
- ^ Пирс Б.А. (2012). Генетика: концептуальный подход (4-е изд.). Нью-Йорк: WH Freeman. стр. 100-1 73–74. ISBN 978-1-4292-3250-0 . OCLC 703739906 .
- ^ Кайзер В.Б., Бахтрог Д. (2010). «Эволюция половых хромосом у насекомых» . Ежегодный обзор генетики . 44 : 91–112. doi : 10.1146/annurev-genet-102209-163600 . ПМК 4105922 . ПМИД 21047257 .
- ^ Деллапорта С.Л., Кальдерон-Урреа А (октябрь 1993 г.). «Определение пола у цветковых растений» . Растительная клетка . 5 (10): 1241–1251. дои : 10.1105/tpc.5.10.1241 . JSTOR 3869777 . ПМК 160357 . ПМИД 8281039 .
- ^ Фуско Дж., Минелли А. (2019). Биология размножения . Издательство Кембриджского университета. стр. 306–308. ISBN 978-1-108-49985-9 .
- ^ Смит Калифорния, Кац М., Синклер А.Х. (февраль 2003 г.). «DMRT1 активируется в гонадах во время смены пола от женского к мужскому у куриных эмбрионов ZW» . Биология размножения . 68 (2): 560–570. дои : 10.1095/biolreprod.102.007294 . ПМИД 12533420 .
- ^ Маджерус М.Э. (2003). Сексуальные войны: гены, бактерии и предвзятое соотношение полов . Издательство Принстонского университета. п. 59. ИСБН 978-0-691-00981-0 .
- ^ Булл Джей-Джей (1983). Эволюция механизмов определения пола . Издательская компания Benjamin/Cummings, Продвинутая книжная программа. п. 17. ISBN 0-8053-0400-2 .
- ^ Тирио-Кьеврё С (2003). «Достижения хромосомных исследований брюхоногих моллюсков» . Журнал исследований моллюсков . 69 (3): 187–202. дои : 10.1093/моллюс/69.3.187 .
- ^ Ёсимура А (2005). «Кариотипы двух американских полевых сверчков: Gryllus Rubens и Gryllus sp. (Orthoptera: Gryllidae)». Энтомологическая наука . 8 (3): 219–222. дои : 10.1111/j.1479-8298.2005.00118.x . S2CID 84908090 .
- ^ Мейер Б.Дж. (1997). «Определение пола и компенсация дозы Х-хромосомы: половой диморфизм» . В Riddle DL, Блюменталь Т., Мейер Б.Дж., Присс-младший (ред.). К. Элеганс II . Лабораторный пресс Колд-Спринг-Харбор. ISBN 978-0-87969-532-3 . Архивировано из оригинала 6 мая 2021 года . Проверено 17 апреля 2021 г.
- ^ Де Принс Дж., Сайто К. (2003). «Кариология и определение пола» . Кристенсен Н. (ред.). Handbuch Der Zoologie / Справочник по зоологии . Том. Членистоногие: Насекомые: чешуекрылые, мотыльки и бабочки. Вальтер де Грюйтер. дои : 10.1515/9783110893724.449 . ISBN 978-3-11-016210-3 . Проверено 29 сентября 2020 г. - через Google Книги.
- ^ Янзен Ф.Дж., ПК Phillips (ноябрь 2006 г.). «Изучение эволюции определения пола в окружающей среде, особенно у рептилий». Журнал эволюционной биологии . 19 (6): 1775–1784. дои : 10.1111/j.1420-9101.2006.01138.x . ПМИД 17040374 .
- ^ Фуско Дж., Минелли А. (2019). Биология размножения . Издательство Кембриджского университета. п. 124. ИСБН 978-1-108-49985-9 .
- ^ Тодд Э.В., Лю Х., Манкастер С., Джеммелл, Нью-Джерси (2016). «Изменение пола: биология естественной смены пола у рыб» . Сексуальное развитие . 10 (5–6): 223–2241. дои : 10.1159/000449297 . hdl : 10536/DRO/DU:30153787 . ПМИД 27820936 . S2CID 41652893 .
- ^ Гонсалес Э.Дж., Мартинес-Лопес М., Моралес-Гардуза М.А., Гарсиа-Моралес Р., Чарруау П., Галлардо-Крус Х.А. (сентябрь 2019 г.). «Модель определения пола у крокодилов: систематический обзор трех десятилетий исследований». Журнал экологии животных . 88 (9): 1417–1427. Бибкод : 2019JAnEc..88.1417G . дои : 10.1111/1365-2656.13037 . ПМИД 31286510 .
- ^ Гилберт С.Ф. (2000). «Определение пола в условиях окружающей среды» . Биология развития. 6-е издание . Архивировано из оригинала 12 июня 2021 года . Проверено 19 мая 2021 г.
- ^ Чарльзворт Б. (август 2003 г.). «Определение пола у пчелы» . Клетка . 114 (4): 397–398. дои : 10.1016/S0092-8674(03)00610-X . ПМИД 12941267 .
- ^ де ла Филия А., Бейн С., Росс Л. (июнь 2015 г.). «Гаплодиплоидия и репродуктивная экология членистоногих» (PDF) . Современное мнение в области науки о насекомых . 9 : 36–43. Бибкод : 2015COIS....9...36D . дои : 10.1016/j.cois.2015.04.018 . hdl : 20.500.11820/b540f12f-846d-4a5a-9120-7b2c45615be6 . ПМИД 32846706 . S2CID 83988416 . Архивировано (PDF) оригинала 25 июня 2021 года . Проверено 25 июня 2021 г.
- ^ Фишер Р.А. (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон Пресс. стр. 141–143 - через Интернет-архив .
- ^ Гамильтон У.Д. (апрель 1967 г.). «Необычайное соотношение полов. Теория соотношения полов для связи полов и инбридинга имеет новые последствия в цитогенетике и энтомологии». Наука . 156 (3774): 477–488. Бибкод : 1967Sci...156..477H . дои : 10.1126/science.156.3774.477 . JSTOR 1721222 . ПМИД 6021675 .
- ^ Кобаяши К., Хасегава Э., Ямамото Ю., Кавацу К., Варго Э.Л., Ёсимура Дж. и др. (2013). «Предвзятость соотношения полов у термитов свидетельствует о родственном отборе» . Природные коммуникации . 4 : 2048. Бибкод : 2013NatCo...4.2048K . дои : 10.1038/ncomms3048 . hdl : 2123/11211 . ПМИД 23807025 .
- ^ «Анализ тенденций соотношения полов при рождении в США» (PDF) . Министерство здравоохранения и социальных служб США, Национальный центр статистики здравоохранения.
- ^ Дэвис Д.Л., Готлиб М.Б., Стампницкий-младший (апрель 1998 г.). «Снижение соотношения рождаемости мужского и женского пола в нескольких промышленно развитых странах: дозорный показатель здоровья?». ДЖАМА . 279 (13): 1018–1023. дои : 10.1001/jama.279.13.1018 . ПМИД 9533502 .
- ^ Перейти обратно: а б «Полевой список — соотношение полов» . Справочник ЦРУ . Центральное разведывательное управление США . Проверено 18 апреля 2024 г. (оценка на 2023 год)
- ^ Уитфилд Дж. (июнь 2004 г.). «Все, что вы всегда хотели знать о полах» . ПЛОС Биология . 2 (6): е183. дои : 10.1371/journal.pbio.0020183 . ПМК 423151 . ПМИД 15208728 .
В одном биологи сходятся во мнении: мужчины и женщины считаются представителями разных полов. И они также согласны с тем, что основная разница между ними заключается в размере гамет: самцы производят много мелких гамет — сперматозоидов у животных и пыльцы у растений, — а самки производят несколько крупных яиц.
- ^ Пирс Б.А. (2012). Генетика: концептуальный подход . У. Х. Фриман. п. 74. ИСБН 978-1-4292-3252-4 .
- ^ Докинз Р. (2016). Эгоистичный ген . Издательство Оксфордского университета. стр. 183–184. ISBN 978-0-19-878860-7 .
Однако есть одна фундаментальная особенность полов, которая может быть использована для обозначения самцов как самцов, а самок как самок у животных и растений. Дело в том, что половые клетки или «гаметы» самцов намного меньше и многочисленнее, чем гаметы самок. Это справедливо независимо от того, имеем ли мы дело с животными или растениями. У одной группы особей половые клетки крупные, и для них удобно использовать слово «самка». Другая группа, которую удобно называть мужскими, имеет мелкие половые клетки. Разница особенно заметна у рептилий и птиц, у которых одна яйцеклетка достаточно велика и питательна, чтобы кормить развивающегося детеныша. Даже у человека, у которого яйцеклетка микроскопическая, она во много раз больше сперматозоида. Как мы увидим, все остальные различия между полами можно интерпретировать как вытекающие из этого основного различия.
- ^ Робин В.В., Синха А., Рамакришнан У (2011). «Определение пола мономорфной, находящейся под угрозой исчезновения, эндемической воробьиной на небесных островах южной Индии с использованием молекулярных и морфометрических методов» . Современная наука . 101 (5): 676–679. ISSN 0011-3891 . JSTOR 24078632 .
Однако многие виды птиц мономорфны, и их трудно определить визуально, особенно в поле, а некоторые даже в руках. Некоторыми примерами являются холмовая майна, Gracula religiosa и черношапочная гаичка, Parus atricapillus .
- ^ Перейти обратно: а б с Чой Дж (2019). «Размер тела и половой диморфизм» . В Кокс Р. (ред.). Энциклопедия поведения животных . Том. 2. Академическая пресса. стр. 7–11. ISBN 978-0-12-813252-4 .
- ^ Перейти обратно: а б Мори Э, Мацца Г, Ловари С (2017). «Половой диморфизм» . В Вонк Дж, Шакелфорд Т. (ред.). Энциклопедия познания и поведения животных . Чам: Международное издательство Springer. стр. 1–7. дои : 10.1007/978-3-319-47829-6_433-1 . ISBN 978-3-319-47829-6 . Проверено 5 июня 2021 г.
- ^ Уилкинсон Г.С., Рейлло П.Р. (22 января 1994 г.). «Реакция женского выбора на искусственный отбор по преувеличенному мужскому признаку у стебельчатой мухи» (PDF) . Труды Королевского общества Б. 225 (1342): 1–6. Бибкод : 1994RSPSB.255....1W . CiteSeerX 10.1.1.574.2822 . дои : 10.1098/rspb.1994.0001 . S2CID 5769457 . Архивировано из оригинала (PDF) 10 сентября 2006 года.
- ^ Дрис Б.М., Джекман Дж (1999). «Южный паук-черная вдова» . Полевое руководство по насекомым Техаса . Хьюстон, Техас: Издательство Gulf Publishing. Архивировано из оригинала 31 августа 2003 года . Проверено 8 августа 2012 г. - через Extension Entomology, Insects.tamu.edu, Техасский университет A&M.
- ^ Стюарт-Смит Дж., Суэйн Р., Стюарт-Смит Р., Вапстра Э. (2007). «Является ли плодовитость основной причиной диморфизма размеров у ящериц-драконов, обусловленного самками?». Журнал зоологии . 273 (3): 266–272. дои : 10.1111/j.1469-7998.2007.00324.x .
- ^ Шоу Эй Джей (2000). «Популяционная экология, популяционная генетика и микроэволюция». В Шоу А.Дж., Гоффине Б. (ред.). Биология мохообразных . Кембридж: Издательство Кембриджского университета. стр. 379–380. ISBN 978-0-521-66097-6 .
- ^ Перейти обратно: а б Шустер Р.М. (1984). «Сравнительная анатомия и морфология печени». Новое руководство по бриологии . Том. 2. Нитинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори. п. 891.
- ^ Крам Х.А., Андерсон Л.Е. (1980). Мхи восточной части Северной Америки . Том. 1. Нью-Йорк: Издательство Колумбийского университета. п. 196. ИСБН 978-0-231-04516-2 .
- ^ Бриггс Д.А. (1965). «Экспериментальная систематика некоторых британских видов рода Dicranum » . Новый фитолог . 64 (3): 366–386. дои : 10.1111/j.1469-8137.1965.tb07546.x .
- ^ Захави А , Захави А (1997). Принцип гандикапа: недостающая часть головоломки Дарвина . Издательство Оксфордского университета. ISBN 978-0-19-510035-8 .
- ^ Климан Р. (2016). Херридж Э.Дж., Мюррей Р.Л., Гвинн Д.Т., Бюссьер Л. (ред.). Энциклопедия эволюционной биологии . Том. 2. Академическая пресса. стр. 453–454. ISBN 978-0-12-800426-5 .
- ^ Хеншоу Дж. М., Фромхейдж Л., Джонс АГ (август 2019 г.). «Половые роли и эволюция специализации родительской заботы» . Слушания. Биологические науки . 286 (1909): 20191312. doi : 10.1098/rspb.2019.1312 . ПМК 6732396 . ПМИД 31455191 .
- ^ Бреннан П. (2010). «Половой отбор | Изучайте науку в Scitable» . Знания о природном образовании . 3 (10): 79. Архивировано из оригинала 9 октября 2021 года . Проверено 25 июля 2021 г.
- ^ Маседо Р.Х., Маника LT (2019). «Системы полового отбора и спаривания: вклад неотропической модели воробьинообразных». В Ребореда Дж.К., Фиорини В.Д., Туеро Д.Т. (ред.). Поведенческая экология неотропических птиц . Спрингер. п. 75. ИСБН 978-3-030-14280-3 .
- ^ Удри-младший (ноябрь 1994 г.). «Природа гендера» (PDF) . Демография . 31 (4): 561–573. дои : 10.2307/2061790 . JSTOR 2061790 . ПМИД 7890091 . Архивировано (PDF) из оригинала 11 декабря 2016 года.
- ^ Хейг Д. (апрель 2004 г.). «Неумолимый рост гендера и упадок пола: социальные изменения в академических званиях, 1945–2001» (PDF) . Архив сексуального поведения . 33 (2): 87–96. CiteSeerX 10.1.1.359.9143 . дои : 10.1023/B:ASEB.0000014323.56281.0d . ПМИД 15146141 . S2CID 7005542 . Архивировано из оригинала (PDF) 25 мая 2011 года.
- ^ Принс, Вирджиния. 2005. «Секс против гендера». Международный журнал трансгендеризма . 8(4).
- ^ Нил Р., Карлсон (2010). Психология: наука о поведении. Четвертое канадское издание . Пирсон . стр. 140–141. ISBN 978-0205702862 .
- ^ «Гендер и генетика» . ВОЗ . Архивировано из оригинала 11 ноября 2012 года . Проверено 31 июля 2020 г.
- ^ «Секс и гендер» . Управление исследований женского здоровья . Архивировано из оригинала 23 июля 2020 года . Проверено 31 июля 2020 г.
- ^ Киммел М.С. (2017). Гендерное общество (Шестое изд.). Нью-Йорк: Издательство Оксфордского университета. п. 3. ISBN 978-0-19-026031-6 . OCLC 949553050 .
- ^ Линдси, Линда Л. (2010). «Глава 1. Социология гендера» (PDF) . Гендерные роли: социологический взгляд . Пирсон. ISBN 978-0-13-244830-7 . Архивировано из оригинала (PDF) 5 апреля 2015 года.
- ^ Киммел 2017 , с. 3.
- ^ Палуди М.А. (2008). Психология женщин на работе: проблемы и решения для нашей женской рабочей силы . АВС-КЛИО. п. 153. ИСБН 978-0-275-99677-2 . Архивировано из оригинала 20 октября 2021 года . Проверено 6 сентября 2021 г.
- ^ О'Халлоран К. (2020). Сексуальная ориентация, гендерная идентичность и международное право прав человека: перспективы общего права . Лондон. стр. 22–28, 328–329. ISBN 978-0-429-44265-0 . OCLC 1110674742 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ «Гендер: определения» . www.euro.who.int . Архивировано из оригинала 25 сентября 2021 года . Проверено 22 августа 2021 г.
Дальнейшее чтение
- Арнквист Г., Роу Л. (2005). Сексуальный конфликт . Издательство Принстонского университета. ISBN 978-0-691-12217-5 .
- Эллис Х (1933). Психология секса . Лондон: Медицинские книги В. Хайнемана. NB .: Одна из многих книг этого авторитета-новатора по аспектам человеческой сексуальности.
- Гилберт С.Ф. (2000). Биология развития (6-е изд.). Sinauer Associates, Inc. ISBN 978-0-87893-243-6 .
- Мейнард-Смит Дж (1978). Эволюция секса . Издательство Кембриджского университета. ISBN 978-0-521-29302-0 .
- Отто С (2008). «Половое размножение и эволюция секса | Изучайте науку в Scitable» . Природное образование . 1 (1): 182. Архивировано из оригинала 8 октября 2023 года.
Внешние ссылки
- Сизоненко ПК. «Половая дифференциация человека» . Женевский фонд медицинского образования и исследований (GFMER) . Архивировано из оригинала 9 февраля 2010 года.
Очерк человеческой сексуальности |
|---|
| Базы данных органов управления : Национальные |
|---|