Секс
| Часть серии на |
| Секс |
|---|
 |
| Биологические термины |
| Сексуальное размножение |
| Сексуальность |
| Сексуальная система |
Секс - это биологическая черта , которая определяет, ли сексуально воспроизводящий производит организм мужского или женского гамета . [ 1 ] [ 2 ] [ 3 ] [ 4 ] Во время сексуального воспроизведения мужчина и женская гамета сформируют зиготу , которая превращается в потомство , которое наследует признаки от каждого родителя. По соглашению, организмы , которые производят меньшие, более мобильные гаметы ( сперматозоиды , сперма ), называются мужчинами , в то время как организмы, которые производят более крупные, не-мобильные гаметы ( OVA , часто называемые яичными клетками), называются женщинами . [ 5 ] Организм, который производит оба типа гамете, - это гермафродит . [ 3 ] [ 6 ]
У нехимафродитных видов пол человека определяется через одну из нескольких биологических систем определения пола . Большинство видов млекопитающих имеют систему определения пола XY , где мужчина обычно несет x и y-хромосому (xy), а самка обычно несет две х хромосомы (xx). Другие системы хромосомного определения пола у животных включают систему ZW у птиц и систему XO у некоторых насекомых. [ 7 ] Различные экологические системы включают определение пола, зависящего от температуры, у рептилий и ракообразных. [ 8 ]
Мужчина и женщина вида могут быть физически как (половой мономорфизм) или иметь физические различия ( половой диморфизм ). У сексуально диморфных видов, в том числе большинство птиц и млекопитающих, пол человека обычно идентифицируется этого человека посредством наблюдения за сексуальными характеристиками . Сексуальный отбор или выбор партнера могут ускорить эволюцию различий между полами.
Термины мужчины и женщины, как правило, не применяются в сексуально недифференцированных видах, в которых люди изоморфны (выглядят так же), а гаметы изогамны (неразличимы по размеру и форме), такие как зеленая водоросли Ulva Lactuca . Некоторые виды функциональных различий между людьми, например, в грибах , [ 9 ] Может упоминаться как типы спаривания . [ 10 ]
Сексуальное размножение
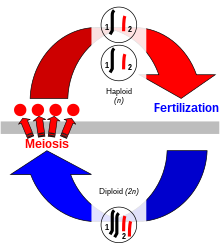
Сексуальное воспроизведение, при котором два человека производят потомство, которое обладает выбором генетических признаков каждого родителя, является исключительным для эукариот . Генетические признаки кодируются в дезоксирибонуклеиновой кислоте (ДНК) хромосом . Клетка эукариота имеет набор парных гомологичных хромосом , по одной от каждого родителя, и эта стадия двойной хромосомы называется « диплоидом ». Во время сексуального размножения диплоидный организм производит специализированные гаплоидные половые клетки, называемые гаметами через мейоз , [ 11 ] каждый из которых имеет один набор хромосом. Мейоз включает в себя стадию генетической рекомбинации через хромосомный кроссовер , в которой области ДНК обмениваются между соответствующими парами хромосом, образуя новые хромосомы, каждый с новой комбинацией генов родителей. Затем хромосомы разделены на отдельные наборы в гаметах. Когда гаметы сливаются во время оплодотворения, в результате зиготы есть половина генетического материала матери и половины отца. [ 12 ] Комбинация хромосомного кроссовера и оплодотворения , объединяя два отдельных набора хромосом вместе для создания новой диплоидной зиготы , приводит к новому организму, который содержит различный набор генетических признаков каждого родителя.
У животных гаплоидная стадия происходит только в гаметах, половые клетки, которые сливаются, чтобы сформировать зиготу, которая развивается непосредственно в новый диплоидный организм. У вида растений диплоидный организм производит тип гаплоидной споры путем мейоза, который способен подвергать повторному делению клеток для получения многоклеточного гаплоидного организма. В любом случае гаметы могут быть внешне одинаковыми ( изогамия ), как в зеленой водоросли Ulva или могут отличаться по размеру и другим аспектам ( анизогамия ). [ 13 ] Разница в размерах является наибольшей в oogamy , типе анизогамии, в которой небольшая подвижная гамета сочетается с гораздо большей, не мотильной гаметой. [ 14 ]
В анизогамических организмах, по соглашению, более крупная гамета (называемая яйцеклеткой или яичной клеткой) считается женской, а меньшая гамета (называемая сперматозоином или сперматозоидами), считается мужчинами. Человек, который производит большие гаметы, является женской, а тот, который производит небольшие гаметы, - это мужчина. [ 15 ] Человек, который производит оба типа гамете, является гермафродитом . У некоторых видов гермафродит может самостоятельно самостоятельно и производить потомство самостоятельно.
Животные

Большинство сексуально воспроизводящих животных проводят свою жизнь как диплоидную, а гаплоидная стадия уменьшена до одноклеточных гамет. [ 16 ] Гаметы животных имеют мужские и женские формы - сперматозоиды и яичные клетки соответственно. Эти гамет сочетаются в формировании эмбрионов , которые превращаются в новые организмы.
Мужская гамета, сперматозон (произведенный у позвоночных в яичках ) представляет собой небольшую ячейку, содержащую один длинный жгутичок , который его продвигает. [ 17 ] Сперматозоиды являются чрезвычайно пониженными клетками, в которых отсутствуют многие клеточные компоненты, которые были бы необходимы для эмбрионального развития. Они специализируются на подвижности, ищут яичную клетку и сливаются с ним в процессе, называемом оплодотворением .
Женские гаметы - яичные клетки. У позвоночных они производятся в яичниках . Это большие, неподвижные клетки, которые содержат питательные вещества и клеточные компоненты, необходимые для развивающегося эмбриона. [ 18 ] Яичные клетки часто ассоциируются с другими клетками, которые поддерживают развитие эмбриона, образуя яйцо . У млекопитающих эмбрион вместо этого развивается у женщин, получая питание непосредственно от матери.
Животные обычно являются мобильными и ищут партнера противоположного пола для спаривания . Животные, которые живут в воде, могут спариваться, используя внешнее оплодотворение , где яйца и сперма высвобождаются и объединяются в окружающую воду. [ 19 ] Большинство животных, которые живут вне воды, однако, используют внутреннее оплодотворение , перенося сперму непосредственно в самку, чтобы предотвратить высыхание гамет.
У большинства птиц как экскреция, так и воспроизводство выполняются с помощью одного заднего отверстия, называемого Cloaca - Малыми и Женщины -птицами касаются Cloaca, чтобы перенести сперму, процесс, называемый «Cloacal Kissing». [ 20 ] У многих других наземных животных мужчины используют специализированные половые органы , чтобы помочь транспортировке спермы - эти мужские половые органы называют индуцитационными органами . У людей и других млекопитающих этот мужской орган известен как пенис , который входит в женский репродуктивный тракт (называемый влагалищем ) для достижения осеменения - процесса, называемого половым актом . Пенис содержит трубку, через которую путешествует сперма (жидкость, содержащая сперму). У женских млекопитающих влагалище соединяется с маткой , органом, который непосредственно поддерживает развитие оплодотворенного эмбриона внутри (процесс, называемый беременностью ).
Из -за их подвижности сексуальное поведение животных может включать принудительный пол. Например, травматическое оплодотворение используется некоторыми видами насекомых для осеменения женщин через рану в брюшной полости - процесс, наносящий ущерб здоровью самки.
Растения
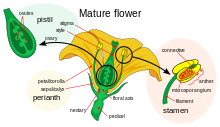
Как и животные, на наземных растениях есть специализированные мужские и женские гаметы. [ 21 ] [ 22 ] У семенных растений мужские гаметы производятся с помощью сниженных мужских гаметофитов , которые содержатся в пыльце , которые имеют твердые пальто, которые защищают мужские гамете, образующие клетки во время транспортировки от пыльников к стигме . Женские гаметы семянных растений содержатся в яйцеклетке . После оплодотворения они образуют семена , которые, как и яйца, содержат питательные вещества, необходимые для начального развития эмбрионального растения.

Цветы . цветущих растений содержат свои половые органы Большинство цветущих растений являются гермафродитными, с мужскими и женскими частями в одном и том же цвете или на одном растении в однополых цветах, около 5% видов растений имеют отдельные растения, которые являются одним полом или другим. [ 23 ] Женские части, в центре гермафродитического или женского цветка, представляют собой плистики , каждый блок, состоящий из ковра , стиля и стигмы . Два или более из этих репродуктивных единиц могут быть объединены, чтобы сформировать единый составной песник , плавные ковры образуют яичник . Внутри ковры находятся яйцеклеты , которые развиваются в семена после оплодотворения. Мужские части цветка - это тычинки : они состоят из длинных нитей, расположенных между плистом и лепестками, которые производят пыльцу в пыльниках на их кончиках. Когда пыльцевое зерно приземляется на стигму на вершине стиля карпеля, он пробуждается производить пыльцевую трубку , которая растет через ткани стиля в ковр, где он доставляет ядра мужского гамета, чтобы оплодотворить яйцу, которая в конечном итоге превращается в семя.
Некоторые гермафродитные растения являются самоопределенными, но растения развивали множественные различные механизмы самообслуживания , чтобы избежать самооплодотворения, включающего последовательный гермафродитизм , системы молекулярного распознавания и морфологические механизмы, такие как гетеростильные . [ 24 ] : 73, 74
У сосен и других хвойных деревьев половые органы производятся в конусах , которые имеют мужские и женские формы. Мужские конусы меньше, чем женщины и производят пыльцу, которая транспортируется ветром в землю в женских конусах. Большие и долгоживущие женские конусы, как правило, более долговечны и содержат яйцеклетки внутри них, которые развиваются в семена после оплодотворения.
Поскольку растения семян неподвижны, они зависят от пассивных методов транспортировки пыльцевых зерен в другие растения. Многие, включая хвойные деревья и травы, производят легкую пыльцу, которая переносится ветром в соседние растения. Некоторые цветущие растения имеют более тяжелую, липкую пыльцу, специализирующуюся на транспортировке насекомыми или более крупными животными, такими как колибри и летучих мышей , которые могут быть привлечены к цветам, содержащим награды нектара и пыльцы. Эти животные транспортируют пыльцу, когда они переходят к другим цветам, которые также содержат женские репродуктивные органы, что приводит к опылению .
Грибы

Большинство видов гриба могут воспроизводить сексуально и иметь жизненные циклы с гаплоидными и диплоидными фазами. Эти виды гриба обычно являются изогамными , т.е. отсутствует специализация мужского и женского пола. Один гаплоидный гриб превращается в контакт с другим, а затем они объединяют свои клетки. В некоторых случаях слияние является асимметричным, и клетка, которая жертвует только ядро (и отсутствие сопровождающего клеточного материала), возможно, можно считать мужчиной. [ 25 ] Грибы также могут иметь более сложные системы аллелейного спаривания, причем другие половые полы не точно описаны как мужские, женские или гермафродитные. [ 26 ]
Некоторые грибы, в том числе дрожжи пекаря , имеют типы спаривания , которые определяют совместимость. Дрожжи с одинаковыми типами спаривания не будут объединяться друг с другом, образуя диплоидные клетки, только с дрожжами, несущими другой тип спаривания. [ 27 ]
Многие виды более высоких грибов производят грибы в рамках их сексуального размножения . В грибах образуются диплоидные клетки, а затем делятся на гаплоидные споры .
Сексуальные системы
Сексуальная система - это распределение мужских и женских функций по организмам у вида. [ 28 ]
Животные
Приблизительно 95% видов животных имеют отдельные мужчины и женщины, и, как говорят, гонохорические . Около 5% видов животных являются гермафродитными. [ 28 ] Этот низкий процент частично связан с очень большим количеством видов насекомых , в которых гермафродитизм отсутствует. [ 29 ] Около 99% позвоночных гонохориальны, а оставшиеся 1% - гермафродиты - почти все рыбы. [ 30 ]
Растения
Большинство растений бисексуалы , [ 31 ] : 212 Либо гермафродит (как с тычинками, так и с плистом в одном цвете), либо однодом . [ 32 ] [ 33 ] У диологичных видов мужчины и женские полов находятся на отдельных растениях. [ 34 ] Около 5% цветущих растений являются диологическими, в результате чего 5000 независимых источников. [ 35 ] Dioecy распространена в гимноселках , в которых около 65% видов являются диологическими, но большинство хвойных деревьев однодом. [ 36 ]
Эволюция секса
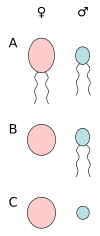
А) анизогамия подвижных клеток, б) огамия (яичная клетка и сперматозоиды), в) анизогамия немотильных клеток (яичные клетки и спермации).
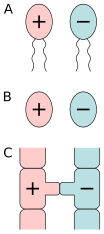
А) изогамия подвижных клеток , б) изогамия немотильных клеток, в) конъюгации.
Общепринято, что изогамия была наследственной для анизогамии [ 37 ] и эта анизогамия развивалась несколько раз независимо в разных группах эукариот, включая протисты, водоросли, растения и животных. [ 29 ] Эволюция анизогамии является синонимом происхождения мужчин и происхождения женщин . [ 38 ] Это также первый шаг к сексуальному диморфизму [ 39 ] и повлиял на эволюцию различных половых различий. [ 40 ]
Неясно, привела ли анизогамия к эволюции гермафродитизма или эволюции гонохоризма , [ 31 ] : 213 И эволюция спермы и яиц не оставила ископаемых доказательств. [ 41 ]
1,2 миллиарда лет окаменелости из Bangiomorpha pubescens предоставила самую старую ископаемость для дифференциации репродуктивных типов мужского и женского пола и показало, что полы развивались на ранних стадиях эукариот. [ 42 ] Исследования на зеленых водорослях предоставили генетические доказательства эволюционной связи между полами и типами спаривания . [ 43 ]
Первоначальной формой секса была внешняя оплодотворение . Внутреннее оплодотворение или пол, как мы его знаем, развивались позже [ 44 ] и стал доминирующим для позвоночных после их появления на земле . [ 45 ]
Адаптивная функция секса
Самым основной ролью мейоза является сохранение целостности генома , который передается родителям. [ 46 ] [ 47 ] Два наиболее фундаментальных аспекта сексуального размножения , мейотическая рекомбинация и скрещивание , вероятно, поддерживаются соответственно в результате адаптивных преимуществ рекомбинационного восстановления геномного повреждения ДНК и генетической комплементации , которые маскируют экспрессию вредных рецессивных мутаций . [ 48 ] Генетические различия , часто производимые в качестве побочного продукта этих процессов, могут обеспечить долгосрочные преимущества в тех сексуальных линиях, которые благоприятствуют скрещиванию . [ 48 ]
Системы определения пола

Биологическая причина организма, превращающегося в один пол или другой, называется определением пола . Причиной может быть генетическая, экологическая, гаплодиплоидия или множество факторов. [ 29 ] У животных и других организмов, которые имеют генетические системы определения пола, определяющим фактором может быть наличие половой хромосомы . В растениях, которые сексуально диморфны, такие как Гинкго Билоба , [ 24 ] : 203 Печеночная полиморфа виды или диологические . [ 49 ] Негенетические системы могут использовать экологические сигналы, такие как температура во время раннего развития в крокодилах , для определения пола потомства. [ 50 ]
Определение пола часто отличается от дифференциации пола . Определение пола является обозначением стадии развития к мужчинам или женщинам, в то время как дифференциация пола - это путь к развитию фенотипа . [ 51 ]
Генетический
XY SEX Определение

Люди и большинство других млекопитающих имеют систему определения пола XY : Y-хромосома в основном на основе наличия или отсутствия хромосомы Y. несет факторы, ответственные за запуск мужского пола, что делает определение пола XY , Это мужская гамета , которая определяет пол потомства. [ 52 ] В этой системе XX млекопитающие обычно являются женскими, а XY, как правило, мужчины. [ 29 ] Тем не менее, люди с XXY или XYY являются мужчинами, в то время как люди с X и XXX являются женщинами. [ 8 ] Необычно, у Плайокопа , монотримного млекопитающего, есть десять половых хромосом; У женщин есть десять х хромосомов, а у мужчин есть пять х хромосомов и пять Y -хромосомов. Все яичные клетки утконос имеют пять х хромосом, тогда как сперматозоиды могут иметь пять х хромосомов или пять лет. [ 53 ]
Определение пола XY обнаруживается в других организмах, включая насекомых, таких как обычная фруктовая муха , [ 54 ] и некоторые растения. [ 55 ] В некоторых случаях именно количество х хромосом определяет пол, а не наличие хромосомы. [ 8 ] В фруктовых мухах люди с XY находятся мужчины, а люди с XX - женщины; Тем не менее, люди с XXY или XXX также могут быть женщинами, а люди с X могут быть мужчин. [ 56 ]
Zw sex определение
У птиц, которые имеют систему определения пола ZW , w-хромосома несет факторы, ответственные за развитие женщин, а развитие по умолчанию-мужчина. [ 57 ] В этом случае люди ZZ являются мужчинами, а ZW - женщины. Это женская гамета определяет пол потомства. Эта система используется птицами, некоторыми рыбами и некоторыми ракообразными . [ 8 ]
Большинство бабочек и мотыльков также имеют систему определения секса ZW. У женщин могут быть Z, ZZW и даже ZZWW. [ 58 ]
Xo sex определение
В системе определения пола XO у мужчин есть одна X-хромосома (XO), в то время как женщины имеют два (xx). Все остальные хромосомы в этих диплоидных организмах в паре, но организмы могут наследовать одну или две х хромосомы. Эта система обнаруживается в большинстве арахнидов , насекомых, таких как серебряные рыбы ( apterygota ), дракозоны ( paleoptera ) и кузнечики ( Exopterygota ), а также некоторые нематоды, ракообразные и гастроподы. [ 59 ] [ 60 ]
в полевых сверчках насекомые с одной X -хромосомой развиваются как мужчины, а люди с двумя развиваются как женщины. Например, [ 61 ]
У нематодного каенорхабдита Elegans большинство червей представляют собой самоопределяющие гермафродиты с XX кариотипом, но случайные аномалии наследования хромосомы могут привести к людям только с одной X-хромосомой-эти люди XO являются плодородными самцами (а половина их отпрыска-мужчина). [ 62 ]
ZO SEX Определение
В системе определения пола Зо у мужчин имеют две Z-хромосомы, тогда как женщины имеют один. Эта система встречается у нескольких видов мотыльков. [ 63 ]
Относящийся к окружающей среде
Для многих видов пол определяется не унаследованными признаками, а вместо этого факторами окружающей среды, такими как температура, испытываемая во время развития или более поздней жизни. [ 64 ]
У Fern Ceratopteris и других гомоспористых видов папоротников по умолчанию пол - гермафродит, но люди, которые растут в почве, которая ранее поддерживала гермафродиты, подвержены влиянию феромона, антеридиогена чтобы развиваться как мужчина. [ 49 ] могут Личинки Bonelliidae развиваться только как мужчины, когда они сталкиваются с женщиной. [ 29 ]
Последовательный гермафродитизм

Некоторые виды могут изменить секс на протяжении всей своей жизни, явление, называемое последовательным гермафродитизмом . [ 65 ]
Телеостры рыб - единственная линия позвоночных , где происходит последовательный гермафродитизм. У клоун рыбы меньшие рыбы - мужчина, а доминирующая и самая большая рыба в группе становится женщиной; Когда доминирующая женщина отсутствует, ее партнер меняет секс с мужчины на женщины. Во многих развевании верно изначально: рыба изначально женщина и становятся мужчинами, когда достигают определенного размера. [ 66 ]
Последовательный гермафродитизм также встречается в таких растениях, как Arisaema Triphyllum .
Определение пола-зависимого пола

Многие рептилии , в том числе все крокодилы и большинство черепах , имеют определение пола в зависимости от температуры . У этих видов температура, испытываемая эмбрионами во время их развития, определяет их пол. [ 29 ]
Например, у некоторых черепах мужчин производится при более низких температурах, чем женщины; Но самок макрокламии продуцируется при температуре ниже 22 ° C или выше 28 ° C, в то время как мужчины продуцируются между этими температурами. [ 68 ]
Гаплодиплоиды
Некоторые насекомые, такие как медоносные пчелы и муравьи , используют галлодиплоидную систему определения пола . [ 69 ] Диплоидные пчелы и муравьи, как правило, являются женскими, а гаплоидные люди (которые развиваются из неоплощенных яиц) мужчины. Эта система определения пола приводит к сильно смещенным соотношениям полов , поскольку пол потомства определяется оплодотворением ( арринотоки или псевдо-архенотоки, что приводит к мужчинам), а не в ассортименте хромосом во время мейоза. [ 70 ]
Соотношение секса
Соотношение полов - это соотношение мужчин к женщинам в популяции . Как объясняется принципом Фишера , по эволюционным причинам это обычно около 1: 1 у видов, которые воспроизводят сексуально . [ 71 ] [ 72 ] Тем не менее, многие виды отклоняются от равномерного полового соотношения, периодически или навсегда. Примеры включают партеногенные виды, периодически спарившиеся организмы, такие как тля, некоторые эусоциальные осы , пчелы , муравья и термиты . [ 73 ]
Соотношение секса человека представляет особый интерес для антропологов и демографов. В обществах человека соотношение полов при рождении может быть значительно искажено такими факторами, как возраст матери при рождении [ 74 ] и селективным абортом и детоубийством. Воздействие пестицидов и других загрязнителей окружающей среды также может быть значительным фактором, способствующим. [ 75 ] По состоянию на 2024 год глобальное соотношение пола при рождении оценивается в 107 мальчиков до 100 девочек (1000 мальчиков на 934 девочки). [ 76 ] По старости соотношение пола меняется, с 81 пожилым мужчиной на каждые 100 пожилых женщин; На протяжении всех возрастов население мира почти сбалансировано, с 101 мужчиной на каждые 100 женщин. [ 76 ]Половые различия
Анизогамия является фундаментальной разницей между мужчинами и женщинами. [ 77 ] [ 78 ] Ричард Докинс заявил, что можно интерпретировать все различия между полами как вытекающие из этого. [ 79 ]
Сексуальные характеристики
Сексуальный диморфизм


У многих животных и некоторых растений люди мужского и женского пола различаются по размеру и внешнему виду, явление, называемое сексуальным диморфизмом . [ 81 ] Сексуальный диморфизм у животных часто ассоциируется с сексуальным отбором : соревнование спаривания между людьми одного пола по сравнению с противоположным полом. [ 82 ] Другие примеры демонстрируют, что именно предпочтение женщин ведет сексуальный диморфизм, например, в случае мухи с глазами . [ 83 ]
Половые различия у людей включают в себя в целом больший размер и больше волос на теле, в то время как женщины имеют большую грудь, более широкие бедра и более высокий процент жира в организме. У других видов могут быть различия в окраске или других особенностях, и могут быть настолько выражены, что различные полов могут быть приняты за два совершенно разных таксона. [ 82 ]
Женщины - более крупный пол у большинства животных. [ 81 ] Например, женщины -пауки южной черной вдовы, как правило, в два раза больше, чем у мужчин. [ 84 ] Это неравенство размера может быть связано с стоимостью производства яичных клеток, что требует большего питания, чем производство сперматозоидов: более крупные женщины способны производить больше яиц. [ 85 ] [ 81 ] Во многих других случаях мужчина вида больше, чем самка. Виды млекопитающих с диморфизмом из крайнего сексуального размера, такими как уплотнения слонов , имеют тенденцию иметь высокополигиновые системы спаривания, предположительно из -за отбора для успеха в конкуренции с другими мужчинами.
Сексуальный диморфизм может быть экстремальным, с мужчинами, такими как некоторые рыболовные рыбы , живя паразитически на женщине. Некоторые виды растений также демонстрируют диморфизм, в котором самки значительно больше, чем у мужчин, например, в роде мха Dicranum [ 86 ] и род печеночного рода Sphaerocarpos . [ 87 ] Есть некоторые доказательства того, что в этих родах диморфизм может быть связан с сексуальной хромосомой, [ 87 ] [ 88 ] или химическую передачу сигналов от женщин. [ 89 ]
У птиц мужчины часто имеют более красочный внешний вид и могут иметь особенности (например, длинный хвост мужских павлина), которые, по -видимому, ставят их в невыгодное положение (например, яркие цвета, кажется, делают птицу более видимой для хищников). Одним из предложенных объяснений для этого является принцип гандикапа . [ 90 ] Эта гипотеза утверждает, что, демонстрируя, что он может выжить с такими недостатками, мужчина рекламирует свою генетическую пригодность для женщин - простых, которые также принесут пользу дочерям, которые не будут обременены такими недостатками.
Половые различия в поведении
Полы среди гонохорических видов обычно различаются по поведению. У большинства видов животных женщины больше инвестируют в родительскую помощь, [ 91 ] Хотя у некоторых видов, таких как некоторые кукалы , мужчины инвестируют больше родительской помощи . [ 92 ] Женщины также, как правило, более разборчивы для того, с кем они спариваются, [ 93 ] такие как большинство видов птиц. [ 94 ] Мужчины, как правило, более конкурентоспособны для спаривания, чем женщины. [ 38 ]
Различие от пола
Смотрите также
Ссылки
- ^ Стивенсон А., Уэйт М. (2011). "Секс" . Краткий Оксфордский английский словарь: набор книг и компакт-дисков . УП Оксфорд . п. 1302. ISBN 978-0-19-960110-3 Полем Получено 23 марта 2018 года .
Секс: любая из двух основных категорий (мужчина и женщины), на которые люди и большинство других живых существ разделяются на основе их репродуктивных функций. Факт принадлежности к одной из этих категорий. Группа всех членов любого пола.
- ^ Mills A (1 января 2018 г.). «Секс и размножение» . Биология секса . Университет Торонто Пресс. С. 43–45. ISBN 978-1-4875-9337-7 Полем Получено 3 октября 2023 года .
- ^ Jump up to: а беременный Purves WK, Sadava DE, Orian GH , Heller HC (2000). Жизнь: наука о биологии . Макмиллан . п. 736. ISBN 978-0-7167-3873-2 Полем Получено 23 марта 2018 года .
Одно тело может функционировать как мужской и женский. Сексуальное воспроизведение требует как мужских, так и женских гаплоидных гамет. У большинства видов эти гаметы производятся людьми, которые являются мужчинами или женщинами. Виды, у которых есть мужские и женские члены, называются диологическими (от греческого языка для «двух домов»). У некоторых видов один человек может обладать как женскими, так и мужскими репродуктивными системами. Такие виды называются однодомным («один дом») или гермафродитом.
- ^ De Loof A (31 января 2018 г.). «Только две половые формы, но несколько гендерных вариантов: как объяснить?» Полем Коммуникативная и интегративная биология . 11 (1): E1427399. doi : 10.1080/194208899.2018.1427399 . ISSN 1942-0889 . PMC 5824932 . PMID 29497472 .
- ^ Кокко Х. , Дженнион М (2012). «Половые различия в родительской помощи» . В Royle NJ, Smiseth Pt, Kölliker M (Eds.). Эволюция родительской помощи . Издательство Оксфордского университета. п. 103. ISBN 978-0-19-969257-6 Полем
Ответ заключается в том, что соглашение о соглашении: люди, производящие меньшие из двух типов гаметных - сперма или пыльцы - являются мужчинами, а люди, производящие большие гаметы - яйца или яйцеклетки - являются женщинами.
- ^ Avise JC (2011). «Два полов в одном» . Гермафродитизм: учебник по биологии, экологии и эволюции двойной сексуальности . Издательство Колумбийского университета. С. 1–7. ISBN 978-0-231-52715-6 Полем Получено 18 сентября 2020 года .
- ^ Blackmon H, Ross L, Bachtrog D (январь 2017 г.). «Определение пола, половые хромосомы и эволюция кариотипа у насекомых» . Журнал наследственности . 108 (1): 78–93. doi : 10.1093/jhered/esw047 . ISSN 0022-1503 . PMC 6281344 . PMID 27543823 .
- ^ Jump up to: а беременный в дюймовый Hake L, O'Connor C (2008). «Генетические механизмы определения пола | изучать науку в Scaital» . Природное образование . 1 (1): 25. Архивировано из оригинала 19 августа 2017 года . Получено 13 апреля 2021 года .
- ^ Мур Д., Робсон Д.Д., Тринси А.П. (2020). Руководство 21 -го века по грибам (2 изд.). Издательство Кембриджского университета. С. 211–228. ISBN 978-1-108-74568-0 .
- ^ Кумар Р., Мина М., Свапнил П (2019). "Анизогамия". В Vonk J, Shackelford T (Eds.). Энциклопедия познания и поведения животных . Cham: Springer International Publishing. С. 1–5. doi : 10.1007/978-3-319-47829-6_340-1 . ISBN 978-3-319-47829-6 Полем
Анизогамия может быть определена как способ сексуального размножения, в котором слияние гамет, образованных участвующими родителями, отличаются по размеру.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). "Мейоз" . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3218-3 Полем Архивировано с оригинала 25 января 2017 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Преимущества секса» . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3218-3 .
- ^ Гилберт С.Ф. (2000). «Многоклеточность: эволюция дифференциации» . Биология развития (6 -е изд.). Sunderland (MA): Sinauer Associates. ISBN 978-0-87893-243-6 Полем Архивировано из оригинала 8 марта 2021 года . Получено 17 апреля 2021 года .
- ^ Аллаби М. (2012). Словарь растительных наук . УП Оксфорд. п. 350. ISBN 978-0-19-960057-1 .
- ^ Джи H (22 ноября 1999 г.). «Размер и одиночная половая клетка» . Природа . Архивировано из оригинала 11 октября 2017 года . Получено 4 июня 2018 года .
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Мендельская генетика в эукариотических жизненных циклах» . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3218-3 Полем Архивировано с оригинала 2 апреля 2017 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). "Сперма" . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3218-3 Полем Архивировано из оригинала 29 июня 2009 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Яйца» . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3218-3 Полем Архивировано из оригинала 29 июня 2009 года.
- ^ Альбертс Б. , Джонсон А. , Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). «Удобрение» . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3218-3 Полем Архивировано из оригинала 19 декабря 2008 года.
- ^ Ритчисон Г. "Птиевой размножение" . Восточный университет Кентукки. Архивировано из оригинала 12 апреля 2008 года . Получено 3 апреля 2008 года .
- ^ Гилберт С.Ф. (2000). «Производство гамета в покрытосеменных» . Биология развития (6 -е изд.). Sunderland (MA): Sinauer Associates. ISBN 978-0-87893-243-6 Полем Архивировано из оригинала 21 апреля 2021 года . Получено 17 апреля 2021 года .
- ^ Dusenbery DB (2009). Жизнь в микромасштабе: неожиданная физика маленькой . Гарвардский университет издательство. С. 308–326. ISBN 978-0-674-03116-6 .
- ^ Уилтон П (12 марта 2009 г.). «Растения, секс и Дарвин» . Oxsciblog . Оксфордский университет . Получено 10 января 2024 года .
- ^ Jump up to: а беременный Judd WS , Campbell CS, Kellogg EA , Stevens PF , Donoghue MJ (2002). Систематика растений, филогенетический подход (2 -е изд.). Sunderland MA: Sinauer Associates Inc. ISBN 0-87893-403-0 .
- ^ Ник Лейн (2005). Власть, пол, самоубийство: митохондрии и значение жизни . Издательство Оксфордского университета. С. 236–237 . ISBN 978-0-19-280481-5 .
- ^ Watkinson SC, Boddy L , Money N (2015). Грибы . Elsevier Science. п. 115. ISBN 978-0-12-382035-8 Полем Получено 18 февраля 2018 года .
- ^ Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). «Спецификация типа ячейки и преобразование типа спаривания у дрожжей» . Молекулярная клеточная биология (четвертое изд.). WH Freeman и Co. ISBN 978-0-7167-4366-8 Полем Архивировано из оригинала 1 июля 2009 года.
- ^ Jump up to: а беременный Леонард Дж.Л. (22 августа 2013 г.). «Парадокс Уильямса и роль фенотипической пластичности в сексуальных системах» . Интегративная и сравнительная биология . 53 (4): 671–688. doi : 10.1093/ICB/ICT088 . ISSN 1540-7063 . PMID 23970358 .
- ^ Jump up to: а беременный в дюймовый и фон Бахтрог Д., Манк Дж. , Пейхель К.Л., Киркпатрик М. , Отто С.П. , Эшман Т.Л. и др. (Июль 2014). "Определение пола: почему так много способов сделать это?" Полем PLOS Биология . 12 (7): E1001899. doi : 10.1371/journal.pbio.1001899 . PMC 4077654 . PMID 24983465 .
- ^ Кувамура Т., Суноб Т., Сакай Ю., Кадота Т., Савада К (1 июля 2020 года). «Гермафродитизм у рыб: аннотированный список видов, филогении и спаривания» . Ихтиологические исследования . 67 (3): 341–360. Bibcode : 2020ichtr..67..341k . doi : 10.1007/s10228-020-00754-6 . ISSN 1616-3915 . S2CID 218527927 .
- ^ Jump up to: а беременный Климан Р.М. (2016). Энциклопедия эволюционной биологии . Тол. 2. Академическая пресса. С. 212–224. ISBN 978-0-12-800426-5 Полем Архивировано из оригинала 6 мая 2021 года . Получено 14 апреля 2021 года .
- ^ Сабат Н., Голдберг Е.Е., Глик Л., Эйнхорн М., Ашман Т.Л., Мин Р. и др. (Февраль 2016 г.). «Dioecy не последовательно ускоряет или медленную диверсификацию линии в разных родах покрытосеменных» . Новый фитолог . 209 (3): 1290–1300. doi : 10.1111/nph.13696 . PMID 26467174 .
- ^ Beentje H (2016). Глоссарий завода Кью (2 изд.). Королевские ботанические сады, Kew: Kew Publishing. ISBN 978-1-84246-604-9 .
- ^ Leite Montalvão AP, Kersten B, Fladung M, Müller NA (2021). «Разнообразие и динамика определения пола у диологичных растений» . Границы в науке о растениях . 11 : 580488. DOI : 10.3389/fpls.2020.580488 . PMC 7843427 . PMID 33519840 .
- ^ Реннер СС (октябрь 2014 г.). «Относительные и абсолютные частоты сексуальных систем покрытосеменных: Dioecy, Monoecy, Gynodioecy и обновленная онлайн -база данных» . Американский журнал ботаники . 101 (10): 1588–1596. doi : 10.3732/ajb.1400196 . PMID 25326608 .
- ^ Walas, Mandryk W, Thomas PA, Tyrała-Wierucka ż, Iszkuło G (2018). «Сексуальные системы в гимноселках: обзор» (PDF) . Основная и прикладная экология . 31 : 1–9. Bibcode : 2018bapec..31 .... 1W . doi : 10.1016/j.baae.2018.05.009 . S2CID 90740232 . Архивировано (PDF) из оригинала 27 января 2022 года . Получено 7 июня 2021 года .
- ^ Awasthi AK. Учебник водорослей . Викас издательство. п. 363. ISBN 978-93-259-9022-7 .
- ^ Jump up to: а беременный Lehtonen J, Kokko H , Parker GA (октябрь 2016 г.). «Чему изучают изогамные организмы о сексе и двух полах?» Полем Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 371 (1706). doi : 10.1098/rstb.2015.0532 . PMC 5031617 . PMID 27619696 .
- ^ Togashi T, Bartelt JL, Yoshimura J, Tainaka K, Cox PA (август 2012 г.). «Эволюционные траектории объясняют диверсифицированную эволюцию изогамии и анизогамии в морских зеленых водорослях» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (34): 13692–13697. BIBCODE : 2012PNAS..10913692T . doi : 10.1073/pnas.1203495109 . PMC 3427103 . PMID 22869736 .
- ^ Székely T , Fairbairn DJ, Blanckenhorn Wu (2007). Пол, размер и гендерные роли: эволюционные исследования диморфизма сексуального размера . УП Оксфорд. с. 167–169, 176, 185. ISBN 978-0-19-920878-4 .
- ^ Pitnick SS, Hosken DJ, Birkhead TR (2008). Биология спермы: эволюционная перспектива . Академическая пресса. С. 43–44. ISBN 978-0-08-091987-4 .
- ^ Hörandl E, Hadacek F (август 2020 г.). «Кислород, формы жизни и эволюция полов у многоклеточных эукариот» . Наследственность . 125 (1–2): 1–14. doi : 10.1038/s41437-020-0317-9 . PMC 7413252 . PMID 32415185 .
- ^ Sawada H, Inoue N, Iwano M, Eds. (2014). Сексуальное размножение у животных и растений . Спрингер. С. 215–227. doi : 10.1007/978-4-431-54589-7 . ISBN 978-4-431-54589-7 .
- ^ Черный Р (19 октября 2014 г.). «Арбинированная рыба стала пионером, как вы его знаете» . Животные National Geographic. Архивировано из оригинала 2 марта 2021 года . Получено 10 июля 2023 года .
- ^ «43.2a: внешнее и внутреннее оплодотворение» . Биология либретекса . 17 июля 2018 года. Архивировано с оригинала 24 мая 2022 года . Получено 9 ноября 2020 года .
- ^ Брандейс М (май 2018). «Идеи нового возраста о вековом полу: отделение мейоза от спаривания может решить вековую головоломку». Биологические обзоры Кембриджского философского общества . 93 (2): 801–810. doi : 10.1111/brv.12367 . PMID 28913952 .
- ^ Hörandl E (июнь 2024 г.). «Апомиксис и парадокс секса в растениях». Анналы ботаники . 134 (1): 1–18. doi : 10.1093/aob/mcae044 . PMC 11161571. PMID 38497809 .
- ^ Jump up to: а беременный Бернштейн Х, Байерли Х.К., Хопф Ф.А., Мичод Р.Е. (сентябрь 1985). «Генетическое повреждение, мутация и эволюция пола». Наука . 229 (4719). Нью -Йорк, Нью -Йорк: 1277–81. doi : 10.1126/science.3898363 . PMID 3898363 .
- ^ Jump up to: а беременный Tanurdzic M, Banks JA (2004). «Механизмы, определяющие половые, на наземных растениях» . Растительная ячейка . 16 (Suppl): S61 - S71. doi : 10.1105/tpc.016667 . PMC 2643385 . PMID 15084718 .
- ^ Warner Da, Shine R (январь 2008 г.). «Адаптивная значимость определения пола, зависящего от температуры в рептилии». Природа . 451 (7178): 566–568. Bibcode : 2008natur.451..566W . doi : 10.1038/nature06519 . PMID 18204437 . S2CID 967516 .
- ^ Beukeboom LW, Perrin N (2014). Эволюция определения пола . Издательство Оксфордского университета. п. 16. ISBN 978-0-19-965714-8 .
- ^ Wallis MC, Waters Pd, Graves JA (октябрь 2008 г.). «Определение пола у млекопитающих - до и после эволюции SRY» . Клеточные и молекулярные науки о жизни . 65 (20): 3182–95. doi : 10.1007/s00018-008-8109-z . PMC 11131626 . PMID 18581056 . S2CID 31675679 .
- ^ Пирс Б.А. (2012). Генетика: концептуальный подход (4 -е изд.). Нью -Йорк: WH Freeman. Стр. 73–7 ISBN 978-1-4292-3250-0 Полем OCLC 703739906 .
- ^ Кайзер В.Б., Бахтрог Д. (2010). «Эволюция половых хромосом у насекомых» . Ежегодный обзор генетики . 44 : 91–112. doi : 10.1146/annurev-genet-102209-163600 . PMC 4105922 . PMID 21047257 .
- ^ Dellaporta SL, Calderon-orrerea A (октябрь 1993 г.). «Определение пола в цветущих растениях» . Растительная ячейка . 5 (10): 1241–1251. doi : 10.1105/tpc.5.10.1241 . JSTOR 3869777 . PMC 160357 . PMID 8281039 .
- ^ Fusco G, Minelli A (2019). Биология размножения . Издательство Кембриджского университета. С. 306–308. ISBN 978-1-108-49985-9 .
- ^ Смит К.А., Кац М., Синклер А.Х. (февраль 2003 г.). «DMRT1 активируется в гонадах во время реверсии пола-мужчины в эмбрионах ZW куриных эмбрионов» . Биология размножения . 68 (2): 560–570. doi : 10.1095/biolreprod.102.007294 . PMID 12533420 .
- ^ Majerus me (2003). Сексуальные войны: гены, бактерии и предвзятые половые соотношения . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. п. 59. ISBN 978-0-691-00981-0 .
- ^ Bull JJ (1983). Эволюция механизмов определения пола . Benjamin/Cummings Publishing Company, Advance Book Program. п. 17. ISBN 0-8053-0400-2 .
- ^ Thiriot-quiévreux C (2003). «Достижения в хромосомных исследованиях гастропод моллюсков» . Журнал Molluscan исследования . 69 (3): 187–202. doi : 10.1093/mollus/69.3.187 .
- ^ Йошимура А. (2005). «Кариотипы двух американских полевых сверчков: Gryllus Rubens и Gryllus sp. (Orthoptera: Gryllidae)». Энтомологическая наука . 8 (3): 219–222. doi : 10.1111/j.1479-8298.2005.00118.x . S2CID 84908090 .
- ^ Мейер Б.Дж. (1997). «Определение пола и хромосомная компенсация: половой диморфизм» . В Riddle DL, Blumenthal T, Meyer BJ, Priess Jr (Eds.). C. Elegans II . Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-532-3 Полем Архивировано из оригинала 6 мая 2021 года . Получено 17 апреля 2021 года .
- ^ De Prins J, Sheithoh K (2003). «Кариология и определение пола» . В Кристенсен N (ред.). Handbuch der Zoologie / Справочник по зоологии . Тол. CHROPODA: Insecta: Lepidoptera, мотыльки и бабочки. Уолтер де Грютер. С. 449–468. doi : 10.1515/9783110893724.449 . ISBN 978-3-11-016210-3 Полем Получено 29 сентября 2020 года - через Google Books.
- ^ Janzen FJ, Phillips PC (ноябрь 2006 г.). «Изучение эволюции определения пола окружающей среды, особенно в рептилиях». Журнал эволюционной биологии . 19 (6): 1775–1784. doi : 10.1111/j.1420-9101.2006.01138.x . PMID 17040374 .
- ^ Fusco G, Minelli A (2019). Биология размножения . Издательство Кембриджского университета. п. 124. ISBN 978-1-108-49985-9 .
- ^ Todd EV, Liu H, Muncaster S, Gemmell NJ (2016). «Изгиб полы: биология естественного пола смены рыбы» . Сексуальное развитие . 10 (5–6): 223–2241. doi : 10.1159/000449297 . HDL : 10536/dro/du: 30153787 . PMID 27820936 . S2CID 41652893 .
- ^ Garcruau-Garduza, Garcruau-Cruru-Cruz JA (сентябрь 2019 г.). Полем Экология Журнал 88 (9): 1417–1427. Bibcode : 2019 ЯНЕК doi : 10.111/1365-2656.1337 . PMID 312865510 .
- ^ Гилберт С.Ф. (2000). «Определение пола окружающей среды» . Биология развития. 6 -е издание . Архивировано из оригинала 12 июня 2021 года . Получено 19 мая 2021 года .
- ^ Чарльзворт Б (август 2003 г.). «Определение пола в пчелах» . Клетка . 114 (4): 397–398. doi : 10.1016/s0092-8674 (03) 00610-x . PMID 12941267 .
- ^ De la Filia A, Bain S, Ross L (июнь 2015 г.). «Гаплодиплоидия и репродуктивная экология членистоногих» (PDF) . Текущее мнение в науке насекомых . 9 : 36–43. Bibcode : 2015cois .... 9 ... 36d . doi : 10.1016/j.cois.2015.04.018 . HDL : 20.500.11820/B540F12F-846D-4A5A-9120-7B2C45615BE6 . PMID 32846706 . S2CID 83988416 . Архивировано (PDF) из оригинала 25 июня 2021 года . Получено 25 июня 2021 года .
- ^ Фишер Р.А. (1930). Генетическая теория естественного отбора . Оксфорд: Clarendon Press. С. 141–143 - через интернет -архив .
- ^ Гамильтон WD (апрель 1967 г.). «Чрезвычайные соотношения половых. Теория пола для сексуальной связи и инбридинга имеет новые последствия для цитогенетики и энтомологии». Наука . 156 (3774): 477–488. Bibcode : 1967sci ... 156..477H . doi : 10.1126/science.156.3774.477 . JSTOR 1721222 . PMID 6021675 .
- ^ Kobayashi K, Hasegawa E, Yamamoto Y, Kawatsu K, Vargo EL, Yoshimura J, et al. (2013). «Предвзятые отношения к соотношению полов в термитах предоставляют доказательства для отбора родственников» . Природная связь . 4 : 2048. Bibcode : 2013natco ... 4.2048K . doi : 10.1038/ncomms3048 . HDL : 2123/11211 . PMID 23807025 .
- ^ «Анализ тенденций пола при рождении в Соединенных Штатах» (PDF) . Министерство здравоохранения и социальных служб США, Национальный центр статистики здравоохранения.
- ^ Дэвис Д.Л., Готлиб М.Б., Сэмпницки младший (апрель 1998 г.). «Снижение соотношения мужчин к женщинам в нескольких промышленных странах: индикатор здоровья стражи?». Джама . 279 (13): 1018–1023. doi : 10.1001/Jama.279.13.1018 . PMID 9533502 .
- ^ Jump up to: а беременный «Полевой листинг - секс -соотношение» . ЦРУ Факт . Центральное разведывательное агентство Соединенных Штатов . Получено 18 апреля 2024 года . (Оценки 2023 г.)
- ^ Уитфилд Дж (июнь 2004 г.). «Все, что вы всегда хотели знать о полах» . PLOS Биология . 2 (6): E183. doi : 10.1371/journal.pbio.0020183 . PMC 423151 . PMID 15208728 .
Одна вещь, с которой биологи согласны, это то, что мужчины и женщины считаются разными полами. И они также согласны с тем, что основное отличие между ними - это размер гамета: мужчины делают много небольших гамет - Sperm у животных, пыльца в растениях - и женщины производят несколько больших яиц.
- ^ Пирс Б.А. (2012). Генетика: концептуальный подход . WH Freeman. п. 74. ISBN 978-1-4292-3252-4 .
- ^ Докинз Р. (2016). Эгоистичный ген . Издательство Оксфордского университета. С. 183–184. ISBN 978-0-19-878860-7 Полем
Тем не менее, существует одна фундаментальная особенность полов, которые можно использовать для маркировки мужчин как мужчин, а женщины - как женщины, по всем животным и растениям. Это то, что половые клетки или «гаметы» мужчин намного меньше и многочисленны, чем гаметы женщин. Это правда, имеем ли мы дело с животными или растениями. Одна группа людей имеет большие половые клетки, и это удобно использовать для них слово. Другая группа, которую удобно называть мужчиной, имеет небольшие половые клетки. Разница особенно выражена в рептилиях и у птиц, где одна яйцеклетка достаточно велика и достаточно питательна, чтобы накормить развивающегося ребенка. Даже у людей, где яйцо является микроскопическим, оно все еще во много раз больше, чем сперма. Как мы увидим, можно интерпретировать все другие различия между полами как вытекающие из этого основного различия.
- ^ Робин В.В., Синха А., Рамакришнан У (2011). «Определение пола мономорфного угрожающего, эндемичного прохожного прохожника на небах островах Южной Индии с использованием молекулярных и морфометрических методов» . Текущая наука . 101 (5): 676–679. ISSN 0011-3891 . JSTOR 24078632 .
Многие виды птиц, однако, являются мономорфными и трудными для визуального секса, особенно в поле, а некоторые даже в руках. Некоторые примеры-Mynah Hill Mynah, Gracula Religiosa и Чикади с черной квалой, Parus aticapillus .
- ^ Jump up to: а беременный в Choe J (2019). «Размер тела и половой диморфизм» . В Cox R (ред.). Энциклопедия поведения животных . Тол. 2. Академическая пресса. С. 7–11. ISBN 978-0-12-813252-4 .
- ^ Jump up to: а беременный Мори Е., Мацца Г., Ловари С. (2017). «Сексуальный диморфизм» . В Vonk J, Shackelford T (Eds.). Энциклопедия познания и поведения животных . Cham: Springer International Publishing. С. 1–7. doi : 10.1007/978-3-319-47829-6_433-1 . ISBN 978-3-319-47829-6 Полем Получено 5 июня 2021 года .
- ^ Wilkinson GS, Reillo PR (22 января 1994 г.). «Женский выбор ответа на искусственный отбор на преувеличенной мужской черте в мухе с ускорительными глазами» (PDF) . Труды Королевского общества б . 225 (1342): 1–6. BIBCODE : 1994RSPSB.255 .... 1W . Citeseerx 10.1.1.574.2822 . doi : 10.1098/rspb.1994.0001 . S2CID 5769457 . Архивировано из оригинала (PDF) 10 сентября 2006 года.
- ^ Drees BM, Jackman J (1999). "Южная черная вдова паук" . Полевое руководство по насекомым Техаса . Хьюстон, Техас: издательская компания Gulf. Архивировано из оригинала 31 августа 2003 года . Получено 8 августа 2012 года - с помощью энтомологии расширения, насекомых. Tamu.edu, Техасский университет A & M.
- ^ Стюарт-Смит Дж., Суэйн Р., Стюарт-Смит Р., Вапстра Е. (2007). «Является ли плодовитость конечной причиной диморфизма с женским размером в ящерице дракона?». Журнал зоологии . 273 (3): 266–272. doi : 10.1111/j.1469-7998.2007.00324.x .
- ^ Шоу А.Дж. (2000). «Экология населения, генетика населения и микроэволюция». В Шоу А.Дж., Гоффинет Б (ред.). Bryophyte Biology . Кембридж: издательство Кембриджского университета. С. 379–380. ISBN 978-0-521-66097-6 .
- ^ Jump up to: а беременный Schuster RM (1984). «Сравнительная анатомия и морфология печени». Новое руководство по браизенту . Тол. 2. Ничинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори. п. 891.
- ^ Crum HA, Anderson Le (1980). Мосс восточной Северной Америки . Тол. 1. Нью -Йорк: издательство Колумбийского университета. п. 196. ISBN 978-0-231-04516-2 .
- ^ Бриггс Д.А. (1965). «Экспериментальная таксономия некоторых британских видов рода Dicranum » . Новый фитолог . 64 (3): 366–386. doi : 10.1111/j.1469-8137.1965.tb07546.x .
- ^ Захави А. , Захави А. (1997). Принцип гандикапа: недостающий кусок головоломки Дарвина . Издательство Оксфордского университета. ISBN 978-0-19-510035-8 .
- ^ Климан Р. (2016). Herridge EJ, Murray RL, Gwynne DT, Bussiere L (Eds.). Энциклопедия эволюционной биологии . Тол. 2. Академическая пресса. С. 453–454. ISBN 978-0-12-800426-5 .
- ^ Henshaw JM, Fromhage L, Jones AG (август 2019 г.). «Секс -роли и эволюция специализации по уходу за родителями» . Разбирательство. Биологические науки . 286 (1909): 20191312. DOI : 10.1098/rspb.2019.1312 . PMC 6732396 . PMID 31455191 .
- ^ Бреннан П. (2010). «Сексуальный отбор | изучать науку в Scileble» . Знание по природе . 3 (10): 79. Архивировано с оригинала 9 октября 2021 года . Получено 25 июля 2021 года .
- ^ Macedo RH, Manica LT (2019). «Системы сексуального отбора и спаривания: вклад неотропической модели прохождения». В Recoreda JC, Fiorini VD, Tuero DT (Eds.). Поведенческая экология неотропических птиц . Спрингер. п. 75. ISBN 978-3-030-14280-3 .
- ^ Удри -младший (ноябрь 1994). «Природа пола» (PDF) . Демография . 31 (4): 561–573. doi : 10.2307/2061790 . JSTOR 2061790 . PMID 7890091 . Архивировано (PDF) из оригинала 11 декабря 2016 года.
- ^ Haig D (апрель 2004 г.). «Неоткрытый рост пола и упадок секса: социальные изменения в академических названиях, 1945–2001» (PDF) . Архивы сексуального поведения . 33 (2): 87–96. Citeseerx 10.1.1.359.9143 . doi : 10.1023/b: aseb.0000014323.56281.0d . PMID 15146141 . S2CID 7005542 . Архивировано из оригинала (PDF) 25 мая 2011 года.
- ^ Принц, Вирджиния. 2005. «Секс против пола». Международный журнал трансгендеризма . 8 (4).
- ^ Нил Р., Карлсон (2010). Психология: наука о поведении. Четвертое канадское издание . Пирсон . С. 140–141. ISBN 978-0205702862 .
- ^ «Пол и генетика» . ВОЗ . Архивировано из оригинала 11 ноября 2012 года . Получено 31 июля 2020 года .
- ^ «Секс и пол» . Управление исследований по здоровью женщин . Архивировано из оригинала 23 июля 2020 года . Получено 31 июля 2020 года .
- ^ Киммел М.С. (2017). Гендерное общество (шестое изд.). Нью -Йорк: издательство Оксфордского университета. п. 3. ISBN 978-0-19-026031-6 Полем OCLC 949553050 .
- ^ Линдси, Линда Л. (2010). «Гл. 1. Социология гендерного» (PDF) . Гендерные роли: социологическая перспектива . Пирсон. ISBN 978-0-13-244830-7 Полем Архивировано из оригинала (PDF) 5 апреля 2015 года.
- ^ Киммел 2017 , с. 3
- ^ Палуди М.А. (2008). Психология женщин на работе: проблемы и решения для нашей женской рабочей силы . ABC-Clio. п. 153. ISBN 978-0-275-99677-2 Полем Архивировано из оригинала 20 октября 2021 года . Получено 6 сентября 2021 года .
- ^ О'Халлоран К (2020). Сексуальная ориентация, гендерная идентичность и международное законодательство о правах человека: перспективы общего права . Лондон С. 22–28, 328–329. ISBN 978-0-429-44265-0 Полем OCLC 1110674742 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ «Пол: определения» . www.euro.who.int . Архивировано из оригинала 25 сентября 2021 года . Получено 22 августа 2021 года .
Дальнейшее чтение
- Arnqvist G, Rowe L (2005). Сексуальный конфликт . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-12217-5 .
- Эллис Х (1933). Психология секса . Лондон: W. Heinemann Medical Books. Nb .: Одна из многих книг этой новаторской власти по аспектам человеческой сексуальности.
- Гилберт С.Ф. (2000). Биология развития (6 -е изд.). Sinauer Associates, Inc. ISBN 978-0-87893-243-6 .
- Мейнард-Смит Дж. (1978). Эволюция секса . Издательство Кембриджского университета. ISBN 978-0-521-29302-0 .
- Отто С. (2008). «Сексуальное воспроизведение и эволюция секса | изучать науку в Scaital» . Природное образование . 1 (1): 182. Архивировано из оригинала 8 октября 2023 года.
Внешние ссылки
- Sizonenko PC. «Человеческая сексуальная дифференциация» . Женевский фонд медицинского образования и исследований (GFMER) . Архивировано из оригинала 9 февраля 2010 года.
Схема человеческой сексуальности |
|---|
| Базы данных управления авторитетом : национальный |
|---|