Мозг
| Мозг | |
|---|---|
 Мозг шимпанзе | |
| Подробности | |
| Часть | Нервная система |
| Идентификаторы | |
| латинский | головность |
| Греческий | Энцефалон |
| Сетка | D001921 |
| Нейроналы | 21 |
| TA98 | A14.1.03.001 |
| TA2 | 5415 |
| Анатомическая терминология | |
Мозг орган - это , который служит центром нервной системы во всех позвоночных и большинстве беспозвоночных животных . Он состоит из нервной ткани и обычно расположен в голове ( головность ), обычно вблизи органов для особых чувств, таких как зрение , слух и обоняние . Будучи наиболее специализированным органом, он отвечает за получение информации от сенсорной нервной системы , обработку этой информации ( мысли , познание и интеллект ) и координацию моторного контроля ( мышечная активность и эндокринная система ).
В то время как мозги беспозвоночных возникают из парных сегментарных ганглиев (каждая из которых ответственна только за соответствующий сегмент тела ) вентрального нервного шнура , мозги позвоночных развиваются в осевом направлении из средней линии дорсального нервного шнура в качестве везикулярного увеличения на росстральном конце труб нейронной с централизованным контролем над всеми сегментами тела. Все мозги позвоночных могут быть эмбрионически разделены на три части: передний мозг (Prosencephalon, подразделение на Telencephalon и Diencephalon ), средний мозг ( Mesencephalon ) и задний мозг ( Rompencephalon , подразделенный в Metencephalon и Myelencephalon ). Спинной мозг , который непосредственно взаимодействует с соматическими функциями под головкой, может рассматриваться как каудальное расширение миленсфалона, заключенного в колонку позвоночника . Вместе мозг и спинной мозг составляют центральную нервную систему у всех позвоночных.
У людей кора головного мозга содержит приблизительно 14–16 миллиардов нейронов, [ 1 ] и предполагаемое количество нейронов в мозжечке составляет 55–70 миллиардов. [ 2 ] Каждый нейрон соединен синапсами с несколькими тысячами других нейронов, обычно общаясь друг с другом посредством корнеподобных выступов, называемых дендритами и длинными волокнистыми растяжениями, называемыми аксонами , которые обычно миелинизируются и переносят поезда из быстрых импульсов микроэлектрического сигнала, называемых потенциалами действия, потенциалы действия. Чтобы нацелиться на конкретные клетки -реципиента в других областях мозга или отдаленных частей тела. Префронтальная кора , которая контролирует исполнительные функции , особенно хорошо развивается у людей.
Физиологически мозг осуществляет централизованный контроль над другими органами тела. Они действуют на остальную часть тела как путем генерации моделей мышечной активности, так и за счет секреции химических веществ, называемых гормонами . Этот централизованный контроль позволяет быстро и скоординировать ответы на изменения в окружающей среде . Некоторые основные типы отзывчивости, такие как рефлексы, могут быть опосредованы спинным мозгом или периферическими ганглиями , но сложный целеустремленный контроль над поведением, основанный на сложном сенсорном вводе, требуют возможности интеграции информации централизованного мозга.
Операции отдельных клеток мозга в настоящее время понимаются в значительной степени, но то, как они сотрудничают в ансамблях миллионов, еще предстоит решить. [ 3 ] Недавние модели в современной нейробиологии рассматривают мозг как биологический компьютер , очень отличающийся по механизму от цифрового компьютера , но сходной в том смысле, что он получает информацию от окружающего мира, хранит его и обрабатывает ее различными способами.
В этой статье сравниваются свойства мозга во всем диапазоне видов животных, с наибольшим вниманием к позвоночным. Он имеет дело с человеческим мозгом , поскольку он разделяет свойства других мозгов. То, как человеческий мозг отличается от других мозгов, рассматриваются в статье человеческого мозга. Несколько тем, которые здесь могут быть рассмотрены, вместо этого покрыты там, потому что о них можно сказать гораздо больше в человеческом контексте. Наиболее важными, которые рассматриваются в статье человека мозга, являются болезнь мозга и последствия повреждения головного мозга .
Структура

Форма и размер мозга сильно различаются между видами, и идентификация общих черт часто затруднена. [ 4 ] Тем не менее, существует ряд принципов архитектуры мозга, которые применяются для широкого спектра видов. [ 5 ] Некоторые аспекты структуры мозга являются общими практически для всего диапазона видов животных; [ 6 ] Другие отличают «продвинутые» мозги от более примитивных или отличают позвоночных от беспозвоночных. [ 4 ]
Самый простой способ получить информацию об анатомии мозга - это визуальный осмотр, но было разработано много более сложных методов. Ткань мозга в его естественном состоянии слишком мягкая, чтобы работать, но она может быть затвердевает погружением в алкоголь или другие фиксированные , а затем нарезается для изучения внутренних дел. Визуально, внутренняя часть мозга состоит из областей так называемого серого вещества , с темным цветом, разделенным областями белого вещества , с более легким цветом. Дополнительную информацию можно получить, окрашивая срезы мозговой ткани с различными химическими веществами, которые выявляют области, где специфические типы молекул присутствуют в высоких концентрациях. Также можно исследовать микроструктуру ткани мозга с использованием микроскопа и отследить паттерн соединений от одной области мозга с другой. [ 7 ]
Клеточная структура

Мозг всех видов состоит в основном из двух широких классов клеток мозга : нейронов и глиальных клеток . Глиальные клетки (также известные как глия или нейроглия ) бывают в нескольких типах, и выполняют ряд критических функций, включая структурную поддержку, метаболическую поддержку, изоляцию и руководство развитием. Однако нейроны обычно считаются наиболее важными клетками в мозге. [ 8 ] Свойство, которое делает нейроны уникальными, является их способностью отправлять сигналы в определенные целевые ячейки на большие расстояния. [ 8 ] Они посылают эти сигналы с помощью аксона, который представляет собой тонкое протоплазматическое волокно, которое простирается от тела клеток и проекта, обычно с многочисленными ветвями, в другие области, иногда поблизости, иногда в отдаленных частях мозга или тела. Длина аксона может быть необычной: например, если пирамидальная клетка (возбуждающий нейрон) коры головного мозга увеличивался так, что его клеточный корпус стал размером с человеческого тела, его аксон, одинаково увеличенное, станет кабелем несколько сантиметров в диаметре, простираясь более чем на километр. [ 9 ] Эти аксоны передают сигналы в форме электрохимических импульсов, называемых потенциалами действия, которые длятся менее тысячи секунды и проходят вдоль аксона со скоростью 1–100 метров в секунду. Некоторые нейроны постоянно испускают потенциал действия, со скоростью 10–100 в секунду, обычно в нерегулярных паттернах; Другие нейроны большую часть времени тихие, но иногда излучают взрыв потенциалов действия. [ 10 ]
Аксоны передают сигналы другим нейронам с помощью специализированных соединений, называемых синапсами . Один аксон может установить несколько тысяч синаптических соединений с другими клетками. [ 8 ] Когда потенциал действия, путешествующий по аксону, прибывает в синапс, он вызывает химического вещества, называемого нейротрансмиттером выпуск . Нейротрансмиттер связывается с рецепторными молекулами в мембране клетки -мишени. [ 8 ]
Синапсы являются ключевыми функциональными элементами мозга. [ 11 ] Основной функцией мозга является коммуникация клеток к клеткам , а синапсы-это точки, в которых происходит связь. По оценкам, человеческий мозг содержит приблизительно 100 триллионов синапсов; [ 12 ] Даже мозг фруктовой мухи содержит несколько миллионов. [ 13 ] Функции этих синапсов очень разнообразны: некоторые являются возбуждающими (захватывающие целевую ячейку); другие запрещены; Другие работают, активируя вторые системы мессенджеров , которые сложными способами меняют внутреннюю химию их целевых клеток. [ 11 ] Большое количество синапсов динамически модифицируется; То есть они способны менять силу таким образом, который контролируется моделями сигналов, которые проходят через них. Широко распространено мнение, что зависимая от активности модификация синапсов является основным механизмом мозга для обучения и памяти. [ 11 ]
Большая часть пространства в мозге занимает аксоны, которые часто объединяются вместе в так называемых нервных волоконно -волоконных трактах . Миелинизированный аксон обернут жирной изоляционной оболочкой миелина , которая служит для значительного увеличения скорости распространения сигнала. (Есть также немиелинизированные аксоны). Миелин белый, что делает части мозга, заполненной исключительно нервными волокнами, выглядят как светло- белое вещество , в отличие от серого вещества темного цвета , которое отмечает области с высокой плотностью нейронных клеточных тел. [ 8 ]
Эволюция
Общая билатерианская нервная система

За исключением нескольких примитивных организмов, таких как губки (которые не имеют нервной системы) [ 14 ] и Cnidarians (которые имеют диффузную нервную систему, состоящую из нервной сети ), [ 14 ] Все живые многоклеточные животные являются билатерианами , что означает животные с двусторонним симметричным планом тела (то есть левой и правой стороны, которые являются приблизительными зеркальными изображениями друг друга). [ 15 ] Считается, что все билатерианцы произошли от общего предка, который появился в конце криогенского периода, 700–650 миллионов лет назад, и было выдвинуто предположение, что у этого общего предка была форма простой трубки с сегментированным телом. [ 15 ] На схематическом уровне эта основная форма червя продолжает отражаться в архитектуре тела и нервной системы всех современных билатерианцев, включая позвоночных. [ 16 ] Фундаментальная двусторонняя форма тела представляет собой трубку с полостью кишки, бегущей от уст до заднего прохода, и нервным шнуром с увеличением ( ганглионом ) для каждого сегмента тела с особенно большим ганглионом спереди, называемого мозгом. Мозг маленький и простой у некоторых видов, таких как нематод черви ; У других видов, таких как позвоночные, это большой и очень сложный орган. [ 4 ] Некоторые типы червей, такие как пиявки , также имеют увеличенный ганглион на заднем конце нервного шнура, известного как «хвостовой мозг». [ 17 ]
Есть несколько типов существующих билатерианцев, которым не хватает узнаваемого мозга, в том числе эхинодермы и туникаты . Не было окончательно установлено, указывает ли существование этих безмозглых видов, что у самых ранних билатерианов не хватало мозга, или их предки развивались таким образом, что привело к исчезновению ранее существующей структуры мозга.
Беспозвоночные

Эта категория включает в себя Тардиграды , членистоногие , моллюски и многочисленные типы червей. Разнообразие планов беспозвоночных тела соответствует равному разнообразию в структурах мозга. [ 18 ]
Две группы беспозвоночных имеют заметно сложный мозг: членистоногие (насекомые, ракообразные , арабониды и другие) и головоногих (осьминоги, кальмары и подобные моллюски). [ 19 ] Мозг членистоногих и головоногих возникает из двойных параллельных нервных шнуров, которые простираются через тело животного. Членистоногие имеют центральный мозг, супересофагеальный ганглион , с тремя подразделениями и большими оптическими долями за каждым глазом для визуальной обработки. [ 19 ] Цефалоподы, такие как осьминог и кальмар, имеют самый большой мозг всех беспозвоночных. [ 20 ]
Есть несколько видов беспозвоночных, мозг которых интенсивно изучался, потому что у них есть свойства, которые делают их удобными для экспериментальной работы:
- Фруктовые мухи ( Drosophila ), из -за большого количества методов, доступных для изучения их генетики , были естественным субъектом для изучения роли генов в развитии мозга. [ 21 ] Несмотря на большое эволюционное расстояние между насекомыми и млекопитающими, было показано, что многие аспекты дрозофилы нейрогенетики имеют отношение к людям. первые биологические часовые гены Например, были идентифицированы путем изучения мутантов Drosophila , которые показали нарушение ежедневных циклов активности. [ 22 ] Поиск в геномах позвоночных выявил набор аналогичных генов, которые, как было установлено, играют сходную роль в биологических часах мыши, и поэтому почти наверняка также в биологических часах человека. [ 23 ] Исследования, проведенные на Drosophila, также показывают, что большинство нейропильных областей мозга постоянно реорганизуются на протяжении всей жизни в ответ на конкретные условия жизни. [ 24 ]
- Червячный червя Caenorhabditis Elegans , как и Drosophila , изучался в основном из -за его важности в генетике. [ 25 ] В начале 1970 -х годов Сидни Бреннер выбрал его в качестве модельного организма для изучения того, как гены контролируют развитие. Одним из преимуществ работы с этим червем является то, что план тела очень стереотипен: нервная система гермафродита содержит ровно 302 нейрона, всегда в одних и тех же местах, что делает идентичные синаптические соединения в каждом червяке. [ 26 ] Команда Бреннера нарезала червей на тысячи ультратонких секций и сфотографировала каждый под электронным микроскопом, а затем визуально соответствовала волокнам от секции к секции, чтобы наметить каждый нейрон и синапс во всем теле. [ 27 ] Полная схема нейронов C.Elegans - его контакт был достигнут. [ 28 ] Ничто, приближающееся к этому уровню детализации, не доступно для любого другого организма, и полученная информация позволила множество исследований, которые в противном случае не были бы невозможны. [ 29 ]
- Sea Slug Aplysia Californica была выбрана Нобелевской премией нейрофизиологом Эриком Канделом в качестве модели для изучения клеточной основы обучения и памяти из-за простоты и доступности ее нервной системы, и она была исследована в сотнях экспериментов. [ 30 ]
Позвоночные

Первые позвоночные появились более 500 миллионов лет назад ( MYA ) в течение кембрийского периода , и, возможно, напоминали современные Hagfish по форме. [ 31 ] Челюстная рыба появилась на 445 млн. Амфибий на 350 мю, рептилий на 310 млн. М и млекопитающих на 200 мюрьев (приблизительно). Каждый вид имеет одинаково длинную эволюционную историю , но мозг современных гагри, миноги , акул , амфибий, рептилий и млекопитающих демонстрируют градиент размера и сложности, которые примерно следуют эволюционной последовательности. Все эти мозги содержат один и тот же набор основных анатомических компонентов, но многие из них являются рудиментарными в Hagfish, тогда как у млекопитающих главная часть ( Telencephalon ) значительно разработана и расширяется. [ 32 ]
Мозги чаще всего сравниваются с точки зрения их размера. Взаимосвязь между размером мозга , размером тела и другими переменными изучалась в широком диапазоне видов позвоночных. Как правило, размер мозга увеличивается с размером тела, но не в простой линейной пропорции. В целом, меньшие животные, как правило, имеют больший мозг, измеряемые как часть размера тела. Для млекопитающих взаимосвязь между объемом мозга и массой тела по сути следует за законом власти с показателем около 0,75. [ 33 ] Эта формула описывает центральную тенденцию, но каждая семья млекопитающих отходит от нее в некоторой степени, в некотором смысле, что частично отражает сложность их поведения. Например, приматы имеют мозг в 5-10 раз больше, чем предсказывает формула. Хищники, как правило, имеют больший мозг, чем их добыча, относительно размера тела. [ 34 ]
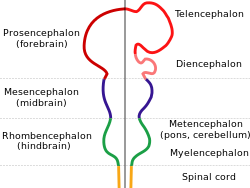
Все мозги позвоночных имеют общую основную форму, которая наиболее четко отображается на ранних стадиях эмбрионального развития. В самой ранней форме мозг появляется как три опухоля на переднем конце нейронной трубки ; Эти отеки в конечном итоге становятся передним, средним мозгом и задним мозгом ( Prosencephalon , Mesencephalon и Rhombencephalon соответственно). На самых ранних этапах развития мозга три области примерно равны по размеру. Во многих классах позвоночных, таких как рыба и амфибии, три части остаются одинаковыми по размеру у взрослого, но у млекопитающих передний мозг становится намного больше, чем другие части, а средний мозг становится очень маленьким. [ 8 ]
Мозг позвоночных изготовлены из очень мягких тканей. [ 8 ] Живая мозговая ткань розовая снаружи и в основном белая внутри, с тонкими вариациями в цвете. Мозг позвоночных окружены системой соединительной ткани мембран , называемые менингами , которые отделяют череп от мозга. Кровеносные сосуды попадают в центральную нервную систему через отверстия в менингеальных слоях. Клетки в стенах кровеносных сосудов тесно соединены друг с другом, образуя гематоэнцефалический барьер , который блокирует проход многих токсинов и патогенов [ 35 ] (Хотя в то же время блокируя антитела и некоторые лекарства, тем самым представляя особые проблемы при лечении заболеваний мозга). [ 36 ]
Нейроанатомисты обычно делят мозг позвоночных на шесть основных областей: телэнсфалон (полушария головного мозга), Diencephalon (Thalamus и Hypothalamus), Mesencephalon (средний мозг), церебеллум , Pons и Medulla Sallongata . Каждая из этих областей имеет сложную внутреннюю структуру. Некоторые части, такие как кора головного мозга и кора мозжечка, состоят из слоев, которые сложены или запутаны, чтобы соответствовать доступному пространству. Другие части, такие как таламус и гипоталамус, состоят из кластеров многих небольших ядер. Тысячи различимых областей могут быть идентифицированы в мозге позвоночных на основе тонких различий нейронной структуры, химии и связности. [ 8 ]

Хотя одни и те же основные компоненты присутствуют во всех мозгах позвоночных, некоторые ветви эволюции позвоночных привели к существенным искажениям геометрии мозга, особенно в области переднего мозга. Мозг акулы показывает основные компоненты простым способом, но в телеострных рыбах (подавляющее большинство существующих видов рыб), передний мозг стал «эверстным», как носок, выброшенный внутрь. У птиц также существуют серьезные изменения в структуре переднего мозга. [ 37 ] Эти искажения могут затруднить соответствие компонентов мозга от одного вида с другими видами. [ 38 ]
Вот список некоторых из наиболее важных компонентов мозга позвоночных, а также краткое описание их функций, как это понятно:
- Целое мозг , наряду со спинным мозгом, содержит много небольших ядер, вовлеченных в широкий спектр сенсорных и непроизвольных моторных функций, таких как рвота, частота сердечных сокращений и процессы пищеварения. [ 8 ]
- Поны . лежат в стволе мозга непосредственно над мозговым веществом Среди прочего, он содержит ядра, которые контролируют часто добровольные, но простые действия, такие как сон, дыхание, глотание, функция мочевого пузыря, равновесие, движение глаз, выражения лица и осанка. [ 39 ]
- Гипоталамус - это небольшая область у основания переднего мозга, сложность и важность которого противоречат его размеру. Он состоит из многочисленных небольших ядер, каждая с различными связями и нейрохимией. Гипоталамус занимается дополнительными непроизвольными или частично добровольными актами, такими как циклы сна и бодрствования, еда и питье, а также высвобождение некоторых гормонов. [ 40 ]
- Thalamus - это набор ядер с разнообразными функциями: некоторые участвуют в передаче информации в полушария головного мозга и обратно, в то время как другие участвуют в мотивации. Похоже, что субталамическая область ( Zona Incerta ) содержит системы, генерирующие действия, для нескольких типов «целесообразных» поведений, таких как еда, питье, дефекация и совокупление. [ 41 ]
- Мозжечок . модулирует выходы других систем мозга, будь то связанные с двигателем или связанные с мысли, чтобы сделать их определенными и точными Удаление мозжечка не мешает животному делать что -то конкретное, но это делает действия нерешительными и неуклюжными. Эта точность не встроена, а изучена методом проб и ошибок. Координация мышц, изученная во время езды на велосипеде, является примером типа нейронной пластичности , который может иметь место в основном внутри мозжечка. [ 8 ] 10% от общего объема мозга состоит из мозжечка, и 50% всех нейронов удерживаются в рамках его структуры. [ 42 ]
- Оптический Tectum позволяет направлять действия на точки в пространстве, чаще всего в ответ на визуальный ввод. У млекопитающих его обычно называют превосходным колликулом , а его наиболее изученная функция-направлять движения глаз. Это также направляет достижение движений и других объектных действий. Он получает сильные визуальные входы, но также входы от других чувств, которые полезны для направления действий, таких как слуховой вход в совы и ввод термочувствительных органов яма у змей. У некоторых примитивных рыб, таких как миноги , эта область является крупнейшей частью мозга. [ 43 ] Верхний колликул является частью среднего мозга.
- Паллий - это слой серого вещества, который лежит на поверхности переднего мозга и является наиболее сложным и последним эволюционным развитием мозга как органа. [ 44 ] У рептилий и млекопитающих это называется корой головного мозга . Многочисленные функции включают паллий, включая запах и пространственную память . У млекопитающих, где он становится настолько большим, чтобы доминировать в мозге, он берет на себя функции из многих других областей мозга. У многих млекопитающих кора головного мозга состоит из сложенных выпуклостей, называемых Giri , которые создают глубокие борозды или трещины, называемые сальками . Складки увеличивают площадь поверхности коры и, следовательно, увеличивают количество серого вещества и объем информации, которая может быть сохранена и обработана. [ 45 ]
- Гиппокамп , строго говоря , встречается только у млекопитающих. Тем не менее, область, из которой он получает, медиальный паллий, имеет аналоги у всех позвоночных. Существуют доказательства того, что эта часть мозга участвует в сложных событиях, таких как пространственная память и навигация у рыб, птиц, рептилий и млекопитающих. [ 46 ]
- Базальные ганглии - это группа взаимосвязанных структур в переднем мозге. Основной функцией базальных ганглиев, по-видимому, является выбор действий : они посылают ингибирующие сигналы на все части мозга, которые могут генерировать моторное поведение, и при правильных обстоятельствах могут выделять ингибирование, так что системы, генерирующие действия, способны выполнять их действия. Награда и наказание оказывают свои наиболее важные нервные последствия, изменяя связи в базальных ганглиях. [ 47 ]
- Обонятельная лампочка - это специальная структура, которая обрабатывает обонятельные сенсорные сигналы и отправляет свои выходные данные в обонятельную часть паллия. Это основной компонент мозга у многих позвоночных, но значительно снижается у людей и других приматов (в чьи чувствах преобладает информация, полученная в зрении, а не запахах). [ 48 ]
Рептилии

Современные рептилии и млекопитающие расходились от общего предка около 320 миллионов лет назад. [ 49 ] Интересно, что число существующих рептилий намного превышает количество видов млекопитающих, с 11 733 признанными видами рептилий [ 50 ] по сравнению с 5884 существующими млекопитающими. [ 51 ] разнообразием видов, рептилии расходились с точки зрения внешней морфологии, от безропоктических планеток Наряду с до бронированных хелонианцев , отражая адаптивное излучение до разнообразного набора сред. [ 52 ] [ 53 ]
Морфологические различия отражаются в фенотипе нервной системы , такие как: отсутствие нейронов боковых моторных колонн у змей, которые иннервируют мышцы конечностей, контролирующие движения конечностей; Отсутствие моторных нейронов, которые иннервируют мышцы туловища в черепах; Наличие иннервации от тройничного нерва до органов ямы, ответственных за инфракрасное обнаружение у змей. [ 52 ] Размер размера, веса и формы мозга можно найти в рептилиях. [ 54 ] Например, крокодильцы имеют наибольший объем мозга до пропорции тела, за которым следуют черепахи, ящерицы и змеи. Рептилии различаются в инвестициях в разные раздела мозга. Крокодильцы имеют самый большой доменфалон, в то время как у змей самый маленький. Черепахи имеют самый большой диенсифалон на вес тела, тогда как крокодильцы имеют наименьшее. С другой стороны, ящерицы имеют самый большой мезенсифалон. [ 54 ]
Тем не менее, их мозг имеет несколько характеристик, выявленных недавними анатомическими, молекулярными и онтогенетическими исследованиями. [ 55 ] [ 56 ] [ 57 ] Позвоночные имеют самые высокие уровни сходства во время эмбриологического развития, контролируемые консервативными факторами транскрипции и сигнальными центрами , включая экспрессию генов, морфологическую дифференцировку и дифференцировку типа клеток. [ 55 ] [ 52 ] [ 58 ] Фактически, высокие уровни транскрипционных факторов могут быть обнаружены во всех областях мозга у рептилий и млекопитающих, с общими нейрональными кластерами, просвещающей эволюцию мозга. [ 56 ] Консервативные факторы транскрипции выясняют, что эволюция действовала в разных областях мозга, либо сохраняя сходную морфологию и функцию, либо диверсифицируя ее. [ 55 ] [ 56 ]
Анатомически, у мозга рептилии меньше подразделений, чем у мозга млекопитающих, однако у него есть многочисленные консервативные аспекты, включая организацию спинного мозга и черепного нерва, а также разработанную схему мозга организации. [ 59 ] Разработанные мозги характеризуются мигрированными нейрональными клеточными телами вдали от перивентрикулярной матрицы, области развития нейронов, образующих организованные ядерные группы. [ 59 ] Помимо рептилий и млекопитающих , другие позвоночные с подробным мозгом включают в себя Hagfish , Galeomorph Sharks , коньки , лучи , телеосты и птицы . [ 59 ] Общие подробные мозги подразделяются в переднем мозге, среднем мозге и заднем мозге.
Задний мозг координирует и интегрирует сенсорные и моторные входы и выходы, ответственные за, но не ограничиваясь, ходьба, плавание или полеты. Он содержит входные и выходные аксоны, соединяющие спинной мозг, средний мозг и передачу переднего мозга из внешних и внутренних сред. [ 59 ] Средний мозг связывает сенсорные, моторные и интегрирующие компоненты, полученные от заднего мозга, соединяя его к переднему мозгу. Tectum, который включает в себя оптический Tectum и Torus semircularis, получает слуховые, визуальные и соматосенсорные входы, образуя интегрированные карты сенсорного и визуального пространства вокруг животного. [ 59 ] Tegmentum получает входящую сенсорную информацию и направляет двигательные ответы на переднюю часть и обратно. Зашивки соединяет задний мозг с средним мозгом. Область переднего мозга особенно хорошо развита, дополнительно разделен на Diencephalon и Telencephalon. Diencephalon связан с регуляцией движения глаз и тела в ответ на визуальные стимулы, сенсорную информацию, циркадные ритмы , обонятельный вход и вегетативную нервную систему . Telencephalon связан с контролем движений, нейротрансмиттеров и нейромодуляторов, ответственных за интеграцию входных данных и передаваемых результатов. , сенсорные системы и когнитивные функции. [ 59 ]
Птицы
является Птиевой мозг центральным органом нервной системы у птиц. Птицы обладают большим, сложным мозгом, которые обрабатывают , интегрируют и координируют информацию, полученную из окружающей среды, и принимают решения о том, как реагировать на остальную часть тела. во всех хордовых , птичий мозг содержится в костях черепа головы Как и .
Мозг птичьего мозга разделен на несколько срезов, каждая из которых с различной функцией. Церебрам . или трентенфалон делятся на два полушария и контролируют более высокие функции В Telencephalon преобладает большой паллий , который соответствует млекопитающих коре головного мозга и отвечает за когнитивные функции птиц. Паллий состоит из нескольких основных структур: гиперпаллий, дорсальной выпуклости паллия, обнаруженного только у птиц, а также нидопаллий, мезопаллий и архипаллий. Ядерная структура птицы, в которой нейроны распределяются в трехмерно расположенных кластерах, без крупномасштабного разделения белого вещества и серого вещества , хотя существуют слои, похожие на слое и столбцы. Структуры в паллиуме связаны с восприятием , обучением и познанием . Под паллиумом находятся два компонента субпаллиума, стриатум и паллидум . Субпаллий соединяет разные части Telencephalon и играет основные роли в ряде критических поведений. В задней части трентенфалона находятся Таламус , средний мозг и мозжечок . соединяет Задний мозг остальную часть мозга с спинным мозгом.
Размер и структура птичьего мозга обеспечивает заметное поведение птиц, таких как полет и вокализация . Выделенные структуры и пути интегрируют слуховые и визуальные чувства, сильные у большинства видов птиц, а также обычно более слабые обонятельные и тактильные чувства. Социальное поведение , широко распространенное среди птиц, зависит от организации и функций мозга. Некоторые птицы демонстрируют сильные способности познания, обеспечиваемых уникальной структурой и физиологией птичьего мозга.Млекопитающие
Наиболее очевидное различие между мозгом млекопитающих и других позвоночных - это их размер. В среднем у млекопитающего есть мозг, примерно в два раза больше, чем у птицы того же размера тела, и в десять раз больше, чем у рептилию того же размера тела. [ 60 ]
Размер, однако, не единственная разница: существуют также существенные различия в форме. Задний мозг и средний мозг млекопитающих, как правило, похожи на те у других позвоночных, но драматические различия появляются в переднем мозге, что значительно увеличено и также изменено по структуре. [ 61 ] Кора головного мозга - это часть мозга, которая наиболее сильно различает млекопитающих. У позвоночных, не являющихся млекопитающими, поверхность головного мозга выстлана сравнительно простой трехслойной структурой, называемой паллиумом . У млекопитающих паллий развивается в сложную шестислойную структуру, называемую неокортекс или изокортекс . [ 62 ] Несколько областей на краю неокортекса, включая гиппокамп и миндалину , также гораздо более широко развиты у млекопитающих, чем у других позвоночных. [ 61 ]
Разработка коры головного мозга несет с ней изменение в других областях мозга. Верхний колликул , который играет важную роль в визуальном контроле поведения у большинства позвоночных, сокращается до небольшого размера у млекопитающих, и многие из его функций воспринимаются визуальными областями коры головного мозга. [ 60 ] Церебелок млекопитающих содержит большую часть ( неоцелбеллум ), посвященную поддержке коры головного мозга, которая не имеет аналога у других позвоночных. [ 63 ]
У плацентарных млекопитающих существует широкий нервный тракт, соединяющий полушарии головного мозга, называемые мозолистым телом .
Приматы
| Разновидность | Уравнение [ 64 ] |
|---|---|
| Человек | 7.4–7.8 |
| Общий шимпанзе | 2.2–2.5 |
| Обезьяна -резус | 2.1 |
| Уничтось дельфин | 4.14 [ 65 ] |
| Слон | 1.13–2.36 [ 66 ] |
| Собака | 1.2 |
| Лошадь | 0.9 |
| Крыса | 0.4 |
Мозг людей и других приматов содержат те же структуры, что и мозг других млекопитающих, но, как правило, больше по размеру тела. [ 67 ] Коэффициент энцефализации (уравнение) используется для сравнения размеров мозга по видам. Это учитывает нелинейность отношений мозга и тела. [ 64 ] Люди имеют средний эквалайзер в диапазоне от 7 до 8, в то время как большинство других приматов имеют эквалайзер в диапазоне от 2 до 3. Дельфины имеют значения выше, чем у приматов, кроме людей, [ 65 ] Но почти у всех других млекопитающих есть значения эквалайзера, которые существенно ниже.
Большая часть увеличения мозга приматов происходит из -за огромного расширения коры головного мозга, особенно префронтальной коры и частей коры, участвующих в зрении . [ 68 ] Сеть визуальной обработки приматов включает в себя не менее 30 различимых областей мозга со сложной сетью взаимосвязи. Было подсчитано, что зоны визуальной обработки занимают более половины общей поверхности приматов неокортекс. [ 69 ] Префронтальная кора выполняет функции, которые включают планирование , рабочую память , мотивацию , внимание и исполнительный контроль . Он занимает гораздо большую долю мозга для приматов, чем для других видов, и особенно большая часть человеческого мозга. [ 70 ]
Разработка
Мозг развивается в запутанно организованной последовательности этапов. [ 71 ] Он меняет форму от простого отек в передней части нервного шнура на самых ранних эмбриональных стадиях, на сложный набор областей и соединений. Нейроны создаются в специальных зонах, которые содержат стволовые клетки , а затем мигрируют через ткань, чтобы достичь их конечных мест. Как только нейроны позиционируют себя, их аксоны прорастают и перемещаются по мозгу, разветвляются и простираются по мере их поступления, пока кончики не достигнут их целей и не образуют синаптические связи. В ряде частей нервной системы нейроны и синапсы производятся в чрезмерных количествах на ранних стадиях, а затем ненужные обрезаны. [ 71 ]
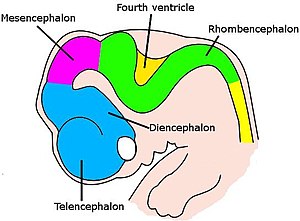
Для позвоночных ранние стадии нейронного развития аналогичны всем видам. [ 71 ] Когда эмбрион превращается из круглой капли клеток в червячную структуру, узкая полоса эктодермы, проходящая вдоль средней линии спины, вызывается , чтобы стать нейронной пластиной , предшественника нервной системы. Нейронная пластина складывается внутрь, образуя нервную канавку , а затем губы, которые выстраивают канавку, слияют, чтобы оградить нейронную трубку , полой шнур клеток с заполненным жидкостью желудочка в центре. На переднем конце желудочки и шнур набухают, образуя три пузырька, которые являются предшественниками Prospencephalon ( передний мозг), Mesencephalon (средний мозг) и Rompencephalon (задний мозг). На следующем этапе передний мозг расщепляется на два пузырька, называемые Telencephalon (который будет содержать кору головного мозга, базальные ганглии и связанные с ними структуры) и Diencephalon (который будет содержать таламус и гипоталамус). Примерно в то же время задний мозг распадается в метингфалон (который будет содержать мозжечок и поны) и миленсфалон (который будет содержать Медулла Илонгата ). Каждая из этих областей содержит пролиферативные зоны, где генерируются нейроны и глиальные клетки; Полученные клетки затем мигрируют, иногда на большие расстояния, на их конечные позиции. [ 71 ]
Как только нейрон на месте, он расширяет дендриты и аксон в область вокруг него. Аксоны, потому что они обычно протягивают большое расстояние от тела ячейки и должны достичь определенных целей, растут особенно сложным образом. Кончик растущего аксона состоит из капли протоплазмы, называемой ростом конуса , усеянного химическими рецепторами. Эти рецепторы ощущают местную среду, вызывая привлечение или отталкиваемое рост конуса различными клеточными элементами, и, следовательно, ее можно тянуть в определенном направлении в каждой точке по его пути. Результатом этого процесса нанесения пути является то, что рост конус проходит через мозг, пока не достигнет своей области назначения, где другие химические сигналы заставляют его начать генерирование синапсов. Принимая во внимание весь мозг, тысячи генов создают продукты, которые влияют на аксональное место для пути. [ 71 ]
Тем не менее, синаптическая сеть, которая, наконец, появляется, только частично определяется генами. Во многих частях мозга аксоны первоначально «перерастают», а затем «обрезаны» механизмами, которые зависят от нейронной активности. [ 71 ] Например, в проекции от глаз до среднего мозга структура у взрослого содержит очень точное отображение, соединяющее каждую точку на поверхности сетчатки с соответствующей точкой в слое среднего мозга. На первых этапах развития каждый аксон от сетчатки направляется в правую общую окрестности в среднем мозге с помощью химических сигналов, но затем очень обильно разветвляется и устанавливает первоначальный контакт с широким полосом нейронов среднего мозга. Сетчатка до рождения содержит специальные механизмы, которые заставляют ее генерировать волны активности, которые спонтанно происходят в случайной точке, а затем медленно распространяются по слону сетчатки. Эти волны полезны, потому что они заставляют соседние нейроны одновременно активными; То есть они производят схему нейронной активности, которая содержит информацию о пространственном расположении нейронов. Эта информация используется в среднем мозге механизмом, который заставляет синапсы ослаблять, и в конечном итоге исчезает, если активность в аксоне не сопровождается активностью целевой клетки. Результатом этого сложного процесса является постепенная настройка и ужесточение карты, оставляя ее, наконец, в своей точной форме для взрослых. [ 72 ]
Подобные вещи случаются в других областях мозга: начальная синаптическая матрица генерируется в результате генетически определенного химического наведения, но затем постепенно уточняется в зависимости от активности механизмов, частично обусловленных внутренней динамикой, частично с помощью внешних сенсорных входов. В некоторых случаях, как и в случае с системой сетчатки сетчатки, паттерны активности зависят от механизмов, которые работают только в развивающемся мозге, и, по-видимому, существуют исключительно для руководства развитием. [ 72 ]
У людей и многих других млекопитающих новые нейроны создаются в основном до рождения, а младенческий мозг содержит значительно больше нейронов, чем взрослый мозг. [ 71 ] Однако есть несколько областей, где новые нейроны продолжают генерироваться на протяжении всей жизни. Две области, для которых хорошо установлен нейрогенез взрослого , - это обонятельная лампочка, которая вовлечена в обоняние и зубчатую извилину гиппокампа, где есть доказательства того, что новые нейроны играют роль в хранении вновь приобретенных воспоминаний. Однако за этими исключениями набор нейронов, присутствующих в раннем детстве, является набором, который присутствует на всю жизнь. Глиальные клетки разные: как и в случае с большинством типов клеток в организме, они генерируются на протяжении всей жизни. [ 73 ]
Давно проводились споры о том, могут ли качества разума , личности и интеллекта отнесены к наследственности или воспитанию . [ 74 ] Хотя многие детали еще предстоит уладить, нейробиология показывает, что оба фактора важны. Гены определяют как общую форму мозга, так и то, как он реагирует на опыт, но для уточнения матрицы синаптических связей требуется опыт, что приводит к значительно повышенной сложности. Наличие или отсутствие опыта имеют решающее значение в ключевые периоды развития. [ 75 ] Кроме того, количество и качество опыта важны. Например, животные, поднятые в обогащенных средах , демонстрируют толстые коры головного мозга, что указывает на высокую плотность синаптических соединений по сравнению с животными с ограниченными уровнями стимуляции. [ 76 ]
Физиология
Функции мозга зависят от способности нейронов передавать электрохимические сигналы другим клеткам и их способность соответствующим образом реагировать на электрохимические сигналы, полученные от других клеток. Электрические свойства нейронов контролируются широким разнообразием биохимических и метаболических процессов, в частности, взаимодействия между нейротрансмиттерами и рецепторами, которые происходят в синапсах. [ 8 ]
Нейротрансмиттеры и рецепторы
Нейротрансмиттеры - это химические вещества, которые высвобождаются при синапсах, когда локальная мембрана деполяризована и 2+ Входит в клетку, как правило, когда потенциал действия приходит в синапс - нейротрансмиттеры присоединяются к молекулам рецепторов на мембране целевой ячейки синапса (или клетки) и тем самым изменяют электрические или химические свойства молекул рецептора. За некоторыми исключениями каждый нейрон в мозге высвобождает один и тот же химический нейротрансмиттер или комбинацию нейротрансмиттеров, во всех синаптических связях, которые он устанавливает с другими нейронами; Это правило известно как принцип Дейла . [ 8 ] Таким образом, нейрон может характеризоваться нейротрансмиттерами, которые он выпускает. Подавляющее большинство психоактивных препаратов оказывают свои эффекты, изменяя специфические нейротрансмиттерные системы. Это относится к таким лекарствам, как каннабиноиды , никотин , героин , кокаин , алкоголь , флуоксетин , хлорпромазин и многие другие. [ 77 ]
Два нейротрансмиттеры, которые наиболее широко обнаруживаются в мозге позвоночных,-это глутамат , который почти всегда оказывает возбуждающее воздействие на нейроны-мишени и гамма-аминобутирическую кислоту (ГАМК), которая почти всегда ингибирует. Нейроны, использующие эти передатчики, могут быть обнаружены почти в каждой части мозга. [ 78 ] Из -за их повсеместного распространения наркотики, которые действуют на глутамат или ГАМК, как правило, оказывают широкие и мощные эффекты. Некоторые общие анестетики действуют, уменьшая эффекты глутамата; Большинство транквилизаторов оказывают свои седативные эффекты, усиливая эффекты ГАМК. [ 79 ]
Существуют десятки других химических нейротрансмиттеров, которые используются в более ограниченных областях мозга, часто областям, посвященной конкретной функции. серотонин Например, - первичная мишень многих антидепрессантов и многих диетических средств - исходит исключительно из небольшой области ствола мозга, называемой ядрами Рафе . [ 80 ] Норепинефрин , который участвует в возбуждении, поступает исключительно из близлежащей небольшой области, называемой локусом coeruleus . [ 81 ] Другие нейротрансмиттеры, такие как ацетилхолин и дофамин, имеют несколько источников в мозге, но не так повсеместно распределены, как глутамат и ГАМК. [ 82 ]
Электрическая активность

В качестве побочного эффекта электрохимических процессов, используемых нейронами для передачи сигналов, ткань мозга генерирует электрические поля, когда он активен. Когда большое количество нейронов демонстрирует синхронизированную активность, электрические поля, которые они генерируют, могут быть достаточно большими для обнаружения вне черепа, используя электроэнцефалографию (ЭЭГ) [ 83 ] или магнитоэнцефалография (MEG). Записи ЭЭГ, наряду с записями, сделанными из электродов, имплантированных в мозг животных, таких как крысы, показывают, что мозг живого животного постоянно активно активно, даже во время сна. [ 84 ] Каждая часть мозга показывает смесь ритмической и неритмической активности, которая может варьироваться в зависимости от поведенческого состояния. У млекопитающих кора головного мозга имеет тенденцию демонстрировать большие медленные дельта-волны во время сна, более быстрые альфа-волны, когда животное бодрствует, но невнимательнее, и хаотика, которая выглядит нерегулярной активностью, когда животное активно участвует в задаче, называемой бета - гамма-волнами . Во время эпилептического захвата механизмы ингибирующего контроля мозга не функционируют, а электрическая активность поднимается до патологических уровней, создавая следы ЭЭГ, которые показывают большие паттерны волны и всплеска, не наблюдаемые в здоровом мозге. Отношение этих моделей уровня населения с вычислительными функциями отдельных нейронов является основным направлением современных исследований в области нейрофизиологии . [ 84 ]
Метаболизм
Все позвоночные имеют барьер крови -мэрию , который позволяет метаболизму внутри мозга работать не так, как метаболизм в других частях тела. Нейвоскулярная единица регулирует церебральный кровоток, так что активированные нейроны могут поставляться с энергией. Глиальные клетки играют важную роль в метаболизме мозга, контролируя химический состав жидкости, которая окружает нейроны, включая уровни ионов и питательных веществ. [ 85 ]
Мозговая ткань потребляет большое количество энергии пропорционально его объему, поэтому большой мозг вызывает серьезные метаболические потребности на животных. Необходимость ограничить массу тела, например, для лета, по -видимому, привела к выбору для уменьшения размера мозга у некоторых видов, таких как летучих мышей . [ 86 ] Большая часть энергопотребления мозга уходит в поддержание электрического заряда ( мембранного потенциала ) нейронов. [ 85 ] Большинство видов позвоночных посвящены 2% и 8% базального метаболизма мозгу. Однако у приматов этот процент намного выше - у людей он вырос до 20–25%. [ 87 ] Потребление энергии мозга не сильно различается с течением времени, но активные области коры головного мозга потребляют несколько больше энергии, чем неактивные области; Это формирует основу для функциональных методов визуализации мозга PET , FMRI , [ 88 ] и нирс . [ 89 ] Мозг обычно получает большую часть своей энергии от кислородно-зависимого метаболизма глюкозы (то есть сахара в крови), [ 85 ] Но кетоны обеспечивают основной альтернативный источник, вместе с вкладами жирных кислот средней цепи ( каприловые и гептановые кислоты), [ 90 ] [ 91 ] лактат , [ 92 ] ацетат , [ 93 ] и, возможно, аминокислоты . [ 94 ]
Функция

Информация из органов смысла собирается в мозге. Там он используется, чтобы определить, какие действия предназначен для организма. Мозг обрабатывает необработанные данные для извлечения информации о структуре окружающей среды. Затем он объединяет обработанную информацию с информацией о текущих потребностях животного и памяти о прошлых обстоятельствах. Наконец, на основе результатов он генерирует моторные шаблоны ответа. Эти задачи обработки сигналов требуют сложного взаимодействия между различными функциональными подсистемами. [ 95 ]
Функция мозга состоит в том, чтобы обеспечить когерентный контроль над действиями животного. Централизованный мозг позволяет коактивировать группы мышц в сложных паттернах; Это также позволяет стимулам, поражающимся в одной части тела вызывать ответы в других частях, и это может помешать различным частям тела действовать на перекрестных целях друг другу. [ 95 ]
Восприятие

Человеческий мозг предоставляется информацией о свете, звуке, химическом составе атмосферы, температуре, положении тела в пространстве ( проприоцепции ), химическом составе крови и многое другое. У других животных присутствуют дополнительные чувства, такие как инфракрасная тепловая точка змей , ощущение магнитного поля некоторых птиц или чувство электрического поля, в основном наблюдаемое у водных животных.
Каждая сенсорная система начинается со специализированных рецепторных клеток, [ 8 ] такие как фоторецепторные клетки в сетчатке глаза волосковые или чувствительные к клетки в улитке уха вибрации . Аксоны сенсорных рецепторных клеток перемещаются в спинной мозг или мозг, где они передают свои сигналы в сенсорное ядро первого порядка, посвященное одной конкретной сенсорной модальности . Это первичное сенсорное ядро отправляет информацию в сенсорные области высшего порядка, которые посвящены тому же модальности. В конечном итоге, посредством станции в таламусе , сигналы отправляются в кору головного мозга, где они обрабатываются для извлечения соответствующих функций и интегрируются с сигналами, поступающими из других сенсорных систем. [ 8 ]
Моторный контроль
Моторные системы - это области мозга, которые участвуют в инициировании движений тела , то есть в активации мышц. За исключением мышц, которые контролируют глаз, которые управляются ядрами в среднем мозге, все добровольные мышцы в организме непосредственно иннервируются моторными нейронами в спинном мозге и заднем мозге. [ 8 ] Спинальные двигательные нейроны контролируются как нейронными цепи, внутренними по отношению к спинному мозгу, так и входными данными, которые спускаются от мозга. Внутренние схемы позвоночника реализуют много рефлекторных реакций и содержат генераторы рисунков для ритмических движений, таких как ходьба или плавание . Нисходящие связи от мозга позволяют обеспечить более сложный контроль. [ 8 ]
Мозг содержит несколько моторных областей, которые проецируются непосредственно в спинном мозге. На самом низком уровне находятся моторные участки в мозге и понах, которые контролируют стереотипные движения, такие как ходьба, дыхание или глотание . На более высоком уровне находятся области в среднем мозге, такие как красное ядро , которое отвечает за координацию движений рук и ног. На более высоком уровне все же находится основная моторная кора , полоска ткани, расположенная на заднем крае лобной доли. Основная моторная кора посылает проекции в области подкорковых двигателей, но также посылает массивную проекцию непосредственно в спинной мозг через пирамидальный тракт . Эта прямая кортикоспинальная проекция обеспечивает точный добровольный контроль мелких деталей движений. Другие моторные области мозга оказывают вторичные эффекты, проецируя на основные моторные области. Среди наиболее важных вторичных областей - премоторная кора , дополнительная моторная зона , базальные ганглии и мозжечок . [ 8 ] В дополнение ко всем вышеперечисленным, мозг и спинной мозг содержат обширную схему для контроля вегетативной нервной системы , которая контролирует движение гладкой мышцы тела. [ 8 ]
| Область | Расположение | Функция |
|---|---|---|
| Вентральный рог | Спинной мозг | Содержит моторные нейроны, которые непосредственно активируют мышцы [ 96 ] |
| Гульмоторные ядра | Средний мозг | Содержит моторные нейроны, которые непосредственно активируют глазные мышцы [ 97 ] |
| Мозжечок | Задних изделий | Калибрует точность и время движений [ 8 ] |
| Базальные ганглии | Передний мозг | Выбор действий на основе мотивации [ 98 ] |
| Моторная кора | Фронтальная доля | Прямая корковая активация позвоночных моторных цепей [ 99 ] |
| Премоторная кора | Фронтальная доля | Группы элементарные движения в скоординированные шаблоны [ 8 ] |
| Дополнительная моторная зона | Фронтальная доля | Последовательности движения во временные узоры [ 100 ] |
| Префронтальная кора | Фронтальная доля | Планирование и другие исполнительные функции [ 101 ] |
Спать
Многие животные чередуются между сна и пробуждения в ежедневном цикле. Пробуждение и бдительность также модулируются в более тонкой масштабе времени сетью областей мозга. [ 8 ] Ключевым компонентом системы сна является супрахиазматическое ядро (SCN), крошечная часть гипоталамуса, расположенной непосредственно над точкой, в которой зрительные нервы от двух глаз пересекаются. SCN содержит центральные биологические часы организма. Там нейроны показывают уровни активности, которые растут и падают с периодом около 24 часов, циркадными ритмами : эти колебания активности обусловлены ритмическими изменениями в экспрессии набора «часовых генов». SCN продолжает сохранять время, даже если он иссекается из мозга и помещается в блюдо с теплым питательным раствором, но обычно он получает вход от зрительных нервов через ретиногипоталамический тракт (RHT), который позволяет ежедневным циклам световых темных калибровать часы. [ 102 ]
SCN проецируется на ряд областей в гипоталамусе, стволе мозга и среднего мозга, которые участвуют в реализации циклов сна и бодрствования. Важным компонентом системы является ретикулярное образование , группа нейрон-кластеров рассеивалась по ядру нижнего мозга. Ретикулярные нейроны посылают сигналы в таламус, что, в свою очередь, посылает сигналы контроля на уровне активности в каждую часть коры. Повреждение ретикулярного образования может привести к постоянному состоянию комы. [ 8 ]
Сон включает в себя большие изменения в активности мозга. [ 8 ] До 1950 -х годов обычно считалось, что мозг по существу отключается во время сна, [ 103 ] Но теперь это известно, что это далеко не правда; Заявление продолжается, но паттерны становятся очень разными. Существует два типа сна: быстрое сон (со сновидением ) и NREM (не-REM, обычно без мечтаний), которые повторяются в слегка различных схемах во всем эпизоде сна. Могут быть измерены три широких типа различных моделей активности мозга: REM, Light NREM и Deep NREM. Во время глубокого сна NREM, также называемого медленным волновым соном , активность в коре в коре принимает форму больших синхронизированных волн, тогда как в состоянии бодрствования это шумно и дезинхронизировано. Уровни нейротрансмиттеров норэпинефрина и серотонина падают во время медленного волнового сна и падают почти до нуля во время быстрого сна; Уровни ацетилхолина показывают обратную схему. [ 8 ]
Гомеостаз

Для любого животного выживаемость требует поддержания различных параметров телесного состояния в пределах ограниченного диапазона вариаций: они включают температуру, содержание воды, концентрацию соли в кровотоке, уровень глюкозы в крови, уровень кислорода в крови и другие. [ 104 ] Способность животного регулировать внутреннюю среду своего тела - интенсивной среды , как назвал его пионерский физиолог Клод Бернард - известна как гомеостаз ( греческий для «стоящего неподвижного»). [ 105 ] Поддержание гомеостаза является решающей функцией мозга. Основным принципом, лежащим в основе гомеостаза, является отрицательная обратная связь : в любое время, когда параметр расходится от своей установки, датчики генерируют сигнал ошибки, который вызывает ответ, который заставляет параметр переключаться обратно к его оптимальному значению. [ 104 ] (Этот принцип широко используется в инженерии, например, в контроле температуры с использованием термостата .)
У позвоночных частью мозга, которая играет наибольшую роль, является гипоталамус , небольшая область у основания переднего мозга, размер которого не отражает его сложность или важность его функции. [ 104 ] Гипоталамус представляет собой набор небольших ядер, большинство из которых участвуют в основных биологических функциях. Некоторые из этих функций связаны с возбуждением или социальными взаимодействиями, такими как сексуальность, агрессия или материнское поведение; Но многие из них относятся к гомеостазу. Несколько ядер гипоталамуса получают вход от датчиков, расположенных в слизистой оболочке кровеносных сосудов, передавая информацию о температуре, уровне натрия, уровне глюкозы, уровне кислорода в крови и других параметрах. Эти ядра гипоталамуса посылают выходные сигналы в моторные области, которые могут генерировать действия для исправления недостатков. Некоторые результаты также идут к гипофизом , крошечной железе, прикрепленной к мозгу непосредственно под гипоталамусом. Гипофик выделяет гормоны в кровоток, где они циркулируют по всему организму и вызывают изменения в клеточной активности. [ 106 ]
Мотивация
Отдельные животные должны выражать поведение, способствующее выживанию, такие как поиск пищи, воды, укрытия и партнера. [ 107 ] Мотивационная система в мозге контролирует текущее состояние удовлетворения этих целей и активирует поведение для удовлетворения любых возникающих потребностей. Мотивационная система работает в основном механизмом вознаграждения. Когда конкретное поведение сопровождаются благоприятными последствиями, активируется механизм вознаграждения в мозге, что индуцирует структурные изменения внутри мозга, которые вызывают повторение того же поведения позже, когда возникает подобная ситуация. И наоборот, когда за поведением следуют неблагоприятные последствия, механизм наказания мозга активируется, что вызывает структурные изменения, которые вызывают подавление поведения, когда в будущем возникают подобные ситуации. [ 108 ]
Большинство организмов, изучаемых на сегодняшний день, используют механизм вознаграждения и выплаты: например, черви и насекомые могут изменить их поведение в поисках источников пищи или избежать опасностей. [ 109 ] У позвоночных система награждения реализована определенным набором структур мозга, в основе которых лежат базальные ганглии, набор взаимосвязанных областей у основания переднего мозга. [ 47 ] Базальные ганглии являются центральным местом, в котором принимаются решения: базальные ганглии оказывают устойчивый ингибирующий контроль над большинством двигательных систем в мозге; Когда это запрещение выделяется, моторная система разрешается выполнять действие, которое он запрограммирован для выполнения. Награды и наказания функционируют, изменяя взаимосвязь между входными данными, которые получают базальные ганглии, и сигналами принятия решений, которые испускаются. Механизм вознаграждения лучше понят, чем механизм наказания, потому что его роль в злоупотреблении наркотиками привела к тому, что его очень интенсивно изучалось. Исследования показали, что дофамин нейротрансмиттера играет центральную роль: наркотики, такие как кокаин, амфетамин и никотин, либо приводят к повышению уровня дофамина, либо вызывают воздействие дофамина внутри мозга. [ 110 ]
Обучение и память
Почти все животные способны изменять свое поведение в результате опыта - даже самые примитивные типы червей. Поскольку поведение обусловлено активностью мозга, изменения в поведении должны каким -то образом соответствовать изменениям внутри мозга. Уже в конце 19 -го века теоретики, такие как Сантьяго Рамон и Каджал, утверждали, что наиболее правдоподобным объяснением является то, что обучение и память выражаются как изменения в синаптических связях между нейронами. [ 111 ] Однако до 1970 года экспериментальные данные, подтверждающие гипотезу синаптической пластичности . В 1971 году Тим Блисс и Терье Лёмо опубликовали статью о явлении, который теперь называется долгосрочным потенцированием : в статье показано явные признаки синаптических изменений, вызванных активностью, которые длились не менее нескольких дней. [ 112 ] С тех пор технические достижения сделали такие эксперименты намного проще, и были проведены тысячи исследований, которые прояснили механизм синаптических изменений и обнаружили другие типы синаптических изменений, основанных на активности в различных областях мозга, в том числе Кора головного мозга, гиппокамп, базальные ганглии и мозжечок. [ 113 ] ) , полученный из мозга ( Нейротрофический фактор ( BDNF BDNF ) и физическая активность , по-видимому, играют полезную роль в процессе. [ 114 ]
Нейробиологи в настоящее время различают несколько типов обучения и памяти, которые реализованы мозгом различными способами:
- Рабочая память - это способность мозга сохранять временное представление информации о задаче, в которой в настоящее время привлечено животное. Считается, что такая динамическая память опосредована формированием клеточных сборок - группы активированных нейронов, которые поддерживают свои активность постоянно стимулируя друг друга. [ 115 ]
- Эпизодическая память - это способность запоминать детали конкретных событий. Такая память может длиться всю жизнь. Много доказательств подразумевает гиппокамп в сыгрании решающей роли: люди с серьезным повреждением гиппокампа иногда демонстрируют амнезию , то есть неспособность сформировать новые длительные эпизодические воспоминания. [ 116 ]
- Семантическая память - это способность изучать факты и отношения. Этот вид памяти, вероятно, хранится в основном в коре головного мозга, опосредованной изменениями в соединениях между ячеями, которые представляют конкретные типы информации. [ 117 ]
- Инструментальное обучение - это способность для вознаграждений и наказаний изменять поведение. Он реализован сетью областей мозга, сосредоточенных на базальных ганглиях. [ 118 ]
- Моторное обучение - это способность уточнить закономерности движения тела, практикуя, или, в более общем плане, путем повторения. Вовлечен ряд областей мозга, в том числе премоторная кора , базальные ганглии и особенно мозжечок, который функционирует как большой банк памяти для микроуджнений параметров движения. [ 119 ]
Исследовать

Поле нейробиологии охватывает все подходы, которые стремятся понять мозг и остальную часть нервной системы. [ 8 ] Психология стремится понять разум и поведение, а неврология - это медицинская дисциплина, которая диагностирует и рассматривает заболевания нервной системы. Мозг также является наиболее важным органом, изученным в психиатрии , отрасли медицины, которая работает для изучения, предотвращения и лечения психических расстройств . [ 120 ] Когнитивная наука стремится объединить нейробиологию и психологию с другими областями, которые касаются мозга, таких как информатика ( искусственный интеллект и подобные области) и философия . [ 121 ]
Самый старый метод изучения мозга является анатомическим , и до середины 20 -го века большая часть прогресса в нейробиологии исходила от развития лучших клеточных пятен и лучших микроскопов. Нейроанатомисты изучают крупномасштабную структуру мозга, а также микроскопическую структуру нейронов и их компонентов, особенно синапсы. Среди других инструментов они используют множество пятен, которые показывают нейронную структуру, химию и связь. В последние годы разработка методов иммуноокрашивания позволила изучить нейроны, которые экспрессируют специфические наборы генов. Кроме того, функциональная нейроанатомия использует методы медицинской визуализации для корреляции вариаций в структуре мозга человека с различиями в познании или поведении. [ 122 ]
Нейрофизиологи изучают химические, фармакологические и электрические свойства мозга: их основными инструментами являются лекарства и записи. Тысячи экспериментально разработанных лекарств влияют на нервную систему, некоторые очень специфичны. Записи активности мозга могут быть сделаны с использованием электродов, либо приклеены к коже головы, как в исследованиях ЭЭГ , либо имплантированы внутри мозга животных для внеклеточных записей, которые могут обнаруживать потенциалы действия, генерируемые отдельными нейронами. [ 123 ] Поскольку мозг не содержит рецепторов боли, возможно использовать эти методы для записи активности мозга от животных, которые бодрствуют и ведут себя, не вызывая дистресса. Те же методы иногда использовались для изучения активности мозга у пациентов с человеческими пациентами с неразрешимой эпилепсией , в тех случаях, когда была медицинская необходимость имплантата электродов для локализации области мозга, ответственной за эпилептические припадки . [ 124 ] Методы функциональной визуализации, такие как МРТ, также используются для изучения мозговой активности; Эти методы в основном использовались с людьми, потому что они требуют сознательного субъекта, чтобы оставаться неподвижным в течение длительных периодов времени, но они имеют большое преимущество в том, что они неинвазивны. [ 125 ]
Другим подходом к функции мозга является изучение последствий повреждения определенных областей мозга. Несмотря на то, что он защищен черепом и менингами , окруженными спинномозговой жидкостью и изолированным из кровотока кровью -мрачным барьером, деликатная природа мозга делает его уязвимым для многочисленных заболеваний и нескольких видов повреждения. У людей эффекты инсультов и других типов повреждения головного мозга были ключевым источником информации о функции мозга. Однако, поскольку нет возможности экспериментально контролировать характер ущерба, эта информация часто трудно интерпретировать. В исследованиях на животных, чаще всего с участием крыс, можно использовать электроды или локально инъецированные химические вещества для получения точных моделей повреждения, а затем изучить последствия для поведения. [ 127 ]
Вычислительная нейробиология охватывает два подхода: во -первых, использование компьютеров для изучения мозга; Во -вторых, изучение того, как мозги выполняют вычисления. С одной стороны, можно написать компьютерную программу для моделирования работы группы нейронов, используя системы уравнений, которые описывают их электрохимическую активность; Такие симуляции известны как биологически реалистичные нейронные сети . С другой стороны, можно изучить алгоритмы для нервных вычислений, моделируя или математически анализируя операции упрощенных «единиц», которые обладают некоторыми свойствами нейронов, но абстрагируют большую часть их биологической сложности. Вычислительные функции мозга изучаются как компьютерными учеными, так и нейробиологом. [ 128 ]
Вычислительное нейрогенетическое моделирование связано с изучением и разработкой динамических нейрональных моделей для моделирования функций мозга в отношении генов и динамических взаимодействий между генами.
В последние годы было увеличено применение генетических и геномных методов для изучения мозга [ 129 ] и акцент на роли нейротрофических факторов и физической активности в нейропластичности . [ 114 ] Наиболее распространенными предметами являются мыши из -за доступности технических инструментов. Теперь это возможно с относительной легкостью «нокаутировать» или мутировать широкий спектр генов, а затем изучить влияние на функцию мозга. Также используются более сложные подходы: например, с использованием рекомбинации Cre-Lox, в определенном времени можно активировать или деактивировать гены в определенных частях мозга. [ 129 ]
В последние годы также наблюдались быстрые достижения в области одноклеточных технологий секвенирования, и они были использованы для использования клеточной гетерогенности мозга в качестве средства лучшего понимания роли различных типов клеток и биологии (а также как геномные варианты влиять на отдельные типы клеток). В 2024 году исследователи изучили большой интегрированный набор данных почти 3 миллиона ядер из префронтального Cortext человека от 388 человек. [ 130 ] При этом они аннотировали 28 типов клеток для оценки экспрессии и изменения хроматина в семействах генов и лекарственных целях. Они идентифицировали около полумиллиона регуляторных элементов, специфичных для типа клеток и около 1,5 миллионов локусов количественных признаков экспрессии в одной клетках (то есть геномных вариантов со сильными статистическими ассоциациями с изменениями экспрессии генов в определенных типах клеток), которые затем использовались для строительства клетки Регуляторные сети-типа (исследование также описывает сети связи с клетками к клеткам). Было обнаружено, что эти сети проявляют клеточные изменения при старении и психоневрологических расстройствах. В рамках того же исследования модель машинного обучения была разработана для точного вменения экспрессии одноклеточной экспрессии (эта модель приоритет ~ 250 генам риска заболевания и мишенями для лекарств с соответствующими типами клеток).
История

Самый старый мозг, который был обнаружен, был в Армении в пещерном комплексе Areni-1 . Мозг, который, по оценкам, более 5000 лет, был найден в черепе от 12 до 14 лет. Хотя мозг был сморщен, они были хорошо сохранены из -за климата, найденного в пещере. [ 131 ]
Ранние философы были разделены на то, лежит ли место души в мозге или в сердце. Аристотель предпочитал сердце и подумал, что функция мозга была просто для того, чтобы охладить кровь. Демократ , изобретатель атомной теории материи, выступал за душу из трех частей, с интеллектом в голове, эмоциями в сердце и жаждой печени. [ 132 ] Неизвестный автор «Священной болезни» , медицинский трактат в корпусе Гиппократа , однозначно вышел в пользу мозга, написание:
Мужчины должны знать, что из -за ничего, кроме мозга, приходят радости, удовольствия, смех и спорт, а также печали, горе, уныние и плаги. ... и по тому же органу мы становимся сумасшедшими и бредовыми, и страхи и ужасы атакуют нас, некоторые ночью, а некоторые днем, и мечты и несвоевременные блуждания, и заботы, которые не подходят, и незнание нынешних обстоятельств, десутде. , и неквалифицированность. Все эти вещи мы переносим из мозга, когда это не здорово ...
- О священном заболевании , приписываемом Гиппократа [ 133 ]

Римский врач Гален также выступал за важность мозга и в некоторой степени теоретизировал, как это может работать. Гален проследил анатомические отношения между мозгом, нервами и мышцами, демонстрируя, что все мышцы в организме соединены с мозгом через разветвленную сеть нервов. Он постулировал, что нервы механически активируют мышцы, неся таинственное вещество, которое он назвал Pneumata Psychikon , обычно переводимый как «духи животных». [ 132 ] Идеи Галена были широко известны в средние века, но не намного дальнейший прогресс достиг до тех пор, пока эпохи Возрождения, когда возобновилось подробное анатомическое исследование, в сочетании с теоретическими предположениями Рене Декарта и тех, кто следовал за ним. Декарт, как Гален, думал о нервной системе в гидравлических терминах. Он полагал, что самые высокие когнитивные функции выполняются нефизическими ресторанами , но что большинство поведения людей и все поведение животных можно было бы объяснить механистически. [ 132 ]
Первый реальный прогресс к современному пониманию нервной функции, тем не менее, произошел из расследований Луиджи Гальвани (1737–1798), который обнаружил, что шок статического электричества, применяемый к открытому нерву мертвой лягушки, может привести к сокращению ноги, чтобы сразиться. Полем С тех пор каждый крупный прогресс в понимании последовал более или менее напрямую от разработки новой техники исследования. До первых лет 20 -го века наиболее важные достижения были получены из новых методов окрашивания клеток. [ 134 ] Особенно критическим было изобретение пятна Гольджи , которое (при правильном использовании) окрашивает лишь небольшую долю нейронов, но окрашивает их полностью, включая тело клеточного, дендрита и аксон. Без такого окрашивания ткань мозга под микроскопом выглядит как непроницаемый клубок протоплазматических волокон, в которых невозможно определить какую -либо структуру. В руках Камилло Гольджи , и особенно испанского нейроанатомиста Сантьяго Рамон и Каджал , новое пятно выявило сотни различных типов нейронов, каждый из которых со своей собственной уникальной дендритной структурой и схемой связности. [ 135 ]

В первой половине 20 -го века достижения в области электроники позволили изучить электрические свойства нервных клеток, кульминационных в работе Алана Ходжкина , Эндрю Хаксли и других о биофизике потенциала действия и работы Бернарда Каца и других на электрохимии синапса. [ 136 ] Эти исследования дополнили анатомическую картину с концепцией мозга как динамической сущности. Размышляя о новом понимании, в 1942 году Чарльз Шеррингтон визуализировал работу мозга, пробуждающегося от сна:
Великий верхний лист массы, в котором вряд ли свет мерцал или двигался, теперь становится блестящим полем ритмичных мигающих точек с поездами путешествий, спешащих сюда и туда -сюда. Мозг пробуждается, и с ним ум возвращается. Как будто Млечный путь вошел в какой -то космический танец. Быстро масса головы становится зачарованным ткацким станками, где миллионы мигающих шаттлов плетения растворяющихся рисунка, всегда значимой схемой, хотя и никогда не упреждающим; Изменяющаяся гармония подчиненных.
- Шеррингтон, 1942, человек на своей природе [ 137 ]
Изобретение электронных компьютеров в 1940 -х годах, наряду с разработкой теории математической информации , привело к осознанию того, что мозги могут быть поняты как системы обработки информации. Эта концепция сформировала основу области кибернетики и в конечном итоге породила область, которая теперь известна как вычислительная нейробиология . [ 138 ] Самые ранние попытки кибернетики были несколько грубыми в том смысле, что они рассматривали мозг как по существу замаскированный цифровой компьютер, как, например, в книге Джона фон Неймана 1958 года «Компьютер и мозг» . [ 139 ] Однако на протяжении многих лет накопление информации о электрических реакциях клеток мозга, зарегистрированных от поведения животных, неуклонно перемещалось теоретические концепции в направлении увеличения реализма. [ 138 ]
Одним из наиболее влиятельных ранних вкладов была статья 1959 года под названием « Что глаз лягушки рассказывает мозгу лягушки : в статье изучались визуальные реакции нейронов в сетчатке и зрительный тектум лягушек и пришли к выводу, что некоторые нейроны в тектуме Лягушка подключена к объединению элементарных ответов таким образом, чтобы они функционировали как «воспринимающие ошибки». [ 140 ] Несколько лет спустя Дэвид Хубель и Торстен Визель обнаружили клетки в первичной визуальной коре обезьян, которые становятся активными, когда острые края перемещаются по конкретным пунктам в области зрения - открытие, за которое они выиграли Нобелевскую премию. [ 141 ] Последующие исследования в областях зрения высшего порядка обнаружили клетки, которые обнаруживают бинокулярное неравенство , цвет, движение и аспекты формы, с областями, расположенными на увеличенных расстояниях от первичной зрительной коры, показывающими все более сложные ответы. [ 142 ] Другие исследования областей мозга, не связанных с зрением, выявили клетки с широким спектром коррелятов ответа, некоторые связанные с памятью, некоторые с абстрактными типами познания, такие как пространство. [ 143 ]
Теоретики работали над пониманием этих шаблонов ответа, построив математические модели нейронов и нейронные сети , которые могут быть смоделированы с использованием компьютеров. [ 138 ] Некоторые полезные модели являются абстрактными, фокусирующиеся на концептуальной структуре нейронных алгоритмов, а не на деталях того, как они реализованы в мозге; Другие модели пытаются включить данные о биофизических свойствах реальных нейронов. [ 144 ] Однако ни одна модель на любом уровне не считается полностью достоверным описанием функции мозга. Основная трудность заключается в том, что сложные вычисления нейронных сетей требуют распределенной обработки, при которой сотни или тысячи нейронов работают совместно - текущие методы записи активности мозга способны выделять потенциалы действия только из нескольких десятков нейронов за раз. [ 145 ]
Кроме того, даже отдельные нейроны, по -видимому, являются сложными и способны выполнять вычисления. [ 146 ] Таким образом, модели мозга, которые не отражают это, слишком абстрактны, чтобы быть репрезентативными для работы мозга; Модели, которые пытаются запечатлеть это, очень дорогие вычислительно и, возможно, неразрешимы с настоящими вычислительными ресурсами. Тем не менее, проект человеческого мозга пытается создать реалистичную, подробную вычислительную модель всего человеческого мозга. Мудрость этого подхода была публично оспаривалась, с высокопоставленными учеными по обе стороны аргумента.
Во второй половине 20 -го века разработки химии, электронной микроскопии, генетики, информатики, функциональной визуализации мозга и других областей постепенно открывали новые окна в структуру и функции мозга. В Соединенных Штатах 1990 -е годы были официально обозначены как « десятилетие мозга », чтобы отметить достижения в области исследований мозга и содействовать финансированию для таких исследований. [ 147 ]
В 21 -м веке эти тенденции продолжались, и несколько новых подходов стали известны, включая многоэлектродную запись , которая позволяет регистрировать активность многих клеток мозга одновременно; [ 148 ] Генетическая инженерия , которая позволяет экспериментально изменять молекулярные компоненты мозга; [ 129 ] Геномика , которая позволяет коррелировать изменения в структуре мозга с вариациями в свойствах ДНК и нейровизуализации . [ 149 ]
Общество и культура
Как еда

Животные мозги используются в качестве пищи в многочисленных кухнях.
В ритуалах
Некоторые археологические данные свидетельствуют о том, что траурные ритуалы европейских неандертальцев также включали потребление мозга. [ 150 ]
Передние , люди Папуа -Новой Гвинеи как известно, едят человеческие мозги. В похоронных ритуалах те, кто близко к мертвым съел мозг умершего, чтобы создать чувство бессмертия . Прионная болезнь , называемая Куру, была прослежена на это. [ 151 ]
Смотрите также
Ссылки
- ^ Саладин, Кеннет (2011). Анатомия человека (3 -е изд.). МакГроу-Хилл. п. 416. ISBN 978-0-07-122207-5 .
- ^ фон Бартельд, CS; Бахни, J; Herculano-Houzel, S (15 декабря 2016 г.). «Поиск истинного числа нейронов и глиальных клеток в мозге человека: обзор 150 -летнего подсчета клеток» . Журнал сравнительной неврологии . 524 (18): 3865–3895. doi : 10.1002/cne.24040 . ISSN 0021-9967 . PMC 5063692 . PMID 27187682 .
- ^ Юсте, Рафаэль; Церковь, Джордж М. (март 2014 г.). «Новый век мозга» (PDF) . Scientific American . 310 (3): 38–45. Bibcode : 2014sciam.310c..38y . doi : 10.1038/Scientificamerican0314-38 . PMID 24660326 . Архивировано из оригинала (PDF) 2014-07-14.
- ^ Jump up to: а беременный в Shepherd, GM (1994). Нейробиология . Издательство Оксфордского университета. п. 3 ISBN 978-0-19-508843-4 .
- ^ Sporns, O (2010). Сети мозга . MIT Press. п. 143. ISBN 978-0-262-01469-4 .
- ^ Başar, E (2010). Благословное тело в туманной картезианской системе: целостный подход по колебаниям . Спрингер. п. 225. ISBN 978-1-4419-6134-1 .
- ^ Сингх, Индербир (2006). «Краткий обзор методов, используемых при изучении нейроанатомии» . Учебник нейроанатомии человека (7 -е изд.). Jaypee Brothers. п. 24. ISBN 978-81-8061-808-6 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В х и Кандель, Эрик Р.; Шварц, Джеймс Харрис; Джесселл, Томас М. (2000). Принципы нейронной науки (4 -е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-8385-7701-1 Полем OCLC 42073108 .
- ^ Дуглас, RJ; Мартин, К.А. (2004). «Нейрональные цепи неокортекс». Ежегодный обзор нейробиологии . 27 : 419–451. doi : 10.1146/annurev.neuro.27.070203.144152 . PMID 15217339 .
- ^ Барнетт, MW; Larkman, PM (2007). «Потенциал действия». Практическая неврология . 7 (3): 192–197. PMID 17515599 .
- ^ Jump up to: а беременный в Шепард, Гордон М. (2004). «1. Введение в синаптические схемы». Синаптическая организация мозга (5 -е изд.). Нью -Йорк, Нью -Йорк: Oxford University Press US. ISBN 978-0-19-515956-1 .
- ^ Уильямс, RW; Эрруп, К (1988). «Контроль номера нейронов». Ежегодный обзор нейробиологии . 11 : 423–453. doi : 10.1146/annurev.ne.11.030188.002231 . PMID 3284447 .
- ^ Хейзенберг, М. (2003). «Мемуары для тела грибов: от карт до моделей». Nature Reports Neuroscience . 4 (4): 266–275. doi : 10.1038/nrn1074 . PMID 12671643 . S2CID 5038386 .
- ^ Jump up to: а беременный Джейкобс, DK; Наканиши, н; Юань, D; и др. (2007). «Эволюция сенсорных структур в базальной метазоа». Интегративная и сравнительная биология . 47 (5): 712–723. Citeseerx 10.1.1.326.2233 . doi : 10.1093/icb/icm094 . PMID 21669752 .
- ^ Jump up to: а беременный Balavoine, G (2003). «Сегментированная урбилатерия: проверенный сценарий» . Интегративная и сравнительная биология . 43 (1): 137–147. doi : 10.1093/icb/43.1.137 . PMID 21680418 .
- ^ Schmidt-Rhaesa, A (2007). Эволюция систем органов . Издательство Оксфордского университета. п. 110 . ISBN 978-0-19-856669-4 .
- ^ Кристан, WB Jr.; Calabrese, RL; Friesen, WO (2005). «Нейрональный контроль над поведением пиявки». Прог -нейробиол . 76 (5): 279–327. doi : 10.1016/j.pneurobio.2005.09.004 . PMID 16260077 . S2CID 15773361 .
- ^ Барнс, Р.Д. (1987). Зоология беспозвоночных (5 -е изд.). Saunders College Pub. п. 1. ISBN 978-0-03-008914-5 .
- ^ Jump up to: а беременный Батлер, А.Б. (2000). «Аккордовая эволюция и происхождение черепников: старый мозг в новой головке» . Анатомическая запись . 261 (3): 111–125. doi : 10.1002/1097-0185 (20000615) 261: 3 <111 :: AID-AR6> 3.0.co; 2-F . PMID 10867629 .
- ^ Буллох, т; Кутч В. (1995). «Являются ли основные оценки мозга различными, главным образом, по количеству соединений или также по качеству?» Полем В Брейдбах О (ред.). Нервные системы беспозвоночных: эволюционный и сравнительный подход . Birkhäuser. п. 439. ISBN 978-3-7643-5076-5 .
- ^ «Flybrain: онлайн -атлас и база данных нервной системы Drosophila » . Архивировано из оригинала на 1998-01-09 . Получено 2011-10-14 .
- ^ Konopka, RJ; Бензер, S (1971). «Часы мутантов Drosophila melanogaster» . Прокурор Нат. Академический Наука США . 68 (9): 2112–2116. Bibcode : 1971pnas ... 68.2112K . doi : 10.1073/pnas.68.9.2112 . PMC 389363 . PMID 5002428 .
- ^ Шин, Хи-Суп; и др. (1985). «Необычная кодирующая последовательность из гена часов Drosophila сохраняется у позвоночных» . Природа . 317 (6036): 445–448. Bibcode : 1985natur.317..445s . doi : 10.1038/317445A0 . PMID 2413365 . S2CID 4372369 .
- ^ Гейзенберг, м; Heusipp, M; Wanke, C. (1995). «Структурная пластичность в мозгу дрозофилы» . J. Neurosci . 15 (3): 1951–1960. doi : 10.1523/jneurosci.15-03-01951.1995 . PMC 6578107 . PMID 7891144 .
- ^ Бреннер, Сидней (1974). «Генетика Caenorhabditis elegans» . Генетика . 77 (1): 71–94. doi : 10.1093/Genetics/77.1.71 . PMC 1213120 . PMID 4366476 .
- ^ Hobert, O (2005). Исследовательское сообщество C. elegans (ред.). «Спецификация нервной системы» . Wormbook : 1–19. doi : 10.1895/Wormbook.1.12.1 . PMC 4781215 . PMID 18050401 .
- ^ Белый, jg; Southgate, E ; Томсон, JN; Бреннер С. (1986). «Структура нервной системы нематодного каенорхабдита элеганс». Философские транзакции Королевского общества б . 314 (1165): 1–340. Bibcode : 1986rsptb.314 .... 1w . doi : 10.1098/rstb.1986.0056 . PMID 22462104 .
- ^ Jabr, Ferris (2012-10-02). "Стоит ли на картирование ума червя?" Полем Scientific American . Получено 2014-01-18 .
- ^ Ходжкин Дж. (2001). « Caenorhabditis elegans ». В Brenner S, Miller JH (Eds.). Энциклопедия генетики . Elsevier. С. 251–256. ISBN 978-0-12-227080-2 .
- ^ Kandel, ER (2007). В поисках памяти: появление новой науки ума . WW Norton. С. 145–150 . ISBN 978-0-393-32937-7 .
- ^ Шу, Д.-Г.; Conway Morris, S.; Хан, Дж.; Zhang, Z.-f.; Yasui, K.; Janvier, P.; Chen, L.; Zhang, X.-L.; Лю, Дж. Н.; и др. (2003). «Голова и магистраль ранних кембрийских позвоночных Хайкухтис ». Природа . 421 (6922): 526–529. Bibcode : 2003natur.421..526s . doi : 10.1038/nature01264 . PMID 12556891 . S2CID 4401274 .
- ^ Стидтер, GF (2005). «Гл. 3: Сохранение в мозге позвоночных». Принципы эволюции мозга . Sinauer Associates. ISBN 978-0-87893-820-9 .
- ^ Армстронг, Е (1983). «Относительный размер мозга и метаболизм у млекопитающих». Наука . 220 (4603): 1302–1304. Bibcode : 1983sci ... 220.1302a . doi : 10.1126/science.6407108 . PMID 6407108 .
- ^ Джерисон, Гарри Дж. (1973). Эволюция мозга и интеллекта . Академическая пресса. С. 55–74. ISBN 978-0-12-385250-2 .
- ^ Родитель, а; Carpenter, MB (1996). "Ch. 1". Нейроанатомия человека плотника . Уильямс и Уилкинс. ISBN 978-0-683-06752-1 .
- ^ Пардридж, В. (2005). «Кроваво-мозговый барьер: узкое место при развитии мозговых лекарств» . Нейрорус . 2 (1): 3–14. doi : 10.1602/neuroRx.2.1.3 . PMC 539316 . PMID 15717053 .
- ^ Northcutt, RG (2008). «Эволюция переднего мозга у костных рыб». Бюллетень исследования мозга . 75 (2–4): 191–205. doi : 10.1016/j.brainresbull.2007.10.058 . PMID 18331871 . S2CID 44619179 .
- ^ Райнер, а; Ямамото, К; Karten, HJ (2005). «Организация и эволюция переднего мозга птиц» . Анатомическая запись Часть A: открытия в молекулярной, клеточной и эволюционной биологии . 287 (1): 1080–1102. doi : 10.1002/ar.a.20253 . PMID 16206213 .
- ^ Зигель, а; Sapru, HN (2010). Основная нейробиология . Липпинкотт Уильямс и Уилкинс. С. 184–189 . ISBN 978-0-7817-8383-5 .
- ^ Swaab, Dick F. (2003). Гипоталамус человека - основные и клинические аспекты: ядра гипоталамуса человека. Часть я . Elsevier. ISBN 9780444514905 Полем Получено 2021-01-22 .
- ^ Джонс, Эдвард Г. (1985). Таламус . Мичиганский университет: Plenum Press. ISBN 9780306418563 .
- ^ Книрим, Джеймс. "Cerebellum (раздел 3, глава 5)" . Нейробиология онлайн . Кафедра нейробиологии и анатомии в Центре медицинских наук Техасского университета в Хьюстоне, Медицинская школа Макговерна. Архивировано из оригинала 2017-11-18 . Получено 22 января 2021 года .
- ^ Сайто, К; Менард, а; Грилнер, S (2007). «Тектальный контроль перед передвижением, рулевым управлением и движениями глаз в миглере». Журнал нейрофизиологии . 97 (4): 3093–3108. doi : 10.1152/jn.00639.2006 . PMID 17303814 .
- ^ Ричард Сванн закладывается; Гарри Берр Феррис; Джордж Ховард Паркер; Джеймс Роуленд Энджелл; Альберт Галлоуэй Келлер; Эдвин Грант Конклин (1922). Эволюция человека: серия лекций, проведенных перед Йельским главой «Сигма XI» в течение 1921–1922 гг . Издательство Йельского университета. п. 50
- ^ Puelles, L (2001). «Мысли о развитии, структуре и эволюции млекопитающего и птичьего телефона» . Философские транзакции Королевского общества б . 356 (1414): 1583–1598. doi : 10.1098/rstb.2001.0973 . PMC 1088538 . PMID 11604125 .
- ^ Salas, c; Broglio, c; Родригес, Ф. (2003). «Эволюция переднего мозга и пространственного познания у позвоночных: сохранение по всему разнообразию». Мозг, поведение и эволюция . 62 (2): 72–82. doi : 10.1159/000072438 . PMID 12937346 . S2CID 23055468 .
- ^ Jump up to: а беременный Грилнер, S; и др. (2005). «Механизмы для выбора основных моторных программ - Roles для стриатума и паллидума». Тенденции в нейронауках . 28 (7): 364–370. doi : 10.1016/j.tins.2005.05.004 . PMID 15935487 . S2CID 12927634 .
- ^ Northcutt, RG (1981). «Эволюция трентенфалона у нельммалей». Ежегодный обзор нейробиологии . 4 : 301–350. doi : 10.1146/annurev.ne.04.030181.001505 . PMID 7013637 .
- ^ Рейтер, Сэм; Лиав, Хуа-Пэн; Ямаваки, Трейси М.; Науманн, Роберт К.; Лоран, Жиль (2017). «О значении рептильных мозгов для картирования эволюции образования гиппокампа» . Мозг, поведение и эволюция . 90 (1): 41–52. doi : 10.1159/000478693 . ISSN 0006-8977 . PMID 28866680 .
- ^ «Статистика видов август 2019» . www.reptile-database.org . Получено 2022-12-06 .
- ^ «Красный список Ужасных видов МСОП. Версия 2022-1 - Сводная статистика» . Красный список IUCN . 2022. ISSN 2307-8235 . Получено 6 декабря 2022 года .
- ^ Jump up to: а беременный в Нура, Тадаши; Кавагучи, это правильно; Oneo, Catchric; Мы прекрасны, Аасаноро (март 2013 г.). Исследование: Resterch Журнал экспериментальной зоологии Часть B: молекулярная эволюция и развитие . 320 (2): 57–7 doi : 10.1002/jez.b.22484 . PMID 23319423 .
- ^ Салас, Косме; Броглио, Кристина; Родригес, Фернандо (2003). «Эволюция переднего мозга и пространственного познания у позвоночных: сохранение по всему разнообразию» . Мозг, поведение и эволюция . 62 (2): 72–82. doi : 10.1159/000072438 . ISSN 0006-8977 . PMID 12937346 .
- ^ Jump up to: а беременный Northcutt, R. Glenn (2013). «Различия в мозге рептилии и познания» . Мозг, поведение и эволюция . 82 (1): 45–54. doi : 10.1159/000351996 . ISSN 0006-8977 . PMID 23979455 .
- ^ Jump up to: а беременный в Науманн, Роберт К.; Ondracek, Janie M.; Рейтер, Самуил; Шейн-Идельсон, Марк; Тошс, Мария Антониетта; Ямаваки, Трейси М.; Лоран, Жиль (2015-04-20). «Рептильный мозг» . Текущая биология . 25 (8): R317 - R321. BIBCODE : 2015CBIO ... 25.R317N . doi : 10.1016/j.cub.2015.02.049 . ISSN 0960-9822 . PMC 4406946 . PMID 25898097 .
- ^ Jump up to: а беременный в Хейн, Дэвид; Gallego-Flores, Tatiana; Клинкманн, Микаэла; Масиас, Анджелес; Ciirdaeva, Елена; Арендс, Анжа; ТУМ, Кристина; Тушев, Георги; Кречмер, Фридрих; Тошс, Мария Антониетта; Лоран, Жиль (2022-09-02). «Молекулярное разнообразие и эволюция типов нейронов в мозге амниота» . Наука . 377 (6610): EABP8202. doi : 10.1126/science.abp8202 . ISSN 0036-8075 . PMID 36048944 .
- ^ Тошс, Мария Антониетта; Ямаваки, Трейси М.; Науманн, Роберт К.; Якоби, Ариэль А.; Тушев, Георги; Лоран, Жиль (2018-05-25). «Эволюция паллия, гиппокампа и типов кортикальных клеток, выявленных с помощью одноклеточной транскриптомики в рептилиях» . Наука . 360 (6391): 881–888. Bibcode : 2018sci ... 360..881t . doi : 10.1126/science.aar4237 . ISSN 0036-8075 . PMID 29724907 .
- ^ Блантон, Марк Г.; Кригштейн, Арнольд Р. (1991-08-22). «Морфологическая дифференциация различных классов нейронов в коре головного мозга эмбриональной черепахи» . Журнал сравнительной неврологии . 310 (4): 550–570. doi : 10.1002/cne.903100405 . ISSN 0021-9967 . PMID 1719040 .
- ^ Jump up to: а беременный в дюймовый и фон Уильям, Батлер, Энн Б. Ходос (2005). Сравнительная нейроанатомия позвоночных: эволюция и адаптация . Wiley-Liss. ISBN 0-471-21005-6 Полем OCLC 489018202 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный Northcutt, RG (2002). «Понимание эволюции мозга позвоночных» . Интегративная и сравнительная биология . 42 (4): 743–756. doi : 10.1093/icb/42.4.743 . PMID 21708771 .
- ^ Jump up to: а беременный Бартон, Ра; Харви, PH (2000). «Мозаичная эволюция структуры мозга у млекопитающих». Природа . 405 (6790): 1055–1058. Bibcode : 2000natur.405.1055b . doi : 10.1038/35016580 . PMID 10890446 . S2CID 52854758 .
- ^ Aboitiz, f; Моралес, D; Монтиэль, Дж. (2003). «Эволюционное происхождение изокортекса млекопитающих: к интегрированному развитию и функциональному подходу». Поведенческие и мозговые науки . 26 (5): 535–552. doi : 10.1017/s0140525x03000128 . PMID 15179935 . S2CID 6599761 .
- ^ Ромер, как; Парсонс, Т.С. (1977). Тело позвоночных . Холт-Саундерс International. п. 531. ISBN 978-0-03-910284-5 .
- ^ Jump up to: а беременный Рот, г; Dicke, U (2005). «Эволюция мозга и интеллекта». Тенденции в когнитивных науках . 9 (5): 250–257. doi : 10.1016/j.tics.2005.03.005 . PMID 15866152 . S2CID 14758763 .
- ^ Jump up to: а беременный Марино, Лори (2004). «Эволюция мозга китообразных: умножение генерирует сложность» (PDF) . Международное общество сравнительной психологии (17): 1–16. Архивировано из оригинала (PDF) 2018-09-16 . Получено 2010-08-29 .
- ^ Шошани, J; Купский, WJ; Marchant, GH (2006). «Слонский мозг Часть I: грубая морфология, функции, сравнительная анатомия и эволюция». Бюллетень исследования мозга . 70 (2): 124–157. doi : 10.1016/j.brainresbull.2006.03.016 . PMID 16782503 . S2CID 14339772 .
- ^ Finlay, Bl; Дарлингтон, РБ; Nicastro, N (2001). «Структура развития в эволюции мозга». Поведенческие и мозговые науки . 24 (2): 263–308. doi : 10.1017/s0140525x01003958 . PMID 11530543 . S2CID 20978251 .
- ^ Calvin, William H. (1996). Как думают мозги (1 -е изд.). Нью -Йорк, Нью -Йорк: Базовые книги. ISBN 978-0-465-07278-1 .
- ^ Серено, Мичиган; Дейл, Ам; Реппас, Ам; Kwong, KK; Belliveau, JW; Брэди, TJ; Розен, Бр; Tootell, RBH (1995). «Границы нескольких визуальных областей у человека, выявленные функциональной магнитно -резонансной томографией» (PDF) . Наука . 268 (5212): 889–893. Bibcode : 1995sci ... 268..889s . doi : 10.1126/science.7754376 . PMID 7754376 . Архивировано (PDF) из оригинала 2006-05-23.
- ^ Fuster, Хоакин М. (2008). Префронтальная кора (4 -е изд.). Elsevier. стр. 1–7 . ISBN 978-0-12-373644-4 .
- ^ Jump up to: а беременный в дюймовый и фон глин Purves, Dale.; Лихтман, Джефф В. (1985). Принципы развития нейронного развития . Sunderland, Mass.: Sinauer Associates. ISBN 978-0-87893-744-8 Полем OCLC 10798963 .
- ^ Jump up to: а беременный Вонг, Ро (1999). «Волны сетчатки и развитие визуальной системы». Ежегодный обзор нейробиологии . 22 Сент -Луис, Миссури: 29–47. doi : 10.1146/annurev.neuro.22.1.29 . PMID 10202531 .
- ^ Ракич, Паско (2002). «Взрослый нейрогенез у млекопитающих: кризис идентичности» . Журнал нейробиологии . 22 (3): 614–618. doi : 10.1523/jneurosci.22-03-00614.2002 . PMC 6758501 . PMID 11826088 .
- ^ Ридли, Мэтт (2004). Природа через воспитание: гены, опыт и то, что делает нас людьми . HarperCollins. С. 1–6. ISBN 978-0-06-000678-5 .
- ^ Визель Т. (1982). «Постнатальное развитие зрительной коры и влияние окружающей среды» (PDF) . Природа . 299 (5884): 583–591. Bibcode : 1982natur.299..583w . Citeseerx 10.1.1.547.7497 . doi : 10.1038/299583a0 . PMID 6811951 . S2CID 38776857 . Архивировано (PDF) из оригинала на 2022-10-09.
- ^ Ван Прааг, ч; Кемперманн, G; Гейдж, Ф.Х. (2000). «Нейронные последствия обогащения окружающей среды». Nature Reports Neuroscience . 1 (3): 191–198. doi : 10.1038/350445558 . PMID 11257907 . S2CID 9750498 .
- ^ Купер, младший; Bloom, Fe; Roth, RH (2003). Биохимическая основа нейрофармакологии . Oxford University Press США. ISBN 978-0-19-514008-8 .
- ^ МакГер, PL; McGeer, EG (1989). «Глава 15, аминокислотные нейротрансмиттеры ». В Г. Сигеле; и др. (ред.). Основная нейрохимия . Мичиганский университет: Raven Press. С. 311–332. ISBN 978-0-88167-343-2 .
- ^ Foster, AC; Кемп, JA (2006). «Глютамат и ГАМК-терапия CNS». Современное мнение о фармакологии . 6 (1): 7–17. doi : 10.1016/j.coph.2005.11.005 . PMID 16377242 .
- ^ Фрейзер, а; Хенслер, JG (1999). «Понимание нейроанатомической организации серотонинергических клеток в мозге дает представление о функциях этого нейротрансмиттера». В Зигеле, GJ (ред.). Основная нейрохимия (шестое изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0-397-51820-3 .
- ^ Мехлер, MF; Purpura, DP (2009). «Аутизм, лихорадка, эпигенетика и локус coeruleus» . Обзоры исследований мозга . 59 (2): 388–392. doi : 10.1016/j.brainresrev.2008.11.001 . PMC 2668953 . PMID 19059284 .
- ^ Rang, HP (2003). Фармакология . Черчилль Ливингстон. С. 476–483. ISBN 978-0-443-07145-4 .
- ^ Speckmann EJ, Elger CE (2004). «Введение в нейрофизиологическую основу потенциалов ЭЭГ и постоянного тока». В Niedermeyer E, Лопес да Силва Ф.Х. (ред.). Электроэнцефалография: основные принципы, клинические применения и связанные с ней области . Липпинкотт Уильямс и Уилкинс. С. 17–31. ISBN 978-0-7817-5126-1 .
- ^ Jump up to: а беременный Бузсаки, Гьорги (2006). Ритмы мозга . Издательство Оксфордского университета. ISBN 9780199828234 .
- ^ Jump up to: а беременный в Newenhuys, R; Темный, HJ; Никулсон, С. (1998). Центральная нервная система или вербов, том 1 . Спрингер. стр. 11–14. ISBN 978-3-540-56013-5 .
- ^ Сафи, К; Seid, MA; Дехманн, Д.К. (2005). «Больше не всегда лучше: когда мозг становится меньше» . Биологические письма . 1 (3): 283–286. doi : 10.1098/rsbl.2005.0333 . PMC 1617168 . PMID 17148188 .
- ^ Норка, JW; Blumenschine, RJ; Адамс, Д.Б. (1981). «Соотношение центральной нервной системы к метаболизму тела у позвоночных: его постоянство и функциональная основа» . Американский журнал физиологии (представленная рукопись). 241 (3): R203–212. doi : 10.1152/ajpregu.1981.241.3.r203 . PMID 7282965 . Архивировано из оригинала 2020-08-17 . Получено 2021-02-10 .
- ^ Райхл, м; Гуснард Д.А. (2002). «Оценка энергетического бюджета мозга» . Прокурор Нат. Академический Наука США . 99 (16): 10237–10239. Bibcode : 2002pnas ... 9910237r . doi : 10.1073/pnas.172399499 . PMC 124895 . PMID 12149485 .
- ^ Мехагнул-Шиппер, диджей; Van der Kallen, bf; Colier, WNJM; Van der sluijs, mc; Ван Эрнинг, LJ; Тиджссен, хо; Озебург, б; Hoefnagels, WH; Янсен, RW (2002). «Одновременные измерения изменений оксигенации головного мозга во время активации мозга с помощью спектроскопии в ближней инфракрасной линии и функциональной магнитно-резонансной томографии у здоровых молодых и пожилых людей» . Гум мозга Мапп . 16 (1): 14–23. doi : 10.1002/hbm.10026 . PMC 6871837 . PMID 11870923 .
- ^ Эберт, Д.; Haller, Rg.; Уолтон, я. (Июль 2003). «Вклад энергетики октанотата в интактный метаболизм головного мозга крысы, измеренный с помощью ядерной магнитно -резонансной спектроскопии 13C» . J Neurosci . 23 (13): 5928–5935. doi : 10.1523/jneurosci.23-13-05928.2003 . PMC 6741266 . PMID 12843297 .
- ^ Marin-Valencia, I.; Хорошо, lb.; MA, Q.; Malloy, Cr.; Pascual, JM. (Февраль 2013 г.). «Гептаноат как нейронное топливо: энергетические и нейротрансмиттерные предшественники в нормальном и глюкозовом транспортере I-дефицитно (G1D) мозга» . J Cereb Metab . 33 (2): 175–182. doi : 10.1038/jcbfm.2012.151 . PMC 3564188 . PMID 23072752 .
- ^ Boumezbeur, F.; Петерсен, Kf.; Cline, GW.; Мейсон, Гф.; Бехар, Кл.; Шульман, Ги.; Ротман, Д.Л. (Октябрь 2010). «Вклад лактата крови в метаболизм энергии мозга у людей, измеренный с помощью динамической 13C -ядерной магнитно -резонансной спектроскопии» . J Neurosci . 30 (42): 13983–13991. doi : 10.1523/jneurosci.2040-10.2010 . PMC 2996729 . PMID 20962220 .
- ^ Deelchand, Dk.; Shestov, AA.; Коски, Д.М.; Uğurbil, K.; Генри, стр. (Май 2009 г.). «Транспорт и использование ацетата в мозге крысы» . J Нейрохим . 109 (Suppl 1): 46–54. doi : 10.1111/j.1471-4159.2009.05895.x . PMC 2722917 . PMID 19393008 .
- ^ Soengas, JL; Aldegunde, M (2002). «Энергетический метаболизм рыбного мозга». Сравнительная биохимия и физиология б . 131 (3): 271–296. doi : 10.1016/s1096-4959 (02) 00022-2 . PMID 11959012 .
- ^ Jump up to: а беременный Carew, TJ (2000). "Ch. 1" . Поведенческая нейробиология: клеточная организация естественного поведения . Sinauer Associates. ISBN 978-0-87893-092-0 .
- ^ Дафни Н. "Анатомия спинного мозга" . Нейробиология онлайн. Архивировано из оригинала 2011-10-08 . Получено 2011-10-10 .
- ^ Dragoi, V. "Окулярная моторная система" . Нейробиология онлайн. Архивировано из оригинала 2011-11-17 . Получено 2011-10-10 .
- ^ Герни, К; Прескотт, TJ; Wickens, Jr; Redgrave, P (2004). «Вычислительные модели базальных ганглиев: от роботов до мембран». Тенденции в нейронауках . 27 (8): 453–459. doi : 10.1016/j.tins.2004.06.003 . PMID 15271492 . S2CID 2148363 .
- ^ Книрим, Джеймс. «Моторная кора (раздел 3, глава 3)» . Нейробиология онлайн . Кафедра нейробиологии и анатомии в Центре медицинских наук Техасского университета в Хьюстоне, Медицинская школа Макговерна . Получено 2021-01-23 .
- ^ Shima, K; Танджи, Дж. (1998). «Как дополнительные, так и предполагаемые двигательные зоны имеют решающее значение для временной организации нескольких движений». Журнал нейрофизиологии . 80 (6): 3247–3260. doi : 10.1152/jn.1998.80.6.3247 . PMID 9862919 .
- ^ Миллер, Эк; Коэн, JD (2001). «Интегративная теория префронтальной функции коры». Ежегодный обзор нейробиологии . 24 (1): 167–202. doi : 10.1146/annurev.neuro.24.1.167 . PMID 11283309 . S2CID 7301474 .
- ^ Анл, MC; Сильвер, Р. (2005). «Время оркестровки: расположение циркадных часов мозга» (PDF) . Тенденции в нейронауках . 28 (3): 145–151. doi : 10.1016/j.tins.2005.01.003 . PMID 15749168 . S2CID 10618277 . Архивировано из оригинала (PDF) на 2008-10-31.
- ^ Клейтман, Натаниэль (1939). Сон и бодрствование . Пересмотренное и расширенное издание 1963 года, переиздание издания 1987 года. Чикаго: Университет Чикагской Прессы, Мидуэй Перепечатка. ISBN 978-0-226-44073-6 .
- ^ Jump up to: а беременный в Догерти, Патрик. «Гипоталамус: структурная организация» . Нейробиология онлайн . Архивировано из оригинала 2011-11-17 . Получено 2011-10-11 .
- ^ Гросс, Чарльз Г. (1998). «Клод Бернард и постоянство внутренней среды» (PDF) . Нейробиолог . 4 (5): 380–385. doi : 10.1177/107385849800400520 . S2CID 51424670 . Архивировано из оригинала (PDF) 2018-12-08.
- ^ Догерти, Патрик. «Гипоталамический контроль гормона гипофиза» . Нейробиология онлайн . Архивировано из оригинала 2011-11-17 . Получено 2011-10-11 .
- ^ Чил, HJ; Пиво, RD (1997). «У мозга есть тело: адаптивное поведение возникает из -за взаимодействия нервной системы, тела и окружающей среды». Тенденции в нейронауках . 20 (12): 553–557. doi : 10.1016/s0166-2236 (97) 01149-1 . PMID 9416664 . S2CID 5634365 .
- ^ Берридж, KC (2004). «Мотивационные концепции в поведенческой нейробиологии». Физиология и поведение . 81 (2): 179–209. doi : 10.1016/j.physbeh.2004.02.004 . PMID 15159167 . S2CID 14149019 .
- ^ Ардиэль, Эль; Ранкин, Ч. (2010). «Элегантный ум: обучение и память у Caenorhabditis elegans » . Обучение и память . 17 (4): 191–201. doi : 10.1101/lm.960510 . PMID 20335372 .
- ^ Хайман, SE; Malenka, RC (2001). «Зависимость и мозг: нейробиология принуждения и ее устойчивость» . Nature Reports Neuroscience . 2 (10): 695–703. doi : 10.1038/35094560 . PMID 11584307 . S2CID 3333114 .
- ^ Ramón Y Cajal, S (1894). «Лекция Круняна: La Fine Structure Des Centres Nerveux» . Труды Королевского общества . 55 (331–335): 444–468. Bibcode : 1894rsps ... 55..444c . doi : 10.1098/rspl.1894.0063 .
- ^ Lømo, T (2003). «Открытие долгосрочного потенцирования» . Философские транзакции Королевского общества б . 358 (1432): 617–620. doi : 10.1098/rstb.2002.1226 . PMC 1693150 . PMID 12740104 .
- ^ Маленка, R; Bear, M (2004). «Ltp и Ltd: смущение богатства» . Нейрон . 44 (1): 5–21. doi : 10.1016/j.neuron.2004.09.012 . PMID 15450156 . S2CID 79844 .
- ^ Jump up to: а беременный Бос, я; De Boever, P; Int panis, l; Meeusen, R (2004). «Физическая активность, загрязнение воздуха и мозг» . Спортивная медицина . 44 (11): 1505–1518. doi : 10.1007/s40279-014-0222-6 . PMID 25119155 . S2CID 207493297 .
- ^ Кертис, CE; D'esposito, M (2003). «Постоянная деятельность в префронтальной коре во время рабочей памяти». Тенденции в когнитивных науках . 7 (9): 415–423. Citeseerx 10.1.1.457.9723 . doi : 10.1016/s1364-6613 (03) 00197-9 . PMID 12963473 . S2CID 15763406 .
- ^ Tulving, e; Markowitsch, HJ (1998). «Эпизодическая и декларативная память: роль гиппокампа» . Гиппокамп . 8 (3): 198–204. doi : 10.1002/(SICI) 1098-1063 (1998) 8: 3 <198 :: AID-HIPO2> 3.0.CO; 2-G . PMID 9662134 . S2CID 18634842 .
- ^ Мартин, а; Chao, LL (2001). «Семантическая память и мозг: структуры и процессы». Современное мнение о нейробиологии . 11 (2): 194–201. doi : 10.1016/s0959-4388 (00) 00196-3 . PMID 11301239 . S2CID 3700874 .
- ^ Balleine, BW; Лилджехольм, Мими; Ostlund, SB (2009). «Интегративная функция базальных ганглиев в инструментальном обучении». Поведенческое исследование мозга . 199 (1): 43–52. doi : 10.1016/j.bbr.2008.10.034 . PMID 19027797 . S2CID 36521958 .
- ^ Доя, К (2000). «Дополнительные роли базальных ганглиев и мозжечка в обучении и моторном контроле». Современное мнение о нейробиологии . 10 (6): 732–739. doi : 10.1016/s0959-4388 (00) 00153-7 . PMID 11240282 . S2CID 10962570 .
- ^ Сторрор, Хью А. (1969). Схема клинической психиатрии . Нью-Йорк: Appleton-Century-Crofts, образовательное подразделение. ISBN 978-0-390-85075-1 Полем OCLC 47198 .
- ^ Thagard, Paul (2007). «Когнитивная наука» . Стэнфордская энциклопедия философии (пересмотрено, 2 -е изд.) . Получено 2021-01-23 .
- ^ Медведь, МФ; Коннорс, BW; Paradiso, MA (2007). "Ch. 2". Нейробиология: изучение мозга . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-6003-4 .
- ^ Доулинг, JE (2001). Нейроны и сети . Гарвардский университет издательство. С. 15–24. ISBN 978-0-674-00462-7 .
- ^ Wyllie, E; Гупта, а; Lachhwani, DK (2005). "Ch. 77". Лечение эпилепсии: принципы и практика . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-4995-4 .
- ^ Laureys S, Boly M, Tononi G (2009). «Функциональное нейровизуализацию». В Laureys S, Tononi G (Eds.). Неврология сознания: когнитивная неврология и невропатология . Академическая пресса. С. 31 –42. ISBN 978-0-12-374168-4 .
- ^ Кармен, JM; и др. (2003). «Обучение контролю над границей мозга и макин для достижения и схватки приматами» . PLOS Биология . 1 (2): 193–208. doi : 10.1371/journal.pbio.0000042 . PMC 261882 . PMID 14624244 .
- ^ Колб, б; Whishaw, I (2008). "Ch. 1". Основы нейропсихологии человека . Макмиллан. ISBN 978-0-7167-9586-5 .
- ^ Эббот, LF; Даян, П (2001). "Предисловие". Теоретическая нейробиология: вычислительное и математическое моделирование нейронных систем . MIT Press. ISBN 978-0-262-54185-5 .
- ^ Jump up to: а беременный в Tonegawa, s; Наказава, K; Уилсон, Массачусетс (2003). «Генетическая нейробиология обучения и памяти млекопитающих» . Философские транзакции Королевского общества б . 358 (1432): 787–795. doi : 10.1098/rstb.2002.1243 . PMC 1693163 . PMID 12740125 .
- ^ Emani, PS; и др. (2024). «Одноклеточная геномика и регуляторные сети для 388 человеческих мозгов» . Наука . 384 . doi : 10.1126/science.adi5199 . PMC 11365579 . PMID 38781369 .
- ^ Бауэр, Брюс (2009-01-12). «Армянская пещера дает древний человеческий мозг» . Sciencenews . Получено 2021-01-23 .
- ^ Jump up to: а беременный в Finger, Stanley (2001). Происхождение нейробиологии . Издательство Оксфордского университета. С. 14–15. ISBN 978-0-19-514694-3 .
- ^ * Гиппократ (2006) [400 г. до н.э.], о священной болезни , перевод Фрэнсиса Адамса, Архив интернет -классики: Библиотека Аделаиды Университета Аделаиды, архивирована из оригинала 26 сентября 2007 г.
- ^ Bloom Fe (1975). Шмидт Ф.О., Уорден Ф.Г., Свази Дж.П., Адельман Г. (ред.). Нейронауки, пути открытия . MIT Press. п. 211 . ISBN 978-0-262-23072-8 .
- ^ Shepherd, GM (1991). "Ch.1: введение и обзор". Основы доктрины нейронов . Издательство Оксфордского университета. ISBN 978-0-19-506491-9 .
- ^ Пикколино, М. (2002). «Пятьдесят лет эпохи Ходжкин-Хаксли». Тенденции в нейронауках . 25 (11): 552–553. doi : 10.1016/s0166-2236 (02) 02276-2 . PMID 12392928 . S2CID 35465936 .
- ^ Шеррингтон, CS (1942). Человек на своей природе . Издательство Кембриджского университета. п. 178 . ISBN 978-0-8385-7701-1 .
- ^ Jump up to: а беременный в Churchland, PS; Кох, C; Sejnowski, TJ (1993). «Что такое вычислительная нейробиология?». В Шварц эль (ред.). Вычислительная нейробиология . MIT Press. С. 46–55. ISBN 978-0-262-69164-2 .
- ^ фон Нейман, J; Черчленд, премьер -министр; Churchland, PS (2000). Компьютер и мозг . Издательство Йельского университета. С. XI - XXII. ISBN 978-0-300-08473-3 .
- ^ Lettvin, JY; Матурна, HR; МакКаллох, WS; Питтс, WH (1959). «Что глаз лягушки говорит мозгу лягушки» (PDF) . Материалы института радиоинженеры . 47 (11): 1940–1951. doi : 10.1109/jrproc.1959.287207 . S2CID 8739509 . Архивировано из оригинала (PDF) 2011-09-28.
- ^ Хубель, DH; Wiesel, TN (2005). Мозг и визуальное восприятие: история 25-летнего сотрудничества . Oxford University Press США. С. 657 –704. ISBN 978-0-19-517618-6 .
- ^ Фара, MJ (2000). Когнитивная нейробиология зрения . Wiley-Blackwell. С. 1–29. ISBN 978-0-631-21403-8 .
- ^ Энгель, Ак; Singer, W (2001). «Временное связывание и нейронные корреляты сенсорного осознания». Тенденции в когнитивных науках . 5 (1): 16–25. doi : 10.1016/s1364-6613 (00) 01568-0 . PMID 11164732 . S2CID 11922975 .
- ^ Даян, P; Эббот, Л.Ф. (2005). «Ch.7: сетевые модели». Теоретическая нейробиология . MIT Press. ISBN 978-0-262-54185-5 .
- ^ Авербек, BB; Ли, Д. (2004). «Кодирование и передача информации с помощью нейронных ансамблей». Тенденции в нейронауках . 27 (4): 225–230. doi : 10.1016/j.tins.2004.02.006 . PMID 15046882 . S2CID 44512482 .
- ^ Forrest, MD (2014). «Внутриклеточная динамика кальция позволяет модели нейрона Purkinje выполнять вычисления и усиления при его входах» . Границы в вычислительной нейробиологии . 8 : 86. doi : 10.3389/fncom.2014.00086 . PMC 4138505 . PMID 25191262 .
- ^ Джонс, например; Менделл, Л.М. (1999). «Оценка десятилетия мозга». Наука . 284 (5415): 739. Bibcode : 1999sci ... 284..739J . doi : 10.1126/science.284.5415.739 . PMID 10336393 . S2CID 13261978 .
- ^ Buzsáki, G (2004). «Крупномасштабная запись нейрональных ансамблей» (PDF) . Nature Neuroscience . 7 (5): 446–451. doi : 10.1038/nn1233 . PMID 15114356 . S2CID 18538341 . Архивировано из оригинала (PDF) на 2006-09-10.
- ^ Geschwind, DH; Konopka, G (2009). «Нейронаука в эпоху функциональной геномики и системной биологии» . Природа . 461 (7266): 908–915. Bibcode : 2009natur.461..908g . doi : 10.1038/nature08537 . PMC 3645852 . PMID 19829370 .
- ^ Коннелл, Эван С. (2001). Ацтек -сокровищница . Counterpoint Press. ISBN 978-1-58243-162-8 .
- ^ Коллинз, S; Маклин СА; Masters CL (2001). «Синдром Герстманн-Страсслер-Схейнкер, смертельная семейная бессонница и Куру: обзор этих менее распространенных человеческих трансмиссивных губчатых энцефалопатий». Журнал клинической нейробиологии . 8 (5): 387–397. doi : 10.1054/jocn.2001.0919 . PMID 11535002 . S2CID 31976428 .
Внешние ссылки
- Мозг сверху донизу , в университете Макгилла
- «Мозг» , обсуждение BBC Radio 4 с Вивиан Наттон, Джонатаном Сэддей и Мариной Уоллес ( в наше время , 8 мая 2008 г.)
- Наше стремление понять мозг - с лекцией Мэтью Кобба Королевского учреждения. Архивировано в Ghostarchive .