Нематода
| Нематода Временной диапазон: Возможное кембрийское возникновение [2] | |
|---|---|
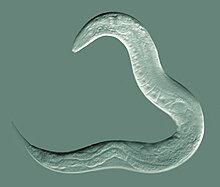 | |
| Ценорхабдитис элегантный , модельный вид круглых червей | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Подкоролевство: | Эуметазоа |
| Клэйд : | ПараХоксозоа |
| Клэйд : | Искатель |
| Клэйд : | Нефрозоа |
| (без рейтинга): | протостомия |
| Супертип: | Экдисозоа |
| Клэйд : | Нематода |
| Тип: | Нематоды Умирающий , 1861 г. |
| Классы | |
(см. текст ) | |
| Синонимы | |
| |
Нематоды ( ; греч нематоды , тип : круглые латынь . Nematoda ) Νηματώδη Nematoda или - составляют Greek: Νηματώδη; Latin: Nematoda), roundworms or eelworms constitute the phylum черви . [3] [4] Это разнообразный тип животных, обитающий в самых разных средах. Большинство видов живут свободно и питаются микроорганизмами виды , но есть и паразитические . [3] Паразитические черви ( гельминты ) являются причиной почвенных гельминтозов .
Они таксономически классифицированы наряду с членистоногими , тихоходками и другими линяющими животными в кладе Ecdysozoa . В отличие от отдаленно похожих плоских червей , нематоды имеют трубчатую пищеварительную систему с отверстиями на обоих концах. Как и тихоходки, они имеют уменьшенное количество Hox-генов , но их сестринский тип Nematomorpha сохранил предковый генотип Hox протостома, что показывает, что сокращение произошло внутри типа нематод. [5]
нематод Виды бывает трудно отличить друг от друга. Следовательно, оценки количества видов нематод неопределенны. Исследование биоразнообразия животных 2013 года, опубликованное в мегажурнале Zootaxa, показывает, что эта цифра превышает 25 000 особей. [6] [7] Оценки общего числа современных видов подвержены еще большим колебаниям. В широко цитируемой статье, опубликованной в 1993 году, говорится, что существует более 1 миллиона видов нематод. [8] Последующая публикация оспорила это утверждение, оценив, что эта цифра составляет не менее 40 000 видов. [9] Хотя самые высокие оценки (до 100 миллионов видов) с тех пор устарели, оценки, подкрепленные кривыми разрежения , [10] [11] вместе с использованием штрих-кодирования ДНК [12] и растущее признание широко распространенных криптических видов среди нематод, [13] приблизили эту цифру к 1 миллиону видов. [14]
Нематоды успешно адаптировались почти к каждой экосистеме : от морской (соленой) до пресной воды, почв, от полярных регионов до тропиков, а также от самых высоких до самых низких возвышенностей. Они повсеместно распространены в пресноводной, морской и наземной среде, где часто превосходят численностью других животных как по количеству особей, так и по видам , и встречаются в таких разнообразных местах, как горы, пустыни и океанические впадины . Земли Они встречаются во всех частях литосферы . [15] даже на больших глубинах, 0,9–3,6 км (3 000–12 000 футов) ниже поверхности Земли на золотых рудниках в Южной Африке. [16] [17] [18] [19] [20] Они представляют 90% всех животных на дне океана . [21] Всего 4,4×10 20 нематоды населяют верхний слой почвы Земли, или примерно 60 миллиардов на каждого человека, при этом самая высокая плотность наблюдается в тундре и бореальных лесах. [22] Их численное преобладание, часто превышающее миллион особей на квадратный метр и составляющее около 80% всех отдельных животных на Земле, разнообразие жизненных циклов и присутствие на различных трофических уровнях указывают на важную роль во многих экосистемах. [22] [23] Было показано, что они играют решающую роль в полярных экосистемах. [24] [25] Примерно 2271 род относится к 256 семействам . [26] Многие паразитические формы включают патогены большинства растений и животных. Треть родов встречается паразиты позвоночных как ; У человека встречается около 35 видов нематод. [26]
Натан Кобб , нематолог , так описал повсеместное распространение нематод на Земле:
Короче говоря, если бы вся материя во вселенной, за исключением нематод, была бы сметена, наш мир по-прежнему оставался бы смутно узнаваемым, и если бы мы, как бестелесные духи, могли бы затем исследовать его, мы должны были бы найти его горы, холмы, долины, реки, озера и океаны представлены пленкой нематод. Местоположение городов можно было бы расшифровать, поскольку каждому скоплению людей соответствовало бы скопление определенных нематод. Деревья по-прежнему стояли призрачными рядами, изображая наши улицы и шоссе. Местоположение различных растений и животных все еще можно было бы расшифровать, и, если бы у нас было достаточно знаний, во многих случаях даже их вид можно было бы определить путем изучения их бывших нематод-паразитов. [27] (стр. 472)
Этимология
[ редактировать ]Слово нематода происходит от современного латинского соединения nemat- «нить» (от греческого nema , родительного падежа nematos «нить», от основы nein «прясть»; см. игла ) + -odes «подобный, по природе» ( см . -oid ).
Таксономия и систематика
[ редактировать ]






История
[ редактировать ]В 1758 году Линней описал некоторые роды нематод (например, Ascaris ), включенные затем в состав Vermes .
Название группы Nematoda, неофициально называемое «нематодами», произошло от Nematoidea , первоначально определенного Карлом Рудольфи (1808 г.), [28] от древнегреческого νῆμα ( nêma, nêmatos , «нить») и -ειδἠς ( -eidēs , «вид»). (1837) отнес его к семейству нематод Бурмейстер . [28]
В состав «Nematoidea» по своему происхождению ошибочно вошли Nematodes и Nematomorpha , приписываемые фон Зибольдом (1843). Вместе с скребнями , трематодами и цестодеями образовали устаревшую группу Entozoa . [29] создан Рудольфи (1808). [30] отнес их вместе с скребнями к устаревшему типу Nemathelminthes Гегенбаур (1859) .
В 1861 г. К. М. Дизинг отнес эту группу к отряду Nematoda. [28] В 1877 году таксон до ранга типа Nematoidea, включающий семейство Gordiidae (черви из конского волоса), был повышен Рэем Ланкестером . Первое четкое различие между немасами и гордиидами было проведено Вейдовским, когда он назвал группу, содержащую конских волосатых червей, отрядом Nematomorpha. В 1919 году Натан Кобб предложил признать нематоды отдельно как отдельный тип. [31] Он утверждал, что по-английски их следует называть «нема», а не «нематоды», и определил таксон Nemate (позже исправленный как Nemata, латинское множественное число от nema ), указав Nematoidea sensustricto как синоним.
Однако в 1910 году Гроббен предложил тип Aschelminthes, и нематоды были включены в класс Nematoda вместе с классом Rotifera, классом Gastrotricha, классом Kinorhyncha, классом Priapulida и классом Nematomorpha (позже этот тип был возрожден и модифицирован Либби Генриеттой Хайман в 1951 году как Pseudoceolomata, но остались похожими). В 1932 году Поттс возвел класс Nematoda на уровень типа, оставив название прежним. Несмотря на то, что классификация Поттса эквивалентна классификации Коббса, оба названия использовались (и используются до сих пор), а нематода стала популярным термином в зоологической науке. [32]
Поскольку Кобб был первым, кто включил нематод в особый тип, отделенный от Nematomorpha, некоторые исследователи считают, что действительным названием таксона является Nemate или Nemata, а не Nematoda. [33] из-за зоологического правила, которое в случае синонимов отдает приоритет первому употребленному термину.
Филогения
[ редактировать ]Филогенетические протостомических взаимоотношения нематод и их близких родственников среди Metazoa не выяснены. Традиционно их считали отдельной линией, но в 1990-х годах было предложено образовать группу Ecdysozoa вместе с линяющими животными, такими как членистоногие . Личность ближайших ныне живущих родственников нематод всегда считалась хорошо установленной. Морфологические признаки и молекулярная филогения согласуются с тем, что круглые черви относятся к сестринскому таксону паразитических Nematomorpha ; вместе они составляют нематоиду . Наряду со Scalidophora (ранее Cephalorhyncha), Nematoida образуют кладу Cycloneuralia , но между имеющимися морфологическими и молекулярными данными возникают большие разногласия. Cycloneuralia или Introverta — в зависимости от обоснованности первого — часто причисляются к супертипу . [34]
Актуальную информацию (по состоянию на 2022 год) см. в разделе « Филогеномный анализ типа Nematoda: конфликты и совпадения с морфологией, 18S рРНК и митогеномами» .
Систематика нематод
[ редактировать ]Из-за отсутствия знаний о многих нематодах их систематика вызывает споры. Ранняя и влиятельная классификация была предложена Читвудом и Читвудом. [35] - позже отредактировано Читвудом [36] — который разделил тип на два класса — афазмидии и фазмидии . Позже они были переименованы в Adenophorea (носители желез) и Secernentea (секреторы) соответственно. [37] Secernentea имеют несколько общих характеристик, в том числе наличие фазмид , пары сенсорных органов, расположенных в латерально-задней области, и это было использовано в качестве основы для этого разделения. Этой схемы придерживались во многих более поздних классификациях, хотя аденофореи не входили в единую группу.
Первоначальные исследования неполных последовательностей ДНК [38] предположил существование пяти клад : [39]
Secernentea, по-видимому, являются естественной группой близких родственников, тогда как «Adenophorea», по-видимому, представляет собой парафилетическое собрание круглых червей, сохраняющих большое количество наследственных черт . Старая Эноплия также не кажется монофилетической, но содержит две отдельные линии. Старая группа « Chromadoria », по-видимому, представляет собой еще одну парафилетическую совокупность, причем Monhysterida представляет собой очень древнюю второстепенную группу нематод. Среди Secernentea Diplogasteria , возможно, потребуется объединить с Rhabditia , тогда как Tylenchia может быть парафилетической с Rhabditia. [40]
Понимание систематики и филогении круглых червей по состоянию на 2002 год резюмировано ниже:
Тип Нематоды
- Базальный порядок Monhysterida
- Класс Дорилаимида
- Класс Эноплея
- Класс Сецернентеа
- Подкласс Diplogasteria (оспаривается)
- Подкласс Rhabditia (парафилетический?)
- Подкласс Спирурия
- Подкласс Tylenchia (оспаривается)
- Комплекс " Хромадория "
Более поздние работы предположили наличие 12 клад. [41] Secernentea — группа, в которую входят практически все основные «нематоды»-паразиты животных и растений — по-видимому, возникла из Adenophorea.
В 2019 году исследование выявило одну консервативную сигнатуру (CSI), обнаруженную исключительно у представителей типа Nematoda посредством сравнительного генетического анализа. [42] CSI состоит из вставки одной аминокислоты в консервативной области белка фактора регулирования обмена Na (+) / H (+) NRFL-1 и является молекулярным маркером, который отличает этот тип от других видов. [42]
Совместная вики-страница под названием «959 геномов нематод» предпринимает серьезные усилия по улучшению систематики этого типа. [43]
Анализ митохондриальной ДНК показывает, что действительны следующие группы: [44]
- подкласс Дорилаимия
- отряды Рабдитиды , Трихинеллиды и Мермитиды.
- подотряд Рабдитина
- инфраотряды Spiruromorpha и Oxyuridomorpha
В 2022 г. новую классификацию всего типа Nematoda представил М. Ходда. Он был основан на современных молекулярных, онтогенетических и морфологических данных. [45] В рамках этой классификации представлены следующие классы и подклассы:
- Класс Эноплея
- Подкласс Эноплия
- Подкласс Онхолаимия
- Подкласс Триплончия
- Класс Дорилаимида
- Подкласс Дорилаимия
- Подкласс Батиодонтия
- Подкласс Трихоцефалия
- Класс Хромадория
- Подкласс Хромадория
- Подкласс Косы
Ископаемая запись
[ редактировать ]Яйца нематод из клад Ascaridina, Spirurina и Trichocephalida были обнаружены в копролитах формации Тремембе олигоценового возраста, которая представляла собой палеолазеро на территории современного Сан-Паулу с разнообразным ископаемым комплексом птиц, рыб и членистоногих, пригодных для содействие высокому разнообразию нематод. [46]
Анатомия
[ редактировать ]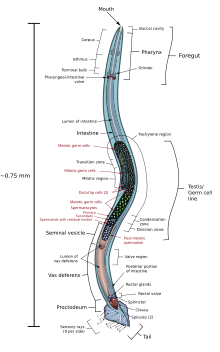
Нематоды — это очень маленькие и тонкие черви: обычно толщиной от 5 до 100 мкм и длиной от 0,1 до 2,5 мм. [47] Самые маленькие нематоды микроскопичны, в то время как свободноживущие виды могут достигать 5 см (2 дюйма), а некоторые паразитические виды еще крупнее, достигая более 1 м (3 футов) в длину. [48] : 271 Тело часто украшено гребнями, кольцами, щетинками или другими характерными структурами. [49]
Голова нематоды относительно отчетливая. В то время как остальная часть тела двусторонне симметрична, голова радиально симметрична, с сенсорными щетинками и, во многих случаях, сплошными «щитами головы», расходящимися наружу вокруг рта. Рот имеет три или шесть губ, на внутренних краях которых часто имеется ряд зубов. На кончике хвоста часто находится клейкая «хвостовая железа». [50]
Эпидермис синцитий представляет собой либо , либо один слой клеток и покрыт толстой коллагеновой кутикулой . Кутикула часто имеет сложную структуру и может состоять из двух или трех отдельных слоев. Под эпидермисом находится слой продольных мышечных клеток. Относительно жесткая кутикула совместно с мышцами создает гидроскелет, поскольку у нематод отсутствуют окружные мышцы. От внутренней поверхности мышечных клеток идут выступы к нервным канатикам ; это уникальное явление в животном мире, при котором нервные клетки обычно протягивают волокна в мышцы, а не наоборот . [50]
Пищеварительная система
[ редактировать ]Полость рта выстлана кутикулой, которая часто укреплена структурами, такими как гребни, особенно у хищных видов, которые могут нести несколько зубов. Рот часто включает в себя острый стилет , которым животное может вонзить в добычу. У некоторых видов стилет полый и может использоваться для высасывания жидкости из растений или животных. [50]
Ротовая полость открывается в мускулистую сосущую глотку , также выстланную кутикулой. В этой области кишечника находятся пищеварительные железы, производящие ферменты , которые начинают расщеплять пищу. У видов, несущих стилеты, их можно даже вводить в жертву. [50]
нет Желудка , глотка соединяется непосредственно с бесмышечной кишкой , которая составляет основную часть кишки. Он производит дополнительные ферменты, а также поглощает питательные вещества через свою оболочку толщиной в одну клетку. Последняя часть кишечника выстлана кутикулой, образующей прямую кишку , которая выбрасывает отходы через отверстие чуть ниже и перед кончиком хвоста. Передвижение пищи по пищеварительной системе является результатом движений тела червя. Кишечник имеет клапаны или сфинктеры на обоих концах, которые помогают контролировать движение пищи по организму. [50]
Выделительная система
[ редактировать ]Азотистые отходы выводятся в виде аммиака через стенки тела и не связываются с какими-либо конкретными органами. Однако структуры выделения соли для поддержания осморегуляции обычно более сложны. [50]
У многих морских нематод одна или две одноклеточные сычужные железы выделяют соль через пору на нижней стороне животного, рядом с глоткой. У большинства других нематод эти специализированные клетки заменены органом, состоящим из двух параллельных протоков, соединенных одним поперечным протоком. Этот поперечный проток открывается в общий канал, идущий к выделительной поре. [50]
Нервная система
[ редактировать ]На переднем конце животного плотное круглое нервное кольцо , служащее мозгом . глотку окружает [50] От этого кольца вперед отходят шесть губно-сосочковых нервных тяжей и шесть нервных тяжей; Кзади отходят большой вентральный, меньший дорсальный и две пары сублатеральных тяжей. [51] Каждый нерв находится внутри тяжа соединительной ткани, лежащего под кутикулой и между мышечными клетками. Вентральный нерв является самым крупным и имеет двойную структуру перед экскреторной порой . Дорсальный нерв отвечает за двигательный контроль, латеральные нервы — сенсорные, а вентральный совмещает обе функции. [50]
Нервная система также является единственным местом в теле нематоды, содержащим реснички , неподвижные и обладающие сенсорной функцией. [52] [53]
Тела нематод покрыты многочисленными сенсорными щетинками и сосочками , которые вместе обеспечивают осязание. За чувствующими щетинками на голове лежат две небольшие ямки, или « амфиды ». Они хорошо снабжены нервными клетками и, вероятно, являются органами хеморецепции . Некоторые водные нематоды обладают чем-то вроде пигментированных глазных пятен, но неясно, действительно ли они имеют сенсорную природу. [50]
Воспроизведение
[ редактировать ]
Большинство видов нематод раздельнополы , с отдельными самцами и самками, хотя некоторые, такие как Caenorhabditis elegans , являются андродиодными , состоящими из гермафродитов и редких самцов. Оба пола обладают одной или двумя трубчатыми гонадами . У мужчин сперматозоиды вырабатываются на конце гонады и мигрируют по ее длине по мере созревания. Яичко открывается в относительно широкий семенной пузырек , а затем во время полового акта в железистый и мышечный семявыбрасывающий проток, связанный с семявыносящим протоком и клоакой . У самок яичники открываются в яйцевод (у гермафродитов яйцеклетки сначала попадают в сперматеку ), а затем в железистую матку . Обе матки открываются в общую вульву/влагалище, обычно расположенную в середине морфологически вентральной поверхности. [50]
Размножение обычно половое, хотя гермафродиты способны к самооплодотворению. Самцы обычно меньше самок или гермафродитов (часто намного меньше) и часто имеют характерно изогнутый или веерообразный хвост. Во время совокупления одна или несколько хитинизированных спикул выходят из клоаки и внедряются в половую пору самки. Амебоидные спермии заползают по спикуле в самку червя. Сперматозоиды нематод считаются единственной эукариотической клеткой, не содержащей глобулярного белка G-актина .
Яйца могут быть с эмбрионом или без эмбриона, когда их проходит самка, а это означает, что их оплодотворенные яйца могут еще не развиться. Известно, что некоторые виды являются яйцеживородящими . Яйца защищены внешней оболочкой, выделяемой маткой. У свободноживущих круглых червей из яиц вылупляются личинки , которые по сути идентичны взрослым особям, за исключением недоразвитой репродуктивной системы; у паразитических круглых червей жизненный цикл часто намного сложнее. [50] Строение яичной скорлупы сложное и включает несколько слоев; для этих слоев в 2023 году предложена подробная анатомо-терминологическая база. [55]
Нематоды в целом обладают широким спектром способов размножения. [56] Некоторые нематоды, такие как Heterorhabditis spp., подвергаются процессу, называемому эндотокиа матрицида : внутриутробные роды, вызывающие материнскую смерть. [57] Некоторые нематоды являются гермафродитами и сохраняют самооплодотворенные яйца внутри матки до тех пор, пока не вылупятся. Затем молодые нематоды поедают родительскую нематоду. Этот процесс значительно ускоряется в средах с низким запасом продовольствия. [57]
Модельные виды нематод C. elegans , C. briggsae и Pristionchus pacificus , среди других видов, демонстрируют андродиоэцию . [58] что в остальном очень редко встречается среди животных. Единственный род Meloidogyne (галловые нематоды) демонстрирует ряд репродуктивных способов, включая половое размножение , факультативную сексуальность (при которой большинство, но не все поколения размножаются бесполым путем), а также мейотический и митотический партеногенез .
Род Mesorhabditis демонстрирует необычную форму партеногенеза, при которой самцы, производящие сперму, совокупляются с самками, но сперматозоиды не сливаются с яйцеклеткой. Контакт со спермой необходим для того, чтобы яйцеклетка начала делиться, но поскольку слияния клеток не происходит, самец не передает генетический материал потомству, которое по сути является клонами самки. [50]
Старение
[ редактировать ]Нематода Caenorhabditis elegans часто используется в качестве модельного организма для изучения старения на молекулярном уровне. Например, у C. elegans старение отрицательно влияет на репарацию ДНК , а мутанты C. elegans обладают повышенной способностью к репарации ДНК. долгоживущие [59] Эти результаты предполагают генетически детерминированную корреляцию между способностью к репарации ДНК и продолжительностью жизни. [59] у самок C. elegans процессы зародышевой линии , которые контролируют репарацию ДНК и образование хромосомных кроссинговеров во время мейоза, с возрастом постепенно ухудшаются. Было показано, что [60]
Свободноживущие виды
[ редактировать ]Различные свободноживущие виды питаются такими разнообразными материалами, как бактерии , водоросли , грибы , мелкие животные, фекалии, мертвые организмы и живые ткани. Свободноживущие морские нематоды — важные и многочисленные представители мейобентоса . Они играют важную роль в процессе разложения, помогают перерабатывать питательные вещества в морской среде и чувствительны к изменениям в окружающей среде, вызванным загрязнением. Один известный круглый червь, C. elegans , живет в почве и нашел широкое применение в качестве модельного организма . Весь геном C. elegans секвенирован, судьба развития каждой клетки определена, а каждый нейрон картирован.
Паразитические виды
[ редактировать ]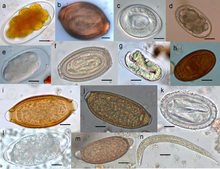
К нематодам, которые обычно паразитируют на людях, относятся аскариды ( Ascaris ), филярии , нематоды , острицы ( Enterobius ) и власоглавы ( Trichuris trichiura ). Вид Trichinellaspiralis , широко известный как «червь-трихинал», встречается у крыс, свиней, медведей и людей и является причиной заболевания трихинеллез . Baylisascaris обычно заражает диких животных, но может быть смертельным и для человека. Dirofilaria immitis известна тем, что вызывает сердечные черви, обитая в сердцах, артериях и легких собак и некоторых кошек. Haemonchus contortus является одним из наиболее распространенных возбудителей инфекции среди овец во всем мире, нанося овцам большой экономический ущерб. Напротив, энтомопатогенные нематоды паразитируют на насекомых и в большинстве случаев считаются полезными для человека, но некоторые из них нападают на полезных насекомых.
Одна из форм нематод полностью зависит от инжирных ос , которые являются единственным источником оплодотворения инжира . Они охотятся на ос, передвигаясь на них от спелого инжира, в котором родилась оса, к фиговому цветку ее смерти, где они убивают осу, а их потомство ожидает рождения следующего поколения ос, пока созревает инжир.

Паразитическая нематода тетрадонематид, открытая в 2005 году, Myrmeconema neotropicum , вызывает мимикрию плодов у тропического муравья Cephalotes atratus . У инфицированных муравьев появляются ярко-красные брюшные полости (живот), они становятся более вялыми и ходят, занимая заметно приподнятое положение. Эти изменения, вероятно, заставляют плодоядных птиц принимать зараженных муравьев за ягоды и поедать их. птиц, Яйца паразитов, попадающие в фекалии впоследствии собираются C. atratus, добывая пищу , и скармливаются их личинкам , таким образом завершая жизненный цикл M. neotropicum . [61]
Аналогичным образом, многочисленные разновидности нематод были обнаружены в брюшной полости примитивной общественной потной пчелы Lasioglossum zephyrus . Внутри самки нематода препятствует развитию яичников и делает пчелу менее активной, а значит, и менее эффективной в сборе пыльцы. [62]
Сельское хозяйство и садоводство
[ редактировать ]В зависимости от вида нематода может быть полезной или вредной для здоровья растений. С точки зрения сельского хозяйства и садоводства , существуют две категории нематод: хищные, убивающие садовых вредителей; и нематоды-вредители, которые поражают растения или действуют как переносчики, распространяющие растительные вирусы между сельскохозяйственными растениями. [63] Хищные нематоды включают Phasmarhabditis hermaphrodita , который является смертельным паразитом брюхоногих моллюсков, таких как слизни и улитки . [64] Некоторые представители рода Steinernema , такие как Steinernema carpocapsae и Steinernema riobrave, являются универсальными паразитами паутинных червей , совков , совков , поясковых червей , некоторых долгоносиков , древоточцев и кукурузной совки . [65] Эти организмы выращиваются в коммерческих целях как биологические средства борьбы с вредителями, которые можно использовать в качестве альтернативы пестицидам ; их использование считается очень безопасным. [66] Нематоды, паразитирующие на растениях, включают несколько групп, вызывающих серьезные потери урожая, ежегодно отбирая 10% посевов во всем мире. [67] Наиболее распространенными родами являются Aphelenchoides ( листовые нематоды ), Ditylenchus , Globodera (картофельные цистообразующие нематоды), Heterodera (соевые цистообразующие нематоды), Longidorus , Meloidogyne ( галловые нематоды ), Nacobbus , Pratylenchus (поражающиеся нематоды), Trichodorus и Xiphinema . кинжальные нематоды). Некоторые виды фитопаразитических нематод вызывают гистологические повреждения корней, включая образование видимых галлов (например, галловыми нематодами), которые являются полезными признаками для их диагностики в полевых условиях. Некоторые виды нематод передают вирусы растений посредством питания на корнях. Один из них — индекс Xiphinema , переносчик вируса веерного листа виноградной лозы , серьезного заболевания винограда, другой — Xiphinema diversicaudatum , переносчик вируса мозаики арабиса . Другие нематоды поражают кору и лесные деревья. Наиболее важным представителем этой группы является Bursaphelenchus xylophilus , сосновая лесная нематода, обитающая в Азии и Америке и недавно обнаруженная в Европе. Эта нематода передается с дерева на дерево жуками-пилорезами ( Монохам ). [68]
Теплицы используют полезных нематод для борьбы с грибковыми комарами . Нематоды проникают в личинки комаров через анус, рот и дыхальца (дыхательные поры), а затем выделяют бактерии , которые убивают личинки комаров; Обычно используемые виды нематод для борьбы с вредителями тепличных культур включают Steinernema Feltiae для грибных комаров и западного цветочного трипса , Steinernema carpocapsae , используемую для борьбы с прибрежными мухами, Steinernema kraussei для борьбы с черными виноградными долгоносиками и Heterorhabditis бактериофора для борьбы с личинками жуков. [69]
Севооборот растений с устойчивыми к нематодам видами или сортами является одним из средств борьбы с заражением паразитическими нематодами. Например, было показано, что посадка бархатцев Tagetes в качестве покровной культуры непосредственно перед посадкой восприимчивого к нематодам растения подавляет нематоды. [70] Другой вариант — лечение природными антагонистами, такими как гриб Gliocladiumroseum . Хитозан , естественный биоконтроль , вызывает защитные реакции растений, уничтожая паразитических нематод- цист на корнях сои , кукурузы , сахарной свеклы , картофеля и томатов , не нанося вреда полезным нематодам в почве. [71] Пропаривание почвы — эффективный метод уничтожения нематод перед посадкой урожая, но он без разбора уничтожает как вредную, так и полезную почвенную фауну.
Золотистая нематода Globodera rostochiensis представляет собой особенно вредоносную разновидность нематод-вредителей, которая стала причиной карантинов и неурожаев во всем мире. CSIRO обнаружила 13-14-кратное снижение плотности популяции нематод на участках, где китайской горчицы Brassica juncea . в почве присутствует зеленое навоз или семенная мука [72]
Эпидемиология
[ редактировать ]
Ряд кишечных нематод вызывают заболевания, поражающие человека, в том числе аскаридоз , трихоцефалез и анкилостомоз . Филяриозные нематоды вызывают филяриатоз . Кроме того, исследования показали, что паразитические нематоды заражают американских угрей, вызывая повреждение их плавательного пузыря. [73] молочные животные, такие как крупный рогатый скот и буйволы, [74] и все виды овец. [75]
Инфекции желудочно-кишечных нематод у людей распространены, им страдают примерно 50% населения мира. Развивающиеся страны страдают больше всего, отчасти из-за отсутствия доступа к медицинской помощи. [76]
Почвенные экосистемы
[ редактировать ]Около 90% нематод обитают в верхних 15 см (6 дюймах) почвы. Нематоды не разлагают органические вещества, а вместо этого являются паразитическими и свободноживущими организмами, питающимися живым материалом. Нематоды могут эффективно регулировать популяцию и сообщество бактерий. состав — они могут поедать до 5000 бактерий в минуту. Кроме того, нематоды могут играть важную роль в круговороте азота посредством минерализации азота. [47]
Одна группа плотоядных грибов — грибы-нематофаги — питаются почвенными нематодами. [77] Они устанавливают приманки для нематод в виде арканов или клейких структур. [78] [79] [80]
Живучесть
[ редактировать ]Черви-нематоды ( C. elegans ), ставшие частью продолжающегося исследовательского проекта, проводимого в 2003 году в рамках космического корабля "Колумбия" миссии STS-107 , пережили распад при входе в атмосферу . Считается, что это первая известная форма жизни, пережившая практически незащищенный атмосферный спуск на поверхность Земли. [81] [82] В исследовательском проекте, опубликованном в 2012 году, было обнаружено, что антарктические нематоды ( P. davidi ) способны противостоять внутриклеточному замораживанию в зависимости от того, насколько хорошо их кормят. [83] В 2023 году сообщалось, что особь Panagrolaimus kolymaensis возродилась после 46 000 лет пребывания в вечной мерзлоте Сибири. [84]
См. также
[ редактировать ]- Биологическая борьба с вредителями - борьба с вредителями с помощью других организмов.
- Capillaria - род круглых червей.
- Nematode.net
- Список тем по органическому садоводству и сельскому хозяйству — обзор и актуальное руководство по органическому садоводству и сельскому хозяйству
- Список паразитов человека
- Почвенная пищевая сеть
- Токсокароз – заболевание человека, вызываемое личинками собачьих или кошачьих круглых червей: Гельминтозная инфекция человека, вызываемая собачьими или кошачьими аскаридами, Toxocara canis или Toxocara cati.
- Мешок с червями - форма живорождения, наблюдаемая у нематод.
Ссылки
[ редактировать ]- ^ Пойнар, Джордж; Керп, Ганс; Хасс, Хаген (январь 2008 г.). «Palaeonema phyticum gen. n., sp. n. (Nematoda: Palaeonematidae fam. n.), девонская нематода, связанная с ранними наземными растениями» . Нематология . 10 (1): 9–14. дои : 10.1163/156854108783360159 .
- ^ Маас, Андреас; Валошек, Дитер; Хауг, Иоахим; Мюллер, Клаус (январь 2007 г.). «Возможная личинка круглого червя из кембрийского периода Орстен и его влияние на филогению Cycloneuralia» . Мемуары Ассоциации палеонтологов Австралии . 34 : 499–519.
- ^ Перейти обратно: а б «Нематоды» . Проверено 11 октября 2023 г.
- ^ Привет, Фрэнк. «Нематоды – хорошие, плохие и ужасные» . Новости и обзоры APS. apsnet.org . Американское фитопатологическое общество . Проверено 28 ноября 2020 г.
- ^ Бейкер, Эмили А.; Вуллард, Элисон (2019). «Насколько странный червь? Эволюция набора генов развития у Caenorhabditis elegans » . Журнал биологии развития . 7 (4): 19. дои : 10.3390/jdb7040019 . ПМК 6956190 . ПМИД 31569401 .
- ^ Ходда, М. (2011). Чжан, З.-Ц. (ред.). «Тип нематод (Кобб, 1932) ». Биоразнообразие животных: обзор классификации более высокого уровня и обзор таксономического богатства. Зоотакса . 3148 : 63–95. дои : 10.11646/zootaxa.3148.1.11 .
- ^ Чжан, З. (2013). Чжан, З.-Ц. (ред.). «Биоразнообразие животных: обновление классификации и разнообразия в 2013 году» . Биоразнообразие животных: обновленная классификация и разнообразие (Дополнения, 2013 г.). Зоотакса . 3703 (1): 5–11. дои : 10.11646/zootaxa.3703.1.3 .
- ^ Ламбсхед, П. Джон Д. (январь 1993 г.). «Последние достижения в исследованиях биоразнообразия морского бентоса» . Океанис . 19 (6): 5–24 . Проверено 5 ноября 2018 г. - через ResearchGate.
- ^ Андерсон, Рой К. (8 февраля 2000 г.). Нематоды-паразиты позвоночных: их развитие и передача . КАБИ. стр. 1–2. ISBN 9780851994215 .
Оценки от 500 000 до миллиона видов на самом деле не имеют под собой никаких оснований.
- ^ Ламбсхед П.Дж., Баучер Дж. (2003). «Глубоководное биоразнообразие морских нематод — гиперразнообразие или шумиха?» . Журнал биогеографии . 30 (4): 475–485. Бибкод : 2003JBiog..30..475L . дои : 10.1046/j.1365-2699.2003.00843.x . S2CID 86504164 .
- ^ Цин X, Берт В. (2019). «Семейство Tylenchidae (Nematoda): обзор и перспективы» . Разнообразие и эволюция организмов . 19 (3): 391–408. дои : 10.1007/s13127-019-00404-4 . S2CID 190873905 .
- ^ Флойд Р., Абебе Э., Паперт А., Блакстер М. (2002). «Молекулярные штрих-коды для идентификации почвенных нематод» . Молекулярная экология . 11 (4): 839–850. Бибкод : 2002MolEc..11..839F . дои : 10.1046/j.1365-294X.2002.01485.x . ПМИД 11972769 . S2CID 12955921 .
- ^ Дерик С., Шейбани Тезерджи Р., Риго А., Моенс Т. (2012). «Исследование экологии и эволюции загадочных видов морских нематод с помощью количественной ПЦР в реальном времени рибосомальной области ITS» . Ресурсы молекулярной экологии . 12 (4): 607–619. дои : 10.1111/j.1755-0998.2012.03128.x . hdl : 1854/LU-3127487 . ПМИД 22385909 . S2CID 4818657 .
- ^ Блакстер, Марк (2016). «Представляя Сизифа счастливым: штрих-кодирование ДНК и неназванное большинство» . Философские труды Лондонского королевского общества Б. 371 (1702): 20150329. doi : 10.1098/rstb.2015.0329 . ПМЦ 4971181 . ПМИД 27481781 .
- ^ Боргони Г., Гарсиа-Мояно А., Литтауэр Д., Берт В., Бестер А., ван Херден Э., Мёллер С., Эразмус М., Онстотт Т.К. (июнь 2011 г.). «Южноафриканская наземная подземная нематода» . Природа . 474 (7349): 79–82. Бибкод : 2011Natur.474...79B . дои : 10.1038/nature09974 . hdl : 1854/LU-1269676 . ПМИД 21637257 . S2CID 4399763 .
- ^ Лемоник, доктор медицины (8 июня 2011 г.). «Могут ли слова «черви из ада» означать, что в космосе есть жизнь?» . Время . ISSN 0040-781X . Архивировано из оригинала 10 июня 2011 года . Проверено 8 июня 2011 г.
- ^ Бхану, С.Н. (1 июня 2011 г.). «Нематода, обнаруженная у меня, — это первый подповерхностный многоклеточный организм» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 13 июня 2011 г.
- ^ «Золотой рудник» . Природа . 474 (7349): 6 июня 2011 г. doi : 10.1038/474006b . ПМИД 21637213 .
- ^ Дрейк Н. (1 июня 2011 г.). «Подземные черви из ада» . Новости природы. Новости природы . дои : 10.1038/news.2011.342 . Проверено 13 июня 2011 г.
- ^ Боргони Г., Гарсиа-Мояно А., Литтауэр Д., Берт В., Бестер А., ван Херден Э., Мёллер С., Эразмус М., Онстотт Т.К. (2 июня 2011 г.). «Южноафриканская наземная подземная нематода» . Природа . 474 (7349): 79–82. Бибкод : 2011Natur.474...79B . дои : 10.1038/nature09974 . hdl : 1854/LU-1269676 . ISSN 0028-0836 . ПМИД 21637257 . S2CID 4399763 .
- ^ Дановаро Р., Гамби С., Делл'Анно А., Коринальдези С., Фраскетти С., Ванреузель А. и др. (январь 2008 г.). «Экспоненциальное снижение функционирования глубоководных экосистем, связанное с утратой донного биоразнообразия» . Курс. Биол . 18 (1): 1–8. Бибкод : 2008CBio...18....1D . дои : 10.1016/j.cub.2007.11.056 . ПМИД 18164201 . S2CID 15272791 .
- «Исчезновение глубоководных видов может привести к коллапсу океанов, предполагает исследование» . ЭврекАлерт! (Пресс-релиз). 27 декабря 2007 г.
- ^ Перейти обратно: а б ван ден Хуген, Йохан; Гейзен, Стефан; Раут, Девин; Феррис, Ховард; Траунспургер, Уолтер; Уордл, Дэвид А.; и др. (24 июля 2019 г.). «Численность почвенных нематод и состав функциональных групп в глобальном масштабе» . Природа . 572 (7768): 194–198. Бибкод : 2019Природа.572..194В . дои : 10.1038/s41586-019-1418-6 . hdl : 20.500.11755/c8c7bc6a-585c-4a13-9e36-4851939c1b10 . ISSN 0028-0836 . ПМИД 31341281 . S2CID 198492891 . Архивировано из оригинала 2 марта 2020 года . Проверено 10 декабря 2019 г.
- ^ Платт Х.М. (1994). «предисловие». В Лоренцен С., Лоренцен С.А. (ред.). Филогенетическая систематика свободноживущих нематод . Лондон, Великобритания: Общество Рэев. ISBN 978-0-903874-22-9 .
- ^ Кэри, С. Крейг; Грин, Т.Г. Аллан; Стори, Брайан С.; Воробей, Эшли Д.; Хогг, Ян Д.; Катурджи, Марван; и др. (15 февраля 2019 г.). «Биотические взаимодействия — это неожиданный, но решающий способ контролировать сложность абиотической полярной экосистемы» . Коммуникационная биология . 2 (1): 62. дои : 10.1038/s42003-018-0274-5 . ISSN 2399-3642 . ПМК 6377621 . ПМИД 30793041 .
- ^ Адамс, Байрон Дж.; Уолл, Диана Х.; Стори, Брайан С.; Грин, Т.Г. Аллан; Барретт, Джон Э.; Кэри, С. Крейг; и др. (15 февраля 2019 г.). «Нематоды в полярной пустыне раскрывают относительную роль биотических взаимодействий в сосуществовании почвенных животных» . Коммуникационная биология . 2 (1): 63. дои : 10.1038/s42003-018-0260-y . ISSN 2399-3642 . ПМК 6377602 . ПМИД 30793042 .
- ^ Перейти обратно: а б Андерсон, Рой К. (8 февраля 2000 г.). Нематоды-паразиты позвоночных: их развитие и передача . КАБИ. п. 1. ISBN 978-0-85199-786-5 .
- ^ Кобб, Натан (1914). «Нематоды и их взаимоотношения» . Ежегодник . Министерство сельского хозяйства США . стр. 472, 457–490. Архивировано из оригинала 9 июня 2016 года . Проверено 25 сентября 2012 г. Цитата на стр. 472
- ^ Перейти обратно: а б с Читвуд Б.Г. (1957). «Английское слово «Нема» пересмотрено». Систематическая биология . 4 (45): 1619. doi : 10.2307/sysbio/6.4.184 .
- ^ Сиддики М.Р. (2000). Тиленхида: Паразиты растений и насекомых . Уоллингфорд, Оксон, Великобритания: Паб CABI. ISBN 978-0-85199-202-0 .
- ^ Шмидт-Раэса А (2014). «Gastrotricha, Cycloneuralia и Gnathifera: общая история и филогения». В Шмидт-Раэса А (ред.). Справочник по зоологии (основатель В. Кюкенталь) . Том. 1 — Nematomorpha, Priapulida, Kinorhyncha, Loricifera. Берлин, Бостон: де Грюйтер.
- ^ Кобб Н.А. (1919). «Отряды и классы немас» . Вклад. наук. Нематол . 8 : 213–216.
- ^ Уилсон, ЭО «Тип Немата» . nematode.unl.edu . Нематоды-паразиты растений и насекомых. Архивировано из оригинала 30 апреля 2018 года . Проверено 29 апреля 2018 г.
- ^ Нематода . Itis.gov (отчет). Отчет ИТИС . Проверено 12 июня 2012 г.
- ^ «Билатерия» . Древо жизни (tolweb.org) . Веб-проект «Древо жизни» . 2002 . Проверено 2 ноября 2008 г.
- ^ Читвуд Б.Г., Читвуд МБ (1933). «Персонажи протонематоды». Дж Паразитол . 20 :130.
- ^ Читвуд Б.Г. (1937). «Пересмотренная классификация нематод » . Статьи по гельминтологии, опубликованные к 30-летию со дня рождения... К.Я. Скрябина.. . Москва: Всесоюзная академия сельскохозяйственных наук имени Ленина. стр. 67–79.
- ^ Читвуд Б.Г. (1958). «Обозначение официальных названий высших таксонов беспозвоночных» . Номенклатура Bull Zool . 15 : 860–895. дои : 10.5962/bhl.part.19410 .
- ^ Коглан, А. (7 сентября 2005 г.). «Эволюция генома нематод» (PDF) . Червячная книга : 1–15. дои : 10.1895/wormbook.1.15.1 . ПМЦ 4781476 . ПМИД 18050393 . Архивировано из оригинала (PDF) 5 марта 2016 года . Проверено 13 января 2016 г.
- ^ Блакстер М.Л., Де Лей П., Гэри-младший, Лю Л.К., Шельдеман П., Виерстрате А., Ванфлетерен-младший, Макки Л.И., Доррис М., Фрисс Л.М., Вида Дж.Т., Томас В.К. (март 1998 г.). «Молекулярная эволюционная основа типа нематод». Природа . 392 (6671): 71–75. Бибкод : 1998Natur.392...71B . дои : 10.1038/32160 . ПМИД 9510248 . S2CID 4301939 .
- ^ «Нематода» . Веб-проект «Древо жизни» . 2002 . Проверено 2 ноября 2008 г.
- ^ Холтерман М., ван дер Вурфф А., ван ден Элсен С., ван Меген Х., Бонгерс Т., Головачев О., Баккер Дж., Хелдер Дж. (2006). «Анализ рДНК SSU в масштабе всего типа выявляет глубокие филогенетические связи между нематодами и ускоренную эволюцию в сторону коронных клад» . Мол Биол Эвол . 23 (9): 1792–1800. дои : 10.1093/molbev/msl044 . ПМИД 16790472 .
- ^ Перейти обратно: а б Хадка, Биджендра; Чаттерджи, Тонука; Гупта, Бхагвати П.; Гупта, Рэдхи С. (24 сентября 2019 г.). «Геномный анализ выявляет новые молекулярные признаки, специфичные для Caenorhabditis и других таксонов нематод, обеспечивая новые средства для генетических и биохимических исследований» . Гены . 10 (10): 739. doi : 10.3390/genes10100739 . ISSN 2073-4425 . ПМК 6826867 . ПМИД 31554175 .
- ^ «959 геномов нематод» . Геномы нематод. Nematodes.org . 11 ноября 2011 года. Архивировано из оригинала 5 августа 2015 года . Проверено 12 июня 2012 г.
- ^ Лю Г.Х., Шао Р., Ли JY, Чжоу Д.Х., Ли Х, Чжу XQ (2013). «Полные митохондриальные геномы трех паразитических нематод птиц: уникальный порядок генов и понимание филогении нематод» . БМК Геномика . 14 (1): 414. дои : 10.1186/1471-2164-14-414 . ПМЦ 3693896 . ПМИД 23800363 .
- ^ Ходда, М. (2022). «Тип Nematoda: классификация, каталог и указатель действительных родов с переписью действительных видов» . Зоотакса . 5114 (1): 1–289. дои : 10.11646/zootaxa.5114.1.1 . ПМИД 35391386 .
- ^ Маседо ду Карму, Густаво; ГАРСИА, Ренато Араужо; Виейра, Фабиано Матос; де Соуза Лима, Суэли; Исмаэль де Араужо-Жуниор, Эрминио; Пиньейру, Ральф Матурано (май 2023 г.). «Палеопаразитологическое исследование окаменелостей птиц из формации Тремембе (олигоцен бассейна Таубате), Сан-Паулу, Бразилия» . Журнал южноамериканских наук о Земле . 125 : 104319. Бибкод : 2023JSAES.12504319M . дои : 10.1016/j.jsames.2023.104319 . Получено 12 апреля 2024 г. - через Elsevier Science Direct.
- ^ Перейти обратно: а б Брэди, Найл К. и Вейл, Рэй Р. (2009). Элементы природы и свойства почв (3-е изд.). Прентис Холл. ISBN 9780135014332 .
- ^ Руперт Э.Э., Фокс Р.С., Барнс Р.Д. (2004). Зоология беспозвоночных: функционально-эволюционный подход (7-е изд.). Бельмонт, Калифорния: Брукс/Коул. ISBN 978-0-03-025982-1 .
- ^ Вейшер Б., Браун DJ (2000). Введение в нематоды: общая нематология . София, Болгария: Pensoft. стр. 75–76. ISBN 978-954-642-087-9 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Барнс Р.Г. (1980). Зоология беспозвоночных . Филадельфия: Колледж Сандерса. ISBN 978-0-03-056747-6 .
- ^ Хуан, Юн; Го, Юйцин (27 ноября 2021 г.). Свободноживущие морские нематоды Восточно-Китайского моря . Спрингер Природа. ISBN 978-981-16-3836-7 – через Google Книги.
- ^ «Чувствующие реснички Caenorhabditis elegans» . www.wormbook.org .
- ^ Кавли, Р.Г.; Кернан, MJ; Эберл, Д.Ф. (май 2010 г.). «Слух дрозофилы требует TilB, консервативного белка, связанного с подвижностью ресничек» . Генетика . 185 (1): 177–88. дои : 10.1534/genetics.110.114009 . ПМК 2870953 . ПМИД 20215474 .
- ^ Лалошевич, В.; Лалошевич, Д.; Капо, И.; Симин В.; Галфи, А.; Траверса, Д. (2013). «Высокий уровень заражения зоонозной инфекцией Eucoleus aerophilus у лисиц из Сербии» . Паразит . 20 :3. дои : 10.1051/parasite/2012003 . ПМЦ 3718516 . ПМИД 23340229 .
- ^ Бонд, Алан Томас; Хаффман, Дэвид Джордж (2023). «Яичная скорлупа нематод: новая анатомическая и терминологическая основа с критическим обзором соответствующей литературы и предлагаемыми рекомендациями по интерпретации и представлению изображений яичной скорлупы» . Паразит . 30 :6. дои : 10.1051/parasite/2023007 . ПМК 10016204 . ПМИД 36920277 .

- ^ Белл Дж. (1982). Шедевр природы: эволюция и генетика сексуальности . Беркли: Издательство Калифорнийского университета. ISBN 978-0-520-04583-5 .
- ^ Перейти обратно: а б Джонигк С.А., Элерс Р.У. (1999). « Endotokia matricida у гермафродитов Heterorhabditis spp. и влияние питания». Нематология . 1 (7–8): 717–726. дои : 10.1163/156854199508748 . ISSN 1388-5545 . S2CID 85279418 .
- ^ Хааг Э.С., Хелдер Дж., Муйджман П.Дж., Инь Д., Ху С. (2018). «Эволюция однородительского размножения нематод Rhabditina: филогенетические закономерности, причины развития и удивительные последствия» . Леонард, Дж.Л. (ред.). Переходы между половыми системами . Спрингер. стр. 99–122. дои : 10.1007/978-3-319-94139-4_4 . ISBN 978-3-319-94137-0 .
- ^ Перейти обратно: а б Хён М., Ли Дж., Ли К., Мэй А., Бор В.А., Ан Б. Долголетие и устойчивость к стрессу коррелируют со способностью восстановления ДНК у Caenorhabditis elegans. Нуклеиновые кислоты Рез. Март 2008 г.;36(4):1380-9. doi: 10.1093/nar/gkm1161. Epub 2008, 18 января. PMID 18203746; PMCID: PMC2275101
- ^ Райс М., Боуман Р., Смоликов С., Яновиц Дж.Л. Старение отрицательно влияет на восстановление ДНК и образование бивалентов в зародышевой линии C. elegans. Front Cell Dev Biol. 4 августа 2021 г.;9:695333. doi: 10.3389/fcell.2021.695333. ПМИД 34422819; PMCID: PMC8371636
- ^ Яновяк С.П., Каспари М., Дадли Р., Пойнар Г. (апрель 2008 г.). «Вызванная паразитами мимикрия фруктов у тропического муравья». Являюсь. Нат . 171 (4): 536–44. дои : 10.1086/528968 . ПМИД 18279076 . S2CID 23857167 .
- ^ Батра, Сюзанна В.Т. (1 октября 1965 г.). «Организмы, связанные с Lasioglossum zephyrum (Hymenoptera: Halictidae)». Журнал Канзасского энтомологического общества . 38 (4): 367–389. JSTOR 25083474 .
- ^ Перселл М., Джонсон М.В., Лебек Л.М., Хара А.Х. (1992). «Биологический контроль Helicoverpa zea (Lepidoptera: Noctuidae) с помощью Steinernema carpocapsae (Rhabditida: Steinernematidae) в кукурузе, используемой в качестве ловушки». Экологическая энтомология . 21 (6): 1441–1447. дои : 10.1093/ee/21.6.1441 .
- ^ Уилсон, MJ; Глен, DM; Джордж, СК (январь 1993 г.). «Рабдитидная нематода Phasmarhabditis hermaphrodita как потенциальный агент биологической борьбы со слизняками». Наука и технология биоконтроля . 3 (4): 503–511. Бибкод : 1993BioST...3..503W . дои : 10.1080/09583159309355306 .
- ^ Раджамани, Минатчи; Неги, Адити (2021). «Биопестициды для борьбы с вредителями». Устойчивая биоэкономика . стр. 239–266. дои : 10.1007/978-981-15-7321-7_11 . ISBN 978-981-15-7320-0 . S2CID 228845133 .
- ^ Элерс, Р.-У.; Хокканен, HMT (сентябрь 1996 г.). «Биоконтроль насекомых с неэндемичными энтомопатогенными нематодами (Steinernema и Heterorhabditis spp.): выводы и рекомендации совместного семинара ОЭСР и COST по вопросам научной и нормативной политики». Наука и технология биоконтроля . 6 (3): 295–302. Бибкод : 1996BioST...6..295E . дои : 10.1080/09583159631280 .
- ^ Смайли Р.В., Дабабат А.А., Икбал С., Джонс М.Г., Маафи З.Т., Пэн Д., Субботин С.А. , Вайенберге Л. (2017). «Злаковые цистообразующие нематоды: сложная и деструктивная группа видов гетеродер » . Болезни растений . 101 (10). Американское фитопатологическое общество : 1692–1720. doi : 10.1094/pdis-03-17-0355-fe . ISSN 0191-2917 . ПМИД 30676930 .
- ^ Гиббс, Дж. Н.; Уэббер, Дж. Ф. (2004), «ПАТОЛОГИЯ | Болезни деревьев, связанные с насекомыми» , Энциклопедия лесных наук , Elsevier, стр. 802–808, doi : 10.1016/b0-12-145160-7/00070-3 , ISBN 978-0-12-145160-8 , получено 21 марта 2023 г.
- ^ Клоостерман, Стивен (апрель 2022 г.). «Маленькие солдаты». Новости о продуктах Green House . Том. 32, нет. 4. С. 26–29.
- ^ Р. Крюгер; К.Е. Дувр; Р. МакСорли; КХ. Ван. «ENY-056/NG045: Бархатцы (Tagetes spp.) для борьбы с нематодами» . Институт пищевых и сельскохозяйственных наук . Проверено 20 ноября 2023 г.
- ^ Заявка США 2008072494 , Стоунер Р.Дж., Линден Дж.К., «Элиситор микроэлементов для лечения нематод в полевых культурах», опубликована 27 марта 2008 г.
- ^ Лутфар Р., Тони С. (22 марта 2005 г.). «Подавление галловой нематоды (Meloidogyne javanica) после внесения горчицы индийской сорта Nemfix в качестве сидерата и семенной муки на виноградниках» . Австралазийская патология растений . 34 (1): 77–83. Бибкод : 2005АуПП...34...77Р . дои : 10.1071/AP04081 . S2CID 24299033 . Проверено 14 июня 2010 г.
- ^ Варшафски, З.Т., Таки, Т.Д., Фогельбейн, В.К., Латур, Р.Дж., и Варго, АР (2019). Временные, пространственные и биологические вариации эпидемиологии нематод у американских угрей. Канадский журнал рыболовства и водных наук, 76 (10), 1808–1818 гг. https://doi.org/10.1139/cjfas-2018-0136
- ^ Джитендран и Бхат, Т. . (1999). Эпидемиология паразитозов молочных животных в северо-западном влажном гималайском регионе Индии с особым упором на желудочно-кишечные нематоды. Здоровье и производство тропических животных, 31(4), 205–214. https://doi.org/10.1023/A:1005263009921
- ^ Морган и ван Дейк, Дж. (2012). Климат и эпидемиология желудочно-кишечных нематодозов овец в Европе. Ветеринарная паразитология, 189 (1), 8–14. https://doi.org/10.1016/j.vetpar.2012.03.028
- ^ Степек, Джиллиан; Баттл, Дэвид Дж; Дуче, Ян Р; Бенке, Ежи М (октябрь 2006 г.). «Инфекции желудочно-кишечных нематод человека: нужны ли новые методы борьбы?» . Международный журнал экспериментальной патологии . 87 (5): 325–341. дои : 10.1111/j.1365-2613.2006.00495.x . ISSN 0959-9673 . ПМК 2517378 . ПМИД 16965561 .
- ^ Носовиц, Фан (8 февраля 2021 г.). «Как калифорнийские культуры боролись с вредителями, не используя пестициды» . Современный фермер . Проверено 15 февраля 2021 г.
- ^ Прамер С (1964). «Грибки, ловящие нематод». Наука . 144 (3617): 382–388. Бибкод : 1964Sci...144..382P . дои : 10.1126/science.144.3617.382 . ПМИД 14169325 .
- ^ Хаузер Дж. Т. (декабрь 1985 г.). «Грибки, ловящие нематод» (PDF) . Информационный бюллетень о плотоядных растениях . 14 (1): 8–11. дои : 10.55360/cpn141.jh945 .
- ^ Арен Д., Урсинг Б.М., Тунлид А. (1998). «Филогения грибов-ловушек нематод на основе последовательностей 18S рДНК». Письма FEMS по микробиологии . 158 (2): 179–184. дои : 10.1111/j.1574-6968.1998.tb12817.x . ПМИД 9465391 .
- ^ «Выжившие в Колумбии» . Журнал «Астробиология» . 1 января 2006 г. Архивировано из оригинала 4 марта 2016 г. Проверено 12 января 2016 г.
{{cite magazine}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Шевчик, Натаниэль Дж.; Манчинелли, Рокко Л.; Макламб, Уильям; Рид, Дэвид; Блумберг, Барух С.; Конли, Кэтрин А. (декабрь 2005 г.). « Caenorhabditis elegans выжил при разрушении атмосферы STS-107 космического корабля «Колумбия»». Астробиология . 5 (6): 690–705. Бибкод : 2005AsBio...5..690S . дои : 10.1089/ast.2005.5.690 . ПМИД 16379525 .
- ^ Раймонд, Мелиани Р.; Уортон, Дэвид А. (февраль 2013 г.). «Способность антарктической нематоды Panagrolaimus davidi пережить внутриклеточное замораживание зависит от статуса питания» . Журнал сравнительной физиологии Б. 183 (2): 181–188. дои : 10.1007/s00360-012-0697-0 . ISSN 0174-1578 . ПМИД 22836298 . S2CID 17294698 .
- ^ Шатилович, Анастасия; Гаде, Вамшидхар р. (27 июля 2023 г.). «Новый вид нематод из вечной мерзлоты Сибири разделяет адаптивные механизмы криптобиотического выживания с личинкой C. elegans dauer» . ПЛОС Генетика . 19 (7): e1010798. дои : 10.1371/journal.pgen.1010798 . ПМЦ 10374039 . ПМИД 37498820 .
Дальнейшее чтение
[ редактировать ]- Аткинсон, HJ (1973). «Дыхательная физиология морских нематод Enoplus brevis (Bastian) и E. communis (Bastian) : I. Влияние напряжения кислорода и размера тела» (PDF) . Журнал экспериментальной биологии . 59 (1): 255–266. дои : 10.1242/jeb.59.1.255 .
- «Черви пережили катастрофу в Колумбии» . news.bbc.co.uk. Новости Би-би-си . 1 мая 2003 года . Проверено 4 ноября 2008 г.
- Губанов, Н. М. (1951). «Гигантские нематоды из плаценты китообразных; Placentonema gigantissima nov. gen., nov. sp». Известия Академии наук СССР (на русском языке). 77 (6): 1123–1125.
- Кайя, Гарри К.; Постельные принадлежности, Робин А.; Ахерст, Раймонд Дж. (1993). «Обзор насекомо-паразитарных и энтомопатогенных нематод» . В Кайе, Гарри К.; Постельные принадлежности, Робин А.; Ахерст, Раймонд Дж. (ред.). Нематоды и биологическая борьба с насекомыми-вредителями . Издательство Csiro. стр. 8–23. ISBN 9780643105911 .
- «Гигантский почечный глист у норок и собак» . Ветеринарное руководство Merck (MVM) (онлайн-изд.). 2006. Архивировано из оригинала 3 марта 2016 года . Проверено 10 февраля 2007 г. - через merckvetmanual.com.
- Уайт Дж.Г., Саутгейт Э., Томсон Дж.Н., Бреннер С. (август 1976 г.). «Строение вентрального нервного канатика Caenorhabditis elegans » . Философские труды Лондонского королевского общества B: Биологические науки . 275 (938): 327–348. Бибкод : 1976RSPTB.275..327W . дои : 10.1098/rstb.1976.0086 . ПМИД 8806 .
- Ли, Дональд Л., изд. (2010). Биология нематод . Лондон, Великобритания: Тейлор и Фрэнсис. ISBN 978-0415272117 . Проверено 16 декабря 2014 г. - через Google Книги.
- де Лей П., Блакстер М. (2004). «Новая система для нематод: объединение морфологических признаков с молекулярными деревьями и перевод клад в ранги и таксоны». В Cook R, Hunt DJ (ред.). Монографии и перспективы нематологии . Том. 2. Лейден, Нидерланды: Э. Дж. Брилл. стр. 633–653.
- Рейн, Элизабет (18 декабря 2023 г.). «Что превращает грибка-падальщика в убийцу (нематод)?» . Арс Техника . Проверено 22 декабря 2023 г.
Внешние ссылки
[ редактировать ]- Нематологические исследования Университетского колледжа Харпера Адамса
- Нематоды/круглые черви человека
- Знакомство с нематодой
- Европейское общество нематологов
- Nematode.net: Репозиторий последовательностей паразитических нематод. Архивировано 14 февраля 2015 г. в Wayback Machine.
- http://webarchive.loc.gov/all/20020914155908/http://www.nematodes.org/
- База данных свободноживущих морских нематод NeMys World
- Виртуальная библиотека нематод
- Международная федерация нематологических обществ
- Общество нематологов
- Австралазийская ассоциация нематологов. Архивировано 26 февраля 2015 г. в Wayback Machine.
- Исследования нематод и долголетия
- Нематода на BBC
- Черви-нематоды в аквариуме
- Тип Nematoda - нематоды на UF / * Веб-сайт IFAS Featured Creatures