Актин
| Актин | |||
|---|---|---|---|
 Ленточная диаграмма G-актина. АДФ, актина связанный с активным центром (разноцветные палочки возле центра рисунка), а также комплекс дикатиона кальция (зеленая сфера) выделены. [ 1 ] | |||
| Идентификаторы | |||
| Символ | Актин | ||
| Пфам | PF00022 | ||
| ИнтерПро | ИПР004000 | ||
| PROSITE | PDOC00340 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 2btf / SCOPe / СУПФАМ | ||
| |||
Актин представляет собой семейство глобулярных , многофункциональных белков которые образуют микрофиламенты в цитоскелете и тонкие нити в мышечных фибриллах . Он обнаружен практически во всех эукариотических клетках , где он может присутствовать в концентрации более 100 мкМ ; его масса составляет примерно 42 кДа , диаметр от 4 до 7 нм.
Белок актин представляет собой мономерную субъединицу двух типов филаментов в клетках: микрофиламентов , одного из трех основных компонентов цитоскелета, и тонких нитей, части сократительного аппарата мышечных клеток. Он может присутствовать либо в виде свободного мономера , называемого G-актином (глобулярным), либо в виде части линейного полимерного микрофиламента, называемого F-актином (нитевидный), оба из которых необходимы для таких важных клеточных функций, как подвижность и сокращение клеток во время деление клеток .
Актин участвует во многих важных клеточных процессах, включая сокращение мышц , подвижность клеток , деление клеток и цитокинез , везикул и органелл движение , передачу сигналов клетками , а также создание и поддержание клеточных соединений и формы клеток. Многие из этих процессов опосредуются обширными и тесными взаимодействиями актина с клеточными мембранами . [ 2 ] У позвоночных три основные группы изоформ актина : альфа , бета и гамма идентифицированы . Альфа-актины, обнаруженные в мышечных тканях, являются основным компонентом сократительного аппарата. Бета- и гамма-актины сосуществуют в большинстве типов клеток как компоненты цитоскелета и как медиаторы внутренней подвижности клеток . Считается, что разнообразие структур, образуемых актином, позволяющее ему выполнять такой широкий спектр функций, регулируется посредством связывания тропомиозина вдоль филаментов. [ 3 ]
Способность клетки динамически формировать микрофиламенты обеспечивает основу, позволяющую ей быстро реконструировать себя в ответ на окружающую среду или на внутренние сигналы организма , например, для увеличения абсорбции клеточной мембраны или увеличения клеточной адгезии для формирования клеточной ткани . Другие ферменты или органеллы , такие как реснички, могут быть прикреплены к этому каркасу, чтобы контролировать деформацию внешней клеточной мембраны , что обеспечивает эндоцитоз и цитокинез . Он также может производить движение либо сам по себе, либо с помощью молекулярных двигателей . Таким образом, актин способствует таким процессам, как внутриклеточный транспорт везикул и органелл, а также мышечное сокращение и клеточная миграция . Поэтому он играет важную роль в эмбриогенезе , заживлении ран и инвазивности раковых клеток. Эволюционное происхождение актина можно проследить до прокариотических клеток , которые имеют эквивалентные белки. [ 4 ] Гомологи актина прокариот и архей полимеризуются в различные спиральные или линейные нити, состоящие из одной или нескольких нитей. Однако внутрицепочечные контакты и сайты связывания нуклеотидов сохраняются у прокариот и архей. [ 5 ] Наконец, актин играет важную роль в контроле экспрессии генов .
Большое количество заболеваний вызвано мутациями аллелей , генов болезней и регулирующих выработку актина или связанных с ним белков. Производство актина также играет ключевую роль в процессе заражения некоторыми патогенными микроорганизмами . Мутации в различных генах, которые регулируют выработку актина у человека, могут вызывать мышечные заболевания , изменения размера и функций сердца , а также глухоту . Строение цитоскелета связано также с патогенностью внутриклеточных бактерий и вирусов , особенно в процессах, связанных с уклонением от действия иммунной системы . [ 6 ]
Функция
[ редактировать ]Основная роль актина в клетке заключается в формировании линейных полимеров, называемых микрофиламентами , которые выполняют различные функции в структуре клетки, сетях транспортировки, миграции и репликации. [ 7 ] Многогранная роль актина зависит от некоторых свойств микрофиламентов: во-первых, образование актиновых нитей обратимо, и их функция часто включает быструю полимеризацию и деполимеризацию. Во-вторых, микрофиламенты поляризованы, то есть два конца нити отличны друг от друга. В-третьих, актиновые нити могут связываться со многими другими белками, которые вместе помогают модифицировать и организовывать микрофиламенты для выполнения их разнообразных функций. [ 7 ]
В большинстве клеток актиновые нити образуют более крупные сети, которые необходимы для многих ключевых функций: [ 8 ]
- Сети актина оказывают механическую поддержку клеткам и обеспечивают маршруты транспортировки через цитоплазму, помогая передаче сигнала.
- Быстрая сборка и разборка актиновой сети позволяет клеткам мигрировать ( миграция клеток ).
Актин чрезвычайно распространен в большинстве клеток и составляет 1–5% от общей белковой массы большинства клеток и 10% мышечных клеток. [ 7 ]
Белок актин содержится как в цитоплазме , так и в ядре клетки . [ 9 ] Его расположение регулируется путями передачи сигнала клеточной мембраны , которые интегрируют стимулы, которые получает клетка, стимулируя в ответ реструктуризацию актиновых сетей. [ 10 ]
Цитоскелет
[ редактировать ]
Существует ряд различных типов актина со слегка различающимися структурами и функциями. α-актин обнаруживается исключительно в мышечных волокнах , тогда как β- и γ-актин обнаруживаются в других клетках. Поскольку последние типы имеют высокую текучесть кадров, большинство из них находятся за пределами постоянных структур. Микрофиламенты, обнаруженные в клетках, отличных от мышечных, присутствуют в трех формах: [ 11 ]
- Сети микрофиламентов . Клетки животных обычно имеют клеточную кору под клеточной мембраной , содержащую большое количество микрофиламентов, что исключает присутствие органелл . Эта сеть связана с многочисленными рецепторами , которые передают сигналы наружу клетки.
- Периодические актиновые кольца обнаружена периодическая структура, состоящая из равномерно расположенных актиновых колец . В аксонах . [ 12 ] В этой структуре актиновые кольца вместе с тетрамерами спектрина , связывающими соседние актиновые кольца, образуют сплоченный цитоскелет , поддерживающий мембрану аксона. Периодичность структуры может также регулировать каналы ионов натрия в аксонах.
Дрожжи
[ редактировать ]Цитоскелет актина играет ключевую роль в процессах эндоцитоза , цитокинеза , определения полярности клеток и морфогенеза у дрожжей . Помимо актина, в этих процессах участвуют 20 или 30 ассоциированных белков, все из которых обладают высокой степенью эволюционной консервативности, а также множество сигнальных молекул. Вместе эти элементы обеспечивают пространственно и временно модулированную сборку, которая определяет реакцию клетки как на внутренние, так и на внешние стимулы. [ 13 ]
Дрожжи содержат три основных элемента, связанных с актином: участки, кабели и кольца. Несмотря на то, что эти структуры существуют недолго, они находятся в состоянии динамического равновесия из-за непрерывной полимеризации и деполимеризации. Они обладают рядом дополнительных белков, включая ADF/кофилин, который имеет молекулярную массу 16 кДа и кодируется одним геном, называемым COF1 ; Aip1, кофактор кофилина, способствующий разборке микрофиламентов; Srv2/CAP, регулятор процесса, связанный с аденилатциклазы белками ; профилин с молекулярной массой приблизительно 14 кДа, родственный/ассоциированный с мономерами актина; и твинфилин, белок массой 40 кДа, участвующий в организации пятен. [ 13 ]
Растения
[ редактировать ]растений Исследования генома выявили существование изовариантов белков в семействе генов актина. Внутри Arabidopsis thaliana , модельного организма , имеется десять типов актина, шесть профилинов и десятки миозинов. Такое разнообразие объясняется эволюционной необходимостью наличия вариантов, незначительно различающихся по своему временному и пространственному выражению. [ 4 ] Большинство этих белков экспрессировались совместно в анализируемых тканях . Актиновые сети распределены по цитоплазме клеток, культивируемых in vitro . Вокруг ядра сосредоточена сеть, которая связана спицами с клеточной корой, эта сеть очень динамична, с непрерывной полимеризацией и деполимеризацией. [ 14 ]

Несмотря на то, что большинство растительных клеток имеют клеточную стенку , определяющую их морфологию, их микрофиламенты могут генерировать достаточную силу для достижения ряда клеточных активностей, таких как цитоплазматические токи, генерируемые микрофиламентами и миозином. Актин также участвует в движении органелл и клеточном морфогенезе, который включает деление клеток, а также удлинение и дифференцировку клетки. [ 16 ]
Наиболее известные белки, связанные с актиновым цитоскелетом растений, включают: [ 16 ] виллин , который принадлежит к тому же семейству, что и гельзолин /северин, и способен разрезать микрофиламенты и связывать мономеры актина в присутствии катионов кальция; фимбрин , способный распознавать и объединять мономеры актина и участвующий в формировании сетей (путем иного процесса регуляции, чем у животных и дрожжей); [ 17 ] формины , которые способны действовать как зародышеобразователь полимеризации F-актина; миозин , типичный молекулярный двигатель, специфичный для эукариот и который у Arabidopsis thaliana кодируется 17 генами двух различных классов; CHUP1, который может связывать актин и участвует в пространственном распределении хлоропластов в клетке; KAM1/MUR3, которые определяют морфологию аппарата Гольджи , а также состав ксилоглюканов клеточной стенки; NtWLIM1, способствующий возникновению актиновых клеточных структур; и ERD10, который участвует в ассоциации органелл внутри мембран и микрофиламентов и, по-видимому, играет роль в реакции организма на стресс .
Ядерный актин
[ редактировать ]Ядерный актин был впервые замечен и описан в 1977 году Кларком и Мерриамом. [ 18 ] Авторы описывают белок, присутствующий в ядерной фракции, полученной из ооцитов Xenopus laevis , который проявляет те же свойства, что и актин скелетных мышц. С тех пор появилось множество научных сообщений о строении и функциях актина в ядре (обзор см.: Hofmann 2009. [ 19 ] ) Контролируемый уровень актина в ядре, его взаимодействие с актин-связывающими белками (АБП) и наличие различных изоформ позволяет актину играть важную роль во многих важных ядерных процессах. [ 20 ]
Транспорт через ядерную мембрану
[ редактировать ]Последовательность актина не содержит сигнала ядерной локализации. Небольшой размер актина (около 43 кДа) позволяет ему проникать в ядро путем пассивной диффузии. [ 21 ] Импорту актина в ядро (вероятно, в комплексе с кофилином) способствует импортный белок импортин 9. [ 22 ]
Низкие уровни актина в ядре, по-видимому, важны, поскольку актин имеет в своей последовательности два сигнала ядерного экспорта (NES). Микроинъецированный актин быстро удаляется из ядра в цитоплазму. Актин экспортируется по крайней мере двумя способами: через экспортин 1 и экспортин 6 . [ 23 ] [ 24 ] Специфические модификации, такие как SUMOylation, позволяют сохранять ядерный актин. Мутация, предотвращающая SUMOylation, вызывает быстрый экспорт бета-актина из ядра. [ 25 ]
Организация
[ редактировать ]Ядерный актин существует главным образом в виде мономера, но может также образовывать динамические олигомеры и короткие полимеры. [ 26 ] [ 27 ] [ 28 ] Организация ядерного актина варьируется в разных типах клеток. Например, в ооцитах Xenopus (с более высоким уровнем ядерного актина по сравнению с соматическими клетками) актин образует нити, которые стабилизируют архитектуру ядра. Эти нити можно наблюдать под микроскопом благодаря окрашиванию фаллоидином, конъюгированным с флуорофором. [ 18 ] [ 21 ]
Однако в ядрах соматических клеток актиновые нити невозможно наблюдать с помощью этого метода. [ 29 ] Анализ ингибирования ДНКазы I, единственный тест, который позволяет количественно оценить полимеризованный актин непосредственно в биологических образцах, показал, что эндогенный ядерный актин действительно существует главным образом в мономерной форме. [ 28 ]
Точно контролируемый уровень актина в ядре клетки, более низкий, чем в цитоплазме, предотвращает образование филаментов. Полимеризация также снижается из-за ограниченного доступа к мономерам актина, которые связаны в комплексы с АБФ, главным образом с кофилином. [ 30 ]
Изоформы актина
[ редактировать ]В ядре клетки присутствуют различные изоформы актина. Уровень изоформ актина может меняться в ответ на стимуляцию роста клеток или остановку пролиферации и транскрипционной активности. [ 31 ] Исследования ядерного актина сосредоточены на бета-изоформе. [ 32 ] [ 33 ] [ 34 ] [ 35 ] Однако использование антител, направленных против различных изоформ актина, позволяет идентифицировать не только цитоплазматический бета-актин в ядре клетки, но также альфа- и гамма-актин в определенных типах клеток. [ 28 ] [ 36 ] [ 37 ] Наличие различных изоформ актина может существенно влиять на его функцию в ядерных процессах, поскольку уровень отдельных изоформ можно контролировать независимо. [ 28 ]
Функции
[ редактировать ]Функции актина в ядре связаны с его способностью полимеризоваться и взаимодействовать с различными АБФ и структурными элементами ядра. Ядерный актин участвует в:
- Архитектура ядра . Взаимодействие актина с альфа II-спектрином и другими белками важно для поддержания правильной формы ядра. [ 38 ] [ 39 ]
- Транскрипция – Актин участвует в реорганизации хроматина, [ 9 ] [ 32 ] [ 40 ] [ 41 ] инициация транскрипции и взаимодействие с транскрипционным комплексом. [ 42 ] Актин принимает участие в регуляции структуры хроматина, [ 43 ] [ 44 ] [ 45 ] взаимодействуя с РНК-полимеразой I, [ 35 ] II [ 33 ] и III. [ 34 ] В транскрипции Pol I актин и миозин ( MYO1C , который связывает ДНК) действуют как молекулярный мотор . Для транскрипции Pol II β-актин необходим для образования преинициаторного комплекса. Pol III содержит β-актин в качестве субъединицы. Актин также может быть компонентом комплексов ремоделирования хроматина, а также частиц пре-мРНП (то есть предшественников информационной РНК, связанной с белками), и участвует в ядерном экспорте РНК и белков. [ 46 ]
- Регуляция активности генов . Актин связывается с регуляторными областями различных типов генов. [ 47 ] [ 48 ] [ 49 ] [ 50 ] Способность актина регулировать активность генов используется в методе молекулярного репрограммирования, который позволяет дифференцированным клеткам вернуться в эмбриональное состояние. [ 49 ] [ 51 ]
- Транслокация активированного фрагмента хромосомы из подмембранной области в эухроматин, где начинается транскрипция. Это движение требует взаимодействия актина и миозина. [ 52 ] [ 53 ]
- Интеграция различных клеточных компартментов . Актин представляет собой молекулу, которая объединяет цитоплазматические и ядерные пути передачи сигналов. [ 54 ] Примером может служить активация транскрипции в ответ на стимуляцию клеток сывороткой in vitro . [ 55 ] [ 56 ] [ 57 ]
- Иммунный ответ . Ядерный актин полимеризуется при стимуляции рецептора Т-клеток и необходим для экспрессии цитокинов и продукции антител in vivo . [ 58 ]
- Восстановление ДНК . Ядерный актин опосредует восстановление двухцепочечных разрывов ДНК . [ 59 ] В клеточном ядре нитевидный полимер актина (F-актин) действует как на пути репарации ДНК негомологичного соединения концов, так и на пути гомологичной рекомбинационной репарации . [ 59 ]
Благодаря своей способности претерпевать конформационные изменения и взаимодействовать со многими белками актин выступает регулятором образования и активности белковых комплексов, таких как транскрипционный комплекс. [ 42 ]
Движение клеток
[ редактировать ]Актин также участвует в движении клеток. Сеть из актиновых нитей отмечает передний край движущейся клетки, и полимеризация новых актиновых нитей выталкивает клеточную мембрану вперед в виде выступов, называемых ламеллиподиями . [ 60 ] [ 61 ] [ 62 ] Эти выступы мембраны затем прикрепляются к субстрату, образуя структуры, известные как фокальные спайки , которые соединяются с актиновой сетью. [ 62 ] После прикрепления задняя часть тела клетки сжимается, выдавливая ее содержимое вперед за точку адгезии. [ 62 ] Как только точка адгезии перемещается к задней части клетки, ячейка разбирает ее, позволяя задней части клетки двигаться вперед. [ 62 ]

Движение актина/миозина
[ редактировать ]В дополнение к физической силе, создаваемой полимеризацией актина, микрофиламенты облегчают движение различных внутриклеточных компонентов, служа дорогой, по которой движется семейство моторных белков, называемых миозинами . [ 63 ]
Сокращение мышц
[ редактировать ]Актин играет особенно важную роль в мышечных клетках, которые состоят в основном из повторяющихся пучков актина и миозина II . [ 64 ] Каждая повторяющаяся единица, называемая саркомером , состоит из двух наборов противоположно ориентированных нитей F-актина («тонких нитей»), переплетенных с пучками миозина («толстых нитей»). Два набора нитей актина ориентированы своими (+) концами, встроенными в оба конца саркомера в разграничивающие структуры, называемые Z-дисками . [ 64 ] Миозиновые фибриллы расположены посередине между наборами актиновых нитей, их нити обращены в обоих направлениях. Когда мышца сокращается, нити миозина движутся вдоль актиновых нитей к (+) концу, сближая концы саркомера и укорачивая его примерно на 70% его длины. [ 64 ] Чтобы двигаться по актиновой нити, миозин должен гидролизовать АТФ; таким образом, АТФ служит источником энергии для сокращения мышц. [ 64 ]
Во время покоя белки тропомиозин и тропонин связываются с актиновыми нитями, предотвращая прикрепление миозина. [ 64 ] Когда сигнал активации (то есть потенциал действия ) поступает в мышечное волокно, он запускает высвобождение Са. 2+ из саркоплазматического ретикулума в цитозоль. Возникающий в результате всплеск цитозольного кальция быстро высвобождает тропомиозин и тропонин из актиновой нити, позволяя миозину связываться и начинать сокращение мышц. [ 65 ]
Деление клеток
[ редактировать ]На заключительных стадиях клеточного деления многие клетки образуют актиновое кольцо в средней точке клетки. Это кольцо, удачно названное « сократительным кольцом », использует тот же механизм, что и мышечные волокна: миозин II тянет актиновое кольцо, заставляя его сокращаться. [ 66 ] Это сокращение расщепляет родительскую клетку на две части, завершая цитокинез . [ 66 ] Сократительное кольцо состоит из актина, миозина, аниллина и α-актинина . [ 67 ] У делящихся дрожжей Schizosaccharomyces pombe актин активно образуется в констрикционном кольце при участии Arp3 , формина Cdc12, профилина и WASp , а также предварительно сформированных микрофиламентов. После того, как кольцо построено, структура поддерживается посредством постоянной сборки и разборки, которая при помощи комплекса Arp2/3 и форминов является ключом к одному из центральных процессов цитокинеза. [ 68 ]
Внутриклеточный трафик
[ редактировать ]Пары актин-миозин также могут участвовать в транспортировке различных мембранных везикул и органелл внутри клетки. Миозин V активируется путем связывания с различными рецепторами груза на органеллах, а затем движется по актиновой нити к (+) концу, увлекая за собой свой груз. [ 69 ]
Эти нетрадиционные миозины используют гидролиз АТФ для направленной транспортировки грузов, таких как везикулы и органеллы, гораздо быстрее, чем диффузия. Миозин V движется к зазубренному концу актиновых нитей, а миозин VI — к заостренному концу. Большинство актиновых нитей расположены зазубренным концом к клеточной мембране, а заостренным концом — внутрь клетки. Такое расположение позволяет миозину V быть эффективным двигателем экспорта грузов, а миозину VI — эффективным двигателем импорта.
Другие биологические процессы
[ редактировать ]Традиционное представление о функции актина связывает его с поддержанием цитоскелета и, следовательно, с организацией и движением органелл, а также с определением формы клетки. [ 11 ] Однако актин играет более широкую роль в физиологии эукариотических клеток, помимо аналогичных функций у прокариот .
- Апоптоз . Во время запрограммированной гибели клеток семейство протеаз ICE/ced-3 (одна из протеаз, конвертирующих интерлейкин-1β) расщепляет актин на два фрагмента in vivo ; один из фрагментов имеет массу 15 кДа, а другой - 31 кДа. Это представляет собой один из механизмов разрушения жизнеспособности клеток, составляющих основу апоптоза. [ 70 ] Также было показано, что протеаза кальпаин участвует в этом типе разрушения клеток; [ 71 ] точно так же, как было показано, использование ингибиторов кальпаина уменьшает протеолиз актина и деградацию ДНК (еще один из характерных элементов апоптоза). [ 72 ] С другой стороны, вызванный стрессом запуск апоптоза вызывает реорганизацию актинового цитоскелета (что также включает его полимеризацию), приводя к образованию структур, называемых стрессовыми волокнами ; это активируется путем MAP-киназы . [ 73 ]

- Клеточная адгезия и развитие . Адгезия между клетками является характеристикой многоклеточных организмов , которая обеспечивает специализацию тканей и, следовательно, увеличивает сложность клеток. В адгезии клеточного эпителия участвует актиновый цитоскелет в каждой из соединяющихся клеток, а также кадгерины, действующие как внеклеточные элементы, связь между которыми опосредована катенинами . [ 74 ] Вмешательство в динамику актина имеет последствия для развития организма, фактически актин является настолько важным элементом, что системы избыточных генов доступны . Например, если α-актинина или фактора гелеобразования был удален ген у Dictyostelium , у особей Dictyostelium не наблюдается аномального фенотипа , возможно, из-за того, что каждый из белков может выполнять функцию другого. развитие двойных мутаций , в которых отсутствуют оба типа генов. Однако это влияет на [ 75 ]
- экспрессии генов Модуляция . Состояние полимеризации актина влияет на характер экспрессии генов . В 1997 году было обнаружено, что деполимеризация, опосредованная цитокалазином D, в шванновских клетках вызывает специфический паттерн экспрессии генов, участвующих в миелинизации этого типа нервных клеток . [ 76 ] Было показано, что F-актин модифицирует транскриптом на некоторых стадиях жизни одноклеточных организмов, таких как гриб Candida albicans . [ 77 ] Кроме того, белки, подобные актину, играют регуляторную роль во время сперматогенеза у мышей. [ 78 ] Считается, что у дрожжей актиноподобные белки играют роль в регуляции экспрессии генов . [ 79 ] Фактически актин способен действовать как инициатор транскрипции, когда он реагирует с типом ядерного миозина, который взаимодействует с РНК-полимеразами и другими ферментами, участвующими в процессе транскрипции. [ 9 ]
- Динамика стереоцилий . На поверхности некоторых клеток образуются тонкие нитевидные выросты, выполняющие механосенсорную функцию. Например, этот тип органелл присутствует в кортиевом органе , который расположен в ухе . Основная особенность этих структур заключается в том, что их длину можно изменять. [ 80 ] Молекулярная архитектура стереоцилий включает паракристаллическое актиновое ядро, находящееся в динамическом равновесии с мономерами, присутствующими в соседнем цитозоле. Миозины типов VI и VIIa присутствуют по всему этому ядру, тогда как миозин XVa присутствует в его концах в количествах, пропорциональных длине стереоцилий. [ 81 ]
- Внутренняя хиральность . Сети актомиозина участвуют в создании внутренней хиральности в отдельных клетках. [ 82 ] Клетки, выращенные на хиральных поверхностях, могут демонстрировать направленное смещение влево/вправо, которое зависит от актомиозина. [ 83 ] [ 84 ]
Структура
[ редактировать ]
Мономерный актин, или G-актин, имеет глобулярную структуру, состоящую из двух долей, разделенных глубокой щелью. [ 85 ] Нижняя часть щели представляет собой «складку АТФазы», структуру, консервативную среди АТФ и ГТФ-связывающих белков, которая связывается с ионом магния и молекулой АТФ. [ 85 ] Связывание АТФ или АДФ необходимо для стабилизации каждого мономера актина; без связывания одной из этих молекул актин быстро денатурируется . [ 85 ]
Рентгеновская кристаллографическая модель актина, полученная Кабшем из поперечно-полосатой мышечной ткани кроликов , наиболее часто используется в структурных исследованиях, поскольку она была очищена первой . G-актин, кристаллизованный Кабшем, имеет примерно 67 x 40 x 37 Å размеры , молекулярную массу 41 785 Да и расчетную изоэлектрическую точку 4,8. Его чистый заряд при pH = 7 равен -7. [ 86 ] [ 87 ]
- Первичная структура
Эльзинга и его коллеги впервые определили полную пептидную последовательность этого типа актина в 1973 году, а более поздняя работа того же автора добавила к модели дополнительные детали. Он содержит 374 аминокислотных остатка. Его N-конец очень кислый и начинается с ацетилированного аспартата в аминогруппе. В то время как его С-конец является щелочным и образован фенилаланином, которому предшествует цистеин , который имеет определенную функциональную важность. Обе крайности находятся в непосредственной близости внутри I-подобласти. Аномальный Н т -метилгистидин расположен в положении 73. [ 87 ]
- Третичная структура — домены
Третичная структура образована двумя доменами, известными как большой и малый, которые разделены щелью, сосредоточенной вокруг места связи с АТФ - АДФ + P i . Ниже есть более глубокая выемка, называемая «паз». В родном штате , несмотря на названия, оба имеют сопоставимую глубину. [ 86 ]
Обычное соглашение в топологических исследованиях означает, что белок показан с самым большим доменом слева и самым маленьким доменом справа. В этом положении меньший домен, в свою очередь, делится на два: субдомен I (нижнее положение, остатки 1–32, 70–144 и 338–374) и субдомен II (верхнее положение, остатки 33–69). Более крупный домен также разделен на две части: субдомен III (нижний, остатки 145–180 и 270–337) и субдомен IV (верхний, остатки 181–269). Открытые области субдоменов I и III называются «колючими» концами, а открытые области доменов II и IV называются «заостренными» концами. Эта номенклатура относится к тому факту, что из-за небольшой массы субдомена II актин является полярным; важность этого будет обсуждаться ниже при обсуждении динамики сборки. Некоторые авторы называют поддомены Ia, Ib, IIa и IIb соответственно. [ 88 ]
- Другие важные структуры
Наиболее заметной супервторичной структурой является пятицепочечный бета-лист , состоящий из β-меандра и β-α-β, направленного по часовой стрелке. Он присутствует в обоих доменах, что позволяет предположить, что белок возник в результате дупликации гена. [ 89 ]
- Сайт связывания аденозиновых нуклеотидов расположен между двумя бета-шпилькообразными структурами, принадлежащими I и III доменам. Участвующие остатки — это Asp11-Lys18 и Asp154-His161 соответственно.
- Сайт связывания двухвалентного катиона расположен чуть ниже сайта связывания аденозинового нуклеотида. In vivo чаще всего образуется Mg 2+ или Ca 2+ в то время как in vitro он образуется хелатной структурой, состоящей из Lys18 и двух атомов кислорода нуклеотида из α- и β- фосфатов . Этот кальций координируется с шестью молекулами воды, которые удерживаются аминокислотами Asp11 , Asp154 и Gln137 . Они образуют комплекс с нуклеотидом, ограничивающим движения так называемой «шарнирной» области, расположенной между остатками 137 и 144. Это сохраняет нативную форму белка до тех пор, пока его удаление не приведет к денатурации мономера актина. Эта область также важна, поскольку она определяет, находится ли щель белка в «открытой» или «закрытой» конформации. [ 1 ] [ 88 ]
- Весьма вероятно, что существуют как минимум три других центра с меньшим сродством (промежуточным) и еще другие с низким сродством к двухвалентным катионам. Было высказано предположение, что эти центры могут играть роль в полимеризации актина, действуя на стадии активации. [ 88 ]
- В субдомене 2 есть структура, которая называется «D-петля», поскольку она связывается с ДНКазой I и расположена между остатками His40 и Gly48 . В большинстве кристаллов он выглядит как неупорядоченный элемент, но в комплексе с ДНКазой I он выглядит как β-лист. Было высказано предположение, что ключевым событием полимеризации, вероятно, является распространение конформационных изменений от центр связи нуклеотида с этим доменом, который меняется от петли к спирали. [ 1 ] Однако эта гипотеза была опровергнута другими исследованиями. [ 90 ]
F-актин
[ редактировать ]
В различных условиях молекулы G-актина полимеризуются в более длинные нити, называемые «нитевидными» или «F-актинами». Эти нити F-актина обычно состоят из двух спиральных нитей актина, намотанных друг на друга, образующих спираль шириной от 7 до 9 нанометров , которая повторяется каждые 72 нанометра (или каждые 14 субъединиц G-актина). [ 92 ] В нитях F-актина все молекулы G-актина ориентированы в одном направлении. Два конца нити F-актина отличны друг от друга. На одном конце, обозначенном как (-), АТФ-связывающая щель терминальной молекулы актина обращена наружу. На противоположном конце, обозначенном (+), АТФ-связывающая щель погружена в нить, контактируя с соседней молекулой актина. [ 92 ] По мере роста нитей F-актина новые молекулы имеют тенденцию присоединяться к (+) концу существующей цепи F-актина. И наоборот, нити имеют тенденцию сжиматься за счет отделения мономеров актина с конца (-) цепи. [ 92 ]
Некоторые белки, такие как кофилин , по-видимому, увеличивают угол поворота, но это опять-таки можно интерпретировать как установление различных структурных состояний. Они могут иметь важное значение в процессе полимеризации. [ 93 ]
Меньше согласия относительно измерений радиуса витка и толщины нити: в то время как первые модели приписывали длину 25 Å, текущие данные рентгеновской дифракции, подтвержденные криоэлектронной микроскопией, предполагают длину 23,7 Å. Эти исследования показали точные точки контакта между мономерами. Некоторые из них образованы звеньями одной и той же цепи между «зазубренным» концом одного мономера и «заостренным» концом следующего. В то время как мономеры в соседних цепях вступают в латеральный контакт через выступы субдомена IV, причем наиболее важными выступами являются те, которые образованы С-концом и гидрофобной связью, образованной тремя телами, включающими остатки 39–42, 201–203 и 286. Это модель предполагает, что филамент образуется из мономеров в виде «листа», в котором субдомены поворачиваются вокруг себя; эта форма также обнаружена у гомолога бактериального актина MreB . [ 94 ]
Термины «заостренный» и «колючий», относящиеся к двум концам микронитей, происходят от их внешнего вида под просвечивающей электронной микроскопией , когда образцы исследуются с использованием метода подготовки, называемого «декорированием». Этот метод заключается в добавлении фрагментов миозина S1 к ткани, зафиксированной дубильной кислотой . Этот миозин образует полярные связи с мономерами актина, образуя конфигурацию, которая выглядит как стрелы с перьевым оперением вдоль стержня, где стержень представляет собой актин, а оперение — миозин. Следуя этой логике, конец микрофиламента, не имеющий выступающего миозина, называется острием стрелки (конец −), а другой конец — зазубренным концом (+ конец). [ 95 ] Фрагмент S1 состоит из головного и шейного доменов миозина II . В физиологических условиях G-актин ( мономерная форма) превращается в F-актин ( полимерная форма) под действием АТФ, где роль АТФ является существенной. [ 96 ]
Спиральная нить F-актина, обнаруженная в мышцах, также содержит молекулу тропомиозина , которая представляет собой белок длиной 40 нанометров , обернутый вокруг спирали F-актина. [ 97 ] Во время фазы покоя тропомиозин покрывает активные участки актина, поэтому взаимодействие актина и миозина не может происходить и вызывать мышечное сокращение. Есть и другие белковые молекулы, связанные с тропомиозиновой нитью, это тропонины , имеющие три полимера: тропонин I , тропонин Т и тропонин С. [ 98 ]
F-актин одновременно силен и динамичен. В отличие от других полимеров , таких как ДНК , составные элементы которых связаны между собой ковалентными связями , мономеры актиновых нитей собираются более слабыми связями. [ 99 ] Латеральные связи с соседними мономерами устраняют эту аномалию, которая теоретически должна ослабить структуру, поскольку они могут быть разрушены термическим перемешиванием. Кроме того, слабые связи дают то преимущество, что концы нитей могут легко высвобождать или включать мономеры. Это означает, что нити могут быстро реконструироваться и изменять клеточную структуру в ответ на раздражитель окружающей среды. Это явление, наряду с биохимическим механизмом его возникновения, известно как «динамика сборки». [ 6 ]
Складной
[ редактировать ]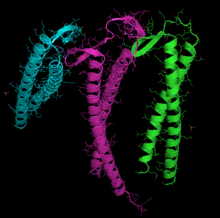
Актин может спонтанно приобретать большую часть своей третичной структуры . [ 101 ] Однако способ, которым он приобретает свою полнофункциональную форму из вновь синтезированной нативной формы, является особенным и почти уникальным в химии белков. Причиной этого особого пути может быть необходимость избежать присутствия неправильно свернутых мономеров актина, которые могут быть токсичными, поскольку могут действовать как неэффективные терминаторы полимеризации. Тем не менее, это ключ к установлению стабильности цитоскелета и, кроме того, это важный процесс координации клеточного цикла . [ 102 ] [ 103 ]
CCT необходим для того, чтобы обеспечить правильное складывание. CCT представляет собой шаперонин группы II, большой белковый комплекс, который способствует сворачиванию других белков. CCT состоит из двойного кольца из восьми различных субъединиц (гетерооктамерных) и отличается от шаперонинов группы I, таких как GroEL , который встречается у эубактерий и в эукариотических органеллах, поскольку ему не требуется ко-шаперон, действующий как крышка. над центральной каталитической полостью. Субстраты связываются с CCT через определенные домены. Первоначально считалось, что он связывается только с актином и тубулином , хотя недавние исследования иммунопреципитации показали, что он взаимодействует с большим количеством полипептидов , которые, возможно, действуют как субстраты . Он действует посредством АТФ-зависимых конформационных изменений, которые иногда требуют нескольких циклов высвобождения и катализа для завершения реакции. [ 104 ]
Чтобы успешно завершить сворачивание, и актину, и тубулину необходимо взаимодействовать с другим белком, называемым префолдином , который представляет собой гетерогексамерный комплекс (образованный шестью отдельными субъединицами), причем взаимодействие настолько специфично, что молекулы эволюционировали совместно. [ нужна ссылка ] . Актин образует комплексы с префолдином, пока он еще формируется, когда его длина составляет примерно 145 аминокислот , особенно на N-конце. [ 105 ]
Для актина и тубулина используются разные субъединицы распознавания, хотя есть некоторое перекрытие. В актине субъединицами, которые связываются с префолдином, вероятно, являются PFD3 и PFD4, которые связываются в двух местах: одно между остатками 60–79, а другое – между остатками 170–198. Актин распознается, загружается и доставляется к цитозольному шаперонину (CCT) в открытой конформации внутренним концом «щупалец» префолдина (см. изображение и примечание). [ 101 ] Контакт при доставке актина настолько краток, что третичный комплекс не образуется, немедленно освобождая префолдин. [ 100 ]

Затем CCT вызывает последовательное сворачивание актина, образуя связи с его субъединицами, а не просто заключая его в свою полость. [ 106 ] Вот почему он обладает специфическими областями узнавания в своем апикальном β-домене. Первый этап сворачивания состоит из узнавания остатков 245–249. Далее другие детерминанты устанавливают контакт. [ 107 ] И актин, и тубулин связываются с CCT в открытых конформациях в отсутствие АТФ. В случае актина при каждом конформационном изменении связываются две субъединицы, тогда как связывание тубулина происходит с четырьмя субъединицами. Актин имеет специфические связывающие последовательности, которые взаимодействуют с субъединицами δ и β-CCT или с δ-CCT и ε-CCT. После связывания AMP-PNP с CCT субстраты перемещаются внутри полости шаперонина. Также кажется, что в случае актина белок CAP необходим в качестве возможного кофактора в финальных состояниях сворачивания актина. [ 103 ]
Точный способ регуляции этого процесса до сих пор до конца не изучен, но известно, что белок PhLP3 (белок, аналогичный фосдуцину ) ингибирует его активность посредством образования третичного комплекса. [ 104 ]
Каталитический механизм АТФазы
[ редактировать ]Актин представляет собой АТФазу , что означает, что это фермент , гидролизующий АТФ. Эта группа ферментов характеризуется медленной скоростью реакции. Известно, что эта АТФаза является «активной», то есть ее скорость увеличивается примерно в 40 000 раз, когда актин входит в состав нити. [ 93 ] Эталонное значение этой скорости гидролиза в идеальных условиях составляет около 0,3 с. −1 . Затем Pi остается связанным с актином рядом с АДФ в течение длительного времени, пока он не будет кооперативно освобожден из внутренней части нити. [ 108 ] [ 109 ]
Точные молекулярные детали каталитического механизма до сих пор полностью не изучены. Хотя по этому вопросу ведется много споров, кажется очевидным, что для гидролиза АТФ необходима «закрытая» конформация и считается, что остатки, участвующие в этом процессе, перемещаются на соответствующее расстояние. [ 93 ] Глутаминовая кислота Glu137 является одним из ключевых остатков, который расположен в субдомене 1. Его функция заключается в связывании молекулы воды, которая производит нуклеофильную атаку АТФ на γ-фосфатную связь , в то время как нуклеотид прочно связан с субдоменами 3 и 4. Медленность каталитического процесса обусловлена большим расстоянием и перекосом положения молекулы воды по отношению к реагенту. Весьма вероятно, что конформационные изменения, вызванные вращением доменов между формами G и F актина, перемещают Glu137 ближе, обеспечивая его гидролиз. Эта модель предполагает, что полимеризация и функция АТФазы сразу же будут разделены. [ 94 ] [ 97 ] Превращение «открытого» в «закрытое» между формами G и F и его влияние на относительное движение нескольких ключевых остатков и образование водных проволок были охарактеризованы с помощью молекулярной динамики и моделирования QM/MM . [ 110 ] [ 111 ]
Динамика сборки
[ редактировать ]
Актиновые нити часто быстро собираются и разбираются, что позволяет им генерировать силу и поддерживать движение клеток. [ 112 ] Сборка классически происходит в три этапа. Во-первых, это «фаза нуклеации», в которой две-три молекулы G-актина медленно соединяются, образуя небольшой олигомер, который будет способствовать дальнейшему росту. Во-вторых, «фаза элонгации», когда актиновая нить быстро растет за счет присоединения множества молекул актина к обоим концам. По мере роста нити молекулы актина добавляются к (+) концу нити примерно в 10 раз быстрее, чем к (-) концу, и поэтому нити имеют тенденцию расти в основном на (+) конце. [ 113 ] В-третьих, «стационарная фаза», когда равновесие достигается, когда молекулы актина присоединяются к нити и покидают ее с одинаковой скоростью, сохраняя длину нити. [ 112 ] Хотя длина нити остается постоянной в стационарной фазе, новые молекулы постоянно добавляются к (+) концу и отпадают от (-) конца. Это явление называется «беговой дорожкой», поскольку данная молекула актина, по-видимому, движется вдоль нее. прядь. [ 114 ] Изолировано, будет ли нить расти или сокращаться и как быстро, определяется концентрацией G-актина вокруг нити; [ 113 ] однако в клетках на динамику актиновых филаментов сильно влияют различные актин-связывающие белки .
Актин-связывающие белки
[ редактировать ]Актиновый цитоскелет in vivo не состоит исключительно из актина, для его формирования, продолжения и функционирования необходимы другие белки. Эти белки называются актин-связывающими белками , и они участвуют в полимеризации, деполимеризации, стабильности и организации актина. [ 115 ] Разнообразие этих белков таково, что актин считается белком, принимающим участие в наибольшем количестве белок-белковых взаимодействий . [ 116 ]

Зародышеобразованию новых актиновых филаментов – этапу, ограничивающему скорость полимеризации актина – способствуют белки, образующие ядра актина, такие как формины (например, формин-2 ) и комплекс Arp2/3 . [ 118 ] Формины помогают образовывать длинные актиновые нити. Они связывают две свободные молекулы актина-АТФ, объединяя их. Затем, когда нить начинает расти, формин движется вдоль (+) конца растущей нити, все время рекрутируя актин-связывающие белки, которые способствуют росту нити, и исключая кэпирующие белки, которые блокируют расширение нити. [ 118 ] Ветви актиновых филаментов обычно зарождаются комплексом Arp2/3 совместно с факторами, способствующими нуклеации . Факторы, способствующие нуклеации, связывают две свободные молекулы G-актина, затем рекрутируют и активируют комплекс Arp2/3. Активированный комплекс Arp2/3 прикрепляется к существующей актиновой нити и использует две связанные молекулы G-актина для образования новой актиновой нити, ответвляющейся от старой под углом 70°. [ 119 ]

По мере роста нитей пул доступных молекул G-актина управляется G-актин-связывающими белками, такими как профилин и тимозин β-4 . Профилин обеспечивает поставку доступного актин-АТФ путем связывания с АДФ-связанным G-актином и способствуя обмену АДФ на АТФ. Связывание профилина с молекулой актина физически блокирует его присоединение к (-) концу нити, но позволяет ему присоединиться к (+) концу. Как только актин-АТФ присоединяется к нити, профилин высвобождает его. [ 114 ] Поскольку формины способствуют зарождению и удлинению новых актиновых нитей, они привлекают профилин в эту область, увеличивая локальную концентрацию актин-АТФ, чтобы стимулировать рост нитей. [ 118 ] Напротив, тимозин β-4 связывает и изолирует актин-АТФ, предотвращая его соединение с микрофиламентами. [ 121 ]
После того как актиновое волокно сформировано, на динамику его роста или коллапса влияют многочисленные белки. Существующие нити могут быть прерваны расщепляющими нити белками, такими как кофилин и гельзолин . Кофилин связывается с двумя молекулами актина-АДФ в нити, вызывая движение, которое дестабилизирует нить и приводит к ее разрыву. [ 122 ] Гельсолин вставляется между молекулами актина в нити, разрушая ее. После разрыва нити гельзолин остается прикрепленным к новому (+) концу, не позволяя ей расти и тем самым заставляя ее разбирать. [ 121 ]

Другие белки связываются с концами актиновых нитей, стабилизируя их. Они называются «кэпирующими белками» и включают CapZ и тропомодулин . CapZ связывает (+) конец нити, предотвращая дальнейшее добавление или потерю актина с этого конца. [ 121 ] Тропомодулин связывается с (-) концом нити, снова предотвращая добавление или потерю молекул на этом конце. Тропомодулин обычно обнаруживается в клетках, которым необходимы чрезвычайно стабильные актиновые нити, например, в мышцах и эритроцитах. [ 121 ]
Эти актин-связывающие белки обычно регулируются различными клеточными сигналами для контроля динамики сборки актина в разных клеточных местах. Формины, например, обычно сворачиваются в неактивную конформацию до тех пор, пока они не активируются связыванием небольшой ГТФазы Rho . [ 118 ] Разветвление актина на клеточной мембране важно для движения клеток, поэтому липид плазматической мембраны PIP 2 активирует фактор, способствующий нуклеации WASp , и ингибирует CapZ. [ 123 ] WASp также активируется небольшой GTPase Cdc42 , в то время как другой фактор, способствующий нуклеации WAVE, активируется GTPase Rac1 . [ 124 ]
Генетика
[ редактировать ]
Хотя большинство дрожжей имеют только один ген актина, высшие эукариоты , как правило, экспрессируют несколько изоформ актина, кодируемых семейством родственных генов. У млекопитающих имеется по крайней мере шесть изоформ актина, кодируемых отдельными генами. [ 125 ] которые делятся на три класса – альфа, бета и гамма – в зависимости от их изоэлектрических точек . В целом альфа-актины обнаруживаются в мышцах ( α-скелетные , α-аортальные гладкие , α-сердечные ), тогда как бета- и гамма-изоформы преобладают в немышечных клетках ( β-цитоплазматические , γ1-цитоплазматические , γ2-кишечные гладкие ). . Хотя аминокислотные последовательности и in vitro свойства изоформ очень схожи, эти изоформы не могут полностью заменить друг друга in vivo . [ 126 ] Растения содержат более 60 актиновых генов и псевдогенов . [ 85 ]
Типичный ген актина имеет примерно 100-нуклеотидную 5'-UTR область из 1200 нуклеотидов , транслируемую и 200-нуклеотидную 3'-UTR . Большинство генов актина прерываются интронами , причем в любом из 19 хорошо изученных мест может находиться до шести интронов. Высокая консервативность семейства делает актин предпочтительной моделью для исследований, сравнивающих модели эволюции интронов с ранними и поздними интронами.
Эволюция
[ редактировать ]Актин и родственные ему белки присутствуют во всех организмах, что позволяет предположить, что у общего предка всей жизни на Земле был актин. [ 127 ] Актин — один из наиболее консервативных белков на протяжении всей эволюции эукариот. Последовательности актиновых белков животных и амеб на 80% идентичны, несмотря на то, что их разделяет примерно один миллиард лет эволюции. [ 85 ] Многие одноклеточные эукариоты имеют один ген актина, тогда как многоклеточные эукариоты часто имеют несколько близкородственных генов, выполняющих специализированные функции. У людей их шесть; растения имеют 10 и более. [ 127 ] Помимо актина, у эукариот есть большое семейство родственных актину белков, или «Arps», которые имеют общего с актином предка и называются Arp1–Arp11, причем Arp1 наиболее тесно связан с актином, а Arp11 — наименее. [ 127 ]
Бактерии кодируют три типа актина: MreB влияет на форму клеток, деление клеток FtsA и ParM разделение крупных плазмид . [ 127 ] Некоторые археи имеют бактериоподобный ген MreB, тогда как другие имеют ген актина, который больше напоминает актин эукариот. [ 127 ]
Эукариотический цитоскелет организмов всех таксономических групп имеет компоненты, сходные с актином и тубулином. Например, белок, кодируемый геном ACTG2 у человека, полностью эквивалентен гомологам , присутствующим у крыс и мышей, хотя на нуклеотидном уровне сходство снижается до 92%. [ 128 ] Однако существуют серьезные различия с эквивалентами у прокариот ( FtsZ и MreB ), где сходство между нуклеотидными последовательностями составляет от 40 до 50% среди разных видов бактерий и архей . Некоторые авторы предполагают, что предковый белок, давший начало модельному эукариотическому актину, напоминает белки, присутствующие в современных бактериальных цитоскелетах. [ 4 ] [ 129 ]

Некоторые авторы отмечают, что поведение актина, тубулина и гистона (белка, участвующего в стабилизации и регуляции ДНК) сходно по своей способности связывать нуклеотиды и способности использовать преимущества броуновского движения . Также было высказано предположение, что все они имеют общего предка. [ 130 ] Таким образом, эволюционные процессы привели к диверсификации предковых белков в разновидности, существующие сегодня, сохраняя, среди прочего, актины как эффективные молекулы, которые были способны решать важные наследственные биологические процессы, такие как эндоцитоз . [ 131 ]
Комплекс Arp2/3 широко обнаружен во всех эукариотических организмах. [ 132 ]
Эквиваленты у прокариот
[ редактировать ]Бактериальный цитоскелет содержит белки, очень похожие на мономеры и полимеры актина. Бактериальный белок MreB полимеризуется в тонкие неспиральные нити, а иногда и в спиральные структуры, подобные F-актину. [ 94 ] [ 133 ] Более того, его кристаллическая структура очень похожа на структуру G-актина (с точки зрения его трехмерной конформации), есть даже сходство между протофиламентами MreB и F-актином. Бактериальный цитоскелет также содержит белки FtsZ , сходные с тубулином . [ 134 ]
Таким образом, бактерии обладают цитоскелетом с элементами, гомологичными актину (например, MreB, AlfA, ParM , FtsA и MamK), хотя аминокислотная последовательность этих белков отличается от таковой, присутствующей в клетках животных. Однако такие белки имеют высокую степень структурного сходства с эукариотическим актином. Высокодинамичные микрофиламенты, образующиеся в результате агрегации MreB и ParM, необходимы для жизнеспособности клеток и участвуют в клеточном морфогенезе, сегрегации хромосом и полярности клеток. ParM представляет собой гомолог актина, который закодирован в плазмиде и участвует в регуляции плазмидной ДНК. [ 4 ] [ 135 ] ParMs из разных бактериальных плазмид могут образовывать удивительно разнообразные спиральные структуры, состоящие из двух [ 136 ] [ 137 ] или четыре [ 138 ] нити для поддержания достоверного наследования плазмиды.
У архей гомолог Та0583 еще больше похож на эукариотические актины. [ 139 ]
Молекулярная патология
[ редактировать ]Большинство млекопитающих обладают шестью различными генами актина . Из них два кодируют цитоскелет ( ACTB и ACTG1 ), а остальные четыре участвуют в скелетных поперечно-полосатых мышцах ( ACTA1 ), гладкой мышечной ткани ( ACTA2 ), кишечных мышцах ( ACTG2 ) и сердечной мышце ( ACTC1 ). Актин цитоскелета участвует в механизмах патогенеза многих инфекционных агентов , включая ВИЧ . Подавляющее большинство мутаций , влияющих на актин, являются точковыми мутациями, имеющими доминантный эффект , за исключением шести мутаций, участвующих в немалиновой миопатии . Это связано с тем, что во многих случаях мутант мономера актина действует как «кэп», предотвращая удлинение F-актина. [ 88 ]
Патология, связанная с ACTA1
[ редактировать ]ACTA1 — это ген, который кодирует α- изоформу актина, которая преобладает в скелетных поперечно-полосатых мышцах человека , хотя он также экспрессируется в сердечной мышце и щитовидной железе . [ 140 ] Его последовательность ДНК состоит из семи экзонов , которые производят пять известных транскриптов . [ 141 ] Большинство из них состоят из точковых мутаций, вызывающих замену аминокислот . Мутации во многих случаях связаны с фенотипом , который определяет тяжесть и течение заболевания. [ 88 ] [ 141 ]
Мутация изменяет структуру и функцию скелетных мышц, вызывая одну из трех форм миопатии : немалиновую миопатию 3-го типа , врожденную миопатию с избытком тонких миофиламентов (СМ) и врожденную миопатию с диспропорцией типов волокон (ВМФТД). Также были обнаружены мутации, которые вызывают основные миопатии . [ 143 ] Хотя их фенотипы схожи, некоторые специалисты помимо типичной немалиновой миопатии выделяют еще один тип миопатии, называемый актинической немалиновой миопатией. В первом случае вместо типичных палочек образуются сгустки актина. Важно отметить, что у пациента может быть выявлено более одного из этих фенотипов при биопсии . [ 144 ] Наиболее распространенные симптомы включают типичную морфологию лица (миопатическое лицо ), мышечную слабость, задержку моторного развития и затруднения дыхания. Течение заболевания, его тяжесть и возраст, в котором оно появляется, варьируются, также встречаются перекрывающиеся формы миопатии. Симптомом немалиновой миопатии является появление «немалиновых стержней» в разных местах мышечных волокон 1 типа. Эти палочки представляют собой непатогномоничные структуры, имеющие сходный состав с Z-дисками, обнаруженными в саркомере . [ 145 ]
Патогенез . этой миопатии весьма разнообразен Многие мутации происходят в области углубления актина рядом с его сайтами связывания нуклеотидов , тогда как другие происходят в домене 2 или в областях, где происходит взаимодействие со связанными белками. Это в некоторой степени объясняет большое разнообразие комков, образующихся в этих случаях, таких как немалиновые, внутриядерные тельца или тельца-зебры. [ 88 ] актина, При немалиновой миопатии происходят изменения в сворачивании а также изменения в его агрегации, а также изменения в экспрессии других ассоциированных белков. В некоторых вариантах, когда обнаруживаются внутриядерные тельца, изменения в складчатости маскируют сигнал экспорта белка из ядра происходит накопление мутантной формы актина , так что в ядре клетки . [ 146 ] С другой стороны, оказывается, что мутации ACTA1 , приводящие к CFTDM, оказывают большее влияние на функцию саркомера, чем на его структуру. [ 147 ] Недавние исследования попытались понять этот очевидный парадокс, который предполагает отсутствие четкой корреляции между количеством палочек и мышечной слабостью. Похоже, что некоторые мутации способны вызывать более высокую скорость апоптоза в мышечных волокнах II типа. [ 102 ]
В гладких мышцах
[ редактировать ]Есть две изоформы, которые кодируют актины гладкой мышечной ткани :
ACTG2 кодирует самую крупную изоформу актина, имеющую девять экзонов , один из которых, расположенный на 5'-конце, не транслируется . [ 128 ] Это γ-актин, который экспрессируется в гладких мышцах кишечника. Никаких мутаций этого гена, соответствующих патологиям, обнаружено не было, хотя микрочипы показали, что этот белок чаще экспрессируется в случаях, устойчивых к химиотерапии с использованием цисплатина . [ 148 ]
ACTA2 кодирует α-актин, расположенный в гладких мышцах, а также в гладких мышцах сосудов. Было отмечено, что мутация MYH11 может быть ответственной по крайней мере за 14% наследственных аневризм грудной аорты, особенно типа 6. Это связано с тем, что мутировавший вариант вызывает неправильную сборку филаментов и снижение способности к сокращению гладких мышц сосудов. деградация среды аорты У этих людей была зафиксирована с участками дезорганизации и гиперплазии , а также стенозом аорты vasa vasorum . [ 149 ] Число заболеваний, в которых замешан этот ген, увеличивается. Это связано с болезнью Моямоя , и вполне вероятно, что определенные мутации при гетерозиготности могут приводить к предрасположенности ко многим сосудистым патологиям, таким как аневризма грудной аорты и ишемическая болезнь сердца . [ 150 ] Альфа-актин, обнаруженный в гладких мышцах, также является интересным маркером для оценки прогресса цирроза печени . [ 151 ]
В сердечной мышце
[ редактировать ]Ген ACTC1 кодирует изоформу α-актина, присутствующую в сердечной мышце. Впервые он был секвенирован Хамадой и его коллегами в 1982 году, когда было обнаружено, что он прерывается пятью интронами. [ 152 ] Это был первый из шести генов, в котором были обнаружены аллели, вовлеченные в патологические процессы. [ 153 ]
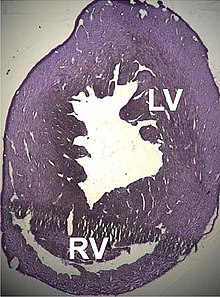
Описан ряд структурных нарушений, связанных с точечными мутациями этого гена, вызывающих нарушение работы сердца, таких как дилатационная кардиомиопатия типа 1R и гипертрофическая кардиомиопатия типа 11 . некоторые дефекты межпредсердной перегородки , которые также могут быть связаны с этими мутациями. Недавно были описаны [ 155 ] [ 156 ]
Были изучены два случая дилатационной кардиомиопатии, включающие замену высококонсервативных аминокислот, принадлежащих белковым доменам , которые связываются и перемежаются с Z-дисками . Это привело к теории, что дилатация возникает из-за дефекта передачи сократительной силы в миоцитах . [ 157 ] [ 153 ]
Мутации ACTC1 ответственны как минимум за 5% гипертрофических кардиомиопатий. [ 158 ] Также было обнаружено существование ряда точковых мутаций: [ 159 ]
- Мутация Е101К: изменение суммарного заряда и образование слабой электростатической связи в актомиозин-связывающем сайте.
- P166A: зона взаимодействия между мономерами актина.
- A333P: зона взаимодействия актина и миозина.
Патогенез, по-видимому, включает компенсаторный механизм: мутировавшие белки действуют как токсины с доминирующим эффектом, снижая способность сердца сокращаться, вызывая аномальное механическое поведение, так что гипертрофия, которая обычно задерживается, является следствием нормальной реакции сердечной мышцы на стресс. . [ 160 ]
Недавние исследования обнаружили мутации ACTC1, которые участвуют в двух других патологических процессах: детской идиопатической рестриктивной кардиомиопатии , [ 161 ] и некомпактность миокарда левого желудочка . [ 162 ]
В цитоплазматических актинах
[ редактировать ]ACTB — очень сложный локус . Существует ряд псевдогенов , которые распределены по всему геному , и их последовательность содержит шесть экзонов, которые могут вызывать до 21 различной транскрипции путем альтернативного сплайсинга , которые известны как β-актины. В соответствии с этой сложностью, его продукты также обнаруживаются в ряде мест и являются частью широкого спектра процессов ( цитоскелет , гистона комплекс -ацилтрансферазы NuA4, ядро клетки ), а также связаны с механизмами большого числа процессов. патологических процессов ( карциномы , ювенильная дистония , механизмы инфекции, пороки развития нервной системы и опухолевая инвазия и др.). [ 163 ] Была обнаружена новая форма актина, каппа-актин, которая, по-видимому, заменяет β-актин в процессах, связанных с опухолями . [ 164 ]

На данный момент открыты три патологических процесса, вызванных прямым изменением последовательности генов:
- Гемангиоперицитома с t(7;12)(p22;q13)-транслокациями является редким заболеванием, при котором транслокационная мутация вызывает слияние гена ACTB с GLI1 в хромосоме 12 . [ 166 ]
- Ювенильная дистония — редкое дегенеративное заболевание , поражающее центральную нервную систему ; в частности, он поражает участки неокортекса и таламуса , палочковидные эозинофильные где образуются включения. Затронутые люди представляют фенотип с деформациями по срединной линии, сенсорной тугоухостью и дистонией. Оно вызвано точечной мутацией, при которой аминокислота триптофан заменяет аргинин в положении 183. Это изменяет взаимодействие актина с системой АДФ/ кофилин , которая регулирует динамику формирования цитоскелета нервных клеток . [ 167 ]
- Также была обнаружена доминантная точечная мутация, которая вызывает дисфункцию нейтрофильных гранулоцитов и рецидивирующие инфекции . Похоже, что мутация модифицирует домен, ответственный за связывание профилина с другими регуляторными белками. Сродство актина к профилину в этом аллеле значительно снижено. [ 168 ]
Локус ACTG1 кодирует цитозольный белок γ-актин, который отвечает за образование микрофиламентов цитоскелета . Он содержит шесть экзонов , дающих начало 22 различным мРНК , которые производят четыре полные изоформы , форма экспрессии которых, вероятно, зависит от типа ткани, в которой они находятся. Он также имеет два разных промотора ДНК . [ 169 ] Было отмечено, что последовательности, транслируемые из этого локуса и из β-актина, очень похожи на предсказанные, что указывает на общую предковую последовательность, претерпевшую дупликацию и генетическую конверсию. [ 170 ]
С точки зрения патологии его связывают с такими процессами, как амилоидоз , пигментный ретинит , механизмами инфекции, заболеваниями почек и различными видами врожденной тугоухости. [ 169 ]
Было обнаружено, что шесть аутосомно-доминантных точковых мутаций в этой последовательности вызывают различные типы потери слуха, в частности нейросенсорную тугоухость, связанную с локусом DFNA 20/26. По-видимому, они влияют на стереоцилии внутреннего уха реснитчатых клеток, присутствующих в кортиевом органе . β-актин является наиболее распространенным белком, обнаруженным в тканях человека, но его мало в реснитчатых клетках, что объясняет локализацию патологии. С другой стороны, похоже, что большинство этих мутаций затрагивают области, участвующие в связывании с другими белками, особенно с актомиозином. [ 88 ] Некоторые эксперименты показали, что патологический механизм этого типа потери слуха связан с тем, что F-актин в мутациях более чувствителен к кофилину, чем обычно. [ 171 ]
Однако, хотя ни одного случая не зарегистрировано, известно, что γ-актин также экспрессируется в скелетных мышцах, и хотя он присутствует в небольших количествах, модельные организмы показали, что его отсутствие может привести к миопатиям. [ 172 ]
Другие патологические механизмы
[ редактировать ]актин, особенно цитоплазматический актин Некоторые инфекционные агенты используют в своем жизненном цикле . присутствуют две основные формы У бактерий :
- Listeria monocytogenes , некоторые виды Rickettsia , Shigella flexneri и другие внутриклеточные микробы покидают фагоцитарные вакуоли, покрываясь капсулой из актиновых нитей. L. monocytogenes и S. flexneri образуют хвост в форме «хвоста кометы», который придает им подвижность. Каждый вид имеет небольшие различия в механизме молекулярной полимеризации своих «хвостов кометы». Наблюдались разные скорости перемещения, например, листерии и шигеллы оказались самыми быстрыми. [ 173 ] Многие эксперименты продемонстрировали этот механизм in vitro . Это указывает на то, что бактерии не используют миозиноподобный белковый мотор, и, по-видимому, их движение достигается за счет давления, оказываемого полимеризацией, которая происходит вблизи клеточной стенки микроорганизма. Бактерии ранее были окружены ABP хозяина, и как минимум покрытие содержит комплекс Arp2/3 , белки Ena/VASP , кофилин, буферный белок и промоторы нуклеации, такие как винкулин комплекс . Благодаря этим движениям они образуют выступы, которые достигают соседних клеток, заражая и их, так что иммунная система может бороться с инфекцией только посредством клеточного иммунитета. Движение могло быть вызвано изменением кривой и разветвлением нитей. [ 174 ] Другие виды, такие как Mycobacterium marinum и Burkholderia pseudomallei , также способны к локализованной полимеризации клеточного актина, чтобы способствовать их движению посредством механизма, сосредоточенного на комплексе Arp2/3. Кроме того, вакцинный вирус коровьей оспы также использует для своего распространения элементы актинового цитоскелета. [ 175 ]
- Pseudomonas aeruginosa способна образовывать защитную биопленку , чтобы избежать защиты организма-хозяина , особенно лейкоцитов и антибиотиков . Биопленка построена с использованием ДНК и актиновых нитей организма-хозяина. [ 176 ]
В дополнение к ранее приведенному примеру, полимеризация актина стимулируется на начальных стадиях интернализации некоторых вирусов, особенно ВИЧ , например, путем инактивации комплекса кофилина. [ 177 ]
Роль актина в процессе инвазии раковых клеток до сих пор не определена. [ 178 ]
Приложения
[ редактировать ]Актин используется в научных и технологических лабораториях в качестве проводника молекулярных моторов, таких как миозин (как в мышечной ткани, так и вне ее), а также как необходимый компонент для функционирования клеток. Его также можно использовать как диагностический инструмент, поскольку несколько его аномальных вариантов связаны с появлением специфических патологий.
- Нанотехнологии . Актин-миозиновые системы действуют как молекулярные моторы, которые обеспечивают транспорт везикул и органелл по цитоплазме. Вполне возможно, что актин может быть применен в нанотехнологиях , поскольку его динамические способности были использованы в ряде экспериментов, в том числе проведенных в бесклеточных системах. Основная идея состоит в том, чтобы использовать микрофиламенты в качестве дорожек для управления молекулярными двигателями, которые могут транспортировать заданный груз. То есть актин можно использовать для определения контура, по которому груз может транспортироваться более или менее контролируемым и направленным образом. Что касается общего применения, его можно использовать для направленного транспорта молекул для осаждения в определенных местах, что позволит осуществлять контролируемую сборку наноструктур. [ 179 ] Эти атрибуты могут быть применены к лабораторным процессам, таким как « лаборатория на чипе» , в механике нанокомпонентов и в нанотрансформаторах, которые преобразуют механическую энергию в электрическую. [ 180 ]
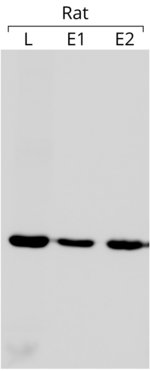
- Актин используется в качестве внутреннего контроля в вестерн-блоттинге, чтобы убедиться, что на каждую дорожку геля загружено одинаковое количество белка. В примере блота, показанном слева, в каждую лунку было загружено 75 мкг общего белка. Блот подвергался реакции с антителом против β-актина (более подробную информацию о блоте см. в ссылке). [ 181 ] )
Использование актина в качестве внутреннего контроля основано на предположении, что его экспрессия практически постоянна и не зависит от условий эксперимента. Сравнивая экспрессию интересующего гена с экспрессией актина, можно получить относительную величину, которую можно сравнивать между различными экспериментами. [ 182 ] всякий раз, когда выражение последнего постоянно. Стоит отметить, что актин не всегда обладает желаемой стабильностью экспрессии генов . [ 183 ]
- Здоровье. Некоторые аллели актина вызывают заболевания; по этой причине были разработаны методы их обнаружения. Кроме того, актин может быть использован в качестве косвенного маркера при хирургической патологии: возможно использование вариаций характера его распределения в тканях в качестве маркера инвазии при неоплазиях , васкулитах и других состояниях. [ 184 ] Кроме того, из-за тесной связи актина с аппаратом мышечного сокращения его уровень в скелетных мышцах снижается при атрофии этих тканей , поэтому его можно использовать в качестве маркера этого физиологического процесса. [ 185 ]
- Пищевая технология . , можно определить Качество некоторых обработанных пищевых продуктов, таких как колбасы путем количественного определения количества актина, присутствующего в составе мяса. Традиционно использовался метод, основанный на обнаружении 3-метилгистидина в гидролизованных образцах этих продуктов, поскольку это соединение присутствует в актине и тяжелой цепи F-миозина (оба являются основными компонентами мышц). Образование этого соединения в мясе происходит в результате метилирования остатков гистидина , присутствующих в обоих белках. [ 186 ] [ 187 ]
История
[ редактировать ]
обнаружен Актин был впервые экспериментально в 1887 году У.Д. Халлибертоном , который извлек из мышц белок, который «коагулировал» препараты миозина , которые он назвал «миозиновым ферментом». [ 188 ] Однако компания Halliburton не смогла уточнить свои выводы, и вместо этого открытие актина приписывают Бруно Ференцу Штраубу , молодому биохимику, работающему в лаборатории Альберта Сент-Дьёрдьи в Институте медицинской химии Сегедского университета , Венгрия .
Вслед за открытием Илоны Банги и Сент-Дьёрдьи в 1941 году, что коагуляция происходит только при некоторых экстракциях миозина и обращается вспять при добавлении АТФ, [ 189 ] Штрауб идентифицировал и очистил актин из тех препаратов миозина, которые коагулировали. Основываясь на оригинальном методе экстракции Банги, он разработал новую технику экстракции мышечного белка, которая позволила ему выделить значительные количества относительно чистого актина, опубликованную в 1942 году. [ 190 ] Метод Штрауба по сути такой же, как тот, который используется сегодня в лабораториях . Поскольку белок Штрауба был необходим для активации коагуляции миозина, его назвали актином . [ 189 ] [ 191 ] Понимая, что коагулирующие препараты миозина Банги содержат также и актин, Сент-Дьёрдьи назвал смесь обоих белков актомиозином . [ 192 ]
Военные действия Второй мировой войны привели к тому, что Сент-Дьёрдьи не смог опубликовать работу своей лаборатории в западных научных журналах . Таким образом, актин стал хорошо известен на Западе только в 1945 году, когда их статья была опубликована в качестве приложения к Acta Physiologica Scandinavica . [ 193 ] Штрауб продолжал работать над актином и в 1950 году сообщил, что актин содержит связанный АТФ. [ 194 ] и что во время гидролизуется до АДФ и полимеризации белка в микрофиламенты нуклеотид неорганического . фосфата ( которые остаются связанными с микрофиламентами) Штрауб предположил, что трансформация АТФ-связанного актина в АДФ-связанный актин играет роль в мышечном сокращении. На самом деле это справедливо только для гладких мышц и не было подтверждено экспериментами до 2001 года. [ 194 ] [ 195 ]
Аминокислотное секвенирование актина было завершено М. Эльзингой и его сотрудниками в 1973 году. [ 86 ] Кристаллическая структура G-актина была раскрыта в 1990 году Кабшем и его коллегами. [ 89 ] В том же году Холмс и его коллеги предложили модель F-актина после экспериментов по совместной кристаллизации с различными белками. [ 91 ] Процедура сокристаллизации с различными белками использовалась неоднократно в последующие годы, пока в 2001 г. выделенный белок не был кристаллизован вместе с АДФ. Однако до сих пор не существует рентгеновской структуры F-актина с высоким разрешением. Кристаллизация G-актина стала возможной благодаря использованию конъюгата родамина , который препятствует полимеризации, блокируя аминокислоту цис-374 . [ 1 ] Кристин Ориол-Аудит умерла в том же году, когда актин был впервые кристаллизован, но именно она была исследователем, который в 1977 году впервые кристаллизовал актин в отсутствие актин-связывающих белков (ABP). Однако полученные кристаллы были слишком малы для доступной технологии того времени. [ 196 ]
Хотя в настоящее время не существует модели нитевидной формы актина с высоким разрешением, в 2008 году команде Савайи удалось создать более точную модель его структуры на основе множества кристаллов димеров актина , которые связываются в разных местах. [ 197 ] Эта модель впоследствии была усовершенствована Савайей и Лоренцем. Другие подходы, такие как использование криоэлектронной микроскопии и синхротронного излучения , недавно позволили повысить разрешение и лучше понять природу взаимодействий и конформационных изменений, участвующих в формировании актиновых нитей. [ 198 ] [ 94 ] [ 97 ]
Исследовать
[ редактировать ]Химические ингибиторы
[ редактировать ]
Ряд природных токсинов , которые влияют на динамику актина, широко используются в исследованиях по изучению роли актина в биологии. Латрункулин – токсин, вырабатываемый губками , – связывается с G-актином, предотвращая его соединение с микрофиламентами. [ 199 ] Цитохалазин D , вырабатываемый некоторыми грибами , служит фактором кэпирования, связываясь с (+) концом нити и предотвращая дальнейшее добавление молекул актина. [ 199 ] Напротив, губчатый токсин ясплакинолид способствует зарождению новых актиновых нитей путем связывания и стабилизации пар молекул актина. [ 200 ] Фаллоидин – из «смертельной шапки» гриба Amanita phalloides – связывается с соседними молекулами актина внутри нити F-актина, стабилизируя нить и предотвращая ее деполимеризацию. [ 200 ]
Фаллоидин часто метят флуоресцентными красителями для визуализации актиновых нитей с помощью флуоресцентной микроскопии . [ 200 ]
См. также
[ редактировать ]- Ремоделирование актина — влияние на структуру и форму клеток
- Активная материя
- Филоподии
- Промежуточная нить
- Ламеллиподий
- Моторный белок — преобразует химическую энергию в механическую работу.
- Нейрон
- фаллотоксин
Ссылки
[ редактировать ]- ^ Jump up to: а б с д ВВП : 1J6Z ; Оттербейн Л.Р., Грасеффа П., Домингес Р. (июль 2001 г.). «Кристаллическая структура незакомплексованного актина в состоянии АДФ». Наука . 293 (5530): 708–711. дои : 10.1126/science.1059700 . ПМИД 11474115 . S2CID 12030018 .
- ^ Доэрти Дж.Дж., МакМахон Х.Т. (2008). «Посредничество, модуляция и последствия взаимодействий мембраны и цитоскелета». Ежегодный обзор биофизики . 37 (1): 65–95. doi : 10.1146/annurev.biophys.37.032807.125912 . ПМИД 18573073 . S2CID 17352662 .
- ^ Виндин Х., Ганнинг П. (август 2013 г.). «Цитоскелетные тропомиозины: хореографы функционального разнообразия актиновых филаментов» . Журнал исследований мышц и подвижности клеток . 34 (3–4): 261–274. дои : 10.1007/s10974-013-9355-8 . ПМЦ 3843815 . ПМИД 23904035 .
- ^ Jump up to: а б с д Ганнинг П.В., Гошдастидер Ю., Уитакер С., Попп Д., Робинсон Р.К. (июнь 2015 г.). «Эволюция композиционно и функционально различных актиновых нитей» . Журнал клеточной науки . 128 (11): 2009–2019. дои : 10.1242/jcs.165563 . ПМИД 25788699 .
- ^ Гошдастидер У., Цзян С., Попп Д., Робинсон Р.К. (июль 2015 г.). «В поисках первичной актиновой нити» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (30): 9150–9151. дои : 10.1073/pnas.1511568112 . ПМЦ 4522752 . ПМИД 26178194 .
- ^ Jump up to: а б Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Глава 16: Цитоскелет» . Молекулярная биология клетки . Нью-Йорк: Garland Science. стр. 907–982. ISBN 978-0-8153-3218-3 .
- ^ Jump up to: а б с Лодиш и др. 2016 , с. 778.
- ^ Хубер Ф., Шнаус Дж., Ренике С., Раух П., Мюллер К., Фюттерер К., Кас Дж. (январь 2013 г.). «Эмерджентная сложность цитоскелета: от одиночных нитей к ткани» . Достижения физики . 62 (1): 1–112. Бибкод : 2013AdPhy..62....1H . дои : 10.1080/00018732.2013.771509 . ПМЦ 3985726 . ПМИД 24748680 .
- ^ Jump up to: а б с Груммт I (апрель 2006 г.). «Актин и миозин как факторы транскрипции». Текущее мнение в области генетики и развития . 16 (2): 191–196. дои : 10.1016/j.где.2006.02.001 . ПМИД 16495046 .
- ^ Экерт Р., Рэндалл Д., Бурггрен В.В., Френч К. (2002). Эккерт Физиология животных: механизмы и приспособления . Нью-Йорк: WH Freeman and CO. ISBN 978-0-7167-3863-3 .
- ^ Jump up to: а б Паниагуа Р., Нистал М., Сесма П., Альварес-Уриа М., Фрайар Б., Анадон Р., Хосе Саес Ф. (2002). Цитология и гистология растений и животных (на испанском языке). McGraw-Hill Inter-American Испании, ISBN SAU 978-84-486-0436-3 .
- ^ Сюй К., Чжун Г, Чжуан Икс (январь 2013 г.). «Актин, спектрин и связанные с ними белки образуют периодическую структуру цитоскелета в аксонах» . Наука . 339 (6118): 452–456. Бибкод : 2013Sci...339..452X . дои : 10.1126/science.1232251 . ПМЦ 3815867 . ПМИД 23239625 .
- ^ Jump up to: а б Мозли Дж. Б., Гуд Б. Л. (сентябрь 2006 г.). «Актиновый цитоскелет дрожжей: от клеточной функции к биохимическому механизму» . Обзоры микробиологии и молекулярной биологии . 70 (3): 605–645. дои : 10.1128/MMBR.00013-06 . ПМЦ 1594590 . ПМИД 16959963 .
- ^ Мигер Р.Б., МакКинни ЕС, Кандасами М.К. (июнь 1999 г.). «Изовариантная динамика расширяет и буферизует реакции сложных систем: разнообразного семейства генов растительного актина» . Растительная клетка . 11 (6): 995–1006. дои : 10.1105/tpc.11.6.995 . ПМК 1464670 . ПМИД 10368172 .
- ^ PDB 1unc ; Вермюлен В., Ванезебрук П., Ван Тройс М., Вершуерен М., Фант Ф., Геталс М., Ампе К., Мартинс Х.К., Борреманс Ф.А. (май 2004 г.). «Структуры раствора С-концевых субдоменов головного мозга виллина и адвиллина человека, оценка требований к связыванию F-актина головного мозга» . Белковая наука . 13 (5): 1276–1287. дои : 10.1110/ps.03518104 . ПМК 2286768 . ПМИД 15096633 .
- ^ Jump up to: а б Хигаки Т., Сано Т., Хасадзава С. (декабрь 2007 г.). «Динамика актиновых микрофиламентов и актиновые боковые белки в растениях». Современное мнение в области биологии растений . 10 (6): 549–556. Бибкод : 2007COPB...10..549H . дои : 10.1016/j.pbi.2007.08.012 . ПМИД 17936064 .
- ^ Ковар Д.Р., Стайгер С.Дж., Уивер Э.А., МакКарди Д.В. (декабрь 2000 г.). «AtFim1 представляет собой белок, сшивающий актиновые нити из Arabidopsis thaliana» . Заводской журнал . 24 (5): 625–636. дои : 10.1046/j.1365-313x.2000.00907.x . ПМИД 11123801 .
- ^ Jump up to: а б Кларк Т.Г., Мерриам Р.В. (декабрь 1977 г.). «Диффузивные и связанные актиновые ядра ооцитов Xenopus laevis». Клетка . 12 (4): 883–891. дои : 10.1016/0092-8674(77)90152-0 . ПМИД 563771 . S2CID 34708250 .
- ^ Хофманн В.А. (1 января 2009 г.). Клеточная и молекулярная биология ядерного актина . Международное обозрение клеточной и молекулярной биологии. Том. 273. С. 219–263. дои : 10.1016/S1937-6448(08)01806-6 . ISBN 9780123748041 . ПМИД 19215906 .
- ^ Ульфертс С., Праджапати Б., Гросс Р., Вартиайнен М.К. (февраль 2021 г.). «Новые свойства и функции актина и актиновых нитей внутри ядра» . Перспективы Колд-Спринг-Харбор в биологии . 13 (3): а040121. doi : 10.1101/cshperspect.a040121 . ПМЦ 7919393 . ПМИД 33288541 .
- ^ Jump up to: а б Бонсак М.Т., Стювен Т., Кун С., Кордес В.К., Герлих Д. (март 2006 г.). «Селективная блокировка экспорта ядерного актина стабилизирует гигантские ядра ооцитов Xenopus». Природная клеточная биология . 8 (3): 257–263. дои : 10.1038/ncb1357 . hdl : 11858/00-001M-0000-0012-E6EB-9 . ПМИД 16489345 . S2CID 16529470 .
- ^ Допи Дж., Скарп К.П., Раякюля Э.К., Танхуанпяя К., Вартиайнен М.К. (февраль 2012 г.). «Активное поддержание ядерного актина импортином 9 поддерживает транскрипцию» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (9): E544–552. дои : 10.1073/pnas.1118880109 . ПМЦ 3295300 . ПМИД 22323606 .
- ^ Вада А., Фукуда М., Мисима М., Нисида Э. (март 1998 г.). «Ядерный экспорт актина: новый механизм, регулирующий субклеточную локализацию основного цитоскелетного белка» . Журнал ЭМБО . 17 (6): 1635–1641. дои : 10.1093/emboj/17.6.1635 . ПМК 1170511 . ПМИД 9501085 .
- ^ Стювен Т., Хартманн Э., Герлих Д. (ноябрь 2003 г.). «Экспортин 6: новый рецептор ядерного экспорта, специфичный для комплексов профилин-актин» . Журнал ЭМБО . 22 (21): 5928–5940. дои : 10.1093/emboj/cdg565 . ПМК 275422 . ПМИД 14592989 .
- ^ Хофманн В.А., Ардуини А., Николь С.М., Камачо С.Дж., Лессард Дж.Л., Фуллер-Пейс Ф.В., де Ланероль П. (июль 2009 г.). «СУМОилирование ядерного актина» . Журнал клеточной биологии . 186 (2): 193–200. дои : 10.1083/jcb.200905016 . ПМЦ 2717643 . ПМИД 19635839 .
- ^ Макдональд Д., Карреро Г., Андрин С., де Врис Г., Хендзель М.Дж. (февраль 2006 г.). «Нуклеоплазматический бета-актин существует в динамическом равновесии между полимерными видами с низкой подвижностью и быстро диффундирующими популяциями» . Журнал клеточной биологии . 172 (4): 541–552. дои : 10.1083/jcb.200507101 . ПМК 2063674 . ПМИД 16476775 .
- ^ Йокуш Б.М., Шененбергер К.А., Стетефельд Дж., Эби У (август 2006 г.). «Отслеживание различных форм ядерного актина». Тенденции клеточной биологии . 16 (8): 391–396. дои : 10.1016/j.tcb.2006.06.006 . ПМИД 16828286 .
- ^ Jump up to: а б с д Мигоцка-Патрзалек М., Маковецка А., Новак Д., Мазур А.Я., Хофманн В.А., Малицка-Блашкевич М. (ноябрь 2015 г.). «β- и γ-актины в ядре клеток меланомы человека А375» . Гистохимия и клеточная биология . 144 (5): 417–428. дои : 10.1007/s00418-015-1349-8 . ПМЦ 4628621 . ПМИД 26239425 .
- ^ Педерсон Т., Эби Ю (1 декабря 2002 г.). «Актин в ядре: какая форма и зачем?». Журнал структурной биологии . 140 (1–3): 3–9. дои : 10.1016/s1047-8477(02)00528-2 . ПМИД 12490148 .
- ^ Чхабра Д., dos Remedios CG (сентябрь 2005 г.). «Кофилин, актин и их комплекс наблюдаются in vivo с использованием резонансного переноса энергии флуоресценции» . Биофизический журнал . 89 (3): 1902–1908. Бибкод : 2005BpJ....89.1902C . дои : 10.1529/biophysj.105.062083 . ПМК 1366693 . ПМИД 15994898 .
- ^ Спенсер, В.А. (сентябрь 2011 г.). «Ядерный актин: ключевой игрок во внеклеточной коммуникации матрикс-ядро» . Коммуникативная и интегративная биология . 4 (5): 511–512. дои : 10.4161/cib.16256 . ПМК 3204115 . ПМИД 22046450 .
- ^ Jump up to: а б Чжао К., Ван В., Рандо О.Дж., Сюэ Ю., Свидерек К., Куо А., Крэбтри Г.Р. (ноябрь 1998 г.). «Быстрое и фосфоинозитол-зависимое связывание SWI/SNF-подобного комплекса BAF с хроматином после передачи сигнала рецептору Т-лимфоцитов» . Клетка . 95 (5): 625–636. дои : 10.1016/s0092-8674(00)81633-5 . ПМИД 9845365 . S2CID 3184211 .
- ^ Jump up to: а б Хофманн В.А., Стоилькович Л., Фуксова Б., Варгас Г.М., Мавромматис Е., Филимоненко В., Кисела К., Гудрич Дж.А., Лессард Дж.Л., Хоуп Т.Дж., Хозак П., де Ланероль П. (ноябрь 2004 г.). «Актин является частью преинициаторных комплексов и необходим для транскрипции РНК-полимеразой II». Природная клеточная биология . 6 (11): 1094–1101. дои : 10.1038/ncb1182 . ПМИД 15502823 . S2CID 23909479 .
- ^ Jump up to: а б Ху П., Ву С., Эрнандес Н. (декабрь 2004 г.). «Роль бета-актина в транскрипции РНК-полимеразы III» . Гены и развитие . 18 (24): 3010–3015. дои : 10.1101/gad.1250804 . ПМК 535912 . ПМИД 15574586 .
- ^ Jump up to: а б Филимоненко В.В., Чжао Дж., Ибен С., Дингова Х., Кисела К., Кале М., Зентграф Х., Хофманн В.А., де Ланероль П., Хозак П., Груммт I (декабрь 2004 г.). «Ядерный актин и миозин I необходимы для транскрипции РНК-полимеразы I». Природная клеточная биология . 6 (12): 1165–1172. дои : 10.1038/ncb1190 . ПМИД 15558034 . S2CID 6633625 .
- ^ Маральди Н.М., Латтанци Дж., Мармироли С., Скуарцони С., Манзоли Ф.А. (1 января 2004 г.). «Новая роль ламинов, белков ядерной оболочки и актина в ядре». Достижения в регуляции ферментов . 44 : 155–172. дои : 10.1016/j.advenzreg.2003.11.005 . ПМИД 15581488 .
- ^ Тонделейр Д, Ламбрехтс А, Мюллер М, Йонкхир В, Долл Т, Вандам Д, Баккали К, Уотершут Д, Лемейстр М, Дебейр О, Декестекер С, Хинц Б, Стас А, Тиммерман Е, Колерт Н, Геварт К, Вандекеркхове Дж , Ampe C (август 2012 г.). «Клетки, лишенные β-актина, генетически перепрограммируются и сохраняют условную миграционную способность» . Молекулярная и клеточная протеомика . 11 (8): 255–271. дои : 10.1074/mcp.M111.015099 . ПМК 3412960 . ПМИД 22448045 .
- ^ Холаска Дж. М., Ковальски А. К., Уилсон К. Л. (сентябрь 2004 г.). «Эмерин покрывает заостренный конец актиновых нитей: свидетельство наличия актиновой кортикальной сети на внутренней мембране ядра» . ПЛОС Биология . 2 (9): Е231. дои : 10.1371/journal.pbio.0020231 . ПМК 509406 . ПМИД 15328537 .
- ^ Пукельварц М., МакНелли Э.М. (1 января 2011 г.). «Мышечная дистрофия Эмери-Дрейфуса». Мышечные дистрофии . Справочник по клинической неврологии. Том. 101. С. 155–166. дои : 10.1016/B978-0-08-045031-5.00012-8 . ISBN 9780080450315 . ПМИД 21496632 .
- ^ Фарранц А.К. (июнь 2008 г.). «Ремоделирование хроматина и организация актина» . Письма ФЭБС . 582 (14): 2041–2050. Бибкод : 2008FEBSL.582.2041F . дои : 10.1016/j.febslet.2008.04.032 . ПМИД 18442483 . S2CID 23147656 .
- ^ Шёлиндер М., Бьорк П., Седерберг Э., Сабри Н., Фаррантс А.К., Visa N (август 2005 г.). «Растущая пре-мРНК рекрутирует факторы, модифицирующие актин и хроматин, к транскрипционно активным генам» . Гены и развитие . 19 (16): 1871–1884. дои : 10.1101/gad.339405 . ПМЦ 1186187 . ПМИД 16103215 .
- ^ Jump up to: а б Перципалле П, Виза N (март 2006 г.). «Молекулярные функции ядерного актина в транскрипции» . Журнал клеточной биологии . 172 (7): 967–971. дои : 10.1083/jcb.200512083 . ПМК 2063754 . ПМИД 16549500 .
- ^ Федорова Е, Цинк Д (ноябрь 2008 г.). «Ядерная архитектура и регуляция генов» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1783 (11): 2174–2184. дои : 10.1016/j.bbamcr.2008.07.018 . ПМИД 18718493 .
- ^ Скарп К.П., Вартиайнен М.К. (август 2010 г.). «Актин на ДНК — древние и динамичные взаимоотношения» . Цитоскелет . 67 (8): 487–495. дои : 10.1002/см.20464 . ПМИД 20593452 . S2CID 37763449 .
- ^ Олав И.А., Рек-Петерсон С.Л., Крэбтри Г.Р. (1 января 2002 г.). «Ядерный актин и родственные актину белки в ремоделировании хроматина». Ежегодный обзор биохимии . 71 : 755–781. doi : 10.1146/annurev.biochem.71.110601.135507 . ПМИД 12045110 .
- ^ Чжэн Б., Хан М., Бернье М., Вэнь Дж. К. (май 2009 г.). «Ядерный актин и актин-связывающие белки в регуляции транскрипции и экспрессии генов» . Журнал ФЭБС . 276 (10): 2669–2685. дои : 10.1111/j.1742-4658.2009.06986.x . ПМК 2978034 . ПМИД 19459931 .
- ^ Феррай С., Наум-Онганиа Г., Лонгобарди Э., Палаццоло М., Дисанса А., Диас В.М., Криппа М.П., Сцита Г., Блази Ф (август 2009 г.). «Индукция транскрипции HoxB ретиноевой кислотой требует полимеризации актина» . Молекулярная биология клетки . 20 (15): 3543–3551. дои : 10.1091/mbc.E09-02-0114 . ПМЦ 2719572 . ПМИД 19477923 .
- ^ Сюй Ю.З., Тюрайсингам Т., Мораис Д.А., Рола-Плещинский М., Радзиох Д. (март 2010 г.). «Ядерная транслокация бета-актина участвует в регуляции транскрипции во время дифференцировки макрофагов клеток HL-60» . Молекулярная биология клетки . 21 (5): 811–820. doi : 10.1091/mbc.E09-06-0534 . ПМЦ 2828967 . ПМИД 20053683 .
- ^ Jump up to: а б Миямото К., Паске В., Жюльен Дж., Гердон Дж.Б. (май 2011 г.). «Ядерная полимеризация актина необходима для транскрипционного перепрограммирования Oct4 ооцитами» . Гены и развитие . 25 (9): 946–958. дои : 10.1101/gad.615211 . ПМК 3084028 . ПМИД 21536734 .
- ^ Хуан В., Гислетти С., Сайджо К., Ганди М., Ауади М., Тес Г.Дж., Чжан Д.С., Яо Дж., член парламента Чехии, Гуд Б.Л., Розенфельд М.Г., Гласс С.К. (февраль 2011 г.). «Коронин 2А опосредует актин-зависимую дерепрессию генов воспалительного ответа» . Природа . 470 (7334): 414–418. Бибкод : 2011Natur.470..414H . дои : 10.1038/nature09703 . ПМЦ 3464905 . ПМИД 21331046 .
- ^ Миямото К., Гердон Дж.Б. (сентябрь 2011 г.). «Ядерный актин и активация транскрипции» . Коммуникативная и интегративная биология . 4 (5): 582–583. дои : 10.4161/cib.16491 . ПМК 3204135 . ПМИД 22046469 .
- ^ Чуанг Ч., Карпентер А.Е., Фуксова Б., Джонсон Т., де Ланеролль П., Бельмонт А.С. (апрель 2006 г.). «Дальнее направленное движение интерфазного участка хромосомы» . Современная биология . 16 (8): 825–831. Бибкод : 2006CBio...16..825C . дои : 10.1016/j.cub.2006.03.059 . ПМИД 16631592 . S2CID 1191289 .
- ^ Хофманн В.А., Варгас Г.М., Рамчандран Р., Стоилькович Л., Гудрич Дж.А., де Ланероль П. (ноябрь 2006 г.). «Ядерный миозин I необходим для образования первой фосфодиэфирной связи во время инициации транскрипции РНК-полимеразой II». Журнал клеточной биохимии . 99 (4): 1001–1009. дои : 10.1002/jcb.21035 . ПМИД 16960872 . S2CID 39237955 .
- ^ Олсон Э.Н., Нордхайм А (май 2010 г.). «Связь динамики актина и транскрипции генов для управления подвижными функциями клеток» . Nature Reviews Молекулярно-клеточная биология . 11 (5): 353–365. дои : 10.1038/nrm2890 . ПМК 3073350 . ПМИД 20414257 .
- ^ Мираллес Ф., Позерн Г., Заромитиду А.И., Трейсман Р. (май 2003 г.). «Динамика актина контролирует активность SRF путем регулирования его коактиватора MAL». Клетка . 113 (3): 329–342. CiteSeerX 10.1.1.327.7451 . дои : 10.1016/s0092-8674(03)00278-2 . ПМИД 12732141 . S2CID 17209744 .
- ^ Вартиайнен МК (июнь 2008 г.). «Ядерная динамика актина - от формы к функции». Письма ФЭБС . 582 (14): 2033–2040. Бибкод : 2008FEBSL.582.2033V . дои : 10.1016/j.febslet.2008.04.010 . ПМИД 18423404 . S2CID 35474838 .
- ^ Кнёлль Б. (июнь 2010 г.). «Актин-опосредованная экспрессия генов в нейронах: связь MRTF-SRF». Биологическая химия . 391 (6): 591–597. дои : 10.1515/BC.2010.061 . ПМИД 20370316 . S2CID 36373214 .
- ^ Цопулидис Н., Кау С., Лакета В., Кутшейдт С., Баарлинк С., Столп Б., Гросс Р., Факлер О.Т. (январь 2019 г.). «Формирование сети ядерного актина, запускаемое рецептором Т-клеток, управляет эффекторными функциями CD4+ Т-клеток» . Наука Иммунология . 4 (31): eaav1987. doi : 10.1126/sciimmunol.aav1987 . ПМИД 30610013 .
- ^ Jump up to: а б Хуан Ю, Чжан С, Пак Цзи. Динамика ядерного актина в экспрессии генов, восстановлении ДНК и раке. Результаты Пробл Ячейка Разн. 2022;70:625-663. дои: 10.1007/978-3-031-06573-6_23. ПМИД 36348125 ; PMCID: PMC9677682
- ^ Пал, Диман Санкар; Банерджи, Татсат; Линь, Иян; де Трогов, Феликс; Борлейс, Джейн; Церкви, Пол А.; Девреотес, Питер Н. (июль 2023 г.). «Активация отдельных нижестоящих узлов в сети факторов роста управляет миграцией иммунных клеток» . Развивающая клетка . 58 (13): 1170–1188.e7. дои : 10.1016/j.devcel.2023.04.019 . ISSN 1534-5807 . ПМЦ 10524337 . ПМИД 37220748 .
- ^ Линь, Иян; Пал, Диман Санкар; Банерджи, Париджат; Банерджи, Татсат; Цинь, Гуанхуэй; Дэн, Ю; Борлейс, Джейн; Церкви, Пол А.; Девреотес, Питер Н. (1 июля 2024 г.). «Подавление Ras усиливает поляризацию и миграцию клеток, обусловленную сократимостью заднего актомиозина» . Природная клеточная биология : 1–15. дои : 10.1038/s41556-024-01453-4 . ISSN 1476-4679 . ПМИД 38951708 .
- ^ Jump up to: а б с д Лодиш и др. 2016 , с. 811–812.
- ^ Лодиш и др. 2016 , с. 796.
- ^ Jump up to: а б с д и Лодиш и др. 2016 , с. 803–805.
- ^ Лодиш и др. 2016 , с. 805–806.
- ^ Jump up to: а б Лодиш и др. 2016 , с. 807.
- ^ Фудзивара К., Портер М.Э., Поллард Т.Д. (октябрь 1978 г.). «Локализация альфа-актинина в борозде расщепления во время цитокинеза» . Журнал клеточной биологии . 79 (1): 268–275. дои : 10.1083/jcb.79.1.268 . ПМК 2110217 . ПМИД 359574 .
- ^ Пелхэм Р.Дж., Чанг Ф. (сентябрь 2002 г.). «Динамика актина в сократительном кольце во время цитокинеза у делящихся дрожжей». Природа . 419 (6902): 82–86. Бибкод : 2002Natur.419...82P . дои : 10.1038/nature00999 . ПМИД 12214236 . S2CID 4389564 .
- ^ Лодиш и др. 2016 , с. 809.
- ^ Машима Т., Наито М., Ногучи К., Миллер Д.К., Николсон Д.В., Цуруо Т. (март 1997 г.). «Расщепление актина с помощью CPP-32/апопаина во время развития апоптоза» . Онкоген . 14 (9): 1007–1012. дои : 10.1038/sj.onc.1200919 . ПМИД 9070648 .
- ^ Ван К.К. (январь 2000 г.). «Кальпаин и каспаза: можете ли вы отличить?». Тенденции в нейронауках . 23 (1): 20–26. дои : 10.1016/S0166-2236(99)01479-4 . ПМИД 10631785 . S2CID 17571984 .
- ^ Вилла П.Г., Хензель В.Дж., Сенсенбреннер М., Хендерсон К.Э., Петтманн Б. (март 1998 г.). «Ингибиторы кальпаина, но не ингибиторы каспаз, предотвращают протеолиз актина и фрагментацию ДНК во время апоптоза». Журнал клеточной науки . 111 (Часть 6): 713–722. дои : 10.1242/jcs.111.6.713 . ПМИД 9472000 .
- ^ Юот Дж., Уль Ф., Руссо С., Дешес Р.Г., Шах Г.М., Лэндри Дж. (ноябрь 1998 г.). «SAPK2/p38-зависимая реорганизация F-актина регулирует раннее пузырение мембран во время апоптоза, вызванного стрессом» . Журнал клеточной биологии . 143 (5): 1361–1373. дои : 10.1083/jcb.143.5.1361 . ПМК 2133090 . ПМИД 9832563 .
- ^ Адамс К.Л., Нельсон В.Дж., Смит С.Дж. (декабрь 1996 г.). «Количественный анализ реорганизации кадгерин-катенин-актин при развитии межклеточной адгезии» . Журнал клеточной биологии . 135 (6, ч. 2): 1899–1911. дои : 10.1083/jcb.135.6.1899 . ПМК 2133977 . ПМИД 8991100 .
- ^ Витке В., Шлейхер М., Ногель А.А. (январь 1992 г.). «Избыточность в системе микрофиламентов: аномальное развитие клеток Dictyostelium, в которых отсутствуют два белка, сшивающего F-актин». Клетка . 68 (1): 53–62. дои : 10.1016/0092-8674(92)90205-Q . ПМИД 1732064 . S2CID 37569656 .
- ^ Фернандес-Валле С., Горман Д., Гомес А.М., Бунге М.Б. (январь 1997 г.). «Актин играет роль как в изменениях формы клеток, так и в экспрессии генов, связанных с миелинизацией шванновских клеток» . Журнал неврологии . 17 (1): 241–250. doi : 10.1523/JNEUROSCI.17-01-00241.1997 . ПМК 6793673 . ПМИД 8987752 .
- ^ Волыняк М.Ю., Сундстрем П. (октябрь 2007 г.). «Роль динамики актинового цитоскелета в активации пути циклического АМФ и экспрессии гена HWP1 у Candida albicans» . Эукариотическая клетка . 6 (10): 1824–1840. дои : 10.1128/EC.00188-07 . ПМК 2043390 . ПМИД 17715368 .
- ^ Танака Х., Игучи Н., Эгидио де Карвалью С., Тадокоро Ю., Йомогада К., Нишимунэ Ю. (август 2003 г.). «Новые актиноподобные белки T-ACTIN 1 и T-ACTIN 2 по-разному экспрессируются в цитоплазме и ядре гаплоидных зародышевых клеток мыши» . Биология размножения . 69 (2): 475–482. дои : 10.1095/biolreprod.103.015867 . ПМИД 12672658 .
- ^ Цзян Ю., DJ Stillman (март 1996 г.). «Эпигенетические эффекты на транскрипцию дрожжей, вызванные мутациями актин-родственного белка, присутствующего в ядре» . Гены и развитие . 10 (5): 604–619. дои : 10.1101/gad.10.5.604 . ПМИД 8598290 .
- ^ Manor U, Качар Б (декабрь 2008 г.). «Динамическая регуляция длины сенсорных стереоцилий» . Семинары по клеточной биологии и биологии развития . 19 (6): 502–510. дои : 10.1016/j.semcdb.2008.07.006 . ПМК 2650238 . ПМИД 18692583 .
- ^ Рзадзинска А.К., Шнайдер М.Е., Дэвис С., Риордан Г.П., Качар Б. (март 2004 г.). «Молекулярная беговая дорожка актина и миозины поддерживают функциональную архитектуру стереоцилий и их самообновление» . Журнал клеточной биологии . 164 (6): 887–897. дои : 10.1083/jcb.200310055 . ПМК 2172292 . ПМИД 15024034 .
- ^ Сюй Дж., Ван Кеймюлен А., Вакида Н.М., Карлтон П., Бернс М.В., Борн Х.Р. (май 2007 г.). «Полярность обнаруживает внутреннюю хиральность клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (22): 9296–9300. Бибкод : 2007PNAS..104.9296X . дои : 10.1073/pnas.0703153104 . ПМК 1890488 . ПМИД 17517645 .
- ^ Тамада А., Кавасе С., Мураками Ф., Камигути Х. (февраль 2010 г.). «Автономное правовинтовое вращение филоподий конуса роста приводит к повороту нейритов» . Журнал клеточной биологии . 188 (3): 429–441. дои : 10.1083/jcb.200906043 . ПМЦ 2819689 . ПМИД 20123994 .
- ^ Ван Л.К., Рональдсон К., Парк М., Тейлор Дж., Чжан Ю., Гимбл Дж.М., Вуняк-Новакович Дж. (июль 2011 г.). «Клетки млекопитающих с микроструктурой демонстрируют фенотипическую лево-правую асимметрию» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (30): 12295–12300. Бибкод : 2011PNAS..10812295W . дои : 10.1073/pnas.1103834108 . ПМЦ 3145729 . ПМИД 21709270 .
- ^ Jump up to: а б с д и Лодиш и др. 2016 , с. 779.
- ^ Jump up to: а б с Эльзинга М., Коллинз Дж. Х., Кюль В. М., Адельштейн Р. С. (сентябрь 1973 г.). «Полная аминокислотная последовательность актина скелетных мышц кролика» . Труды Национальной академии наук Соединенных Штатов Америки . 70 (9): 2687–2691. Бибкод : 1973PNAS...70.2687E . дои : 10.1073/pnas.70.9.2687 . ПМК 427084 . ПМИД 4517681 .
- ^ Jump up to: а б Коллинз Дж. Х., Эльзинга М. (август 1975 г.). «Первичная структура актина из скелетных мышц кролика. Пополнение и анализ аминокислотной последовательности» . Журнал биологической химии . 250 (15): 5915–5920. дои : 10.1016/S0021-9258(19)41139-3 . ПМИД 1150665 .
- ^ Jump up to: а б с д и ж г Дос Ремедиос К.Г., Чабра Д. (2008). Актин-связывающие белки и болезни . Спрингер. ISBN 978-0-387-71747-0 .
- ^ Jump up to: а б Кабш В., Мангерц Х.Г., Сак Д., Пай Э.Ф., Холмс К.К. (сентябрь 1990 г.). «Атомная структура комплекса актин: ДНКаза I». Природа . 347 (6288): 37–44. Бибкод : 1990Natur.347...37K . дои : 10.1038/347037a0 . ПМИД 2395459 . S2CID 925337 .
- ^ Роулд М.А., Ван К., Джоэл П.Б., Лоуи С., Трибус К.М. (октябрь 2006 г.). «Кристаллические структуры экспрессируемого неполимеризующегося мономерного актина в состояниях АДФ и АТФ» . Журнал биологической химии . 281 (42): 31909–31919. дои : 10.1074/jbc.M601973200 . ПМИД 16920713 .
- ^ Jump up to: а б Холмс К.К., Попп Д., Гебхард В., Кабш В. (сентябрь 1990 г.). «Атомная модель актиновой нити». Природа . 347 (6288): 44–49. Бибкод : 1990Natur.347...44H . дои : 10.1038/347044a0 . ПМИД 2395461 . S2CID 4317981 .
- ^ Jump up to: а б с Лодиш и др. 2016 , с. 780.
- ^ Jump up to: а б с Рейслер Э., Эгельман Э.Х. (декабрь 2007 г.). «Структура и функция актина: то, что мы до сих пор не понимаем» . Журнал биологической химии . 282 (50): 36133–36137. дои : 10.1074/jbc.R700030200 . ПМИД 17965017 .
- ^ Jump up to: а б с д Ода Т., Иваса М., Айхара Т., Маэда Ю., Нарита А. (январь 2009 г.). «Природа перехода глобулярного актина в фиброзный». Природа . 457 (7228): 441–445. Бибкод : 2009Natur.457..441O . дои : 10.1038/nature07685 . ПМИД 19158791 . S2CID 4317892 .
- ^ Бегг Д.А., Родевальд Р., Ребхун Л.И. (декабрь 1978 г.). «Визуализация полярности актиновых нитей на тонких срезах. Доказательства однородной полярности мембран-ассоциированных нитей» . Журнал клеточной биологии . 79 (3): 846–852. дои : 10.1083/jcb.79.3.846 . ПМК 2110270 . ПМИД 569662 .
- ^ Генезер Ф (1981). Гистология . Мунксгаард. п. 105. ИСБН 978-87-16-08418-7 .
- ^ Jump up to: а б с фон дер Экен Дж., Мюллер М., Леман В., Манштейн Д.Д., Пенчек П.А., Раунсер С. (май 2015 г.). «Строение комплекса F-актин-тропомиозин» . Природа . 519 (7541): 114–117. Бибкод : 2015Природа.519..114В . дои : 10.1038/nature14033 . ПМЦ 4477711 . ПМИД 25470062 .
- ^ Холл Дж. Э., Гайтон AC (2006). Учебник медицинской физиологии . Сент-Луис, Миссури: Эльзевир Сондерс. п. 76 . ISBN 978-0-7216-0240-0 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П., ред. (2002). «Самосборка и динамическая структура цитоскелетных нитей» . Молекулярная биология клетки (4-е изд.). Гирляндная наука. ISBN 978-0-8153-3218-3 .
- ^ Jump up to: а б Саймонс К.Т., Стас А., Роммелер Х., Ампе С., Льюис С.А., Коуэн, штат Нью-Джерси (февраль 2004 г.). «Избирательный вклад субъединиц префолдина эукариот в связывание актина и тубулина» . Журнал биологической химии . 279 (6): 4196–4203. дои : 10.1074/jbc.M306053200 . ПМИД 14634002 .
- ^ Jump up to: а б Мартин-Бенито Дж., Боскович Дж., Гомес-Пуэртас П., Карраскоса Х.Л., Саймонс К.Т., Льюис С.А., Бартолини Ф., Коуэн Н.Дж., Вальпуэста Х.М. (декабрь 2002 г.). «Структура эукариотического префолдина и его комплексов с развернутым актином и цитозольным шаперонином CCT» . Журнал ЭМБО . 21 (23): 6377–6386. дои : 10.1093/emboj/cdf640 . ПМК 136944 . ПМИД 12456645 .
- ^ Jump up to: а б Вандам Д., Ламберт Э., Уотершут Д., Коньяр К., Вандекерхове Дж., Ампе К., Константин Б., Роммелер Х. (июль 2009 г.). «Мутанты немалиновой миопатии альфа-скелетных мышц вызывают гибель клеток в культивируемых мышечных клетках» (PDF) . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1793 (7): 1259–1271. дои : 10.1016/j.bbamcr.2009.04.004 . ПМИД 19393268 .
- ^ Jump up to: а б Брэкли К.И., Грэнтэм Дж. (январь 2009 г.). «Активность шаперонина, содержащего TCP-1 (CCT): значение для развития клеточного цикла и организации цитоскелета» . Клеточные стрессы и шапероны . 14 (1): 23–31. дои : 10.1007/s12192-008-0057-x . ПМЦ 2673901 . ПМИД 18595008 .
- ^ Jump up to: а б Стирлинг ПК, Куэльяр Дж., Альфаро Г.А., Эль Хадали Ф., Бех К.Т., Вальпуэста Х.М., Мелки Р., Леру М.Р. (март 2006 г.). «PhLP3 модулирует опосредованное CCT сворачивание актина и тубулина посредством тройных комплексов с субстратами» . Журнал биологической химии . 281 (11): 7012–7021. дои : 10.1074/jbc.M513235200 . ПМИД 16415341 .
- ^ Хансен В.Дж., Коуэн, Нью-Джерси, Уэлч В.Дж. (апрель 1999 г.). «Комплексы префолдин-возникающая цепь при сворачивании белков цитоскелета» . Журнал клеточной биологии . 145 (2): 265–277. дои : 10.1083/jcb.145.2.265 . ПМК 2133115 . ПМИД 10209023 .
- ^ Мартин-Бенито Дж., Грэнтэм Дж., Боскович Дж., Брэкли К.И., Карраскоса Дж.Л., Уиллисон К.Р., Вальпуэста Дж.М. (март 2007 г.). «Межкольцевое расположение цитозольного шаперонина CCT» . Отчеты ЭМБО . 8 (3): 252–257. дои : 10.1038/sj.embor.7400894 . ПМК 1808031 . ПМИД 17304242 .
- ^ Нейринк К., Уотершут Д., Вандекерхове Дж., Ампе К., Роммелер Х. (январь 2006 г.). «Актин взаимодействует с CCT через дискретные сайты связывания: модель перехода-высвобождения связывания для сворачивания актина, опосредованного CCT». Журнал молекулярной биологии . 355 (1): 124–138. дои : 10.1016/j.jmb.2005.10.051 . ПМИД 16300788 .
- ^ Вавилонис Д., Ян К., О'Шонесси Б. (июнь 2005 г.). «Кинетика полимеризации актина, структура кепки и колебания» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (24): 8543–8548. arXiv : q-bio/0404004 . Бибкод : 2005PNAS..102.8543V . дои : 10.1073/pnas.0501435102 . ПМЦ 1150824 . ПМИД 15939882 .
- ^ Каткар Х.Х., Давтян А., Дурумерик А.Е., Хокки ГМ, Шрамм А., Энрике М., Вот Г.А. (сентябрь 2018 г.). «Понимание кооперативной природы гидролиза АТФ в актиновых нитях» . Биофизический журнал . 115 (8): 1589–1602. Бибкод : 2018BpJ...115.1589K . дои : 10.1016/j.bpj.2018.08.034 . ПМК 6260209 . ПМИД 30249402 .
- ^ МакКаллах М., Сондерс М.Г., Вот Г.А. (сентябрь 2014 г.). «Разгадка тайны гидролиза АТФ в актиновых нитях» . Журнал Американского химического общества . 136 (37): 13053–13058. дои : 10.1021/ja507169f . ПМК 4183606 . ПМИД 25181471 .
- ^ Сондерс М.Г., Вот Г.А. (октябрь 2011 г.). «Молекулы воды в нуклеотидсвязывающей щели актина: влияние на конформацию субъединицы и значение для гидролиза АТФ». Журнал молекулярной биологии . 413 (1): 279–291. дои : 10.1016/j.jmb.2011.07.068 . ПМИД 21856312 .
- ^ Jump up to: а б Лодиш и др. 2016 , с. 781.
- ^ Jump up to: а б Лодиш и др. 2016 , с. 783.
- ^ Jump up to: а б Лодиш и др. 2016 , с. 784.
- ^ Клеточная биология (на испанском языке). Эльзевир Испания. 2002. с. 132. ИСБН 978-84-458-1105-4 .
- ^ Домингес Р. (ноябрь 2004 г.). «Актин-связывающие белки - объединяющая гипотеза». Тенденции биохимических наук . 29 (11): 572–578. дои : 10.1016/j.tibs.2004.09.004 . ПМИД 15501675 .
- ^ Робинсон Р.К., Турбедски К., Кайзер Д.А., Маршан Дж.Б., Хиггс Х.Н., Чоу С., Поллард Т.Д. (ноябрь 2001 г.). «Кристаллическая структура комплекса Arp2/3». Наука . 294 (5547): 1679–1684. Бибкод : 2001Sci...294.1679R . дои : 10.1126/science.1066333 . ПМИД 11721045 . S2CID 18088124 .
- ^ Jump up to: а б с д Лодиш и др. 2016 , с. 786.
- ^ Лодиш и др. 2016 , с. 787–788.
- ^ ВВП : 2BTF ; Шутт С.Э., Мыслик Дж.К., Розицки, доктор медицинских наук, Гунесекере, Северная Каролина, Линдберг У (октябрь 1993 г.). «Строение кристаллического профилина-бета-актина». Природа . 365 (6449): 810–816. Бибкод : 1993Natur.365..810S . дои : 10.1038/365810a0 . ПМИД 8413665 . S2CID 4359724 .
- ^ Jump up to: а б с д Лодиш и др. 2016 , с. 785.
- ^ Лодиш и др. 2016 , стр. 784–785.
- ^ Лодиш и др. 2016 , стр. 785, 788.
- ^ Лодиш и др. 2016 , стр. 788–789.
- ^ Вандекерхове Дж., Вебер К. (декабрь 1978 г.). «По крайней мере шесть различных актинов экспрессируются у высших млекопитающих: анализ, основанный на аминокислотной последовательности аминоконцевого триптического пептида». Журнал молекулярной биологии . 126 (4): 783–802. дои : 10.1016/0022-2836(78)90020-7 . ПМИД 745245 .
- ^ Хаитлина С.Ю. (2001). Функциональная специфичность изоформ актина . Международный обзор цитологии. Том. 202. С. 35–98. дои : 10.1016/S0074-7696(01)02003-4 . ISBN 9780123646064 . ПМИД 11061563 .
- ^ Jump up to: а б с д и Поллард 2016 , «Гены, сохранение последовательности, распространение и численность».
- ^ Jump up to: а б Мива Т., Манабе Ю., Курокава К., Камада С., Канда Н., Брунс Г., Уэяма Х., Какунага Т. (июнь 1991 г.). «Структура, расположение хромосом и экспрессия гена гамма-актина гладкой мускулатуры человека (энтерального типа): эволюция шести генов актина человека» . Молекулярная и клеточная биология . 11 (6): 3296–3306. дои : 10.1128/mcb.11.6.3296 . ПМК 360182 . ПМИД 1710027 .
- ^ Эриксон HP (июль 2007 г.). «Эволюция цитоскелета» . Биоэссе . 29 (7): 668–677. doi : 10.1002/bies.20601 . ПМЦ 2630885 . ПМИД 17563102 .
- ^ Гардинер Дж., МакГи П., Общий Р., Марк Дж. (2008). «Являются ли гистоны, тубулин и актин производными от общего предкового белка?». Протоплазма . 233 (1–2): 1–5. дои : 10.1007/s00709-008-0305-z . ПМИД 18615236 . S2CID 21765920 .
- ^ Галлетта Б.Дж., Купер Дж.А. (февраль 2009 г.). «Актин и эндоцитоз: механизмы и филогения» . Современное мнение в области клеточной биологии . 21 (1): 20–27. дои : 10.1016/j.ceb.2009.01.006 . ПМК 2670849 . ПМИД 19186047 .
- ^ Маллинз Р.Д., Поллард Т.Д. (апрель 1999 г.). «Структура и функции комплекса Arp2/3». Современное мнение в области структурной биологии . 9 (2): 244–249. дои : 10.1016/S0959-440X(99)80034-7 . ПМИД 10322212 .
- ^ Попп Д., Нарита А., Маэда К., Фудзисава Т., Гошдастидер У., Иваса М., Маэда Ю., Робинсон Р.К. (май 2010 г.). «Структура, организация и динамика нитей в листах MreB» . Журнал биологической химии . 285 (21): 15858–15865. дои : 10.1074/jbc.M109.095901 . ПМЦ 2871453 . ПМИД 20223832 .
- ^ ван ден Энт Ф., Амос Л.А., Лёве Дж. (сентябрь 2001 г.). «Прокариотическое происхождение актинового цитоскелета». Природа . 413 (6851): 39–44. Бибкод : 2001Natur.413...39V . дои : 10.1038/35092500 . ПМИД 11544518 . S2CID 4427828 .
- ^ Карбаллидо-Лопес Р. (декабрь 2006 г.). «Бактериальный актиноподобный цитоскелет» . Обзоры микробиологии и молекулярной биологии . 70 (4): 888–909. дои : 10.1128/MMBR.00014-06 . ПМК 1698507 . ПМИД 17158703 .
- ^ Попп Д., Сюй В., Нарита А., Бржоска А.Дж., Скуррей Р.А., Ферт Н., Гошдастидер Ю., Гошдастидер Ю., Маэда Ю., Робинсон Р.К., Шумахер М.А. (март 2010 г.). «Структура и динамика филаментов актиноподобного белка ParM pSK41: значение для сегрегации плазмидной ДНК» . Журнал биологической химии . 285 (13): 10130–10140. дои : 10.1074/jbc.M109.071613 . ПМЦ 2843175 . ПМИД 20106979 .
- ^ Попп Д., Нарита А., Гошдастидер У., Маэда К., Маэда Ю., Ода Т., Фудзисава Т., Ониши Х., Ито К., Робинсон Р.К. (апрель 2010 г.). «Полимерные структуры и динамические свойства бактериального актина AlfA». Журнал молекулярной биологии . 397 (4): 1031–1041. дои : 10.1016/j.jmb.2010.02.010 . ПМИД 20156449 .
- ^ Попп Д., Нарита А., Ли Л.Дж., Гошдастидер У., Сюэ Б., Шринивасан Р., Баласубраманиан М.К., Танака Т., Робинсон Р.К. (июнь 2012 г.). «Новая актиноподобная структура нитей Clostridium tetani» . Журнал биологической химии . 287 (25): 21121–21129. дои : 10.1074/jbc.M112.341016 . ПМЦ 3375535 . ПМИД 22514279 .
- ^ Хара Ф., Ямасиро К., Немото Н., Охта Ю., Ёкобори С., Ясунага Т., Хисанага С., Ямагиши А. (март 2007 г.). «Гомолог актина археи Thermoplasma acidophilum, сохраняющий древние характеристики эукариотического актина» . Журнал бактериологии . 189 (5): 2039–2045. дои : 10.1128/JB.01454-06 . ПМЦ 1855749 . ПМИД 17189356 .
- ^ Су А.И., Уилтшир Т., Баталов С., Лапп Х., Чинг К.А., Блок Д, Чжан Дж., Соден Р., Хаякава М., Крейман Г., Кук М.П., Уокер Дж.Р., Хогенеш Дж.Б. (апрель 2004 г.). «Атлас генов транскриптомов, кодирующих белки мыши и человека» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (16): 6062–6067. Бибкод : 2004PNAS..101.6062S . дои : 10.1073/pnas.0400782101 . ПМЦ 395923 . ПМИД 15075390 .
- ^ Jump up to: а б "ACTS_HUMAN" . Р68133 . Консорциум ЮниПрот. Архивировано из оригинала 5 ноября 2012 г. Проверено 21 января 2013 г.
- ^ Jump up to: а б Бате Ф.С., Роммелер Х., Мачески Л.М. (2007). «Фенотипы мутантов актина, связанных с миопатией, в дифференцированных мышечных трубках C2C12» . Клеточная биология BMC . 8 (1): 2. дои : 10.1186/1471-2121-8-2 . ПМЦ 1779783 . ПМИД 17227580 .
- ^ Кайндл А.М., Рюшендорф Ф., Краузе С., Гебель Х.Х., Келер К., Беккер С., Понгратц Д., Мюллер-Хёккер Дж., Нюрнберг П., Столтенбург-Дидингер Г., Лохмюллер Х., Хюбнер А. (ноябрь 2004 г.). «Миссенс-мутации ACTA1 вызывают доминантную врожденную миопатию с ядрами» . Журнал медицинской генетики . 41 (11): 842–848. дои : 10.1136/jmg.2004.020271 . ПМЦ 1735626 . ПМИД 15520409 .
- ^ Воробей Дж.К., Новак К.Дж., Дурлинг Х.Дж., Беггс А.Х., Валлгрен-Петтерссон С., Ромеро Н., Нонака И., Лэнг Н.Г. (сентябрь 2003 г.). «Мышечные заболевания, вызванные мутациями в гене альфа-актина скелетных мышц (ACTA1)». Нервно-мышечные расстройства . 13 (7–8): 519–531. дои : 10.1016/S0960-8966(03)00101-9 . ПМИД 12921789 . S2CID 20716 .
- ^ Норт К., Райан М.М. (2002). «Немалиновая миопатия» . В Пагон Р.А., Берд Т.Д., Долан Ч.Р., Стивенс К., Адам М.П. (ред.). GeneReviews [Интернет] . Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл. ПМИД 20301465 . Архивировано из оригинала 18 января 2017 г.
- ^ Ильковски Б., Новак К.Дж., Домазетовска А., Максвелл А.Л., Клемент С., Дэвис К.Э., Лэнг Н.Г., Норт К.Н., Купер С.Т. (август 2004 г.). «Доказательства доминантно-негативного эффекта при немалиновой миопатии ACTA1, вызванной аномальным сворачиванием, агрегацией и измененной полимеризацией мутантных изоформ актина» . Молекулярная генетика человека . 13 (16): 1727–1743. дои : 10.1093/hmg/ddh185 . ПМИД 15198992 .
- ^ Кларк Н.Ф., Илковски Б., Купер С., Валова В.А., Робинсон П.Дж., Нонака И., Фэн Дж.Дж., Марстон С., Норт К. (июнь 2007 г.). «Патогенез врожденной диспропорции типов волокон, связанной с ACTA1». Анналы неврологии . 61 (6): 552–561. дои : 10.1002/ana.21112 . ПМИД 17387733 . S2CID 11746835 .
- ^ Уотсон М.Б., Линд М.Дж., Смит Л., Дрю П.Дж., Коуквелл Л. (2007). «Анализ экспрессионного микрочипа выявляет гены, связанные с устойчивостью in vitro к цисплатину на модели клеточной линии» . Акта Онкологика . 46 (5): 651–658. дои : 10.1080/02841860601156157 . ПМИД 17562441 . S2CID 7163200 .
- ^ Го Д.С., Панну Х., Тран-Фадулу В., Папке С.Л., Ю.РК., Авидан Н., Буржуа С., Эстрера А.Л., Сафи Х.Дж., Спаркс Е., Амор Д., Адес Л., МакКоннелл В., Уиллоби С.Э., Абуэло Д., Уиллинг М., Льюис Р.А., Ким Д.Х., Шерер С., Тунг П.П., Ан С., Буджа Л.М., Раман К.С., Шете С.С., Милевич Д.М. (декабрь 2007 г.). «Мутации в гладкомышечном альфа-актине (ACTA2) приводят к аневризмам и расслоениям грудной аорты». Природная генетика . 39 (12): 1488–1493. дои : 10.1038/ng.2007.6 . ПМИД 17994018 . S2CID 62785801 .
- ^ Го Д.С., Папке С.Л., Тран-Фадулу В., Регаладо Э.С., Авидан Н., Джонсон Р.Дж., Ким Д.Х., Панну Х., Уиллинг М.С., Спаркс Э., Пьеритц Р.Э., Сингх М.Н., Далман Р.Л., Гротта Дж.К., Мэриан Эй.Дж., Бурвинкль Э.А., Фрейзер Л.К., ЛеМэр С.А., Коселли Дж.С., Эстрера А.Л., Сафи Х.Дж., Вирарагаван С., Музный Д.М., Уилер Д.А., Уиллерсон Дж.Т., Ю.Р.К., Шете С.С., Шерер С.Е., Раман К.С., Бужа Л.М., Милевич Д.М. (май 2009 г.). «Мутации в гладкомышечном альфа-актине (ACTA2) вызывают ишемическую болезнь сердца, инсульт и болезнь Моямоя, а также заболевание грудной аорты» . Американский журнал генетики человека . 84 (5): 617–627. дои : 10.1016/j.ajhg.2009.04.007 . ПМК 2680995 . ПМИД 19409525 .
- ^ Акполат Н., Яхси С., Годекмердан А., Ялниз М., Демирбаг К. (сентябрь 2005 г.). «Значение альфа-СМА в оценке тяжести фиброза печени при инфекции гепатита В и развитии цирроза: гистопатологическое и иммуногистохимическое исследование». Гистопатология . 47 (3): 276–280. дои : 10.1111/j.1365-2559.2005.02226.x . ПМИД 16115228 . S2CID 23800095 .
- ^ Хамада Х., Петрино М.Г., Какунага Т. (октябрь 1982 г.). «Молекулярная структура и эволюционное происхождение гена актина сердечной мышцы человека» . Труды Национальной академии наук Соединенных Штатов Америки . 79 (19): 5901–5905. Бибкод : 1982PNAS...79.5901H . дои : 10.1073/pnas.79.19.5901 . ПМК 347018 . ПМИД 6310553 .
- ^ Jump up to: а б Олсон Т.М., Михелс В.В., Тибодо С.Н., Тай Ю.С., Китинг М.Т. (май 1998 г.). «Мутации актина при дилатационной кардиомиопатии, наследственной форме сердечной недостаточности». Наука . 280 (5364): 750–752. Бибкод : 1998Sci...280..750O . дои : 10.1126/science.280.5364.750 . ПМИД 9563954 . S2CID 26971894 .
- ^ Ся XG, Чжоу Х, Сампер Э, Мелов С, Сюй З (январь 2006 г.). «Экспрессируемая Pol II shRNA подавляет экспрессию гена Sod2 и вызывает фенотипический нокаут гена у мышей» . ПЛОС Генетика . 2 (1): е10. дои : 10.1371/journal.pgen.0020010 . ПМЦ 1358942 . ПМИД 16450009 .
- ^ Интернет-менделевское наследование у человека (OMIM): Актин, альфа, сердечная мышца; ACTC1 - 102540
- ^ Матссон Х, Исон Дж, Буквалтер К.С., Клар Дж., Густавссон П., Суннегард Дж., Энелл Х., Джонсон А., Виккула М., Гутьеррес И., Гранадос-Риверон Дж., Поуп М., Бу'Лок Ф., Кокс Дж., Робинсон Т.Э., Сонг Ф., Брук DJ, Марстон С., Трибус К.М., Даль Н. (январь 2008 г.). «Мутации альфа-сердечного актина вызывают дефекты межпредсердной перегородки» . Молекулярная генетика человека . 17 (2): 256–265. дои : 10.1093/hmg/ddm302 . ПМИД 17947298 .
- ^ Девлин Т.М. (2006). Биохимия . Барселона: Реверте. ISBN 978-84-291-7208-9 .
- ^ Кабаева З. (11 ноября 2002 г.). Генетический анализ при гипертрофической кардиомиопатии (Диссертация). дои : 10.18452/14800 .
- ^ Олсон Т.М., Доан Т.П., Кишимото Нью-Йорк, Уитби Ф.Г., Акерман М.Дж., Фананапазир Л. (сентябрь 2000 г.). «Наследственные и de novo мутации в гене сердечного актина вызывают гипертрофическую кардиомиопатию». Журнал молекулярной и клеточной кардиологии . 32 (9): 1687–1694. дои : 10.1006/jmcc.2000.1204 . ПМИД 10966831 .
- ^ Рамирес CD, Падрон Р. (2004). «Семейная гипертрофическая кардиомиопатия: гены, мутации и модели на животных. Обзор» [Семейная гипертрофическая кардиомиопатия: гены, мутации и модели на животных. просмотреть]. Клинические исследования (на испанском языке). 45 (1): 69–100.
- ^ Каски Дж.П., Сиррис П., Берч М., Томе-Эстебан М.Т., Фентон М., Кристиансен М., Андерсен П.С., Себир Н., Эшворт М., Динфилд Дж.Э., Маккенна У.Дж., Эллиотт П.М. (ноябрь 2008 г.). «Идиопатическая рестриктивная кардиомиопатия у детей вызвана мутациями в генах белков сердечного саркомера». Сердце . 94 (11): 1478–1484. дои : 10.1136/hrt.2007.134684 . ПМИД 18467357 . S2CID 44257334 .
- ^ Пиготт Т.Дж., Джефферсон Д. (1991). «Идиопатический паралич общего малоберцового нерва - обзор тринадцати случаев». Британский журнал нейрохирургии . 5 (1): 7–11. дои : 10.3109/02688699108998440 . ПМИД 1850600 .
- ^ «Ген: ACTB» . ЭйсВью . Национальный центр биотехнологической информации США (NCBI). Архивировано из оригинала 18 июня 2013 г. Проверено 21 января 2013 г.
- ^ Чанг К.В., Ян П.И., Лай ХИ, Йе ТС, Чен ТС, Йе КТ (сентябрь 2006 г.). «Идентификация новой изоформы актина при гепатоцеллюлярной карциноме». Гепатологические исследования . 36 (1): 33–39. дои : 10.1016/j.hepres.2006.05.003 . ПМИД 16824795 .
- ^ Уильямс К.Л., Рахимтула М., Мироу К.М. (2005). «Hsp27 и рост аксонов во взрослых сенсорных нейронах in vitro» . BMC Нейронаука . 6 (1): 24. дои : 10.1186/1471-2202-6-24 . ПМЦ 1087488 . ПМИД 15819993 .
- ^ «Опухоли мягких тканей: Перицитома с t(7;12)» . Атлас генетики и цитогенетики в онкологии и гематологии . Университетская больница Пуатье. Архивировано из оригинала 30 декабря 2008 г. Проверено 21 января 2013 г.
- ^ Прокаччо В., Салазар Г., Оно С., Стайерс М.Л., Гиринг М., Давила А., Хименес Р., Юнкос Дж., Гутекунст К.А., Мерони Г., Фонтанелла Б., Зонтаг Е., Зонтаг Дж.М., Фаундез В., Вайнер Б.Х. (июнь 2006 г.). «Мутация бета-актина, которая изменяет динамику деполимеризации, связана с аутосомно-доминантными пороками развития, глухотой и дистонией» . Американский журнал генетики человека . 78 (6): 947–960. дои : 10.1086/504271 . ПМЦ 1474101 . ПМИД 16685646 .
- ^ Нунои Х., Ямадзаки Т., Цучия Х., Като С., Малех Х.Л., Мацуда И., Канегасаки С. (июль 1999 г.). «Гетерозиготная мутация бета-актина, связанная с дисфункцией нейтрофилов и рецидивирующей инфекцией» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (15): 8693–8698. Бибкод : 1999PNAS...96.8693N . дои : 10.1073/pnas.96.15.8693 . ПМК 17578 . ПМИД 10411937 .
- ^ Jump up to: а б «Ген: ACTG1» . ЭйсВью . Национальный центр биотехнологической информации США (NCBI). Архивировано из оригинала 18 июня 2013 г. Проверено 21 января 2013 г.
- ^ Эрба Х.П., Ганнинг П., Кедес Л. (июль 1986 г.). «Нуклеотидная последовательность мРНК гамма-цитоскелетного актина человека: аномальная эволюция генов немышечного актина позвоночных» . Исследования нуклеиновых кислот . 14 (13): 5275–5294. дои : 10.1093/нар/14.13.5275 . ПМК 311540 . ПМИД 3737401 .
- ^ Брайан К.Э., Рубинштейн П.А. (июль 2009 г.). «Аллель-специфические эффекты мутаций гамма-актина человека при глухоте (DFNA20/26) на взаимодействие актина и кофилина» . Журнал биологической химии . 284 (27): 18260–18269. дои : 10.1074/jbc.M109.015818 . ПМК 2709362 . ПМИД 19419963 .
- ^ Зоннеманн К.Дж., Фицсаймонс Д.П., Патель Дж.Р., Лю Й., Шнайдер М.Ф., Мосс Р.Л., Эрвасти Дж.М. (сентябрь 2006 г.). «Цитоплазматический гамма-актин не требуется для развития скелетных мышц, но его отсутствие приводит к прогрессирующей миопатии» . Развивающая клетка . 11 (3): 387–397. дои : 10.1016/j.devcel.2006.07.001 . ПМИД 16950128 .
- ^ Гуэн Э., Гантеле Х., Эжиль С., Ласа И., Охайон Х., Вильерс В. и др. (июнь 1999 г.). «Сравнительное исследование актиновой подвижности патогенных бактерий Listeria monocytogenes, Shigella flexneri и Rickettsia conorii». Журнал клеточной науки . 112 (11): 1697–1708. дои : 10.1242/jcs.112.11.1697 . ПМИД 10318762 .
- ^ Ламбрехтс А., Геварт К., Коссарт П., Вандекеркхове Дж., Ван Тройс М. (май 2008 г.). «Хвосты комет Listeria: механизм движения на основе актина в действии». Тенденции клеточной биологии . 18 (5): 220–227. дои : 10.1016/j.tcb.2008.03.001 . ПМИД 18396046 .
- ^ Гуэн Э., Уэлч, доктор медицинских наук, Коссарт П. (февраль 2005 г.). «Актиновая подвижность внутриклеточных патогенов». Современное мнение в микробиологии . 8 (1): 35–45. дои : 10.1016/j.mib.2004.12.013 . ПМИД 15694855 .
- ^ Паркс К.М., Янг Р.Л., Поч К.Р., Малкольм К.К., Васил М.Л., Ник Дж.А. (апрель 2009 г.). «Усиление нейтрофилами развития биопленки Pseudomonas aeruginosa: человеческий F-актин и ДНК как мишени для терапии» . Журнал медицинской микробиологии . 58 (Часть 4): 492–502. дои : 10.1099/jmm.0.005728-0 . ПМЦ 2677169 . ПМИД 19273646 .
- ^ Лю Ю., Белкина Н.В., Шоу С. (2009). «ВИЧ-инфекция Т-клеток: актин-ин и актин-аут» . Научная сигнализация . 2 (66): пэ23. дои : 10.1126/scisignal.266pe23 . ПМИД 19366992 . S2CID 30259258 .
- ^ Мачески Л.М., Тан Х.Р. (июль 2009 г.). «Выступы на основе актина: промоторы или ингибиторы инвазии рака?» . Раковая клетка . 16 (1): 5–7. дои : 10.1016/j.ccr.2009.06.009 . ПМИД 19573806 .
- ^ Хесс Х., Клемменс Дж., Цинь Д., Ховард Дж., Фогель В. (2001). «Управляемые светом молекулярные челноки, созданные из моторных белков, перевозящие груз по инженерным поверхностям». Нано-буквы . 1 (5): 235–239. Бибкод : 2001NanoL...1..235H . дои : 10.1021/nl015521e .
- ^ Манссон А., Сундберг М., Банк Р., Балаз М., Николлс И.А., Омлинг П., Тегенфельдт Дж.О., Тагеруд С., Монтелиус Л. (2005). «Молекулярные двигатели на основе актина для грузоперевозок в нанотехнологиях - потенциал и проблемы». Транзакции IEEE в расширенной упаковке . 28 (4): 547–555. дои : 10.1109/TADVP.2005.858309 . S2CID 33608087 .
- ^ Шарма С., Ханукоглу I (апрель 2019 г.). «Картирование мест локализации эпителиальных натриевых каналов (ENaC) и CFTR в сегментах придатка яичка млекопитающих». Журнал молекулярной гистологии . 50 (2): 141–154. дои : 10.1007/s10735-019-09813-3 . ПМИД 30659401 . S2CID 58026884 .
- ^ Вандесомпель Дж., Де Претер К., Паттин Ф., Поппе Б., Ван Рой Н., Де Паепе А., Спелеман Ф. (июнь 2002 г.). «Точная нормализация количественных данных RT-PCR в реальном времени путем геометрического усреднения нескольких генов внутреннего контроля» . Геномная биология . 3 (7): ИССЛЕДОВАНИЕ0034. doi : 10.1186/gb-2002-3-7-research0034 . ПМЦ 126239 . ПМИД 12184808 .
- ^ Селви С., Томпсон Э.В., Маттеи К., Леа Р.А., Ирвинг М.Г., Гриффитс Л.Р. (октябрь 2001 г.). «Бета-актин — неподходящий внутренний контроль для RT-PCR» . Молекулярные и клеточные зонды . 15 (5): 307–311. дои : 10.1006/mcpr.2001.0376 . ПМИД 11735303 .
- ^ Мукаи К., Шольмейер Й.В., Розай Дж. (январь 1981 г.). «Иммуногистохимическая локализация актина: применение в хирургической патологии». Американский журнал хирургической патологии . 5 (1): 91–97. дои : 10.1097/00000478-198101000-00013 . ПМИД 7018275 .
- ^ Хаддад Ф., Рой Р.Р., Чжун Х., Эдгертон В.Р., Болдуин К.М. (август 2003 г.). «Атрофические реакции на мышечную бездеятельность. II. Молекулярные маркеры дефицита белка». Журнал прикладной физиологии . 95 (2): 791–802. doi : 10.1152/japplphysicalol.01113.2002 . ПМИД 12716877 . S2CID 8268572 .
- ^ Хоккетт Дж. Ф., Ленерт С., Барендсе В., Кассар-Малек И., Пикард Б. (2006). «Современные достижения в протеомном анализе и его использование для определения качества мяса птицы». Мировой научный журнал по птицеводству . 62 (1): 123–130. дои : 10.1079/WPS200589 . S2CID 86189373 .
- ^ Ноллет Л. (2004). «Методы и инструменты прикладного анализа пищевых продуктов». Справочник по анализу пищевых продуктов . Том. 3 (2-е изд.). Нью-Йорк, штат Нью-Йорк: Марсель Деккер. стр. 1741–2226. ISBN 978-0-8247-5039-8 .
- ^ Halliburton WD (август 1887 г.). «О мышечной плазме» . Журнал физиологии . 8 (3–4): 133–202. doi : 10.1113/jphysicalol.1887.sp000252 . ПМЦ 1485127 . ПМИД 16991477 .
- ^ Jump up to: а б Банга I (1942). Сент-Дьёрдьи А (ред.). «Получение и свойства миозина А и Б». Исследования Института медицинской химии Университета Сегеда. 1941-1942 гг . Я : 5–15.
- ^ Штрауб Б.Ф. (1942). Сент-Дьёрдьи А (ред.). «Актин» . Исследования Института медицинской химии Сегедского университета. 1942 год . II : 3–15.
- ^ Буги Б., Келлермайер М. (март 2020 г.). «Открытие актина: «видеть то, что видели все остальные, и думать так, как никто не думал» » . Журнал исследований мышц и подвижности клеток . 41 (1): 3–9. дои : 10.1007/s10974-019-09515-z . ПМК 7109165 . ПМИД 31093826 .
- ^ Сент-Дьёрдьи А (1942). Сент-Дьёрдьи А (ред.). «Дискуссия» . Исследования Института медицинской химии Сегедского университета. 1941-1942 гг . Я : 67–71.
- ^ Сент-Дьёрдьи А (1945). «Исследования о мышцах». Acta Physiol Scandinav . 9 (Приложение): 25.
- ^ Jump up to: а б Штрауб Ф.Б., Фойер Г (1989). «Аденозитрифосфат. Функциональная группа актина. 1950». Биохимика и биофизика Acta . 1000 : 180–195. дои : 10.1016/0006-3002(50)90052-7 . ПМИД 2673365 .
- ^ Барань М., Бэррон Дж.Т., Гу Л., Барань К. (декабрь 2001 г.). «Обмен связанного с актином нуклеотида в интактных гладких мышцах артерий» . Журнал биологической химии . 276 (51): 48398–48403. дои : 10.1074/jbc.M106227200 . ПМИД 11602582 .
- ^ Ориол С., Дюбор С., Лэндон Ф. (январь 1977 г.). «Кристаллизация нативного актина поперечнополосатых мышц» . Письма ФЭБС . 73 (1): 89–91. Бибкод : 1977FEBSL..73...89O . дои : 10.1016/0014-5793(77)80022-7 . ПМИД 320040 . S2CID 5142918 .
- ^ Савая М.Р., Кудряшов Д.С., Пашков И., Адисетийо Х., Рейслер Э., Йейтс Т.О. (апрель 2008 г.). «Множественные кристаллические структуры димеров актина и их значение для взаимодействий в актиновой нити» . Acta Crystallographica Раздел D. 64 (Часть 4): 454–465. Бибкод : 2008AcCrD..64..454S . дои : 10.1107/S0907444908003351 . ПМЦ 2631129 . ПМИД 18391412 .
- ^ Нарита А., Такеда С., Ямасита А., Маэда Ю. (ноябрь 2006 г.). «Структурная основа покрытия актиновых нитей на зазубренном конце: исследование криоэлектронной микроскопии» . Журнал ЭМБО . 25 (23): 5626–5633. дои : 10.1038/sj.emboj.7601395 . ПМЦ 1679762 . ПМИД 17110933 .
- ^ Jump up to: а б Лодиш и др. 2016 , стр. 791–792.
- ^ Jump up to: а б с Лодиш и др. 2016 , стр. 792.
Цитируемые работы
[ редактировать ]- Лодиш Х.Ф., Берк А., Кайзер С., Кригер М., Бретчер А., Плох Х. и др. (2016). «Организация и движение клеток I: микрофиламенты». Молекулярно-клеточная биология (Восьмое изд.). Нью-Йорк: WH Freeman. ISBN 978-1-4641-8339-3 .
- Поллард Т.Д. (август 2016 г.). «Актин и актин-связывающие белки» . Колд Спринг Харб Перспектива Биол . 8 (8): а018226. doi : 10.1101/cshperspect.a018226 . ПМЦ 4968159 . ПМИД 26988969 .
Внешние ссылки
[ редактировать ]- Методы окрашивания актина (окрашивание живых и фиксированных клеток)
- ресурса Eukaryotic Linear Motif Класс мотива LIG_Actin_RPEL_3
- ресурсов Eukaryotic Linear Motif Класс мотивов LIG_Actin_WH2_1
- ресурсов Eukaryotic Linear Motif Класс мотивов LIG_Actin_WH2_2
- 3D макромолекулярные структуры актиновых нитей из Банка данных EM (EMDB)