Спинозавр
| Спинозавр Временной диапазон: поздний мел ( сеноман ),
| |
|---|---|

| |
| Реконструированный скелет по образцу FSAC-KK-11888 в плавательной позе, Полевой музей естественной истории. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Спинозавриды |
| Племя: | † Спинозаврины |
| Род: | † Спинозавр Стромер , 1915 год. |
| Типовой вид | |
| † Спинозавр aegyptiacus Стромер, 1915 год.
| |
| Синонимы | |
| |
Спинозавр ( / ˌ s p aɪ n ə ˈ s ɔːr ə s / ; букв. « позвоночная ящерица » ) — род динозавров , -спинозавридов живших на территории нынешней Северной Африки в сеноманском этапе позднего мелового периода , около 100-100 лет назад. 94 миллиона лет назад . [ 2 ] Род был впервые известен по египетским останкам, обнаруженным в 1912 году и описанным немецким палеонтологом Эрнстом Стромером в 1915 году . Оригинальные останки были уничтожены во время Второй мировой войны , но в начале 21 века были обнаружены дополнительные материалы. Неясно, представлены ли в окаменелостях, представленных в научной литературе, один или два вида. Самый известный вид — S. aegyptiacus из Египта, хотя потенциальный второй вид, S. maroccanus , был обнаружен в Марокко. Современный род спинозаврид Sigilmassasaurus также был синонимом некоторых авторов с S. aegyptiacus , хотя другие исследователи предполагают, что это отдельный таксон . Другой возможный младший синоним — Oxalaia из формации Алькантара в Бразилии .
Спинозавр — старейшее из известных наземных хищников ; другие крупные хищники, сравнимые со спинозавром, включают тероподов, таких как тираннозавр , гиганотозавр и кархародонтозавр . Самое последнее исследование показывает, что предыдущие оценки размера тела завышены и что S. aegyptiacus достигал 14 метров (46 футов) в длину и 7,4 метрических тонны (8,2 коротких тонны) массы тела. [ 3 ] Череп спинозавра был длинным, низким и узким, похожим на череп современного крокодила , с прямыми коническими зубами без зазубрин . У него были большие, крепкие передние конечности с трехпалыми руками и увеличенным когтем на первом пальце . Отличительные нервные отростки спинозавра или позвоночника), вырастали по меньшей мере до 1,65 метра (5,4 фута) в , которые представляли собой длинные продолжения позвонков ( длину и, вероятно, имели соединяющую их кожу, образуя структуру, подобную парусу , хотя некоторые авторы предположили, что шипы были покрыты жиром и образовывали горб. [ 4 ] Бедренные кости спинозавра были редуцированы, а ноги были очень короткими по сравнению с телом. Его длинный и узкий хвост был углублен высокими тонкими нервными отростками и удлиненными шевронами , образующими гибкую плавниковую или лопастную структуру.
Известно, что спинозавр ел рыбу, и большинство ученых полагают, что он охотился как на наземную, так и на водную добычу. Имеющиеся данные свидетельствуют о том, что он вел полуводный образ жизни ; насколько он был способен плавать, вызывает серьезные споры. остеосклероз Кости ног спинозавра имели ( высокая плотность костей), что позволяло лучше контролировать плавучесть . Для спинного паруса было предложено множество функций, включая терморегуляцию и отображение ; либо для запугивания соперников, либо для привлечения товарищей. Он жил во влажной среде приливных отмелей и мангровых лесов вместе со многими другими динозаврами, а также рыбами, крокодиломорфами , ящерицами, черепахами, птерозаврами и плезиозаврами .
Открытие и присвоение имени
Именование видов

два вида спинозавра Spinosaurus Были названы : Spinosaurus aegyptiacus (что означает «египетская позвоночная ящерица») и спорный maroccanus (что означает «марокканская позвоночная ящерица»). [ 5 ] [ 6 ] Первые описанные останки спинозавра были найдены и описаны в начале 20 века. В 1912 году Ричард Маркграф обнаружил частичный скелет гигантского динозавра-теропода в формации Бахария на западе Египта. В 1915 году немецкий палеонтолог Эрнст Штромер опубликовал статью, отнесшую этот экземпляр к новому роду и виду Spinosaurus aegyptiacus . [ 7 ] [ 5 ]
Фрагментарные дополнительные останки Бахарии, включая позвонки и кости задних конечностей, были обозначены Стромером как « Спинозавр B » в 1934 году. [ 8 ] Стромер считал их достаточно разными, чтобы принадлежать к другому виду, и это подтвердилось. Учитывая большее количество экспедиций и материалов, похоже, что они относятся либо к кархародонтозаврам, либо к кархародонтозавру. [ 9 ] или Сигилмассазавра . [ 6 ]
S. maroccanus был первоначально описан Дейлом Расселом в 1996 году как новый вид на основании длины его шейных позвонков. В частности, Рассел утверждал, что отношение длины центра ( тела позвонка) к высоте задней суставной фасетки составляло 1,1 у S. aegyptiacus и 1,5 у S. maroccanus . [ 6 ] Позже авторы разделились по этой теме. Некоторые авторы отмечают, что длина позвонков может варьироваться от человека к человеку, что образец голотипа был уничтожен и, следовательно, его нельзя сравнивать напрямую с экземпляром S. maroccanus , и что неизвестно, какие шейные позвонки S. maroccanus представляют экземпляры . Поэтому, хотя некоторые и сохранили этот вид как действительный без особых комментариев, [ 10 ] [ 11 ] [ 12 ] большинство исследователей считают S. maroccanus nomen dubium (сомнительное название). [ 13 ] [ 14 ] [ 15 ] или как младший синоним S. aegyptiacus . [ 9 ]
Образцы

шесть основных частичных экземпляров спинозавра Описано был BSP 1912 VIII 19, описанный Стромером в 1915 году из формации Бахария . Голотипом . [ 7 ] [ 5 ] Материал состоял из следующих предметов, большинство из которых были неполными: правая и левая зубные кости и сплениалы нижней челюсти длиной 75 сантиметров (30 дюймов); прямой кусок левой верхней челюсти , описанный, но не нарисованный; 20 зубов; 2 шейных позвонка; 7 спинных (туловищных) позвонков; 3 крестцовых позвонка; 1 хвостовой позвонок; 4 грудных ребра; и гастралия . Из девяти нервных отростков, высота которых указана, самый длинный («i», связанный со спинным позвонком), составлял 1,65 метра (5,4 фута) в длину. Стромер утверждал, что этот образец принадлежал раннему сеноману, около 97 миллионов лет назад. [ 7 ] [ 5 ]
Он был разрушен во время Второй мировой войны , а именно «в ночь с 24 на 25 апреля 1944 года во время британской бомбардировки Мюнхена», в результате которой было серьезно повреждено здание, в котором находится Палеонтологический музей Мюнхена (Баварская государственная коллекция палеонтологии). Однако подробные чертежи и описания экземпляра сохранились. Сын Штромера передал архивы Стромера в дар Paläontologische Staatssammlung München в 1995 году, а Смит и его коллеги проанализировали две фотографии экземпляра голотипа Spinosaurus BSP 1912 VIII 19, обнаруженного в архивах в 2000 году. На основе фотографии нижней челюсти и фотографии весь экземпляр в установленном виде, Смит пришел к выводу, что оригинал Стромера 1915 года рисунки были немного неточными. [ 5 ] Стромера спинозавра В 2003 году Оливер Раухут предположил, что голотип был химерой , состоящей из позвонков и нервных отростков кархародонтозаврид, похожих на акрокантозавра и зубной кости барионикса , или зухомима . [ 15 ] Анализ был отклонен по крайней мере в одной последующей статье. [ 13 ]
NMC 50791, хранящийся в Канадском музее природы , представляет собой среднешейный позвонок длиной 19,5 см (7,7 дюйма) из пластов Кем-Кем в Марокко . Это голотип Spinosaurus maroccanus, описанный Расселом в 1996 году. Другими экземплярами, упомянутыми в той же статье к S. maroccanus, были два других среднешейных позвонка (NMC 41768 и NMC 50790), фрагмент передней зубной кости (NMC 50832), средний зубной фрагмент (NMC 50833) и передний дорсальный нервный фрагмент арка (НМК 50813). Рассел заявил, что «может быть предоставлена только общая информация о местоположении» этого экземпляра, и поэтому его можно датировать только «возможно» альбским периодом. [ 6 ]

MNHN SAM 124, хранящийся в Национальном музее естественной истории , представляет собой морду (состоящую из частичных предчелюстных костей, частичных верхних челюстей, сошников и фрагмента зубной кости). Описанный Таке и Расселом в 1998 году, экземпляр имеет ширину от 13,4 до 13,6 см (5,3–5,4 дюйма); длина не указана. Экземпляр находился в Алжире и «имеет альбский возраст». Таке и Рассел полагали, что образец вместе с фрагментом предчелюстной кости (SAM 125), двумя шейными позвонками (SAM 126–127) и дорсальной нервной дугой (SAM 128) принадлежал S. maroccanus . [ 16 ]
BM231 (в коллекции Национального горного управления, Тунис) был описан Бюффето и Уахой в 2002 году. Он состоит из частичной передней зубной кости длиной 11,5 см (4,5 дюйма) из слоя раннего альба формации Шенини в Тунисе . Фрагмент зубной кости, включавший четыре альвеолы и два частичных зуба, был «чрезвычайно похож» на существующий материал S. aegyptiacus . [ 14 ]
UCPC-2 из палеонтологической коллекции Чикагского университета состоит в основном из двух узких соединенных носовых пазух с рифленым (гребневым) гребнем из области между глазами. Образец длиной 18,0 см (7,1 дюйма) был обнаружен в ранней сеноманской части марокканских слоев Кем Кем в 1996 году и описан в научной литературе в 2005 году Криштиану Даль Сассо из Гражданского музея естественной истории в Милане и его коллегами. . [ 13 ]
MSNM V4047 (в Museo di Storia Naturale di Milano ), описанный Далем Сассо и его коллегами в 2005 году как Spinosaurus cf. S. aegyptiacus состоит из рыла (предчелюстные кости, части челюстей и части носа) длиной 98,8 см (38,9 дюйма) из кем-кемских слоев. [ 13 ] Изолированный рыбий позвонок, предварительно отнесенный к Onchopristis , был связан с альвеолой зуба этого экземпляра. [ 13 ] Точно так же фрагмент зубной кости Spinosaurus aegyptiacus , MPDM 31, связан с ростральным зубом Onchopristis . [ 17 ] Считается, что, как и UCPC-2, он произошел из раннего сеномана. Арден и его коллеги в 2018 году предварительно отнесли этот экземпляр к Sigilmassasaurus brevicollis, учитывая его размер. [ 18 ] Однако позже это назначение было отвергнуто другими исследователями, которые посчитали, что уникальность этого экземпляра основана на неправильных интерпретациях и плохой сохранности другого экземпляра, NHMUK R16665, еще одного морда, хранящегося в Музее естественной истории в Лондоне . [ 1 ]
FSAC-KK 11888 представляет собой частичный скелет полувзрослой особи, обнаруженный в отложениях Кем-Кем в Северной Африке. Он был описан Ибрагимом и его коллегами в 2014 году и обозначен как образец неотипа. [ 19 ] хотя Эверс и его коллеги отклонили неотипическое обозначение FSAC-KK-11888 в 2015 году. [ 20 ] В его состав входят шейные позвонки, спинные позвонки, нервные отростки, весь крестец, бедра, голени, педальные фаланги, хвостовые позвонки, несколько спинных ребер и фрагменты черепа. [ 19 ] Пропорции тела экземпляра являются спорными, поскольку задние конечности у этого экземпляра непропорционально короче, чем на предыдущих реконструкциях. Однако несколько палеонтологов продемонстрировали, что этот экземпляр не является химерой, а действительно является экземпляром спинозавра , что позволяет предположить, что у животного были гораздо меньшие задние конечности, чем считалось ранее. [ 21 ] [ 22 ] [ 23 ]
Другие известные экземпляры состоят в основном из очень фрагментарных останков и разрозненных зубов. К ним относятся:
- В статье 1986 года описаны призматические структуры зубной эмали двух зубов спинозавра из Туниса. [ 24 ]
- три экземпляра из Института и музея геологии и палеонтологии Геттингенского университета в Германии Баффетаут (1989, 1992) отнес к спинозавру : фрагмент правой верхней челюсти IMGP 969–1, фрагмент челюсти IMGP 969–2 и зуб IMGP 969. –3. [ 25 ] [ 26 ] Они были обнаружены в отложениях нижнего сеномана или верхнего альба на юго-востоке Марокко в 1971 году. [ 25 ]
- Келлнер и Мадер (1997) описали два зуба спинозаврид без зазубрин из Марокко (LINHM 001 и 002), которые были «очень похожи» на зубы голотипа S. aegyptiacus . [ 27 ]
- Зубы из формации Шенини в Тунисе, которые «узкие, несколько закругленные в поперечном сечении и лишены передних и задних зазубренных краев, характерных для теропод и базальных архозавров », были отнесены к спинозавру в 2000 году. [ 28 ]
- Зубы из формации Эчкар в Нигере были предварительно отнесены к спинозавру в 2007 году. [ 29 ]
- Частичный зуб длиной 8 сантиметров (3,1 дюйма), купленный на выставке окаменелостей, предположительно из слоя Кем-Кем в Марокко и приписываемый Spinosaurus maroccanus , имел продольные бороздки и микроструктуры шириной от 1 до 5 миллиметров (0,039-0,197 дюйма). неправильные гребни) среди полосок в статье 2010 года. [ 12 ]
MHNM.KK374–.KK378 — это пять изолированных квадратов (костей черепа) разных размеров, которые были собраны местными жителями и приобретены на коммерческой основе в регион Кем-Кем на юго-востоке Марокко, предоставленный Франсуа Эскуилье и хранящийся в коллекциях Музея естественной истории в Марракеше. Квадраты имеют две разные морфологии, что позволяет предположить существование двух спинозавринов в Марокко. [ 30 ] Однако исследование вариаций внутри спинозавра, проведенное в 2020 году , считает, что эти различия в морфологии указывают на вариации морфологии черепа внутри одного вида, как в случае с аллозавром. [ 1 ]
Возможные образцы
Сообщалось , что материал, возможно, принадлежащий спинозавру, был обнаружен в районе Туркана Гриц в Кении . [ 31 ]
Некоторые ученые считали род Sigilmassasaurus младшим синонимом спинозавра . В работе Ибрагима и его коллег (2014) экземпляры Sigilmassasaurus были отнесены к Spinosaurus aegyptiacus вместе со «Spinosaurus B» как неотип , а Spinosaurus maroccanus считался nomen dubium согласно выводам других статей. [ 13 ] [ 19 ] [ 9 ] Повторное описание Sigilmassasaurus в 2015 году оспорило эти выводы и признало род действительным. [ 20 ] Этот вывод был дополнительно поддержан в 2018 году Арденом и его коллегами, которые считают Sigilmassasaurus отдельным родом, хотя и очень близким родственником Spinosaurus , двух объединенных в трибу Spinosaurini, выведенное в исследовании. [ 18 ]
В статье 2020 года, написанной Symth et al. При оценке образцов спинозавра из группы Кем Кем предположил, что бразильский спинозавр Oxalaia может быть потенциальным младшим синонимом Spinosaurus aegyptiacus . Это было основано на рассмотрении экземпляров, отнесенных к Oxalaia , и предполагаемых аутапоморфиях этого таксона как незначительных и попадающих в гиподигму Spinosaurus aegyptiacus . Если это подтвердится будущими исследованиями, это будет означать, что Spinosaurus aegyptiacus имел более широкое распространение и поддерживал обмен фауной между Южной Америкой и Африкой в это время. Кроме того, исследование также указывает на синонимичность между спинозавром и сигилмассазавром . [ 1 ]
Описание
Размер
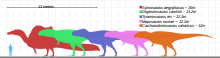
С момента своего открытия спинозавр был претендентом на звание крупнейшего динозавра-теропода. [ 32 ] Оба Фридриха фон Хюне в 1926 году [ 33 ] и Дональд Ф. Глут в 1982 году назвал его одним из самых массивных теропод в своих исследованиях: его длина составляла 15 метров (49 футов), а вес - более 6 метрических тонн (6,6 коротких тонн). [ 34 ] В 1988 году Грегори С. Пол также назвал его самым длинным тероподом - 15 метров (49 футов), но дал меньшую оценку массы - 4 метрических тонны (4,4 коротких тонны). [ 35 ]
В 2005 году Даль Сассо и его коллеги предположили, что спинозавр и спинозавр Зухомимус имели одинаковые пропорции тела по отношению к длине черепа, и таким образом рассчитали, что спинозавр имел длину от 16 до 18 метров (от 52 до 59 футов) и вес от 7 до 9 метрических тонн. (от 7,7 до 9,9 коротких тонн) по весу. [ 13 ] Оценки подверглись критике, поскольку оценка длины черепа была неточной, и (при условии, что масса тела увеличивается пропорционально кубу длины тела) масштабирование Зухомима, длина которого составляла 11 метров (36 футов), а масса - 3,8 метрических тонны (4,2 коротких тонны), В диапазоне предполагаемой длины спинозавр будет иметь предполагаемую массу тела от 11,7 до 16,7 метрических тонн (от 12,9 до 18,4 коротких тонн). [ 36 ]

Франсуа Терриен и Дональд Хендерсон в статье 2007 года, использующей масштабирование на основе длины черепа, оспорили предыдущие оценки размера спинозавра , найдя длину слишком большой, а вес слишком маленьким. Основываясь на предполагаемой длине черепа от 1,5 до 1,75 метра (от 4,9 до 5,7 футов), их оценки включают длину тела от 12,6 до 14,3 метра (от 41 до 47 футов) и массу тела от 12 до 20,9 метрических тонн (от 13,2 до 23,0 коротких тонн). ). Более низкие оценки спинозавра подразумевают, что животное было короче и легче, чем кархародонтозавр и гиганотозавр . [ 36 ] Исследование Терриена и Хендерсона подверглось критике за выбор теропод, использованных для сравнения (например, большинство теропод, использованных для установления первоначальных уравнений, были тираннозавридами и карнозаврами , которые имеют телосложение, отличное от спинозавридов), а также за предположение, что спинозавриды Длина черепа могла составлять всего 1,5 метра (4,9 фута). [ 37 ] [ 38 ]
В 2014 году Ибрагим и его коллеги предположили, что Spinosaurus aegyptiacus может достигать более 15 метров (49 футов) в длину. [ 19 ] Однако в 2022 году Пол Серено и его коллеги предположили, что Spinosaurus aegyptiacus достиг максимальной длины тела 14 метров (46 футов) и максимальной массы тела 7,4 метрических тонны (8,2 коротких тонны), построив трехмерную модель скелета на основе компьютерной томографии». с осевой колонной в нейтральном положении». [ 3 ] Они утверждали, что 2D-графическая реконструкция водной гипотезы Ибрагима и его коллег в 2020 году [ 17 ] переоценил длину пресакральной колонны на 10%, глубину грудной клетки на 25% и длину передних конечностей на 30% по сравнению с размерами, основанными на окаменелостях, полученных с помощью компьютерной томографии; эти пропорциональные завышенные оценки смещают центр масс вперед при переводе на модель тела, и поэтому оценку Ибрагима и его коллег нельзя считать надежной оценкой размера тела. [ 3 ]
Череп

Его череп имел узкую морду с прямыми коническими зубами без зазубрин. Зубов было по шесть-семь с каждой стороны самой передней части верхней челюсти, в предчелюстных костях , и еще двенадцать в обеих челюстях позади них. Второй и третий зубы с каждой стороны были заметно крупнее остальных зубов предчелюстной кости, что создавало пространство между ними и крупными зубами передней части верхней челюсти; К этому пространству обращены крупные зубы нижней челюсти. Самый кончик морды, на котором находились несколько крупных передних зубов, был расширен, а перед глазами имелся небольшой гребень. Используя размеры трех экземпляров, известных как MSNM V4047, UCPC-2 и BSP 1912 VIII 19, и предполагая, что заглазничная часть черепа MSNM V4047 имела форму, аналогичную заглазничной части черепа Ирритатора , Даля Сассо и коллеги (2005) подсчитали, что череп спинозавра составлял 1,75 метра (5,7 футов). длинный, [ 13 ] но по более поздним оценкам длина составляет 1,6–1,68 метра (5,2–5,5 футов). [ 30 ] [ 39 ] Оценка длины черепа Даля Сассо и его коллег подвергается сомнению, поскольку форма черепа может различаться у разных видов спинозаврид, а также потому, что MSNM V4047 может не принадлежать самому спинозавру . [ 36 ] [ 18 ]
Посткраниальный скелет

Будучи спинозавридом, спинозавр имел длинную мускулистую шею, изогнутую в сигмовидной или S-образной форме. Его плечи были выдающимися, а передние конечности большие и коренастые, с тремя когтистыми пальцами на каждой руке. Первый палец (или «большой») был бы самым большим. У спинозавра были длинные фаланги (кости пальцев) и лишь несколько загнутые когти , что позволяет предположить, что его руки были длиннее по сравнению с руками других спинозавридов. [ 19 ] [ 18 ] [ 40 ]
Очень высокие нервные отростки, растущие на задних позвонках спинозавра, » животного легли в основу того, что обычно называют « парусом . Длина нервных отростков более чем в 10 раз превышала диаметры центров ( тел позвонков), от которых они отходили. [ 41 ] [ 42 ] Нервные отростки были немного длиннее спереди назад у основания, чем выше, и отличались от тонких стержней, наблюдаемых у пеликозавров плавников Edaphosaurus и Dimetrodon , а также контрастировали с более толстыми шипами у игуанодонтийного Ouranosaurus . [ 41 ]

Паруса спинозавра были необычными, хотя другие динозавры, а именно уранозавр , который жил несколькими миллионами лет назад в том же регионе, что и спинозавр , и южноамериканский зауропод амаргазавр , возможно, развили аналогичные структурные адаптации своих позвонков. Парус может быть аналогом паруса пермского синапсида диметродона , жившего ещё до появления динозавров, созданного путём конвергентной эволюции . [ 41 ]
Структура, возможно, также была больше похожа на горб, чем на парус, как отмечал Стромер в 1915 году («можно скорее подумать о существовании большого горба жира [ нем . Fettbuckel ], которому [нервные отростки] давали внутренняя поддержка") [ 7 ] и Джек Боуман Бэйли в 1997 году. [ 41 ] В поддержку своей гипотезы «буйволиной спины» Бэйли утверждал, что у спинозавра , уранозавра и других динозавров с длинными нервными шипами шипы были относительно короче и толще, чем шипы пеликозавров (которые, как известно, имеют паруса); вместо этого нервные отростки динозавров были похожи на нервные отростки вымерших горбатых млекопитающих, таких как Megacerops и Bison latifrons . [ 41 ] [ 43 ] В 2014 году Ибрагим и его коллеги вместо этого предположили, что шипы были плотно покрыты кожей, как у хохлатого хамелеона , учитывая их компактность, острые края и, вероятно, плохой кровоток . [ 19 ]
У спинозавра был значительно меньший таз (тазовая кость), чем у других гигантских теропод, а площадь поверхности подвздошной кости (основная часть таза) была вдвое меньше, чем у большинства членов клады. Задние конечности были короткими, составляли чуть более 25 процентов от общей длины тела, при этом большеберцовая кость (икроножная кость) была длиннее бедренной кости (бедренная кость). В отличие от других теропод, большой палец (или четвертый палец) спинозавра касался земли, а фаланги костей пальцев ног были необычайно длинными и хорошо построенными. На их концах располагались неглубокие клешни с плоским дном. Этот тип морфологии стоп также наблюдается у куликов , что указывает на то, что ступни спинозавра развились для ходьбы по нестабильному субстрату и что они, возможно, были перепончатыми . [ 19 ]
Из хвостовых позвонков хвоста отходили значительно удлиненные тонкие нервные отростки, подобные состоянию, наблюдаемому у некоторых других спинозаврид. [ 18 ] хотя и в более крайней степени. В сочетании с удлиненными шевронными костями на нижней стороне хвостового плавника это привело к образованию глубокого и узкого хвоста веслообразной или плавниковой формы, сравнимого с хвостами тритонов и крокодилов . [ 44 ]
Классификация

Спинозавр динозавров дал свое название семейству Spinosauridae , которое включает два подсемейства: Baryonychinae и Spinosaurinae. Baryonychinae включает Baryonyx южной Англии иuchomimus из из Нигера в центральной Африке . Spinosaurinae включает Spinosaurus , Sigilmassasaurus , Oxalaia , Siamosaurus , Ichthyovenator , Irritator из Бразилии и Angaturama (что может быть синонимом Irritator ) из Бразилии. [ 13 ] [ 18 ] У спинозавринов общие прямые зубы без зазубрин, которые широко расположены (например, 12 на одной стороне верхней челюсти), в отличие от барионихин, у которых много зазубренных изогнутых зубов (например, 30 на одной стороне верхней челюсти). [ 13 ] [ 10 ]
Анализ спинозавридов, проведенный Арденом и его коллегами (2018), назвал кладу Spinosaurini и определил ее как всех спинозавридов, более близких к Spinosaurus aegyptiacus, чем к Irritator Challengeri или Oxalaia quilombensis ; он также обнаружил, что Siamosaurus suteethorni и Icthyovenator laosensis являются членами Spinosaurinae. [ 18 ]
Филогения
Подсемейство Spinosaurinae было названо Серено в 1998 году и определено Хольцем и его коллегами (2004) как все таксоны, более близкие к Spinosaurus aegyptiacus, чем к Baryonyx walkeri . Подсемейство Baryonychinae было названо Чаригом и Милнером в 1986 году. Они выделили как подсемейство, так и семейство Baryonychidae для недавно открытого Baryonyx , прежде чем его отнесли к Spinosauridae. Их подсемейство было определено Хольцем и его коллегами в 2004 году как дополнительная клада всех таксонов, более близких к Baryonyx walkeri, чем к Spinosaurus aegyptiacus . Исследования Маркоса Сейлса, Сезара Шульца и коллег (2017) показывают, что южноамериканские спинозавриды Angaturama , Irritator и Oxalaia занимали промежуточное положение между Baronychinae и Spinosaurinae на основании их краниодентальных особенностей и кладистического анализа. Это указывает на то, что Baryonychinae на самом деле могут быть немонофилетическими. Их кладограмму можно увидеть ниже. [ 45 ]

| Спинозавриды |
| ||||||||||||||||||||||||
На кладограмме ниже отражены выводы Ардена и его коллег (2018): [ 18 ]
| Спинозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Функция нервных отростков

Функция паруса или горба динозавра неясна; ученые предложили несколько гипотез, включая регулирование и отображение тепла. Кроме того, такая заметная особенность на его спине могла заставить его казаться даже больше, чем он был на самом деле, отпугивая других животных. [ 41 ]
Возможно, конструкция использовалась для терморегуляции . Если бы в структуре было много кровеносных сосудов, животное могло бы использовать большую площадь поверхности паруса для поглощения тепла. Это означало бы, что животное было в лучшем случае лишь частично теплокровным и жило в климате, где ночные температуры были прохладными или низкими, а небо обычно не облачным. Также возможно, что конструкция использовалась для излучения избыточного тепла от тела, а не для его сбора. Крупные животные из-за относительно небольшого соотношения площади поверхности их тела по сравнению с общим объемом ( принцип Холдейна ) сталкиваются с гораздо большими проблемами рассеивания избыточного тепла при более высоких температурах, чем получения его при более низких. Паруса крупных динозавров значительно увеличили площадь кожи их тел при минимальном увеличении объема. Более того, если бы парус был повернут от солнца или расположен под углом 90 градусов к прохладному ветру, животное вполне эффективно охладило бы себя в теплом климате меловой Африки. [ 46 ] Однако Бэйли (1997) придерживался мнения, что парус мог поглощать больше тепла, чем излучать. Вместо этого Бейли предположил, что у спинозавров и других динозавров с длинными нервными шипами были жировые горбы на спине для хранения энергии, изоляции и защиты от жары. [ 41 ]
Многие сложные структуры тела современных животных служат для привлечения представителей противоположного пола во время спаривания. Возможно, для ухаживания спинозавр использовал парус, похожий на павлина хвост . Стромер предположил, что размер нервных отростков мог различаться у мужчин и женщин. [ 7 ]

Гимса и его коллеги (2015) предполагают, что спинной парус спинозавра был аналогичен спинным плавникам парусника и служил гидродинамической цели. [ 47 ] Гимса и другие отмечают, что у более базальных, длинноногих спинозавридов спинные паруса имели круглую или серповидную форму, тогда как у спинозавров спинные нервные отростки имели примерно прямоугольную форму, похожую по форме на спинные плавники парусников. Поэтому они утверждают, что спинозавр использовал свой спинной нервный парус так же, как рыба-парусник, и что он также использовал свой длинный узкий хвост, чтобы оглушать добычу, как современная акула-молотилка . Парусник использует свои спинные плавники, чтобы собрать косяки рыбы в « шарик с приманкой », где они вместе ловят рыбу в определенной области, где рыба-парусник может схватить рыбу своими клювами. Парус мог бы уменьшить вращение рыскания, противодействуя боковой силе в направлении, противоположном удару, как предположили Гимса и его коллеги (2015). [ 47 ]
В анатомии спинозавра есть еще одна особенность, которая может иметь современную аналогию: его длинный хвост напоминал хвост акулы-молотилки, которую использовали для того, чтобы шлепать по воде стада и оглушать косяки рыб перед тем, как их съесть (Оливер и коллеги, 2013). Стратегии, которые используют парусники и акулы-молотилы против стайной рыбы, более эффективны, когда косяк сначала концентрируется в «шар приманки» (Helfman, Collette & Facey, 1997; Oliver et al., 2013; Domenici et al., 2014). Поскольку отдельным хищникам трудно добиться этого, они сотрудничают в этих усилиях. Выпасая косяк рыб или кальмаров, парусник также поднимает паруса, чтобы казаться больше. Когда они протирают клювами стайную рыбу, поворачивая голову, их спинной парус и плавники вытягиваются, чтобы стабилизировать тело гидродинамически (Lauder & Drucker, 2004). Доменичи и его коллеги (2014) предполагают, что эти удлинения плавников повышают точность постукивания и рубящих ударов. Парус может уменьшить вращение рыскания, противодействуя боковой силе в направлении, противоположном удару. Это означает, что жертва с меньшей вероятностью узнает в массивном хоботе часть приближающегося хищника (Маррас и коллеги, 2015; Уэбб и Вейхс, 2015). [ 47 ]
Спинозавр обладал анатомическими особенностями, необходимыми для объединения всех трех охотничьих стратегий: парус для более эффективного выпаса добычи, а также гибкий хвост и шея, позволяющие ударять по воде для оглушения, ранения или убийства добычи. Погруженный спинной парус мог бы обеспечить мощное противодействие мощным движениям сильной шеи и длинного хвоста в стороны, как это делают парусники (Доменичи и коллеги, 2014) или акулы-молотилки (Оливер и коллеги, 2013). В то время как меньшие спинные паруса или плавники делают спинной объем воды более доступным для разрезания, можно предположить, что их меньший стабилизирующий эффект делает боковое разрезание менее эффективным (например, для акул-молотил). Образуя гидродинамическую точку опоры и гидродинамически стабилизируя туловище вдоль дорсовентральной оси, парус спинозавра компенсировал бы также инерцию боковой шеи движениями хвоста и наоборот не только при хищничестве, но и при ускоренном плавании. Такое поведение также могло быть одной из причин Мускулистая грудь и шея спинозавра , о которых сообщили Ибрагим и его коллеги (2014). [ 47 ]
Диета и кормление

Неясно, был ли спинозавр в первую очередь наземным хищником или рыбоядным животным , о чем свидетельствуют его удлиненные челюсти, конические зубы и приподнятые ноздри. Гипотеза о спинозаврах как о специализированных пожирателях рыбы была предложена ранее А.Дж. Чаригом и А.С. Милнером для барионикса . Они основывают это на анатомическом сходстве с крокодилами и наличии протравленной пищеварительной кислотой рыбьей чешуи в грудной клетке типового экземпляра . [ 48 ] Крупные рыбы известны из фаун, содержащих другие спинозавриды, в том числе Mawsonia , в середине мела Северной Африки и Бразилии. Прямые доказательства диеты спинозавров получены из родственных европейских и южноамериканских таксонов. Барионикс был найден с рыбьей чешуей и костями молодого игуанодона в его желудке, а зуб, встроенный в кость южноамериканского птерозавра, позволяет предположить, что спинозавры иногда охотились на птерозавров. [ 49 ] но спинозавр, вероятно, был обобщенным и оппортунистическим хищником, возможно, меловым эквивалентом крупных медведей гризли , склонным к рыбной ловле, хотя он, несомненно, питался мусором и ловил много видов мелкой и средней добычи. [ 35 ]

В 2009 году Даль Сассо и коллеги. сообщили о результатах рентгеновской компьютерной томографии морды MSNM V4047. Поскольку все внешние отверстия сообщались с пространством внутри морды, авторы предположили, что у спинозавра внутри этого пространства были рецепторы давления , которые позволяли ему удерживать морду на поверхности воды, чтобы обнаруживать плавающие виды добычи, не видя их. . [ 50 ] Исследование, проведенное в 2013 году Эндрю Р. Каффом и Эмили Дж. Рэйфилд, пришло к выводу, что биомеханические данные свидетельствуют о том, что спинозавр не был обязательным рыбоядным животным и что его диета более тесно связана с размером каждой особи. Характерная ростральная морфология спинозавра позволяла его челюстям сопротивляться изгибу в вертикальном направлении, но его челюсти были плохо приспособлены к сопротивлению боковому изгибу по сравнению с другими представителями этой группы ( бариониксами ) и современными аллигаторами. Это говорит о том, что спинозавры чаще охотились на рыбу, чем на наземных животных, хотя и считались хищниками первых. [ 51 ] В 2022 году Сакамото подсчитал, что спинозавра сила переднего укуса составляла 4829 ньютонов, а сила заднего укуса - 11 936 ньютонов. Основываясь на этой оценке, он утверждал, что челюсти спинозавра приспособлены для создания относительно более высоких скоростей смыкания с меньшей мышечной силой, что указывает на то, что животное, вероятно, убивало свою добычу быстро щелкающими челюстями, а не медленными сокрушительными укусами, что обычно наблюдается. у животных, ведущих полуводный образ жизни. [ 52 ]
Водные привычки

проведенный Роменом Амио и его коллегами в 2010 году, Изотопный анализ, показал, что соотношение изотопов кислорода в зубах спинозаврид, включая зубы спинозавра , указывает на полуводный образ жизни. Соотношения изотопов зубной эмали и других частей спинозавра (обитаемого в Марокко и Тунисе) и других хищников из того же региона, таких как кархародонтозавр, сравнивались с изотопным составом современных теропод, черепах и крокодилов. Исследование показало, что зубы спинозавра из пяти из шести выбранных мест имели соотношение изотопов кислорода, более близкое к таковому у черепах и крокодилов по сравнению с зубами других теропод из тех же мест. Авторы предположили, что спинозавр переключался между наземной и водной средой обитания, чтобы конкурировать за пищу с крупными крокодилами и другими крупными тероподами соответственно. [ 53 ] Однако исследование Дональда Хендерсона, проведенное в 2018 году, опровергает утверждение о том, что спинозавр вел полуводный образ жизни. Изучая плавучесть легких крокодилов и сравнивая ее с расположением легких у спинозавра , было обнаружено, что спинозавр не мог тонуть или нырять под поверхность воды. Он также был способен держать всю голову над поверхностью воды во время плавания, как и другие неводные тероподы. Кроме того, исследование показало, что спинозавру приходилось постоянно грести задними ногами, чтобы не допустить опрокидывания на бок, чего не нужно делать современным полуводным животным. Поэтому Хендерсон предположил, что спинозавр , вероятно, не охотился полностью погруженным в воду, как предполагалось ранее, а вместо этого проводил большую часть своего времени на суше или на мелководье. [ 54 ] [ 55 ]
Недавние исследования хвостовых позвонков спинозавра опровергают предположение Хендерсона о том, что спинозавр в основном населял участки суши вблизи и на мелководье и был слишком плавучим, чтобы погружаться в воду. Исследования хвоста, проведенные благодаря окаменелостям, обнаруженным и проанализированным Ибрагимом, Пирсом, Лаудером, Серено и их коллегами в 2018 году, показывают, что у спинозавра был килевидный хвост, который был хорошо приспособлен для передвижения животного по воде. Удлиненные нервные отростки и шевроны, доходящие до конца хвоста как на спинной, так и на брюшной стороне, указывают на то, что спинозавр был способен плавать так же, как современные крокодилы. В результате экспериментов Лаудера и Пирса было обнаружено, что хвост спинозавра имеет в восемь раз большую тягу вперед, чем хвосты наземных теропод, таких как целофиз и аллозавр , а также в два раза эффективнее обеспечивает тягу вперед. Открытие указывает на то, что спинозавр, возможно, вел образ жизни, сравнимый с образом жизни современных аллигаторов и крокодилов, оставаясь в воде в течение длительного периода времени во время охоты. [ 44 ]

Дэвид Хоун и Томас Хольц опубликовали в 2021 году статью, в которой утверждают, что анатомия спинозавра больше соответствует прибрежному образу жизни, а не активному хищнику, преследующему воду, как предположил Ибрагим. [ 56 ] Они подчеркивают расположение ноздрей и глазниц как одну из причин, почему крокодиловый образ жизни маловероятен: они расположены вентрально таким образом, что всю голову придется неэффективно поднимать из воды, чтобы дышать. Кроме того, они утверждают, что общая форма тела спинозавра плохо приспособлена для такого образа жизни, что связано с сопротивлением воды и водной нестабильностью. [ 55 ] от паруса, а также от жесткого туловища и, казалось бы, едва мускулистого хвоста. Таким животным, как крокодилы, требуется гибкое тело, чтобы передвигаться по воде и делать резкие повороты при преследовании добычи, а это прямо противоречит выводам Хоуна и Хольца.
Исследование Fabbri et al. , проведенное в 2022 году. , сравнил спинозавра костную структуру и сравнил ее со структурой барионикса и зухомима . Исследование показало, что у спинозавра и барионикса были плотные кости, что позволяло им нырять и преследовать добычу под водой. По сравнению с ними, у Зухомима было больше полых костей, что позволяет предположить, что он предпочитал охотиться на мелководье. Эти данные также предполагают, что различные роды спинозавридов были более экологически разными, чем считалось ранее, поскольку некоторые из них лучше подходили для охоты в подводной среде, чем другие, близкородственные роды. [ 57 ] [ 58 ] [ 59 ]
В том же году, вопреки исследованию Фаббри и его коллег, Серено и его коллеги предположили, что спинозавр был полностью двуногим на суше и неустойчивым, медленно передвигающимся поверхностным пловцом в глубокой воде. Их результаты взяты из реконструкции компьютерной модели скелета с последующим добавлением внутреннего воздуха и мышц. Их результаты, в сочетании с окаменелостями спинозавра , которые показали, что он также жил дальше вглубь страны, вдоль рек и озер, позволяют предположить, что это было полуводное рыбоядное животное, которое предпочитало водную среду как вдоль побережья, так и дальше вглубь страны, вдоль рек и озер. Одновременно они предположили, что большой хвостовой плавник, вероятно, использовался больше для демонстрации, чем для плавания, поскольку хвосты у современных животных выполняют ту же функцию, когда они обладают сравнительно высокими нервными отростками. [ 3 ]
Передвижение и осанка

традиционно изображался в научном сообществе как двуногое , Хотя спинозавр в середине 20-го века его время от времени изображали как облигатное четвероногое животное, родственное диметродону . [ 60 ] Начиная с середины 1970-х годов предполагалось, что спинозавр, по крайней мере, иногда был четвероногим . [ 34 ] [ 46 ] подкреплено открытием Барионикса , родственника с крепкими руками. [ 61 ] Из-за массы предполагаемых жировых горбов спинозавра Бэйли (1997) предположил возможность четвероногого положения. [ 41 ] что приводит к новым реставрациям его как такового. [ 61 ] Тероподы, в том числе спинозавриды, не могли пронационировать руки (поворачивать предплечье так, чтобы ладонь была обращена к земле). [ 62 ] но положение покоя на боковой стороне руки было возможным, как показывают отпечатки окаменелостей раннеюрского теропода. [ 63 ] Гипотеза о том, что спинозавры имели типичную четвероногие походки, с тех пор потеряла популярность, однако все еще считалось, что спинозавриды могли приседать в четвероногой позе из-за биологических и физиологических ограничений. [ 48 ] [ 62 ]

Возможность существования четвероногого спинозавра была возрождена в статье Ибрагима и его коллег в 2014 году, в которой описывался новый материал об этом животном. В статье было обнаружено, что задние конечности спинозавра были намного короче, чем считалось ранее, и что его центр массы располагался в средней точке туловища , а не возле бедра, как у типичных двуногих теропод. Поэтому было высказано предположение, что спинозавр был плохо приспособлен к двуногому наземному передвижению и, должно быть, был обязательным четвероногим на суше. Реконструкция, использованная в исследовании, представляла собой экстраполяцию, основанную на особях разного размера, масштабированных до пропорций, которые считались правильными. [ 19 ] Палеонтолог Джон Хатчинсон из Королевского ветеринарного колледжа Лондонского университета выразил скептицизм по поводу новой реконструкции и предупредил, что использование разных образцов может привести к получению неточных химер. [ 64 ] Скотт Хартман также выразил критику, поскольку, по его мнению, размеры ног и таза были неточно (слишком короткими на 27%) и не соответствовали опубликованным длинам. [ 65 ] Однако Марк Уиттон выразил согласие с пропорциями, указанными в статье. [ 66 ] В своем переописании Sigilmassasaurus в 2015 году Эверс и его коллеги утверждали, что Sigilmassasaurus на самом деле был отдельным родом от Spinosaurus , и поэтому сомневались в том, что материал, отнесенный к Spinosaurus Ибрагимом и др. следует отнести к спинозавру или сигилмассазауру . [ 20 ] В 2018 году анализ, проведенный Хендерсоном, показал, что спинозавр, вероятно, был способен передвигаться по земле на двух ногах; вместо этого было обнаружено, что центр массы находился близко к бедрам, что позволяло спинозавру стоять прямо, как и другие двуногие тероподы. [ 55 ]
В статье 2024 года, соавтором которой был Серено, говорилось, что предыдущие расчеты Серено, которые использовались для обоснования четвероногости спинозавра, ошибочно сместили центр массы перед бедрами. Вместо этого они предположили, что динозавр соответствует критериям гравипортального (или медленно движущегося) двуногого существа. [ 67 ] [ 68 ]
Онтогенез
Ногтевая фаланга размером 21 миллиметр (0,83 дюйма), принадлежащая очень молодому молодому спинозавру, указывает на то, что теропод развил свои полуводные адаптации в очень молодом возрасте или при рождении и сохранял их на протяжении всей своей жизни. Образец, найденный в 1999 году и описанный Симоне Магануко, Криштиану Даль Сассо и коллегами, как полагают, произошел от животного размером 1,78 метра (5,8 футов) (при условии, что он напоминал уменьшенную версию взрослой особи), что делает его самым маленьким экземпляром. настоящее время спинозавров известных в . [ 69 ]
Палеопатология
Ср. Спинозавр сп. зуб из формации Ифесуан демонстрирует усиленную язычную кривизну коронки зуба, развитие трех глубоких борозд, идущих от соединения коронки и корня в направлении вершины коронки, ослабленный киль, который не распространяется ни апикально, ни до основания зуба, и фасетка износа на кончике. [ 70 ]
Палеосреда

Среда, в которой обитают спинозавры , изучена лишь частично и охватывает большую часть территории современной Северной Африки. Регион Африки Спинозавр сохранился в датах от 112 до 93,5 миллионов лет назад, [ 71 ] [ 5 ] [ 72 ] хотя потенциальный образец был найден в кампана . отложениях [ 73 ] [ 74 ] Исследование 1996 года на основе марокканских окаменелостей пришло к выводу, что спинозавр , кархародонтозавр и дельтадромеус «распространялись по Северной Африке в позднем меловом периоде (сеномане)». [ 75 ] Те спинозавры , которые жили в формации Бахария на территории современного Египта, возможно, боролись с условиями береговой линии на приливных отмелях и каналах, живя в мангровых лесах вместе с такими же крупными хищниками-динозаврами Бахариазавром и Кархародонтозавром , -титанозаврами зауроподами Paralititan и Aegyptosaurus , крокодиломорфами , костными и хрящевыми. рыбы, черепахи, ящерицы, и плезиозавры . [ 76 ] В засушливый сезон он мог прибегнуть к охоте на птерозавров . [ 77 ] Эта ситуация напоминает ситуацию в позднеюрской формации Моррисон в Северной Америке , которая может похвастаться до пяти родов теропод весом более 1 метрической тонны (1,1 коротких тонны), а также несколькими более мелкими родами (Henderson, 1998; Holtz et al. , 2004). . Различий в форме головы и размере тела у крупных североафриканских теропод, возможно, было достаточно, чтобы обеспечить разделение ниш , как это наблюдается среди многих различных видов хищников, обитающих сегодня в африканской саванне (Farlow & Pianka, 2002).
В популярной культуре

Спинозавр появился в фильме 2001 года «Парк Юрского периода III» , заменив Тираннозавра в качестве главного антагониста. [ 78 ] Палеонтолог-консультант фильма Джон Р. Хорнер сказал: «Если мы будем основывать фактор свирепости на длине животного, то на этой планете не было ничего, что когда-либо жило, что могло бы сравниться с этим существом [ спинозавром ]. Также моя гипотеза состоит в том, что Ти-рекс на самом деле был падальщиком, а не убийцей. Спинозавр на самом деле был хищным животным». [ 79 ] С тех пор он отказался от заявления о том, что Ти-рекс был мусорщиком. В фильме Спинозавр был изображен более крупным и могущественным, чем Тираннозавр : в сцене, изображающей битву между двумя воскресшими хищниками, Спинозавр выходит победителем, сломав Тираннозавру шею . [ 80 ] В четвертом фильме « Мир Юрского периода » есть отсылка к этому бою, где тираннозавр пробивает скелет спинозавра в решающей битве ближе к концу фильма. [ 81 ] Спинозавр появлялся во многих играх «Парка Юрского периода», особенно в «Jurassic World Evolution» и ее продолжении . Тот же спинозавр из третьего фильма возвращается в четвертом и пятом сезонах « Мира Юрского периода, Лагерь меловой период» , на этот раз сражаясь с двумя тираннозаврами . [ 82 ] [ 83 ]
Спинозавр издавна изображался в популярных книгах о динозаврах, хотя лишь в последнее время информации о спинозавридах стало достаточно для точного изображения. После влиятельной реконструкции скелета 1955 года, проведенной Лаппарентом и Лавокатом. [ 84 ] на основе диаграммы Стромера 1936 года, [ 85 ] его считали обобщенным прямоходящим тероподом с черепом, похожим на череп других крупных теропод, и парусом на спине, а также с четырехпалыми руками. [ 61 ]
Помимо фильмов, фигурок, видеоигр и книг, Спинозавр был изображен на почтовых марках таких стран, как Ангола , Гамбия и Танзания . [ 86 ] [ 87 ]
См. также
Ссылки
- ^ Перейти обратно: а б с д Смит, Роберт Ш.; Ибрагим, Низар; Мартилл, Дэвид М. (октябрь 2020 г.). « Сигилмассазавр - это спинозавр : переоценка африканских спинозавринов». Меловые исследования . 114 : 104520. Бибкод : 2020CrRes.11404520S . дои : 10.1016/j.cretres.2020.104520 . S2CID 219487346 .
- ^ Огюстен, Ф.Дж.; Хартунг, Дж.; Кампуридис, П. (2023). «Фауна динозавров Египта - Летопись наземных позвоночных позднего мела». В Хамими, З.; Хозем, Х.; Адатте, Т.; Надер, Фади Х.; Обох-Икуэнобе, Ф.; Зобба, Мохамед К.; Атфи, Хайтам Эль (ред.). Фанерозойская геология и природные ресурсы Египта . Достижения в области науки, технологий и инноваций. Спрингер, Чам. стр. 253–284. дои : 10.1007/978-3-030-95637-0_9 . ISBN 978-3-030-95636-3 .
- ^ Перейти обратно: а б с д Серено, Пол С.; Мирволд, Натан; Хендерсон, Дональд М.; Фиш, Фрэнк Э.; Видаль, Дэниел; Баумгарт, Стефани Л.; Кейллор, Тайлер М.; Формозо, Кирстен К.; Конрой, Лорен Л. (2022). «Спинозавр не является водным динозавром» . электронная жизнь . 11 . е80092. doi : 10.7554/eLife.80092 . ПМЦ 9711522 . ПМИД 36448670 .
 Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Молина-Перес и Ларраменди, 2016. Записи и курьезы динозавров, тероподов и других динозавроморфов , Ларусс. Барселона, Испания с. 259
- ^ Перейти обратно: а б с д и ж Смит, Дж. Б.; Ламанна, MC; Майр, Х.; и Лаковара, К.Дж. (2006). «Новая информация о голотипе Spinosaurus aegyptiacus Stromer, 1915». Журнал палеонтологии . 80 (2): 400–406. doi : 10.1666/0022-3360(2006)080[0400:NIRTHO]2.0.CO;2 . S2CID 130989487 .
- ^ Перейти обратно: а б с д Рассел, Д.А. (1996). «Изолированные кости динозавров из среднего мела Тафилальта, Марокко» . Бюллетень Национального музея естественной истории, Париж, раздел C. 4е серия. 18 (2–3): 349–402.
- ^ Перейти обратно: а б с д и Стромер, Э. (1915). «Результаты исследовательских поездок проф. Э. Стромера по пустыням Египта. II. Остатки позвоночных яруса Бахарие (нижний сеноман). 3. Оригинал теропода Spinosaurus aegyptiacus nov. gen., nov. spec» . Трактаты Королевской Баварской академии наук, математико-физический класс (на немецком языке). 28 (3): 1–32. [ постоянная мертвая ссылка ]
- ^ Стромер, Э. (1934). «Результаты исследовательских поездок проф. Э. Стромера по пустыням Египта. II. Остатки позвоночных яруса Бахарие (нижний сеноман). 13. Динозаврия». Трактаты Отделения математики и естественных наук Баварской академии наук . Новый эпизод (на немецком языке). 22 :1–79.
- ^ Перейти обратно: а б с Серено, ПК; Бек, Алабама; Дютей, Д.Б.; Гадо, Б.; Ларссон, ХК; Лион, GH; Маркот, доктор юридических наук; Раухут, OWM; Садлер, RW; Сидор, Калифорния; Варриккио, диджей; Уилсон, врач общей практики; Уилсон, Дж. А. (1998). «Длиннорылый хищный динозавр из Африки и эволюция спинозаврид» . Наука . 282 (5392): 1298–1302. Бибкод : 1998Sci...282.1298S . дои : 10.1126/science.282.5392.1298 . ПМИД 9812890 .
- ^ Перейти обратно: а б Хольц, Томас Р.; Мольнар, Ральф Э.; Карри, Филип Дж. (2004). «Базальная столбняк». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. стр. 71–110. ISBN 978-0-520-24209-8 . JSTOR 10.1525/j.ctt1pn61w.10 .
- ^ Малер, Люк (11 марта 2005 г.). «Запись об Abelisauridae (Dinosauria: Theropoda) из сеномана Марокко». Журнал палеонтологии позвоночных . 25 (1): 236–239. doi : 10.1671/0272-4634(2005)025[0236:ROADTF]2.0.CO;2 . S2CID 4974444 .
- ^ Перейти обратно: а б Хасэгава, Ю.; Танака, Г.; Такакува, Ю.; Койке, С. (2010). «Прекрасные скульптуры на зубе спинозавра (Dinosauria, Theropoda) из Марокко» (PDF) . Вестник Музея естественной истории Гумма . 14 :11–20. Архивировано из оригинала (PDF) 25 марта 2012 года . Проверено 11 сентября 2010 г.
- ^ Перейти обратно: а б с д и ж г час я дж даль Сассо, К.; Магануко, С.; Бюффето, Э.; Мендес, Массачусетс (2005). «Новая информация о черепе загадочного теропода спинозавра с замечаниями о его размерах и сходстве». Журнал палеонтологии позвоночных . 25 (4): 888–896. doi : 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2 . S2CID 85702490 .
- ^ Перейти обратно: а б Баффето, Эрик; Уаджа, Мохамед (1 сентября 2002 г.). «Новый экземпляр спинозавра (Dinosauria, Theropoda) из нижнего мела Туниса с замечаниями по эволюционной истории спинозавридов» (PDF) . Бюллетень геологического общества Франции . 173 (5): 415–421. дои : 10.2113/173.5.415 . HDL : 2042/216 .
- ^ Перейти обратно: а б Раухут, OWM (2003). Взаимоотношения и эволюция базальных теропод динозавров . Специальные статьи по палеонтологии. Том. 69. стр. 1–213. ISBN 978-0-901702-79-1 .
- ^ Таке, П.; и Рассел, Д.А. (1998). «Новые данные о динозаврах-спинозавридах раннего мела Сахары» (PDF) . Comptes Rendus de l'Académie des Sciences, Série IIA . 327 (5): 347–353. Бибкод : 1998CRASE.327..347T . дои : 10.1016/S1251-8050(98)80054-2 . Проверено 22 сентября 2010 г.
- ^ Перейти обратно: а б Ибрагим, Н.; Серено, ПК; Варрачио, диджей; Мартилл, DM; Анвин, DM; Байддер, Л.; Ларссон, HCE; Зухри, С.; Каукайя, У. (2020). «Геология и палеонтология верхнемеловой Кем-Кемской группы восточного Марокко» . ZooKeys (928): 1–216. Бибкод : 2020ЗооК..928....1И . дои : 10.3897/zookeys.928.47517 . ПМЦ 7188693 . ПМИД 32362741 .
- ^ Перейти обратно: а б с д и ж г час Арден, ТМС; Кляйн, КГ; Зухри, С.; Лонгрич, Северная Каролина (2018). «Водная адаптация черепа хищных динозавров (Theropoda: Spinosauridae) и эволюция водных привычек у спинозавров ». Меловые исследования . 93 : 275–284. дои : 10.1016/j.cretres.2018.06.013 . S2CID 134735938 .
- ^ Перейти обратно: а б с д и ж г час Ибрагим, Низар; Серено, Пол С.; Даль Сассо, Криштиану; Магануко, Симоне; Фаббри, Маттео; Мартилл, Дэвид М.; Зухри, Самир; Мирволд, Натан; Юрино, Давид А. (26 сентября 2014 г.). «Полуводные адаптации гигантского хищного динозавра» . Наука . 345 (6204): 1613–1616. Бибкод : 2014Sci...345.1613I . дои : 10.1126/science.1258750 . ПМИД 25213375 . S2CID 34421257 . Дополнительная информация
- ^ Перейти обратно: а б с Эверс, Юго-Запад; Раухут, OWM; Милнер, AC; Макфитерс, Б.; Аллен, Р. (2015). «Переоценка морфологии и систематического положения тероподного динозавра Sigilmassasaurus из «среднего» мела Марокко» . ПерДж . 3 : е1323. дои : 10.7717/peerj.1323 . ПМЦ 4614847 . ПМИД 26500829 .
- ^ Доктор философии, Андреа Кау (20 сентября 2014 г.). «Теропода: Революция спинозавров, Эпизод IV: Решение всех загадок?» . Проверено 20 декабря 2019 г.
- ^ Доктор философии, Андреа Кау (21 сентября 2014 г.). «Теропода: Революция спинозавров, Эпизод V: Сигилмассазавр против спинозавра: тафономическая битва» . Получено 20 . декабря
- ^ Мортимер, Микки (11 сентября 2014 г.). «Блог базы данных Theropod: сюрприз спинозавра» . Проверено 20 декабря 2019 г.
- ^ Бюффето, Э.; Дофин, Ю.; Йегер, Ж.-Ж.; Мартин, М.; Мазин, Ж.-М.; и Тонг, Х. (1986). «Призматическая зубная эмаль тероподовых динозавров». естественные науки . 73 (6): 326–327. Бибкод : 1986NW.....73..326B . дои : 10.1007/BF00451481 . ПМИД 3748191 . S2CID 11701295 .
- ^ Перейти обратно: а б Баффето, Э. (1989). «Новые останки загадочного динозавра спинозавра из мелового периода Марокко и сходство между спинозавром и бариониксом ». Новогодний ежегодник по геологии и палеонтологии, ежемесячные выпуски . 1989 (2): 79–87. дои : 10.1127/njgpm/1989/1989/79 .
- ^ Баффето, Э. (1992). «Заметки о меловых динозаврах-тероподах спинозаврах и бариониксах ». Новогодний ежегодник по геологии и палеонтологии, ежемесячные выпуски . 1992 (2): 88-96. дои : 10.1127/njgpm/1992/1992/88 .
- ^ Келлнер, AWA; и Мэдер, Б.Дж. (1997). «Зубы архозавра из мелового периода Марокко». Журнал палеонтологии . 71 (3): 525–527. Бибкод : 1997JPal...71..525K . дои : 10.1017/S0022336000039548 . JSTOR 1306632 . S2CID 132508192 .
- ^ Бентон, MJ; Буазиз, С.; Бюффето, Э.; Мартилль, Д.; Уаха, М.; Сусси, М.; и Труман, К. (2000). «Динозавры и другие ископаемые позвоночные животные из речных отложений нижнего мела на юге Туниса» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 157 (3–4): 227–246. Бибкод : 2000PPP...157..227B . дои : 10.1016/S0031-0182(99)00167-4 .
- ^ Брусатте, Стивен Л.; Серено, Пол К. (12 декабря 2007 г.). «Новый вид Carcharodontosaurus (Dinosauria: Theropoda) из сеномана Нигера и ревизия рода». Журнал палеонтологии позвоночных . 27 (4): 902–916. doi : 10.1671/0272-4634(2007)27[902:ANSOCD]2.0.CO;2 . S2CID 86202969 .
- ^ Перейти обратно: а б Хендрикс, К.; Матеус, О.; Баффето, Э. (2016). «Морфофункциональный анализ квадрата спинозавридов (Dinosauria: Theropoda) и наличие спинозавров и второго таксона спинозавров в сеномане Северной Африки» . ПЛОС ОДИН . 11 (1): e0144695. Бибкод : 2016PLoSO..1144695H . дои : 10.1371/journal.pone.0144695 . ПМК 4703214 . ПМИД 26734729 .
- ^ Вейшампель, Дэвид Б.; Барретт, Пол М.; Кориа, Родольфо А.; Ле Лефф, Жан; Син, Сюй; Сицзинь, Чжао; Санхи, Ашок; Гомани, член парламента Элизабет; Ното, Кристофер Р. (2004). «Распространение динозавров» В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. стр. 100-1 517–606. ISBN 978-0-520-24209-8 . JSTOR 10.1525/j.ctt1pn61w.31 .
- ^ « Речное чудовище: 50-футовый спинозавр» . Архивировано из оригинала 13 сентября 2014 г. – на сайте video.nationalgeographic.com.
- ^ Хюне, Фридрих фон (1926). «Плотоядные ящерицы в юрских и меловых формациях, главным образом в Европе» . Ревиста Музея Ла-Платы . 29 :35–167.
- ^ Перейти обратно: а б Перенасыщение, Д.Ф. (1982). Новый словарь динозавров . Секаукус, Нью-Джерси: Citadel Press. стр. 226–228 . ISBN 978-0-8065-0782-8 .
- ^ Перейти обратно: а б Пол, GS (1988). «Семейство спинозавриды» . Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 271–274 . ISBN 978-0-671-61946-6 .
- ^ Перейти обратно: а б с Терриен, Ф.; Хендерсон, DM (2007). «Мой теропод больше вашего… или нет: оцениваем размер тела теропод по длине черепа». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . S2CID 86025320 .
- ^ Мортимер, М. (25 марта 2007 г.). «Комментарии к новой статье Терриена и Хендерсона» . Список рассылки динозавров. Архивировано из оригинала 11 октября 2007 года . Проверено 22 сентября 2010 г.
- ^ Харрис, доктор юридических наук (26 марта 2007 г.). «Re: Комментарии к новой статье Терриена и Хендерсона» . Список рассылки динозавров. Архивировано из оригинала 14 апреля 2011 года . Проверено 22 сентября 2010 г.
- ^ Молина-Перес и Ларраменди, 2016. Записи и курьезы динозавров, тероподов и других динозавроморфов , Ларусс. Барселона, Испания с. 142
- ^ Хоун, Дэвид Уильям Эллиотт; Хольц, Томас Ричард (июнь 2017 г.). «Век спинозавров - обзор и пересмотр спинозавридов с комментариями по их экологии» . Acta Geologica Sinica — английское издание . 91 (3): 1120–1132. Бибкод : 2017AcGlS..91.1120H . дои : 10.1111/1755-6724.13328 . S2CID 90952478 .
- ^ Перейти обратно: а б с д и ж г час Бейли, Джек Боуман (1997). «Удлинение нервного отдела позвоночника у динозавров: парусники или буйволы?». Журнал палеонтологии . 71 (6): 1124–1146. Бибкод : 1997JPal...71.1124B . дои : 10.1017/S0022336000036076 . JSTOR 1306608 . S2CID 130861276 . ПроКвест 231041667 .
- ^ Манн, Арьян; Рейс, Роберт Р. (2020). «Древность гиперудлинения нервного отдела позвоночника с парусом у предшественников млекопитающих» . Границы в науках о Земле . 8 : 83. Бибкод : 2020FrEaS...8...83M . дои : 10.3389/feart.2020.00083 .
- ^ Блэк, Райли (6 июня 2011 г.). «Был ли спинозавр динозавром со спиной бизона?» . Смитсоновский журнал .
- ^ Перейти обратно: а б Ибрагим, Низар; Магануко, Симоне; Даль Сассо, Криштиану; Фаббри, Маттео; Аудиторе, Марко; Бинделлини, Габриэле; Мартилл, Дэвид М.; Зухри, Самир; Маттарелли, Диего А.; Анвин, Дэвид М.; Виманн, Жасмин; Бонадонна, Давиде; Амане, Аюб; Якубчак, Юлиана; Джогер, Ульрих; Лаудер, Джордж В.; Пирс, Стефани Э. (7 мая 2020 г.). «Водное передвижение тероподового динозавра с помощью хвоста» . Природы . 581 (7806): 67–70. Бибкод : 2020Natur.581...67I . дои : 10.1038/s41586-020-2190-3 . ПМИД 32376955 . S2CID 216650535 .
- ^ Продажи, МАФ; Шульц, CL (2017). «Таксономия спинозавров и эволюция краниодентальных особенностей: данные из Бразилии» . ПЛОС ОДИН . 12 (11): e0187070. Бибкод : 2017PLoSO..1287070S . дои : 10.1371/journal.pone.0187070 . ПМЦ 5673194 . ПМИД 29107966 .
- ^ Перейти обратно: а б Холстед, Л.Б. (1975). Эволюция и экология динозавров . Лондон: Eurobook Limited. стр. 1–116. ISBN 978-0-85654-018-9 .
- ^ Перейти обратно: а б с д Гимса, Ян; Сани, Роберт; Гимса, Ульрика (май 2016 г.). «Загадка спинного паруса Spinosaurus aegyptiacus» . Геологический журнал . 153 (3): 544–547. Бибкод : 2016ГеоМ..153..544Г . дои : 10.1017/S0016756815000801 . S2CID 51999370 .
- ^ Перейти обратно: а б Чариг, Эй Джей; Милнер, AC (1997). « Baryonyx walkeri , рыбоядный динозавр из Уилдена в графстве Суррей». Вестник Музея естественной истории, серия «Геология» . 53 : 11–70.
- ^ Бюффето, Э.; Мартилль, Д.; Эскуилье, Ф. (2004). «Птерозавры как часть диеты спинозавров» . Природа . 430 (6995): 33. Бибкод : 2004Natur.429...33B . дои : 10.1038/430033а . ПМИД 15229562 . S2CID 4398855 .
- ^ Даль Сассо, К.; Магануко, С.; Чоффи, А. (26 мая 2009 г.). «Нервно-сосудистая полость на морде хищного динозавра спинозавра » (PDF) . 1-й Международный конгресс по палеонтологии позвоночных Северной Африки . Национальный музей естественной истории. Архивировано из оригинала (PDF) 21 июля 2011 года . Проверено 22 сентября 2010 г.
- ^ Манжета, Арканзас; Рэйфилд, Э.Дж. (2013). Фарке, Эндрю А. (ред.). «Механика питания спинозавровых теропод и современных крокодилов» . ПЛОС ОДИН . 8 (5): e65295. Бибкод : 2013PLoSO...865295C . дои : 10.1371/journal.pone.0065295 . ПМЦ 3665537 . ПМИД 23724135 .
- ^ Сакамото, Манабу (2022 г.). «Оценка силы укуса у вымерших динозавров с использованием филогенетически предсказанных физиологических площадей поперечного сечения приводящих мышц челюсти» . ПерДж . 10 : е13731. дои : 10.7717/peerj.13731 . ПМЦ 9285543 . ПМИД 35846881 .
- ^ Амиот, Р.; Бюффето, Э.; Лекюйер, К.; Ван, X.; Будад, Л.; Дин, З.; Фурель, Ф.; Хатт, С.; Мартино, Ф.; Медейрос, А.; Мо, Дж.; Саймон, Л.; Сутитхорн, В.; Свитман, С.; Тонг, Х.; Чжан, Ф.; Чжоу, З. (2010). «Изотопы кислорода свидетельствуют о полуводном образе жизни спинозавридных теропод». Геология . 38 (2): 139–142. Бибкод : 2010Geo....38..139A . дои : 10.1130/G30402.1 .
- ^ Палеонтология, Королевский Тиррелловский музей (16 августа 2018 г.). «Новое исследование опровергает утверждения о том, что спинозавр был полуводным человеком» . WordPress.com .
- ^ Перейти обратно: а б с Хендерсон, DM (2018). «Вызов плавучести, равновесия и устойчивости гипотезе полуводного спинозавра Стромера, 1915 (Dinosauria: Theropoda)» . ПерДж . 6 : е5409. дои : 10.7717/peerj.5409 . ПМК 6098948 . ПМИД 30128195 .
- ^ Отточить; Хольц, Томас Р. младший (2021). «Оценка экологии спинозавра: специалист по береговой линии или специалист по водным преследователям?» . Электронная палеонтология . 24 (1): а03. дои : 10.26879/1110 . hdl : 1903/28570 .
- ^ Фаббри, Маттео; Навалон, Гильермо; Бенсон, Роджер Б.Дж.; Пол, Диего; О'Коннор, Цзинмай; Бхуллар, Бхарт-Анджан С.; Эриксон, Грегори М.; Норелл, Марк А.; Оркнейские острова, Эндрю; Ламанна, Мэтью С.; Зухри, Самир; Беккер, Жюстин; Эмке, Аманда; Даль Сассо, Криштиану; Бинделлини, Габриэле; Магануко, Симоне; Аудиторе, Марко; Ибрагим, Низар (23 марта 2022 г.). «Подводное кормодобывание среди хищных динозавров» . Природы . 603 (7903): 852–857. Бибкод : 2022Natur.603..852F . дои : 10.1038/s41586-022-04528-0 . ПМИД 35322229 . S2CID 247630374 .
- ^ Грешко Михаил (23 марта 2022 г.). «У спинозавра были кости, похожие на пингвиньи, что являлось признаком подводной охоты» . Нэшнл Географик . Архивировано из оригинала 23 марта 2022 года.
- ^ Полевой музей (23 марта 2022 г.). «Плотные кости позволяли спинозавру охотиться под водой, как показывают исследования» . физ.орг .
- ^ «Когда динозавры правили разумом № 8: Динозавры на протяжении многих лет: Спинозавр» . Когда динозавры правили разумом . 25 августа 2014 года . Проверено 30 апреля 2020 г.
- ^ Перейти обратно: а б с Перенасыщение, Д.Ф. (2000). «Спинозавр». Динозавры: Энциклопедия. 1-е дополнение . Джефферсон, Северная Каролина: McFarland & Company, Inc., стр. 329–333. ISBN 978-0-7864-0591-6 .
- ^ Перейти обратно: а б Карпентер, К. (2002). «Биомеханика передних конечностей нептичьих динозавров-теропод в условиях хищничества». Сенкенбергиана Летея . 82 (1): 59–76. дои : 10.1007/BF03043773 . S2CID 84702973 .
- ^ Милнер, Арканзас; Харрис, доктор юридических наук; Локли, МГ; Киркланд, Дж.И.; Мэтьюз, Северная Каролина (2009). Харпендидинг, Генри (ред.). «Анатомия, поза и поведение птиц, выявленные по следам покоящегося динозавра-теропода ранней юры» . ПЛОС ОДИН . 4 (3): e4591. Бибкод : 2009PLoSO...4.4591M . дои : 10.1371/journal.pone.0004591 . ПМЦ 2645690 . ПМИД 19259260 .
- ^ Витце, А. (2014). «Плавающий динозавр найден в Марокко». Природа . дои : 10.1038/nature.2014.15901 . S2CID 131487243 .
- ^ Скотт Хартман (12 сентября 2014 г.). «В спинозавре есть что-то подозрительное» . Skeletaldrawing.com . Проверено 20 сентября 2014 г.
- ^ Марк Уиттон (22 сентября 2014 г.). «Спор о задних конечностях спинозавра: подробный ответ авторов» . markwitton-com.blogspot.de . Проверено 22 сентября 2014 г.
- ^ Мирволд, Натан П.; Баумгарт, Стефани Л.; Видаль, Дэниел; Фиш, Фрэнк Э.; Хендерсон, Дональд М.; Сайтта, Эван Т.; Серено, Пол К. (2024). «Ныряющие динозавры? Предостережения относительно использования компактности костей и pFDA для определения образа жизни» . ПЛОС ОДИН . 19 (3): e0298957. Бибкод : 2024PLoSO..1998957M . дои : 10.1371/journal.pone.0298957 . ПМЦ 10917332 . ПМИД 38446841 .
- ^ Чанг, Кеннет (6 марта 2024 г.). «Плавающий динозавр? Возможно, нет, говорится в исследовании. Новая статья бросает вызов идее о том, что большой плотоядный спинозавр нырял за добычей, а не шел вброд и выдергивал ее из воды» . Нью-Йорк Таймс . Архивировано из оригинала 6 марта 2024 года . Проверено 8 марта 2024 г.
- ^ Магануко, Симоне; Даль Сассо, Криштиану (30 мая 2018 г.). «Самый маленький и самый большой динозавр-теропод: крошечный когтевой ноготь молодого спинозавра из мелового периода Марокко» . ПерДж . 6 : е4785. дои : 10.7717/peerj.4785 . ПМЦ 5984586 . ПМИД 29868253 .
- ^ Смит, Рой Э.; Мартилл, Дэвид М. (июнь 2023 г.). «Необычная стоматологическая патология в зубе спинозавра (Dinosauria, Theropoda) из среднего мела Марокко» . Меловые исследования . 146 : 105499. Бибкод : 2023CrRes.14605499S . дои : 10.1016/j.cretres.2023.105499 . Получено 6 мая 2024 г. - через Elsevier Science Direct.
- ^ Градштейн, FM; Огг, Дж.Г.; Смит, А.Г., ред. (2004). Геологическая шкала времени, 2004 г. Кембридж и Нью-Йорк: Издательство Кембриджского университета. п. 380. ИСБН 978-0-521-78673-7 .
- ^ Хольц, Томас Р. младший (2012) Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов, зима 2011 г., Приложение.
- ^ Черчер, К.С.; Де Юлиис, Г (2001). «Новый вид Protopterus и ревизия Ceratodus humei (Dipnoi: Ceratodontiformes) из позднемеловой мутной формации восточного оазиса Дахле, Западная пустыня Египта» . Палеонтология . 44 (2): 305–323. Бибкод : 2001Palgy..44..305C . дои : 10.1111/1475-4983.00181 .
- ^ «Утес Двоякодышащей рыбы, Вади-эль-Баттих (Мел Египта)» . ПБДБ .
- ^ Серено, ПК; Дютейл, Д.Б.; Ярошене, М; Ларссон, HCE; Лион, GH; Магвене, премьер-министр; Сидор, Калифорния; Варриккио, диджей; Уилсон, Дж. А. (1996). «Хищные динозавры Сахары и дифференциация фауны позднего мела» (PDF) . Наука . 272 (5264): 986–991. Бибкод : 1996Sci...272..986S . дои : 10.1126/science.272.5264.986 . ПМИД 8662584 . S2CID 39658297 .
- ^ Смит, Дж. Б.; Ламанна, MC; Лаковара, К.Дж.; Додсон, П.; Смит, младший; Пул, Джей Си; Гигенгак, Р.; Аттиа, Ю. (2001). «Гигантский динозавр-зауропод из мангровых отложений верхнего мела в Египте» (PDF) . Наука . 292 (5522): 1704–1706. Бибкод : 2001Sci...292.1704S . дои : 10.1126/science.1060561 . ПМИД 11387472 . S2CID 33454060 .
- ^ Нэйш, Даррен (2012). Планета динозавров: следующее поколение гигантов-убийц . Книги Светлячка. стр. 80–85 . ISBN 978-1-77085-049-1 .
- ^ Элли, Дерек (17 июля 2001 г.). «Парк Юрского периода III» . Разнообразие . Проверено 9 июля 2007 г.
- ^ Портман, Дж. (11 июля 2001 г.). «Спинозавр делает тираннозавра похожим на кошечку: когда дело доходит до «Парка Юрского периода III», размер имеет значение». Гражданин Оттавы .
- ^ Чендлер, Г. (август 2001 г.). «Краткий путеводитель по динозаврам из нового фильма «Парк Юрского периода III». Национальный географический мир .
- ^ Романо, Ник (июнь 2015 г.). «Мир Юрского периода пробрался в сладкий реверанс в сторону Парка Юрского периода 3» . СинемаБленд .
- ^ Оркиола, Джон (23 июля 2022 г.). «В мире юрского периода наконец-то состоялся достойный матч-реванш тираннозавра против спинозавра» . Экранная ругань . Проверено 7 февраля 2024 г.
- ^ Матадин, Ренальдо (9 декабря 2021 г.). «Мир юрского периода: лагерь меловых отходов самого разрушительного динозавра» . CBR.com . Проверено 7 февраля 2024 г.
- ^ Лаппарент, AF de; и Лавокат Р. (1955). «Динозавры». В: Пивето, Ж., редактор. Трактат по палеонтологии. Том V. Выход вод. Рождение Тетраподии. Буйство растительной жизни. Покорение воздуха. Земноводные. Рептилии. Птицы. Париж: Masson et Cie, стр. 785–962.
- ^ Стромер, Э. (1936). «Результаты исследовательских поездок профессора Э. Стромера по пустыням Египта. VII. Котел Бахарие и сцена с их фауной и флорой. Дополнительное резюме». Трактаты Баварской академии наук, отделение математики и естественных наук . Новый эпизод (на немецком языке). 33 :1–102.
- ^ Хатри, В.С. (9 июня 2006 г.). «Из прошлого» . Индуист . Архивировано из оригинала 18 июня 2006 года . Проверено 12 сентября 2010 г.
- ^ Перенасыщение, DF; и Бретт-Сурман, МК (2000). «Динозавры и СМИ» (PDF) . В Фарлоу, Джо; и Уолтерс, Р.Х. (ред.). Полный динозавр . Блумингтон, Индиана: Издательство Университета Индианы. стр. 673–706. ISBN 978-0-253-21313-6 . Архивировано из оригинала (PDF) 27 марта 2012 года . Проверено 12 сентября 2010 г.
Дальнейшее чтение
- Перенасыщение Д.Ф. «В поисках спинозавра ». В: Юрская классика: сборник очерков ящеров и мезозойских размышлений , стр. 77–85. Джефферсон, Северная Каролина: МакФарланд, 2001. ISBN 0-7864-0961-4 .
- Нотдерфт, В.; и Смит Дж. Затерянные динозавры Египта. Нью-Йорк: Рэндом Хаус, 2002. ISBN 0-375-50795-7 .
- Дань уважения Эрнсту Штромеру: сто лет открытия Spinosaurus aegyptiacus : Саубхик Гош
Внешние ссылки
- «Странный динозавр, возможно, переплывал реки Африки» . Профиль спинозавра, автор Кеннет Чанг в NY Times, 29 апреля 2020 г.
- Хартман, Скотт. Сравнение спинозавров. SkeletalDrawing.com , 2006.
- Мортимер, Микки. Спинозавр Стромер, 1915 г. (Список экземпляров из базы данных Theropod).
- Музей естественной истории . Каталог динозавров: Спинозавр.
- Прендергаст, Джон. Потерянные и найденные динозавры. The Pennsylvania Gazette, журнал выпускников Пенсильванского университета , июль/август 2001 г.















- Спинозавриды
- Раннемеловые динозавры Африки.
- Позднемеловые динозавры Африки.
- Первые появления рода Albian
- Сеноманское вымирание родов
- Окаменелости Алжира
- Окаменелости Египта
- Формация Бахария
- Окаменелости Марокко
- Окаменелости Туниса
- Ископаемые таксоны описаны в 1915 году.
- Таксоны, названные Эрнстом Стромером
- Полуводные животные
- неологизмы 1910-х годов