Baryonyx
| Baryonyx Временный диапазон: Барреман ,
| |
|---|---|

| |
| Реконструированная скелетная гора в Национальном музее природы и науки , Токио | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | Сауриския |
| Клада : | Теропода |
| Семья: | † Spinosauridae |
| Подсемейство: | † Baryonychinae |
| Род: | † Baryonyx Charig & Milner , 1986 |
| Разновидность: | † Б. Уокер
|
| Биномиальное название | |
| † Baryonyx Walker Charig & Milner, 1986
| |
Baryonyx ( / ˌ B ær i ˈ ɒ n ɪ k s / ) является родом динозавра теропода , который жил на барремйской стадии , раннего мела около 130–125 миллионов лет назад. Первый скелет был обнаружен в 1983 году в глиной яме Smokejack , Суррея , Англия, в отложениях глинистой формирования Weald и стал голотипом Baryonyx Walkeri , названным палеонтологами Аланом Дж. Чаригом и Анжелой С. Милнер в 1986 году . Общее название, Baryonyx , означает «тяжелый коготь» и намекает на очень большой когтя животного на первом пальце; Конкретное имя , Уокери , относится к своему открытию, любительскому коллектору ископаемых Уильяма Дж. Уокера. Образец голотипа является одним из самых полных скелетов в области теропода из Великобритании (и остается наиболее полным спинозавридом ), и его открытие привлекло внимание средств массовой информации. Образцы, позже обнаруженные в других частях Соединенного Королевства и Иберии, также были назначены роду, хотя многие с тех пор были перемещены в новые роды.
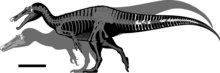
Образец голотипа, который, возможно, не был полностью выращен, был оценен в длину от 7,5 до 10 метров (25 и 33 фута) и весил от 1,2 до 2 метрических тонн (1,3 и 2,2 тонны ; 1,2 и 2,0 длиной тонны ). У Baryonyx была длинная, низкая и узкая морда, которая сравнивалась с гариалом . Кончик морды расширился до сторон в форме розетки . За этим верхняя челюсть имела выемку, которая была установлена в нижней челюсти (которая изогнулась вверх в той же области). У него был треугольный гребень на вершине носовых костей . У Baryonyx было большое количество тонко зубчатых , конических зубов, с самыми большими зубами впереди. Шея образовала S-образную форму, а нервные шипы дорсальных позвонков увеличились по высоте спереди к спине. Один удлиненный нейронный позвоночник указывает на то, что у него, возможно, был горб или гребень вдоль центра спины. У него были надежные передние конечности, с одноименным когтями первого пальца длиной около 31 сантиметров (12 дюймов).
В настоящее время признанный членом семейства Spinosauridae , Baryonyx аффинность была неясной, когда она была обнаружена. Некоторые исследователи предположили, что Suchosaurus cultridens является старшим синонимом (будучи более старым именем), и что Sucyomimus tenerensis принадлежит к одному роду; Последующие авторы держали их отдельными. Baryonyx был первым динозавром Theropod, который, как показали, был писцидным (поедание рыбы), о чем свидетельствуют рыбные масштабы в области желудка образца голотипа. Это также мог быть активным хищником более крупной добычи и мусорщика , поскольку в нем также содержались кости ювенильного игуанодонтида . Существо поймало бы и обработало бы свою добычу в первую очередь с помощью передних конечностей и больших когтей. У Baryonyx , возможно, были полуакотические привычки и сосуществовали с другими динозаврами Theropod, Ornithopod и Sauropod , а также с птерозаврами , крокодилами , черепахами и рыбами в речной среде.
История открытия
[ редактировать ]
сантехника и любителя В январе 1983 года сборщик ископаемых Уильям Дж. Уокер исследовал глиняную яму Smokejack , глиняную яму в форме глины Weald возле Окли в Суррее, Англия. Он нашел камень, в котором он обнаружил большой когти, но после того, как он собрал его дома, он понял, что кончик коготь отсутствовал. Уокер вернулся в то же место в яме через несколько недель и нашел недостающую часть после поиска часа. Он также нашел кость фаланги и часть ребра . Зять Уолкера позже принес Коготь в Лондонский музей естественной истории , где он был рассмотрен палеонтологами Аланом Дж. Чаригом и Анжелой С. Милнер , которые определили его как принадлежащую к Теропода динозавру . [ 1 ] [ 2 ] Палеонтологи обнаружили больше фрагментов кости в этом месте в феврале, но весь скелет не мог быть собран до мая и июня из -за погодных условий в яме. [ 3 ] [ 2 ] Команда из восьми сотрудников музея и нескольких добровольцев выкопала 2 метрических тонн (2,2 коротких тонн ; 2,0 тонны ) каменной матрицы в 54 блоках в течение трехнедельного периода. Уокер пожертвовал когти в музей, а компания Ockley Brick Company (владельцы ямы) пожертвовала остальную часть скелета и предоставила оборудование. [ 3 ] [ 4 ] [ 2 ] Область была исследована в течение 200 лет, но ранее не было обнаружено подобных останков. [ 5 ] [ 2 ]
Большинство собравшихся костей были заключены в алочные узелки, окруженные мелким песком и илом, а остальные лежали в глине. Кости были рассеяны и разбросаны по районе 5 на 2 метра (16,4 на 6,6 фута), но большинство из них были недалеко от их естественных позиций. Положение некоторых костей было нарушено бульдозером , а некоторые были нарушены механическим оборудованием до того, как они были собраны. [ 3 ] [ 1 ] [ 6 ] Подготовка образца была затруднена из -за твердости матрицы алеза и присутствия сидерита ; Кислотная препарат был предпринят, но большая часть матрицы была удалена механически. Потребовалось шесть лет почти постоянной подготовки, чтобы вытащить все кости из скалы, и к концу зубные инструменты и воздушные молотки должны были использоваться под микроскопом. Образец представляет около 65 процентов скелета и состоит из частичных костей черепа, в том числе преднасиллы (первые кости верхней челюсти); левые челюсти (вторая кость верхней челюсти); оба носовые кости; левый ласковый ; левый префронтальный ; левый посторбитал ; мозговой пропуск, включая затылку ; Оба зубные зубные данные (передние кости нижней челюсти); различные кости от задней части нижней челюсти; зубы; шейки матки (шея), дорсальный (спина) и каудальные (хвостовые) позвонки ; ребра; грудина ; Оба лопатки (лопатки); оба коракоида ; Оба гуммери (кости верхней части руки); левый радиус и локтевая кости (кости нижней руки); кости пальцев и Unguals (кости когтей); модные кости; Верхний конец левой бедренной кости (бедра) и нижний конец правого; правая малоберцовая кость (из нижней ноги); и кости ноги, включая Ungual. [ 3 ] [ 1 ] [ 7 ] [ 2 ] Первоначальный номер образца был BMNH R9951, но позже он был повторно рассмотрен как NHMUK PV R9951. [ 1 ] [ 8 ]



В 1986 году Чариг и Милнер назвали новый род и виды со скелетом в качестве образца голотипа : Baryonyx Walkeri . Общее название происходит от древнегреческого ; βαρύς ( barys ) означает «тяжелый» или «сильный», а ὄνυξ ( Onyx ) означает «когтя» или «ког». Конкретное название чествует Уокера, чтобы открыть образец. большой ког В то время авторы не знали , принадлежал ли [ 9 ] ) Динозавр был представлен ранее в том же году во время лекции на конференции о систематике динозавров в Драмхеллере , Канада. Из -за продолжающейся работы над костями (в то время было подготовлено 70 процентов), они назвали свою статью предварительной и пообещали более подробное описание позднее. Baryonyx был первым крупным ранним меловым тероподом, найденным в любом месте мира к тому времени. [ 1 ] [ 6 ] Перед открытием Baryonyx последней значимой находкой теропод в Соединенном Королевстве была Eustreptospondylus в 1871 году, а в интервью 1986 года Хариг под названием Baryonyx «Лучшая находка века» в Европе. [ 3 ] [ 4 ] Baryonyx был широко распространен в международных СМИ и был назван «Когтями» журналистами, наказаниями на название фильма « Челюсти » . Его открытие было предметом документального фильма BBC 1987 года , а актерский состав скелета установлен в Музее естественной истории в Лондоне. В 1997 году Чариг и Милнер опубликовали монографию, в которой подробно описывается голотип скелет. [ 3 ] [ 5 ] [ 10 ] Образец голотипа остается наиболее известным скелетом спинозауридов . [ 11 ]
Назначенные образцы
[ редактировать ]Окаменелости из других частей Великобритании и Иберии , в основном изолированных зубов, впоследствии были связаны с Baryonyx или подобными животными. [ 3 ] Изолированные зубы и кости с острова Уайт , в том числе ручные кости, зарегистрированные в 1998 году, и позвонка, о котором сообщили палеонтологи Стива Хатта и Пенни Ньюбери в 2004 году, были приписаны этому роду. [ 12 ] Фрагмент верхней челюсти из Ла -Риоджи, Испания , был приписан Барионекс палеонтологами Луисом И. Виерой и Хосе Ангелом Торресом в 1995 году [ 13 ] (Хотя палеонтолог Томас Р. Хольц и его коллеги повысили вероятность того, что он мог бы принадлежать Сучимусу в 2004 году). [ 14 ] В 1999 году посторбитальный, сквомольный , зубной, позвоночные останки, ручные кости) и фаланга от депозита Салас -де -Лос в провинции Бургос , Испания метакарпалы ( Голотип) от палеонтолога Каролины Фуэнтес Видарте и его коллег. [ 15 ] [ 16 ] треки динозавров Также было предложено, что возле Бургоса принадлежат Baryonyx или аналогичному тероподу. [ 17 ]

В 2011 году образец (каталогизировался как ML1190 в музее da lourinhã ) из формирования Papo Seco в Бока -до Чапиме , Португалия, с фрагментарной зубной зубной зоной, зубами, позвонками, ребрами, костями бедра, лопат Baryonyx и коллег, наиболее полных иберийских от палеонтолога Octávio Mateus остатков животного. Скелетные элементы этого образца также представлены в более полном голотипе (который был одинаковый размер), за исключением позвонков в середине шея. [ 18 ] В 2018 году палеонтолог Томас г -жа Арден и его коллеги обнаружили, что португальский скелет не принадлежал Baryonyx , поскольку передняя часть его зубной кости не была сильно перевернута. [ 19 ] Это испанцы [ 20 ] Многочисленные исследования показали, что дополнительные остатки спинозауридов от Иберии могут принадлежать к таксонам , отличным от Baryonyx , таких как Vallibonavenatrix и Protathlitis , или может быть неопределенным. [ 21 ] [ 22 ] [ 23 ] [ 24 ] В статье 2024 года палеонтолога Эрика Исасменди и его коллег рассмотрены спинозауридные ископаемые записи Иберии и пришли к выводу, что ни один из образцов оттуда не может быть назначен Baryonyx . Они перенесли образец, ранее назначенный в Baryonyx из La Rioja в новый род Riojavenatrix . [ 25 ]
В 2021 году палеонтолог Крис Т. Баркер и его коллеги описали два новых рода спинозауридов из формирования Уэссекса острова Уайт, Цератосучопов и Рипироввестита (последний по имени Р. Милнере в честь Милнера за ее вклад в исследования спинозавридов) и заявил, что спинозаврид Материал оттуда, который ранее был приписан современному Baryonyx, мог бы принадлежать к другим таксонам. [ 26 ] Эти образцы ранее были назначены в Baryonyx в аннотации конференции 2017 года. [ 27 ] Баркер и его коллеги заявили, что распознавание образцов формирования Wessex в качестве новых родов делает присутствие Baryonyx там неоднозначным, и поэтому большинство ранее назначенных изолированных материалов из супергруппы Wealden являются неопределенными. [ 26 ] Исследование изолированного зуба Баркера и коллег в 2023 году показало, что он и другие зубы из супергруппы Wealden, которые ранее были назначены в Baryonyx, вероятно, не принадлежат к роду, основываясь на их морфологии и возрасте. [ 28 ]
Возможные синонимы
[ редактировать ]

В 2003 году Милнер отметил, что некоторые зубы в музее естественной истории, ранее идентифицированные как принадлежащие к родам Sicosaurus (первый названный спинозаурид), и мегалозавр, вероятно, принадлежал Baryonyx . [ 7 ] [ 29 ] Типовые виды Sighosaurus Гидеоном А. , S. Cultridens , был назван биологом Ричардом Оуэном в 1841 году на основе зубов, обнаруженных геологом Мантеллом в Форесте Тилгейт , Суссекс . Оуэн изначально думал, что зубы принадлежали крокодилу ; Ему еще предстоит назвать группу Dinosauria, которая произошла в следующем году. Второй вид, С. Жирарди , был назван палеонтологом Анри Эмил Соваге в 1897 году на основе фрагментов челюсти и зуба из Бока -до Чапима, Португалия. В 2007 году палеонтолог Эрик Баффета считал зубы С. Жирарди, очень похожие на зубы барионикса (и S. cultridens ), за исключением более сильного развития зубных флейт (или «ребра»; продольные хребты), предполагая, что останки принадлежал тому же роду. Баффетаут согласился с Милнером, что зубы С. Калтрисенса были почти идентичны зубам Б. Уокери , но с более грудной поверхностью. Бывший таксон может быть старшим синонимом последнего (так как он был опубликован в первую очередь), в зависимости от того, были ли различия в таксоне или между разными. По словам Баффетаута, так как вида голотипа S. Cultridens - это единственный зуб, а B. Walkeri - скелет, было бы более практичным сохранить новое имя. [ 30 ] [ 31 ] [ 32 ] В 2011 году Матеус и его коллеги согласились с тем, что Сучозавр был тесно связан с Baryonyx , но считал оба вида в бывшем роде Nomina Dubia (сомнительные названия), поскольку их образцы голотипа не считались диагностическими (не имея различительных признаков) и не могли определенно приравниваться к другим таксоны [ 18 ] Баркер и его коллеги согласились с этим в 2023 году. [ 28 ]

В 1997 году Чариг и Милнер отметили, что две фрагментарные спинозавридные морды из формирования Нигера Элрхаза (сообщили палеонтолог Филипп Таке в 1984 году) были достаточно похожи на барионикс , что они считали их принадлежащим к неопределенным видам рода (несмотря на их много Младший геологический геологический возраст). [ 3 ] В 1998 году эти окаменелости стали основой рода и видов Cristatusaurus lapparenti , названные Таке и палеонтологом Дейлом Расселом . [ 33 ] Палеонтолог Пол Серено и его коллеги назвали новый род и виды Sucomomimus tenerensis позже в 1998 году, основываясь на более полных окаменелостях из формирования Elrhaz. В 2002 году немецкий палеонтолог Ханс-Дилер Свейес и коллеги предположили, что Suchomimus tenerensis был достаточно похож на Baryonyx Walkeri , чтобы считаться видом в пределах того же рода (как B. tenerensis ), и что Сухомимус был идентичен Cristatusaurus . [ 34 ] Милнер согласился с тем, что материал из Нигера был неотличим от Baryonyx в 2003 году. [ 7 ] В реферате конференции 2004 года Хатт и Ньюберри поддержали синонимию, основанную на большом позвонке с тероподом с острова Уайт, который они приписали животному, тесно связанному с Барионисом и Сухомусом . [ 35 ] Более поздние исследования удерживали Baryonyx и Сухомимуса отдельными, тогда как Cristatusaurus , как было предложено, является либо номенованным дубием , либо, возможно, отличным от обоих. [ 18 ] [ 36 ] [ 37 ] [ 38 ] [ 39 ] в 2017 году В документе обзора палеонтолога Карлоса Роберто А. Кэндиро и его коллег заявили, что эта дебаты были в большей степени в сфере семантики, чем науки, поскольку обычно согласны с тем, что Б. Уокри и С. Тенериенсис являются отдельными, родственными видами. [ 40 ] Баркер и его коллеги обнаружили, что Сухомус ближе связан с британским родам Riparovenator и Ceratosuchops , чем с Baryonyx в 2021 году. [ 26 ]
Описание
[ редактировать ]
По оценкам, Baryonyx был длиной от 7,5 до 10 м (25 и 33 фута), 2,5 м (8,2 фута) по высоте бедра и весом от 1,2 до 2 т (1,3 и 2,2 тонны; 1,2 и 2,0 тонны ) Тот факт, что элементы черепа и позвоночной колонны образец голотипа B. Walkeri (NHM R9951), по-видимому, не совпадали ( слитые), предполагает, что человек не был полностью выращен, и зрелое животное, возможно, было гораздо больше (как в случае некоторых других спинозавридов). С другой стороны, плавленная грудина образца указывает, что она могла быть зрелым. [ 3 ] [ 41 ] [ 42 ] [ 43 ]
Череп
[ редактировать ]

Череп Baryonyx является не полностью известным, и большая часть средних и задних порций не сохраняется. По оценкам, полная длина черепа составляла длину 91–95 сантиметров (36–37 дюймов), в зависимости от сравнения с родным родом Sucomomimus (который был на 20% больше). [ 3 ] [ 38 ] Он был удлинен, а передняя 17 см (6,7 дюйма) преднамерье сформировала длинную, узкую и низкую морду ( рострум ) с плавно округлой верхней поверхностью. [ 3 ] Внешние NARES (Bony Nostrils) были длинными, низкими и расположены далеко назад от кончика морды. Передняя 13 см (5,1 дюйма) морда расширилась на лопатую (ложную), «терминальную розетку », форму, похожую на рострум современного гариала . Передние 7 см (2,8 дюйма) нижнего края предчезгилей были спарены (или подключены), тогда как часть передней части челюсти была поднята. Эта морфология привела к сигмоидному или S-образному краю нижнего верхнего ряда зубов, в котором зубы от передней части верхней челюсти выступали вперед. Мвина была особенно узкой прямо за розеткой; Эта область получила большие зубы нижней челюсти. Максилла и предчезгила Baryonyx сочетаются в сложной артикуляции , а результирующий разрыв между верхней и нижней челюстью известен как подрострная выемка . [ 3 ] Продолжительная предчелюстная и сигмоидальная нижняя маржа верхнего зубного ряда также присутствовал в отдаленно связанных тероподах, таких как Dilophosaurus . У морды было обширное отверстие (отверстия), которое было бы выходом для кровеносных сосудов и нервов, [ 3 ] и верхняя челюсть, по -видимому, размещала пазухи . [ 3 ] [ 38 ] [ 42 ]

У Baryonyx был рудиментарный вторичный небо , похожее на крокодилы, но в отличие от большинства динозавров. [ 44 ] Поверхность с морщинистыми (грубо морщинистыми) предполагает присутствие возбужденной накладки в крыше рта. Носовые кости были слиты, которые отличали барионикс от других спинозавридов, а над глазами присутствовал сагиттальный гребень , на верхней средней линии ножных. Этот гребень был треугольным, узким и острым в своей передней части и отличался от других спинозавридов в конце задних палах в перекрестном процессе. Лесковая кость перед глазом, по -видимому, сформировала ядро рога, аналогичное тем, которые наблюдались, например, в аллозавре , и отличалась от других спинозавридов, которые были твердыми и почти треугольными. Затылка была узкой, с парокципиталивыми процессами, указывающими наружу горизонтально, а базиптеригоидные процессы удлинялись, спускающиеся далеко под базиокципитал (самая нижняя кость затылка). [ 3 ] [ 42 ] [ 45 ] Серено и его коллеги предположили, что некоторые из были черепных костей Baryonyx неправильно идентифицированы Коригом и Милнером, в результате чего затылка была реконструирована как слишком глубокая, и что череп вместо этого был таким же низким, длинным и узким, как и у Сухомуса . [ 45 ] Передние 14 см (5,5 дюйма) зубного помещения в нижней челюсти наклонились вверх к кривой морды. Значение было очень длинным и мелким, с выдающейся мекелейской канавкой на внутренней стороне. Симфиз нижней челюсти , где две половинки нижней челюсти соединены спереди, был особенно коротким. Остальная часть нижней челюсти была хрупкой; Задние третьи были намного тоньше, чем передняя, с лезвием, похожим на лезвия. Передняя часть зубной зубья изогнута наружу, чтобы разместить большие передние зубы, и эта область образовала нижнечелюстную часть розетки. Значение, как морда - у меня было много отверстий. [ 3 ] [ 38 ]

Большая часть зубов, найденных с образцом голотипа, не была в артикуляции с черепом; Несколько остались в верхней челюсти, и только небольшие замены зубов все еще носили нижнюю челюсть. У зубов была форма рецищренных конусов, где слегка сплющивалась из боковых путей, а их кривизна была почти однородной. Корни были очень длинными и сужались к их конечности. [ 3 ] Карины (острые края передней и задней части) были мелко зазубренными зубцами спереди и сзади и простирались по всей короне. Было от шести до восьми зубьев на мм (0,039 дюйма), что гораздо большее число, чем у теропод с крупным телом, таким как Torvosaurus и Tyrannosaurus . Некоторые из зубов были рифлены, с шестью до восьми хребтами по длине их внутренних сторон и мелкозернистой эмалью (самый внешний слой зубов), в то время как другие не содержали флейты; Их присутствие, вероятно, связано с положением или онтогеном (развитие во время роста). [ 3 ] [ 38 ] Внутренняя сторона каждого зубного ряда имела костную стену. Количество зубов было большим по сравнению с большинством других теропод, с шестью до семи зубьев в каждой премактилле и тридцатью двумя в каждом зубном состоянии. Основываясь на более близкой упаковке и меньшем размере зубов зубов по сравнению с таковыми в соответствующей длине предчезгиллы, разница между количеством зубов в верхних и нижних челюстях, по -видимому, была более выраженной, чем в других тероподах. [ 3 ] Терминальная розетка в верхней челюсти голотипа имела тринадцать стоматологических альвеол (зубные розетки), шесть слева и семь с правой стороны, показывающие асимметрию количества зубов. Первые четыре верхних зуба были большими (со вторым и третьим самым большим), в то время как четвертый и пятый постепенно уменьшались в размерах. [ 3 ] Диаметр самого большого был вдвое больше, чем у самого маленького. Первые четыре альвеолы зубного мозга (соответствующие кончику верхней челюсти) были самыми большими, а остальные более регулярны по размеру. небольшие субтриангольные межзубные пластины . Между альвеолами присутствовали [ 3 ] [ 38 ]
Посткраниальный скелет
[ редактировать ]

Первоначально считал, что у него не хватало сигмоидной кривой, типичной для теропод, [ 3 ] Шея Baryonyx , по -видимому, сформировала форму S, хотя и прямой, чем в других тероподах. [ 46 ] Шейные позвонки шеи сунулись к голове и постепенно стали дольше спереди назад. Их зигапофизы (процессы, которые соединяли позвонки) были плоскими, и их эпипофизы (процессы, к которым прикреплены мышцы шеи) были хорошо развиты. Ось развитый (второй позвонок шеи) была небольшой по сравнению с размером черепа и имела хорошо гипосен . Нейронные арки шейных позвонков не всегда были зашиты в центр (тела позвонков), а нервные шипы были там низкими и тонкими. Ребра шейки матки были короткими, похожими на ребра с крокодилами, и, возможно, несколько перекрывали друг друга. Центра дорсальных позвонков спины были одинаковыми по размеру. Как и в других тероподах, скелет Baryonyx показал скелетную пневматичность , снижая его вес через фенестра (отверстия) в нейронных арках и плеорокоэле (полые депрессии) в центре (в первую очередь вблизи поперечных процессов ). Спереди к спине нервные шипы дорсальных позвонков изменились с короткого и крепкого на высокий и широкий. Один изолированный дорсальный нервный позвоночник был умеренно вытянутый и стройный, что указывает на то, что У Baryonyx , возможно, был горб или гребень вдоль центра его спины (хотя и растущий по сравнению с другими спинозавридами). Baryonyx был уникальным среди спинозавридов, имея заметное сужение из стороны в сторону в позвонке, который либо принадлежал к крестцу , либо передней части хвоста. [ 3 ] [ 6 ] [ 45 ]

Коракоидные конические награды за задние запасы при просмотре в профиле, и, уникально среди спинозавридов, связанные с лопатой в артикуляции с привязкой и ночкой. Лопатки были надежными, а кости передней основы были короткими по отношению к размеру животного, но широким и крепким. Компания плеча была короткой и прочной, с его концов широко расширены и сплющены - верхняя сторона для дельтопекторного гребня и прикрепления мышц и нижняя часть артикуляции с радиусом и локтевой костью. Радиус был коротким, крепким и прямым, и менее чем половина длины плечевой кости, в то время как локтевая кость была немного длиннее. Умеяница имела мощный олекранон и расширенный нижний конец. У рук было три пальца; Первый палец имел большой когти, размером около 31 см (12 дюймов) вдоль своей кривой в образце голотипа. Коготь был бы удлинен кератиновой (возбужденной) оболочкой в жизни. Помимо его размера, пропорции коготь были довольно типичными для теропода, то есть он был двусторонним симметричным , слегка сжатым, плавно округлым и резко заостренным. Канавка для оболочки проходила по длине когтя. Другие когти руки были намного меньше. А Подвздош (основная бедра) таза имела выдающийся супрацетабулярный гребень , передний процесс, который был стройным и вертикально расширенным, и задний процесс, который был длинным и прямым. У подвздошного костюма также был заметный шельф Бревиса и глубокая роща, которая столкнулась вниз. Втранство . (гнездо для бедренной кости) была длинной спереди к спине Ишай (нижняя и самая задняя бедренная кость) имел хорошо развитый обтюраторный процесс в верхней части. Маржа лобкового лезвия в нижнем конце была выведена наружу, а лобковая нога не была расширена. У бедренной кости не хватало канавки на мыщелке с фибулярным мыщелком, и, уникально среди спинозавридов, у малоберцовой ямки была очень мелкая ямка (депрессия). [ 3 ] [ 42 ] [ 6 ] [ 45 ]
Классификация
[ редактировать ]
В своем первоначальном описании, Чариг и Милнер [ 1 ] Нашел Baryonyx, достаточно уникальный, чтобы оправдать новую семью динозавров Theropod: Baryonychidae. Они обнаружили, что Baryonyx отличается от любой другой группы Theropod, и рассмотрели вероятность того, что это был TheCodont (группировка ранних архозавров, которые теперь считаются неестественными ), из -за того, что имея, по -видимому, примитивные особенности, [ 1 ] Но отметил, что артикуляция верхней челюсти и предчезгиллы была похожа на дилофозавра . Они также отметили, что две морды из Нигера (которая впоследствии стала основой Cristatusaurus ), назначенных семейным Spinosauridae Taquet в 1984 году, казались почти идентичны основанию Baryonyx , и вместо этого они направили их в Baryonchidae. [ 1 ] В 1988 году палеонтолог Грегори С. Пол согласился с Таке, что Спинозавр , описанный в 1915 году на основе фрагментарных останков из Египта, которые были уничтожены во Второй мировой войне , и Барионикс были схожи и (из-за их странных мордов), возможно, поздние выживающие динофозары. [ 43 ] Buffetaut также поддержал эти отношения в 1989 году. [ 47 ] В 1990 году Чариг и Милнер отклонили сродство спинозауридов Baryonyx , поскольку они не нашли, что их остаются достаточно похожими. [ 6 ] В 1997 году они согласились с тем, что Baryonhichidae и Spinosauridae были связаны, но не согласились с тем, что первое имя должно стать синонимом последнего, потому что полнота Baryonyx по сравнению со Спинозавром сделала его лучшим типом родом для семьи, и потому что они не нашли Сходство между двумя достаточно значимыми. [ 3 ] Хольц и его коллеги перечислили Baryonychidae как синоним Spinosauridae в 2004 году. [ 14 ]
Открытия в 1990 -х годах пролили больше света на отношения Baryonyx и его родственников. В 1996 году была названа морда из Марокко в спинозавре , а раздражение и ангатура из Бразилии (эти два - возможные синонимы). [ 48 ] [ 34 ] Cristatusaurus и Shucomimus были названы на основе окаменелостей из Нигера в 1998 году. В своем описании Сучимуса , Серено и его коллег поместили его и Baryonyx в новую подсемейство Baryonychinae в Spinosauridae; Спинозавра и раздражение были помещены в подсемейство Spinosaurinae . Baryonychinae отличался небольшим размером и большим количеством зубов в зубном состоянии за терминальной розеткой, глубоко кирэтированными передними спинными позвонками и с зубчатыми зубами. Спинозауринаэ был отличался их прямыми зубными коронами без зубцов, маленьким первым зубом в предчелюсении, увеличенным расстоянием зубов в челюстях и, возможно, путем того, как их ноздри помещают дальше назад и наличие глубокого нейронного паруса позвоночника . [ 45 ] [ 33 ] Они также объединили спинозавриды и их ближайшие родственники в суперсемействе Spinosauroidea, но в 2010 году палеонтолог Роджер Бенсон считал это младшим синонимом Megalosauroidea (более старое имя). [ 49 ] В реферате на конференции 2007 года палеонтолог Денвер В. Фаулер предположил, что, поскольку судозавр является первым названным родом в своей группе, клады называют Spinosauroidea, Spinosauridae и Baryonychinae, должны быть заменены Sicosauroidea, SicoSauridae, и Sucosaurinae, независимо от того, или нет, или нет. Имя Baryonyx сохраняется. [ 29 ] Исследование, проведенное в 2017 году палеонтологов Маркосом AF Sales и Cesar L. Schultz, показало, что Clade Baryonychinae не поддерживалась, поскольку зубчатые зубы могут быть наследственной чертой среди спинозавридов. [ 39 ]


Баркер и его коллеги нашли поддержку для разделения Baryonychinae-Spinosaurinae в 2021 году, а следующая кладограмма показывает положение Baryonyx в Spinosauridae в соответствии с их исследованием: [ 26 ]
| Spinosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Эволюция
[ редактировать ]
Спинозавриды, по -видимому, были широко распространены от барремана к сеноманской стадии мелового периода средней , около 130-95 миллионов лет назад, в то время как самый старый известный спинозаврид остается датой до юры . [ 50 ] Они поделились такими функциями, как длинные узкие, крокодилоподобные черепа; субкругальные зубы, с тонкими или без зубцов; Терминальная розетка морды; и вторичное небо, которое сделало их более устойчивыми к крути. Напротив, примитивным и типичным условием для теропод представляли собой высокую узкую морду с лезвия, похожие на лезвия (Ziphodont) зубы с зубчатыми каринами. [ 51 ] Адаптация спинозавридов черепа сходилась с адаптацией крокодилов ; У ранних членов последней группы были черепа, похожие на типичные тероподы, впоследствии развивая удлиненные морды, конические зубы и вторичные вкусы. Эти адаптации, возможно, были результатом диетического изменения от наземной добычи на рыбу. В отличие от крокодилов, посткраниальные скелеты барионишиновых спинозавридов, по-видимому, не имеют водных адаптаций. [ 52 ] [ 51 ] Серено и его коллеги предложили в 1998 году, что крупные пальцы и надежные передние конечности спинозавридов развивались в средней юре, до удлинения черепа и других адаптаций, связанных с поеданием рыбы, поскольку первые особенности поделились их родственниками мегалозавридов . Они также предположили, что спинозаурины и барионихины расходились до баремического эпоха раннего мела. [ 45 ]
Было предложено несколько теорий о биогеографии спинозавридов. Поскольку SUCOMIMUS был более тесно связан с Baryonyx (из Европы), чем с Спинозавром , хотя этот род также жил в Африке, распределение спинозавридов не может быть объяснено как викарий, возникающий в результате континентального рифтинга . [ 45 ] Серено и коллеги [ 45 ] предположил, что спинозавриды были первоначально распределены по суперконтиненту Пангея , но разделены с открытием моря Тетис . Спинозаурины тогда развивались бы на юге (Африка и Южная Америка: в Гондване ) и барионихины на севере (Европа: в Лауразии ), с Сухомимусом, результатом одного события рассеивания севера на юг . [ 45 ] Бафтаут и тунисский палеонтолог Мохамед Уаджа также предположили в 2002 году, что барионихины могут быть предками спинозауринов, которые, по -видимому, заменили первое в Африке. [ 53 ] Милнер предположил в 2003 году, что спинозавриды возникли в Лауразии во время юры и рассеялись через иберийский сухопутный мост в Гондвану, где они излучали . [ 7 ] В 2007 году Баффетат указал, что палеогеографические исследования показали, что Иберия была недалеко от Северной Африки во время раннего мелового, которое, как он обнаружил, подтверждает идею Милнера, что Иберийский регион был ступенькой между Европой и Африкой, которая подтверждает присутствие барионихинов в Иберии. Направление рассеивания между Европой и Африкой до сих пор неизвестно, [ 30 ] и последующие открытия спинозаврида остаются в Азии, и, возможно, Австралия указывает на то, что это могло быть сложным. [ 18 ]
Кандейро и его коллеги предположили в 2017 году, что спинозавриды северной гондваны были заменены другими хищниками, такими как абелизавроиды , поскольку не существенные ископаемые спинозауриды не известны после сеномана в мире. Они объяснили исчезновение спинозавридов и других сдвигов в фауне Гондваны с изменениями в окружающей среде, возможно, вызванных нарушением уровня моря . [ 40 ] Малафайя и его коллеги заявили в 2020 году, что Baryonyx остается самым старым бесспорным спинозавридом, признавая, что пожилые останки также были предварительно назначены в группу. [ 23 ] Баркер и его коллеги нашли поддержку европейского происхождения спинозавридов в 2021 году с расширением в Азию и Гондвану в первой половине раннего мелового. В отличие от Серено, эти авторы предположили, что было по меньшей мере два события разгона из Европы в Африку, что привело к Сукомимусу и африканской части Спинозаурины. [ 26 ]
Палеобиология
[ редактировать ]
Диета и кормление
[ редактировать ]Чариг и Милнер предположили, что его удлиненная морда со многими мелко зубчатыми зубами указывала на то, что Baryonyx был разярил (рыба), предполагая, что он присел на берегу реки и использовал свой ког В 1986 году Гризли медведь ). [ 1 ] Два годами ранее, таке [ 54 ] указал, что спинозавриды из Нигера были похожи на те у современного гариала и предложили поведение, похожее на цапли или аисты . [ 3 ] [ 1 ] В 1987 году биолог Эндрю Китченер оспорил писципальное поведение Baryonyx и предположил, что он был бы мусорщиком , используя свою длинную шею, чтобы питаться на земле, его когти, чтобы разразиться в тушу, и его длинная морда (с ноздрями для дыхания) для исследования полости тела. [ 55 ] Китченер утверждал, что и зубы Baryonyx челюсти были слишком слабы, чтобы убить других динозавров и слишком тяжелые, чтобы ловить рыбу, со слишком большим количеством адаптаций для Рычности. [ 55 ] По словам палеонтолога Робина Э. Э. Рейда, утопленная каркасная каркаса была бы разбита его хищником, а крупные животные, способные сделать это - такие как медведи -гризли - также способны ловить рыбу (по крайней мере, в мелководье). [ 56 ]


В 1997 году Чариг и Милнер продемонстрировали прямые диетические доказательства в области желудка голотипа Б. Уокери . Он содержал первое доказательство речи в тероподном динозавре, с кислотой масштабами и зубами общей рыбы Scheenstia Mantelli (затем классифицируется в роде Lepidotes [ 57 ] ), а также искаженные или протяженные кости молодого игуанодонтида . Они также представляли косвенные доказательства речи, такие как крокодилоподобные адаптации для ловли и проглатывания добычи: длинные узкие челюсти с их «терминальной розеткой», аналогичной таковой у гариала, а также спаренная кончика и ступенька. По их мнению, эти адаптации предположили, что Baryonyx поймал бы небольшую и среднюю рыбу в виде крокодильца: схватить их с выемкой морды (давая зубам «функция ударов»), наклоняя голову назад, и и глотать их головой. [ 3 ] Большая рыба будет разбита с помощью когтей. То, что зубы в нижней челюсти были меньше, более многолюдны и многочисленны, чем в верхней челюсти, могли бы помочь животным схватить пищу. Количество и Милнер утверждали, что Baryonyx в первую очередь съел бы рыбу (хотя это также был бы активным хищником и оппортунистическим мусорщиком), но он не был оборудован как макро-преседатор, как Allosaurus . Они предположили, что Baryonyx в основном использовал свои передние конечности и большие когти, чтобы поймать, убить и разорвать большую добычу. Очевидный гастролит ( Gizzard Stone) также был обнаружен с образцом. [ 3 ] Немецкий палеонтолог Оливер Вингс предположил в 2007 году, что низкое количество камней, обнаруженных в тероподах, таких как Baryonyx и Allosaurus, могло проглотить случайно. [ 58 ] В 2004 году позвонок шейки птерозавра из Бразилии с зубом спинозавридов, встроенный в него, о котором сообщили Buffetaut, а коллеги подтвердили, что последние не были исключительно неровными. [ 59 ]

Исследование теории луча, проведенное палеонтологом Франсуа Терриен и его коллегами, не смогли восстановить профили силы Baryonyx , но обнаружили, что связанный сухомимус использовал бы переднюю часть ее челюстей, чтобы захватить добычу, и предположили, что челюсти спискозаридов были бы адаптированы. для охоты на меньшую земную добычу в дополнение к рыбе. Они предполагали, что спинозавриды могли запечатлеть меньшую добычу с розеткой зубов в передней части челюстей, и закончить ее, встряхивая. Вместо этого большая добыча была бы захвачена и убита своими передниками вместо их укуса, поскольку их черепа не смогут противостоять изгибному стрессу. Они также согласились с тем, что конические зубы спинозавридов были хорошо разработаны для пронзания и удержания добычи, а их форма позволила им выдерживать изгибы нагрузки со всех сторон. [ 60 ] в 2007 году Анализ конечных элементов Сканированные морды палеонтолога Эмили Дж. Рэйфилд и его коллеги показал, что наиболее похожа биомеханика барионекса на биологические и не похожие на американские аллигаторы и более нечеенные тероподы, поддерживая писци-иродную диету для спинозавриды. Их второстепенное небо помогло им сопротивляться изгибе и скручиванию трубчатых мордов. [ 44 ] в 2013 году Исследование теории луча от палеонтологов Эндрю Р. Каффа и Рейфилда сравнивало биомеханику КТ-бортовых бундов с таковыми у существующих крокодилов и обнаружило, что морды барионекс и спинозавра, сходные по их сопротивлению из-за изгиба и отрыва. Было обнаружено, что у Baryonyx относительно высокая устойчивость в морде до дорсовентрального изгиба по сравнению со спинозавром и гариалом. Авторы пришли к выводу (в отличие от исследования 2007 года), что Baryonyx работал иначе, чем Gharial; Спинозавриды не были эксклюзивными писцистанциями, а их диета определялась их индивидуальным размером. [ 8 ]
В реферате на конференции 2014 года палеонтолог Дэнни Андуза и Фаулер отметили, что медведи гризли не вытаскивают рыбу из воды, как было предложено для Baryonyx , а также исключило, что динозав из спинозавридов не были сильно заглушены, а их глаза не были хорошо расположены для бинокулярного зрения . Вместо этого они предположили, что челюсти сделают в сторону развернуть рыбу, как гариальные, с когтиками, которые, вероятно, использовались, чтобы отпечатать и пронзительную большую рыбу, тогда как они манипулировали ими своими челюстями, аналогично медведям гризли и рыбалке кошки Они не нашли зубы спинозавридов, подходящими для расчленения добычи, из -за отсутствия зубцов, и предположили, что они проглотили бы добычу целую (при этом отмечая, что они также могли бы использовать свои когти для расчленения). [ 61 ]
Исследование, проведенное палеонтологом Кристофа Хендриккса в 2016 году, показало, что взрослые спинозавры могут вытеснить свои нижнечелюстные рами (половинки нижней челюсти ) в боковой Полем Эта челюстная артикуляция аналогична той, которая наблюдается у птерозавров и живых пеликанов , и также позволила бы спинозавридам проглотить большую добычу, такую как рыба и другие животные. некоторые возможные португальские окаменелости Baryonyx Они также сообщили, что были обнаружены , связанные с изолированными зубами игуанодона , и перечислили их вместе с другими подобными ассоциациями в качестве поддержки оппортунистического поведения кормления в спинозаврах. [ 38 ] Еще одно исследование, проведенное палеонтологом Романом Вулло и его коллегами, показало, что челюсти спинозавридов были сходящимися с шлюхами Пайк Конгера ; У этих рыб также есть челюсти, которые сжаты от стороны в сторону (в то время как челюсти крокодилов сжаты сверху вниз), удлиненную морду с «терминальной розеткой», которая несет увеличенные зубы, и выемка за розеткой с меньшими зубами. Такие челюсти, вероятно, развивались для захвата добычи в водных условиях с низким светом, и, возможно, помогли в обнаружении добычи. [ 62 ]
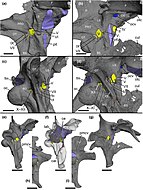

Исследование 2023 года, проведенное Баркером и коллегами, основанное на КТ-мозговых значениях барионекс и цератосухопов, показало, что анатомия мозга этих барионихинов была аналогична аналогии другим не манирапторным тероподам. Их нейросенсорные возможности, такие как слух и обоняние (обоняние), были необычными, а их стабилизация взгляда менее развита, чем у спинозавринов, поэтому их поведенческие адаптации, вероятно, были сопоставимы с возможностями других наземных тероподов с крупным телом. Это говорит о том, что их переход от наземных гиперкарноводов к полу-кватическим «универсалам» во время их эволюции не потребовал существенной модификации их мозга и сенсорных систем. Это может означать, что спинозавриды были либо предварительно адаптированы для обнаружения и захвата водной добычи, либо что их переход к полу-квартирному образу жизни требовал лишь модификации костей, связанных с ртом. Их рептилий энцефализационные ценности подразумевают, что когнитивная способность и поведенческая изощренность барионихинов не сильно отклонились от других других базальные тероподы. [ 63 ]
Движение и полу-квадратные привычки
[ редактировать ]В своем первоначальном описании Чариг и Милнер не считали Baryonyx водным (из-за того, что его ноздри находятся на сторонах ее морды-Far от наконечника-и формы посткраниального скелета), но думал, что он способен на Плавание, как и большинство земельных позвоночных. [ 1 ] Они предположили, что удлиненный череп, длинная шея и сильная плечевая кость Baryonyx указывали на то, что животное было факультативным четвероногим , уникальным среди теропод. [ 1 ] В своей статье 1997 года они не обнаружили для этого не поддержки скелета, но утверждали, что передние конечности были бы достаточно сильны для четверопедтной позы, и это, вероятно, поймало бы водную добычу во время приседания - или на всех четвертях - более популярной (или в) водой. [ 3 ] Повторное описание спинозавра в 2014 году Палеонтологом Низаром Ибрагимом и его коллегами, основанная на новых останках, предположила, что он был четвероногим, основанным на его переднем центре массы тела . Авторы обнаружили, что четвероделия маловероятной для Baryonyx , поскольку более известные ноги тесно связанного Сухомуса не поддерживали эту позу. [ 52 ]
В 2017 году палеонтологи Дэвид Э. Хон и Хольц предположили, что головные гребни спинозавридов, вероятно, использовались для сексуального или угрозы. Авторы также отметили, что (как и другие тероподы) не было никаких оснований полагать, что передние конечности барионекса были способны пронатеть (пересечение радиуса и костей локтевой зоны нижней руки, чтобы повернуть руку), и тем самым сделать его способным отдыхать Или ходить по ладоням. Можно было бы быть возможным для отдыха или использования передней части передвижения (как указывается треки покоящегося теропода), но если бы это было нормой, передние мыслители, вероятно, показали бы адаптацию для этого. Hone и Holtz Кроме того, предположили, что передние мыши спинозавридов не кажутся оптимальными для захвата добычи, но вместо этого выглядят похожими на передние конечности копающих животных. Они предположили, что способность копать была бы полезно при раскопке гнезд, копании воды или достижению некоторых видов добычи. Хон и Хольц также полагали, что спинозавриды будут проходить и окунаться в воде, а не погрузить себя из -за их разреженности водной адаптации. [ 11 ]


Исследование, проведенное в 2010 году палеонтологом Романом Амиотом и его коллегами, предположило, что спинозавриды были полуаколичными, основываясь на изотопном составе кислорода зубов спинозавридов со всего мира по сравнению с другими тероподами и существующими животными. Спинозавриды, вероятно, провели большую часть дня в воде, таких как крокодилы и бегемоты , и имели диету, похожую на первую; Оба были оппортунистическими хищниками. Поскольку большинство спинозавридов, по -видимому, не имеют анатомической адаптации для водного образа жизни, авторы предположили, что погружение в воду было средством терморегуляции, аналогичного таковым у крокодилов и бегемота. Спинозавриды также могли обратиться к водным местам обитания и речи, чтобы избежать конкуренции с большими, более терроподными тероподами. [ 64 ] В 2016 году продажи и коллеги статистически изучили ископаемое распределение спинозавридов, абелисавридов и карчадонтозавридов и пришли к выводу, что спинозавриды оказывают самую сильную поддержку в связи с прибрежными палеоэрадонами. Спинозавриды также, по -видимому, населяли внутреннюю среду (с их распространением, сопоставимым с кархадонтозавридами), что указывает на то, что они могли быть более обобщенными, чем обычно думали. [ 65 ]
Продажи и Шульц в 2017 году согласились с тем, что спинозавриды были полуаколичными и частично неровными, основываясь на таких функциях черепа, как конические зубы, морды, которые были сжаты из стороны в сторону, и втянутые ноздри. Они интерпретировали тот факт, что гистологические данные указывают на то, что некоторые спинозавриды были более наземными, чем другие, как отражающие экологическое разделение ниши среди них. Поскольку у некоторых спинозавридов есть меньшие ноздри, чем другие, их обонятельные способности были, по -видимому, меньше, как у современных писци иродных животных, и вместо этого они могут использовать другие чувства (такие как зрение и механорецепция ) при охоте рыбы. Огнетание, возможно, было более полезным для спинозавридов, которые также питались наземной добычей, такой как барионихины. [ 39 ] Исследование 2022 года, проведенное палеонтологом Маттео Фаббри и его коллегами, показало, что Baryonyx обладал плотными костями, которые позволили бы ему нырнуть под водой. Эта же адаптация была выявлена в связанном спинозавре , и, как полагают, они были подковистыми фуражами, которые нырнули после водной добычи, в то время как сухомимус был лучше адаптирован к непакологическому образу жизни в соответствии с предоставленным анализом. Это открытие также демонстрирует уникальный и экологически разрозненный образ жизни, которые были спинозавриды, с большим количеством впальных родов, предпочитающих охотиться в более мелкой воде. [ 66 ] [ 67 ] [ 68 ]
Палеоэрона
[ редактировать ]
Формирование глины Weald состоит из отложений гаутеривийской (нижней глины Weald) до барремавой (верхней глины Weald) , возраста около 130–125 миллионов лет. Первоначальный образец Baryonyx был обнаружен в последнем, в глине, представляющей немаренную все еще воду, которая была интерпретирована как речная или грязная среда с мелкой водой, лагунами и болотами . [ 3 ] Во время раннего мелового района Суррей, Сассекс и Кент был частично покрыт большим, свежим- солончатым озером Уилдена . Две большие реки истощили северную территорию (где сейчас стоит Лондон), входящие в озеро через дельту реки ; Англо-паррис бассейн был на юге. Его климат был субтропическим , похожим на нынешний средиземноморский регион . Поскольку яма для глины SmokeJack состоит из разных стратиграфических уровней , ископаемые таксоны обнаружили, что они не обязательно существуют одновременно. [ 3 ] [ 69 ] [ 70 ] Динозавры из местности включают Ornithopods Mantellisaurus , Iguanodon и небольшие сауроподы . [ 71 ] Другие позвоночные из глины Weald включают крокодилов, птерозавров, ящериц (такие как Dorsetisaurus ), амфибии, акулы (такие как Hybodus ) и костные рыбы (включая Scheenstia ). [ 72 ] [ 73 ] Были идентифицированы члены десяти заказов насекомых, в том числе Valditermes , Archisphex и Pterinoblattina . Другие беспозвоночные включают остракоды , изоподы , конхостраканы и двустворчатые моллюски . [ 74 ] [ 75 ] Растения Вайчселия и водные травянистые Бевральстия были обычными. Другие найденные растения включают папоротники , хвощи , клубные мохи и хвойные . [ 76 ] [ 77 ]
Другие динозавры из формирования Wessex острова Уайт, где, возможно, произошел Baryonyx , включают в себя теропод Riparovenator , Ceratosuchops , Neovanator , Eotyrannus , Aristosuchus , Thecocoelurus , Calamospondylus и Ornithodesmus ; Ornithopods Iguanodon , Hypsilophodon и Valdosaurus ; Sauropods Ornithopsis , Eucamerotus и Chondrosteosaurus ; и анкилозавра Polacanthus . [ 78 ] [ 26 ] Баркер и его коллеги заявили в 2021 году, что идентификация двух дополнительных спинозавридов из супергруппы Wealden, Riparovenator и Ceratosuchops имеет значение для потенциального экологического разделения в Spinosauridae, если эти и барионикс были современными и взаимодействующими. Они предупредили, что возможно, что формации верхней глины и Wearsex и спинозавриды, известные от них, были разделены во времени и расстоянии. [ 26 ]
Как правило, считается, что крупные хищники происходят с небольшим таксономическим разнообразием в любой области из -за экологических требований, однако многие мезозойские объединения включают два или более симпатрических теропод, которые были сопоставимы по размеру и морфологии, и, по -видимому, это было в случае спинозаридов. Баркер и его коллеги предположили, что высокое разнообразие в Spinosauridae в данной области, возможно, было результатом экологических обстоятельств, приносящих пользу их нише. Хотя в целом предполагалось, что только идентифицируемые анатомические признаки, связанные с разделением ресурсов, разрешены для сосуществования крупных теропод, Баркер и коллеги отметили, что это не исключает, что аналогичные и тесно связанные таксоны могут сосуществовать и совпадать с экологическими требованиями. Возможное нешевое распределение может быть во времени (сезонное или ежедневное), в космосе (между средами обитания в тех же экосистемах) или в зависимости от условий, и они также могли быть разделены по выбору среды обитания в своих регионах (которые могли иметь место в климат). [ 26 ]
Тафономия
[ редактировать ]
Через Гиг и Милнер представили возможный сценарий, объясняющий тафономию (изменения во время распада и окаменелости) образца голотипа B. Walkeri . [ 3 ] Мелкозернистые отложения вокруг скелета и тот факт, что кости были найдены близко друг к другу (элементы черепа и передняя часть передней части на одном конце зоны раскопок, а также элементы таза и задних конечностей) указывает на то, что окружающая среда была тихой Во время осаждения, и водные токи не перевозили тушу далеко - возможно, потому что вода была мелкой. Район, где умер образ, кажется, подходил для писцидного животного. Возможно, он поймал рыбу и поглотил на грязной равнине, становясь погрязкой до смерти и был похоронен. Поскольку кости хорошо сохранились и не имели грызей, а также нетронувшись, а также нетронутые мусорщиками (предполагая, что он был быстро покрыт отложением). [ 3 ]
Рассылка костей, возможно, была результатом разложения мягких тканей. Части скелета, по -видимому, вывещались в разных градусах, возможно, потому, что уровень воды изменился или сдвинулись отложения (обнажая части скелета). Пояс и кости конечности, зубные и ребро были сломаны перед окаменелостью, возможно, из -за вытапления крупными животными во время похороненных. Большая часть хвоста, по -видимому, была потеряна до окаменелости, возможно, из -за удаления или гниения и выпадения. Ориентация костей указывает на то, что каркас лежала на спине (возможно, слегка наклонилась влево, с правой стороной вверх), что может объяснить, почему все нижние зубы выпали из их гнезда, и некоторые верхние зубы все еще были на месте Полем [ 3 ] [ 2 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Charig, aj; Милнер, AC (1986). « Baryonyx , замечательный новый динозавр Theropod». Природа . 324 (6095): 359–361. Bibcode : 1986natur.324..359c . doi : 10.1038/324359a0 . PMID 3785404 . S2CID 4343514 .
- ^ Jump up to: а беременный в дюймовый и фон Psihoyos, L.; Knoebber, J. (1994). Охота на динозавров . Лондон: Касселл. С. 176–179 . ISBN 978-0679431244 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В х и С аа Аб и объявление Но из в нравиться Charig, aj; Милнер, AC (1997). « Baryonyx Walkeri , динозавр, питающий рыбу из Wealden of Surrey» . Бюллетень музея естественной истории Лондона . 53 : 11–70.
- ^ Jump up to: а беременный Эдвардс, Д.Д. (1986). «Ископаемое когтя обнаруживает новое семейное древо». Science News . 130 (23): 356. doi : 10.2307/3970849 . JSTOR 3970849 .
- ^ Jump up to: а беременный Муди, RTJ; Найш Д. (2010). «Алан Джек Чариг (1927–1997): обзор его академических достижений и роли в мире исследований ископаемых рептилий». Геологическое общество, Лондон, Специальные публикации . 343 (1): 89–109. BIBCODE : 2010GSLSP.343 ... 89M . doi : 10.1144/sp343.6 . S2CID 129586311 .
- ^ Jump up to: а беременный в дюймовый и Charig, aj; Милнер, AC (1990). «Систематическое положение Baryonyx Walkeri , в свете реклассификации Gauthier Theropoda» . В Карпентере, К.; Currie, PJ (ред.). Систематика динозавра: перспективы и подходы . Кембридж: издательство Кембриджского университета. С. 127–140. doi : 10.1017/cbo9780511608377.012 . ISBN 978-0-521-43810-0 .
- ^ Jump up to: а беременный в дюймовый Милнер, AC (2003). «Тероподы, питающие рыбу: краткий обзор системы, биологии и палаобеогеографии спиносур». Actas de Las II Jornadas Internacionales Sobre Paleontologico de Dinosaurios Y на Entorno : 129–138.
- ^ Jump up to: а беременный Манжета, AR; Рэйфилд, EJ (2013). Фарке, Эндрю А (ред.). «Механика кормления в спинозауридах и существующих крокодильцах» . Plos один . 8 (5): E65295. BIBCODE : 2013PLOSO ... 865295C . doi : 10.1371/journal.pone.0065295 . PMC 3665537 . PMID 23724135 .
- ^ Норман, Д.Б. (1985). «Дромаозавриды». Иллюстрированная энциклопедия динозавров: оригинальное и убедительное понимание жизни в королевстве динозавров . Нью -Йорк: Полумесяц. С. 57–58. ISBN 978-0-517-46890-6 .
- ^ Остерлофф Э. (2018). «Разрушение мифов динозавров и заблуждений в фильмах» . www.nhm.ac.uk. Получено 20 октября 2018 года .
- ^ Jump up to: а беременный Hone, DWE; Хольц, Т.Р. (2017). «Столетие спинозавров - обзор и пересмотр Spinosauridae с комментариями об их экологии» . Acta Geologica Sinica - английское издание . 91 (3): 1120–1132. Bibcode : 2017acgls..91.1120h . doi : 10.1111/1755-6724.13328 . S2CID 90952478 .
- ^ Clabby, SM (2005). " Baryonyx Charig and Milner 1986" . Dinowight . Получено 12 октября 2015 года .
- ^ Виера, Ли; Башни, JA (1995). « Сауриша, Теропода ) в Weald of La Rioja (Испания) » Естественные науки (на испанском). 47 : 57–61. ISSN 0214-7
- ^ Jump up to: а беременный Хольц, Тр; Молнар, Re; Currie, PJ (2004). «Базальные столбцы». В Вайшампеле, дБ; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2 изд.). Беркли: Университет Калифорнийской прессы. С. 71–110. ISBN 978-0-520-24209-8 .
- ^ Видарте, CF; Лысый, мм; Meijide, M.; Слева, Лос -Анджелес; Монтеро, Д.; Pérez, G.; Torcida, F.; Uriero, v.; Источники, FM; Fuentes, MM (2001). « Остатки ископаемых Baryonyx (Dinosauria, Theopoda) в нижнем меловом фоне младенцев Салас (Бургос, Испания)». Материалы международных дней I по палеонтологии динозавров и их окрестностям. Младшие комнаты, Бургос (на испанском языке): 349–359.
- ^ Pereda-suberbiola, x.; Руиз-Оменья, Джи; Canudo, Ji; Torcida, F.; Sanz, JL (2012). «Фауны динозавров из раннего мела (авалангинско-албийца) Испании» . В Godfroit, P. (ed.). Берниссарт Диносурс . Издательство Университета Индианы. стр. 389–390. ISBN 978-0-253-00570-0 .
- ^ Pérez-Lance, F. (2015). Следы динозавров и дорожки La Rioja Жизнь прошлого. Индиана: издательство Университета Индианы. П. 325. ISBN 978-0-253-01515-0 .
- ^ Jump up to: а беременный в дюймовый Матус, О.; AROWJO, R.; Натарио, C.; Кастинханха Р. (2011). «Новый образец динозавра Theropod Baryonyx из раннего мела Португалии и таксономической достоверности SicoSaurus » (PDF) . Zootaxa . 2827. 2827 : 54–68. doi : 10.11646/Zootaxa .
- ^ Арден, TMS; Кляйн, CG; Zouhri, S.; Лонгрич, Н.Р. (2018). «Водная адаптация в черепе плотоядных динозавров (Theropoda: Spinosauridae) и эволюция водных привычек в спинозавра». Мерашные исследования . 93 : 275–284. Bibcode : 2019crres..93..275a . doi : 10.1016/j.cretres.2018.06.013 . S2CID 134735938 .
- ^ Мэтью, Октавио; Стравиз-Лопес, Дарио (2022). «Новый динозавр Theropod из раннего мела (Барремаана) Кабо Эспикеля, Португалия: последствия для эволюции спинозауридов » . Plos один . 17 (2): E0262614. Bibcode : 2022ploso..1762614M . Doi : 10.1371/journal.pone.0262614 . ISSN 1932-6203 . PMC 8849621 . PMID 35171930 .
- ^ Malafaia, E.; Gasulla, JM; Вскоре, ф.; Narvaez, i.; Sanz, JL; Ортега, Ф. (2018). Архиллы Мореллы Формирование Мерашные исследования 92 : 174–183. Bibcode 2018CrRes..92..174M: два 10.1016/j.cretres.2018.08.006: 134572693S2CID
- ^ Jump up to: а беременный Malafaia, E.; Gasulla, JM; Escaso, F.; Narvaéz, я.; Ортега, Ф. (2020). «Обновление спинозауридов (Dinosauria: Theropoda) ископаемых записей из нижнего мела на иберийском полуострове: распределение, разнообразие и эволюционная история». Журнал иберийской геологии . 46 (4): 431–444. Bibcode : 2020jibg ... 46..431M . doi : 10.1007/s41513-020-00138-9 . S2CID 222149842 .
- ^ Santos-Cubed, Андрес; Сантистебан, Чарльз; Поза, Бегонья; Meseguer, Sergi (2023). Испания) ( Научные отчеты 13 1 ( ) : два 10.1038/s41598-023-33418-2: 10195869PMC 37202441PMID
- ^ Isasmendi, Erik (2024). «Увеличение тероподного отчета о Европе: новая базальная спинозаврида из группы ENCISO бассейна Камероса (La Rioja, Испания). Эволюционные последствия и палеобиоооохождение». Зоологический журнал Линневого общества . XX : 34. doi : 10.1093/Zoolinnean/Zlad193 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Barker, CT; Hone, DWE; Найш, Д.; Кау, а.; Локвуд, JAF; Фостер, Б.; Кларкин, CE; Schneider, P.; Гостолинг, Нью -Джерси (2021). «Новые спинозавриды из формирования Wessex (ранний меловый, Великобритания) и европейское происхождение Spinosauridae» . Научные отчеты . 11 (1): 19340. Bibcode : 2021natsr..1119340b . doi : 10.1038/s41598-021-97870-8 . PMC 8481559 . PMID 34588472 .
- ^ Munt, MC; Блэквелл, Г.; Кларк, Дж.; Фостер, Б. (2017). «Новый спинозауридный динозавр находит из формирования Wessex (Wealden Group, Раннее меловое) острова Уайт». SVPCA . 65 : 1. doi : 10.13140/rg.2.2.17925.86242 .
- ^ Jump up to: а беременный Баркер, Крис Т.; Найш, Даррен; Гостолинг, Нил Дж. (2023). «Изолированный зуб выявляет скрытое разнообразие динозавров спинозауридов в британской супергруппе Wealden (нижние меловые)» . ПЕРЕЙ . 11 : E15453. doi : 10.7717/peerj.15453 . PMC 10239232 . PMID 37273543 .
- ^ Jump up to: а беременный Fowler, DW (2007). «Недавно заново открыли зубы барионишина (Dinosauria: Theropoda): новые морфологические данные, расширение диапазона и сходство с Ceratosaurus » . Журнал палеонтологии позвоночных . 27 (3): 3.
- ^ Jump up to: а беременный Buffetaut, E. (2007). «Динозавр спинозауридов Baryonyx (Saurischia, Theropoda) в раннем меловом виде Португалии» (PDF) . Геологический журнал . 144 (6): 1021–1025. Bibcode : 2007geom..144.1021b . doi : 10.1017/s0016756807003883 . S2CID 130212901 .
- ^ Hendrickx, C.; Матеус, О.; Araújo, R. (2015). «Предложенная терминология зубов теропод (Dinosauria, Saurischia)» . Журнал палеонтологии позвоночных . 35 (5): E982797. Bibcode : 2015jvpal..35e2797h . doi : 10.1080/02724634.2015.982797 . S2CID 85774247 .
- ^ Hendrickx, C. (2008). "Spinosauridae - Historique des decouvertes" . Spinosauridae.fr.gd (по -французски) . Получено 22 октября 2018 года .
- ^ Jump up to: а беременный Таке, П.; Рассел, из (1998). «Новые данные о спинозауридах динозавров из раннего мела Сахары». Comptes rendus de l'Académie des Sciences, Série Iia . 327 (5): 347–353. Bibcode : 1998crase.327..347t . Doi : 10.1016/s1251-8050 (98) 80054-2 .
- ^ Jump up to: а беременный Суды, HD; Frey, E.; Martill, M.; Скотт, Д.М. (2002). « Иртатор Challengeri , спинозаурид (Dinosauria: Theropoda) из нижнего мела Бразилии». Журнал палеонтологии позвоночных . 22 (3): 535–547. doi : 10.1671/0272-4634 (2002) 022 [0535: icasdt] 2.0.co; 2 . S2CID 131050889 .
- ^ Hutt, S.; Ньюбери, П. (2004). «Новый взгляд на Baryonyx Walkeri (Parig and Milner, 1986) на основе недавней ископаемой находки из Wealden» . Симпозиум палеонтологии позвоночных и сравнительной анатомии . Архивировано из оригинала 5 октября 2015 года.
- ^ Allain, R.; Xaisanavong, T.; Ричир, П.; Хентавонг Б. (2012). «Первая окончательная азиатская спинозаврида (Dinosauria: Theropoda) из раннего мелового чуда Лаоса». Naturwissenschaften . 99 (5): 369–377. Bibcode : 2012nw ..... 99..369a . doi : 10.1007/s00114-012-0911-7 . PMID 22528021 . S2CID 2647367 .
- ^ Бенсон, RBJ; Каррано, MT; Brusatte, SL (2009). «Новая клада архаичных хищных динозавров с крупным телом (Theropoda: Allosauroidea), которые выжили до последнего мезозоя» (PDF) . Naturwissenschaften (представленная рукопись). 97 (1): 71–78. Bibcode : 2010nw ..... 97 ... 71b . doi : 10.1007/s00114-009-0614-x . PMID 19826771 . S2CID 22646156 .
- ^ Jump up to: а беременный в дюймовый и фон глин Hendrickx, C.; Матеус, О.; Buffetaut, E.; Эванс, AR (2016). «Морфофункциональный анализ квадратных спинозауридов (Dinosauria: Theropoda) и наличия спинозавра и второго таксона спинозавра в сеномане Северной Африки» . Plos один . 11 (1): E0144695. BIBCODE : 2016PLOSO..1144695H . doi : 10.1371/journal.pone.0144695 . PMC 4703214 . PMID 26734729 .
- ^ Jump up to: а беременный в Продажи, маф; Schultz, CL (2017). «Таксономия спинозавра и эволюция краниодиентационных черт: доказательства из Бразилии» . Plos один . 12 (11): E0187070. BIBCODE : 2017PLOSO..1287070S . doi : 10.1371/journal.pone.0187070 . PMC 5673194 . PMID 29107966 .
- ^ Jump up to: а беременный Кандейро, CRA; Брусатте, SL; Souza, AL (2017). «Динозавры спинозауридов из раннего мела Северной Африки и Европы: ископаемые записи, биогеография и вымирание » . Ежегодник Института героян . 40 (3): 294–302. Doi : 10.11137/2017_3_294_302 . HDL : 20.500.11820/1BC143B5-5BD5-416B-BF10-DD28304D2C9B .
- ^ Терриен, Ф.; Хендерсон, Д.М. (2007). «Мой теропод больше вашего… или нет: оценка размера тела по длине черепа в тероподах». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634 (2007) 27 [108: mtibty] 2.0.co; 2 . S2CID 86025320 .
- ^ Jump up to: а беременный в дюймовый Пол, GS (2010). Полевой гид Принстона по динозаврам . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 87–88 . ISBN 978-0-691-13720-9 .
- ^ Jump up to: а беременный Пол, Г.С. (1988). Хищные динозавры мира . Нью -Йорк: Саймон и Шустер. С. 271–274 . ISBN 978-0-671-61946-6 .
- ^ Jump up to: а беременный Рэйфилд, EJ; Милнер, AC; Сюань, Вб; Янг, PG (2007). «Функциональная морфология спинозавра 'крокодила-мимических« динозавров ». Журнал палеонтологии позвоночных . 27 (4): 892–901. doi : 10.1671/0272-4634 (2007) 27 [892: fmoscd] 2.0.co; 2 . S2CID 85854809 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Серено, ПК; Бек, Ал; DuTheil, DB; Gado, B.; Ларссон, HCE; Лион, GH; Марко, JD; Раухут, Оум; Садлейр, RW; Сидор, Калифорния; Varricchio, DD; Уилсон, GP; Уилсон, JA (1998). «Давно выпущенный хищный динозавр из Африки и эволюция спинозавридов». Наука . 282 (5392): 1298–1302. Bibcode : 1998sci ... 282.1298s . doi : 10.1126/science.282.5392.1298 . PMID 9812890 .
- ^ Evers, SW; Раухут, Оум; Милнер, AC; McFeeters, B.; Allain, R. (2015). «Переоценка морфологии и систематического положения сигилмасасавра динозавров Theropod из« среднего »мела Марокко» . ПЕРЕЙ . 3 : E1323. doi : 10.7717/peerj.1323 . PMC 4614847 . PMID 26500829 .
- ^ Buffetaut, E. (1989). «Новые остатки загадочного спинозавра динозавров из мела Марокко и аффинности между спинозавром и бариониксом ». Новый ежегодный книга по геологии и палеонтологии - ежемесячные виды . 1989 (2): 79–87. Doi : 10.1127/njgpm/1989/189/79 .
- ^ Рассел, Д.А. (1996). «Изолированные кости динозавров от среднего мела тафилальта, Марокко». Бюллетень du muséum national d'istoire Naturelle, Париж, раздел c . 4e Série. 18 (2–3): 349–402.
- ^ Бенсон, RBJ (2010). «Описание Megalosaurus Bucklandii (Dinosauria: Theropoda) от батониана Великобритании и отношения средних юрских теропод» . Зоологический журнал Линневого общества . 158 (4): 882–935. doi : 10.1111/j.1096-3642.2009.00569.x . S2CID 84266680 .
- ^ Serrano-Martínez, A.; Vidal, D.; Sciscio, L.; Ортега, Ф.; Knoll, F. (2015). "Мой изолирован. Acta Poolonological Польша doi : 10.4202/app.00101.2014 . HDL : 10261/152148 . S2CID 53331040 .
- ^ Jump up to: а беременный Holtz Jr., TR (1998). «Спинозавры как крокодил имитирует». Наука . 282 (5392): 1276–1277. doi : 10.1126/science.282.5392.1276 . S2CID 16701711 .
- ^ Jump up to: а беременный Ибрагим, Н .; Серено, ПК; Dal Sasso, C.; Maganuco, S.; Фабри, М.; Martill, DM; Zouhri, S.; Myhrvold, N.; Lurino, DA (2014). «Полуватические адаптации в гигантском хищническом динозавре» . Наука . 345 (6204): 1613–1616. Bibcode : 2014sci ... 345.1613i . doi : 10.1126/science.1258750 . PMID 25213375 . S2CID 34421257 . Дополнительная информация
- ^ Buffetaut, E.; Ouaja, M. (2002). «Новый образец спинозавра (Dinosauria, Theropoda) из нижнего мела Туниса, с замечаниями об эволюционной истории Spinosauridae» (PDF) . Бюллетень de la Société Géologique de France . 173 (5): 415–421. doi : 10.2113/173.5.415 . HDL : 2042/216 . S2CID 53519187 .
- ^ Таке, П. (1984). «Любопытная специализация черепа некоторых плотоядных меловых динозавров: длинная и узкая морда спинозауриды». Отчеты Академии наук (по -французски). 299 : 217–222.
- ^ Jump up to: а беременный Китченер, А. (1987). «Функция когти». Природа . 325 (6100): 114. Bibcode : 1987natur.325..114K . doi : 10.1038/325114A0 . S2CID 4264665 .
- ^ Рейд, Рех (1987). «Когти» . Природа . 325 (6104): 487. Bibcode : 1987natur.325..487r . doi : 10.1038/325487b0 . S2CID 4259887 .
- ^ López-Arbarello, A. (2012). «Филогенетические взаимосвязи гиллинских рыб (Actinopterygii: neopterygii)» . Plos один . 7 (7): E39370. Bibcode : 2012ploso ... 739370L . doi : 10.1371/journal.pone.0039370 . PMC 3394768 . PMID 22808031 .
- ^ Wings, O. (2007). «Обзор функции гастролита с последствиями для ископаемых позвоночных и пересмотренной классификации» . Acta Palaeontologica Polonica . 52 (1): 1–16.
- ^ Buffetaut, E.; Martill, D.; Escuillié, F. (2004). «Птерозавры как часть диеты спинозавра» . Природа . 429 (6995): 33. Bibcode : 2004natur.429 ... 33b . doi : 10.1038/430033a . PMID 15229562 . S2CID 4398855 .
- ^ Терриен, Ф.; Хендерсон, Д.; Ruff, C. (2005). «Укусить меня - биомеханические модели тероподных мандиблей и последствия для кормления». В Карпентере, К. (ред.). Плотноядные динозавры . Издательство Университета Индианы. С. 179–230. ISBN 978-0-253-34539-4 .
- ^ Anduza, D.; Fowler, DW (2014). «Как спинозавриды могут поймать рыбу? Тестирование поведенческих выводов посредством сравнения с современными тетраподами, питающимися рыбой» . Геологическое общество Америки тезисы с программами . 46 (5): 12.
- ^ Vullo, R.; Allain, R.; Cavin, L. (2016). «Конвергентная эволюция челюстей между спинозавридными динозаврами и щуковыми угрями» . Acta Palaeontologica Polonica . 61 . doi : 10.4202/app.00284.2016 . S2CID 54514619 .
- ^ Баркер, Крис Тиджани; Найш, Даррен; Тенденция, Джейкоб; Мишэльс, Лизанна Вирле; Витмер, Лоуренс; Риджли, Райан; Ранкин, Кэти; Кларкин, Клэр Э.; Шнайдер, Филипп; Гостолинг, Нил Дж. (2023). «Модифицированные черепа, но консервативный мозг? Палеонерология и эндокраниальная анатомия барионишиновых динозавров (Theropoda: spinosauridae)» . Журнал анатомии . 242 (6): 1124–1145. doi : 10.1111/joa.13837 . PMC 10184548 . PMID 36781174 .
- ^ Амиот, Р.; Buffetaut, E.; Lecuyer, C.; Ван, х.; Будад, Л.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Медейрос, Массачусетс; Мо, Дж.; Simon, L.; Suteethorn, v.; Sweetman, S.; Тонг, H.; Чжан, Ф.; Zhou, Z. (2010). «Изотопные доказательства кислорода для полуаватических привычек среди спинозавридов». Геология . 38 (2): 139–142. Bibcode : 2010geo .... 38..139a . doi : 10.1130/g30402.1 .
- ^ Продажи, маф; Lacerda, MB; Рог, блд; de Oliveira, IAP; Schultz, cl; Биби, Ф. (2016). «« Χ »вопроса: проверка взаимосвязи между палеоокружением и тремя тероподскими кладами» . Plos один . 11 (2): E0147031. BIBCODE : 2016PLOSO..1147031S . doi : 10.1371/journal.pone.0147031 . PMC 4734717 . PMID 26829315 .
- ^ Фаббри, Маттео; Навалон, Гильермо; Бенсон, Роджер Б.Дж.; Пол, Диего; О'Коннор, Цзингмай; Bhullar, Bhart-Anjan S.; Эриксон, Грегори М.; Norell, Mark A.; Оркней, Эндрю; Ламанна, Мэтью С.; Зухри, Самир; Беккер, Джастин; Эмке, Аманда; От камня, христианин; Биндиллини, Габриэле; Магануко, Симона; Аудитор, Марко; Ибрагим, Низар (23 марта 2022 г.). «Подвековочное предназначение среди плотоядных динозавров» . Природа . 603 (7903): 852–857. Bibcode : 2022nater.603..852f . Doi : 10.1038/s41586-022-04528-0 . ISSN 1476-4687 . PMID 35322229 . S2CID 247630374 .
- ^ «Спинозавр имел кости, похожие на пингвин, признак охоты под водой» . Наука . 23 марта 2022 года. Архивировано с оригинала 23 марта 2022 года.
- ^ Музей, поле (23 марта 2022 г.). «Плотные кости позволили Спинозауру охотиться под водой, показывает исследование» . Phys.org .
- ^ Рэдли, JD; Аллен, П. (2012). «Южный английский Wealden (не-лайн нижний меловый цвет): обзор палеоэрабов и палеоэкологии». Материалы Ассоциации геологов . 123 (2): 382–385. Bibcode : 2012prga..123..382r . doi : 10.1016/j.pgeola.2011.12.005 .
- ^ Radley, JD (2006). "Wealden Guide I: The Weald Sub-Basin". Геология сегодня . 22 (3): 109–118. Bibcode : 2006geolt..22..109r . doi : 10.1111/j.1365-2451.2006.00563.x . S2CID 129247105 .
- ^ Blows, WT (1998). «Обзор нижних и средних меловых динозавров Англии» . Бюллетени из музея естественной истории Нью -Мексико . 14 : 29–38.
- ^ Кук, E.; Росс, AJ (1996). «Стратиграфия, седиментология и палеонтология нижней глины Weald (Hauterivian) в Keymer Tileworks, Западная Суссекс, Южная Англия». Материалы Ассоциации геологов . 107 (3): 231–239. Bibcode : 1996prga..107..231c . doi : 10.1016/s0016-7878 (96) 80031-9 .
- ^ Милнер, AC; Эванс, SE (1998). «Первый отчет о амфибиях и ящерицах из Wealden (нижний меловой) в Англии» . Бюллетени из музея естественной истории Нью -Мексико . 14 : 173–176.
- ^ Nye, E.; Feist-Burkhardt, S.; Хорн, DJ; Росс, AJ; Whittaker, JE (2008). «Палеоэнакружение, связанное с частичным скелетом игуанодона из верхней глины Weald (Barremian, Early Breatacelous) в Smokejacks Brickworks (Окли, Суррей, Великобритания), на основе палиноморфов и остракод». Мерашные исследования . 29 (3): 417–444. Bibcode : 2008crres..29..417n . doi : 10.1016/j.cretres.2008.01.004 .
- ^ Кук, Э. (1997). «Седиментология и позвоночные тафономии из костей, несущих фации, из магазина Clockhouse Rock (Weald Clay, покойный хаутеривиан), Капель, Суррей, Великобритания». Материалы Ассоциации геологов . 108 (1): 49–56. Bibcode : 1997prga..108 ... 49c . doi : 10.1016/s0016-7878 (97) 80005-3 .
- ^ Росс, AJ; Кук, Э. (1995). «Стратиграфия и палеонтология глины Верхнего Уилда (Barremian) в Smokejacks Brickworks, Окли, Суррей, Англия» (PDF) . Мерашные исследования . 16 (6): 705–716. Bibcode : 1995crres..16..705r . doi : 10.1006/cres.1995.1044 .
- ^ Баттен, DJ (1998). «Палеоэкологические последствия растений, насекомых и других микрофоссилий с органическими стенами в образовании глины Weald (нижние меловые) юго-восточной Англии». Мерашные исследования . 19 (3–4): 279–315. Bibcode : 1998crres..19..279b . doi : 10.1006/cres.1998.0116 .
- ^ Martill, DM; Хатт С. (1996). «Возможные зубы динозавров барионихиды из формирования Wessex (нижний меловой, барремйский) острова Уайт, Англия». Материалы Ассоциации геологов . 107 (2): 81–84. Bibcode : 1996prga..107 ... 81M . doi : 10.1016/s0016-7878 (96) 80001-0 .
Внешние ссылки
[ редактировать ]- Музей естественной истории-« Baryonyx : открытие удивительного динозавра, питающего рыбу» -четырехминутное видео, представленное Анжелой С. Милнер















![]() Эта статья была представлена в Wikijournal of Science для внешнего академического рецензирования в 2018 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на странице Википедии под лицензией CC-By-SA-3.0 ( 2018 ). Версия записи как рецензировано:
Майкл Бех; и др. (2019). "Baryonyx" (PDF) . Викиджурнал науки . 2 (1): 3. DOI : 10.15347/WJS/2019.003 . ISSN 2470-6345 . Wikidata Q63252951 .
Эта статья была представлена в Wikijournal of Science для внешнего академического рецензирования в 2018 году ( отчеты рецензентов ). Обновленный контент был реинтегрирован на странице Википедии под лицензией CC-By-SA-3.0 ( 2018 ). Версия записи как рецензировано:
Майкл Бех; и др. (2019). "Baryonyx" (PDF) . Викиджурнал науки . 2 (1): 3. DOI : 10.15347/WJS/2019.003 . ISSN 2470-6345 . Wikidata Q63252951 .
- Статьи Википедии, опубликованные в рецензируемой литературе
- Статьи Википедии, опубликованные в Wikijournal of Science
- Внешне рецензируемые статьи
- Статьи Википедии, опубликованные в рецензируемой литературе (W2J)
- Спинозавриды
- Монотипические роды динозавров
- Барремальная жизнь
- Ранние меловые динозавры Европы
- Брицеловая Англия
- Окаменелости Англии
- Ископаемые таксоны описаны в 1986 году
- Таксоны, названные Аланом Дж. Чаригом
- Таксоны, названные Анжелой Милнер