Цистатионин-бета-синтаза
| CBS | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CBS , HIP4, цистатионин-бета-синтаза, CBSL, цистатионин-бета-синтаза | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 613381 ; МГИ : 88285 ; Гомологен : 37258 ; GeneCards : CBS ; ОМА : CBS - ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||




Цистатионин-β-синтаза , также известная как CBS , представляет собой фермент ( EC 4.2.1.22 ), который у человека кодируется CBS геном . Он катализирует первый этап пути транссульфурации от гомоцистеина к цистатионину : [ 5 ]

CBS использует кофактор пиридоксальфосфат (PLP) и может аллостерически регулироваться эффекторами, такими как вездесущий кофактор S -аденозил- L -метионин (adoMet). Этот фермент принадлежит к семейству лиаз , а именно гидролиаз, расщепляющих углерод-кислородные связи.
CBS представляет собой многодоменный фермент, состоящий из N-концевого ферментативного домена и двух доменов CBS . Ген CBS является наиболее распространенным локусом мутаций, связанных с гомоцистинурией . [ 6 ]
Номенклатура
[ редактировать ]Систематическое название этого класса ферментов — L -серингидролиаза (добавляющая гомоцистеин; L -цистатионинобразующая). Другие широко используемые имена включают:
- β-тионаза,
- цистеинсинтаза,
- L -серингидролиаза (с добавлением гомоцистеина),
- метилцистеинсинтаза,
- серинсульфгидраза и
- серинсульфгидрилаза.
В 1961 году метилцистеинсинтазе был присвоен номер ЕС EC 4.2.1.23. Это вызвало побочная реакция CBS. Номер ЕС EC 4.2.1.23 был удален в 1972 году. [ 7 ]
Структура
[ редактировать ]
Фермент цистатионин-β-синтаза человека представляет собой тетрамер и состоит из 551 аминокислоты с молекулярной массой субъединицы 61 кДа. Он представляет собой модульную организацию из трех модулей с N-концевым гемовым доменом, за которым следует ядро, содержащее кофактор PLP . [ 9 ] Кофактор находится глубоко в гемовом домене и связан основанием Шиффа. [ 10 ] Основание Шиффа представляет собой функциональную группу, содержащую связь C=N с атомом азота, соединенным с арильной или алкильной группой. Гемовый домен состоит из 70 аминокислот, и похоже, что гем существует только в CBS млекопитающих и отсутствует в CBS дрожжей и простейших . На С-конце регуляторный домен CBS содержит тандемный повтор двух доменов CBS β-α-β-β-α, мотива вторичной структуры, обнаруженного в других белках. [ 9 ] CBS имеет C-концевой ингибирующий домен. С-концевой домен цистатионин-β-синтазы регулирует ее активность посредством как внутристерических, так и аллостерических эффектов и важен для поддержания тетрамерного состояния белка. [ 9 ] Это ингибирование облегчается связыванием аллостерического эффектора adoMet или удалением регуляторного домена; однако масштабы эффектов различаются. [ 9 ] Мутации в этом домене коррелируют с наследственными заболеваниями . [ 11 ]
Гемовый домен содержит N-концевую петлю, которая связывает гем и обеспечивает аксиальные лиганды C52 и H65. Расстояние гема от сайта связывания PLP предполагает его неучастие в катализе, однако удаление гема-домена приводит к потере окислительно-восстановительной чувствительности, поэтому предполагается, что гем является окислительно-восстановительным сенсором. [ 10 ] Присутствие протопорфирина IX в CBS является уникальным PLP-зависимым ферментом и обнаруживается только в CBS млекопитающих. D. melanogaster и D. diskoides имеют усеченные N-концевые отростки и, следовательно, предотвращают образование консервативных гистидиновых и цистеиновых гемовых лигандов остатков . Однако последовательность Anopheles gambiae имеет более длинное N-концевое удлинение, чем человеческий фермент, и содержит консервативные остатки гистидинового и цистеинового гема- человека лиганда, такие как гем . Следовательно, возможно, что CBS у слизевиков и насекомых представляют собой гемепротеины, что позволяет предположить, что гемовый домен является ранней эволюционной инновацией, возникшей до разделения животных и слизевиков. [ 9 ] PLP с K119 представляет собой внутренний альдимин и образует основание Шиффа в активном центре. Между каталитическим и регуляторным доменами существует сверхчувствительный сайт, который вызывает протеолитическое расщепление и продуцирует укороченный димерный фермент, который более активен, чем исходный фермент. Как укороченный фермент, так и фермент, обнаруженный в дрожжах, не регулируются adoMet. Дрожжевой фермент также активируется за счет удаления С-конца с образованием димерного фермента. [ 9 ]
По состоянию на конец 2007 года две структуры для этого класса ферментов были решены PDB с кодами доступа 1JBQ и 1M54 .
Ферментативная активность
[ редактировать ]| цистатионин бета-синтаза | |||
|---|---|---|---|
 Гомодимер цистатионин-бета-синтазы, человек | |||
| Идентификаторы | |||
| Номер ЕС. | 4.2.1.22 | ||
| Номер CAS. | 9023-99-8 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
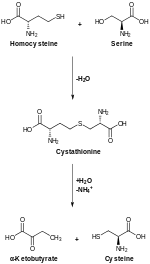
Транссульфурация, катализируемая CBS, превращает гомоцистеин в цистатионин , который цистатион-гамма-лиаза превращает в цистеин . [ 12 ]
CBS занимает ключевую позицию в метаболизме серы у млекопитающих в гомоцистеиновом соединении, где принимается решение сохранить метионин или превратить его в цистеин через путь транссульфурации . Более того, путь транссульфурации является единственным путем, способным удалять серосодержащие аминокислоты в условиях избытка. [ 9 ]
По аналогии с другими β-замещающими ферментами, в реакции, катализируемой CBS, предполагается участие ряда промежуточных продуктов, связанных с adoMet . Добавление серина приводит к реакции трансшиффизации , в результате которой образуется внешний альдимин . Альдимин промежуточного подвергается отщеплению протона у α-углерода с последующим отщеплением с образованием аминоакрилата . Нуклеофильная атака тиолата гомоцистеина на аминоакрилат и репротонирование Cα приводят к образованию внешнего альдимина цистатионина . Заключительная реакция трансальдиминирования высвобождает конечный продукт — цистатионин. [ 9 ] Конечный продукт, L -цистатионин, также может образовывать промежуточный аминоакрилат, что указывает на то, что вся реакция CBS обратима. [ 13 ]
Измеренное значение V 0 ферментативно-катализируемой реакции, как правило, отражает устойчивое состояние (где [ES] является постоянным), даже несмотря на то, что V 0 ограничено ранней частью реакции, и анализ этих начальных скоростей называется как стационарная кинетика. Стационарный кинетический анализ дрожжевого CBS дает параллельные линии. Эти результаты согласуются с предложенным механизмом пинг-понга, в котором за связыванием серина и высвобождением воды следует связывание гомоцистеина и высвобождение цистатионина. Напротив, кинетика фермента CBS в устойчивом состоянии крысы дает пересекающиеся линии, что указывает на то, что β-заместитель серина не высвобождается из фермента до связывания гомоцистеина. [ 9 ]
Одной из альтернативных реакций с участием CBS является конденсация цистеина с гомоцистеином с образованием цистатионина и сероводорода (H 2 S). [ 13 ] H 2 S в мозге вырабатывается из L -цистеина под действием CBS. Этот альтернативный метаболический путь также зависит от adoMet . [ 14 ]
Ферментативная активность CBS обнаруживается не во всех тканях и клетках. Он отсутствует в сердце, легких, семенниках, надпочечниках и селезенке у крыс. Было показано, что у людей он отсутствует в сердечной мышце и первичных культурах эндотелиальных клеток аорты человека. Отсутствие CBS в этих тканях означает, что эти ткани не способны синтезировать цистеин и что цистеин должен поступать из внеклеточных источников. Это также предполагает, что эти ткани могут иметь повышенную чувствительность к токсичности гомоцистеина, поскольку они не могут катаболизировать избыток гомоцистеина посредством транссульфурации. [ 13 ]
Регулирование
[ редактировать ]Аллостерическая активация CBS с помощью adoMet определяет метаболическую судьбу гомоцистеина . CBS млекопитающих активируется AdoMet в 2,5-5 раз с константой диссоциации 15 мкМ. [ 6 ] AdoMet представляет собой аллостерический активатор, который увеличивает V max реакции CBS, но не влияет на K m субстратов. Другими словами, AdoMet стимулирует активность CBS за счет увеличения скорости оборота, а не за счет связывания субстратов с ферментом. [ 9 ] Этот белок может использовать морфеиновую модель аллостерической регуляции . [ 15 ]
Человеческий CBS выполняет решающий этап биосинтетического пути цистеина , обеспечивая точку регуляторного контроля для AdoMet. Гомоцистеин после метилирования в метионин может быть преобразован в AdoMet, который передает метильные группы различным субстратам, например, нейротрансмиттерам , белкам и нуклеиновым кислотам . AdoMet действует как аллостерический активатор CBS и контролирует его биосинтез: низкие концентрации AdoMet приводят к низкой активности CBS, тем самым направляя гомоцистеин в цикл трансметилирования для образования AdoMet. Напротив, высокие концентрации adoMet направляют гомоцистеин по пути транссульфурации к биосинтезу цистеина . [ 16 ]
У млекопитающих CBS представляет собой строго регулируемый фермент, который содержит гем- кофактор, который действует как окислительно-восстановительный сенсор. [ 11 ] который может модулировать его активность в ответ на изменения окислительно-восстановительного потенциала. Если в покоящейся форме CBS в клетке имеется железо (Fe 2+ ) гем, существует возможность активации фермента в окислительных условиях путем превращения в трехвалентное железо (Fe 3+ ) состояние. [ 9 ] Фе 2+ форма фермента ингибируется при связывании CO или оксида азота, тогда как активность фермента удваивается, когда Fe 2+ окисляется до Fe 3+ . Окислительно-восстановительное состояние гема зависит от pH, при этом происходит окисление Fe. 2+ –CBS в Fe 3+ – CBS предпочтителен в условиях низкого pH. [ 17 ]
Поскольку CBS млекопитающих содержит кофактор гема, тогда как ферменты дрожжей и простейших Trypanosoma cruzi не имеют кофакторов гема, исследователи предположили, что гем не требуется для активности CBS. [ 9 ]
CBS регулируется на уровне транскрипции с помощью NF-Y , SP-1 и SP-3 . Кроме того, его транскрипционная регуляция усиливается глюкокортикоидами и гликогеном и подавляется инсулином . Метионин активирует CBS на посттранскрипционном уровне.
Болезнь человека
[ редактировать ]Синдром Дауна — это заболевание, характеризующееся сверхэкспрессией цистатионин-бета-синтазы (CBS) и низким уровнем гомоцистеина в крови. Было высказано предположение, что сверхэкспрессия цистатионин-бета-синтазы может быть основным виновником этого заболевания (наряду с дисфункцией GabaA и Dyrk1a). Фенотип синдрома Дауна противоположен гипергомоцистеинемии (описанной ниже). Фармакологические ингибиторы CBS запатентованы Фондом Жерома Лежена (ноябрь 2011 г.) и запланированы испытания (на животных и людях).
Гипергомоцистеинемия — это заболевание, характеризующееся аномально высоким уровнем гомоцистеина в крови. Мутации CBS являются единственной наиболее распространенной причиной наследственной гипергомоцистеинемии. Генетические дефекты, влияющие на ферментные пути MTHFR , MTR и MTRR/MS, также могут способствовать повышению уровня гомоцистеина. Врожденные ошибки CBS приводят к гипергомоцистеинемии с осложнениями на сердечно-сосудистую систему, приводящими к раннему и агрессивному заболеванию артерий. Гипергомоцистеинемия также поражает три другие основные системы органов, включая глазную, центральную нервную и скелетную. [ 9 ]
Гомоцистинурия, обусловленная дефицитом CBS, представляет собой особый тип гипергомоцистеинемии. Это редкое наследственное рецессивное аутосомное заболевание, которое обычно диагностируется в детстве. Всего была идентифицирована 131 различная мутация, вызывающая гомоцистинурию. Общей функциональной особенностью мутаций в доменах CBS является то, что мутации отменяют или сильно снижают активацию adoMet . [ 16 ] Специфического лечения гомоцистинурии не обнаружено; однако многие люди лечатся высокими дозами витамина B6 , который является кофактором CBS.
Биоинженерия
[ редактировать ]Цистатионин-бета-синтаза (CBS) участвует в развитии ооцитов . Однако мало что известно о региональных и клеточных паттернах экспрессии CBS в яичниках, и в настоящее время исследования сосредоточены на определении местоположения и экспрессии во время развития фолликула в яичниках. [ 18 ]
Отсутствие цистатионин-бета-синтазы у мышей провоцирует бесплодие из-за потери экспрессии белка в матке. [ 19 ]
Мутации
[ редактировать ]Гены, контролирующие экспрессию фермента CBS, могут не работать со 100% эффективностью у людей, у которых есть один из SNP ( однонуклеотидный полиморфизм , тип мутаций ), влияющий на этот ген. Известные варианты включают SNP A360A, C699T, I278T, N212N и T42N (среди прочих). Эти SNP, которые по-разному влияют на эффективность фермента, можно обнаружить с помощью стандартных методов тестирования ДНК.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000160200 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000024039 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ген Энтреза: цистатионин-бета-синтаза CBS» .
- ^ Jump up to: а б Яношик М., Кери В., Гаустаднес М., Маклин К.Н., Краус Дж.П. (сентябрь 2001 г.). «Регуляция цистатионин-бета-синтазы человека с помощью S-аденозил-L-метионина: доказательства наличия двух каталитически активных конформаций, включающих аутоингибирующий домен в С-концевой области». Биохимия . 40 (35): 10625–33. дои : 10.1021/bi010711p . ПМИД 11524006 .
- ^ ЕС 4.2.1.23
- ^ PDB : 1JBQ ; Мейер М., Яношик М., Кери В., Краус Дж. П., Буркхард П. (август 2001 г.). «Структура человеческой цистатионин-бета-синтазы: уникальный пиридоксаль-5'-фосфат-зависимый гем-белок» . Журнал ЭМБО . 20 (15): 3910–6. дои : 10.1093/emboj/20.15.3910 . ПМК 149156 . ПМИД 11483494 .
- ^ Jump up to: а б с д и ж г час я дж к л м Банерджи Р., Цзоу К.Г. (январь 2005 г.). «Окислительно-восстановительная регуляция и механизм реакции цистатионин-бета-синтазы человека: PLP-зависимый гемесенсорный белок». Архив биохимии и биофизики . 433 (1): 144–56. дои : 10.1016/j.abb.2004.08.037 . ПМИД 15581573 .
- ^ Jump up to: а б Яманиши М., Кабил О., Сен С., Банерджи Р. (декабрь 2006 г.). «Структурные данные о патогенных мутациях гем-зависимой цистатионин-бета-синтазы». Журнал неорганической биохимии . 100 (12): 1988–95. дои : 10.1016/j.jinorgbio.2006.08.020 . ПМИД 17069888 .
- ^ Jump up to: а б Кабил О., Чжоу Ю., Банерджи Р. (ноябрь 2006 г.). «Человеческая цистатионин-бета-синтаза является мишенью сумойлирования». Биохимия . 45 (45): 13528–36. дои : 10.1021/bi0615644 . ПМИД 17087506 .
- ^ Нодзаки Т., Сигета Ю., Сайто-Накано Ю., Имада М., Крюгер В.Д. (март 2001 г.). «Характеристика транссульфурации и путей биосинтеза цистеина у простейших гемофлагеллят Trypanosoma cruzi. Выделение и молекулярная характеристика цистатионин-бета-синтазы и серин-ацетилтрансферазы из Trypanosoma» . Журнал биологической химии . 276 (9): 6516–23. дои : 10.1074/jbc.M009774200 . ПМИД 11106665 .
- ^ Jump up to: а б с Джи К.Х., Крюгер В.Д. (2005). «Роль цистатионин-бета-синтазы в метаболизме гомоцистеина». Антиоксиданты и окислительно-восстановительная сигнализация . 7 (5–6): 813–22. дои : 10.1089/ars.2005.7.813 . ПМИД 15890029 .
- ^ Это К., Кимура Х. (ноябрь 2002 г.). «Новый механизм усиления сероводородпродуцирующей активности цистатионин-бета-синтазы» . Журнал биологической химии . 277 (45): 42680–5. дои : 10.1074/jbc.M205835200 . ПМИД 12213817 .
- ^ Т. Селвуд и Э. К. Яффе (2011). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Арх. Биохим. Биофиз . 519 (2): 131–43. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Jump up to: а б Иньул С., Эггермонт Дж. (декабрь 2005 г.). «Домены CBS: структура, функции и патология в белках человека» . Американский журнал физиологии. Клеточная физиология . 289 (6): C1369–78. doi : 10.1152/ajpcell.00282.2005 . ПМИД 16275737 .
- ^ Пураник М., Уикс С.Л., Лахайе Д., Кабил О., Таока С., Нильсен С.Б., Гровс Дж.Т., Банерджи Р., Спиро Т.Г. (май 2006 г.). «Динамика связывания монооксида углерода с бета-синтазой цистатионина» . Журнал биологической химии . 281 (19): 13433–8. дои : 10.1074/jbc.M600246200 . ПМЦ 2745537 . ПМИД 16505479 .
- ^ Лян Р., Ю В.Д., Ду Дж.Б., Ян Л.Дж., Шан М., Го Цз.З. (ноябрь 2006 г.). «Локализация цистатионин-бета-синтазы в яичниках мышей и профиль ее экспрессии во время развития фолликулов» . Китайский медицинский журнал . 119 (22): 1877–83. дои : 10.1097/00029330-200611020-00006 . ПМИД 17134586 . S2CID 23891500 .
- ^ Гусман М.А., Наварро М.А., Карнисер Р., Саррия А.Дж., Асин С., Арнал С., Муниеса П., Сурра Х.К., Арбонес-Майнар Х.М., Маеда Н., Осада Дж. (ноябрь 2006 г.). «Цистатионин-бета-синтаза необходима для женской репродуктивной функции» . Молекулярная генетика человека . 15 (21): 3168–76. дои : 10.1093/hmg/ddl393 . ПМИД 16984962 .
Дальнейшее чтение
[ редактировать ]- Краус Дж. П. (1994). «Лекция Комровера. Молекулярные основы проявления фенотипа при гомоцистинурии». Дж. Наследовать. Метаб. Дис . 17 (4): 383–90. дои : 10.1007/BF00711354 . ПМИД 7967489 . S2CID 42317828 .
- Краус Дж.П., Яношик М., Козич В. и др. (1999). «Мутации цистатионин-бета-синтазы при гомоцистинурии». Хм. Мутировать . 13 (5): 362–75. doi : 10.1002/(SICI)1098-1004(1999)13:5<362::AID-HUMU4>3.0.CO;2-K . ПМИД 10338090 . S2CID 86447340 .
- Джонс А.Л. (1999). «Локализация и взаимодействие хантингтина» . Филос. Пер. Р. Сок. Лонд. Б Биол. Наука . 354 (1386): 1021–7. дои : 10.1098/rstb.1999.0454 . ПМК 1692601 . ПМИД 10434301 .
- Гриффитс Р., Тадбол Н. (1977). «Молекулярный дефект в случае гомоцистинурии с дефицитом (цистатионин-бета-синтазы)» . Евро. Дж. Биохим . 74 (2): 269–73. дои : 10.1111/j.1432-1033.1977.tb11390.x . ПМИД 404147 .
- Краус Дж., Пакман С., Фаулер Б., Розенберг Л.Е. (1978). «Очистка и свойства цистатионин-бета-синтазы из печени человека. Доказательства идентичных субъединиц» . Ж. Биол. Хим . 253 (18): 6523–8. дои : 10.1016/S0021-9258(19)46963-9 . ПМИД 681363 .
- Лонги Р.К., Флейшер Л.Д., Таллан Х.Х., Галл Г.Е. (1977). «Дефицит цистатионин-бета-синтазы: качественное нарушение дефицитного фермента, модифицированное терапией витамином B6 » . Педиатр. Рез . 11 (2): 100–3. дои : 10.1203/00006450-197702000-00003 . ПМИД 840498 .
- Козич В., Краус Дж. П. (1993). «Скрининг мутаций путем экспрессии сегментов кДНК пациента в E. coli: гомоцистинурия из-за дефицита цистатионин-бета-синтазы» . Хм. Мутат . 1 (2): 113–23. дои : 10.1002/humu.1380010206 . ПМИД 1301198 . S2CID 36663527 .
- Мюнке М, Краус Дж. П., Охура Т., Франк У (1988). «Ген цистатионин-бета-синтазы (CBS) картируется в субтеломерной области хромосомы 21q человека и в проксимальной хромосоме 17 мыши» . Являюсь. Дж. Хум. Жене . 42 (4): 550–9. ПМЦ 1715237 . ПМИД 2894761 .
- Ху Ф.Л., Гу З., Козич В. и др. (1994). «Молекулярные основы дефицита цистатионин-бета-синтазы при пиридоксин-чувствительной и нечувствительной гомоцистинурии». Хм. Мол. Жене . 2 (11): 1857–60. дои : 10.1093/hmg/2.11.1857 . ПМИД 7506602 .
- Сперандео М.П., Панико М., Пепе А. и др. (1995). «Молекулярный анализ пациентов, страдающих гомоцистинурией из-за дефицита цистатионин-бета-синтазы: сообщение о новой мутации в экзоне 8 и делеции в интроне 11». Дж. Наследовать. Метаб. Дис . 18 (2): 211–4. дои : 10.1007/BF00711769 . ПМИД 7564249 . S2CID 40407615 .
- Шассе Ж.Ф., Пали Э., Пэрис Д. и др. (1995). «Геномная организация гена цистатионин-бета-синтазы человека: данные о различных кДНК». Биохим. Биофиз. Рез. Коммун . 211 (3): 826–32. дои : 10.1006/bbrc.1995.1886 . ПМИД 7598711 .
- Ши В.Е., Фрингер Дж.М., Манделл Р. и др. (1995). «Миссенс-мутация (I278T) в гене цистатионин-бета-синтазы, распространенная при пиридоксин-чувствительной гомоцистинурии и связанная с легким клиническим фенотипом» . Являюсь. Дж. Хум. Жене . 57 (1): 34–9. ПМК 1801250 . ПМИД 7611293 .
- Клюйтманс Л.А., Блом Х.Дж., Бурс Г.Х. и др. (1995). «Две новые миссенс-мутации в гене цистатионин-бета-синтазы у пациентов с гомоцистинурией». Хм. Жене . 96 (2): 249–50. дои : 10.1007/BF00207394 . ПМИД 7635485 . S2CID 6642338 .
- Себастио Г., Сперандео М.П., Панико М. и др. (1995). «Молекулярная основа гомоцистинурии из-за дефицита цистатионин-бета-синтазы в итальянских семьях и сообщение о четырех новых мутациях» . Являюсь. Дж. Хум. Жене . 56 (6): 1324–33. ПМК 1801112 . ПМИД 7762555 .
- Марбл М., Джерати М.Т., де Франшис Р. и др. (1995). «Характеристика аллеля цистатионин-бета-синтазы с тремя цис-мутациями у пациента с невосприимчивой к B 6 гомоцистинурией». Хм. Мол. Жене . 3 (10): 1883–6. дои : 10.1093/hmg/3.10.1883 . ПМИД 7849717 .
- Краус Дж.П., Ле К., Сваруп М. и др. (1994). «КДНК цистатионин-бета-синтазы человека: последовательность, альтернативный сплайсинг и экспрессия в культивируемых клетках». Хм. Мол. Жене . 2 (10): 1633–8. дои : 10.1093/hmg/2.10.1633 . ПМИД 7903580 .
- де Франшис Р., Козич В., Макиннес Р.Р., Краус Дж.П. (1995). «Идентичные генотипы у братьев и сестер с разными гомоцистинурическими фенотипами: идентификация трех мутаций в цистатионин-бета-синтазе с использованием улучшенной бактериальной системы экспрессии». Хм. Мол. Жене . 3 (7): 1103–8. дои : 10.1093/hmg/3.7.1103 . ПМИД 7981678 .
- Крюгер В.Д., Кокс Д.Р. (1994). «Дрожжевая система экспрессии цистатионин-бета-синтазы человека: структурная и функциональная консервативность генов человека и дрожжей» . Учеб. Натл. акад. наук. США . 91 (14): 6614–8. Бибкод : 1994PNAS...91.6614K . дои : 10.1073/pnas.91.14.6614 . ПМК 44253 . ПМИД 8022826 .
- Козич В., де Франшис Р., Краус Дж. П. (1993). «Молекулярный дефект у пациента с пиридоксин-чувствительной гомоцистинурией». Хм. Мол. Жене . 2 (6): 815–6. дои : 10.1093/hmg/2.6.815 . ПМИД 8353501 .
Внешние ссылки
[ редактировать ]- Главная страница CBS в Центре медицинских наук Университета Колорадо
- Цистатионин-бета-синтаза в BRENDA: Комплексная информационная система ферментов [ постоянная мертвая ссылка ]
- Цистатионин-бета-синтаза: запись в банке данных белков
галерея PDB |
|---|

