Карнотавр
| Карнотавр | |
|---|---|

| |
| Установленный слепок скелета в музее Хлупача в Праге. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Абелизавриды |
| Клэйд : | † Фурилеусаура |
| Племя: | † Карнотаурини |
| Род: | † Карнотавр Бонапарт , 1985 год. |
| Разновидность: | † К. састрей
|
| Биномиальное имя | |
| † Карнотавр састрей Бонапарт, 1985 год.
| |
Карнотавр ( / ˌ k ɑːr n oʊ ˈ t ɔːr ə s / ; букв. « мясной бык » ) — род динозавров теропод - , который жил в Южной Америке в позднемеловой период, вероятно, где-то между 72 и 69 миллионами лет назад. Единственный вид — Carnotaurus sastrei . Известный по единственному хорошо сохранившемуся скелету, это один из наиболее изученных теропод Южного полушария . Скелет, найденный в 1984 году, был обнаружен в провинции Чубут в Аргентине в породах формации Ла Колония . Карнотавр — производный член Abelisauridae , группы крупных теропод, занимавших большую хищную нишу на южных территориях Гондваны в позднем меловом периоде . В составе Abelisauridae этот род часто считается членом Brachyrostra, клады короткорылых форм, обитающих в Южной Америке.
Карнотавр был легким двуногим хищником длиной от 7,5 до 8 м (от 24,6 до 26,2 футов) и весом 1,3–2,1 метрических тонны (1,4–2,3 коротких тонны; 1,3–2,1 длинных тонны). Как теропод, Карнотавр был узкоспециализированным и самобытным. У него были толстые рога над глазами, особенность, невиданная у всех других хищных динозавров, и очень глубокий череп, сидящий на мускулистой шее. Карнотавр также характеризовался маленькими рудиментарными передними конечностями и длинными тонкими задними конечностями. Скелет сохранился с обширными отпечатками кожи, представляющими собой мозаику из мелких неперекрывающихся чешуек диаметром примерно 5 мм. Мозаика прерывалась крупными бугорками по бокам животного, никаких намеков на перья нет.
Отличительные рога и мускулистая шея, возможно, использовались в борьбе с сородичами . Согласно отдельным исследованиям, соперничающие особи могли драться друг с другом быстрыми ударами по голове, медленными толчками верхней частью черепа или таранами друг друга в лоб, используя рога в качестве амортизаторов. Пищевые привычки карнотавра остаются неясными: некоторые исследования показали, что животное могло выслеживать очень крупную добычу, такую как зауроподы , в то время как другие исследования показали, что оно охотилось в основном на относительно мелких животных. Полость его мозга предполагает острое обоняние, тогда как слух и зрение были менее развиты. Карнотавр , вероятно, был хорошо приспособлен к бегу и, возможно, был одним из самых быстрых крупных теропод.
Открытие
[ редактировать ]
Единственный скелет ( голотип MACN -CH 894) был раскопан в 1984 году экспедицией под руководством аргентинского палеонтолога Хосе Бонапарта . [ А ] Эта экспедиция также обнаружила своеобразного колючего зауропода Amargasaurus . [ 3 ] Это была восьмая экспедиция в рамках проекта «Юрские и меловые наземные позвоночные Южной Америки», стартовавшего в 1976 году и спонсируемого Национальным географическим обществом . [ 3 ] [ Б ] Скелет хорошо сохранился и сочленён (все еще соединен вместе), при этом только задние две трети хвоста, большая часть голени и задние лапы были разрушены в результате выветривания . [ С ] [ 5 ] Скелет принадлежал взрослой особи, о чем свидетельствуют сросшиеся швы в черепной коробке . [ 6 ] Его нашли лежащим на правом боку в типичной позе смерти с согнутой назад шеей над туловищем. [ 7 ] Необычно то, что он сохранился с обширными отпечатками кожи. [ Д ] Учитывая значимость этих отпечатков, была начата вторая экспедиция по повторному исследованию первоначального места раскопок, в результате которой было обнаружено несколько дополнительных участков кожи. [ 7 ] Череп был деформирован во время окаменения: кости морды левой стороны смещены вперед относительно правой стороны, носовые кости выдвинуты вверх, а предчелюстные кости выдвинуты назад на носовые кости . Деформация также усилила искривление верхней челюсти вверх. [ Э ] Морда пострадала от деформации сильнее, чем задняя часть черепа, возможно, из-за большей жесткости последней. При виде сверху или снизу верхние челюсти имели менее U-образную форму, чем нижние, что приводило к явному несоответствию. Это несоответствие является результатом действующей с боков деформации, которая затронула верхние челюсти, но не затронула нижние, возможно, из-за большей гибкости суставов внутри последних. [ 1 ]

Скелет был собран на ферме под названием «Почо Састре» недалеко от Бахада Морено в департаменте Тельсен провинции Чубут , Аргентина. [ 5 ] Поскольку он был заключен в крупную гематитовую конкрецию , очень твердую породу, подготовка была сложной и продвигалась медленно. [ 9 ] [ 5 ] В 1985 году Бонапарт опубликовал заметку, в которой Carnotaurus sastrei представлялся как новый род и вид и кратко описывал череп и нижнюю челюсть. [ 5 ] Родовое название Carnotaurus происходит от латинского carno [carnis] («плоть») и taurus («бык») и может быть переведено как «бык-мясоед», что является намеком на бычьи рога животного. [ 10 ] Видовое название sastrei дано в честь Анхеля Састре, владельца ранчо, где был найден скелет. [ 11 ] Подробное описание всего скелета последовало в 1990 году. [ 4 ] После Абелизавра . Карнотавр был вторым открытым представителем семейства Abelisauridae [ 12 ] В течение многих лет он был, безусловно, самым изученным представителем своего семейства, а также самым изученным тероподом из Южного полушария . [ 13 ] [ 14 ] Лишь в 21 веке подобные хорошо сохранившиеся абелизавриды были описаны , в том числе ауказавр , майюнгазавр и скорпиовенатор , что позволило ученым по-новому оценить некоторые аспекты анатомии карнотавра . [ Ф ] Скелет-голотип выставлен в музее естественных наук Аргентинском Бернардино Ривадавиа ; [ Г ] реплики можно увидеть в этом и других музеях мира. [ 15 ] Скульпторы Стивен и Сильвия Черкас изготовили скульптуру Карнотавра в натуральную величину , которая ранее выставлялась в Музее естественной истории округа Лос-Анджелес . Эта скульптура, заказанная музеем в середине 1980-х годов, вероятно, является первой реставрацией теропода, демонстрирующей точную кожу. [ 7 ] [ 16 ]
Описание
[ редактировать ]
Карнотавр был крупным, но легковесным хищником. [ 17 ] Единственная известная особь имела длину около 7,5–8 м (24,6–26,2 футов). [ Ч ] [ я ] [ 19 ] что делает Карнотавра одним из крупнейших абелизаврид. [ Дж ] [ К ] [ 19 ] Экриксинатозавр и, возможно, абелизавр , которые являются крайне неполными, могли быть такими же или даже больше по размеру. [ Л ] [ М ] [ Н ] Исследование 2016 года показало, что только пикнонемозавр длиной 8,9 м (29,2 фута) был длиннее карнотавра ; его высота оценивалась в 7,8 м (25,6 футов). [ 21 ] Его масса оценивается в 1350 кг (1,33 длинных тонны; 1,49 коротких тонны). [ О ] 1500 кг (1,5 длинных тонны; 1,7 коротких тонны), [ П ] 2000 кг (2,0 длинных тонны; 2,2 коротких тонны), [ 19 ] 2100 кг (2,1 длинных тонны; 2,3 коротких тонны), [ Вопрос ] и 1306–1743 кг (1,285–1,715 длинных тонн; 1,440–1,921 коротких тонн) [ 24 ] в отдельных исследованиях, в которых использовались разные методы оценки. Карнотавр был узкоспециализированным тероподом , что особенно заметно по характеристикам черепа , позвонков и передних конечностей. [ Р ] и таз С другой стороны, задние конечности оставались относительно консервативными, напоминая таковые у более базального цератозавра . И таз, и задние конечности были длинными и тонкими. Длина левой бедренной кости (бедренной кости) человека составляет 103 см, но средний диаметр составляет всего 11 см. [ С ]
Череп
[ редактировать ]
Череп длиной 59,6 см (23,5 дюйма) был пропорционально короче и глубже, чем у любого другого крупного плотоядного динозавра. [ Т ] [ В ] Морда была умеренно широкой, а не такой сужающейся, как у более базальных теропод, таких как цератозавр , а челюсти были изогнуты вверх. [ 25 ] Выдающаяся пара рогов наискосок выступала над глазами. Эти рога, образованные лобными костями, [ V ] были толстыми, конусообразными, твердыми внутри, несколько сплюснутыми по вертикали в поперечном сечении и имели длину 15 см (5,9 дюйма). [ 6 ] [ 1 ] Бонапарт в 1990 году предположил, что эти рога, вероятно, образовывали костные ядра гораздо более длинных кератиновых оболочек. [ В ] Маурисио Черрони и его коллеги в 2020 году согласились с тем, что рога поддерживают кератиновые оболочки, но утверждали, что эти оболочки не были намного длиннее костных ядер. [ 1 ]
Как и у других динозавров, череп был продырявлен шестью крупными отверстиями с каждой стороны. Самое переднее из этих отверстий, наружная ноздря (костная ноздря), была подпрямоугольной и направлена в стороны и вперед, но не была наклонной при взгляде сбоку, как у некоторых других цератозавров, таких как Ceratosaurus . Это отверстие было образовано только носовой и предчелюстной костью, тогда как у некоторых родственных цератозавров этому отверстию способствовала и верхняя челюсть. Между костной ноздрей и глазницей (глазничным отверстием) располагалось анторбитальное окно . У Карнотавра это отверстие было выше длины, а у родственных форм, таких как Скорпиовенатор и Майюнгазавр , оно было длиннее высоты . Анторбитальное окно было ограничено более крупным углублением — анорбитальной ямкой , образованной утопленными частями верхней челюсти спереди и слезной частью сзади. Как и у всех абелизаврид, у Карнотавра это углубление было небольшим . В нижнем переднем углу предглазничной ямки имелось меньшее отверстие — промаксиллярное окно , которое вело в заполненную воздухом полость внутри верхней челюсти. [ 1 ] Глаз располагался в верхней части глазницы в форме замочной скважины. [ Х ] Эта верхняя часть была пропорционально небольшой, субкруглой и отделена от нижней части глазницы выступающей вперед заглазничной костью . [ 1 ] Он был слегка повёрнут вперёд, вероятно, обеспечивая некоторую степень бинокулярного зрения . [ И ] Форма глазницы, напоминающая замочную скважину, возможно, была связана с заметным укорочением черепа и также встречается у родственных короткорылых абелизаврид. [ 1 ] Как и у всех абелизаврид, лобная кость (на крыше черепа между глазами) была исключена из орбиты. Позади глазницы находились два отверстия: подвисочное окно сбоку и надвисочное окно на верхушке черепа. Подвисочное окно было высоким, коротким и почковидным, тогда как надвисочное окно было коротким и квадратной формы. Другое отверстие, нижнечелюстное окно , располагалось в нижней челюсти – у карнотавра это отверстие было сравнительно большим. [ 1 ]

На каждой стороне верхних челюстей было по четыре предчелюстных и двенадцать верхнечелюстных зубов. [ С ] в то время как нижние челюсти были оснащены пятнадцатью зубными зубами с каждой стороны. [ АА ] [ 1 ] Зубы были описаны как длинные и тонкие. [ 9 ] в отличие от очень коротких зубов, наблюдаемых у других абелизаврид. [ 25 ] Однако Черрони и его коллеги в своем описании черепа в 2020 году заявили, что все прорезавшиеся зубы были сильно повреждены во время раскопок и позже были реконструированы с помощью гипса (Бонапарте в 1990 году отметил только, что некоторые зубы нижней челюсти были фрагментированы). [ 1 ] [ АБ ] Таким образом, надежная информация о форме зубов ограничивается замененными зубами и корнями зубов, которые все еще закрыты челюстью, и могут быть изучены с помощью компьютерной томографии. [ 1 ] Замещающие зубы имели низкие, уплощенные коронки , были близко расположены и наклонены вперед примерно на 45°. [ 1 ] В своем описании 1990 года Бонапарт отметил, что нижняя челюсть неглубокая и слабо построенная, а зубная кость (передняя челюстная кость) соединена с задними челюстными костями только двумя точками контакта; это контрастирует с крепким на вид черепом. [ 9 ] [ И ] Вместо этого Черрони и его коллеги обнаружили множественные, но слабые связи между зубной костью и костями задней челюсти. Таким образом, эта артикуляция была очень гибкой, но не обязательно слабой. [ 1 ] Нижний край зубной кости был выпуклым, тогда как у Majungasaurus он был прямым . [ 1 ]

На нижней челюсти были обнаружены окостеневшие подъязычные кости в том положении, в котором они находились бы, если бы животное было живо. Эти тонкие кости, поддерживающие мускулатуру языка и некоторые другие мышцы, редко встречаются у динозавров, поскольку они часто хрящевые , не соединены с другими костями и поэтому легко теряются. [ ОБЪЯВЛЕНИЕ ] [ 26 ] [ 1 ] У карнотавра сохранились три подъязычные кости: пара изогнутых стержневидных цератобранхиальных костей, которые сочленяются с единственным трапециевидным элементом — базихиелем. Карнотавр - единственный известный нептичий теропод, от которого известен базихял. [ 1 ] Задняя часть черепа имела хорошо развитые, заполненные воздухом камеры, окружающие черепную коробку, как и у других абелизаврид. Имелись две отдельные системы камер: паратимпанальная система, сообщающаяся с полостью среднего уха , а также камеры, образующиеся из выростов воздушных мешков шеи. [ 24 ]
В черепе можно обнаружить ряд аутапоморфий (отличительных черт), в том числе пару рогов и очень короткий и глубокий череп. На верхней челюсти имелись выемки над промаксиллярным окном, в которых должен был располагаться анторбитальный воздушный синус (воздушные проходы в морде). Носослезный проток, транспортирующий глазную жидкость, выходил на медиальную (внутреннюю) поверхность слезной железы через канал неопределенной функции. Другие предполагаемые аутапоморфии включают глубокую и длинную, наполненную воздухом выемку в квадратной кости и удлиненную депрессию на крыловидной неба кости . [ 1 ]
Позвонки
[ редактировать ]
Позвоночник состоял из десяти шейных (шейных), двенадцати спинных и шести сросшихся крестцовых. [ НО ] и неизвестное количество хвостовых (хвостовых) позвонков. [ 4 ] Шея была почти прямой, а не S-образной, как у других теропод, а также необычно широкой, особенно к основанию. [ 27 ] В верхней части позвоночника имеется двойной ряд увеличенных, направленных вверх костных отростков, называемых эпипофизами , образующих гладкую впадину на вершине шейных позвонков. Эти отростки представляли собой высшие точки позвоночника, возвышающиеся над необычайно низкими остистыми отростками . [ 4 ] [ 26 ] Эпипофизы, вероятно, служили местами прикрепления заметно сильной мускулатуры шеи. [ ВЫКЛЮЧЕННЫЙ ] Подобный двойной ряд присутствовал и в хвосте, образованном там сильно видоизмененными хвостовыми ребрами , на виде спереди выступающими вверх V-образной формы, их внутренние стороны создавали гладкую, плоскую верхнюю поверхность передних хвостовых позвонков. Конец каждого хвостового ребра имел выступающее вперед крючковидное расширение, которое соединялось с хвостовым ребром предыдущего позвонка. [ 26 ] [ 28 ]
Передние конечности
[ редактировать ]

Передние конечности были пропорционально короче, чем у любых других крупных хищных динозавров, включая тираннозаврид. [ АГ ] Предплечье было всего лишь в четверть размера плеча. не было запястья На руке , так что пястные кости сочленялись непосредственно с предплечьем. [ 29 ] Рука показывала четыре основные цифры, [ 4 ] хотя, по-видимому, только две средние из них заканчивались костями пальцев, а четвертый состоял из единственной пястной кости, похожей на шину, которая могла представлять собой внешнюю «шпору». Сами пальцы были сросшимися и неподвижными, возможно, у них не было когтей. [ 30 ] Карнотавр отличался от всех других абелизаврид тем, что у него были пропорционально более короткие и крепкие передние конечности, а также наличие четвертой, похожей на шину пястной кости, самой длинной кости руки. [ 29 ] Исследование 2009 года предполагает, что у абелизаврид руки были рудиментарными , поскольку нервные волокна, ответственные за передачу стимулов, были редуцированы до такой степени, как у современных эму и киви , у которых также есть рудиментарные передние конечности. [ 31 ]
Кожа
[ редактировать ]Карнотавр был первым динозавром-тероподом, обнаруженным со значительным количеством отпечатков окаменелой кожи. [ 7 ] Эти отпечатки, обнаруженные под правой стороной скелета, происходят из разных частей тела, включая нижнюю челюсть, [ 7 ] передняя часть шеи, плечевой пояс и грудная клетка . [ ах ] Самый большой участок кожи соответствует передней части хвоста. [ ИИ ] Первоначально правая сторона черепа также была покрыта большими участками кожи — при подготовке черепа это не было распознано, и эти участки были случайно уничтожены. [ 7 ] Однако текстура поверхности некоторых костей черепа позволяет сделать выводы об их вероятном покрытии. Бугристая поверхность с бороздками, ямками и небольшими отверстиями находится по бокам и в передней части морды и указывает на чешуйчатый покров, возможно, с плоскими чешуйками, как у современных крокодилов. Верхняя часть рыла была украшена многочисленными мелкими отверстиями и шипами – эту текстуру, вероятно, можно соотнести с ороговевшей подушечкой (роговым покровом). Такая подушечка также встречалась у Majungasaurus , но отсутствовала у Abelisaurus и Rugops . Ряд крупных чешуек, вероятно, окружал глаз, о чем свидетельствует бугристая поверхность с продольными бороздками на слезной и заглазничной костях. [ 1 ] [ Эй Джей ]

Кожа состоит из мозаики многоугольных непересекающихся чешуек диаметром примерно 5–12 мм (0,20–0,47 дюйма). Эта мозаика была разделена тонкими параллельными бороздками. [ И ] Чешуйки на разных частях тела были одинаковыми, за исключением головы, на которой, по-видимому, наблюдался разный, неравномерный рисунок чешуек. [ АЛ ] [ 15 ] Никаких следов перьев нет. [ 7 ] Более крупные бугоркообразные образования были расположены по бокам шеи, спины и хвоста неправильными рядами. Эти шишки имели диаметр от 4 до 5 см (от 1,6 до 2,0 дюйма) и высоту до 5 см (2,0 дюйма) и часто имели низкий срединный гребень. Они были расположены на расстоянии 8–10 см (3,1–3,9 дюйма) друг от друга и становились больше к верху животного. Шишки, вероятно, представляют собой характерные чешуйки – скопления уплотненных щитков – подобные тем, которые можно увидеть на мягком жабо, идущем вдоль средней линии тела у гадрозаврид («утконосых») динозавров. Эти структуры не содержали кости. [ ЯВЛЯЮСЬ ] [ 7 ] [ 32 ] Стивен Черкас (1997) предположил, что эти структуры могли защищать бока животного во время борьбы с представителями того же вида ( сородичей ) и другими тероподами, утверждая, что подобные структуры можно найти на шее современной игуаны , где они обеспечивают ограниченную защиту в бою. . [ 7 ]
Более поздние исследования кожи карнотавра, опубликованные в 2021 году, показывают, что предыдущие изображения чешуи на теле были неточными, а более крупные чешуйки были распределены вдоль тела случайным образом, а не отдельными рядами, как в старых художественных изображениях и иллюстрациях. Также нет признаков прогрессивного изменения размеров в масштабах функций на разных участках тела. По сравнению с этим, базальные чешуи карнотавра были весьма разнообразными: от маленьких и удлиненных до больших и многоугольных, от круглых до двояковыпуклых в грудной, лопаточной и хвостовой областях соответственно. Эта дифференциация масштабов могла быть связана с регулированием тепла тела и отводом избыточного тепла посредством терморегуляции из-за большого размера тела и активного образа жизни. [ 33 ]
Классификация
[ редактировать ]

Carnotaurus — один из наиболее изученных родов Abelisauridae южном , семейства крупных теропод, обитающих на древнем суперконтиненте Гондвана . Абелизавриды были доминирующими хищниками в позднем мелу Гондваны, заменив кархародонтозаврид и заняв экологическую нишу, заполненную тираннозавридами на северных континентах. [ 17 ] Несколько примечательных черт, развившихся внутри этого семейства, в том числе укорочение черепа и рук, а также особенности шейных и хвостовых позвонков, были более выражены у карнотавра , чем у любого другого абелизаврида. [ АН ] [ К ] [ 28 ]
Хотя взаимоотношения внутри Abelisauridae обсуждаются, кладистический анализ неизменно показывает, что Carnotaurus является одним из наиболее производных членов семейства. [ АП ] Его ближайшим родственником мог быть ауказавр. [ 34 ] [ 35 ] [ 36 ] [ 37 ] или Майюнгазавр . [ 38 ] [ 39 ] [ 40 ] Обзор 2008 года, напротив, показал, что Carnotaurus не был тесно связан ни с одним из родов, и вместо этого предложил Ilokelesia в качестве его сестринского таксона . [ АК ] Хуан Канале и его коллеги в 2009 году создали новую кладу Brachyrostra, включив в нее Карнотавра , но не Майюнгазавра ; С тех пор за этой классификацией последовал ряд исследований. [ 34 ] [ 37 ] [ 41 ]
Carnotaurus является одноименным для двух подгрупп Abelisauridae: Carnotaurinae и Carnotaurini . Палеонтологи не всегда признают эти группы. В состав Carnotaurinae были включены все производные абелизавриды, за исключением Abelisaurus , который в большинстве исследований считается базальным представителем. [ 42 ] Однако обзор 2008 года показал, что абелизавр был производным абелизаврида. [ АР ] Карнотаурини было предложено назвать кладу , образованную Карнотавром и Ауказавром ; [ 35 ] Эту группу используют только те палеонтологи, которые считают ауказавра ближайшим родственником карнотавра . [ 43 ] Исследование 2024 года выявило Карнотаурини как действительную кладу, состоящую Карнотавра , Ауказавра , Тумана и Коллекции. из [ 44 ]
Ниже представлена кладограмма, опубликованная Канале и его коллегами в 2009 году. [ 34 ]
Палеобиология
[ редактировать ]Функция рогов
[ редактировать ]
Карнотавр — единственное известное хищное двуногое животное с парой рогов на лобной кости. [ 45 ] Использование этих рогов не совсем понятно. Несколько интерпретаций вращались вокруг использования в борьбе с сородичами или для убийства добычи, хотя возможно и использование для демонстрации для ухаживания или распознавания представителей того же вида. [ 1 ]
Грег Пол (1988) предположил, что рога бодали оружие и что маленькая орбита сводила к минимуму возможность повредить глаза во время боя. [ 9 ] Херардо Маццетта и его коллеги (1998) предположили, что Карнотавр использовал свои рога так же, как бараны. Они подсчитали, что мускулатура шеи была достаточно сильной, чтобы выдержать силу двух человек, столкнувшихся головами фронтально со скоростью 5,7 м/с каждый. [ 22 ] Фернандо Новас (2009) интерпретировал некоторые особенности скелета как приспособления для нанесения ударов головой. [ КАК ] Он предположил, что короткий череп мог бы ускорить движения головы за счет уменьшения момента инерции , а мускулистая шея позволяла бы наносить сильные удары головой. Он также отметил повышенную жесткость и прочность позвоночного столба, который, возможно, развился, чтобы выдерживать удары, проводимые головой и шеей. [ В ]
Другие исследования показывают, что соперничающие карнотары не наносили быстрых ударов головой, а медленно толкали друг друга верхними сторонами черепов. [ 45 ] [ 46 ] Маццетта и его коллеги в 2009 году утверждали, что рога могли быть устройством для распределения сил сжатия без повреждения мозга. В пользу этого говорят уплощенные верхние стороны рогов, сильно сросшиеся кости верхней части черепа и неспособность черепа выдерживать быстрые удары головой. [ 45 ] Рафаэль Делькур в 2018 году предположил, что рога можно было использовать либо для медленных ударов головой и толчков, как у современной морской игуаны , либо для ударов по шее и бокам противника, как у современного жирафа . [ 37 ] Последняя возможность была ранее предложена для родственного майюнгазавра в документе конференции 2011 года. [ 47 ]
Херардо Маццетта и его коллеги (1998) предполагают, что рога также могли использоваться для ранения или убийства мелкой добычи. Хотя сердцевины рогов тупые, они могли иметь форму, похожую на рога современных быков, если бы у них было ороговевшее покрытие. Однако это единственный зарегистрированный пример использования рогов в качестве охотничьего оружия у животных. [ 22 ]
Функция челюсти и диета
[ редактировать ]
Анализ строения челюстей карнотавра , проведенный Маццеттой и его коллегами в 1998, 2004 и 2009 годах, позволяет предположить, что животное было способно к быстрым, но не сильным укусам. [ 22 ] [ 23 ] [ 45 ] Быстрые укусы важнее сильных при поимке мелкой добычи , как показали исследования современных крокодилов . [ 45 ] Эти исследователи также отметили высокую степень гибкости ( кинезиса ) внутри черепа и особенно нижней челюсти, чем-то напоминающую современных змей . Эластичность челюсти позволила бы карнотаврам проглатывать мелкую добычу целиком. Кроме того, передняя часть нижней челюсти была шарнирной и благодаря этому могла двигаться вверх и вниз. При нажатии вниз зубы выступали вперед, позволяя Карнотаврам протыкать мелкую добычу; когда зубы были изогнуты вверх, выступающие назад зубы препятствовали бы побегу пойманной добычи. [ 22 ] Маццетта и его коллеги также обнаружили, что череп способен противостоять силам, возникающим при перетаскивании крупных предметов добычи. [ 45 ] Таким образом, карнотавр , возможно, питался в основном относительно мелкой добычей, но также был способен охотиться на крупных динозавров. [ 45 ] В 2009 году Маццетта и его коллеги оценили силу укуса примерно в 3341 ньютон. [ 45 ] Исследование 2022 года, оценивающее силу укуса 33 различных динозавров, показывает, что сила укуса у карнотавра составляла около 3392 ньютонов в передней части челюстей; немного выше предыдущей оценки. Между тем сила заднего укуса в задней части челюстей оценивалась в 7172 ньютона. [ 48 ]
Эта интерпретация была подвергнута сомнению Франсуа Терриеном и его коллегами (2005), которые обнаружили, что сила укуса Карнотавра была вдвое больше, чем у американского аллигатора , который, возможно, имеет самый сильный укус среди всех ныне живущих четвероногих . Эти исследователи также отметили аналогии с современными драконами Комодо : сила изгиба нижней челюсти линейно уменьшается к кончику, что указывает на то, что челюсти приспособлены не для высокоточной ловли мелкой добычи, а для нанесения режущих ран, чтобы ослабить крупную добычу. Как следствие, согласно этому исследованию, карнотавр, должно быть, охотился в основном на крупных животных, возможно, из засады. [ 49 ] Черрони и его коллеги в 2020 году утверждали, что гибкость была ограничена нижней челюстью, в то время как утолщенная крыша черепа и окостенение нескольких черепных суставов позволяют предположить, что череп не имел кинезиса или имел лишь небольшой кинезис. [ 1 ]
Роберт Баккер (1998) обнаружил, что карнотавр в основном питался очень крупной добычей, особенно зауроподами . Как он отметил, несколько адаптаций черепа — короткая морда, относительно маленькие зубы и сильная задняя часть черепа ( затылок ) — независимо развились у аллозавра . Эти особенности позволяют предположить, что верхняя челюсть использовалась как зазубренная дубинка для нанесения ран; большие зауроподы были бы ослаблены повторяющимися атаками. [ 50 ]
Передвижение
[ редактировать ]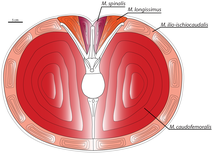

Маццетта и его коллеги (1998, 1999) предположили, что Карнотавр был быстрым бегуном, утверждая, что бедренная кость приспособлена выдерживать сильные изгибающие моменты во время бега; Способность ноги животного противостоять этим силам ограничивает его максимальную скорость. Адаптация Карнотавра к бегу была бы лучше, чем у человека, хотя и далеко не так хороша, как у страуса . [ В ] [ 51 ] Ученые подсчитали, что Карнотавр развивал максимальную скорость до 48–56 км (30–35 миль) в час. [ 52 ]
У динозавров самая важная двигательная мышца располагалась в хвосте. Эта мышца, называемая хвостобедренной костью , прикрепляется к четвертому вертелу , выступающему выступу на бедренной кости, и при сокращении тянет бедренную кость назад. Скотт Персонс и Фил Карри (2011) утверждали, что в хвостовых позвонках карнотавра хвостовые ребра не выступали горизонтально («Т-образно»), а были расположены под углом к вертикальной оси позвонков, образуя букву «V». Это обеспечило бы дополнительное пространство для хвостатофеморальной мышцы, большей, чем у любого другого теропода: мышечная масса оценивалась от 111 до 137 килограммов (от 245 до 302 фунтов) на ногу. Следовательно, Карнотавр мог быть одним из самых быстрых крупных теропод. [ 28 ] В то время как хвостобедренная мышца была увеличена, эпаксиальные мышцы, расположенные над хвостовыми ребрами, были пропорционально меньше. Эти мышцы, называемые длиннейшей и спинной мышцей, отвечали за движение и устойчивость хвоста. Чтобы сохранить устойчивость хвоста, несмотря на сокращение этих мышц, хвостовые ребра выступают вперед, сцепляя позвонки друг с другом и с тазом, придавая хвосту жесткость. Как следствие, способность совершать крутые повороты была бы уменьшена, поскольку бедро и хвост приходилось поворачивать одновременно, в отличие от других теропод. [ 28 ]
Мозг и чувства
[ редактировать ]Черрони и Паулина-Карабахал в 2019 году использовали компьютерную томографию для изучения эндокраниальной полости, в которой находится мозг. Объем эндокраниальной полости составил 168,8 см. 3 , хотя мозг занимал бы лишь часть этого пространства. Авторы использовали две разные оценки размера мозга, предполагая, что размер мозга составляет 50% и 37% эндокраниальной полости соответственно. Это приводит к тому, что коэффициент энцефализации рептилий (показатель интеллекта) выше, чем у родственного майюнгазавра , но меньше, чем у тираннозаврид . , Шишковидная железа которая вырабатывает гормоны , возможно, была меньше, чем у других абелизаврид, о чем свидетельствует небольшое расширение твердой мозговой оболочки — пространство в верхней части переднего мозга, в котором, как полагают, располагалась шишковидная железа. [ 24 ]
, Обонятельные луковицы отвечающие за обоняние, были большими, а зрительные доли , отвечающие за зрение, были относительно небольшими. Это указывает на то, что обоняние могло быть развито лучше, чем зрение, тогда как у современных птиц дело обстоит наоборот. Передний конец обонятельных путей и луковиц был изогнут вниз, что характерно только для индозавра ; у других абелизаврид эти структуры были ориентированы горизонтально. По предположению Черрони и Паулины-Карабахал, эта кривизна вниз вместе с большим размером луковиц может указывать на то, что карнотавр больше полагался на обоняние, чем другие абелизавриды. Флоккулус , доля мозга , которая, как считается, связана со стабилизацией взгляда (координацией между глазами и телом), была большой у карнотавра и других южноамериканских абелизаврид. Это может указывать на то, что эти формы часто использовали быстрые движения головы и тела. Слух мог быть плохо развит у карнотавра. и других абелизаврид, на что указывает короткая лагена внутреннего уха . По оценкам, диапазон слышимости был ниже 3 кГц. [ 24 ]
Возраст и палеосреда
[ редактировать ]
Первоначально породы, в которых был найден Карнотавр , были отнесены к верхней части формации Горро Фриджио , возраст которой считался примерно 100 миллионов лет ( альбский или сеноманский этап). [ 5 ] [ ВЫКЛЮЧЕННЫЙ ] Позже выяснилось, что они относятся к гораздо более молодой формации Ла Колония . [ 13 ] датируемый кампанским и маастрихтским этапами (от 83,6 до 66 миллионов лет назад). [ 1 ] Новас в книге 2009 года дал более узкий временной интервал. 72–69,9 млн лет назад (нижний маастрихтский этап). [ АУ ] Таким образом, карнотавр был последним известным южноамериканским абелизавридом. [ 28 ] К позднему мелу Южная Америка уже была изолирована как от Африки, так и от Северной Америки. [ 53 ]
Формация Ла Колония обнажена на южном склоне Северо-Патагонского массива . [ 54 ] Большинство окаменелостей позвоночных, включая карнотавра , происходят из средней части формации (так называемая ассоциация средней фации ). [ 54 ] Эта часть, вероятно, представляет собой отложения среды эстуариев , приливных отмелей или прибрежных равнин . [ 54 ] Климат должен был быть сезонным, с сухими и влажными периодами. [ 54 ] Наиболее распространенные собираемые позвоночные включают цератодонтидных двоякодышащих рыб , черепах, плезиозавров , крокодилов, динозавров, ящериц, змей и млекопитающих. [ 55 ] Другие динозавры включают Koleken inakayali , который тесно связан с Carnotaurus ; [ 44 ] сальтазавроид - титанозавр Titanomachya gimenezi ; [ 56 ] безымянный анкилозавр ; и безымянный гадрозавроид , среди других. Некоторые из найденных змей принадлежат к семействам Boidae и Madtsoidae, например Alamitophis argentinus . [ 57 ] Черепахи представлены как минимум пятью таксонами , четырьмя из Chelidae ( Pleurodira ) и одним из Meiolaniidae ( Cryptodira ). [ 58 ] К плезиозаврам относятся два эласмозавра ( Kwanectes и Chubutinectes ) и поликотилид ( Sulcusuchus ). [ 59 ] [ 60 ] Млекопитающие представлены Reigitherium Bunodontum и Coloniatherium cilinski , первый из которых считается первой находкой южноамериканского докодонта . [ 54 ] [ 61 ] и возможные гондванатерии или мультитуберкуляты Argentoditescoliensis и Ferugliotheriumwindhauseni . [ 62 ] [ 63 ] останки энантиорнитина и, возможно, неорнитина птицы . Обнаружены [ 64 ] [ 65 ]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ с. 276 в новостях (2009) [ 2 ]
- ^ с. 2 в Бонапарте (1990) [ 4 ]
- ^ с. 2 в Бонапарте (1990) [ 4 ]
- ^ с. 2 в Бонапарте (1990) [ 4 ]
- ^ с. 191 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ с. 191 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ с. 3 в Бонапарте (1990) [ 4 ]
- ^ с. 38 в Бонапарте (1990) [ 4 ]
- ^ с. 162 в деле Хуареса Валиери и др. (2010) [ 18 ]
- ^ с. 191 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ с. 162 в деле Хуареса Валиери и др. (2010) [ 18 ]
- ^ с. 163 в Хуаресе Валиери и др. (2010) [ 18 ]
- ^ с. 556 в Calvo et al. (2004) [ 20 ]
- ^ с. 191 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ с. 30 в Бонапарте (1990) [ 4 ]
- ^ с. 187 в Mazzetta et al. (1998) [ 22 ]
- ^ с. 79 в Mazzetta et al. (2004) [ 23 ]
- ^ с. 276 в новостях (2009) [ 2 ]
- ^ с. 28–32 в Бонапарте (1990) [ 4 ]
- ^ с. 8 в Бонапарте (1990) [ 4 ]
- ^ с. 191 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ с. 4–5 в Бонапарте (1990) [ 4 ]
- ^ с. 5 в Бонапарте (1990) [ 4 ]
- ^ с. 3 в Бонапарте (1990) [ 4 ]
- ^ с. 191 в Mazzetta et al. (1998) [ 22 ]
- ^ с. 255 в новостях (2009) [ 2 ]
- ^ с. 6 в Бонапарте (1990) [ 4 ]
- ^ с. 6 в Бонапарте (1990) [ 4 ]
- ^ с. 6 в Бонапарте (1990) [ 4 ]
- ^ с. 6 в Бонапарте (1990) [ 4 ]
- ^ с. 191 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ стр. 257 в новостях (2009) [ 2 ]
- ^ с. 1276 г. у Руиса и др. (2011) [ 29 ]
- ^ с. 32 в Бонапарте (1990) [ 4 ]
- ^ с. 32 в Бонапарте (1990) [ 4 ]
- ^ с. 3 в Бонапарте (1990) [ 4 ]
- ^ стр. 264–299 в новостях (2009). [ 2 ]
- ^ стр. 264–299 в новостях (2009). [ 2 ]
- ^ с. 32 в Бонапарте (1990) [ 4 ]
- ^ с. 276–277 в новостях (2009). [ 2 ]
- ^ стр. 256–261 в новостях (2009). [ 2 ]
- ^ стр. 188–189 и 202 в Каррано и Сэмпсоне (2008). [ 8 ]
- ^ с. 202 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ с. 202 в Каррано и Сэмпсоне (2008) [ 8 ]
- ^ стр. 259–261 в новостях (2009). [ 2 ]
- ^ стр. 260–261 в новостях (2009). [ 2 ]
- ^ с. 186 и 190 у Mazzetta et al. (1998) [ 22 ]
- ^ с. 3 в Бонапарте (1990) [ 4 ]
- ^ с. 276 в новостях (2009) [ 2 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в Черрони, Массачусетс; Канале, Дж.И.; Новас, FE (2020). «Пересмотр черепа Carnotaurus sastrei Bonaparte, 1985 год: данные черепно-лицевых костей, неба и нижней челюсти» . Историческая биология . 33 (10): 2444–2485. дои : 10.1080/08912963.2020.1802445 . S2CID 225374445 .
- ^ Jump up to: а б с д и ж г час я дж к Новас, Фернандо Э. (2009). Эпоха динозавров в Южной Америке . Блумингтон: Издательство Университета Индианы. ISBN 978-0-253-35289-7 .
- ^ Jump up to: а б Сальгадо, Леонардо; Бонапарт, Хосе Ф. (1991). «Новый зауропод Dicraeosauridae, Amargasaurus chunkui gen. et sp. nov., из формации Ла-Амарга, неоком, провинция Неукен, Аргентина». Амегиниана (на испанском языке). 28 (3–4): 334.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х Бонапарт, Хосе Ф.; Новас, Фернандо Э.; Кориа, Родольфо А. (1990). « Carnotaurus sastrei Bonaparte, рогатый, легкого телосложения карнозавр из среднего мела Патагонии» (PDF) . Вклад в науку . 416 : 1–41. дои : 10.5962/стр.226819 . S2CID 132580445 . Архивировано из оригинала (PDF) 21 июля 2010 г.
- ^ Jump up to: а б с д и Бонапарт, Хосе Ф. (1985). «Рогатый карнозавр мелового периода из Патагонии». Национальное географическое исследование . 1 (1): 149–151.
- ^ Jump up to: а б Полина Карабахал, Ариана (2011). «Анатомия черепной коробки Carnotaurus sastrei (Theropoda: Abelisauridae) из верхнего мела Патагонии». Журнал палеонтологии позвоночных . 31 (2): 378–386. Бибкод : 2011JVPal..31..378P . дои : 10.1080/02724634.2011.550354 . S2CID 129933997 .
- ^ Jump up to: а б с д и ж г час я Черкас, Стивен А.; Черкас, Сильвия Дж. (1997). «Покровы и восстановление жизни карнотавра». В Вольберге, ДИ; Стамп, Э.; Розенберг, Г.Д. (ред.). Динофест Международный . Академия естественных наук, Филадельфия. стр. 155–158.
- ^ Jump up to: а б с д и ж г час я Каррано, Мэтью Т.; Сэмпсон, Скотт Д. (январь 2008 г.). «Филогения цератозавров (Dinosauria: Theropoda)». Журнал систематической палеонтологии . 6 (2): 183–236. Бибкод : 2008JSPal...6..183C . дои : 10.1017/S1477201907002246 . S2CID 30068953 .
- ^ Jump up to: а б с д Пол, Грегори С. (1988). Хищные динозавры мира . Саймон и Шустер. стр. 284–285 . ISBN 978-0-671-61946-6 .
- ^ Йонг, Эд (18 октября 2011 г.). «Буч-хвост сделал Карнотавра чемпионом среди динозавров-спринтеров» . Нэшнл Географик . Архивировано из оригинала 29 октября 2018 года . Проверено 31 июля 2019 г.
- ^ Хедден, Джейми А. (19 сентября 2006 г.). «Re: Carnotaurus sastrei Этимология » . Список рассылки динозавров. Архивировано из оригинала 22 февраля 2012 года . Проверено 27 декабря 2012 г.
- ^ Бонапарт, Хосе Ф. (1991). «Семейства гондванских теропод Abelisauridae и Noasauridae». Историческая биология . 5 (1): 1. Бибкод : 1991HBio....5....1B . дои : 10.1080/10292389109380385 .
- ^ Jump up to: а б Бонапарт, Хосе Ф. (1996). «Меловые четвероногие Аргентины». Мюнхенский геонаучный трактат . А (30): 89.
- ^ Перенасыщение, Дональд Ф. (1997). « Карнотавр » . Динозавры, энциклопедия . Джефферсон, Северная Каролина: Издательство McFarland & Company, Inc. стр. 256–259 . ISBN 978-0-375-82419-7 .
- ^ Jump up to: а б Перенасыщение, Дональд Ф. (2003). « Карнотавр » . Динозавры: Энциклопедия. 3-е дополнение . Джефферсон, Северная Каролина: McFarland & Company, Inc., стр. 274–276 . ISBN 978-0-7864-1166-5 .
- ^ Перенасыщение, Дональд Ф. (2000). « Карнотавр ». Динозавры: Энциклопедия. 1-е дополнение . Джефферсон, Северная Каролина: McFarland & Company, Inc., стр. 165–167. ISBN 978-0-7864-0591-6 .
- ^ Jump up to: а б Кандейру, Карлуш Роберту душ Аньос; МАРТИНЕЛЛИ, Агустин Гильермо. «Abelisauroidea и carchardontosauridae (тероподы, динозаврии) в меловом периоде Южной Америки. Палеогеографические и геохронологические значения». Уберландия . 17 (33): 5–19.
- ^ Jump up to: а б с Хуарес Вальери, Рубен Д.; Порфири, Хуан Д.; Кальво, Хорхе О. (2010). «Новая информация о Ekrixinatosaurus novasi Calvo et al. 2004, гигантском и массово построенном абелизавроиде из «среднего мела» Патагонии». Палеонтология и динозавры в Латинской Америке : 161–169.
- ^ Jump up to: а б с Пол, Грегори С. (2010). Принстонский полевой справочник по динозаврам (1-е изд.). Издательство Принстонского университета. ISBN 9780691137209 .
- ^ Кальво, Хорхе О.; Рубилар-Роджерс, Дэвид; Морено, Карен (2004). «Новые Abelisauridae (Dinosauria: Theropoda) из северо-западной Патагонии» . Амегиниана . 41 (4): 555–563.
- ^ Грилло, Онтарио; Делькур, Р. (2016). «Аллометрия и длина тела абелизавроидных теропод: Pycnonemosaurus nevesi — новый король». Меловые исследования . 69 : 71–89. Бибкод : 2017CrRes..69...71G . дои : 10.1016/j.cretres.2016.09.001 .
- ^ Jump up to: а б с д и ж г Маццетта, Херардо В.; Фаринья, Ричард А.; Вискайно, Серджио Ф. (1998). «К палеобиологии южноамериканского рогатого теропода Carnotaurus sastrei Bonaparte» (PDF) . Гайя . 15 : 185–192.
- ^ Jump up to: а б Маццетта, Херардо В.; Кристиансен, Пер; Фаринья, Ричард А. (июнь 2004 г.). «Гиганты и причуды: размер тела некоторых южно-южноамериканских меловых динозавров». Историческая биология . 16 (2–4): 71–83. Бибкод : 2004HBio...16...71M . CiteSeerX 10.1.1.694.1650 . дои : 10.1080/08912960410001715132 . S2CID 56028251 .
- ^ Jump up to: а б с д Черрони, Маурисио А.; Паулина-Карабахал, Ариана (2019). «Новая информация об эндокраниальной морфологии теропода-абелизаврида Carnotaurus sastrei ». Comptes Рендус Палевол . 18 (8): 985–995. Бибкод : 2019CRPal..18..985C . дои : 10.1016/j.crpv.2019.09.005 .
- ^ Jump up to: а б Сэмпсон, Скотт Д.; Уитмер, Лоуренс М. (2007). «Черепно-лицевая анатомия Majungasaurus crenatissimus (Theropoda: Abelisauridae) из позднего мела Мадагаскара». Журнал палеонтологии позвоночных . 27 (сп8): 95–96. doi : 10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2 . S2CID 129240095 .
- ^ Jump up to: а б с Хартман, Скотт (2012). «Карнотавр – углубляемся в самопародию?» . Проверено 7 декабря 2012 г.
- ^ Мендес, Ариэль (2014). «Шейные позвонки позднемелового динозавра-абелизаврида Carnotaurus sastrei» . Acta Palaeontologica Polonica . 59 (1): 99–107. дои : 10.4202/app.2011.0129 . hdl : 11336/32909 . S2CID 67792084 .
- ^ Jump up to: а б с д и Лица, WS; Карри, Пи Джей (2011). Фарке, Эндрю Аллен (ред.). «Демон скорости динозавров: хвостовая мускулатура Carnotaurus sastrei и значение для эволюции южноамериканских абелизаврид» . ПЛОС ОДИН . 6 (10): e25763. Бибкод : 2011PLoSO...625763P . дои : 10.1371/journal.pone.0025763 . ПМК 3197156 . ПМИД 22043292 .
- ^ Jump up to: а б с д Руис, Хавьер; Торичес, Анжелика; Серрано, Умберто; Лопес, Валле (2011). «Строение рук Carnotaurus sastrei (Theropoda, Abelisauridae): значение для разнообразия рук и эволюции абелизаврид» (PDF) . Палеонтология . 54 (6): 1271–1277. Бибкод : 2011Palgy..54.1271R . дои : 10.1111/j.1475-4983.2011.01091.x . S2CID 43168700 . Архивировано из оригинала (PDF) 22 сентября 2017 года . Проверено 18 декабря 2018 г.
- ^ Аньолин, Федерико Л.; Кьярелли, Пабло (июнь 2010 г.). «Положение когтей у Noasauridae (Dinosauria: Abelisauroidea) и его значение для эволюции абелизавроидной кисти». Палеонтологический журнал . 84 (2): 293–300. Бибкод : 2010PalZ...84..293A . дои : 10.1007/s12542-009-0044-2 . S2CID 84491924 .
- ^ Сентер, П. (2010). «Рудиментарные скелетные структуры динозавров». Журнал зоологии . 280 (4): 60–71. дои : 10.1111/j.1469-7998.2009.00640.x .
- ^ Кампионе, Николас Э.; Барретт, Пол М.; Эванс, Дэвид С. (2020). «О происхождении перьев мезозойских динозавров». В Кристиане Фоте; Оливер В.М. Раухут (ред.). Эволюция перьев . Спрингер. стр. 213–243. ISBN 978-3-030-27222-7 .
- ^ Хендрикс, Кристоф; Белл, Фил Р. (август 2021 г.). «Чешуйчатая кожа абелизаврида Carnotaurus sastrei (Theropoda: Ceratosauria) из верхнего мела Патагонии». Меловые исследования . 128 : 104994. Бибкод : 2021CrRes.12804994H . дои : 10.1016/j.cretres.2021.104994 .
- ^ Jump up to: а б с Канале, Хуан И.; Сканферла, Карлос А.; Аньолин, Федерико; Новас, Фернандо Э. (2009). «Новый плотоядный динозавр из позднего мела северо-западной Патагонии и эволюция теропод абелизаврид». Naturwissenschaften . 96 (3): 409–14. Бибкод : 2009NW.....96..409C . дои : 10.1007/s00114-008-0487-4 . hdl : 11336/52024 . ПМИД 19057888 . S2CID 23619863 .
- ^ Jump up to: а б Кориа, Родольфо А.; Кьяппе, Луис М.; Дингус, Лоуэлл (2002). «Новый близкий родственник Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) из позднего мела Патагонии». Журнал палеонтологии позвоночных . 22 (2): 460. doi : 10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2 . S2CID 131148538 .
- ^ Эскурра, Мартин Д.; Агнолин, Фредерик Л.; Новас, Фердинанд Э. (10 мая 2010 г.). «Абелизавроидный динозавр с неатрофированной костью кисти из позднемеловой формации Пари Айке на юге Патагонии» Зоотаксы 2450 (1): 1. дои : 10.11646/zootaxa.2450.1.1 .
- ^ Jump up to: а б с Делькур, Рафаэль (2018). «Палеобиология цератозавров: новый взгляд на эволюцию и экологию южных правителей» . Научные отчеты . 8 (1): 9730. Бибкод : 2018НатСР...8.9730Д . дои : 10.1038/s41598-018-28154-x . ПМК 6021374 . ПМИД 29950661 .
- ^ Серено, Пол С .; Уилсон, Джеффри А.; Конрад, Джек Л. (7 июля 2004 г.). «Новые динозавры связывали южные массивы суши в середине мелового периода» . Труды Королевского общества B: Биологические науки . 271 (1546): 1325–1330. дои : 10.1098/rspb.2004.2692 . ПМК 1691741 . ПМИД 15306329 .
- ^ Тыкоски, Рональд Б.; Роу, Тимоти (2004). «Цератозаврия». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (Второе изд.). Беркли: Издательство Калифорнийского университета. п. 65 . ISBN 978-0-520-24209-8 .
- ^ Уилсон, Джеффри А.; Серено, Пол С.; Шривастава, Суреш; Бхатт, Девендра К.; Хосла, Ашу; Сахни, Ашок (2003). «Новый абелизаврид (Dinosauria, Theropoda) из формации Ламета (мел, маастрихт) Индии». Материалы Музея палеонтологии . 31 (1): 25. HDL : 2027,42/48667 .
- ^ Ван, Шуо; Штиглер, Йозеф; Амио, Ромен; Ван, Сюй; Ду, Го-хао; Кларк, Джеймс М.; Сюй, Син (январь 2017 г.). «Чрезвычайные онтогенетические изменения у цератозаврового теропода» . Современная биология . 27 (1): 144–148. Бибкод : 2017CBio...27..144W . дои : 10.1016/j.cub.2016.10.043 . ПМИД 28017609 .
- ^ Серено, Пол (2005). «Карнотаурины» . Таксоновый поиск . Архивировано из оригинала 16 мая 2012 года . Проверено 29 декабря 2012 г.
- ^ Серено, Пол (2005). «Карнотаурини» . Таксоновый поиск . Архивировано из оригинала 16 мая 2012 года . Проверено 29 декабря 2012 г.
- ^ Jump up to: а б Пол, Диего; Баяно, Маттиа Антонио; Блэк, Дэвид; Новас, Фернандо; Серда, Игнасио А. (21 мая 2024 г.). «Новый динозавр-абелизаврид конца мела Патагонии и темпы эволюции среди цератозавров» . Кладистика . 40 (3): 307–356. дои : 10.1111/cla.12583 . ПМИД 38771085 .
- ^ Jump up to: а б с д и ж г час Маццетта, Херардо В.; Чисилино, Адриан П.; Бланко, Р. Эрнесто; Кальво, Нестор (2009). «Краниальная механика и функциональная интерпретация рогатого хищного динозавра Carnotaurus sastrei ». Журнал палеонтологии позвоночных . 29 (3): 822–830. Бибкод : 2009JVPal..29..822M . дои : 10.1671/039.029.0313 . hdl : 11336/34937 . S2CID 84565615 .
- ^ Чуре, Дэниел Дж. (1998). «На орбите тероподовых динозавров». Гайя . 15 : 233–240.
- ^ Снайвли, Эрик; Коттон, Джон Р.; Уитмер, Лоуренс; Риджели, Райан; Теодор, Джессика (2011). «Конечно-элементное сравнение функции черепной пазухи у динозавра майюнгазавра и жирафов, бьющих головой». Летняя биоинженерная конференция . Том. 54587. Американское общество инженеров-механиков. стр. 1075–1076.
- ^ Сакамото, Манабу (2022 г.). «Оценка силы укуса у вымерших динозавров с использованием филогенетически предсказанных физиологических площадей поперечного сечения приводящих мышц челюсти» . ПерДж . 10 : е13731. дои : 10.7717/peerj.13731 . ПМЦ 9285543 . ПМИД 35846881 .
- ^ Терриен, Франсуа; Хендерсон, Дональд; Рафф, Кристофер (2005). «Укуси меня - биомеханические модели нижних челюстей теропод и их влияние на пищевое поведение». В Карпентер, Кеннет (ред.). Плотоядные динозавры . Издательство Университета Индианы. стр. 179–198 , 228. ISBN. 978-0-253-34539-4 .
- ^ Баккер, Роберт Т. (1998). «Убийцы бронтозавров: позднеюрские аллозавриды как аналоги саблезубых кошек» (PDF) . Гайя . 15 : 145–158.
- ^ Маццетта, Херардо В.; Фарина, Ричард А. (1999). «Оценка спортивных способностей Amargasaurus cazaui Salgado и Bonaparte, 1991 г. и Carnotaurus sastrei Bonaparte, 1985 г. (Saurischia, Sauropoda-Theropoda)». XIV Jornadas Argentinas de Paleontologia de Vertebrados, Ameghiniana (на испанском языке). 36 (1): 105–106.
- ^ «Хищный динозавр был устрашающе быстр» . Новости ЦБК . 21 октября 2011 года . Проверено 22 апреля 2017 г.
- ^ Ле Лефф, Жан (1997). «Биогеография». В Падиане, Кевин; Карри, Филип Дж. (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса . стр. 51–56 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с д и Паскуаль, Розендо; Гоин, Франциско Дж.; Гонсалес, Пабло; Ардолино, Альберто; Пуэрта, Пабло Ф. (2000). «Высокоразвитый докодонт из патагонского позднего мела: эволюционные последствия для млекопитающих Гондваны» . Геодиверситас . 22 (3): 395–414.
- ^ Стерли, Джулиана; Де ла Фуэнте, Марсело С. (2011). «Новая черепаха из формации Ла Колония (кампан-маастрихт), Патагония, Аргентина, с замечаниями об эволюции позвоночного столба черепах» . Палеонтология . 54 (1): 65. Бибкод : 2011Palgy..54...63S . дои : 10.1111/j.1475-4983.2010.01002.x .
- ^ Перес-Морено, А.; Сальгадо, Л.; Карбаллидо, JL; Отеро, А.; Пол, Д. (2024). «Новый титанозавр из формации Ла Колония (кампан-маастрихт), провинция Чубут, Аргентина» . Историческая биология: Международный журнал палеобиологии : 1–20. дои : 10.1080/08912963.2024.2332997 .
- ^ Альбинос, Адриана М. (2000). «Новая находка змей из мела Патагонии (Аргентина)» . Геодиверситас . 22 (2): 247–253.
- ^ Гаспарини, Зульма; Де ла Фуэнте, Марсело (2000). «Черепахи и плезиозавры из формации Ла Колония (верхний мел) в Патагонии, Аргентина». Испанский журнал палеонтологии (на испанском языке). 15 (1): 23.
- ^ О'Горман, Джозеф П.; Кариньяно, Анна Паула; Балд-Маркилезе, Лидия; Перес Панера, Джон Пол (10 августа 2023 г.). «Новый эласмозаврид (Sauropterygia, Plesiosauria) из верхних слоев формации Ла Колония (верхний маастрихт), провинция Чубут, Аргентина» Меловые исследования . 152 : 105674. Бибкод : 2023CrRes.15205674O . дои : 10.1016/j.crests.2023.105674 . ISSN 0195-6671 . S2CID 260830333 .
- ^ О'Горман, JP; Гаспарини, З. (2013). «Ревизия Sulcusuchus erraini (Sauropterygia, Polycotylidae) из верхнего мела Патагонии, Аргентина». Алчеринга: Австралазийский журнал палеонтологии . 37 (2): 163–176. Бибкод : 2013Алч...37..163О . дои : 10.1080/03115518.2013.736788 . hdl : 11336/2489 . S2CID 131429825 .
- ^ Ружье, GW; Тураццинни, Г.Ф.; Кардозо, Миссисипи; Харпер, Т.; Лирес, А.И.; Канесса, Луизиана (2021). «Новые образцы Reigitherium Bunodontum из позднемеловой формации Ла Колония, Патагония, Аргентина и разнообразие Меридиолестидана в Южной Америке». Журнал эволюции млекопитающих . 28 (4): 1051–1081. дои : 10.1007/s10914-021-09585-2 . S2CID 254704047 .
- ^ Килан-Яворовска, Зофья; Ортис-Хорегисар, Эдгардо; Виейтес, Каролина; Паскуаль, Розендо; Гоин, Франсиско Дж. (2007). «Первое цимолодонтанное мультитуберкулезное млекопитающее из Южной Америки» (PDF) . Acta Palaeontologica Polonica . 52 (2): 257–262.
- ^ Гурович Ю.; Бек, Р. (2009). «Филогенетическое сходство загадочной клады млекопитающих Gondwanatheria». Журнал эволюции млекопитающих . 16 (1): 25–49. дои : 10.1007/s10914-008-9097-3 . S2CID 42799370 .
- ^ Ловер, Дэниел Р.; Деби, Эй Джей М.; Кларк, Джулия А.; Ружье, Гильермо В. (1 января 2011 г.). «Новая энантиорнитиновая птица из верхнемеловой формации Ла Колония в Патагонии, Аргентина». Анналы музея Карнеги . 80 (1): 35–42. дои : 10.2992/007.080.0104 . S2CID 85015365 .
- ^ Акоста Госпиталече, К.; О'Горман, JP; Панзери, КМ (2023 г.). «Новая меловая птица из маастрихтской формации Ла Колония (Патагония, Аргентина)». Меловые исследования . 150 . 105595. Бибкод : 2023CrRes.15005595A . дои : 10.1016/j.cretres.2023.105595 . S2CID 259059084 .
Внешние ссылки
[ редактировать ]- Укус Карнотавра в Национальном университете Мар-дель-Плата. (на испанском языке)
- Скелетная реконструкция Скотта Хартмана














