Жгутичок
| Жгутичок | |
|---|---|
 Структура бактериального жгутика | |
 | |
| Идентификаторы | |
| Сетка | D005407 |
| Тур | H1.00.01.1.01032 |
| FMA | 67472 |
| Анатомическая терминология | |
Жгутичок « ( / f l ə ˈ dʒ ɛ l əm / ; pl.: Жгутичок ) для (латынь кнута» или «бич») - это приложение для волос, которое выступает из определенных растений животных и сперматозоидов , от грибковых споров ( зооспоры ) и из широкого спектра микроорганизмов для обеспечения подвижности . [ 1 ] [ 2 ] [ 3 ] [ 4 ] Многие протисты с жгутиками известны как жгутики .
Микроорганизм может иметь от одного до многих жгутиков. Helicobacter грамотрицательная бактерия pylori Например, использует свою жгутику, чтобы продемонстрировать себя через желудок, чтобы достичь слизистой оболочки , где он может колонизировать эпителий и потенциально вызвать гастрит, а также язвы -фактор риска развития рака желудка . [ 5 ] В некоторых роящих бактериях жгутичок также может функционировать как сенсорная органелла , чувствительная к влажности вне клетки. [ 6 ]
В трех доменах бактерий . , археи и эукариоты жгутиковой флагель имеет другую структуру, состав белка и механизм движения, но имеет ту же функцию обеспечения подвижности Латинский », чтобы описать словесный жгутичок означает « кнут его плавание, похожее на ресничные движения. Плаватка в археи называется архауллом, чтобы отметить его отличие от бактериального жгутика. [ 7 ] [ 8 ]
Эукариотические жгутики и реснички идентичны по структуре, но имеют различные длины и функции. [ 9 ] Прокариотические фимбрии и пили являются меньше, и более тонкие придатки с различными функциями.
Типы
[ редактировать ]
Три типа жгутиков являются бактериальными, архиальными и эукариотическими.
У жгутиков у эукариот есть динеин и микротрубочки , которые движутся с изгибающим механизмом. Бактерии и археи не имеют в их жгутике динеина или микротрубочек, и они движутся с помощью вращающегося механизма. [ 11 ]
Другие различия между этими тремя типами:
- Бактериальные жгутики - это спиральные филаменты, каждая с вращающимся двигателем у его основания, которая может поворачиваться по часовой стрелке или против часовой стрелки. [ 12 ] [ 13 ] [ 14 ] Они обеспечивают два из нескольких видов бактериальной подвижности . [ 15 ] [ 16 ]
- Архаальные жгутики ( архаэлла ) поверхностно похожи на бактериальную жгутику в том смысле, что она также имеет вращающийся мотор, но отличается во многих деталях и считается не гомологичным . [ 17 ] [ 18 ] [ 19 ]
- Эукариотическая жгутика - те животных, растений и протести -клеток - сложные клеточные проекции, которые набрасываются взад -вперед. Эукариотические жгутики и подвижные реснички идентичны по структуре, но имеют различные длины, формы волны и функции. Первичные реснички являются нестильными и имеют структурно различную аксонеему 9+0, а не аксоксия 9+2, обнаруженная как в жгутиках, так и в мотивных ресничках.
Бактериальная жгутика
[ редактировать ]Структура и композиция
[ редактировать ]Бактериальный жгутичок состоит из белковых субъединиц флагеллина . [ 11 ] Его форма - 20 -нанометровая полая трубка. Он спиральный и имеет резкий изгиб прямо за пределами внешней мембраны; Этот «крючок» позволяет ось спирали указывать непосредственно от ячейки. Шахта проходит между крючком и базальным телом , проходя через белковые кольца в мембране клетки, которые действуют как подшипники. Грамположительные организмы имеют два из этих базальных колец тела, один в слое пептидогликана и один в плазматической мембране . Грам-негативные организмы имеют четыре таких кольца: кольцо L связано с липополисахаридами , кольцо P ассоциируется со слоем пептидогликана , кольцо М встроено в плазматическую мембрану , а кольцо S прямо прикреплено к цитоплазме . Филамента заканчивается с белком с покрытием. [ 20 ] [ 21 ]
Платаж флажки - это длинный спиральный винт, который продвигает бактерию при вращении двигателя через крючок. У большинства изучаемых бактерий, в том числе грамотрицательная Escherichia coli , Salmonella typhimurium , Caulobacter Crescentus и Vibrio alginolyticus , нить состоит из 11 протофиламентов, приблизительно параллельных оси. Каждое протофиламент представляет собой серию тандемных белковых цепей. Тем не менее, Campylobacter Jejuni имеет семь протофиламентов. [ 22 ]
Базальное тело имеет несколько общих черт с некоторыми типами секреторных пор , таких как пустого, похожая на стержень, в их центрах, простирающихся через плазматическую мембрану. Сходство между бактериальными жгутиками и структурными структурами и белками бактериальной секреторной системы предоставляет научные доказательства, подтверждающие теорию, что бактериальная жгутика эволюционировала из системы секреции три типа (TTSS).
Атомная структура как бактериальных жгутиков, так и инъекции TTSS была выяснена в значительной степени, особенно с развитием криоэлектронной микроскопии . Лучшими понятными частями являются части между внутренней и внешней мембраной , то есть кольцами лесов внутренней мембраны (IM), пары лесов внешней мембраны (ОМ) и стержня/иглы (инъекционная) или стержень/ Крюк (жгутистый) разделы. [ 23 ]
Мотор
[ редактировать ]
Бактериальный жгутичок управляется вращающимся двигателем ( MOT -комплексом ), состоящим из белка, расположенного в якорной точке жгутиков на внутренней клеточной мембране. Двигатель оснащен протонной силой , т. Е. Поток протонов (ионов водорода) через мембрану бактериальных клеток из-за градиента концентрации, установленного метаболизмом клетки ( виды вибраоны имеют два вида жгутиков, боковой и полярной, а некоторые управляются ионным насосом натрия, а не протонным насосом [ 25 ] ) Ротор транспортирует протоны через мембрану и поворачивается в процессе. Только ротор может работать со скоростью от 6000 до 100 000 об / мин , [ 26 ] Но с жгутикой, прикрепленной, обычно достигает от 200 до 1000 об / мин. Направление вращения может быть изменено флагванным переключателем двигателя почти мгновенно, вызванным небольшим изменением положения белка, Flig , в роторе. [ 27 ] Жгутичок очень энергоэффективен и использует очень мало энергии. [ 28 ] [ ненадежный источник? ] Крутящий момент передается из мотаба в крутящую спираль на домене FLIG D5, и с увеличением потребности в крутящем моменте или скорости больше MOTAB. [ 24 ] Поскольку жгутиковой двигатель не имеет выключателя, белок EPSE используется в качестве механической муфты для отключения двигателя от ротора, тем самым останавливая жгутичок и позволяя бактерии оставаться в одном месте. [ 29 ]
| Часть серии на |
| Микробное движение и микробот |
|---|
 |
| Microswimmers |
| Молекулярные двигатели |
Цилиндрическая форма жгутиков подходит для локомоции микроскопических организмов; Эти организмы работают с низким числом Рейнольдса , где вязкость окружающей воды гораздо важнее, чем ее масса или инерция. [ 30 ]
Скорость вращения жгутиков варьируется в ответ на интенсивность протонной силы, тем самым позволяя определенным формам контроля скорости, а также позволяет некоторым типам бактерий достигать замечательных скоростей пропорционально их размеру; Некоторые достигают примерно 60 длины ячейки в секунду. На такой скорости бактерия займет около 245 дней, чтобы покрыть 1 км; Хотя это может показаться медленным, перспектива изменяется при введении концепции масштаба. По сравнению с макроскопическими формами жизни, это действительно очень быстро, когда выражается с точки зрения количества длины тела в секунду. Например, гепарда достигает всего 25 длины тела в секунду. [ 31 ]
Благодаря использованию их жгутиков, бактерии способны быстро двигаться в сторону аттрактантов и вдали от репеллентов, посредством предвзятой случайной прогулки , с пробежками и падающими, приводимыми путем вращения флагеля против часовой стрелки и по часовой стрелке соответственно. Два направления вращения не идентичны (относительно движения жгутика) и выбираются молекулярным переключателем. [ 32 ] Вращение по часовой стрелке называется режим тяги с корпусом, следуя жгутиком. Вращение против часовой стрелки называется режимом двигателя с жгутией, отставая за телом. [ 33 ]
Сборка
[ редактировать ]Во время сборки жгутиков компоненты жгутика проходят через полые ядра базального тела и зарождающуюся нить. Во время сборки компоненты белка добавляются на жгутиковой кончике, а не в основании. [ 34 ] In vitro , жгутиковые нити спонтанно собираются в растворе, содержащем очищенный флагеллин в качестве единственного белка. [ 35 ]
Эволюция
[ редактировать ]По меньшей мере 10 белковых компонентов бактериального жгутика имеют гомологичные белки с системой секреции три типа (T3SS), обнаруженной во многих грамотрицательных бактериях, [ 36 ] Следовательно, один, вероятно, развился от другого. Поскольку T3SS имеет одинаковое количество компонентов, как жгутиковый аппарат (около 25 белков), который из них сначала изменил трудно определить. Тем не менее, жгутиковая система, по -видимому, включает в себя в целом больше белков, включая различные регуляторы и шапероны, следовательно, утверждается, что жгутики эволюционировали из T3SS. Однако это также было предложено [ 37 ] что жгутичок мог развиваться первым, или две структуры развивались параллельно. Потребность в ранних одноклеточных организмах в подвижности (подвижность) поддерживает, что больше мобильной жгутики будет выбран в первую очередь Evolution, [ 37 ] Но T3SS, развивающиеся из жгутика, можно рассматривать как «восстановительная эволюция» и не получает топологической поддержки от филогенетических деревьев. [ 38 ] Гипотеза о том, что две структуры развивались отдельно от общего предка, объясняет сходство белка между двумя структурами, а также их функциональное разнообразие. [ 39 ]
Жгности и интеллектуальные дебаты в дизайне
[ редактировать ]Некоторые авторы утверждают, что жгутики не могут развиваться, предполагая, что они могут функционировать должным образом только тогда, когда все белки на месте. Другими словами, жгутиковый аппарат « неповрежденно сложный ». [ 40 ] Тем не менее, многие белки могут быть удалены или мутированы, а жгутичок все еще работает, хотя иногда при сниженной эффективности. [ 41 ] Более того, со многими белками, уникальными для некоторого количества по разным видам, разнообразие композиции бактериальной жгутики было выше, чем ожидалось. [ 42 ] Следовательно, жгутиковый аппарат явно очень гибкий в эволюционных терминах и совершенно способен потерять или получить белковые компоненты. Например, был обнаружен ряд мутаций, которые увеличивают подвижность кишечной палочки . [ 43 ] Дополнительные доказательства эволюции бактериальной жгутики включают существование рудиментарной жгутики, промежуточные формы жгутиков и паттерны сходства среди последовательностей жгутиков белков, включая наблюдение, что почти все ядра жгутиков имеют известные гомологии с нефлагеллярными белками. [ 36 ] Кроме того, было идентифицировано несколько процессов как играющие важные роли в эволюции жгутиков, включая самосборку простых повторяющихся субъединиц, дублирование генов с последующей дивергенцией, рекрутирование элементов из других систем («молекулярный бриколаж») и рекомбинация. [ 44 ]
Платежеса
[ редактировать ]Различные виды бактерий имеют разные числа и расположение жгутиков, [ 45 ] [ 46 ] Назван с использованием термина Tricho , от греческих триходов, означающих волосы . [ 47 ]
- Монотрихозные бактерии, такие как vibrio cholerae, имеют единый полярный жгутичок . [ 48 ]
- У амфитрихазных бактерий есть один жгутичок на каждом из двух противоположных концов (например, Campylobacter jejuni или Alcaligenes faecalis ) - обоих жгутиков вращаются, но координируются для получения когерентной тяги.
- Lophotrichous Bacteriaia ( Lopho Greek объединяет термин, означающий гребень или пучок ) [ 49 ] Иметь несколько жгутиков, расположенных на одном месте на бактериальной поверхности, таких как Helicobacter pylori , которые действуют согласованно, чтобы управлять бактериями в одном направлении. Во многих случаях основания множественных жгутиков окружены специализированной областью клеточной мембраны, называемой полярной органеллой . [ Цитация необходима ]
- Перитрихозные бактерии имеют жгутиковые жгутики во всех направлениях (например, E. coli ).
Вращение против часовой стрелки монотрихозного полярного жгутика толкает ячейку вперед с отставкой жгутиков, очень похожий на штопок, движущийся внутри пробки. Вода в микроскопическом масштабе очень вязкая , в отличие от обычной воды .
Спирохеты , напротив, имеют жгутиков, называемую эндофлагеллой, возникающей из противоположных полюсов клетки, и расположены в периплазматическом пространстве, как показано путем разрыва внешней мембраны, а также с помощью электронной криотомографической микроскопии. [ 50 ] Вращение филаментов относительно тела клеток заставляет всю бактерию двигаться вперед в движении, подобном штополю, даже через материал достаточно вязкий, чтобы предотвратить проход нормально жгутированных бактерий.
В некоторых крупных формах селеномона , более 30 отдельных жгутиков организованы вне тела клеток, спирально сужая друг вокруг друга, образуя толстую структуру (легко видно с помощью светового микроскопа), называемой « пучкой ».
В некотором вибрации Spp. (особенно Vibrio parahaemolyticus [ 51 ] ) и родственные бактерии , такие как Aeromonas , две жгутиковые системы сосуществуют, используя различные наборы генов и различные градиенты ионов для энергии. Полярные жгутики выражаются конститутивно и обеспечивают подвижность в объемной жидкости, в то время как латеральная жгутика выражается, когда полярная жгутика встречается слишком большим сопротивлением повороту. [ 52 ] [ 53 ] [ 54 ] [ 55 ] [ 56 ] [ 57 ] Они обеспечивают подвижность роя на поверхностях или в вязких жидкостях.
Связывание
[ редактировать ]Связывание-это событие, которое может произойти в мультифлагетированных клетках, объединяя жгутику вместе и заставляя их вращаться скоординированной манерой.
Планировка-это левша спирали, и при вращении против часовой стрелки их роторы могут объединиться и вращаться вместе. Когда роторы обратного направления, тем самым вращающееся по часовой стрелке, жгутичок разматывается от пачка. Это может привести к тому, что ячейка остановила свое направление вперед и вместо этого начнет дергать на месте, называемое падением . Упалование приводит к стохастической переориентации клетки, заставляя ее изменить направление своего прямого плавания.
Неизвестно, какие стимулы приводят к переключению между связкой и падением, но двигатель очень адаптивен к различным сигналам. В модели, описывающей хемотаксис («движение нарочно») вращение по часовой стрелке жгутиков подавляется химическими соединениями, благоприятными для клетки (например, пищи). При движении в благоприятном направлении концентрация таких химических аттрактантов увеличивается, и, следовательно, падающие постоянно подавляются, что позволяет прямое движение; Аналогичным образом, когда направление движения клетки является неблагоприятной (например, вдали от химического аттракта), падающие больше не подавляются и встречаются гораздо чаще, с вероятностью того, что клетка будет переориентироваться в правильном направлении.
Однако, даже если все жгутики будут вращаться по часовой стрелке, они часто не могут образовывать пакет по геометрическим и гидродинамическим причинам. [ 58 ] [ 59 ]
Эукариотическая жгутика
[ редактировать ]



Терминология
[ редактировать ]Стремившись подчеркнуть различие между бактериальной жгутикой и эукариотической ресничной ресничкой и жгутиками, некоторые авторы попытались заменить название этих двух эукариотических структур на « Unmulipodia » (например, все документы Margulis с 1970 -х годов)) [ 60 ] или «реснички» для обоих (например, Хюльсманн, 1992; [ 61 ] Adl et al., 2012; [ 62 ] Большинство документов кавалер-смита ), сохраняя «жгутиковые» для бактериальной структуры. Тем не менее, дискриминационное использование терминов «ресничек» и «жгутиков» для эукариот, принятых в этой статье (см. § Плавания по сравнению с ресничками ниже), все еще является распространенным явлением (например, Andersen et al., 1991; [ 63 ] Leadbeater et al., 2000). [ 64 ]
Внутренняя структура
[ редактировать ]Ядро эукариотического жгутика, известного как аксония, представляет собой пачку из девяти слитых пар микротрубочек, известных как дублеты , окружающие две центральные отдельные микротрубочки ( синглетки ). Эта аксонема 9+2 характерна для эукариотического жгутика. У основания эукариотического жгутика находится базальный корпус , «блефаропласт» или кинетосома, которая представляет собой центр организации микротрубочек для жгутиковых микротрубочек и длиной около 500 нанометров. Базальные тела структурно идентичны центриотам . клетки Жгутичок заключен в плазматическую мембрану клетки , так что внутренняя часть жгутика доступна для цитоплазмы .
Помимо аксонеми и базального тела, относительно постоянного по морфологии, другие внутренние структуры жгутикового аппарата - это переходная зона (где встречаются аксония и базальное тело) и корневая система (микротрубочные или фибриляные структуры, которые простираются от базальных тел в цитоплазму ), более вариабельный и полезный в качестве показателей филогенетических взаимосвязей эукариот. Другими структурами, более необычными, являются парафлагеллярным (или параксиальным, параксосовой) стержнем, R Fiber и S Fiber. [ 65 ] : 63–84 Для поверхностных структур см. Ниже.
Механизм
[ редактировать ]Каждая из наружных 9 -дублетовых микротрубочек распространяет пару рук динеина («внутреннее» и «внешнюю» руку) до соседней микротрубочки; Они производят силу через гидролиз АТФ. Плавательная аксония также содержит радиальные спицы , полипептидные комплексы, простирающиеся от каждой из внешних девяти дублетов микротрубочек в направлении центральной пары, с «головой» спица, обращенной внутрь. Считается, что радиальные выступления участвуют в регуляции жгутикового движения, хотя его точная функция и метод действия еще не поняты. [ 66 ]
Кнопку к ресницам
[ редактировать ]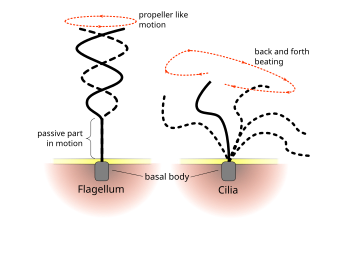
Регулярные рисунки эукариотических ресничек и жгутиков генерируют движение на клеточном уровне. Примеры варьируются от движения отдельных клеток, таких как плавание сперматозоидов , до транспортировки жидкости вдоль стационарного слоя клеток, таких как в дыхательных путях . [ 67 ]
Хотя эукариотические реснички и жгутики в конечном итоге одинаковы, они иногда классифицируются по своему шаблону движения, традиция до того, как их структуры были известны. В случае жгутиков движение часто является плоским и волнообразным, тогда как подвижные реснички часто выполняют более сложное трехмерное движение с силой и ударом по восстановлению. [ 67 ] Еще одна традиционная форма различия - число 9+2 органелл на клетке. [ 66 ]
Внутренний транспорт
[ редактировать ]Внутренний транспорт , процесс, посредством которого аксинемальные субъединицы, трансмембранные рецепторы и другие белки перемещаются вверх и вниз по длине жгутиков, необходим для правильного функционирования жгутика, как при подвижности, так и в трансдукции сигнала. [ 68 ]
Эволюция и возникновение
[ редактировать ]Эукариотическая жгутика или реснички, вероятно, родовая характеристика, [ 69 ] широко распространены почти во всех группах эукариот, как относительно многолетнее состояние или в качестве стадии жизненного цикла жгутиков (например, зооиды , гаметы , зооспоры , которые могут постоянно или нет). [ 70 ] [ 71 ] [ 62 ]
Первая ситуация обнаружена либо в специализированных клетках многоклеточных организмов (например, хоаноциты губки . , либо ресничные эпителии метазой уровень ), как в цилиатах и многих эукариотах с «жгутиковым состоянием» (или «монадоидный организации » Флагеллата , искусственная группа).
Стадии жизненного цикла жгутикового цикла встречаются во многих группах, например, многие зеленые водоросли (зооспоры и мужские гаметы), бриофиты (мужские гаметы), птеридофиты (мужские гаметы), некоторые гимнозерм ( цикады и гинкго , мужские гаметы), сосредоточенные диатомы (мужские гейты. ), коричневые водоросли (зооспоры и гаметы), Oomycetes (ассексуальные зооспоры и гаметы), гифохитриды (зооспоры), лабиринтуломицеты (Zooprose), некоторые апикомпетки (гаметы), некоторые радиоловые (вероятно, гаметы), некоторые [ 72 ] Foraminiferans (гаметы), плазмодиофоромицеты (зооспоры и гаметы), миксогастриды (зооспоры), метазоанцы (мужские гаметы) и грибы читрида (зооспоры и гаметы).
Жакония или реснички полностью отсутствуют в некоторых группах, вероятно, из -за потери, а не примитивного состояния. Потеря ресничек произошла в красных водорослях , некоторых зеленых водорослях ( Zygnematophyceae ), гимноскусах , кроме циад и гинкго , покрытосеменных , типовые диатомовые лица , некоторые апикомпетканы, некоторые амибозоны, в сперме некоторых метазой, некоторые апикомпетканы , некоторые амибозоны , в сперме некоторых метазой , некоторые апикомплексан [ 73 ] и в грибах (кроме хитридов ).
Типология
[ редактировать ]Ряд терминов, связанных с жгутиками или ресничками, используется для характеристики эукариот. [ 71 ] [ 74 ] [ 65 ] : 60–63 [ 75 ] [ 76 ] В соответствии с присутствующими поверхностными структурами, жг.
- жг. Challash
- Волосатая жгутика (= мишура, флэммер, плевронематическая жгутика): с волосками (= мастигонемы Sensu Lato ), разделенная на:
- С тонкими волосками (= не тубулярными или простыми волосками): встречается в Euglenophyceae , Dinoflagellata , некоторых гаптофицеаэ ( Pavlovales )
- с жесткими волосками (= трубчатые волосы, ретронемы, мастигонемы Sensu Stricto ), разделены на:
- Двухпартные волосы: с двумя регионами. Встречается в Cryptophyceae , Prasinophyceae и некоторых гетероконтах
- Трехсторонние (= Straminipilous) волосы: с тремя областями (основание, трубчатый вал и один или несколько терминальных волосков). Встречается в большинстве гетероконты
- Стихондематическая жгутика: с одним рядом волос
- пантонематическая жгутика: с двумя рядами волос
- Acronematic: жгутичок с одним, терминальной мастигонемой или жгутиковыми волосами (например, Bodonids ); [ 77 ] Некоторые авторы используют термин в качестве синонима Whiplash
- С весы: например, Prasinophyceae
- с шипами: например, некоторые коричневые водоросли
- с волнистой мембраной: например, некоторые кинетопластиды , некоторые парабазалиды
- с хоботком (туловище, выступая клетки): например, апусомонады , некоторые бодониды [ 78 ]
Согласно количеству жг. [ 62 ] [ 79 ]
- Uniflagerted: EG, большинство Opisthoconta
- раздваивается: например, все Dinoflagellata , гаметы Charophyceae , большинство бриофитов и некоторых метазоан [ 73 ]
- трифлагетирован: например, гаметы некоторых фораминиферов
- квадрифлагетован: например, некоторые Prasinophyceae , Collodictyonidae
- Octoflagellated: например, некоторые дипломада , некоторые Prasinophyceae
- Многогагетированный: например, Opalinata , Ciliophora , Stephanopogon , Parabasalida , Hemimastigophora , Caryoblastea , Multicilia , гаметы (или зооиды ) Oedogoniales ( Chlorophyta ), некоторые птерофиты и некоторые гимнаспермы
Согласно месту введения жгутиков: [ 80 ]
- Opisthokont: клетки с жгутиками, вставленными сзади, например, в Opisthokonta (Vischer, 1945). В Happophyceae жг. [ 81 ]
- Akrokont: клетки с жгутиками вставлены апикально
- Subakrokont: клетки с жгутиками вставлены субапически
- Pleurokont: клетки с жгутиками вставлены в боковом направлении
Согласно рисунку избиения:
- скольжение: жгутичок, который тянется на подложке [ 78 ]
- Гетеродинамический: жгутики с различными рисунками взбивания (обычно с одним жгутиком, функционирующим в захвате пищи, а другой функционирует в скольжении, якорре, движении или «рулевом управлении») [ 82 ]
- Изодинамическая: бьют жгутиков с теми же узорами
Другие термины, связанные с типом жгутиков:
- Изоконт: ячейки с жгутией равной длины. Это также раньше использовалось для обозначения хлорофиты
- Anisokont: клетки с жгутией неравной длины, например, некоторые Euglenophyceae и Prasinophyceae
- Гетероконт: термин, введенный Лютером (1899), для обозначения ксантофики , из -за пары жгутиков неравной длины. Он принял определенное значение при ссылке на клетки с передним страминипированным жгутиком (с трехсторонними мастигонемами, в одном или двух рядах) и задней, обычно гладкой жгутиков. Он также используется для обозначения таксона Гетероконты
- Стефаноконт: клетки с коронкой жгутиков возле его передней части, например, гаметы и споры эдиагониала , споры некоторых брайопсидал . Термин, представленный Blackman & Tansley (1902) для обозначения Oedogoniales
- Akont: ячейки без жгутиков. Он также использовался для обозначения таксономических групп, как Aconta или Akonta: Zygnematophyceae и Bacillariophyceae (Oltmanns, 1904) или Rhodophyceae (Christensen, 1962)
Архаальные жгутики
[ редактировать ]Архелум , обладающий некоторыми видами архей , поверхностно похож на бактериальный жгутичок; В 1980 -х годах они считались гомологичными на основе грубой морфологии и поведения. [ 83 ] И жгутики, и архаэлла состоят из филаментов, простирающихся за пределами ячейки, и вращаются, чтобы выдвинуть ячейку. Архаальные жгутики имеют уникальную структуру, которой не хватает центрального канала. Подобно пилинам бактериального типа IV , архейные белки (архаэллины) изготовлены с сигнальными пептидами класса 3, и они обрабатываются препаратевым пептидазоподобным ферментом типа IV. Архаэллины обычно модифицируются путем добавления N-связанных гликанов , которые необходимы для правильной сборки или функции. [ 3 ]
Открытия в 1990 -х годах выявили многочисленные подробные различия между архаальными и бактериальными жгутиками. К ним относятся:
- Вращение бактериального жгутиков питается мотивой протона - поток H + Ионы или время от времени при силе, мощной натрия- поток NA + ионы; Архаальное вращение жгутиков питается АТФ . [ 84 ]
- В то время как бактериальные клетки часто имеют много жгутиковых филаментов, каждая из которых вращается независимо, архаальный жгутичок состоит из пачка многих филаментов, которые вращаются в виде одной сборки.
- Бактериальная жгутика растут путем добавления субъединиц флагеллина на кончике; Архаальные жгутики растут благодаря добавлению субъединиц к основанию.
- Бактериальные жгутики толще, чем архаэлла, а бактериальная нить имеет достаточно большую полая «трубка», в которой субъединицы флагеллина могут течь по внутренней части нити и добавляться на кончике; Архелум слишком тонкий (12-15 нм), чтобы допустить это. [ 85 ]
- Многие компоненты бактериальных жгутиков разделяют сходство последовательности с компонентами систем секреции типа III , но компоненты бактериальной жгутики и архаэлле не имеют сходства последовательностей. Вместо этого, некоторые компоненты архаэллы разделяют последовательность и морфологическое сходство с компонентами PILI типа IV , которые собираются посредством действия систем секреции типа II (номенклатура систем секреции PILI и белков не является согласованной). [ 85 ]
Эти различия подтверждают теорию о том, что бактериальная жгутика и архаэлла являются классическим случаем биологической аналогии или сходящейся эволюции , а не гомологии . [ 86 ] [ 87 ] [ 88 ] Исследование структуры архаэллы добилось значительного прогресса, начиная с начала 2010 -х годов, с первой структурой атомного разрешения белка архаэллы, открытием дополнительных функций археэллы и первыми сообщениями о архаэлле в Наноархиоте и Таумархите. [ 89 ] [ 90 ]
Гриб
[ редактировать ]Единственными грибами , имеющими один жених на своих спорах, являются хитриды . В Batrachochytrium dendrobatidis жгутиковой флаголит длиной 19–20 мкм. [ 91 ] Нефункциональная центриоль лежит рядом с кинетосомой . Девять взаимосвязанных реквизитов прикрепляют кинетосому к плазмалемме , а в переходной зоне присутствует терминальная пластина. Внутренняя структура, похожая на кольцо, прикрепленную к канальцам флагуллических дублетов в переходной зоне, наблюдалась в поперечном сечении. [ 91 ]
Дополнительные изображения
[ редактировать ]-
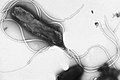 Множественные жгутики в лофотрихазном расположении на поверхности Helicobacter pylori
Множественные жгутики в лофотрихазном расположении на поверхности Helicobacter pylori -
 Физическая модель бактериального жгутика
Физическая модель бактериального жгутика


Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Барди, Соня Л.; Нг, Сэнди Им; Джаррелл, Кен Ф. (1 февраля 2003 г.). «Прокариотические структуры подвижности» . Микробиология . 149 (2): 295–304. doi : 10.1099/mic.0.25948-0 . PMID 12624192 .
- ^ Silflow, Carolyn D.; Лефевр, Пол А. (1 декабря 2001 г.). «Сборка и подвижность эукариотических ресничек и жгутиков. Уроки от chlamydomonas reinhardtii» . Физиология растений . 127 (4): 1500–1507. doi : 10.1104/pp.010807 . PMC 1540183 . PMID 11743094 .
- ^ Jump up to: а беременный Джаррелл, Кен Ф., изд. (2009). Pili и жгутики: текущие исследования и будущие тенденции . Норфолк: Caister Academic Press. ISBN 978-1-904455-48-6 .
- ^ Мало, Аурелио Ф; Гомендио, Монтсеррат; Гарде, Джулиан; Ланг-Лентон, Барбара; Soler, Ana J; Ролдан, Эдуардо Р.С. (22 июня 2006 г.). «Дизайн спермы и функция спермы» . Биологические письма . 2 (2): 246–249. doi : 10.1098/rsbl.2006.0449 . PMC 1618917 . PMID 17148374 .
- ^ Лейси, будь; Rosemore, J (октябрь 2001 г.). «Helicobacter pylori: язвы и многое другое: начало эпохи» . Журнал питания . 131 (10): 2789S - 2793S. doi : 10.1093/jn/131.10.2789s . PMID 11584108 . Архивировано из оригинала (абстрактная страница) 7 февраля 2009 года . Получено 2 июня 2008 года .
- ^ Ван, Цинфенг; Сузуки, Асака; Мариконда, Сусана; Порволлик, Штеффен; Харши, Расика М (1 июня 2005 г.). «Ощущение влажности: новая роль для бактериального жгутика» . Embo Journal . 24 (11): 2034–2042. doi : 10.1038/sj.emboj.7600668 . PMC 1142604 . PMID 15889148 .
- ^ Альберс, Соня-Верена; Джаррелл, Кен Ф. (27 января 2015 г.). «Архаулм: как плавать архаи» . Границы в микробиологии . 6 : 23. DOI : 10.3389/fmicb.2015.00023 . PMC 4307647 . PMID 25699024 .
- ^ Quax, Tef; Albers, SV; Pfeiffer, F (14 декабря 2018 г.). «Такси в археи» . Новые темы в науках о жизни . 2 (4): 535–546. doi : 10.1042/etls20180089 . PMC 7289035 . PMID 33525831 .
- ^ Haimo, Lt; Розенбаум, JL (1 декабря 1981 г.). "Реснички, жгутики и микротрубочки" . Журнал клеточной биологии . 91 (3): 125S - 130S. doi : 10.1083/jcb.91.3.125s . PMC 2112827 . PMID 6459327 .
- ^ Стрейф, Стефан; Стаудингер, Уилфрид Франц; Марван, Вольфганг; Oesterhelt, Dieter (декабрь 2008 г.). «Вращение жгутиков в археонах Halobacterium salinarum зависит от АТФ». Журнал молекулярной биологии . 384 (1): 1–8. doi : 10.1016/j.jmb.2008.08.057 . PMID 18786541 .
- ^ Jump up to: а беременный Альбертс, Брюс (2015). Молекулярная биология клетки (шестое изд.). Нью -Йорк, Нью -Йорк. п. 942. ISBN 9780815344643 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Сильверман, Майкл; Саймон, Мелвин (май 1974). «Вращение жгутиков и механизм бактериальной подвижности». Природа . 249 (5452): 73–74. Bibcode : 1974natur.249 ... 73S . doi : 10.1038/249073a0 . PMID 4598030 . S2CID 10370084 .
- ^ Лоу, Грэм; Мейстер, Маркус; Берг, Говард С. (февраль 1987 г.). «Быстрое вращение жгутиковых пучков в плавательных бактериях». Природа . 325 (6105): 637–640. Bibcode : 1987natur.325..637L . doi : 10.1038/325637A0 . S2CID 4242129 .
- ^ Берг, Говард С.; Андерсон, Роберт А. (октябрь 1973 г.). «Бактерии плавают, вращая их жгутиковые нити». Природа . 245 (5425): 380–382. Bibcode : 1973natur.245..380b . doi : 10.1038/245380a0 . PMID 4593496 . S2CID 4173914 .
- ^ Jahn, TL; Bovee, EC (октябрь 1965 г.). «Движение и локомоция микроорганизмов». Ежегодный обзор микробиологии . 19 (1): 21–58. doi : 10.1146/annurev.mi.19.100165.000321 . PMID 5318439 .
- ^ Харши, RM (2003). «Бактериальная подвижность на поверхности: много способов к общей цели». Ежегодный обзор микробиологии . 57 : 249–73. doi : 10.1146/annurev.micro.57.030502.091014 . PMID 14527279 .
- ^ Нг, Сэнди Им; Чабан, Бонни; Джаррелл, Кен Ф. (2006). «Архаальные жгутики, бактериальная жгутика и Pili типа IV: сравнение генов и посттрансляционных модификаций». Микробная физиология . 11 (3–5): 167–191. doi : 10.1159/000094053 . PMID 16983194 . S2CID 30386932 .
- ^ Metlina, AL (ноябрь 2004 г.). «Бактериальная и архаальная жгутика как прокариотическая подвижность органелл». Биохимия (Москва) . 69 (11): 1203–1212. doi : 10.1007/s10541-005-0065-8 . PMID 15627373 . S2CID 632440 .
- ^ Джаррелл, К. (2009). "Архаальная жгутика и пили". Pili и жгутики: текущие исследования и будущие тенденции . Caister Academic Press. ISBN 978-1-904455-48-6 .
- ^ Макнаб, Роберт М. (октябрь 2003 г.). «Как бактерии собирают жгутики». Ежегодный обзор микробиологии . 57 (1): 77–100. doi : 10.1146/annurev.micro.57.030502.090832 . PMID 12730325 .
- ^ Diószeghy, Zoltán; Заводский, Петер; Намба, Кейичи; Вондервист, Ференк (18 июня 2004 г.). «Стабилизация жгутиковых фильмов от HAP2 Capping». Письма Febs . 568 (1-3): 105-109. Bibcode : 2004febsl.568..105d . Doi : 10.1016/j.febslet.2004.05.029 . PMID 15196929 . S2CID 33886010 .
- ^ Galkin, Vitold E.; Ю, Xiong; Bielnicki, Jakub; Heuser, Джон; Юинг, Шерил П.; Герри, Патриция; Эгельман, Эдвард Х. (18 апреля 2008 г.). «Дивергенция четвертичных структур среди бактериальных жгутиков». Наука . 320 (5874): 382–385. Bibcode : 2008Sci ... 320..382G . doi : 10.1126/science.1155307 . PMID 18420936 . S2CID 7702002 .
- ^ Уорралл, Лиам Дж.; Мажьюски, Дороти Д.; Странадка, Натали С.Дж. (15 сентября 2023 г.). «Структурное понимание систем секреции типа III бактериального жгутика и инъекции» . Ежегодный обзор микробиологии . 77 (1): 669–698. doi : 10.1146/annurev-micro-032521-025503 . ISSN 0066-4227 . PMID 37713458 . S2CID 261963968 .
- ^ Jump up to: а беременный Сингх, Прашант К.; Шарма, Панкадж; Афанзар, Ошри; Goldfarb, Margo H.; Маклашина, Елена; Эйзенбах, Майкл; Чеккини, Гэри; Айверсон, ТМ (17 апреля 2024 г.). «Криоэмные структуры показывают, как бактериальный жгутичок вращается и переключает направление» . Природная микробиология . 9 (5): 1271–1281. doi : 10.1038/s41564-024-01674-1 . ISSN 2058-5276 . PMC 11087270 . PMID 38632342 .
- ^ Atsumi, Tatsuo; Маккартерт, Линда; Imae, Yasuo (январь 1992 г.). «Полярные и боковые жгутиковые двигатели морского вибрации управляются различными иоонными силами». Природа . 355 (6356): 182–184. Bibcode : 1992natur.355..182a . doi : 10.1038/355182A0 . PMID 1309599 . S2CID 4315167 .
- ^ Кодзима, Сейджи; Блэр, Дэвид Ф. (2004), «Бактериальный жгутиковый двигатель: структура и функция сложной молекулярной машины» , Международный обзор цитологии , 233 , Elsevier: 93–134, doi : 10.1016/s0074-7696 (04) 33003-2 , ISBN 978-0-12-364637-8 , PMID 15037363 , извлечен 23 апреля 2024 г.
- ^ Дин, Тим (2 августа 2010 г.). «Внутри самый эффективный мотор природы: жгутиковый» . Австралийский жизненный ученый .
- ^ Нагата, Йошио (июнь 2014 г.). «Разблокируя секреты наномотора природы» . Nikkei Asian Review .
- ^ Уитфилд, Джон (19 июня 2008 г.). «Бактериальные двигатели имеют свое собственное сцепление» . Nature News : News.2008.903. doi : 10.1038/news.2008.903 . Получено 17 мая 2017 года .
- ^ Dusenbery, DB (2009). «Глава 13». Жизнь в микромасштабе: неожиданная физика маленькой . Кембридж: издательство Гарвардского университета. ISBN 978-0-674-03116-6 .
- ^ Хильдебранд, Милтон (ноябрь 1959 г.). «Движения бегущего гепарда и лошади». Журнал млекопитающих . 44 (4): 481–495. doi : 10.2307/1376265 . JSTOR 1376265 . Хотя по Охотник, Люк; Хэмман, Дэйв (2003). Гепарда . Struik Publishers. С. 37–38.
Самая быстро записанная скорость гепарда составила 110 км/ч (68 миль в час)
- ^ Медоуз, Робин (10 мая 2011 г.). «Как бактерии переключают шестерни» . PLOS Биология . 9 (5): E1001061. doi : 10.1371/journal.pbio.1001061 . PMC 3091840 . PMID 21572986 .
- ^ Солнце, Qifang; Юань, Ченгжи; Чжоу, Сайнань; Лу, Цзин; Зенг, Мейан; Cai, Xiong; Песня, Хупан (19 октября 2023 г.). «Helicobacter Pylori Infection: динамический процесс от диагностики до лечения» . Границы в клеточной и инфекционной микробиологии . 13 doi : 10.3389/fcimb.2023.1257817 . PMC 10621068 . PMID 37928189 .
- ^ Минамино, Тору; Имада, Кацуми; Намба, Кейичи (2008). «Механизмы экспорта белка типа III для бактериальной жгутиковой сборки». Молекулярные биосистемы . 4 (11): 1105–1115. doi : 10.1039/b808065h . PMID 18931786 .
- ^ Асакура, Шо; Эгучи, Горо; Iino, Tetsuo (октябрь 1964 г.). «Восстановление бактериальной жгутики in vitro». Журнал молекулярной биологии . 10 (1): 42 - in9. doi : 10.1016/s0022-2836 (64) 80026-7 . PMID 14222895 .
- ^ Jump up to: а беременный Паллен, Марк Дж.; Мацке, Николас Дж. (Октябрь 2006 г.). «От происхождения видов до происхождения бактериальной жгутики». Nature Reviews Microbiology . 4 (10): 784–790. doi : 10.1038/nrmicro1493 . PMID 16953248 . S2CID 24057949 .
- ^ Jump up to: а беременный Saier, M (март 2004 г.). «Эволюция систем секреции белка бактериального типа III». Тенденции в микробиологии . 12 (3): 113–115. doi : 10.1016/j.tim.2004.01.003 . PMID 15001186 .
- ^ Гофна, Ури; Рон, Элиора З.; Граур, Дэн (июль 2003 г.). «Системы секреции бактериального типа III древние и развиваются с помощью множественных горизонтальных событий». Ген . 312 : 151–163. doi : 10.1016/s0378-1119 (03) 00612-7 . PMID 12909351 .
- ^ Макканн, честь С.; Гутман, Дэвид С. (январь 2008 г.). «Эволюция системы секреции типа III и ее эффекторов во взаимодействиях растений -микробов» . Новый фитолог . 177 (1): 33–47. doi : 10.1111/j.1469-8137.2007.02293.x . PMID 18078471 .
- ^ Бег, Майкл Дж. (2007). Край эволюции: поиск границ дарвинизма . Нью -Йорк, Нью -Йорк: Свободная пресса. ISBN 978-0-7432-9620-5 .
- ^ Раджагопала, Seesandra V; Тиц, Бьорн; Голл, Йоханнес; Пэрриш, Джоди Р; Wohlbold, Katrin; Маккевитт, Мэтью Т; Палцкилл, Тимоти; Мори, Хиротада; Финли, Рассел Л; Уэц, Питер (январь 2007 г.). «Белковая сеть бактериальной подвижности» . Биология молекулярных систем . 3 (1): 128. doi : 10.1038/msb4100166 . PMC 1943423 . PMID 17667950 .
- ^ Тиц, Бьорн; Раджагопала, Сайесандра В.; Эстер, Клаудия; Häuser, Roman; Уэц, Питер (ноябрь 2006 г.). «Новый консервативный коэффициент сборки бактериального жгутика» . Журнал бактериологии . 188 (21): 7700–7706. doi : 10.1128/jb.00820-06 . PMC 1636259 . PMID 16936039 .
- ^ Kakkanat, Asha; Фан, Мин-Дюй; Ло, Элвин У.; Битсон, Скотт А.; Schembri, Mark A. (10 мая 2017 г.). «Новые гены, связанные с повышенной подвижностью Escherichia coli ST131» . Plos один . 12 (5): E0176290. Bibcode : 2017ploso..1276290K . doi : 10.1371/journal.pone.0176290 . PMC 5425062 . PMID 28489862 .
- ^ Паллен, MJ; Гофна, У. (2007). «Бактериальная жгутика и секреция типа III: тематические исследования в области эволюции сложности». Динамика генома . 3 : 30–47. doi : 10.1159/000107602 . ISBN 978-3-8055-8340-4 Полем PMID 18753783 .
- ^ «Бактериальная жгутика» (PDF) . Архивировано (PDF) из оригинала 9 октября 2022 года . Получено 29 декабря 2021 года .
- ^ Руан, Хуанфанг; Като, Такаюки; Сантини, Клэр-Лиз; Мията, Томоко; Кавамото, Акихиро; Чжан, Вэй-Цзия; Бернадак, Ален; WU, Long-Fei; Намба, Кейичи (11 декабря 2012 г.). «Архитектура жгутикового аппарата в быстро плавающей магнитотаксической бактерии MO-1» . Труды Национальной академии наук . 109 (50): 20643–20648. BIBCODE : 2012PNAS..10920643R . doi : 10.1073/pnas.1215274109 . PMC 3528567 . PMID 23184985 .
- ^ «Трихо-префикс» . Получено 26 марта 2022 года .
- ^ Echazarreta, MA; Klose, KE (2019). « жгутика Синтез варинового » . Границы в клеточной и инфекционной микробиологии . 9 : 131. doi : 10.3389/fcimb.2019.00131 . PMC 6504787 . PMID 31119103 .
- ^ "Лофо" . Получено 26 марта 2022 года .
- ^ Кудришев, Михаил; Сирклафф, Марек; Baumeister, Wolfgang; Саймон, Маркус М.; Уоллих, Рейнхард; Фришкнехт, Фридрих (март 2009 г.). «Сравнительная криоэлектронная томография патогенных спирохетов болезни Лайма» . Молекулярная микробиология . 71 (6): 1415–1434. doi : 10.1111/j.1365-2958.2009.06613.x . PMID 19210619 . S2CID 19650892 .
- ^ Ким, Юн-Кейонг; Маккартер, Линда Л. (июль 2000 г.). «Анализ полярной жгутиковой генной системы Vibrio parahaemolyticus» . Журнал бактериологии . 182 (13): 3693–3704. doi : 10.1128/jb.182.13.3693-3704.2000 . PMC 94540 . PMID 10850984 .
- ^ Atsumi, t; Маакава, y; Ямада, т; Кававиши, я; Imae, y; Homma, M (август 1996 г.). «Влияние вискозитного на боковое и полярное жгутиков вибраолитического Algolyticus » Журнал бактериологии 178 (16): 5024–5 Doi : 10.1128/ jb.178.16.5024-5026.1 178290PMC PMID 8759871
- ^ Маккартер, Линда Л. (2004). «Двойные жгутиковые системы обеспечивают подвижность в разных обстоятельствах». Микробная физиология . 7 (1–2): 18–29. doi : 10.1159/000077866 . PMID 15170400 . S2CID 21963003 .
- ^ Мерино, Сюзанна; Шоу, Джонатан Дж.; Том, Джон М. (октябрь 2006 г.). "Бактериальная боковая жгутика: невидимая система жгутиков " FMS Микробиологические письма 263 (2): 127–1 Doi : 10.1111/j.1574-6968.2006.00403.x . PMID 16978346
- ^ Белас, R; Саймон, м; Сильверман, М (июль 1986 г.). «Регуляция транскрипции гена бокового жгутиков в Vibrio parahaemolyticus» . Журнал бактериологии . 167 (1): 210–218. doi : 10.1128/jb.167.1.210-218.1986 . PMC 212863 . PMID 3013835 .
- ^ Каналы, Роцио; Альтернала, Мэри; Вильч, Сильвия; Хорсбург, Гэвин; Шоу, Джонатан Дж.; Томас, Джон М.; Мерино, Сюзанна (февраль 2006 г.). Аролиз Гидрофила Журнал бактерологии 188 (3): 852–8 doi : 10.1128/ jb.188.352-8 PMC 1347325 . PMID 16428388 .
- ^ Каналы, Роцио; Рамирес, Сильвия; Вильч, Сильвия; Хорсбург, Гэвин; Шоу, Джонатан Дж.; Томас, Джон М.; Мерино, Сюзанна (15 января 2006 г.). гидрофила " Журнал бактерологии 188 (2): 542–555. doi : 10.1128/ jb.188.542-555.200 PMC 1347287 . PMID 163385045 .
- ^ Ким, Мунджу; Птица, Джеймс С.; Ван Парис, Аннемари Дж.; Breuer, Kenneth S.; Пауэрс, Томас Р. (23 декабря 2003 г.). «Макроскопическая масштабная модель бактериальной жгутиковой связки» . Труды Национальной академии наук . 100 (26): 15481–15485. arxiv : cond-mat/0312562 . Bibcode : 2003pnas..10015481K . doi : 10.1073/pnas.2633596100 . PMC 307593 . PMID 14671319 .
- ^ MacNab, RM (январь 1977 г.). «Бактериальная жгутика, вращающаяся в пучках: исследование в спиральной геометрии» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (1): 221–5. Bibcode : 1977pnas ... 74..221M . doi : 10.1073/pnas.74.1.221 . PMC 393230 . PMID 264676 .
- ^ Тейлор, FJR Max (1 ноября 2003 г.). «Крысок системы с двумя ценностями, рост профитологии и основание Международного общества эволюционной протистологии (ISEP)» . Международный журнал систематической и эволюционной микробиологии . 53 (6): 1707–1714. doi : 10.1099/ijs.0.02587-0 . PMID 14657097 .
- ^ Хюльсманн, Норберт (август 1992 г.). «Undulipodium: конец бесполезного обсуждения». Европейский журнал профистологии . 28 (3): 253–257. doi : 10.1016/s0932-4739 (11) 80231-2 . PMID 23195228 .
- ^ Jump up to: а беременный в Adl, Sina M.; Симпсон, Аластер Г.Б.; Лейн, Кристофер Э.; Люкш, Юлиус; Басс, Дэвид; Баузер, Сэмюэль С.; Браун, Мэтью В.; Бурки, Фабен; Данторн, Мика; Хэмпл, Владимир; Хейсс, Аарон; Хоппенрат, Мона; Лара, Энрике; Le Gall, Line; Линн, Денис Х.; Макманус, Хилари; Митчелл, Эдвард А.Д.; Mozley-Stanridge, Sharon E.; Parfrey, Laura W.; Павловский, Ян; Рукерт, Соня; Шадвик, Лора; Schoch, Conrad L.; Смирнов, Алексей; Spiegel, Frederick W. (сентябрь 2012 г.). «Пересмотренная классификация эукариот» . Журнал эукариотической микробиологии . 59 (5): 429–514. doi : 10.1111/j.1550-7408.2012.00644.x . PMC 3483872 . PMID 23020233 .
- ^ Андерсен, Ра; Барр, диджеи; Линн, DH; Melkonian, M.; Moestroup, Ø.; Sleigh, MA (февраль 1991 г.). «Терминология и номенклатура элементов цитоскелета, связанных с жгутиковым/ресничным аппаратом в протистах». Протоплазма . 164 (1–3): 1–8. doi : 10.1007/bf01320809 . S2CID 40755371 .
- ^ Leadbeater, Barry SC; Грин, Джон С., ред. (2000). Жгутики: единство, разнообразие и эволюция . Специальный том систематической ассоциации. Тол. 59. Тейлор и Фрэнсис. ISBN 978-1-4822-6822-5 .
- ^ Jump up to: а беременный Барсанти, Лора; Gualtieri, Paolo (2006). Ворот: анатомия, биохимия и биотехнология . Флорида, США: CRC Press. ISBN 9780203492598 .
- ^ Jump up to: а беременный Линдеманн, CB; Лесич, Ка (15 февраля 2010 г.). «Бламенное и ресничное избиение: проверенное и возможное». Журнал сотовой науки . 123 (Pt 4): 519–28. doi : 10.1242/jcs.051326 . PMID 20145000 . S2CID 18673550 .
- ^ Jump up to: а беременный Lodish, Харви; Берк, Арнольд; Зипурский, С. Лоуренс; Мацудайра, Пол; Балтимор, Дэвид; Дарнелл, Джеймс (2000). «Раздел 19.4cilia и жгутики: структура и движение». Реснички и жгутики: структура и движение . ISBN 0-7167-3136-3 .
- ^ Пазур, Грегори Дж. (Октябрь 2004 г.). «Внутренний транспорт и ресничка-зависимые почечные заболевания: цилиарная гипотеза о поликистозном заболевании почек» . Журнал Американского общества нефрологии . 15 (10): 2528–2536. doi : 10.1097/01.asn.0000141055.57643.e0 . PMID 15466257 .
- ^ Юбуки, Наоджи; Леандер, Брайан С. (июль 2013 г.). «Эволюция центров организации микротрубочек через дерево эукариот» . Заводский журнал . 75 (2): 230–244. doi : 10.1111/tpj.12145 . PMID 23398214 .
- ^ Ворон, JA (2000). «Условие жгутиков» . Leadbeater & Green 2000 , с. 27–48 . CRC Press. ISBN 9781482268225 .
- ^ Jump up to: а беременный Вебстер, Джон; Вебер, Роланд (25 января 2007 г.). «Споры грибов» . 2007 (3 -е изд.). Кембридж: издательство Кембриджского университета. С. 23–24. ISBN 9781139461504 .
- ^ Лар, Даниэль Дж.Г.; Парфри, Лора Вегенер; Митчелл, Эдвард А.Д.; Кац, Лора А.; Лара, Энрике (22 июля 2011 г.). «Целомудрие амебы: переоценка доказательств секса в амебоидных организмах» . Труды Королевского общества B: Биологические науки . 278 (1715): 2081–2090. doi : 10.1098/rspb.2011.0289 . PMC 3107637 . PMID 21429931 .
- ^ Jump up to: а беременный Остин, Кр (1995). Грудзинскас, Юргис Гедиминас; Йович, JL (ред.). Гамет - сперматозон . Кембридж: издательство Кембриджского университета. ISBN 9780521479967 .
- ^ Юг, Гр; Whittick, A (1987). Введение в фикологию . Оксфорд: Blackwell Scientific Publications. п. 65. ISBN 9781444314205 .
- ^ Dodge, JD (1973). Тонкая структура клеток водорослей . Лондон: академическая пресса. С. 57–79. ISBN 9780323158237 .
- ^ Lee, Re (2008). Фикология (4 -е изд.). Издательство Кембриджского университета. п. 7 ISBN 9781139469876 Полем
Ли трубчатые волосы.
- ^ Корлисс, Джо; Lom, J (2000). «Аннотированный глоссарий протозоологических терминов». В Ли, JJ; Leedale, GF; Брэдбери, П. (ред.). Иллюстрированное руководство по простейшим . Тол. 2 (2 -е изд.). Общество простейших. С. 1346–85. ISBN 1891276239 .
- ^ Jump up to: а беременный Иоук, Александра; Arndt, Hartmut (ноябрь 2013 г.). «Короткое руководство по общим гетеротрофным жгутикам пресноводных мест обитания на основе морфологии живых организмов» . Профист . 164 (6): 842–860. doi : 10.1016/j.protis.2013.08.003 . PMID 24239731 .
- ^ Sleigh, M (1989). Протозои и другие профисты . Лондон: Эдвард Арнольд. С. 98–99. ISBN 9780521428057 .
- ^ Sparrow, FK (1960). Водные фикомицеты (2 -е изд.). Энн Арбор: Мичиган: Университет Мичиганской прессы. п. 15
- ^ Хибберд, DJ (1976). «Ультраструктура и таксономия Chrysophyceae и Prymnesiophyceae (Haptophyceae): обследование с некоторыми новыми наблюдениями по ультраструктуре Chrysophyceae». Журнал Лондонского общества Линни, ботаника . 72 (2): 55–80. doi : 10.1111/j.1095-8339.1976.tb01352.x .
- ^ Sleigh, MA (1985). «Происхождение и эволюция жгутикового движения» . Cell Motil . 5 : 137–138. Архивировано с оригинала 3 марта 2016 года . Получено 21 февраля 2016 года .
- ^ Cavalier-Smith, T (1987). «Происхождение эукариотических и архебактериальных клеток» . Анналы нью -йоркской академии наук . 503 (1): 17–54. Bibcode : 1987nyasa.503 ... 17c . doi : 10.1111/j.1749-6632.1987.tb40596.x . PMID 3113314 . S2CID 38405158 . [ Постоянная мертвая ссылка ]
- ^ Мэдиган, Майкл Т. (2019). Брок биология микроорганизмов (пятнадцатая, глобальная изд.). Нью -Йорк, Нью -Йорк. С. 70–71. ISBN 9781292235103 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Jump up to: а беременный Гош, Абраджёти; Альберс, Соня-Верена (1 февраля 2011 года). «Сборка и функция архаального жгутика». Биохимическое общество транзакций . 39 (1): 64–69. doi : 10.1042/bst0390064 . PMID 21265748 .
- ^ Томас, Нихил А.; Барди, Соня Л.; Джаррелл, Кен Ф. (апрель 2001 г.). «Архаальная жгутика: другой вид прокариотической структуры подвижности». Обзоры микробиологии FEMS . 25 (2): 147–174. doi : 10.1111/j.1574-6976.2001.tb00575.x . PMID 11250034 . S2CID 34411164 .
- ^ Чимильский, Скотт; Папке, Р. Тайн (2015). «Получив архейный PILI типа IV: расширяющийся репертуар клеточных придаток, подразумевает сложную регуляцию и разнообразные функции» . Границы в микробиологии . 6 : 362. DOI : 10.3389/fmicb.2015.00362 . ISSN 1664-302X . PMC 4419858 . PMID 25999922 .
- ^ Де Соуза Мачадо, Дж. Нуно; Воллмар, Леони; Шимпф, Джулия; Чаудхури, Паушали; Кумария, Рашми; Ван дер делает, Крис; Хьюсель, Торстен; Альберс, Соня-Верена (2021). «Аутофосфорилирование каоподобного белка Arlh ингибирует олигомеризацию и взаимодействие с Arli, моторной Атпазой архаулма» . Молекулярная микробиология . 116 (3): 943–956. doi : 10.1111/mmi.14781 . ISSN 0950-382X . PMID 34219289 .
- ^ Нуно де Соуза Мачадо, Жоао; Альберс, Соня-Верена; Даум, Бертрам (2022). «На пути к выяснению вращающегося механизма архаллума» . Границы в микробиологии . 13 Doi : 10.3389/fmicb.2022.848597 . ISSN 1664-302X . PMC 8978795 . PMID 35387068 .
- ^ Джаррелл, Кен Ф; Альберс, Соня-Верена; Мачадо, Дж. Нуно де Соуза (2021). «Комплексная история подвижности и археэлляции в археи» . FEMS микробы . 2 : XTAB002. doi : 10.1093/femsmc/xtab002 . ISSN 2633-6685 . PMC 10117864 . PMID 37334237 .
- ^ Jump up to: а беременный Longcore, Joyce E.; Пессиер, Аллан П.; Николс, Дональд К. (март 1999 г.). « Batrachochytrium dendrobatidis Gen. Et sp. Nov. , Хитридный патогенный для амфибий». Микология . 91 (2): 219–227. doi : 10.2307/3761366 . JSTOR 3761366 .
Дальнейшее чтение
[ редактировать ]- Берг, Говард С. (январь 2000 г.). «Модильное поведение бактерий» . Физика сегодня . 53 (1): 24–29. Bibcode : 2000pht .... 53a..24b . doi : 10.1063/1,882934 . S2CID 178516210 .
- Линдеманн, Чарльз (4 апреля 2008 г.). «Механизмы подвижности спермы» . Оклендский университет. Архивировано из оригинала 16 мая 2008 года . Получено 18 мая 2008 года .
- Purcell, EM (1977). «Жизнь на низком чисел Рейнольдса» (PDF) . Американский журнал физики . 45 (1): 3–11. Bibcode : 1977AMJPH..45 .... 3P . doi : 10.1119/1.10903 . HDL : 2433/226838 . Архивировано из оригинала (PDF) 5 июня 2011 года . Получено 19 октября 2009 года .
- Мацке, Нью -Джерси (10 ноября 2003 г.). «Эволюция в (Браунском) пространстве: модель для происхождения бактериального жгутика» . talkdesign.org.
Внешние ссылки
[ редактировать ]
![]() Эта статья включает в себя текст из публикации, который сейчас в общественном доступе : Chambers, Ephraim , ed. (1728). Циклопдия, или универсальный словарь искусств и наук (1 -е изд.). Джеймс и Джон Кнаптон и др.
Эта статья включает в себя текст из публикации, который сейчас в общественном доступе : Chambers, Ephraim , ed. (1728). Циклопдия, или универсальный словарь искусств и наук (1 -е изд.). Джеймс и Джон Кнаптон и др. {{cite encyclopedia}}: Отсутствует или пусто |title= ( помощь )