Рецептор клеточной поверхности

Рецепторы клеточной поверхности ( мембранные рецепторы , трансмембранные рецепторы ) — рецепторы встроенные в плазматическую мембрану клетки , . [1] Они участвуют в передаче сигналов в клетках , получая (связываясь) внеклеточные молекулы . Это специализированные интегральные мембранные белки , обеспечивающие связь между клеткой и внеклеточным пространством . Внеклеточные молекулы могут быть гормонами , нейротрансмиттерами , цитокинами , факторами роста , молекулами клеточной адгезии или питательными веществами ; они реагируют с рецептором, вызывая изменения в метаболизме и активности клетки. В процессе сигнала передачи связывание лиганда влияет на каскадные химические изменения через клеточную мембрану.
Структура и механизм
[ редактировать ]Многие мембранные рецепторы представляют собой трансмембранные белки . Существуют различные виды, включая гликопротеины и липопротеины . [2] Известны сотни различных рецепторов, и многие еще предстоит изучить. [3] [4] Трансмембранные рецепторы обычно классифицируются на основе их третичной (трехмерной) структуры. Если трехмерная структура неизвестна, их можно классифицировать на основе топологии мембраны . В простейших рецепторах полипептидные цепи пересекают липидный бислой один раз, тогда как другие, такие как рецепторы, связанные с G-белком , пересекают целых семь раз. Каждая клеточная мембрана может иметь несколько типов мембранных рецепторов с различным распределением на поверхности. Один и тот же рецептор также может по-разному распределяться в разных положениях мембраны, в зависимости от типа мембраны и клеточной функции. Рецепторы часто группируются на поверхности мембраны, а не распределяются равномерно. [5] [6]
Механизм
[ редактировать ]Для объяснения механизма действия трансмембранных рецепторов были предложены две модели.
- Димеризация . Модель димеризации предполагает, что до связывания лиганда рецепторы существуют в мономерной форме. Когда происходит связывание агониста, мономеры объединяются, образуя активный димер .
- Вращение : связывание лиганда с внеклеточной частью рецептора вызывает вращение (конформационное изменение) части трансмембранных спиралей рецептора. Вращение изменяет, какие части рецептора открываются на внутриклеточной стороне мембраны, изменяя то, как рецептор может взаимодействовать с другими белками внутри клетки. [7]
Домены
[ редактировать ]P = плазматическая мембрана
Я = внутриклеточное пространство
Трансмембранные рецепторы плазматической мембраны обычно можно разделить на три части.
Внеклеточные домены
[ редактировать ]Внеклеточный домен находится снаружи клетки или органеллы . Если полипептидная цепь пересекает бислой несколько раз, внешний домен содержит петли, переплетенные через мембрану. По определению, основная функция рецептора — распознавать тип лиганда и реагировать на него. Например, нейротрансмиттер , гормон или атомарные ионы могут связываться с внеклеточным доменом в качестве лиганда, связанного с рецептором. Клото — это фермент, который воздействует на рецептор, распознавая лиганд ( FGF23 ).
Трансмембранные домены
[ редактировать ]Двумя наиболее распространенными классами трансмембранных рецепторов являются GPCR и однопроходные трансмембранные белки . [8] [9] В некоторых рецепторах, таких как никотиновый ацетилхолиновый рецептор , трансмембранный домен образует белковую пору через мембрану или вокруг ионного канала . При активации внеклеточного домена путем связывания соответствующего лиганда пора становится доступной для ионов, которые затем диффундируют. В других рецепторах трансмембранные домены претерпевают конформационные изменения при связывании, что влияет на внутриклеточные условия. В некоторых рецепторах, таких как члены суперсемейства 7TM , трансмембранный домен включает лиганд-связывающий карман.
Внутриклеточные домены
[ редактировать ]Внутриклеточный (или цитоплазматический ) домен рецептора взаимодействует с внутренней частью клетки или органеллы, передавая сигнал. Существует два основных пути такого взаимодействия:
- Внутриклеточный домен взаимодействует посредством белок-белковых взаимодействий с эффекторными белками , которые, в свою очередь, передают сигнал по назначению.
- В случае рецепторов, связанных с ферментами , внутриклеточный домен обладает ферментативной активностью . Часто это активность тирозинкиназы . Ферментативная активность также может быть обусловлена ферментом, связанным с внутриклеточным доменом.
Преобразование сигнала
[ редактировать ]
Процессы передачи сигнала через мембранные рецепторы включают внешние реакции, при которых лиганд связывается с мембранным рецептором, и внутренние реакции, при которых запускается внутриклеточный ответ. [10] [11]
Передача сигнала через мембранные рецепторы состоит из четырех частей:
- Внеклеточная сигнальная молекула: внеклеточная сигнальная молекула вырабатывается одной клеткой и, по крайней мере, способна перемещаться в соседние клетки.
- Рецепторный белок: клетки должны иметь рецепторные белки клеточной поверхности, которые связываются с сигнальной молекулой и сообщаются внутрь клетки.
- Внутриклеточные сигнальные белки: они передают сигнал органеллам клетки. Связывание сигнальной молекулы с белком-рецептором активирует внутриклеточные сигнальные белки, которые инициируют сигнальный каскад.
- Белки-мишени: конформации или другие свойства белков-мишеней изменяются, когда сигнальный путь активен и меняет поведение клетки. [11]

Мембранные рецепторы в основном делятся по структуре и функциям на 3 класса: рецепторы, связанные с ионными каналами ; Фермент -связанный рецептор ; и рецептор, связанный с G-белком .
- Рецепторы, связанные с ионными каналами, имеют ионные каналы для анионов и катионов и составляют большое семейство многопроходных трансмембранных белков. Они участвуют в событиях быстрой передачи сигналов, обычно обнаруживаемых в электрически активных клетках, таких как нейроны . Их еще называют лиганд-управляемыми ионными каналами . Открытие и закрытие ионных каналов контролируется нейромедиаторами .
- Рецепторы, связанные с ферментами, либо сами являются ферментами, либо непосредственно активируют ассоциированные ферменты. Обычно это однопроходные трансмембранные рецепторы, при этом ферментативный компонент рецептора остается внутриклеточным. Большинство фермент-связанных рецепторов представляют собой протеинкиназы или связаны с ними.
- Рецепторы, связанные с G-белком, представляют собой интегральные мембранные белки, имеющие семь трансмембранных спиралей. Эти рецепторы активируют G-белок при связывании агониста , а G-белок опосредует действие рецептора на внутриклеточные сигнальные пути.
Рецептор, связанный с ионным каналом
[ редактировать ]Во время передачи сигнала в нейроне нейромедиатор связывается с рецептором и изменяет конформацию белка. Это открывает ионный канал, пропуская внеклеточные ионы в клетку. Изменяется ионная проницаемость плазматической мембраны, и это преобразует внеклеточный химический сигнал во внутриклеточный электрический сигнал, который изменяет возбудимость клетки . [12]
Рецептор ацетилхолина представляет собой рецептор, связанный с катионным каналом. Белок состоит из четырех субъединиц: альфа (α), бета (β), гамма (γ) и дельта (δ). Есть две α-субъединицы, каждая с одним сайтом связывания ацетилхолина . Этот рецептор может существовать в трех конформациях. Закрытое и незанятое состояние является нативной конформацией белка. Поскольку обе молекулы ацетилхолина связываются с сайтами связывания на α-субъединицах, конформация рецептора изменяется, и ворота открываются, позволяя проникать многим ионам и небольшим молекулам. Однако это открытое и занятое состояние длится лишь незначительное время, а затем ворота закрываются, переходя в закрытое и занятое состояние. Две молекулы ацетилхолина вскоре отделятся от рецептора, вернув его в нативное закрытое и незанятое состояние. [13] [14]
Ферментасвязанные рецепторы
[ редактировать ]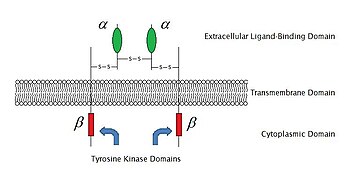
По состоянию на 2009 год известно 6 типов фермент-связанных рецепторов : рецепторные тирозинкиназы ; рецепторы, ассоциированные с тирозинкиназой; Рецептоподобные тирозинфосфатазы ; Рецепторные серин / треониновые киназы ; Рецепторы гуанилилциклазы и рецепторы, ассоциированные с гистидинкиназой . Рецепторные тирозинкиназы имеют самую большую популяцию и самое широкое применение. Большинство этих молекул являются рецепторами для факторов роста, таких как эпидермальный фактор роста (EGF), фактор роста тромбоцитов (PDGF), фактор роста фибробластов (FGF), фактор роста гепатоцитов (HGF), фактор роста нервов (NGF) и гормоны. например, инсулин .Большинство этих рецепторов димеризуются после связывания со своими лигандами, чтобы активировать дальнейшую сигнальную трансдукцию. Например, после того, как рецептор эпидермального фактора роста (EGF) связывается со своим лигандом EGF, два рецептора димеризуются, а затем подвергаются фосфорилированию остатков тирозина в ферментной части каждой молекулы рецептора. Это активирует тирозинкиназу и катализирует дальнейшие внутриклеточные реакции.
Рецепторы, связанные с G-белком
[ редактировать ]Рецепторы, связанные с G-белком, составляют большое семейство белков трансмембранных рецепторов. Они встречаются только у эукариот . [15] Лиганды , которые связывают и активируют эти рецепторы, включают: светочувствительные соединения, запахи , феромоны , гормоны и нейротрансмиттеры . Они различаются по размеру: от небольших молекул до пептидов и крупных белков . Рецепторы, связанные с G-белком, участвуют во многих заболеваниях и поэтому являются мишенью многих современных медицинских препаратов. [16]
Существует два основных пути передачи сигнала с участием рецепторов, связанных с G-белком: сигнальный путь цАМФ и сигнальный путь фосфатидилинозитола . [17] Оба опосредованы активацией G-белка . G-белок представляет собой тримерный белок с тремя субъединицами, обозначенными как α, β и γ. В ответ на активацию рецептора α-субъединица высвобождает связанный гуанозиндифосфат (GDP), который заменяется гуанозинтрифосфатом (GTP), тем самым активируя α-субъединицу, которая затем диссоциирует от β- и γ-субъединиц. Активированная α-субъединица может дополнительно влиять на внутриклеточные сигнальные белки или непосредственно на функциональные белки-мишени.
Заболевания, связанные с мембранными рецепторами
[ редактировать ]Если мембранные рецепторы денатурированы или недостаточны, передача сигнала может быть затруднена и вызвать заболевания. Некоторые заболевания вызваны нарушениями функции мембранных рецепторов. Это происходит из-за дефицита или деградации рецептора из-за изменений в генах, которые кодируют и регулируют рецепторный белок. Мембранный рецептор TM4SF5 влияет на миграцию печеночных клеток и гепатомы . [18] Кроме того, кортикальный NMDA-рецептор влияет на текучесть мембран и изменяется при болезни Альцгеймера. [19] Когда клетка заражается вирусом без оболочки, вирус сначала связывается со специфическими мембранными рецепторами, а затем передает себя или субвирусный компонент на цитоплазматическую сторону клеточной мембраны. В случае полиовируса известно in vitro, что взаимодействия с рецепторами вызывают конформационные перестройки, которые высвобождают вирионный белок, называемый VP4. N-конец VP4 миристилирован и, следовательно, гидрофобен ( миристиновая кислота =CH 3 (CH 2 ) 12 COOH). Предполагается, что конформационные изменения, вызванные связыванием рецептора, приводят к присоединению миристиновой кислоты к VP4 и образованию канала для РНК.
Структурно-ориентированный дизайн лекарств
[ редактировать ]
Благодаря таким методам, как рентгеновская кристаллография и ЯМР-спектроскопия , информация о трехмерных структурах целевых молекул резко возросла, как и структурная информация о лигандах. Это стимулирует быстрое развитие разработки лекарств на основе структуры . Некоторые из этих новых препаратов нацелены на мембранные рецепторы. Современные подходы к разработке лекарств на основе структуры можно разделить на две категории. Первая категория касается определения лигандов для данного рецептора. Обычно это достигается посредством запросов к базе данных, биофизического моделирования и создания химических библиотек. В каждом случае проверяется большое количество потенциальных молекул-лигандов, чтобы найти те, которые подходят для связывающего кармана рецептора. Этот подход обычно называют разработкой лекарств на основе лигандов. Ключевое преимущество поиска в базе данных заключается в том, что он экономит время и силы для получения новых эффективных соединений. Другой подход к разработке лекарств на основе структуры заключается в комбинаторном картировании лигандов, который называется дизайном лекарств на основе рецепторов. В этом случае молекулы-лиганды конструируются в рамках связывающего кармана путем поэтапной сборки небольших частей. Эти части могут быть либо атомами, либо молекулами. Ключевым преимуществом такого метода является возможность открытия новых структур. [20] [21] [22]
Другие примеры
[ редактировать ]- Адренергический рецептор
- Обонятельные рецепторы
- Рецепторные тирозинкиназы
- Рецептор эпидермального фактора роста
- Инсулиновый рецептор
- Рецепторы фактора роста фибробластов ,
- Высокоаффинные рецепторы нейротрофинов
- Рецепторы эфрина
- Интегрины
- Рецептор фактора роста нервов с низким сродством
- NMDA-рецептор
- Несколько иммунных рецепторов
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «9.3: Сигнальные молекулы и клеточные рецепторы – типы рецепторов» . Свободные тексты по биологии . 12 июля 2018 года . Проверено 24 июля 2023 г.
- ^ Куатрекасас П. (1974). «Мембранные рецепторы». Ежегодный обзор биохимии . 43 : 169–214. дои : 10.1146/annurev.bi.43.070174.001125 . ПМИД 4368906 . S2CID 44727052 .
- ^ Даутценберг Ф.М., Хаугер Р.Л. (февраль 2002 г.). «Семейство пептидов CRF и их рецепторы: обнаружено еще больше партнеров». Тренды Фармакол. наук. 23 (2): 71–7. дои : 10.1016/S0165-6147(02)01946-6 . ПМИД 11830263 .
- ^ Ривьер С., Шалле Л., Флюгге Д., Шпер М., Родригес I (май 2009 г.). «Белки, подобные рецептору формилпептида, представляют собой новое семейство вомероназальных хемосенсоров». Природа . 459 (7246): 574–7. дои : 10.1038/nature08029 . ПМИД 19387439 .
- ^ Ротберг К.Г.; Ин Ю.С.; Камен Б.А.; Андерсон Р.Г. (1990). «Холестерин контролирует кластеризацию мембранного рецептора 5-метилтетрагидрофолата, закрепленного на гликофосфолипиде» . Журнал клеточной биологии . 111 (6): 2931–2938. дои : 10.1083/jcb.111.6.2931 . ПМК 2116385 . ПМИД 2148564 .
- ^ Джейкобсон К.; Коте ПД; Росси С.Г.; Ротундо РЛ; Карбонетто С. (2001). «Дистрогликановый комплекс необходим для стабилизации кластеров рецепторов ацетилхолина в нервно-мышечных соединениях и формирования синаптической базальной мембраны» . Журнал клеточной биологии . 152 (3): 435–450. дои : 10.1083/jcb.152.3.435 . ПМК 2195998 . ПМИД 11157973 .
- ^ Маруяма, Ичиро Н. (1 сентября 2015 г.). «Активация трансмембранных рецепторов клеточной поверхности по общему механизму? «Модель вращения» » . Биоэссе . 37 (9): 959–967. doi : 10.1002/bies.201500041 . ISSN 1521-1878 . ПМК 5054922 . ПМИД 26241732 .
- ^ Суперсемейства однопроходных трансмембранных рецепторов в базе данных Membranome.
- ^ Суперсемейства однопроходных трансмембранных белковых лигандов и регуляторов рецепторов в базе данных Membranome.
- ^ Ульриха А., Шлессингерб Й.; Шлессинджер, Дж (1990). «Передача сигнала рецепторами с тирозинкиназной активностью». Клетка . 61 (2): 203–212. дои : 10.1016/0092-8674(90)90801-К . ПМИД 2158859 .
- ^ Jump up to: а б Кеннет Б. Стори (1990). Функциональный метаболизм . Wiley-IEEE. стр. 87–94. ISBN 978-0-471-41090-4 .
- ^ Хилле Б. (2001). Ионные каналы возбудимых мембран . Сандерленд, Массачусетс. ISBN 978-0-87893-321-1 .
- ^ Миядзава А.; Фудзиёси Ю.; Анвин Н. (2003). «Структура и механизм открытия поры рецептора ацетилхолина». Природа . 423 (6943): 949–955. дои : 10.1038/nature01748 . ПМИД 12827192 .
- ^ Акабас М.Х.; Штауффер Д.А.; Сюй М.; Карлин А. (1992). «Структура канала рецептора ацетилхолина исследована у мутантов с цистеиновым замещением». Наука . 258 (5080): 307–310. дои : 10.1126/science.1384130 . ПМИД 1384130 .
- ^ Кинг Н., Хиттингер К.Т., Кэрролл С.Б. (2003). «Эволюция семейств ключевых клеточных сигнальных белков и белков адгезии предшествует животному происхождению». Наука . 301 (5631): 361–3. дои : 10.1126/science.1083853 . ПМИД 12869759 .
- ^ Филмор, Дэвид (2004). «Это мир GPCR» . Современное открытие лекарств . 2004 (ноябрь): 24–28.
- ^ Гилман А.Г. (1987). «G-белки: преобразователи сигналов, генерируемых рецепторами». Ежегодный обзор биохимии . 56 : 615–649. дои : 10.1146/annurev.bi.56.070187.003151 . ПМИД 3113327 . S2CID 33992382 .
- ^ Мюллер-Пиллаша Ф.; Валраппа К.; Лачера У.; Фрисс Х.; Бюхлерб М.; Адлера Г.; Гресс ТМ (1998). «Идентификация нового опухолеассоциированного антигена TM4SF5 и его экспрессия при раке человека». Джин . 208 (1): 25–30. дои : 10.1016/S0378-1119(97)00633-1 . ПМИД 9479038 .
- ^ Шойер К.; Марасб А.; Гаттазб ВФ; Кэрнс Н.; Фёрстльб Х.; Мюллер В.Е. (1996). «Свойства кортикальных рецепторов NMDA и текучесть мембран изменяются при болезни Альцгеймера». Деменция 7 (4): 210–214. дои : 10.1159/000106881 . ПМИД 8835885 .
- ^ Ван Р.; Гао Ю.; Лай Л. (2000). «LigBuilder: многоцелевая программа для разработки лекарств на основе структуры». Журнал молекулярного моделирования . 6 (7–8): 498–516. дои : 10.1007/s0089400060498 .
- ^ Шнайдер Г.; Фехнер У. (2005). «Компьютерный дизайн молекул, подобных лекарствам, de novo». Nature Reviews Открытие лекарств . 4 (8): 649–663. дои : 10.1038/nrd1799 . ПМИД 16056391 .
- ^ Йоргенсен В.Л. (2004). «Множество ролей вычислений в открытии лекарств». Наука . 303 (5665): 1813–1818. дои : 10.1126/science.1096361 . ПМИД 15031495 . S2CID 1307935 .
Внешние ссылки
[ редактировать ]- База данных IUPHAR GPCR, заархивированная 23 марта 2019 г. на Wayback Machine.
- Клетка + поверхность + рецепторы в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|