Гуанилатциклаза
| Гуанилатциклаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 4.6.1.2 | ||
| Номер CAS. | 9054-75-5 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Гуанилатциклаза (EC 4.6.1.2, также известная как гуанилциклаза , гуанилилциклаза или GC ; систематическое название GTP-дифосфат-лиаза (циклизующая; 3',5'-циклическая-GMP-образующая) ) представляет собой лиазы фермент , который превращает гуанозинтрифосфат. (GTP) на циклический гуанозинмонофосфат (цГМФ) и пирофосфат : [1]
- GTP = 3',5'-циклический GMP + дифосфат
Часто он является частью G-белка сигнального каскада , который активируется низкими уровнями внутриклеточного кальция и ингибируется высокими уровнями внутриклеточного кальция. В ответ на уровень кальция гуанилатциклаза синтезирует цГМФ из ГТФ. , управляемые цГМФ цГМФ сохраняет открытыми каналы , позволяя кальцию проникать в клетку. [2]
Подобно цАМФ , цГМФ является важным вторичным мессенджером , который интернализует информацию, передаваемую межклеточными мессенджерами, такими как пептидные гормоны и оксид азота , а также может функционировать как аутокринный сигнал . [1] В зависимости от типа клеток он может вызывать изменения в адаптации/развитии, требующие синтеза белка . В гладких мышцах цГМФ является сигналом к расслаблению и связан со многими гомеостатическими механизмами, включая регуляцию вазодилатации , голосового тонуса, секреции инсулина и перистальтики . После образования цГМФ может расщепляться фосфодиэстеразами , которые сами находятся под разными формами регуляции, в зависимости от ткани.
Реакция
[ редактировать ]Гуанилатциклаза катализирует реакцию гуанозинтрифосфата (ГТФ) с 3',5'-циклическим гуанозинмонофосфатом (цГМФ) и пирофосфатом :

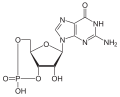
Эффекты
[ редактировать ]Гуанилатциклаза обнаруживается в сетчатке (RETGC) и модулирует зрительную фототрансдукцию в палочках и колбочках . Это часть системы отрицательной обратной связи кальция , которая активируется в ответ на гиперполяризацию фоторецепторов светом . Это приводит к уменьшению внутриклеточного кальция, что стимулирует белки, активирующие гуанилатциклазу (GCAP). Исследования показали, что синтез цГМФ в колбочках примерно в 5-10 раз выше, чем в палочках, что может играть важную роль в модуляции адаптации колбочек к свету. [3] Кроме того, исследования показали, что рыбки данио экспрессируют большее количество GCAP, чем млекопитающие, и что GCAP рыбок данио могут связывать как минимум три иона кальция. [4]
Гуанилатциклаза 2C (GC-C) представляет собой фермент, экспрессирующийся преимущественно в нейронах кишечника. Активация GC-C усиливает возбуждающий клеточный ответ, который модулируется рецепторами глутамата и ацетилхолина . GC-C, известный главным образом своей секреторной регуляцией в эпителии кишечника , также экспрессируется в головном мозге. Точнее, он обнаруживается в соматах и дендритах дофаминергических нейронов вентральной покрышки (ВТА) и черной субстанции . Некоторые исследования предполагают, что этот путь играет роль в дефиците внимания и гиперактивном поведении . [5]
Растворимая гуанилатциклаза содержит молекулу гема и активируется в первую очередь связыванием оксида азота (NO) с этим гемом. [6] рГЦ является первичным рецептором NO, газообразного мембранорастворимого нейромедиатора . Было показано, что экспрессия sGC является самой высокой в полосатом теле по сравнению с другими областями мозга и рассматривается как возможный кандидат на восстановление дисфункции полосатого тела при болезни Паркинсона . sGC действует как внутриклеточный посредник для регуляции дофамина и глутамата. Повышение регуляции цГМФ в полосатом теле, обедненном дофамином, которое создает чувствительность нейронов, связано с симптомами болезни Паркинсона. Показано, что повышение внутриклеточного цГМФ способствует чрезмерной возбудимости нейронов и двигательной активности. Активация этого пути может также стимулировать пресинаптическое высвобождение глутамата и вызывать постсинаптическое усиление АМРА- рецепторов. [7]
Типы
[ редактировать ]Существуют мембраносвязанные (тип 1, гуанилатциклазный рецептор ) и растворимые (тип 2, растворимая гуанилатциклаза ) формы гуанилатциклазы.
Мембраносвязанные гуанилатциклазы включают внешний лиганд-связывающий домен (например, для пептидных гормонов, таких как BNP и ANP ), трансмембранный домен и внутренний каталитический домен, гомологичный аденилатциклазам . [8] Недавно у водных грибов была обнаружена непосредственно светозависимая гуанилатциклаза. [9] [10]
В сетчатке млекопитающих были идентифицированы две формы гуанилатциклазы, каждая из которых кодируется отдельными генами; РЕТГК-1 и РЕТГК-2 . Было обнаружено, что RETGC-1 экспрессируется в колбочках на более высоких уровнях по сравнению с клетками-палочками. Исследования также показали, что мутации в гене RETGC-1 могут привести к дистрофии колбочек-палочек за счет нарушения процессов фототрансдукции.
Мутации
[ редактировать ]Колбочковая дистрофия (ХПК) представляет собой деградацию функции фоторецепторов сетчатки, при которой функция колбочек утрачивается в начале дистрофии, но функция палочек сохраняется почти до конца. ХПК связана с несколькими генетическими мутациями, включая мутации активатора гуанилатциклазы 1A (GUCA1A) и гуанилатциклазы 2D (GUY2D) среди других ферментов. Точнее, GUY2D кодирует RETGC-1, который участвует в адаптации колбочек и чувствительности фоторецепторов путем синтеза цГМФ. Низкие концентрации кальция вызывают димеризацию белков RETGC-1 за счет стимуляции белков, активирующих гуанилатциклазу (GCAP). Этот процесс происходит в аминокислотах 817–857, и мутации в этой области увеличивают сродство RETGC-1 к GCAP. Это позволяет изменить чувствительность нейрона к кальцию, позволяя мутантному RETGC-1 активироваться с помощью GCAP при более высоких уровнях кальция, чем у дикого типа. Поскольку RETGC-1 вырабатывает цГМФ, который сохраняет открытыми циклические нуклеотидные каналы, обеспечивая приток кальция, эта мутация вызывает чрезвычайно высокие внутриклеточные уровни кальция. Кальций, который играет множество ролей в клетке и жестко регулируется, разрушает мембрану, когда появляется в избытке. Кроме того, кальций связан с апоптоз , вызывая высвобождение цитохрома с . Следовательно, мутации в RETGC-1 могут вызывать ХПК за счет повышения внутриклеточных уровней кальция и стимуляции гибели фоторецепторов колбочек. [11]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Мартин, Эмиль; Берка, Владимир; Цай, Ах-Лим; Мурад, Ферид (2005). «Растворимая гуанилилциклаза: рецептор оксида азота». Методы энзимологии . Том. 396. Эльзевир. стр. 478–492. дои : 10.1016/s0076-6879(05)96040-0 . ISBN 978-0-12-182801-1 . ISSN 0076-6879 . ПМИД 16291255 .
Растворимая гуанилатциклаза признана наиболее чувствительным физиологическим рецептором к оксиду азота. Связывание оксида азота с гемовой частью циклазы индуцирует ее способность синтезировать второй мессенджер цГМФ.
- ^ Сакурай К.; Чэнь Дж.; Кефалов В. (2011). «Роль модуляции гуанилатциклазы в фототрансдукции конуса мыши» . Журнал неврологии . 31 (22): 7991–8000. doi : 10.1523/jneurosci.6650-10.2011 . ПМК 3124626 . ПМИД 21632921 .
- ^ Такемото Н., Татибанаски С., Кавамура С. (2009). «Высокая синтетическая активность цГМФ в шишках карпа» . Proc Natl Acad Sci США . 106 (28): 11788–11793. Бибкод : 2009PNAS..10611788T . дои : 10.1073/pnas.0812781106 . ПМК 2710672 . ПМИД 19556550 .
- ^ Схолтен А, Кох К (2011). «Дифференциальная передача сигналов кальция с помощью колбочек-специфичных белков, активирующих гуанилатциклазу, из сетчатки рыбок данио» . ПЛОС ОДИН . 6 (8): e23117. Бибкод : 2011PLoSO...623117S . дои : 10.1371/journal.pone.0023117 . ПМК 3149064 . ПМИД 21829700 .
- ^ Гонг Р., Дин С., Ху Дж., Лу Й., Лю Ф., Манн Э., Сюй Ф., Коэн М., Луо М. (2011). «Роль мембранного рецептора гуанилатциклазы-с в дефиците внимания и гиперактивном поведении».
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Дербишир ER, Марлетта Массачусетс (2009). «Биохимия растворимой гуанилатциклазы». CGMP: Генераторы, эффекторы и терапевтические последствия . Справочник по экспериментальной фармакологии. Том. 191. стр. 17–31. дои : 10.1007/978-3-540-68964-5_2 . ISBN 978-3-540-68960-7 . ПМИД 19089323 . S2CID 25597341 .
- ^ Ценг К., Кабальеро А., Дек А., Касс Д., Симак Н., Суну Э., Парк М., Блюм С., Саммут С., Парк Д., Уэст (2011). «Ингибирование передачи сигналов стриарной растворимой гуанилатциклазы-цГМФ обращает вспять дисфункцию базальных ганглиев и акинезию при экспериментальном паркинсонизме» . ПЛОС ОДИН . 6 (11): e27187. Бибкод : 2011PLoSO...627187T . дои : 10.1371/journal.pone.0027187 . ПМК 3206945 . ПМИД 22073284 .
- ^ Кун М (2003). «Структура, регуляция и функция рецепторов гуанилатциклазы мембран млекопитающих с акцентом на гуанилатциклазу-А» . Исследование кровообращения . 93 (8): 700–709. дои : 10.1161/01.res.0000094745.28948.4d . ПМИД 14563709 .
- ^ Гао С.К., Нагпал Дж., Шнайдер М.В., Козьяк-Павлович В., Нагель Г., Готшалк А. (июль 2015 г.). «Оптогенетическое манипулирование цГМФ в клетках и животных с помощью жестко регулируемого светом гуанилатциклазного опсина CyclOp» . Природные коммуникации . 6 (8046): 8046. Бибкод : 2015NatCo...6.8046G . дои : 10.1038/ncomms9046 . ПМЦ 4569695 . ПМИД 26345128 .
- ^ Шейб У., Штефест К., Джи К.Э., Кёршен Х.Г., Фудим Р., Ортнер Т.Г., Хегеманн П. (2015). «Родопсин-гуанилатциклаза водного гриба Blastoladiella emersonii обеспечивает быстрый оптический контроль передачи сигналов цГМФ» . Научная сигнализация . 8 (389): р8. doi : 10.1126/scisignal.aab0611 . ПМИД 26268609 . S2CID 13140205 .
- ^ Ойос-Гарсия М., Ауз-Александр С., Альмогуера Б., Канталапьедра Д., Ривейро-Альварес Р., Лопес-Мартинес А. и др. (2011). «Анализ мутаций в кодоне 838 гена гуанилатциклазы 2D в испанских семьях с аутосомно-доминантной колбочкой, колбочкой-стержнем и макулярной дистрофией» . Молекулярное видение . 17 : 1103–1109. ПМК 3087450 . ПМИД 21552474 .
Внешние ссылки
[ редактировать ]- Гуанилат + циклаза Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|