Дайсер
| ДАЙСЕР1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | DICER1 , DCR1, Dicer, Dicer1e, HERNA, MNG1, RMSE2, K12H4.8-LIKE, dicer 1, рибонуклеаза III, GLOW | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 606241 ; МГИ : 2177178 ; Гомологен : 13251 ; GeneCards : DICER1 ; OMA : DICER1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Dicer , также известная как эндорибонуклеаза Dicer или хеликаза с мотивом РНКазы , представляет собой фермент , который у человека кодируется DICER1 геном . Являясь частью семейства РНКаз III , Dicer расщепляет двухцепочечную РНК (дсРНК) и пре-микроРНК (пре-миРНК) на короткие фрагменты двухцепочечной РНК, называемые малыми интерферирующими РНК и микроРНК соответственно. Эти фрагменты имеют длину примерно 20–25 пар оснований с выступом из двух оснований на 3'-конце . Dicer способствует активации РНК-индуцированного комплекса молчания (RISC), который необходим для РНК-интерференции . RISC имеет каталитический компонент Argonaute , который представляет собой эндонуклеазу, способную расщеплять информационную РНК (мРНК).
Открытие
[ редактировать ]Название Dicer было дано в 2001 году аспиранткой -Брук Стоуни Эмили Бернштейн во время проведения исследований в Грегори Хэннона лаборатории в лаборатории Колд-Спринг-Харбор . Бернштейн стремился открыть фермент, ответственный за образование небольших фрагментов РНК из двухцепочечной РНК. Способность Дайсера генерировать около 22 нуклеотидных фрагментов РНК была обнаружена путем отделения его от ферментного комплекса RISC после инициации пути RNAi с помощью трансфекции дцРНК . Этот эксперимент показал, что RISC не несет ответственности за генерацию наблюдаемых небольших нуклеотидных фрагментов. Последующие эксперименты по проверке способности ферментов семейства РНКазы III создавать фрагменты РНК сузили поиск до Drosophila CG4792, теперь называемого Dicer. [ 5 ]
Dicer Ортологи присутствуют во многих других организмах. [ 6 ] У мха Physcomitrella patens DCL1b, один из четырех белков DICER, участвует не в биогенезе микроРНК, а в нарезке целевых транскриптов микроРНК. Таким образом, был открыт новый механизм регуляции экспрессии генов — эпигенетическое молчание генов с помощью микроРНК. [ 7 ]
Что касается кристаллической структуры, первым исследованным дайцером был кристалл простейшего Giardia кишечная . Работа была выполнена Иэном Макреем во время проведения исследований в качестве постдокторанта в Дженнифер Дудны лаборатории в Калифорнийском университете в Беркли . Домен PAZ и два домена РНКазы III были обнаружены с помощью рентгеновской кристаллографии . Размер белка составляет 82 кДа , что представляет собой консервативное функциональное ядро, которое впоследствии было обнаружено в более крупных белках Dicer в других организмах; например, у человека он составляет 219 кДа. Разница в размерах между человеческими и G. кишечными Dicer обусловлена присутствием по меньшей мере пяти различных доменов в человеческом Dicer. Эти домены важны для регуляции активности Dicer, процессинга дцРНК и функционирования белкового фактора РНК-интерференции. [ 8 ]
Функциональные домены
[ редактировать ]
Человеческий дайцер (также известный как hsDicer или DICER1 ) классифицируется как рибонуклеаза III, поскольку он расщепляет двухцепочечную РНК. Помимо двух доменов РНКазы III, он содержит домен геликазы PAZ ( Piwi / Argonaute /Zwille) , домен , [ 10 ] [ 11 ] и два двухцепочечных РНК-связывающих домена (DUF283 и dsRBD). [ 8 ] [ 12 ]
Текущие исследования показывают, что домен PAZ способен связывать 2-нуклеотидный 3'-конец дцРНК, в то время как каталитические домены РНКазы III образуют псевдодимер вокруг дцРНК, инициируя расщепление цепей. Это приводит к функциональному укорочению цепи дцРНК. Расстояние между доменами PAZ и RNaseIII определяется углом спирали соединителя и влияет на длину продукта микроРНК. [ 9 ] Домен dsRBD связывает dsRNA, хотя специфический сайт связывания домена не определен. Возможно, что этот домен работает в составе комплекса с другими белками-регуляторами (TRBP у человека, R2D2, Loqs у дрозофилы ), чтобы эффективно позиционировать домены РНКазы III и, таким образом, контролировать специфичность продуктов мРНК. [ 13 ] Домен хеликазы участвует в обработке длинных субстратов. [ 13 ]
Роль в интерференции РНК
[ редактировать ]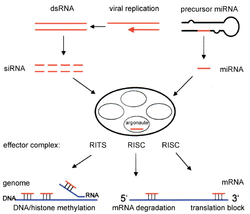
Микро РНК
[ редактировать ]РНК-интерференция — это процесс, при котором распад молекул РНК на микроРНК ингибирует экспрессию генов определенных последовательностей мРНК хозяина. МикроРНК вырабатывается внутри клетки , начиная с первичной микроРНК (при-миРНК) в ядре . Эти длинные последовательности расщепляются на более мелкие предшественники микроРНК (пре-миРНК), которые обычно состоят из 70 нуклеотидов и имеют шпилечную структуру . При-миРНК идентифицируются DGCR8 и расщепляются Дрошей с образованием пре-миРНК, процесс, который происходит в ядре. Эти пре-миРНК затем экспортируются в цитоплазму, где они расщепляются Dicer с образованием зрелой микроРНК. [ 15 ]
Малая интерферирующая РНК
[ редактировать ]Малые интерферирующие РНК (миРНК) производятся и функционируют аналогично микроРНК путем расщепления двухцепочечной РНК с помощью Dicer на более мелкие фрагменты длиной от 21 до 23 нуклеотидов. [ 13 ] И микроРНК, и миРНК активируют РНК-индуцированный комплекс молчания (RISC), который находит комплементарную последовательность мРНК-мишени и расщепляет РНК с помощью РНКазы. [ 16 ] Это, в свою очередь, заставляет конкретный ген замолчать посредством РНК-интерференции. [ 17 ] siRNA и miRNA отличаются тем, что siRNA обычно специфичны по отношению к последовательности мРНК, тогда как miRNA не полностью комплементарны последовательности мРНК. микроРНК могут взаимодействовать с мишенями, имеющими схожие последовательности, что ингибирует трансляцию различных генов. [ 18 ] В целом, РНК-интерференция является неотъемлемой частью нормальных процессов внутри таких организмов, как человек, и эта область исследуется как диагностический и терапевтический инструмент для борьбы с раком. [ 15 ]

Болезнь
[ редактировать ]Дегенерация желтого пятна
[ редактировать ]Возрастная дегенерация желтого пятна является распространенной причиной слепоты в развитых странах. Роль Дайсера в этом заболевании стала очевидной после того, как было обнаружено, что у пораженных пациентов наблюдалось снижение уровня Дайсера в пигментном эпителии сетчатки (ПЭС). Мыши с нокаутом Дайсера, у которых Дайсер отсутствовал только в их RPE, демонстрировали аналогичные симптомы. Однако у других мышей, у которых отсутствуют важные белки пути RNAi, такие как Drosha и Pasha , не наблюдалось симптомов дегенерации желтого пятна, как у мышей с нокаутом Dicer. Это наблюдение предполагает специфическую роль Dicer в здоровье сетчатки, которая не зависит от пути RNAi и, следовательно, не является функцией генерации si/miRNA. форма РНК, называемая Alu РНК (транскрипты РНК элементов alu Было обнаружено, что ), повышена у пациентов с недостаточным уровнем Dicer. Эти некодирующие цепи РНК могут образовывать петли, образующие структуры дцРНК, которые будут разрушены Дайсером в здоровой сетчатке. Однако при недостаточном уровне Dicer накопление alu РНК приводит к дегенерации РПЭ в результате воспаления. [ 19 ] [ 20 ]
Рак
[ редактировать ]Измененные профили экспрессии микроРНК при злокачественных опухолях предполагают ключевую роль микроРНК и, следовательно, более важную роль в развитии рака и прогнозе. miRNAs могут действовать как супрессоры опухолей, и поэтому их измененная экспрессия может приводить к онкогенезу . [ 21 ] При анализе рака легких и яичников плохой прогноз и уменьшение времени выживаемости пациентов коррелируют со снижением экспрессии dicer и drosha . Снижение уровня мРНК dicer коррелирует с поздней стадией опухоли. Однако высокая экспрессия dicer наблюдается при других видах рака, таких как рак простаты. [ 22 ] и пищевода коррелирует с плохим прогнозом для пациентов. Это несоответствие между типами рака предполагает, что уникальные процессы регуляции РНКи с участием дайсера различаются среди разных типов опухолей. [ 15 ]
Дайсер также участвует в восстановлении ДНК . Повреждение ДНК увеличивается в клетках млекопитающих со снижением экспрессии Dicer в результате снижения эффективности репарации повреждений ДНК и других механизмов. Например, siRNA из двухцепочечных разрывов (продуцируемых Dicer) может действовать как проводник для белковых комплексов, участвующих в механизмах репарации двухцепочечных разрывов, а также может направлять хроматина модификации . Кроме того, паттерны экспрессии микроРНК изменяются в результате повреждения ДНК, вызванного ионизирующим или ультрафиолетовым излучением . Механизмы RNAi ответственны за молчание транспозонов , и в их отсутствие, например, когда Dicer нокаутирован/выключен, это может привести к активации транспозонов, которые вызывают повреждение ДНК. Накопление повреждений ДНК может привести к появлению в клетках онкогенных мутаций и, как следствие, к развитию опухоли. [ 15 ]
Другие условия
[ редактировать ]многоузловой зоб с шванноматозом Было показано, что является аутосомно-доминантным заболеванием, связанным с мутациями в этом гене. [ 23 ]
Вирусный патогенез
[ редактировать ]Заражение РНК-вирусами может запустить каскад РНКи. Вероятно, дайсер участвует в вирусном иммунитете , поскольку вирусы, поражающие как растительные, так и животные клетки, содержат белки, предназначенные для ингибирования ответа РНКи. У людей вирусы ВИЧ-1 , гриппа и коровьей оспы кодируют такие белки, подавляющие РНКи. Ингибирование дайсера полезно для вируса, поскольку дайсер способен расщеплять вирусную дцРНК и загружать продукт в RISC, что приводит к целенаправленной деградации вирусной мРНК; таким образом борясь с инфекцией. Другим потенциальным механизмом вирусного патогенеза является блокада дайсера как способ ингибирования путей клеточной микроРНК. [ 24 ]
У насекомых
[ редактировать ]У дрозофилы Dicer-1 генерирует микроРНК (миРНК) путем процессинга пре-миРНК, Dicer-2 отвечает за производство малых интерферирующих РНК (миРНК) из длинной двухцепочечной РНК (дцРНК). [ 25 ] Насекомые могут использовать Dicer как мощное противовирусное средство . Это открытие особенно важно, учитывая, что комары ответственны за передачу многих вирусных заболеваний, включая потенциально смертельные арбовирусы : вирус Западного Нила , лихорадку денге и желтую лихорадку . [ 26 ] Хотя комары, а точнее вид Aedes aegypti , служат переносчиками этих вирусов, они не являются предполагаемыми хозяевами вируса. Передача происходит в результате потребности самки комара в крови позвоночных для развития яиц. Путь РНКи у насекомых очень похож на путь РНКи у других животных; Dicer-2 расщепляет вирусную РНК и загружает ее в комплекс RISC, где одна цепь служит матрицей для продукции продуктов РНКи, а другая разрушается. Насекомые с мутациями, приводящими к нефункциональным компонентам пути РНКи, демонстрируют повышенную вирусную нагрузку для вирусов, которые они несут, или повышенную восприимчивость к вирусам, для которых они являются хозяевами. Подобно людям, вирусы насекомых выработали механизмы, позволяющие избегать пути РНКи. Например, дрозофилы C вирус кодирует белок 1A, который связывается с дцРНК, тем самым защищая ее от расщепления дайсером, а также от загрузки RISC. 3a Heliothis virescens Асковирус кодирует фермент РНКазу III, аналогичный доменам РНКазы III дайсера, который может конкурировать за субстрат дцРНК, а также разрушать дуплексы миРНК для предотвращения загрузки RISC. [ 27 ]
Диагностическое и терапевтическое применение
[ редактировать ]Dicer можно использовать для определения наличия опухолей в организме на основе уровня экспрессии фермента. Исследование показало, что у многих пациентов, больных раком, уровень экспрессии Dicer снизился. То же исследование показало, что более низкая экспрессия Dicer коррелирует с более низкой продолжительностью выживания пациентов. [ 15 ] Помимо того, что Dicer является диагностическим инструментом , его можно использовать для лечения пациентов путем внутривенного введения чужеродной миРНК , чтобы вызвать подавление генов. [ 28 ]
Было показано, что миРНК доставляется двумя способами у видов млекопитающих, таких как мыши. Одним из способов было бы прямое внедрение в систему, для чего не требовалась бы функция Dicer. Другой способ — ввести его с помощью плазмид, кодирующих короткую шпилечную РНК, которая расщепляется Dicer на siRNA. [ 29 ]
Одним из преимуществ использования Dicer для терапевтического производства siRNA является специфичность и разнообразие мишеней, на которые он может воздействовать, по сравнению с тем, что используется в настоящее время, например, с антителами или низкомолекулярными ингибиторами. В целом, низкомолекулярные ингибиторы сложны с точки зрения специфичности и имеют непереносимые побочные эффекты. Антитела столь же специфичны, как и миРНК, но их можно использовать только против лигандов или поверхностных рецепторов . С другой стороны, низкая эффективность внутриклеточного поглощения является основным препятствием для инъекции миРНК. [ 15 ] Введенная SiRNA имеет плохую стабильность в крови и вызывает стимуляцию неспецифического иммунитета . [ 30 ] Кроме того, терапевтическому производству микроРНК недостает специфичности, поскольку для прикрепления микроРНК к мРНК требуется всего лишь 6-8 пар нуклеотидных оснований. [ 31 ]
Белки, подобные дайсеру
[ редактировать ]Геномы растений кодируют белки, подобные кубикам, с теми же функциями и белковыми доменами, что и кубики животных и насекомых. Например, в модельном организме Arabidopsis thaliana образуются четыре белка, подобные дайсерам, которые обозначаются от DCL1 до DCL4. DCL1 участвует в генерации микроРНК и продукции мРНК из инвертированных повторов. DCL2 создает миРНК из цис-действующих антисмысловых транскриптов, которые способствуют вирусному иммунитету и защите. DCL3 генерирует миРНК, которая способствует модификации хроматина, а DCL4 участвует в транс-действующем метаболизме миРНК и подавлении транскриптов на посттранскрипционном уровне. Кроме того, DCL 1 и 3 важны для цветения арабидопсиса . У Arabidopsis нокаут DCL не вызывает серьезных проблем в развитии.
Рис и виноград также производят DCL, поскольку механизм нарезки кубиков является общей защитной стратегией многих организмов. Рис развил другие функции для пяти DCL, которые он производит, и они играют более важную роль в функционировании и развитии, чем у Arabidopsis . Кроме того, характер экспрессии различается у разных типов растительных клеток риса, тогда как экспрессия у арабидопсиса более однородна . На экспрессию DCL в рисе могут влиять условия биологического стресса, включая засуху, засоление и холод. Таким образом, эти стрессоры могут снизить устойчивость растения к вирусам. В отличие от арабидопсиса , потеря функции белков DCL вызывает дефекты развития у риса. [ 32 ]
См. также
[ редактировать ]- экспрессия генов
- РИСК
- РНК-интерференция
- микроРНК
- Малая интерферирующая РНК
- Дроша
- Рибонуклеаза III
- мРНК
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000100697 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000041415 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Бернштейн Э., Коди А.А., Хаммонд С.М., Хэннон Г.Дж. (2001). «Роль бидентатной рибонуклеазы на этапе инициации РНК-интерференции». Природа . 409 (6818): 363–6. Бибкод : 2001Natur.409..363B . дои : 10.1038/35053110 . ПМИД 11201747 . S2CID 4371481 .

- ^ Яскевич Л., Филипович В. (2008). «Роль Дайсера в посттранскрипционном подавлении РНК». РНК-интерференция . Актуальные темы микробиологии и иммунологии. Том. 320. стр. 77–97. дои : 10.1007/978-3-540-75157-1_4 . ISBN 978-3-540-75156-4 . ПМИД 18268840 .
- ^ Храйвеш Б., Ариф М.А., Сеумель Г.И., Оссовски С., Вейгель Д., Рески Р. и др. (январь 2010 г.). «Транскрипционный контроль экспрессии генов с помощью микроРНК» . Клетка . 140 (1): 111–22. дои : 10.1016/j.cell.2009.12.023 . ПМИД 20085706 .
- ^ Перейти обратно: а б Лау П.В., Поттер К.С., Каррагер Б., Макрей И.Дж. (октябрь 2009 г.). «Структура комплекса Dicer-TRBP человека по данным электронной микроскопии» . Структура . 17 (10): 1326–32. дои : 10.1016/j.str.2009.08.013 . ПМК 2880462 . ПМИД 19836333 .
- ^ Перейти обратно: а б Макрэ И.Дж., Чжоу К., Ли Ф., Репик А., Брукс А.Н., Канде В.З. и др. (январь 2006 г.). «Структурная основа процессинга двухцепочечной РНК с помощью Dicer». Наука . 311 (5758): 195–8. Бибкод : 2006Sci...311..195M . дои : 10.1126/science.1121638 . ПМИД 16410517 . S2CID 23785494 .
- ^ «Ген Энтреза: DICER1 Dicer1, гомолог Dcr-1 (дрозофила)» .
- ^ Мацуда С., Ичиготани Ю., Окуда Т., Иримура Т., Накацугава С., Хамагути М. (январь 2000 г.). «Молекулярное клонирование и характеристика нового гена человека (HERNA), который кодирует предполагаемую РНК-геликазу». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1490 (1–2): 163–9. дои : 10.1016/S0167-4781(99)00221-3 . ПМИД 10786632 .
- ^ Хаммонд С.М. (октябрь 2005 г.). «Нарезка кубиками и нарезка: основной механизм пути РНК-интерференции». Письма ФЭБС . 579 (26): 5822–9. дои : 10.1016/j.febslet.2005.08.079 . ПМИД 16214139 . S2CID 14495726 .
- ^ Перейти обратно: а б с Ченик Е.С., Фукунага Р., Лу Г., Датчер Р., Ван Ю., Танака Холл Т.М. и др. (апрель 2011 г.). «Фосфат и R2D2 ограничивают субстратную специфичность Dicer-2, АТФ-управляемой рибонуклеазы» . Молекулярная клетка . 42 (2): 172–84. doi : 10.1016/j.molcel.2011.03.002 . ПМК 3115569 . ПМИД 21419681 .
- ^ Хаммонд С.М., Бернштейн Э., Бич Д., Хэннон Г.Дж. (март 2000 г.). «РНК-направленная нуклеаза опосредует посттранскрипционное молчание генов в клетках дрозофилы». Природа . 404 (6775): 293–6. Бибкод : 2000Natur.404..293H . дои : 10.1038/35005107 . ПМИД 10749213 . S2CID 9091863 .
- ^ Перейти обратно: а б с д и ж Мерритт В.М., Бар-Эли М., Суд А.К. (апрель 2010 г.). «Рискованная роль Dicer: последствия для терапии RNAi» . Исследования рака . 70 (7): 2571–4. дои : 10.1158/0008-5472.CAN-09-2536 . ПМК 3170915 . ПМИД 20179193 .
- ^ Вермюлен А., Белен Л., Рейнольдс А., Вольфсон А., Маршалл В.С., Карпилов Дж. и др. (май 2005 г.). «Вклад структуры дцРНК в специфичность и эффективность Dicer» . РНК . 11 (5): 674–82. дои : 10.1261/rna.7272305 . ПМК 1370754 . ПМИД 15811921 .
- ^ Уотсон Джей Ди (2008). Молекулярная биология гена . Сан-Франциско, Калифорния: Лабораторное издательство Колд-Спринг-Харбор. стр. 641–648. ISBN 978-0-8053-9592-1 .
- ^ Цзэн Ю, Йи Р, Каллен Б.Р. (август 2003 г.). «МикроРНК и малые интерферирующие РНК могут ингибировать экспрессию мРНК с помощью аналогичных механизмов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (17): 9779–84. Бибкод : 2003PNAS..100.9779Z . дои : 10.1073/pnas.1630797100 . ПМК 187842 . ПМИД 12902540 .
- ^ Мейстер Г. (март 2011 г.). «Видение: Дайсер появляется в поле зрения» . Природа . 471 (7338): 308–9. Бибкод : 2011Natur.471..308M . дои : 10.1038/471308a . ПМИД 21412326 .
- ^ Таралло В., Хирано Ю., Гельфанд Б.Д., Дриди С., Керур Н., Ким Ю. и др. (май 2012 г.). «Потеря DICER1 и РНК Alu вызывают возрастную дегенерацию желтого пятна через воспалительную сому NLRP3 и MyD88» . Клетка . 149 (4): 847–59. дои : 10.1016/j.cell.2012.03.036 . ПМК 3351582 . ПМИД 22541070 .
- ^ Тан К.Ф., Рен Х (2012). «Роль дайсера в восстановлении повреждений ДНК» . Международный журнал молекулярных наук . 13 (12): 16769–78. дои : 10.3390/ijms131216769 . ПМЦ 3546719 . ПМИД 23222681 .
- ^ Чиосеа С., Железцова Е., Чандран У., Acquafondata M., Макхейл Т., Соболь Р.В. и др. (ноябрь 2006 г.). «Повышающая регуляция dicer, компонента механизма микроРНК, при аденокарциноме простаты» . Американский журнал патологии . 169 (5): 1812–20. дои : 10.2353/ajpath.2006.060480 . ПМК 1780192 . ПМИД 17071602 .
- ^ Ривера Б, Надаф Дж, Фахиминия С, Апелланис-Руис М, Саскин А, Чонг А.С., Шарма С, Вагенер Р, Ревиль Т, Конделло В, Харра З, Хамель Н, Саббагиан Н, Мучантеф К, Томас С, де Кок Л, Эбер-Блуэн М.Н., Бассенден А.В., Рабенштайн Х., Мете О., Пашке Р., Пустазери М.П., Паулюс В., Бергюс А., Рагуссис Дж., Никифоров Ю.Е., Зиберт Р., Альбрехт С., Тюркотт Р., Хассельблатт М., Фабиан М.Р., Фоулкс В.Д. (2019) Дефект микропроцессора DGCR8 характеризует семейный многоузловой зоб с шванноматозом. Джей Клин Инвест
- ^ Берхаут Б., Хааснут Дж. (май 2006 г.). «Взаимодействие между вирусной инфекцией и механизмом интерференции клеточной РНК» . Письма ФЭБС . 580 (12): 2896–902. дои : 10.1016/j.febslet.2006.02.070 . ПМЦ 7094296 . ПМИД 16563388 .
- ^ Ченик Е.С., Фукунага Р., Лу Г., Датчер Р., Ван Ю., Танака Холл Т.М. и др. (апрель 2011 г.). «Фосфат и R2D2 ограничивают субстратную специфичность Dicer-2, АТФ-управляемой рибонуклеазы» . Молекулярная клетка . 42 (2): 172–184. doi : 10.1016/j.molcel.2011.03.002 . ПМК 3115569 . ПМИД 21419681 .
- ^ «Болезни, передающиеся комарами» . Национальный центр инфекционных заболеваний, Центр по контролю и профилактике заболеваний. Архивировано из оригинала 31 января 2014 года . Проверено 22 апреля 2014 г.
- ^ Бронхорст А.В., ван Рей Р.П. (август 2014 г.). «Противовирусная защита в целом: иммунитет насекомых на основе малой РНК». Современное мнение в вирусологии . 7 :19–28. дои : 10.1016/j.coviro.2014.03.010 . ПМИД 24732439 .
- ^ Камлах Ф., Юл Б.Г., Ли С., Ланг Н., Марш Л.М., Сигер В. и др. (март 2009 г.). «Внутривенная инъекция миРНК, направленная против факторов, индуцируемых гипоксией, продлевает выживаемость на модели рака легких Льюиса» . Генная терапия рака . 16 (3): 195–205. дои : 10.1038/cgt.2008.71 . ПМИД 18818708 .
- ^ «Замалчивание генов с помощью РНК-интерференции регулярно используется для изучения функции генов в культивируемых клетках млекопитающих» . Технологии жизни . Проверено 23 апреля 2014 г.
- ^ Шиффелерс Р.М., Ансари А., Сюй Дж., Чжоу К., Тан К., Сторм Г. и др. (2004). «Терапия миРНК рака путем селективной доставки в опухоль пространственно стабилизированных наночастиц, нацеленных на лиганд» . Исследования нуклеиновых кислот . 32 (19): е149. дои : 10.1093/нар/gnh140 . ПМК 528817 . ПМИД 15520458 .
- ^ Чи С.В., Занг Дж.Б., Меле А., Дарнелл Р.Б. (июль 2009 г.). «Argonaute HITS-CLIP декодирует карты взаимодействия микроРНК-мРНК» . Природа . 460 (7254): 479–86. Бибкод : 2009Natur.460..479C . дои : 10.1038/nature08170 . ПМЦ 2733940 . ПМИД 19536157 .
- ^ Лю Ц, Фэн Ю, Чжу Цзы (август 2009 г.). «Dicer-подобные (DCL) белки в растениях». Функциональная и интегративная геномика . 9 (3): 277–86. дои : 10.1007/s10142-009-0111-5 . ПМИД 19221817 . S2CID 28801338 .