микроРНК

МикроРНК ( миРНК ) представляют собой небольшие одноцепочечные некодирующие молекулы РНК, содержащие от 21 до 23 нуклеотидов . [ 1 ] МикроРНК, обнаруженные в растениях, животных и некоторых вирусах, участвуют в подавлении РНК и посттранскрипционной регуляции экспрессии генов . [ 2 ] [ 3 ] микроРНК пара оснований к комплементарным последовательностям в молекулах мРНК , [ 4 ] затем заставляют молчать указанные молекулы мРНК с помощью одного или нескольких из следующих процессов: [ 1 ] [ 5 ]
- Расщепление цепи мРНК на две части.
- Дестабилизация мРНК за счет укорочения ее поли(А)-хвоста , или
- Снижение трансляции мРНК в белки.
В клетках человека и других животных микроРНК в первую очередь действуют путем дестабилизации мРНК. [ 6 ] [ 7 ]
микроРНК напоминают малые интерферирующие РНК (миРНК) пути РНК-интерференции (РНКи) , за исключением того, что микроРНК происходят из областей транскриптов РНК, которые складываются сами по себе, образуя короткие шпильки, тогда как миРНК происходят из более длинных областей двухцепочечной РНК . [ 2 ] Геном человека может кодировать более 1900 микроРНК. [ 8 ] [ 9 ] Однако только около 500 человеческих микроРНК представляют собой настоящие микроРНК в вручную курируемой базе данных генов микроРНК MirGeneDB . [ 10 ]
микроРНК широко распространены во многих типах клеток млекопитающих. [ 11 ] [ 12 ] МикроРНК, по-видимому, нацелены примерно на 60% генов человека и других млекопитающих. [ 13 ] [ 14 ] Многие микроРНК эволюционно консервативны, что означает, что они выполняют важные биологические функции. [ 15 ] [ 1 ] Например, со времен общего предка млекопитающих и рыб сохранилось 90 семейств микроРНК, и большинство из этих консервативных микроРНК выполняют важные функции, как показали исследования, в которых были нокаутированы гены одного или нескольких членов семейства. у мышей. [ 1 ]
История
[ редактировать ]Первая микроРНК была открыта в начале 1990-х годов. [ 16 ] Однако микроРНК не были признаны отдельным классом биологических регуляторов до начала 2000-х годов. [ 17 ] [ 18 ] [ 19 ] [ 20 ] [ 21 ] Исследование микроРНК выявило разные наборы микроРНК, экспрессируемые в разных типах клеток и тканях. [ 12 ] [ 22 ] и множественная роль микроРНК в развитии растений и животных, а также во многих других биологических процессах. [ 23 ] [ 24 ] [ 25 ] [ 26 ] [ 27 ] [ 28 ] [ 29 ] Аберрантная экспрессия микроРНК вовлечена в болезненные состояния. Терапия на основе микроРНК находится в стадии изучения. [ 30 ] [ 31 ] [ 32 ] [ 33 ]
Первая микроРНК была открыта в 1993 году группой под руководством Амброса , в которую входили Ли и Фейнбаум. Однако для дополнительного понимания способа его действия потребовалась одновременно опубликованная работа команды Рувкуна , включая Вайтмана и Ха. [ 16 ] [ 34 ] Эти группы опубликовали подряд статьи о гене lin-4 , который, как известно, контролирует время C. elegans развития личинок путем репрессии гена lin-14 . Когда Ли и др. изолировав микроРНК lin-4 , они обнаружили, что вместо производства мРНК, кодирующей белок, она производит короткие некодирующие РНК , одна из которых представляла собой ~22-нуклеотидную РНК, содержащую последовательности, частично комплементарные множественным последовательностям в 3'-UTR. мРНК lin-14 . [ 16 ] Было высказано предположение, что эта комплементарность ингибирует трансляцию мРНК lin-14 в белок LIN-14. В то время lin-4 считалось, что малая РНК является проявлением идиосинкразии нематод .
В 2000 году была охарактеризована вторая малая РНК: let-7 РНК , которая репрессирует lin-41 , способствуя более позднему переходу в развитии у C. elegans . [ 17 ] Было обнаружено, что РНК let-7 консервативна у многих видов, что привело к предположению, что РНК let-7 и дополнительные «малые временные РНК» могут регулировать сроки развития у различных животных, включая человека. [ 18 ]
Год спустя было обнаружено, что РНК lin-4 и let-7 являются частью большого класса малых РНК, присутствующих в C. elegans , Drosophila и клетках человека. [ 19 ] [ 20 ] [ 21 ] Многие РНК этого класса напоминали РНК lin-4 и let-7 , за исключением того, что характер их экспрессии обычно не соответствовал роли в регуляции сроков развития. Это предполагает, что большинство из них могут функционировать в других типах регуляторных путей. В этот момент исследователи начали использовать термин «микроРНК» для обозначения этого класса малых регуляторных РНК. [ 19 ] [ 20 ] [ 21 ]
Первым заболеванием человека, связанным с нарушением регуляции микроРНК, был хронический лимфоцитарный лейкоз . При этом заболевании микроРНК играют двойную роль, выступая как супрессорами опухолей, так и онкогенами. [ 35 ]
Номенклатура
[ редактировать ]В соответствии со стандартной номенклатурной системой экспериментально подтвержденным микроРНК перед публикацией присваиваются имена. [ 36 ] [ 37 ] За префиксом «миР» следует тире и цифра, последняя часто указывает порядок именования. Например, миР-124 была названа и, вероятно, открыта раньше миР-456. Написанная с заглавной буквы «миР-» относится к зрелой форме микроРНК, тогда как написанная с заглавной буквы «мир-» относится к пре-микроРНК и при-миРНК. [ 38 ] Гены, кодирующие микроРНК, также называются с использованием того же трехбуквенного префикса в соответствии с правилами номенклатуры генов организма. Например, официальные названия генов микроРНК у некоторых организмов: mir-1 у C. elegans и Drosophila, Mir1 у Rattus norvegicus и MIR25 у человека.
микроРНК с почти идентичными последовательностями, за исключением одного или двух нуклеотидов, помечены дополнительной строчной буквой. Например, миР-124a тесно связана с миР-124b. Например:
- hsa-миР-181а : аааукаACgcugucggugAgu
- hsa-миР-181b : аааукаУУгкугуггуггуГгу
Пре-микроРНК, при-миРНК и гены, которые приводят к образованию 100% идентичных зрелых микроРНК, но которые расположены в разных местах генома, обозначаются дополнительным суффиксом в виде тире. Например, пре-миРНК hsa-mir-194-1 и hsa-mir-194-2 приводят к идентичной зрелой микроРНК (hsa-miR-194), но происходят из генов, расположенных в разных областях генома.
Виды происхождения обозначаются трехбуквенным префиксом, например, hsa-miR-124 представляет собой микроРНК человека ( Homo sapiens ), а oar-miR-124 представляет собой микроРНК овцы ( Ovis aries ). Другие распространенные префиксы включают «v» для вирусного (миРНК, кодируемая вирусным геномом) и «d» для микроРНК дрозофилы (дрозофилы, обычно изучаемой в генетических исследованиях).
Когда две зрелые микроРНК происходят из противоположных плеч одной и той же пре-миРНК и обнаруживаются примерно в одинаковых количествах, они обозначаются суффиксом -3p или -5p. (Раньше это различие также проводилось с помощью «s» ( смысл ) и «as» (антисмысл)). Однако зрелой микроРНК, обнаруженной на одном плече шпильки, обычно гораздо больше, чем на другом плече. [ 2 ] в этом случае звездочка после названия указывает на зрелые виды, обнаруженные на низких уровнях на противоположном плече шпильки. Например, миР-124 и миР-124* имеют общую шпильку пре-миРНК, но в клетке обнаруживается гораздо больше миР-124.
Цели
[ редактировать ]Растительные микроРНК обычно имеют почти идеальное спаривание со своими мРНК-мишенями, что вызывает репрессию генов за счет расщепления целевых транскриптов. [ 23 ] [ 39 ] Напротив, микроРНК животных способны распознавать свои целевые мРНК, используя всего лишь 6–8 нуклеотидов (затравочная область) на 5'-конце микроРНК. [ 13 ] [ 40 ] [ 41 ] которого недостаточно спаривания, чтобы вызвать расщепление целевых мРНК. [ 4 ] Комбинаторная регуляция является особенностью регуляции микроРНК у животных. [ 4 ] [ 42 ] Данная микроРНК может иметь сотни различных мишеней мРНК, и данная мишень может регулироваться несколькими микроРНК. [ 14 ] [ 43 ]
Оценки среднего количества уникальных информационных РНК, которые являются мишенями для репрессии типичной микроРНК, варьируются в зависимости от метода оценки. [ 44 ] но многочисленные подходы показывают, что микроРНК млекопитающих могут иметь множество уникальных мишеней. Например, анализ высококонсервативных микроРНК у позвоночных показывает, что каждая из них имеет в среднем примерно 400 консервативных мишеней. [ 14 ] Аналогичным образом эксперименты показывают, что один вид микроРНК может снизить стабильность сотен уникальных информационных РНК. [ 45 ] Другие эксперименты показывают, что один вид микроРНК может подавлять продукцию сотен белков, но эта репрессия часто относительно мягкая (менее чем в 2 раза). [ 46 ] [ 47 ]
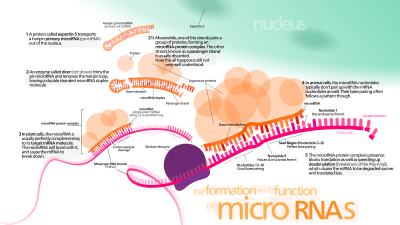
Биогенез
[ редактировать ]
До 40% генов микроРНК могут находиться в интронах или даже экзонах других генов. [ 48 ] Обычно, хотя и не исключительно, они обнаруживаются в чувственной ориентации, [ 49 ] [ 50 ] и, таким образом, обычно регулируются вместе с генами хозяина. [ 48 ] [ 51 ] [ 52 ]
Матрица ДНК не является последним словом в производстве зрелых микроРНК: 6% человеческих микроРНК демонстрируют редактирование РНК ( IsomiRs ), сайт-специфическую модификацию последовательностей РНК с получением продуктов, отличных от тех, которые кодируются их ДНК. Это увеличивает разнообразие и масштаб действия микроРНК за пределы того, что подразумевается только в геноме.
Транскрипция
[ редактировать ]Гены микроРНК обычно транскрибируются РНК-полимеразой II (Pol II). [ 53 ] [ 54 ] Полимераза часто связывается с промотором, обнаруженным рядом с последовательностью ДНК, и кодирует то, что станет шпилькой пре-миРНК. Полученный транскрипт кэпирован специально модифицированным нуклеотидом на 5'-конце, полиаденилированным множеством аденозинов (поли(А)-хвост). [ 53 ] [ 49 ] и сращены . МикроРНК животных первоначально транскрибируются как часть одного плеча стволовой петли РНК из ~80 нуклеотидов , которая, в свою очередь, образует часть предшественника микроРНК длиной в несколько сотен нуклеотидов, называемого при-миРНК. [ 53 ] [ 49 ] Когда предшественник «стебель-петля» обнаруживается в 3'-UTR, транскрипт может служить в качестве при-миРНК и мРНК. [ 49 ] РНК-полимераза III (Pol III) транскрибирует некоторые микроРНК, особенно те, которые имеют расположенные выше последовательности Alu , транспортные РНК (тРНК) и с широкими вкраплениями повторов млекопитающих (MWIR). промоторные единицы [ 55 ]
Ядерная обработка
[ редактировать ]
Одна при-миРНК может содержать от одного до шести предшественников микроРНК. Эти структуры шпилек-петлей состоят примерно из 70 нуклеотидов каждая. Каждая шпилька окружена последовательностями, необходимыми для эффективной обработки.
Структура двухцепочечной РНК (дсРНК) шпилек в при-микроРНК распознается ядерным белком, известным как критическая область 8 синдрома ДиДжорджа (DGCR8 или «Паша» у беспозвоночных), названная в честь его связи с синдромом ДиДжорджа . DGCR8 связывается с ферментом Дроша , белком, который разрезает РНК, образуя микропроцессорный комплекс . [ 56 ] [ 57 ] В этом комплексе DGCR8 ориентирует каталитический домен РНКазы III Drosha на высвобождение шпилек из pri-миРНК путем отщепления РНК примерно на одиннадцать нуклеотидов от основания шпильки (одна спиральная дцРНК превращается в стебель). [ 58 ] [ 59 ] Полученный продукт имеет выступающий конец из двух нуклеотидов на своем 3'-конце; он имеет 3'-гидроксильные и 5'-фосфатные группы. Ее часто называют пре-миРНК (предшественник-миРНК). Были идентифицированы мотивы последовательности, расположенные ниже пре-миРНК, которые важны для эффективного процессинга. [ 60 ] [ 61 ] [ 62 ]
Пре-миРНК, которые сплайсируются непосредственно из интронов, минуя микропроцессорный комплекс, известны как « миртроны ». [ 63 ] Миртроны были обнаружены у дрозофилы , C. elegans и млекопитающих. [ 64 ] [ 65 ]
До 16% пре-миРНК могут быть изменены посредством редактирования ядерной РНК . [ 66 ] [ 67 ] [ 68 ] Чаще всего ферменты , известные как аденозиндезаминазы, действующие на РНК (ADAR), катализируют переходы аденозина в инозин (от A до I). Редактирование РНК может остановить ядерный процессинг (например, при-миР-142, что приводит к деградации рибонуклеазой Tudor-SN) и изменить последующие процессы, включая цитоплазматический процессинг миРНК и целевую специфичность (например, путем изменения затравочной области миР-376). в центральной нервной системе). [ 66 ]
Ядерный экспорт
[ редактировать ]
Шпильки пре-миРНК экспортируются из ядра в процессе, вовлекающем нуклеоцитоплазматический челночный транспортер Экспортин-5 . Этот белок, член семейства кариоферинов , распознает двухнуклеотидный выступ, оставленный ферментом РНКазы III Дроша на 3'-конце шпильки пре-миРНК. Транспорт, опосредованный экспортином-5, в цитоплазму энергозависимый, с использованием гуанозинтрифосфата (GTP), связанного с белком Ran . [ 69 ]
Цитоплазматический процессинг
[ редактировать ]В цитоплазме шпилька пре-миРНК расщепляется ферментом РНКазой III Dicer . [ 70 ] Эта эндорибонуклеаза взаимодействует с 5'- и 3'-концами шпильки. [ 71 ] и отсекает петлю, соединяющую 3'- и 5'-плечи, образуя несовершенный дуплекс микроРНК:миРНК* длиной около 22 нуклеотидов. [ 70 ] Общая длина шпильки и размер петли влияют на эффективность обработки Dicer. Несовершенная природа спаривания микроРНК:миРНК* также влияет на расщепление. [ 70 ] [ 72 ] Некоторые из G-богатых пре-миРНК потенциально могут принять структуру G-квадруплекса в качестве альтернативы канонической структуре «стебель-петля». Например, пре-микроРНК 92b человека имеет структуру G-квадруплекса , которая устойчива к расщеплению, опосредованному Dicer, в цитоплазме . [ 73 ] Хотя любая цепь дуплекса потенциально может действовать как функциональная микроРНК, только одна цепь обычно включается в РНК-индуцируемый комплекс молчания (RISC), где взаимодействуют микроРНК и ее мишень мРНК.
Хотя большинство микроРНК расположены внутри клетки, некоторые микроРНК, широко известные как циркулирующие микроРНК или внеклеточные микроРНК, также были обнаружены во внеклеточной среде, включая различные биологические жидкости и среды для культивирования клеток. [ 74 ] [ 75 ]
Биогенез у растений
[ редактировать ]Биогенез микроРНК у растений отличается от биогенеза животных главным образом этапами ядерного процессинга и экспорта. Вместо расщепления двумя разными ферментами, один раз внутри и один раз снаружи ядра, оба расщепления микроРНК растения выполняются гомологом Dicer, называемым Dicer-like1 (DL1). DL1 экспрессируется только в ядре растительных клеток, что указывает на то, что обе реакции протекают внутри ядра. Прежде чем дуплексы микроРНК:миРНК* растений будут транспортированы из ядра, их 3'-концы метилируются РНК-метилтрансферазным белком, называемым Hua-Enhancer1 (HEN1). Затем дуплекс транспортируется из ядра в цитоплазму с помощью белка под названием Hasty (HST), гомолога экспортина 5, где они разбираются, и зрелая микроРНК включается в RISC. [ 76 ]
РНК-индуцированный комплекс молчания
[ редактировать ]Зрелая микроРНК является частью активного РНК-индуцированного комплекса молчания (RISC), содержащего Dicer и многие связанные белки. [ 77 ] RISC также известен как комплекс микроРНК-рибонуклеопротеина (миРНП); [ 78 ] RISC со встроенной микроРНК иногда называют «миРИСК».
Считается, что процессинг пре-миРНК Dicer связан с раскручиванием дуплекса. Обычно в miRISC включена только одна цепь, выбранная на основании ее термодинамической нестабильности и более слабого спаривания оснований на 5'-конце по сравнению с другой цепью. [ 79 ] [ 80 ] [ 81 ] Положение стебля-петли также может влиять на выбор пряди. [ 82 ] Другая цепь, называемая пассажирской цепью из-за ее более низких уровней в устойчивом состоянии, отмечена звездочкой (*) и обычно подвергается деградации. В некоторых случаях обе цепи дуплекса жизнеспособны и становятся функциональными микроРНК, нацеленными на разные популяции мРНК. [ 83 ]
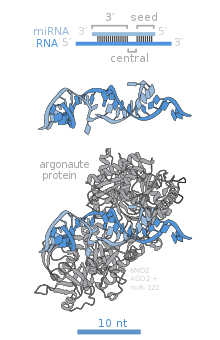
Члены семейства белков Argonaute (Ago) играют центральную роль в функции RISC. Аргонавты необходимы для молчания, индуцированного микроРНК, и содержат два консервативных РНК-связывающих домена: домен PAZ, который может связывать одноцепочечный 3'-конец зрелой микроРНК, и домен PIWI , который структурно напоминает рибонуклеазу-H и функционирует для взаимодействия с 5'-концом зрелой микроРНК. конец направляющей пряди. Они связывают зрелую микроРНК и ориентируют ее для взаимодействия с целевой мРНК. Некоторые аргонавты, например Ago2 человека, напрямую расщепляют целевые транскрипты; argonautes также могут рекрутировать дополнительные белки для достижения репрессии трансляции. [ 84 ] Геном человека кодирует восемь белков-аргонавтов, разделенных по сходству последовательностей на два семейства: AGO (четыре члена присутствуют во всех клетках млекопитающих и называются E1F2C/hAgo у человека) и PIWI (обнаружены в зародышевых и гемопоэтических стволовых клетках). [ 78 ] [ 84 ]
Дополнительные компоненты RISC включают TRBP [белок, связывающий трансактивирующую ответную РНК вируса иммунодефицита человека (TAR)]. [ 85 ] PACT (белок-активатор интерферон -индуцированной протеинкиназы ), комплекс SMN, хрупкий X-белок умственной отсталости (FMRP), белок, содержащий домен стафилококковой нуклеазы Tudor (Tudor-SN), предполагаемая ДНК- геликаза MOV10 и распознавание РНК мотив, содержащий белок TNRC6B . [ 69 ] [ 86 ] [ 87 ]
Режим глушения и регуляторные петли
[ редактировать ]Замалчивание генов может происходить либо за счет деградации мРНК, либо за счет предотвращения трансляции мРНК. Например, miR16 содержит последовательность, комплементарную AU-богатому элементу, обнаруженному в 3'UTR многих нестабильных мРНК, таких как TNF-альфа или GM-CSF . [ 88 ] Было продемонстрировано, что при полной комплементарности между микроРНК и последовательностью целевой мРНК Ago2 может расщеплять мРНК и приводить к прямой деградации мРНК. При отсутствии комплементарности замалчивание достигается за счет предотвращения трансляции. [ 45 ] Взаимосвязь микроРНК и ее целевой мРНК может быть основана на простой негативной регуляции целевой мРНК, но, по-видимому, распространенным сценарием является использование «когерентной петли прямой связи », «петли взаимной отрицательной обратной связи» (также называемой двойной отрицательный контур) и «контур положительной обратной связи/упреждающей связи». Некоторые микроРНК работают как буферы случайных изменений экспрессии генов, возникающих из-за стохастических событий в транскрипции, трансляции и стабильности белков. Такая регуляция обычно достигается за счет петель отрицательной обратной связи или некогерентной петли прямой связи, разобщающей выход белка из транскрипции мРНК.
Оборот
[ редактировать ]Обмен зрелых микроРНК необходим для быстрых изменений в профилях экспрессии микроРНК. Считается, что во время созревания микроРНК в цитоплазме поглощение белком Argonaute стабилизирует направляющую цепь, в то время как противоположная (* или «пассажирская») цепь преимущественно разрушается. В рамках так называемой стратегии «Используй или потеряй» Argonaute может преимущественно сохранять микроРНК со многими мишенями, а не микроРНК с небольшим количеством мишеней или без них, что приводит к деградации нецелевых молекул. [ 89 ]
Распад зрелых микроРНК у Caenorhabditis elegans опосредован 5'-3' экзорибонуклеазой XRN2 , также известной как Rat1p. [ 90 ] У растений члены семейства SDN (нуклеазы, расщепляющие малые РНК) расщепляют микроРНК в противоположном направлении (от 3' до 5'). Подобные ферменты кодируются в геномах животных, но их роль не описана. [ 89 ]
Несколько модификаций микроРНК влияют на стабильность микроРНК. Как показали работы на модельном организме Arabidopsis thaliana (кресс-салат), микроРНК зрелых растений, по-видимому, стабилизируются за счет добавления метильных фрагментов на 3'-конце. 2'-O-конъюгированные метильные группы блокируют добавление остатков урацила (U) ферментами уридилтрансферазами , модификация, которая может быть связана с деградацией микроРНК. Однако уридилирование может также защищать некоторые микроРНК; последствия этой модификации до конца не понятны. Сообщалось об уридилировании некоторых микроРНК животных. Как растительные, так и животные микроРНК могут быть изменены путем добавления остатков аденина (А) к 3'-концу микроРНК. Дополнительный А, добавленный к концу миР-122 млекопитающих , миРНК, обогащенной печенью, важной при гепатите С , стабилизирует молекулу, и растительные миРНК, заканчивающиеся остатком аденина, имеют более медленную скорость распада. [ 89 ]
Клеточные функции
[ редактировать ]
Функция микроРНК, по-видимому, заключается в регуляции генов. Для этой цели микроРНК комплементарна части одной или нескольких информационных РНК (мРНК). МикроРНК животных обычно комплементарны участку в 3'-UTR, тогда как микроРНК растений обычно комплементарны кодирующим областям мРНК. [ 92 ] Идеальное или почти идеальное спаривание оснований с целевой РНК способствует расщеплению РНК. [ 93 ] Это основной способ растительных микроРНК. [ 94 ] У животных совпадения несовершенны.
Чтобы частично комплементарные микроРНК могли распознавать свои мишени, нуклеотиды 2–7 микроРНК (ее «зародышевая область» [ 13 ] [ 40 ] ) должны быть идеально дополняющими друг друга. [ 95 ] МикроРНК животных ингибируют трансляцию белка целевой мРНК. [ 96 ] (это присутствует, но реже встречается у растений). [ 94 ] Частично комплементарные микроРНК также могут ускорять деаденилирование , вызывая более раннюю деградацию мРНК. [ 97 ] Хотя деградация мРНК, нацеленной на микроРНК, хорошо документирована, горячо обсуждается, осуществляется ли репрессия трансляции посредством деградации мРНК, ингибирования трансляции или их комбинации. Недавние работы с миР-430 у рыбок данио, а также с бантам-миРНК и миР-9 в культивируемых клетках дрозофилы показали, что репрессия трансляции вызвана нарушением инициации трансляции , независимо от деаденилирования мРНК. [ 98 ] [ 99 ]
микроРНК иногда также вызывают гистонов и метилирование ДНК промоторных модификацию сайтов, что влияет на экспрессию генов-мишеней. [ 100 ] [ 101 ]
Девять механизмов действия микроРНК описаны и собраны в единую математическую модель: [ 91 ]
- ингибирование инициации Cap-40S;
- ингибирование присоединения 60S рибосомальной единицы;
- Ингибирование удлинения;
- Отпадение рибосом (преждевременное прекращение);
- Котрансляционная деградация возникающего белка;
- Секвестрация в P-телах ;
- распад мРНК (дестабилизация);
- расщепление мРНК;
- Ингибирование транскрипции посредством реорганизации хроматина, опосредованной микроРНК, с последующим молчанием генов.
Разобраться в этих механизмах, используя экспериментальные данные о стационарных скоростях реакций, зачастую невозможно. Тем не менее, они дифференцированы по динамике и имеют разные кинетические характеристики . [ 91 ]
В отличие от микроРНК растений, микроРНК животных нацелены на разнообразные гены. [ 40 ] Однако гены, участвующие в функциях, общих для всех клеток, таких как экспрессия генов, имеют относительно меньше сайтов-мишеней микроРНК и, по-видимому, находятся в стадии отбора, чтобы избежать нацеливания микроРНК. [ 102 ] Существует сильная корреляция между регуляцией генов ITPR и mir-92 и mir-19. [ 103 ]
дсРНК может также активировать экспрессию генов , механизм, который получил название «активация гена, индуцированная малой РНК» или РНКа . дцРНК, нацеленные на промоторы генов, могут индуцировать мощную активацию транскрипции связанных генов. Это было продемонстрировано на клетках человека с использованием синтетических дцРНК, называемых малыми активирующими РНК (саРНК). [ 104 ] но также было продемонстрировано для эндогенных микроРНК. [ 105 ]
Считается , что взаимодействия между микроРНК и комплементарными последовательностями генов и даже псевдогенов , которые имеют гомологию последовательностей, являются обратным каналом связи, регулирующим уровни экспрессии между паралогичными генами (гены, имеющие сходную структуру, указывающую на отличие от общего предкового гена). Получив название «конкурирующие эндогенные РНК» ( ceRNA ), эти микроРНК связываются с «элементами ответа микроРНК» на генах и псевдогенах и могут дать другое объяснение существования некодирующей ДНК . [ 106 ]
микроРНК также обнаруживаются в виде внеклеточных циркулирующих микроРНК . [ 107 ] Циркулирующие микроРНК высвобождаются в жидкости организма, включая кровь и спинномозговую жидкость , и потенциально могут быть доступны в качестве биомаркеров при ряде заболеваний. [ 107 ] [ 108 ] Некоторые исследования показывают, что мРНК экзосом может играть роль в имплантации: они могут разрушать адгезию между трофобластом и эндометрием или поддерживать адгезию путем понижающей или повышающей регуляции экспрессии генов, участвующих в адгезии/инвазии. [ 109 ]
Более того, микроРНК в виде миР-183/96/182, по-видимому, играет ключевую роль в циркадном ритме . [ 110 ]
Эволюция
[ редактировать ]МикроРНК хорошо консервативны как у растений, так и у животных и считаются жизненно важным и эволюционно древним компонентом регуляции генов. [ 111 ] [ 112 ] [ 113 ] [ 114 ] [ 115 ] Хотя основные компоненты пути микроРНК консервативны у растений и животных , репертуары микроРНК в этих двух царствах, по-видимому, возникли независимо с разными первичными способами действия. [ 116 ] [ 117 ]
микроРНК являются полезными филогенетическими маркерами из-за их явно низкой скорости эволюции. [ 118 ] Происхождение микроРНК как регуляторного механизма развилось из предыдущего механизма РНКи, который первоначально использовался для защиты от экзогенного генетического материала, такого как вирусы. [ 119 ] Их происхождение, возможно, позволило развить морфологические инновации и, сделав экспрессию генов более специфичной и «тонко настраиваемой», позволило создать сложные органы. [ 120 ] и, возможно, в конечном итоге, сложная жизнь. [ 115 ] Быстрые всплески морфологических инноваций обычно связаны с высокой скоростью накопления микроРНК. [ 118 ] [ 120 ]
Новые микроРНК создаются разными способами. Новые микроРНК могут возникать в результате случайного образования шпилек в «некодирующих» участках ДНК (т.е. в интронах или межгенных областях), а также в результате дупликации и модификации существующих микроРНК. [ 121 ] МикроРНК также могут образовываться из инвертированных дупликаций белково-кодирующих последовательностей, что позволяет создавать структуру складной шпильки. [ 122 ] Скорость эволюции (т.е. замены нуклеотидов) в недавно возникших микроРНК сравнима со скоростью в других частях некодирующей ДНК, что подразумевает эволюцию за счет нейтрального дрейфа; однако более старые микроРНК имеют гораздо меньшую скорость изменений (часто менее одной замены за сто миллионов лет), [ 115 ] предполагая, что как только микроРНК обретает функцию, она подвергается очищающему отбору. [ 121 ] Отдельные области внутри гена микроРНК сталкиваются с разным эволюционным давлением, при этом области, жизненно важные для процессинга и функционирования, имеют более высокий уровень консервации. [ 123 ] На этом этапе микроРНК редко теряется из генома животного. [ 115 ] хотя более новые микроРНК (таким образом, предположительно нефункциональные) часто теряются. [ 121 ] По прогнозам, у Arabidopsis thaliana чистый поток генов микроРНК составит от 1,2 до 3,3 генов на миллион лет. [ 124 ] Это делает их ценным филогенетическим маркером, и их рассматривают как возможное решение нерешенных филогенетических проблем, таких как взаимоотношения членистоногих . [ 125 ] С другой стороны, во многих случаях микроРНК плохо коррелируют с филогенией, и возможно, что их филогенетическое соответствие в значительной степени отражает ограниченную выборку микроРНК. [ 126 ]
МикроРНК присутствуют в геномах большинства эукариотических организмов, включая бурые водоросли. [ 127 ] животным. Однако разница в том, как функционируют эти микроРНК и способы их процессинга, позволяет предположить, что микроРНК возникли независимо у растений и животных. [ 128 ]
Сосредоточившись на животных, геном Mnemiopsis leidyi. [ 129 ] по-видимому, отсутствуют узнаваемые микроРНК, а также ядерные белки Drosha и Pasha , которые имеют решающее значение для канонического биогенеза микроРНК. На данный момент это единственное животное, у которого отсутствует Дроша. МикроРНК играют жизненно важную роль в регуляции экспрессии генов у всех животных, не являющихся гребневиками, исследованных до сих пор, за исключением Trichoplax adhaerens , первого известного представителя типа Placozoa . [ 130 ]
К марту 2010 года у всех видов было идентифицировано более 5000 различных микроРНК. [ 131 ] Хотя у бактерий встречаются короткие последовательности РНК (50–сотни пар оснований) с широко сопоставимыми функциями, у бактерий отсутствуют настоящие микроРНК. [ 132 ]
Экспериментальное обнаружение и манипуляции
[ редактировать ]В то время как исследователи сосредоточились на экспрессии микроРНК в физиологических и патологических процессах, появились различные технические переменные, связанные с выделением микроРНК. Стабильность хранящихся образцов микроРНК подвергается сомнению. [ 75 ] микроРНК деградируют гораздо легче, чем мРНК, отчасти из-за их длины, но также и из-за повсеместного присутствия РНКаз . Это приводит к необходимости охлаждать образцы на льду и использовать оборудование, не содержащее РНКазы . [ 133 ]
Экспрессию микроРНК можно количественно оценить с помощью двухэтапной полимеразной цепной реакции модифицированной RT-PCR с последующей количественной ПЦР . Вариации этого метода обеспечивают абсолютную или относительную количественную оценку. [ 134 ] МикроРНК также можно гибридизовать с микрочипами , предметными стеклами или чипами с зондами для сотен или тысяч мишеней микроРНК, так что относительные уровни микроРНК можно определить в различных образцах. [ 135 ] МикроРНК можно как обнаружить, так и профилировать с помощью методов высокопроизводительного секвенирования ( секвенирование микроРНК ). [ 136 ] Активность микроРНК можно экспериментально ингибировать с помощью нуклеиновой кислоты (LNA) олигозаблокированной , олигоморфолино . [ 137 ] [ 138 ] или 2'-O-метил олигоРНК. [ 139 ] Специфическая микроРНК может подавлять комплементарный антагомир . Созревание микроРНК может быть ингибировано в нескольких точках с помощью стерически блокирующих олигонуклеотидов. [ 140 ] [ 141 ] Целевой сайт микроРНК транскрипта мРНК также может быть заблокирован стерически блокирующим олигонуклеотидом. [ 142 ] Для обнаружения микроРНК «in situ» LNA [ 143 ] или Морфолино [ 144 ] можно использовать зонды. Заблокированная конформация LNA приводит к усилению свойств гибридизации и повышению чувствительности и селективности, что делает ее идеальной для обнаружения коротких микроРНК. [ 145 ]
Высокопроизводительная количественная оценка микроРНК подвержена ошибкам из-за большей дисперсии (по сравнению с мРНК ), что сопряжено с методологическими проблемами. Поэтому экспрессию мРНК часто анализируют для проверки влияния микроРНК на их уровни (например, в [ 146 ] ). Базы данных могут использоваться для сопряжения мРНК- и микроРНК-данных, которые предсказывают мишени-мишени на основе их базовой последовательности. [ 147 ] [ 148 ] Хотя это обычно делается после того, как интересующие микроРНК были обнаружены (например, из-за высоких уровней экспрессии), мРНК и микроРНК. были предложены идеи инструментов анализа, которые интегрируют информацию об экспрессии [ 149 ] [ 150 ]
Болезни человека и животных
[ редактировать ]Подобно тому, как микроРНК участвует в нормальном функционировании эукариотических клеток, нарушение регуляции микроРНК связано с заболеваниями. В общедоступной базе данных miR2Disease, созданной вручную, документируются известные взаимосвязи между нарушением регуляции микроРНК и заболеваниями человека. [ 151 ]
Наследственные заболевания
[ редактировать ]Мутация в зародышевой области миР-96 вызывает наследственную прогрессирующую потерю слуха. [ 152 ]
Мутация в семенной области миР-184 вызывает наследственный кератоконус с передней полярной катарактой. [ 153 ]
Удаление кластера miR-17~92 вызывает дефекты скелета и роста. [ 154 ]
Рак
[ редактировать ]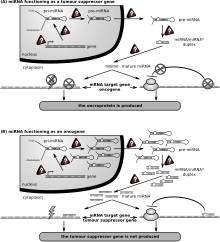
Первым заболеванием человека, которое, как известно, связано с нарушением регуляции микроРНК, был хронический лимфоцитарный лейкоз . [ 155 ] Многие другие микроРНК также связаны с раком и поэтому иногда называются « онкомирами ». [ 156 ] В злокачественных B-клетках микроРНК участвуют в путях, фундаментальных для развития B-клеток, таких как передача сигналов B-клеточного рецептора (BCR), миграция/адгезия B-клеток, межклеточные взаимодействия в иммунных нишах, а также выработка и переключение классов иммуноглобулинов. МикроРНК влияют на созревание В-клеток, образование пре-, маргинальной зоны, фолликулярных, В1-клеток, плазматических В-клеток и В-клеток памяти. [ 157 ]
Другая роль микроРНК при раке заключается в использовании уровня их экспрессии для прогноза. В НМРЛ образцах низкие уровни миР-324а могут служить индикатором плохой выживаемости. [ 158 ] Как высокие уровни миР-185, так и низкие уровни миР-133b могут коррелировать с метастазированием и плохой выживаемостью при колоректальном раке . [ 159 ]
Более того, специфические микроРНК могут быть связаны с определенными гистологическими подтипами колоректального рака. Например, было показано, что уровни экспрессии миР-205 и миР-373 повышены при муцинозном колоректальном раке и муцинпродуцирующем раке толстой кишки, связанном с язвенным колитом, но не при спорадической аденокарциноме толстой кишки, в которой отсутствуют муциновые компоненты. [ 160 ] Исследования in vitro показали, что миР-205 и миР-373 могут функционально индуцировать различные особенности муцин-ассоциированной неопластической прогрессии в эпителиальных клетках кишечника. [ 160 ]
Пролиферация клеток гепатоцеллюлярной карциномы может возникать в результате взаимодействия миР-21 с MAP2K3, геном-репрессором опухоли. [ 161 ] Оптимальное лечение рака предполагает точное определение пациентов для терапии со стратификацией риска. Те, у кого быстрый ответ на первоначальное лечение, могут получить пользу от усеченных схем лечения, что показывает ценность точных мер реагирования на заболевание. Бесклеточные циркулирующие микроРНК (цимиРНК) очень стабильны в крови, сверхэкспрессируются при раке и поддаются количественному определению в диагностической лаборатории. При классической лимфоме Ходжкина миР-21, миР-494 и миР-1973 плазмы являются многообещающими биомаркерами ответа на заболевание. [ 162 ] Циркулирующие микроРНК могут помочь в принятии клинических решений и интерпретации данных позитронно-эмиссионной томографии в сочетании с компьютерной томографией . Их можно проводить на каждой консультации для оценки реакции заболевания и выявления рецидива.
МикроРНК потенциально могут быть использованы в качестве инструментов или мишеней для лечения различных видов рака. [ 163 ] В нескольких исследованиях было обнаружено, что специфическая микроРНК, miR-506, действует как антагонист опухоли. Было обнаружено, что в значительном количестве образцов рака шейки матки экспрессия миР-506 снижена. Кроме того, миР-506 способствует апоптозу клеток рака шейки матки посредством своего прямого транскрипционного фактора пути hedgehog, Gli3. [ 164 ] [ 165 ]
Восстановление ДНК и рак
[ редактировать ]Многие микроРНК могут напрямую нацеливаться на гены клеточного цикла и ингибировать их , контролируя пролиферацию клеток . Новая стратегия лечения опухолей заключается в ингибировании пролиферации опухолевых клеток путем восстановления дефектного пути микроРНК в опухолях. [ 166 ] Рак вызывается накоплением мутаций в результате повреждения ДНК или неисправленных ошибок репликации ДНК . [ 167 ] Дефекты репарации ДНК вызывают накопление мутаций, которые могут привести к раку. [ 168 ] Некоторые гены, участвующие в репарации ДНК, регулируются микроРНК. [ 169 ]
Зародышевые мутации в генах репарации ДНК вызывают только 2–5% случаев рака толстой кишки . [ 170 ] Однако измененная экспрессия микроРНК, вызывающая нарушения репарации ДНК, часто связана с раком и может быть важным причинным фактором. Среди 68 спорадических случаев рака толстой кишки со сниженной экспрессией репарации несоответствия ДНК белка MLH1 у большинства был обнаружен дефицит из-за эпигенетического метилирования CpG - островка гена MLH1 . [ 171 ] Однако до 15% дефицитов MLH1 при спорадическом раке толстой кишки, по-видимому, обусловлены сверхэкспрессией микроРНК миР-155, которая подавляет экспрессию MLH1. [ 172 ]
В 29–66% [ 173 ] [ 174 ] В глиобластомах репарация ДНК недостаточна из-за эпигенетического метилирования гена MGMT , что снижает экспрессию белка MGMT. Однако у 28% глиобластом белок MGMT дефицитен, но промотор MGMT не метилирован. [ 173 ] В глиобластомах без метилированных промоторов MGMT уровень микроРНК miR-181d обратно коррелирует с экспрессией белка MGMT, а прямой мишенью miR-181d является 3'UTR мРНК MGMT ( три основных нетранслируемых участка мРНК MGMT). [ 173 ] Таким образом, в 28% глиобластом причинным фактором может быть повышенная экспрессия миР-181d и сниженная экспрессия фермента репарации ДНК MGMT.
Белки HMGA (HMGA1a, HMGA1b и HMGA2) участвуют в развитии рака, и экспрессия этих белков регулируется микроРНК. Экспрессия HMGA практически не обнаруживается в дифференцированных тканях взрослых, но повышена при многих видах рака. Белки HMGA представляют собой полипептиды из ~100 аминокислотных остатков, характеризующиеся модульной организацией последовательностей. Эти белки имеют три сильно положительно заряженных участка, называемых АТ-крючками , которые связывают малую бороздку богатых АТ участков ДНК в определенных участках ДНК. Неоплазии человека, включая рак щитовидной железы, предстательной железы, шейки матки, колоректальный рак, поджелудочной железы и яичников, демонстрируют значительное увеличение белков HMGA1a и HMGA1b. [ 175 ] У трансгенных мышей с HMGA1, нацеленным на лимфоидные клетки, развивается агрессивная лимфома, что показывает, что высокая экспрессия HMGA1 связана с раком и что HMGA1 может действовать как онкоген. [ 176 ] Белок HMGA2 специфически нацелен на промотор ERCC1 , тем самым снижая экспрессию этого гена репарации ДНК. [ 177 ] Экспрессия белка ERCC1 была недостаточной в 100% из 47 исследованных случаев рака толстой кишки (хотя степень участия HGMA2 неизвестна). [ 178 ]
Полиморфизмы одиночных нуклеотидов (SNP) могут изменять связывание микроРНК с 3'UTR, например, в случае hsa-mir181a и hsa-mir181b с геном-супрессором опухолей CDON. [ 179 ]
Сердечное заболевание
[ редактировать ]Глобальная роль функции микроРНК в сердце была решена путем условного ингибирования созревания микроРНК в сердце мышей . Это показало, что микроРНК играют важную роль в его развитии. [ 180 ] [ 181 ] Исследования профиля экспрессии микроРНК показывают, что уровни экспрессии специфических микроРНК изменяются при заболеваниях сердца человека, что указывает на их участие в кардиомиопатиях . [ 182 ] [ 183 ] [ 184 ] Более того, исследования специфических микроРНК на животных выявили различные роли микроРНК как во время развития сердца, так и при патологических состояниях, включая регуляцию ключевых факторов, важных для кардиогенеза, реакции гипертрофического роста и сердечной проводимости. [ 181 ] [ 185 ] [ 186 ] [ 187 ] [ 188 ] [ 189 ] Другая роль микроРНК при сердечно-сосудистых заболеваниях заключается в использовании уровней их экспрессии для диагностики, прогноза или стратификации риска. [ 190 ] МикроРНК на животных моделях также были связаны с метаболизмом и регуляцией холестерина.
миРНК-712
[ редактировать ]Мышиная микроРНК-712 является потенциальным биомаркером (т.е. предиктором) атеросклероза , сердечно-сосудистого заболевания артериальной стенки, связанного с задержкой липидов и воспалением. [ 191 ] Неламинарный кровоток также коррелирует с развитием атеросклероза, поскольку механосеноры эндотелиальных клеток реагируют на силу сдвига нарушенного кровотока (d-поток). [ 192 ] Ряд проатерогенных генов, включая матриксные металлопротеиназы (ММП), активируются d-потоком, [ 192 ] опосредуя провоспалительные и проангиогенные сигналы. Эти результаты наблюдались на перевязанных сонных артериях мышей, чтобы имитировать эффекты d-потока. В течение 24 часов ранее существовавшая незрелая миР-712 образовала зрелую миР-712, что позволяет предположить, что миР-712 чувствительна к потоку. [ 192 ] В соответствии с этими результатами, миР-712 также активируется в эндотелиальных клетках, подвергающихся естественному d-потоку в большой кривизне дуги аорты. [ 192 ]
Источник
[ редактировать ]Последовательность пре-мРНК миР-712 генерируется из мышиного рибосомального гена RN45s во внутренней транскрибируемой спейсерной области 2 (ITS2). [ 192 ] XRN1 представляет собой экзонуклеазу, которая разрушает область ITS2 во время процессинга RN45. [ 192 ] Таким образом, снижение XRN1 в условиях d-потока приводит к накоплению миР-712. [ 192 ]
Механизм
[ редактировать ]МиР-712 нацелена на тканевой ингибитор металлопротеиназ 3 (TIMP3). [ 192 ] ТИМП обычно регулируют активность матриксных металлопротеиназ (ММП), которые разрушают внеклеточный матрикс (ECM). Артериальный ЕСМ в основном состоит из коллагеновых и эластиновых волокон, обеспечивающих структурную поддержку и противооткатные свойства артерий. [ 193 ] Эти волокна играют решающую роль в регуляции воспаления и проницаемости сосудов, которые важны для развития атеросклероза. [ 194 ] TIMP3, экспрессируемый эндотелиальными клетками, является единственным TIMP, связанным с ЕСМ. [ 193 ] Снижение экспрессии TIMP3 приводит к усилению деградации ЕСМ в присутствии d-потока. В соответствии с этими данными ингибирование пре-миР712 увеличивает экспрессию TIMP3 в клетках даже при воздействии турбулентного потока. [ 192 ]
TIMP3 также снижает экспрессию TNFα (провоспалительного регулятора) во время турбулентного потока. [ 192 ] Активность TNFα в турбулентном потоке измеряли по экспрессии TNFα-превращающего фермента (TACE) в крови. TNFα снижался, если миР-712 была ингибирована или сверхэкспрессия TIMP3, [ 192 ] предполагая, что миР-712 и TIMP3 регулируют активность TACE в условиях турбулентного потока.
Анти-миР-712 эффективно подавляет экспрессию миР-712, индуцированную d-потоком, и увеличивает экспрессию TIMP3. [ 192 ] Анти-миР-712 также ингибирует гиперпроницаемость сосудов, тем самым значительно снижая развитие атеросклероза и инфильтрацию иммунных клеток. [ 192 ]
Человеческий гомолог микроРНК-205
[ редактировать ]Человеческий гомолог миР-712 был обнаружен в гене гомолога RN45s, который поддерживает сходные с мышами миРНК. [ 192 ] МиР-205 человека имеет схожие последовательности с миР-712 мышей и консервативна у большинства позвоночных. [ 192 ] МиР-205 и миР-712 также разделяют более 50% мишеней клеточной сигнализации, включая TIMP3. [ 192 ]
При тестировании d-поток снижал экспрессию XRN1 у людей, как и в эндотелиальных клетках мышей, что указывает на потенциально общую роль XRN1 у людей. [ 192 ]
Болезнь почек
[ редактировать ]Направленная делеция Dicer в почечных клетках-предшественниках, происходящих из FoxD1 , на мышиной модели приводила к сложному фенотипу почек, включая увеличение количества предшественников нефронов , меньшее количество клеток ренина , гладкомышечных артериол , прогрессирующую мезангиальную потерю и клубочковые аневризмы. [ 195 ] Высокопроизводительное профилирование всего транскриптома на мышиной модели с нокаутом FoxD1-Dicer выявило эктопическую активацию проапоптотического гена Bcl2L11 (Bim) и нарушение регуляции пути p53 с увеличением эффекторных генов p53, включая Bax , Trp53inp1 , Jun, Cdkn1a , Mmp2 и Арид3а . Уровни белка p53 остались неизменными, что позволяет предположить, что стромальные микроРНК FoxD1 непосредственно репрессируют эффекторные гены p53. Используя подход отслеживания клонов с последующей флуоресцентно-активируемой сортировкой клеток , профилирование микроРНК клеток, происходящих из FoxD1, не только всесторонне определило транскрипционный ландшафт микроРНК, которые имеют решающее значение для развития сосудов, но также идентифицировало ключевые микроРНК, которые, вероятно, будут модулировать почечный фенотип. в его отсутствие. Эти микроРНК включают микроРНК-10a, 18a, 19b, 24, 30c, 92a, 106a, 130a, 152, 181a, 214, 222, 302a, 370 и 381, которые регулируют Bcl2L11 (Bim), а также микроРНК-15b, 18a, 21, 30c, 92a, 106a, 125b-5p, 145, 214, 222, 296-5p и 302a, которые регулируют эффекторные гены p53. В соответствии с результатами профилирования, внематочная апоптоз наблюдался в клеточных производных линии предшественников FoxD1 и подтверждает важность почечных стромальных микроРНК в клеточном гомеостазе. [ 195 ]
Нервная система
[ редактировать ]МикроРНК имеют решающее значение для здорового развития и функционирования нервной системы . [ 196 ] Предыдущие исследования показали, что микроРНК могут регулировать дифференцировку и созревание нейронов на различных стадиях. [ 197 ] МикроРНК также играют важную роль в развитии синапсов. [ 198 ] (например, дендритогенез или морфогенез шипов) и синаптическая пластичность [ 199 ] (способствует обучению и памяти). Устранение образования микроРНК у мышей путем экспериментального подавления активности Dicer привело к патологическим результатам, таким как уменьшение размера нейронов, двигательные нарушения (при подавлении активности полосатого тела) . нейронов [ 200 ] ) и нейродегенерация (когда замалчиваются переднего мозга нейроны [ 201 ] ). Измененная экспрессия микроРНК была обнаружена при нейродегенеративных заболеваниях (таких как болезнь Альцгеймера , болезнь Паркинсона и болезнь Хантингтона). [ 202 ] ), а также многие психические расстройства (в том числе эпилепсия , [ 203 ] шизофрения , большая депрессия , биполярное расстройство и тревожные расстройства. [ 204 ] [ 205 ] [ 206 ] ).
Гладить
[ редактировать ]По данным Центра по контролю и профилактике заболеваний, инсульт является одной из основных причин смертности и долгосрочной инвалидности в Америке. 87% случаев — это ишемические инсульты , возникающие в результате закупорки артерии головного мозга, несущей богатую кислородом кровь. Препятствие кровотоку означает, что мозг не может получать необходимые питательные вещества, такие как кислород и глюкоза, и удалять отходы, такие как углекислый газ. [ 207 ] [ 208 ] miRNAs играют роль в посттрансляционном молчании генов путем нацеливания на гены в патогенезе церебральной ишемии, такие как воспалительный, ангиогенез и апоптотический путь. [ 209 ]
Алкоголизм
[ редактировать ]Жизненно важная роль микроРНК в экспрессии генов важна для развития зависимости , особенно алкоголизма . [ 210 ] Хроническое злоупотребление алкоголем приводит к стойким изменениям в функциях мозга, частично опосредованным изменениями в экспрессии генов . [ 210 ] Глобальная регуляция микроРНК многих нижестоящих генов считается важной в отношении реорганизации синаптических связей или долгосрочных нейронных адаптаций, включающих изменение поведения от потребления алкоголя к абстиненции и/или зависимости. [ 211 ] Было обнаружено, что до 35 различных микроРНК были изменены в посмертном мозге алкоголика, все из которых нацелены на гены, которые включают в себя регуляцию клеточного цикла , апоптоз , клеточную адгезию , развитие нервной системы и передачу сигналов в клетках . [ 210 ] Измененные уровни микроРНК были обнаружены в медиальной префронтальной коре мышей, зависимых от алкоголя, что позволяет предположить роль микроРНК в организации трансляционного дисбаланса и создании дифференциально экспрессируемых белков в области мозга, где, вероятно, возникают сложные когнитивные действия и принятие решений . [ 212 ]
Уровень микроРНК может повышаться или понижаться в ответ на хроническое употребление алкоголя. Экспрессия миР-206 увеличилась в префронтальной коре алкогользависимых крыс, воздействуя на фактор транскрипции нейротрофического фактора головного мозга ( BDNF ) и в конечном итоге снижая его экспрессию. BDNF играет решающую роль в формировании и созревании новых нейронов и синапсов, что предполагает возможное участие в росте синапсов/ синаптической пластичности у лиц, злоупотребляющих алкоголем. [ 213 ] , вызванных алкоголем нейровоспаления Было обнаружено, что миР-155, важная в регуляции реакций , активируется, что указывает на роль микроглии и воспалительных цитокинов в патофизиологии алкоголя. [ 214 ] Понижение уровня миР-382 было обнаружено в прилежащем ядре , структуре в базальном отделе переднего мозга, играющей важную роль в регуляции чувства вознаграждения , которое влияет на мотивационные привычки. миР-382 является мишенью для дофаминового рецептора D1 (DRD1), и ее сверхэкспрессия приводит к усилению регуляции DRD1 и дельта- fosB , фактора транскрипции, который активирует серию транскрипционных событий в прилежащем ядре , что в конечном итоге приводит к аддиктивному поведению. [ 215 ] Альтернативно, сверхэкспрессия миР-382 приводила к ослаблению употребления алкоголя и ингибированию повышения регуляции DRD1 и дельта -fosB на крысиных моделях алкоголизма, демонстрируя возможность использования фармацевтических препаратов, нацеленных на миРНК, в лечении. [ 215 ]
Ожирение
[ редактировать ]miRNAs играют решающую роль в регуляции предшественников стволовых клеток, дифференцирующихся в адипоциты . [ 216 ] Исследования по определению роли плюрипотентных стволовых клеток в адипогенезе были проведены на иммортализованной костного мозга человека. полученной из линии стромальных клеток hMSC-Tert20, [ 217 ] Снижение экспрессии миР-155 , миР-221 и миР-222 было обнаружено во время адипогенного программирования как иммортализованных, так и первичных hMSCs, что позволяет предположить, что они действуют как негативные регуляторы дифференцировки. Напротив, эктопическая экспрессия miRNAs 155 , 221 и 222 значительно ингибирует адипогенез и подавляет индукцию главных регуляторов PPARγ и CCAAT/энхансер-связывающего белка альфа ( CEBPA ). [ 218 ] Это открывает путь к возможному лечению генетического ожирения.
Другой класс микроРНК, которые регулируют резистентность к инсулину , ожирение и диабет , — это семейство let-7 . Let-7 накапливается в тканях человека в процессе старения . [ 219 ] Когда let-7 подвергался эктопической сверхэкспрессии для имитации ускоренного старения, мыши становились инсулинорезистентными и, таким образом, более склонными к ожирению и диабету, вызванным диетой с высоким содержанием жиров . [ 220 ] Напротив, когда let-7 ингибировался инъекциями антагомиров , специфичных для let-7 , мыши становились более чувствительными к инсулину и удивительно устойчивыми к ожирению и диабету, вызванным диетой с высоким содержанием жиров. Ингибирование let-7 может не только предотвратить ожирение и диабет, но также обратить вспять и вылечить это заболевание. [ 221 ] Эти экспериментальные результаты позволяют предположить, что ингибирование let-7 может представлять собой новую терапию ожирения и диабета 2 типа.
Гемостаз
[ редактировать ]микроРНК также играют решающую роль в регуляции сложных ферментативных каскадов, включая систему гемостатического свертывания крови. [ 222 ] Крупномасштабные исследования функционального нацеливания микроРНК недавно открыли обоснованные терапевтические цели в системе гемостаза. [ 223 ] [ 224 ] Они напрямую связаны с гомеостазом кальция в эндоплазматическом ретикулуме, который имеет решающее значение для дифференцировки клеток на раннем этапе развития. [ 225 ]
Растения
[ редактировать ]МикроРНК считаются ключевыми регуляторами многих процессов развития, гомеостатических и иммунных процессов у растений. [ 226 ] Их роль в развитии растений включает развитие апикальной меристемы побега, рост листьев, образование цветов, образование семян или расширение корней. [ 227 ] [ 228 ] [ 229 ] [ 230 ] Кроме того, они играют сложную роль в ответах на различные абиотические стрессы, включая тепловой стресс, низкотемпературный стресс, стресс засухи, световой стресс или воздействие гамма-излучения. [ 226 ]
Вирусы
[ редактировать ]Вирусные микроРНК играют важную роль в регуляции экспрессии генов вируса и/или генов хозяина, принося пользу вирусу. Следовательно, микроРНК играют ключевую роль во взаимодействии хозяина с вирусом и патогенезе вирусных заболеваний. [ 231 ] [ 232 ] Считается, что экспрессия активаторов транскрипции ДНК вируса герпеса-6 человека регулируется вирусной микроРНК. [ 233 ]
Целевой прогноз
[ редактировать ]МикроРНК могут связываться с транскриптами целевой информационной РНК (мРНК) генов, кодирующих белок, и отрицательно контролировать их трансляцию или вызывать деградацию мРНК. Крайне важно точно идентифицировать мишени микроРНК. [ 234 ] сравнение прогнозной производительности восемнадцати алгоритмов in silico . Доступно [ 235 ] Крупномасштабные исследования нацеливания функциональных микроРНК предполагают, что многие функциональные микроРНК могут быть пропущены алгоритмами прогнозирования мишени. [ 223 ]
См. также
[ редактировать ]- Олигонуклеотиды анти-миРНК
- Экспрессия генов
- Список инструментов прогнозирования генов микроРНК
- Список инструментов прогнозирования целей микроРНК
- микроДНК
- Биосенсоры микроРНК
- МИРНЕСТ
- МИР222
- миР-324-5п
- РНК-интерференция
- Малая интерферирующая РНК
- Малая микроРНК, полученная из ядрышковой РНК
- Кластер микроРНК C19MC
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Бартель Д.П. (март 2018 г.). «МикроРНК многоклеточных животных» . Клетка . 173 (1): 20–51. дои : 10.1016/j.cell.2018.03.006 . ПМК 6091663 . ПМИД 29570994 .
- ^ Jump up to: а б с Бартель Д.П. (январь 2004 г.). «МикроРНК: геномика, биогенез, механизм и функции» . Клетка . 116 (2): 281–297. дои : 10.1016/S0092-8674(04)00045-5 . ПМИД 14744438 .
- ^ Куреши А., Тхакур Н., Монга И., Тхакур А., Кумар М. (1 января 2014 г.). «VIRmiRNA: комплексный ресурс экспериментально подтвержденных вирусных микроРНК и их мишеней» . База данных . 2014 : бау103. дои : 10.1093/база данных/bau103 . ПМК 4224276 . ПМИД 25380780 .
- ^ Jump up to: а б с Бартель Д.П. (январь 2009 г.). «МикроРНК: целевое распознавание и регуляторные функции» . Клетка . 136 (2): 215–33. дои : 10.1016/j.cell.2009.01.002 . ПМЦ 3794896 . ПМИД 19167326 .
- ^ Йонас С., Изаурральде Э. (июль 2015 г.). «На пути к молекулярному пониманию молчания генов, опосредованного микроРНК». Обзоры природы. Генетика . 16 (7): 421–433. дои : 10.1038/nrg3965 . ПМИД 26077373 . S2CID 24892348 .
- ^ Йонас С., Изаурральде Э. (июль 2015 г.). «На пути к молекулярному пониманию молчания генов, опосредованного микроРНК». Обзоры природы. Генетика . 16 (7): 421–433. дои : 10.1038/nrg3965 . ПМИД 26077373 . S2CID 24892348 .
- ^ Го Х, Инголия НТ, Вайсман Дж.С., Бартель Д.П. (август 2010 г.). «МикроРНК млекопитающих преимущественно снижают уровни целевой мРНК» . Природа . 466 (7308): 835–840. Бибкод : 2010Natur.466..835G . дои : 10.1038/nature09267 . hdl : 1721.1/72447 . ПМК 2990499 . ПМИД 20703300 .
- ^ МикроРНК Homo sapiens в miRBase в Манчестерском университете
- ^ Аллес Дж., Фельманн Т., Фишер У., Бакес С., Галата В., Минет М. и др. (апрель 2019 г.). «Оценка общего количества истинных микроРНК человека» . Исследования нуклеиновых кислот . 47 (7): 3353–3364. дои : 10.1093/nar/gkz097 . ПМК 6468295 . ПМИД 30820533 .
- ^ Фромм Б., Доманска Д., Хойе Е., Овчинников В., Канг В., Апарисио-Пуэрта Е. и др. (январь 2020 г.). «MirGeneDB 2.0: комплемент микроРНК многоклеточных животных» . Исследования нуклеиновых кислот . 48 (Д1): Д132–Д141. дои : 10.1093/nar/gkz885 . ПМК 6943042 . ПМИД 31598695 .
- ^ Лим Л.П., Лау Н.К., Вайнштейн Э.Г., Абдельхаким А., Йекта С., Роудс М.В. и др. (апрель 2003 г.). «МикроРНК Caenorhabditis elegans» . Гены и развитие . 17 (8): 991–1008. дои : 10.1101/gad.1074403 . ЧВК 196042 . ПМИД 12672692 .
- ^ Jump up to: а б Лагос-Кинтана М., Раухут Р., Ялчин А., Мейер Дж., Лендекель В., Тушль Т. (апрель 2002 г.). «Идентификация тканеспецифичных микроРНК у мышей» . Современная биология . 12 (9): 735–9. Бибкод : 2002CBio...12..735L . дои : 10.1016/S0960-9822(02)00809-6 . ПМИД 12007417 .
- ^ Jump up to: а б с Льюис Б.П., Бердж CB, Бартель Д.П. (январь 2005 г.). «Консервативные пары семян, часто окруженные аденозинами, указывают на то, что тысячи человеческих генов являются мишенями микроРНК» . Клетка . 120 (1): 15–20. дои : 10.1016/j.cell.2004.12.035 . ПМИД 15652477 .
- ^ Jump up to: а б с Фридман Р.К., Фарх К.К., Бердж CB, Бартель Д.П. (январь 2009 г.). «Большинство мРНК млекопитающих являются консервативными мишенями микроРНК» . Геномные исследования . 19 (1): 92–105. дои : 10.1101/гр.082701.108 . ПМЦ 2612969 . ПМИД 18955434 .
- ^ Фромм Б., Биллипп Т., Пек Л.Е., Йохансен М., Тарвер Дж.Э., Кинг Б.Л. и др. (2015). «Единая система аннотации генов микроРНК позвоночных и эволюция микроРНК человека» . Ежегодный обзор генетики . 49 : 213–42. doi : 10.1146/annurev-genet-120213-092023 . ПМЦ 4743252 . ПМИД 26473382 .
- ^ Jump up to: а б с Ли Р.К., Фейнбаум Р.Л., Амброс В. (декабрь 1993 г.). «Гетерохронный ген lin-4 C. elegans кодирует малые РНК, антисмысловые, комплементарные lin-14» . Клетка . 75 (5): 843–54. дои : 10.1016/0092-8674(93)90529-Y . ПМИД 8252621 .
- ^ Jump up to: а б Рейнхарт Б.Дж., Слэк Ф.Дж., Бассон М., Паскинелли А.Е., Беттингер Дж.К., Ругви А.Е. и др. (февраль 2000 г.). «21-нуклеотидная РНК let-7 регулирует сроки развития Caenorhabditis elegans». Природа . 403 (6772): 901–6. Бибкод : 2000Natur.403..901R . дои : 10.1038/35002607 . ПМИД 10706289 . S2CID 4384503 .
- ^ Jump up to: а б Паскинелли А.Е., Рейнхарт Б.Дж., Слэк Ф., Мартиндейл М.К., Курода М.И., Маллер Б. и др. (ноябрь 2000 г.). «Сохранение последовательности и временной экспрессии гетерохронной регуляторной РНК let-7». Природа . 408 (6808): 86–9. Бибкод : 2000Natur.408...86P . дои : 10.1038/35040556 . ПМИД 11081512 . S2CID 4401732 .
- ^ Jump up to: а б с Лагос-Кинтана М., Раухут Р., Лендекель В., Тушль Т. (октябрь 2001 г.). «Идентификация новых генов, кодирующих малые экспрессируемые РНК». Наука . 294 (5543): 853–8. Бибкод : 2001Sci...294..853L . дои : 10.1126/science.1064921 . hdl : 11858/00-001M-0000-0012-F65F-2 . ПМИД 11679670 . S2CID 18101169 .
- ^ Jump up to: а б с Лау, Северная Каролина, Лим Л.П., Вайнштейн Э.Г., Бартель Д.П. (октябрь 2001 г.). «Обильный класс крошечных РНК, играющих вероятную регуляторную роль у Caenorhabditis elegans». Наука . 294 (5543): 858–62. Бибкод : 2001Sci...294..858L . дои : 10.1126/science.1065062 . ПМИД 11679671 . S2CID 43262684 .
- ^ Jump up to: а б с Ли Р.К., Амброс В. (октябрь 2001 г.). «Обширный класс малых РНК Caenorhabditis elegans». Наука . 294 (5543): 862–4. Бибкод : 2001Sci...294..862L . дои : 10.1126/science.1065329 . ПМИД 11679672 . S2CID 33480585 .
- ^ Винхолдс Е., Клоостерман В.П., Миска Е., Альварес-Сааведра Е., Березиков Е., де Брёйн Е. и др. (июль 2005 г.). «Экспрессия микроРНК в эмбриональном развитии рыбок данио». Наука . 309 (5732): 310–1. Бибкод : 2005Sci...309..310W . дои : 10.1126/science.1114519 . ПМИД 15919954 . S2CID 38939571 .
- ^ Jump up to: а б Джонс-Роудс М.В., Бартель Д.П., Бартель Б. (2006). «МикроРНК и их регуляторная роль в растениях». Ежегодный обзор биологии растений . 57 : 19–53. doi : 10.1146/annurev.arplant.57.032905.105218 . ПМИД 16669754 .
- ^ Бреннеке Дж., Хипфнер Д.Р., Старк А., Рассел Р.Б., Коэн С.М. (апрель 2003 г.). «Бантам кодирует микроРНК, регулируемую в процессе развития, которая контролирует пролиферацию клеток и регулирует проапоптотический ген, скрытый у дрозофилы» . Клетка . 113 (1): 25–36. дои : 10.1016/S0092-8674(03)00231-9 . ПМИД 12679032 .
- ^ Куэльяр Т.Л., Макманус М.Т. (декабрь 2005 г.). «МикроРНК и эндокринная биология» . Журнал эндокринологии . 187 (3): 327–32. дои : 10.1677/joe.1.06426 . ПМИД 16423811 .
- ^ Пой М.Н., Элиассон Л., Круцфельдт Дж., Кувадзима С., Ма Х., Макдональд П.Е. и др. (ноябрь 2004 г.). «МикроРНК, специфичная для островков поджелудочной железы, регулирует секрецию инсулина». Природа . 432 (7014): 226–30. Бибкод : 2004Natur.432..226P . дои : 10.1038/nature03076 . ПМИД 15538371 . S2CID 4415988 .
- ^ Чен Ч.З., Ли Л., Лодиш Х.Ф., Бартель Д.П. (январь 2004 г.). «МикроРНК модулируют дифференцировку гемопоэтических линий». Наука . 303 (5654): 83–6. Бибкод : 2004Sci...303...83C . дои : 10.1126/science.1091903 . hdl : 1721.1/7483 . ПМИД 14657504 . S2CID 7044929 .
- ^ Уилфред БР, Ван В.С., Нельсон П.Т. (июль 2007 г.). «Энергичные исследования микроРНК: обзор роли микроРНК в метаболизме липидов с прогнозом, что микроРНК-103/107 регулирует метаболические пути человека» . Молекулярная генетика и обмен веществ . 91 (3): 209–17. дои : 10.1016/j.ymgme.2007.03.011 . ПМК 1978064 . ПМИД 17521938 .
- ^ Харф Б.Д., Макманус М.Т., Мэнсфилд Дж.Х., Хорнштейн Э., Табин С.Дж. (август 2005 г.). «Фермент РНКаза III Dicer необходим для морфогенеза, но не для формирования паттерна конечностей позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (31): 10898–903. Бибкод : 2005PNAS..10210898H . дои : 10.1073/pnas.0504834102 . ПМК 1182454 . ПМИД 16040801 .
- ^ Транг П., Вайдхас Дж.Б., Слак Ф.Дж. (декабрь 2008 г.). «МикроРНК как потенциальные средства лечения рака» . Онкоген . 27 (Приложение 2): S52–7. дои : 10.1038/onc.2009.353 . ПМЦ 10033140 . ПМИД 19956180 .
- ^ Ли С., Фэн Ю., Кукос Г., Чжан Л. (декабрь 2009 г.). «Терапевтические стратегии микроРНК при раке человека» . Журнал AAPS . 11 (4): 747–57. дои : 10.1208/s12248-009-9145-9 . ПМК 2782079 . ПМИД 19876744 .
- ^ Фасанаро П., Греко С., Иван М., Капогросси М.К., Мартелли Ф. (январь 2010 г.). «микроРНК: новые терапевтические мишени при острых ишемических заболеваниях». Фармакология и терапия . 125 (1): 92–104. doi : 10.1016/j.pharmthera.2009.10.003 . ПМИД 19896977 .
- ^ Гидбринг П., Бадалян-Вери Дж. (август 2013 г.). «Клиническое применение микроРНК» . F1000Исследования . 2 : 136. doi : 10.12688/f1000research.2-136.v2 . ПМЦ 3917658 . ПМИД 24627783 .
- ^ Вайтман Б., Ха И, Рувкун Г. (декабрь 1993 г.). «Посттранскрипционная регуляция гетерохронного гена lin-14 с помощью lin-4 опосредует формирование временного паттерна у C. elegans» . Клетка . 75 (5): 855–62. дои : 10.1016/0092-8674(93)90530-4 . ПМИД 8252622 .
- ^ Гиза Д.Э., Калин Г.А. (2015). «МикроРНК и хронический лимфоцитарный лейкоз». МикроРНК: Рак . Достижения экспериментальной медицины и биологии. Том. 889. стр. 23–40. дои : 10.1007/978-3-319-23730-5_2 . ISBN 978-3-319-23729-9 . ПМИД 26658994 .
- ^ Амброс В., Бартель Б., Бартель Д.П., Бердж С.Б., Кэррингтон Дж.К., Чен X и др. (март 2003 г.). «Единая система аннотации микроРНК» . РНК . 9 (3): 277–9. дои : 10.1261/rna.2183803 . ПМЦ 1370393 . ПМИД 12592000 .
- ^ Гриффитс-Джонс С., Грокок Р.Дж., ван Донген С., Бейтман А., Энрайт А.Дж. (январь 2006 г.). «miRBase: последовательности микроРНК, мишени и номенклатура генов» . Исследования нуклеиновых кислот . 34 (Проблема с базой данных): D140–4. дои : 10.1093/нар/gkj112 . ПМЦ 1347474 . ПМИД 16381832 .
- ^ Райт М.В., Бруфорд Э.А. (январь 2011 г.). «Именование «мусора»: номенклатура генов небелковой РНК (нкРНК) человека» . Геномика человека . 5 (2): 90–8. дои : 10.1186/1479-7364-5-2-90 . ПМК 3051107 . ПМИД 21296742 .
- ^ Хант М., Банерджи С., Сурана П., Лю М., Фюрст Г., Матиони С. и др. (2019). «Открытие малых РНК при взаимодействии ячменя и возбудителя мучнистой росы» . БМК Геномика . 20 (1): 19–53. дои : 10.1186/s12864-019-5947-z . ПМК 6657096 . ПМИД 31345162 .
- ^ Jump up to: а б с Льюис Б.П., Ши И.Х., Джонс-Роудс М.В., Бартель Д.П., Бердж CB (декабрь 2003 г.). «Прогнозирование целей микроРНК млекопитающих» . Клетка . 115 (7): 787–98. дои : 10.1016/S0092-8674(03)01018-3 . ПМИД 14697198 .
- ^ Эльвангер, округ Колумбия, Бюттнер Ф.А., Мьюс Х.В., Штюмпфлен В. (май 2011 г.). «Достаточный минимальный набор типов семян микроРНК» . Биоинформатика . 27 (10): 1346–50. doi : 10.1093/биоинформатика/btr149 . ПМК 3087955 . ПМИД 21441577 .
- ^ Раевский Н. (июнь 2006 г.). «Предсказания целевых микроРНК у животных». Природная генетика . 38 (6с): С8–13. дои : 10.1038/ng1798 . ПМИД 16736023 . S2CID 23496396 .
- ^ Крек А., Грюн Д., Пой М.Н., Вольф Р., Розенберг Л., Эпштейн Э.Дж. и др. (май 2005 г.). «Комбинаторные предсказания целей микроРНК». Природная генетика . 37 (5): 495–500. дои : 10.1038/ng1536 . ПМИД 15806104 . S2CID 22672750 .
- ^ Томсон Д.В., Бракен С.П., Гудолл Дж.Дж. (сентябрь 2011 г.). «Экспериментальные стратегии идентификации мишеней микроРНК» . Исследования нуклеиновых кислот . 39 (16): 6845–53. дои : 10.1093/nar/gkr330 . ПМК 3167600 . ПМИД 21652644 .
- ^ Jump up to: а б Лим Л.П., Лау Н.К., Гарретт-Энгеле П., Гримсон А., Шелтер Дж.М., Касл Дж. и др. (февраль 2005 г.). «Анализ на микрочипах показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа . 433 (7027): 769–73. Бибкод : 2005Natur.433..769L . дои : 10.1038/nature03315 . ПМИД 15685193 . S2CID 4430576 .
- ^ Зельбах М., Шванхойссер Б., Тирфельдер Н., Фанг З., Ханин Р., Раевски Н. (сентябрь 2008 г.). «Широко распространенные изменения в синтезе белка, вызванные микроРНК». Природа . 455 (7209): 58–63. Бибкод : 2008Natur.455...58S . дои : 10.1038/nature07228 . ПМИД 18668040 . S2CID 4429008 .
- ^ Пэк Д., Виллен Дж., Шин С., Камарго Ф.Д., Гиги С.П., Бартель Д.П. (сентябрь 2008 г.). «Влияние микроРНК на выход белка» . Природа . 455 (7209): 64–71. Бибкод : 2008Natur.455...64B . doi : 10.1038/nature07242 (неактивен 26 апреля 2024 г.). ПМК 2745094 . ПМИД 18668037 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Jump up to: а б Родригес А., Гриффитс-Джонс С., Ашерст Дж.Л., Брэдли А. (октябрь 2004 г.). «Идентификация генов-хозяев микроРНК млекопитающих и единиц транскрипции» . Геномные исследования . 14 (10А): 1902–10. дои : 10.1101/гр.2722704 . ПМК 524413 . ПМИД 15364901 .
- ^ Jump up to: а б с д Кай X, Хагедорн CH, Каллен BR (декабрь 2004 г.). «Человеческие микроРНК производятся из кэпированных полиаденилированных транскриптов, которые также могут функционировать как мРНК» . РНК . 10 (12): 1957–66. дои : 10.1261/rna.7135204 . ПМК 1370684 . ПМИД 15525708 .
- ^ Вебер М.Дж. (январь 2005 г.). «Новые гены микроРНК человека и мыши, обнаруженные методом поиска гомологии» . Журнал ФЭБС . 272 (1): 59–73. дои : 10.1111/j.1432-1033.2004.04389.x . ПМИД 15634332 . S2CID 32923462 .
- ^ Ким Ю.К., Ким В.Н. (февраль 2007 г.). «Процессинг интронных микроРНК» . Журнал ЭМБО . 26 (3): 775–83. дои : 10.1038/sj.emboj.7601512 . ПМЦ 1794378 . ПМИД 17255951 .
- ^ Баскервиль С., Бартель Д.П. (март 2005 г.). «Профилирование микроРНК на микрочипах показывает частую коэкспрессию с соседними микроРНК и генами хозяина» . РНК . 11 (3): 241–7. дои : 10.1261/rna.7240905 . ПМК 1370713 . ПМИД 15701730 .
- ^ Jump up to: а б с Ли Ю, Ким М, Хан Дж, Ём К.Х., Ли С., Пэк Ш. и др. (октябрь 2004 г.). «Гены микроРНК транскрибируются РНК-полимеразой II» . Журнал ЭМБО . 23 (20): 4051–60. дои : 10.1038/sj.emboj.7600385 . ПМК 524334 . ПМИД 15372072 .
- ^ Чжоу X, Жуань Дж, Ван Г, Чжан В (март 2007 г.). «Характеристика и идентификация основных промоторов микроРНК у четырех модельных видов» . PLOS Вычислительная биология . 3 (3): е37. Бибкод : 2007PLSCB...3...37Z . дои : 10.1371/journal.pcbi.0030037 . ПМЦ 1817659 . ПМИД 17352530 .
- ^ Фаллер М., Го Ф (ноябрь 2008 г.). «Биогенез микроРНК: есть несколько способов снять шкуру с кошки» . Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1779 (11): 663–7. дои : 10.1016/j.bbagrm.2008.08.005 . ПМЦ 2633599 . ПМИД 18778799 .
- ^ Ли Ю, Ан С, Хан Дж, Чой Х, Ким Дж, Йим Дж и др. (сентябрь 2003 г.). «Ядерная РНКаза III Дроша инициирует процессинг микроРНК». Природа . 425 (6956): 415–9. Бибкод : 2003Natur.425..415L . дои : 10.1038/nature01957 . ПМИД 14508493 . S2CID 4421030 .
- ^ Грегори Р.И., Чендримада Т.П., Шихаттар Р. (2006). «Биогенез микроРНК: выделение и характеристика микропроцессорного комплекса». Протоколы микроРНК . Методы молекулярной биологии. Том. 342. стр. 33–47. дои : 10.1385/1-59745-123-1:33 . ISBN 978-1-59745-123-9 . ПМИД 16957365 .
- ^ Хан Дж., Ли Ю., Ём К.Х., Ким Ю.К., Джин Х., Ким В.Н. (декабрь 2004 г.). «Комплекс Дроша-DGCR8 в первичном процессинге микроРНК» . Гены и развитие . 18 (24): 3016–27. дои : 10.1101/gad.1262504 . ПМК 535913 . ПМИД 15574589 .
- ^ Хан Дж., Ли Ю., Йом К.Х., Нам Дж.В., Хо И., Ри Дж.К. и др. (июнь 2006 г.). «Молекулярные основы распознавания первичных микроРНК комплексом Дроша-DGCR8» . Клетка . 125 (5): 887–901. дои : 10.1016/j.cell.2006.03.043 . ПМИД 16751099 .
- ^ Конрад Т., Марсико А., Гере М., Ором У.А. (октябрь 2014 г.). «Активность микропроцессора контролирует дифференциальный биогенез микроРНК в Vivo» . Отчеты по ячейкам . 9 (2): 542–54. дои : 10.1016/j.celrep.2014.09.007 . ПМИД 25310978 .
- ^ Ауён В.К., Улицкий И., Макгири С.Э., Бартель Д.П. (февраль 2013 г.). «Помимо вторичной структуры: детерминанты первичной последовательности лицензируют шпильки при-миРНК для обработки» . Клетка . 152 (4): 844–58. дои : 10.1016/j.cell.2013.01.031 . ПМК 3707628 . ПМИД 23415231 .
- ^ Али П.С., Гошдастидер У., Хоффманн Дж., Брутши Б., Филипек С. (ноябрь 2012 г.). «Распознавание предшественника микроРНК let-7g человеческим Lin28B» . Письма ФЭБС . 586 (22): 3986–90. дои : 10.1016/j.febslet.2012.09.034 . ПМИД 23063642 . S2CID 28899778 .
- ^ Руби Дж.Г., Ян CH, Бартель Д.П. (июль 2007 г.). «Предшественники интронных микроРНК, обходящие процессинг Дроши» . Природа . 448 (7149): 83–86. Бибкод : 2007Natur.448...83R . дои : 10.1038/nature05983 . ПМЦ 2475599 . ПМИД 17589500 .
- ^ Руби Дж.Г., Ян CH, Бартель Д.П. (июль 2007 г.). «Предшественники интронных микроРНК, обходящие процессинг Дроши» . Природа . 448 (7149): 83–86. Бибкод : 2007Natur.448...83R . дои : 10.1038/nature05983 . ПМЦ 2475599 . ПМИД 17589500 .
- ^ Березиков Э., Чанг В.Дж., Уиллис Дж., Куппен Э., Лай Э.К. (октябрь 2007 г.). «Миртронные гены млекопитающих» . Молекулярная клетка . 28 (2): 328–336. doi : 10.1016/j.molcel.2007.09.028 . ПМЦ 2763384 . ПМИД 17964270 .
- ^ Jump up to: а б Кавахара Ю., Меграу М., Крайдер Э., Иисаса Х., Валенте Л., Хацигеоргиу А.Г. и др. (сентябрь 2008 г.). «Частота и судьба редактирования микроРНК в мозге человека» . Исследования нуклеиновых кислот . 36 (16): 5270–80. дои : 10.1093/нар/gkn479 . ПМЦ 2532740 . ПМИД 18684997 .
- ^ Винтер Дж., Юнг С., Келлер С., Грегори Р.И., Дидерихс С. (март 2009 г.). «Многие пути к зрелости: пути биогенеза микроРНК и их регуляция». Природная клеточная биология . 11 (3): 228–34. дои : 10.1038/ncb0309-228 . ПМИД 19255566 . S2CID 205286318 .
- ^ Оман М. (октябрь 2007 г.). «Претендент или союзник процесса редактирования микроРНК». Биохимия . 89 (10): 1171–6. дои : 10.1016/j.biochi.2007.06.002 . ПМИД 17628290 .
- ^ Jump up to: а б Мерчисон Э.П. , Хэннон Дж.Дж. (июнь 2004 г.). «МиРНК в движении: биогенез микроРНК и механизм РНКи». Современное мнение в области клеточной биологии . 16 (3): 223–9. дои : 10.1016/j.ceb.2004.04.003 . ПМИД 15145345 .

- ^ Jump up to: а б с Лунд Э., Дальберг Дж. Э. (2006). «Субстратная селективность экспортина 5 и Dicer в биогенезе микроРНК» . Симпозиумы Колд-Спринг-Харбор по количественной биологии . 71 : 59–66. дои : 10.1101/sqb.2006.71.050 . ПМИД 17381281 .
- ^ Пак Дж.Э., Хо И., Тянь Ю., Симаншу Д.К., Чанг Х., Джи Д. и др. (июль 2011 г.). «Dicer распознает 5'-конец РНК для эффективной и точной обработки» . Природа . 475 (7355): 201–5. дои : 10.1038/nature10198 . ПМЦ 4693635 . ПМИД 21753850 .
- ^ Джи Икс (2008). «Механизм действия РНКазы III: как Dicer Dices». РНК-интерференция . Актуальные темы микробиологии и иммунологии. Том. 320. стр. 99–116. дои : 10.1007/978-3-540-75157-1_5 . ISBN 978-3-540-75156-4 . ПМИД 18268841 .
- ^ Мирихана Араччилаге Дж., Дассанаяке AC, Басу С. (февраль 2015 г.). «Зависимый от ионов калия структурный переключатель РНК регулирует созревание пре-микроРНК 92b человека» . Химия и биология . 22 (2): 262–72. doi : 10.1016/j.chembiol.2014.12.013 . ПМИД 25641166 .
- ^ Сохель М.Х. (2016). «Внеклеточные/циркулирующие микроРНК: механизмы высвобождения, функции и проблемы» . Достижения в области наук о жизни . 10 (2): 175–186. дои : 10.1016/j.als.2016.11.007 .
- ^ Jump up to: а б Бекель Й.Н., Рейс С.М., Лейстнер Д., Томе К.Э., Зейхер А.М., Фихтльшерер С. и др. (апрель 2014 г.). «От сердца до пят: вклад сердца в уровень периферических микроРНК». Международный журнал кардиологии . 172 (3): 616–7. doi : 10.1016/j.ijcard.2014.01.082 . ПМИД 24508494 .
- ^ Леландэ-Брьер С., Сорин С., Деклерк М., Бенслиман А., Креспи М., Хартманн С. (март 2010 г.). «Разнообразие малых РНК у растений и его влияние на развитие» . Современная геномика . 11 (1): 14–23. дои : 10.2174/138920210790217918 . ПМЦ 2851111 . ПМИД 20808519 .
- ^ Рана ТМ (январь 2007 г.). «Освещая тишину: понимание структуры и функции малых РНК». Nature Reviews Молекулярно-клеточная биология . 8 (1): 23–36. дои : 10.1038/nrm2085 . ПМИД 17183358 . S2CID 8966239 .
- ^ Jump up to: а б Шварц Д.С., Замор П.Д. (май 2002 г.). «Почему микроРНК живут в микроРНК?» . Гены и развитие . 16 (9): 1025–31. дои : 10.1101/gad.992502 . ПМИД 12000786 .
- ^ Крол Дж., Собчак К., Вильчинска У., Драт М., Ясинска А., Качиньска Д. и др. (октябрь 2004 г.). «Структурные особенности предшественников микроРНК (миРНК) и их значение для биогенеза микроРНК и дизайна малых интерферирующих РНК/коротких шпилек РНК» . Журнал биологической химии . 279 (40): 42230–9. дои : 10.1074/jbc.M404931200 . ПМИД 15292246 .
- ^ Хворова А, Рейнольдс А, Джаясена С.Д. (октябрь 2003 г.). «Функциональные миРНК и микроРНК демонстрируют смещение цепи» . Клетка . 115 (2): 209–16. дои : 10.1016/S0092-8674(03)00801-8 . ПМИД 14567918 .
- ^ Шварц Д.С., Хутвагнер Г., Ду Т., Сюй З., Аронин Н., Замор П.Д. (октябрь 2003 г.). «Асимметрия сборки ферментного комплекса РНКи» . Клетка . 115 (2): 199–208. дои : 10.1016/S0092-8674(03)00759-1 . ПМИД 14567917 .
- ^ Линь С.Л., Чанг Д., Ин С.Ю. (август 2005 г.). «Асимметрия интронных структур пре-миРНК в функциональной сборке RISC» . Джин . 356 : 32–8. дои : 10.1016/j.gene.2005.04.036 . ПМК 1788082 . ПМИД 16005165 .
- ^ Окамура К., Чунг В.Дж., Лай Э.К. (сентябрь 2008 г.). «Длинные и короткие гены с инвертированными повторами у животных: микроРНК, миртроны и шпильковые РНК» . Клеточный цикл . 7 (18): 2840–5. дои : 10.4161/cc.7.18.6734 . ПМК 2697033 . ПМИД 18769156 .
- ^ Jump up to: а б Пратт А.Дж., Макрей И.Дж. (июль 2009 г.). «Комплекс молчания, индуцированный РНК: универсальная машина для подавления генов» . Журнал биологической химии . 284 (27): 17897–901. дои : 10.1074/jbc.R900012200 . ПМК 2709356 . ПМИД 19342379 .
- ^ Макрей И.Дж., Ма Э., Чжоу М., Робинсон К.В., Дудна Дж.А. (январь 2008 г.). «Восстановление in vitro человеческого RISC-загрузочного комплекса» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (2): 512–7. Бибкод : 2008PNAS..105..512M . дои : 10.1073/pnas.0710869105 . ПМК 2206567 . ПМИД 18178619 .
- ^ Мурелатос З., Дости Дж., Паушкин С., Шарма А., Шарру Б., Абель Л. и др. (март 2002 г.). «МиРНП: новый класс рибонуклеопротеинов, содержащий многочисленные микроРНК» . Гены и развитие . 16 (6): 720–8. дои : 10.1101/gad.974702 . ПМК 155365 . ПМИД 11914277 .
- ^ Мейстер Г., Ландталер М., Петерс Л., Чен П.Ю., Урлауб Х., Люрманн Р. и др. (декабрь 2005 г.). «Идентификация новых белков, связанных с аргонавтами» . Современная биология . 15 (23): 2149–55. Бибкод : 2005CBio...15.2149M . дои : 10.1016/j.cub.2005.10.048 . hdl : 11858/00-001M-0000-0012-E763-B . ПМИД 16289642 .
- ^ Цзин Ц, Хуан С., Гут С., Зарубин Т., Мотояма А., Чен Дж. и др. (март 2005 г.). «Участие микроРНК в нестабильности мРНК, опосредованной AU-богатыми элементами» . Клетка . 120 (5): 623–34. дои : 10.1016/j.cell.2004.12.038 . PMID 15766526 .
- ^ Jump up to: а б с Кай З.С., Паскинелли А.Е. (январь 2010 г.). «Убийцы микроРНК: факторы, регулирующие исчезновение микроРНК» . Структурная и молекулярная биология природы . 17 (1): 5–10. дои : 10.1038/nsmb.1762 . ПМК 6417416 . ПМИД 20051982 .
- ^ Чаттерджи С., Гроссханс Х. (сентябрь 2009 г.). «Активный оборот модулирует активность зрелых микроРНК у Caenorhabditis elegans». Природа . 461 (7263): 546–9. Бибкод : 2009Natur.461..546C . дои : 10.1038/nature08349 . ПМИД 19734881 . S2CID 4414841 .
- ^ Jump up to: а б с Морозова Н., Зиновьев А., Нонне Н., Притчард Л.Л., Горбан А.Н., Харель-Беллан А. (сентябрь 2012 г.). «Кинетические признаки действия микроРНК» . РНК . 18 (9): 1635–55. дои : 10.1261/rna.032284.112 . ПМЦ 3425779 . ПМИД 22850425 .
- ^ Ван XJ, Рейес Дж.Л., Чуа Н.Х., Гаастерланд Т. (2004). «Прогнозирование и идентификация микроРНК Arabidopsis thaliana и их мишеней мРНК» . Геномная биология . 5 (9): 65 р. дои : 10.1186/gb-2004-5-9-r65 . ПМК 522872 . ПМИД 15345049 .
- ^ Кавасаки Х, Тайра К (2004). «МикроРНК-196 ингибирует экспрессию HOXB8 при миелоидной дифференцировке клеток HL60» . Серия симпозиумов по нуклеиновым кислотам . 48 (1): 211–2. дои : 10.1093/насс/48.1.211 . ПМИД 17150553 .
- ^ Jump up to: а б Моксон С., Цзин Р., Шиття Г., Швах Ф., Рашолм Пилчер Р.Л., Моултон В. и др. (октябрь 2008 г.). «Глубокое секвенирование коротких РНК томатов идентифицирует микроРНК, нацеленные на гены, участвующие в созревании фруктов» . Геномные исследования . 18 (10): 1602–9. дои : 10.1101/гр.080127.108 . ПМК 2556272 . ПМИД 18653800 .
- ^ Мазьер П., Энрайт А.Дж. (июнь 2007 г.). «Прогнозирование мишеней микроРНК». Открытие наркотиков сегодня . 12 (11–12): 452–8. дои : 10.1016/j.drudis.2007.04.002 . ПМИД 17532529 .
- ^ Уильямс А.Е. (февраль 2008 г.). «Функциональные аспекты микроРНК животных» . Клеточные и молекулярные науки о жизни . 65 (4): 545–62. дои : 10.1007/s00018-007-7355-9 . ПМЦ 11131689 . ПМИД 17965831 . S2CID 5708394 .
- ^ Эулалио А., Ханцингер Э., Нишихара Т., Ревинкель Дж., Фаузер М., Изаурральде Э. (январь 2009 г.). «Деаденилирование является широко распространенным эффектом регуляции микроРНК» . РНК 15 (1): 21–3 дои : 10.1261/rna.1399509 . ПМЦ 2612776 . ПМИД 19029310 .
- ^ Баззини А.А., Ли М.Т., Хиральдес А.Дж. (апрель 2012 г.). «Профилирование рибосом показывает, что миР-430 снижает трансляцию, прежде чем вызвать распад мРНК у рыбок данио» . Наука . 336 (6078): 233–7. Бибкод : 2012Sci...336..233B . дои : 10.1126/science.1215704 . ПМЦ 3547538 . ПМИД 22422859 .
- ^ Джуранович С., Нахви А., Грин Р. (апрель 2012 г.). «миРНК-опосредованное замалчивание генов путем репрессии трансляции с последующим деаденилированием и распадом мРНК» . Наука . 336 (6078): 237–40. Бибкод : 2012Sci...336..237D . дои : 10.1126/science.1215691 . ПМЦ 3971879 . ПМИД 22499947 .
- ^ Тан Ю, Чжан Б, Ву Т, Скогербё Г, Чжу X, Го X и др. (февраль 2009 г.). «Транскрипционное ингибирование экспрессии Hoxd4 с помощью миРНК-10а в клетках рака молочной железы человека» . BMC Молекулярная биология . 10 (1): 12. дои : 10.1186/1471-2199-10-12 . ПМК 2680403 . ПМИД 19232136 .
- ^ Хокинс П.Г., Моррис К.В. (март 2008 г.). «РНК и транскрипционная модуляция экспрессии генов» . Клеточный цикл . 7 (5): 602–7. дои : 10.4161/cc.7.5.5522 . ПМЦ 2877389 . ПМИД 18256543 .
- ^ Старк А., Бреннеке Дж., Бушати Н., Рассел Р.Б., Коэн С.М. (декабрь 2005 г.). «МикроРНК животных придают устойчивость экспрессии генов и оказывают значительное влияние на эволюцию 3'UTR» . Клетка . 123 (6): 1133–46. дои : 10.1016/j.cell.2005.11.023 . ПМИД 16337999 .
- ^ Хе Л., Хэннон Дж.Дж. (июль 2004 г.). «МикроРНК: малые РНК, играющие большую роль в регуляции генов». Обзоры природы. Генетика . 5 (7): 522–531. дои : 10.1038/nrg1379 . ПМИД 15211354 . S2CID 5270062 .
- ^ Ли Л.К. (2008). «Активация генов, опосредованная малыми РНК» . В Моррисе К.В. (ред.). РНК и регуляция экспрессии генов: скрытый уровень сложности . Горизонт Научная Пресса. ISBN 978-1-904455-25-7 .
- ^ Плейс РФ, Ли Л.К., Пукот Д., Нунан Э.Дж., Дахия Р. (февраль 2008 г.). «МикроРНК-373 индуцирует экспрессию генов с комплементарными последовательностями промотора» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (5): 1608–13. Бибкод : 2008PNAS..105.1608P . дои : 10.1073/pnas.0707594105 . ПМК 2234192 . ПМИД 18227514 . (Ошибка: два : 10.1073/pnas.1803343115 , PMID 29555737 , Часы втягивания )
- ^ Салмена Л., Полисено Л., Тай Ю., Кац Л., Пандольфи П.П. (август 2011 г.). «Гипотеза ceRNA: Розеттский камень скрытого языка РНК?» . Клетка . 146 (3): 353–8. дои : 10.1016/j.cell.2011.07.014 . ПМК 3235919 . ПМИД 21802130 .
- ^ Jump up to: а б Кумар С., Редди П.Х. (сентябрь 2016 г.). «Являются ли циркулирующие микроРНК периферическими биомаркерами болезни Альцгеймера?» . Биохим Биофиз Акта . 1862 (9): 1617–27. дои : 10.1016/j.bbadis.2016.06.001 . ПМК 5343750 . ПМИД 27264337 .
- ^ ван ден Берг М.М., Краускопф Дж., Рамаекерс Дж.Г. и др. (февраль 2020 г.). «Циркулирующие микроРНК как потенциальные биомаркеры психиатрических и нейродегенеративных расстройств» . Прога Нейробиол . 185 : 101732. doi : 10.1016/j.pneurobio.2019.101732 . ПМИД 31816349 .
- ^ Куман С., Ван Синдерен М., Гантье М.П., Раинчук К., Сорби К., Ромбаутс Л. и др. (октябрь 2015 г.). «Секретируемые бластоцистой человека микроРНК регулируют адгезию эпителиальных клеток эндометрия» . Электронная биомедицина . 2 (10): 1528–1535. дои : 10.1016/j.ebiom.2015.09.003 . ПМЦ 4634783 . ПМИД 26629549 .
- ^ Чжоу Л., Миллер С., Миралья Л.Дж., Ромеро А., Муре Л.С., Панда С. и др. (январь 2021 г.). «Полногеномный скрининг микроРНК идентифицирует кластер микроРНК-183/96/182 как модулятор циркадных ритмов» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (1): e2020454118. Бибкод : 2021PNAS..11820454Z . дои : 10.1073/pnas.2020454118 . ПМЦ 7817116 . ПМИД 33443164 . S2CID 230713808 .
- «МикроРНК играют ключевую роль в регуляции циркадных ритмов» . Новости науки . 6 января 2021 г.
- ^ Axtell MJ, Bartel DP (июнь 2005 г.). «Древность микроРНК и их мишени в наземных растениях» . Растительная клетка . 17 (6): 1658–73. дои : 10.1105/tpc.105.032185 . ПМК 1143068 . ПМИД 15849273 .
- ^ Танцер А., Штадлер ПФ (май 2004 г.). «Молекулярная эволюция кластера микроРНК». Журнал молекулярной биологии . 339 (2): 327–35. CiteSeerX 10.1.1.194.1598 . дои : 10.1016/j.jmb.2004.03.065 . ПМИД 15136036 .
- ^ Чен К., Раевски Н. (февраль 2007 г.). «Эволюция регуляции генов с помощью факторов транскрипции и микроРНК». Обзоры природы Генетика . 8 (2): 93–103. дои : 10.1038/nrg1990 . ПМИД 17230196 . S2CID 174231 .
- ^ Ли К.Т., Рисом Т., Штраус В.М. (апрель 2007 г.). «Эволюционная консервативность регуляторных цепей микроРНК: исследование сложности генов микроРНК и консервативных взаимодействий микроРНК-мишень в филогении многоклеточных животных». ДНК и клеточная биология . 26 (4): 209–18. дои : 10.1089/dna.2006.0545 . PMID 17465887 .
- ^ Jump up to: а б с д Петерсон К.Дж., Дитрих М.Р., МакПик М.А. (июль 2009 г.). «МикроРНК и макроэволюция многоклеточных животных: понимание канализации, сложности и кембрийского взрыва» . Биоэссе . 31 (7): 736–47. doi : 10.1002/bies.200900033 . ПМИД 19472371 . S2CID 15364875 .
- ^ Шабалина С.А., Кунин Е.В. (октябрь 2008 г.). «Происхождение и эволюция эукариотической РНК-интерференции» . Тенденции в экологии и эволюции . 23 (10): 578–87. дои : 10.1016/j.tree.2008.06.005 . ПМЦ 2695246 . ПМИД 18715673 .
- ^ Экстелл М.Дж., Вестхольм Дж.О., Лай Э.К. (2011). «Vive la différence: биогенез и эволюция микроРНК у растений и животных» . Геномная биология . 12 (4): 221. doi : 10.1186/gb-2011-12-4-221 . ПМК 3218855 . ПМИД 21554756 .
- ^ Jump up to: а б Уилер Б.М., Хаймберг А.М., Мой В.Н., Сперлинг Е.А., Хольштейн Т.В., Хибер С. и др. (2009). «Глубокая эволюция микроРНК многоклеточных животных». Эволюция и развитие . 11 (1): 50–68. дои : 10.1111/j.1525-142X.2008.00302.x . ПМИД 19196333 . S2CID 14924603 .
- ^ Пашковский ПП, Рязанский СС (июнь 2013 г.). «Биогенез, эволюция и функции растительных микроРНК». Биохимия. Биохимия . 78 (6): 627–37. дои : 10.1134/S0006297913060084 . ПМИД 23980889 . S2CID 12025420 .
- ^ Jump up to: а б Хеймберг А.М., Семпере Л.Ф., Мой В.Н., Донохью ПК, Петерсон К.Дж. (февраль 2008 г.). «МикроРНК и появление морфологической сложности позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (8): 2946–50. Бибкод : 2008PNAS..105.2946H . дои : 10.1073/pnas.0712259105 . ПМК 2268565 . ПМИД 18287013 .
- ^ Jump up to: а б с Нозава М., Миура С., Ней М. (июль 2010 г.). «Происхождение и эволюция генов микроРНК у видов дрозофилы» . Геномная биология и эволюция . 2 : 180–9. дои : 10.1093/gbe/evq009 . ПМК 2942034 . ПМИД 20624724 .
- ^ Аллен Э., Се З., Густафсон А.М., Сунг Г.Х., Спатафора Дж.В., Кэррингтон Дж.К. (декабрь 2004 г.). «Эволюция генов микроРНК путем инвертированного дупликации последовательностей целевых генов у Arabidopsis thaliana». Природная генетика . 36 (12): 1282–90. дои : 10.1038/ng1478 . ПМИД 15565108 . S2CID 11997028 .
- ^ Вартманн Н., Дас С., Ланц С., Вейгель Д. (май 2008 г.). «Сравнительный анализ локуса микроРНК MIR319a у Arabidopsis и родственных Brassicaceae» . Молекулярная биология и эволюция . 25 (5): 892–902. дои : 10.1093/molbev/msn029 . ПМИД 18296705 .
- ^ Фальгрен Н., Джогдео С., Касшау К.Д., Салливан С.М., Чепмен Э.Дж., Лаубингер С. и др. (апрель 2010 г.). «Эволюция генов микроРНК у Arabidopsis lyrata и Arabidopsis thaliana» . Растительная клетка . 22 (4): 1074–89. дои : 10.1105/tpc.110.073999 . ПМЦ 2879733 . ПМИД 20407027 .
- ^ Каравас Дж., Фридрих М. (июнь 2010 г.). «О клещах и многоножках: недавний прогресс в разрешении основания дерева членистоногих». Биоэссе . 32 (6): 488–95. doi : 10.1002/bies.201000005 . ПМИД 20486135 . S2CID 20548122 .
- ^ Кенни Нью-Джерси, Намигай Э.К., Марлетаз Ф., Хуэй Дж.Х., Шимелд С.М. (декабрь 2015 г.). «Проект сборок генома и прогнозируемые комплементы микроРНК приливных лофотрохозойцев Patella vulgata (Mollusca, Patellogastropoda) и Spirobranchus (Pomatoceros) lamarcki (Annelida, Serpulida)» . Морская геномика . 24 (2): 139–46. Бибкод : 2015Март..24..139К . дои : 10.1016/j.margen.2015.07.004 . ПМИД 26319627 .
- ^ Кок Дж.М., Стерк Л., Рузе П., Скорнет Д., Аллен А.Е., Амуциас Г. и др. (июнь 2010 г.). «Геном Ectocarpus и независимая эволюция многоклеточности бурых водорослей» . Природа . 465 (7298): 617–21. Бибкод : 2010Natur.465..617C . дои : 10.1038/nature09016 . ПМИД 20520714 .
- ^ Куперус Дж.Т., Фальгрен Н., Кэррингтон Дж.К. (февраль 2011 г.). «Эволюция и функциональное разнообразие генов MIRNA» . Растительная клетка . 23 (2): 431–42. дои : 10.1105/tpc.110.082784 . ПМК 3077775 . ПМИД 21317375 .
- ^ Райан Дж. Ф., Панг К., Шницлер С. Э., Нгуен А. Д., Морленд РТ, Симмонс Д. К. и др. (декабрь 2013 г.). «Геном гребневика Mnemiopsis leidyi и его значение для эволюции типов клеток» . Наука . 342 (6164): 1242592. doi : 10.1126/science.1242592 . ПМЦ 3920664 . ПМИД 24337300 .
- ^ Максвелл Э.К., Райан Дж.Ф., Шницлер К.Э., Браун В.Е., Баксеванис А.Д. (декабрь 2012 г.). «МикроРНК и важные компоненты механизма обработки микроРНК не закодированы в геноме гребневика Mnemiopsis leidyi» . БМК Геномика . 13 (1): 714. дои : 10.1186/1471-2164-13-714 . ПМЦ 3563456 . ПМИД 23256903 .
- ^ Даймонд П.Ф. (15 марта 2010 г.). «Терапевтический потенциал микроРНК» . Новости генной инженерии и биотехнологии . 30 (6): 1. Архивировано из оригинала 19 июля 2010 года . Проверено 10 июля 2010 г.
- ^ Тьяден Б., Гудвин С.С., Опдайк Дж.А., Гильер М., Фу Д.С., Готтесман С. и др. (2006). «Целевой прогноз для малых некодирующих РНК в бактериях» . Исследования нуклеиновых кислот . 34 (9): 2791–802. дои : 10.1093/нар/gkl356 . ПМЦ 1464411 . ПМИД 16717284 .
- ^ Лю К.Г., Калин Г.А., Волиния С., Кроче С.М. (2008). «Профилирование экспрессии микроРНК с использованием микрочипов». Протоколы природы . 3 (4): 563–78. дои : 10.1038/nprot.2008.14 . ПМИД 18388938 . S2CID 2441105 .
- ^ Чен С., Ридзон Д.А., Брумер А.Дж., Чжоу З., Ли Д.Х., Нгуен Дж.Т. и др. (ноябрь 2005 г.). «Количественное определение микроРНК в реальном времени с помощью RT-PCR на основе стволовой петли» . Исследования нуклеиновых кислот . 33 (20): e179. дои : 10.1093/nar/gni178 . ПМК 1292995 . ПМИД 16314309 .
- ^ Шингара Дж., Кейгер К., Шелтон Дж., Лаосинчай-Вольф В., Пауэрс П., Конрад Р. и др. (сентябрь 2005 г.). «Оптимизированная платформа выделения и маркировки для точного профилирования экспрессии микроРНК» . РНК . 11 (9): 1461–70. дои : 10.1261/rna.2610405 . ПМЦ 1370829 . ПМИД 16043497 .
- ^ Буерманс Х.П., Ариюрек Ю., ван Оммен Г., ден Даннен Дж.Т., 'т Хоэн П.А. (декабрь 2010 г.). «Новые методы определения профиля экспрессии микроРНК на основе секвенирования следующего поколения» . БМК Геномика . 11 : 716. дои : 10.1186/1471-2164-11-716 . ПМК 3022920 . ПМИД 21171994 .
- ^ Клоостерман В.П., Винхольдс Э., Кеттинг Р.Ф., Пластерк Р.Х. (2004). «Требования к субстрату для функционирования let-7 в развивающемся эмбрионе рыбки данио» . Исследования нуклеиновых кислот . 32 (21): 6284–91. дои : 10.1093/nar/gkh968 . ПМК 535676 . ПМИД 15585662 .
- ^ Флинт А.С., Ли Н., Тэтчер Э.Дж., Солника-Крезель Л., Паттон Дж.Г. (февраль 2007 г.). «МиР-214 рыбки данио модулирует передачу сигналов Hedgehog, чтобы определить судьбу мышечных клеток» . Природная генетика . 39 (2): 259–63. дои : 10.1038/ng1953 . ПМЦ 3982799 . ПМИД 17220889 .
- ^ Мейстер Г., Ландталер М., Дорсетт Ю., Тушл Т. (март 2004 г.). «Последовательность-специфическое ингибирование молчания РНК, индуцированного микроРНК и миРНК» . РНК . 10 (3): 544–50. дои : 10.1261/rna.5235104 . ПМК 1370948 . ПМИД 14970398 .
- ^ Клоостерман В.П., Лагендейк А.К., Кеттинг Р.Ф., Моултон Дж.Д., Пластерк Р.Х. (август 2007 г.). «Направленное ингибирование созревания миРНК с помощью морфолино раскрывает роль миР-375 в развитии островков поджелудочной железы» . ПЛОС Биология . 5 (8): е203. doi : 10.1371/journal.pbio.0050203 . ЧВК 1925136 . ПМИД 17676975 . (В настоящее время в этом документе выражается обеспокоенность , см. дои : 10.1371/journal.pbio.3001631 , ПМИД 35486652 )
- ^ Геберт Л.Ф., Ребхан М.А., Кривелли С.Е., Дензлер Р., Стоффель М., Холл Дж. (январь 2014 г.). «Миравирсен (SPC3649) может ингибировать биогенез миР-122» . Исследования нуклеиновых кислот . 42 (1): 609–21. дои : 10.1093/нар/gkt852 . ПМЦ 3874169 . ПМИД 24068553 .
- ^ Чой Вайоминг, Хиральдес А.Дж., Шир А.Ф. (октябрь 2007 г.). «Защитники мишени демонстрируют подавление и балансировку агониста и антагониста узла с помощью миР-430» . Наука . 318 (5848): 271–4. Бибкод : 2007Sci...318..271C . дои : 10.1126/science.1147535 . ПМИД 17761850 . S2CID 30461594 .
- ^ Ю Ю, Морейра Б.Г., Бельке М.А., Овзарзи Р. (май 2006 г.). «Разработка МШУ-зондов, улучшающих распознавание несоответствий» . Исследования нуклеиновых кислот . 34 (8): е60. дои : 10.1093/нар/gkl175 . ПМЦ 1456327 . ПМИД 16670427 .
- ^ Лагендейк А.К., Моултон Дж.Д., Баккерс Дж. (июнь 2012 г.). «Показательные детали: протокол гибридизации in situ микроРНК целиком для эмбрионов рыбок данио и взрослых тканей» . Биология Открытая . 1 (6): 566–9. дои : 10.1242/bio.2012810 . ПМК 3509442 . ПМИД 23213449 .
- ^ Каур Х., Арора А., Венгель Дж., Маити С. (июнь 2006 г.). «Термодинамические, противоионные и гидратационные эффекты для включения заблокированных нуклеотидов нуклеиновой кислоты в дуплексы ДНК». Биохимия . 45 (23): 7347–55. дои : 10.1021/bi060307w . ПМИД 16752924 .
- ^ Нильсен Дж.А., Лау П., Марик Д., Баркер Дж.Л., Хадсон Л.Д. (август 2009 г.). «Интеграция профилей экспрессии микроРНК и мРНК предшественников нейронов для выявления регуляторных сетей, лежащих в основе начала коркового нейрогенеза» . BMC Нейронаука . 10:98 . дои : 10.1186/1471-2202-10-98 . ПМЦ 2736963 . ПМИД 19689821 .
- ^ Гримсон А., Фарх К.К., Джонстон В.К., Гаррет-Энгеле П., Лим Л.П., Бартель Д.П. (июль 2007 г.). «Специфичность нацеливания микроРНК у млекопитающих: детерминанты, выходящие за рамки спаривания семян» . Молекулярная клетка . 27 (1): 91–105. doi : 10.1016/j.molcel.2007.06.017 . ПМК 3800283 . ПМИД 17612493 .
- ^ Гриффитс-Джонс С., Сайни Х.К., ван Донген С., Энрайт А.Дж. (январь 2008 г.). «miRBase: инструменты для геномики микроРНК» . Исследования нуклеиновых кислот . 36 (Проблема с базой данных): D154–8. дои : 10.1093/нар/gkm952 . ПМК 2238936 . ПМИД 17991681 .
- ^ Нам С., Ли М., Чой К., Балч С., Ким С., Племянник К.П. (июль 2009 г.). «Интегрированный анализ микроРНК и мРНК (MMIA): веб-инструмент для изучения биологических функций экспрессии микроРНК» . Исследования нуклеиновых кислот . 37 (проблема с веб-сервером): W356–62. дои : 10.1093/nar/gkp294 . ПМК 2703907 . ПМИД 19420067 .
- ^ Артманн С., Юнг К., Блекманн А., Байсбарт Т. (2012). Проверо П. (ред.). «Обнаружение одновременных групповых эффектов в экспрессии микроРНК и связанных с ней наборах целевых генов» . ПЛОС ОДИН . 7 (6): e38365. Бибкод : 2012PLoSO...738365A . дои : 10.1371/journal.pone.0038365 . ПМЦ 3378551 . ПМИД 22723856 .
- ^ Цзян Ц, Ван Ю, Хао Ю, Хуан Л, Тенг М, Чжан Икс и др. (январь 2009 г.). «miR2Disease: созданная вручную база данных для дерегуляции микроРНК при заболеваниях человека» . Исследования нуклеиновых кислот . 37. 37 (Проблема с базой данных): D98–104. дои : 10.1093/нар/gkn714 . ПМК 2686559 . ПМИД 18927107 .
- ^ Менсия А., Модамио-Хойбьер С., Редшоу Н., Морин М., Майо-Мерино Ф., Олаварриета Л. и др. (май 2009 г.). «Мутации в семенной области человеческой миР-96 ответственны за несиндромальную прогрессирующую потерю слуха». Природная генетика . 41 (5): 609–13. дои : 10.1038/ng.355 . ПМИД 19363479 . S2CID 11113852 .
- ^ Хьюз А.Е., Брэдли Д.Т., Кэмпбелл М., Лехнер Дж., Дэш Д.П., Симпсон Д.А. и др. (ноябрь 2011 г.). «Мутация, изменяющая семенную область миР-184, вызывает семейный кератоконус с катарактой» . Американский журнал генетики человека . 89 (5): 628–33. дои : 10.1016/j.ajhg.2011.09.014 . ПМЦ 3213395 . ПМИД 21996275 .
- ^ де Понтуаль Л., Яо Э., Калье П., Фавр Л., Друэн В., Кариу С. и др. (сентябрь 2011 г.). «Удаление зародышевой линии кластера миР-17~92 вызывает дефекты скелета и роста у людей» . Природная генетика . 43 (10): 1026–30. дои : 10.1038/ng.915 . ПМК 3184212 . ПМИД 21892160 .
- ^ Калин Г.А., Лю К.Г., Севиньяни С., Феррацин М., Фелли Н., Думитру К.Д. и др. (август 2004 г.). «Профилирование микроРНК выявляет отчетливые признаки В-клеточного хронического лимфоцитарного лейкоза» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (32): 11755–11760. Бибкод : 2004PNAS..10111755C . дои : 10.1073/pnas.0404432101 . ПМК 511048 . ПМИД 15284443 .
- ^ Велле А., Песенти С., Грасси Т., Бельтрам Л., Мартини П., Якони М. и др. (май 2023 г.). «Комплексное исследование гистотип-специфичных микроРНК и их вариантов при эпителиальном раке яичников I стадии» . Международный журнал рака . 152 (9): 1989–2001. дои : 10.1002/ijc.34408 . ПМИД 36541726 . S2CID 255034585 .
- ^ Кириакидис И, Кириакидис К, Цезу А (август 2022 г.). «МикроРНК и диагностика острого лимфобластного лейкоза у детей: систематический обзор, метаанализ и повторный анализ с использованием новых инструментов секвенирования малых РНК» . Раки . 14 (16): 3976. doi : 10.3390/cancers14163976 . ПМЦ 9406077 . ПМИД 36010971 .
- ^ Выса У, Воодер Т, Кольде Р, Фишер К, Вялк К, Тыниссон Н и др. (октябрь 2011 г.). «Идентификация миР-374a как прогностического маркера выживаемости у пациентов с немелкоклеточным раком легкого на ранней стадии» . Гены, хромосомы и рак . 50 (10): 812–22. дои : 10.1002/gcc.20902 . ПМИД 21748820 . S2CID 9746594 .
- ^ Акчакая П., Экелунд С., Колосенко И., Карамута С., Озата Д.М., Се Х. и др. (август 2011 г.). «Нарушение регуляции миР-185 и миР-133b связано с общей выживаемостью и метастазированием при колоректальном раке» . Международный журнал онкологии . 39 (2): 311–8. дои : 10.3892/ijo.2011.1043 . ПМИД 21573504 .
- ^ Jump up to: а б Эйкинг А., Рейс Х., Франк М., Геркен Г., Шмид К.В., Карио Э. (2016). «МиР-205 и МиР-373 связаны с агрессивным муцинозным колоректальным раком человека» . ПЛОС ОДИН . 11 (6): e0156871. Бибкод : 2016PLoSO..1156871E . дои : 10.1371/journal.pone.0156871 . ПМЦ 4894642 . ПМИД 27271572 .
- ^ МикроРНК-21 способствует пролиферации клеток гепатоцеллюлярной карциномы HepG2 посредством репрессии митоген-активируемой протеинкиназы-киназы 3. Guangxian Xu et al., 2013
- ^ Джонс К., Нурс Дж.П., Кин С., Бхатнагар А., Ганди М.К. (январь 2014 г.). «МикроРНК плазмы являются биомаркерами реакции на заболевание при классической лимфоме Ходжкина» . Клинические исследования рака . 20 (1): 253–64. дои : 10.1158/1078-0432.CCR-13-1024 . ПМИД 24222179 .
- ^ Хоссейнали Н., Агапур М., Дуйф П.Х., Барадаран Б. (август 2018 г.). «Лечение рака заместительной терапией микроРНК: обзор литературы» . Журнал клеточной физиологии . 233 (8): 5574–5588. дои : 10.1002/jcp.26514 . ПМИД 29521426 . S2CID 3766576 .
- ^ Лю Дж., Сунь Ю., Цзи П., Ли Х., Когделл Д., Ян Д. и др. (июль 2014 г.). «МиР-506 подавляет пролиферацию и индуцирует старение, напрямую воздействуя на ось CDK4/6-FOXM1 при раке яичников» . Журнал патологии . 233 (3): 308–18. дои : 10.1002/путь.4348 . ПМК 4144705 . ПМИД 24604117 .
- ^ Вэнь С.Ю., Линь Ю., Ю Ю.К., Цао С.Дж., Чжан Р., Ян С.М. и др. (февраль 2015 г.). «МиР-506 действует как супрессор опухоли, напрямую воздействуя на фактор транскрипции Gli3 пути hedgehog при раке шейки матки человека» . Онкоген . 34 (6): 717–25. дои : 10.1038/onc.2014.9 . ПМИД 24608427 . S2CID 20603801 .
- ^ Пекинский университет (13 сентября 2022 г.). «Ученые разрабатывают новое мощное оружие для борьбы с раком» . Клетка . 185 (11). SciTech Daily : 1888–1904.e24. дои : 10.1016/j.cell.2022.04.030 . ПМИД 35623329 . S2CID 249070106 . Проверено 15 сентября 2022 г.
- ^ Леб К.Р., Леб Л.А. (март 2000 г.). «Значение множественных мутаций при раке» . Канцерогенез . 21 (3): 379–385. дои : 10.1093/carcin/21.3.379 . PMID 10688858 .
- ^ Лодиш Х., Берк А., Кайзер К.А., Кригер М., Бретшер А., Плох Х. и др. (2016). Молекулярно-клеточная биология (8-е изд.). Нью-Йорк: WH Freeman and Company. п. 203. ИСБН 978-1-4641-8339-3 .
- ^ Ху Х, Гатти Р.А. (июнь 2011 г.). «МикроРНК: новые игроки в реакции на повреждение ДНК» . Журнал молекулярно-клеточной биологии . 3 (3): 151–158. дои : 10.1093/jmcb/mjq042 . ПМК 3104011 . ПМИД 21183529 .
- ^ Джасперсон К.В., Туохи Т.М., Некласон Д.В., Берт Р.В. (июнь 2010 г.). «Наследственный и семейный рак толстой кишки» . Гастроэнтерология . 138 (6): 2044–58. дои : 10.1053/j.gastro.2010.01.054 . ПМК 3057468 . ПМИД 20420945 .
- ^ Трунингер К., Менигатти М., Луз Дж., Рассел А., Хайдер Р., Гебберс Дж.О. и др. (май 2005 г.). «Иммуногистохимический анализ выявляет высокую частоту дефектов PMS2 при колоректальном раке» . Гастроэнтерология . 128 (5): 1160–71. дои : 10.1053/j.gastro.2005.01.056 . ПМИД 15887099 .
- ^ Валери Н., Гаспарини П., Фаббри М., Бракони С., Веронезе А., Ловат Ф. и др. (апрель 2010 г.). «Модуляция репарации ошибочных спариваний и стабильности генома с помощью миР-155» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (15): 6982–7. Бибкод : 2010PNAS..107.6982V . дои : 10.1073/pnas.1002472107 . ПМЦ 2872463 . ПМИД 20351277 .
- ^ Jump up to: а б с Чжан В., Чжан Дж., Ходли К., Кушваха Д., Рамакришнан В., Ли С. и др. (июнь 2012 г.). «миР-181d: прогнозирующий биомаркер глиобластомы, который подавляет экспрессию MGMT» . Нейроонкология . 14 (6): 712–9. дои : 10.1093/neuonc/nos089 . ПМЦ 3367855 . ПМИД 22570426 .
- ^ Шпигль-Крейнекер С., Пиркер С., Филипитс М., Лётч Д., Бухройтнер Дж., Пихлер Дж. и др. (январь 2010 г.). «Экспрессия белка O6-метилгуанин-ДНК-метилтрансферазы в опухолевых клетках предсказывает результат терапии темозоломидом у пациентов с глиобластомой» . Нейроонкология . 12 (1): 28–36. дои : 10.1093/neuonc/nop003 . ПМЦ 2940563 . ПМИД 20150365 .
- ^ Сгарра Р., Рустиги А., Тессари М.А., Ди Бернардо Дж., Альтамура С., Фуско А. и др. (сентябрь 2004 г.). «Ядерные фосфопротеины HMGA и их связь со структурой хроматина и раком». Письма ФЭБС . 574 (1–3): 1–8. дои : 10.1016/j.febslet.2004.08.013 . ПМИД 15358530 . S2CID 28903539 .
- ^ Сюй Ю, Самтер Т.Ф., Бхаттачарья Р., Тесфай А., Фукс Э.Дж., Вуд Л.Дж. и др. (май 2004 г.). «Онкоген HMG-I вызывает высоко проникающие агрессивные лимфоидные злокачественные новообразования у трансгенных мышей и сверхэкспрессируется при лейкемии человека» . Исследования рака . 64 (10): 3371–5. дои : 10.1158/0008-5472.CAN-04-0044 . ПМИД 15150086 .
- ^ Боррманн Л., Шванбек Р., Хейдук Т., Зеебек Б., Рогалла П., Буллердик Дж. и др. (декабрь 2003 г.). «Белок группы А2 с высокой подвижностью и его производные связывают конкретную область промотора гена репарации ДНК ERCC1 и модулируют его активность» . Исследования нуклеиновых кислот . 31 (23): 6841–51. дои : 10.1093/nar/gkg884 . ПМК 290254 . ПМИД 14627817 .
- ^ Фациста А., Нгуен Х., Льюис С., Прасад А.Р., Рэмси Л., Зайтлин Б. и др. (апрель 2012 г.). «Недостаточная экспрессия ферментов репарации ДНК на ранней стадии развития спорадического рака толстой кишки» . Целостность генома . 3 (1): 3. дои : 10.1186/2041-9414-3-3 . ПМК 3351028 . ПМИД 22494821 .
- ^ Жибер Б., Деллуа-Буржуа С., Гаттолья Ш., Меретт О., Ле Герневель С., Фомбонн Дж. и др. (ноябрь 2014 г.). «Регуляция с помощью семейства miR181 опухолевой супрессорной активности рецептора зависимости CDON при нейробластоме» . Журнал Национального института рака . 106 (11). дои : 10.1093/jnci/dju318 . ПМИД 25313246 .
- ^ Чен Дж.Ф., Мерчисон Э.П. , Тан Р., Каллис Т.Э., Тацугучи М., Дэн З. и др. (февраль 2008 г.). «Направленное удаление Dicer в сердце приводит к дилатационной кардиомиопатии и сердечной недостаточности» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (6): 2111–6. Бибкод : 2008PNAS..105.2111C . дои : 10.1073/pnas.0710228105 . ПМК 2542870 . ПМИД 18256189 .
- ^ Jump up to: а б Чжао Ю., Рэнсом Дж.Ф., Ли А., Ведантам В., фон Дреле М., Мут А.Н. и др. (апрель 2007 г.). «Нарушение регуляции кардиогенеза, сердечной проводимости и клеточного цикла у мышей, лишенных микроРНК-1-2» . Клетка . 129 (2): 303–17. дои : 10.1016/j.cell.2007.03.030 . ПМИД 17397913 .
- ^ Тум Т., Галуппо П., Вольф С., Фидлер Дж., Кнейтц С., ван Лааке Л.В. и др. (июль 2007 г.). «МикроРНК в сердце человека: ключ к перепрограммированию генов плода при сердечной недостаточности» . Тираж . 116 (3): 258–67. doi : 10.1161/CIRCULATIONAHA.107.687947 . ПМИД 17606841 .
- ^ ван Рой Э., Сазерленд Л.Б., Лю Н., Уильямс А.Х., Макэналли Дж., Джерард Р.Д. и др. (ноябрь 2006 г.). «Характерный образец стресс-чувствительных микроРНК, которые могут вызвать гипертрофию сердца и сердечную недостаточность» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (48): 18255–60. Бибкод : 2006PNAS..10318255V . дои : 10.1073/pnas.0608791103 . ПМЦ 1838739 . ПМИД 17108080 .
- ^ Тацугути М., Сок Х.И., Каллис Т.Э., Томсон Дж.М., Чен Дж.Ф., Ньюман М. и др. (июнь 2007 г.). «Экспрессия микроРНК динамически регулируется во время гипертрофии кардиомиоцитов» . Журнал молекулярной и клеточной кардиологии . 42 (6): 1137–41. дои : 10.1016/j.yjmcc.2007.04.004 . ЧВК 1934409 . ПМИД 17498736 .
- ^ Чжао Ю, Самал Э, Шривастава Д (июль 2005 г.). «Фактор ответа сыворотки регулирует специфическую для мышц микроРНК, которая нацелена на Hand2 во время кардиогенеза». Природа . 436 (7048): 214–20. Бибкод : 2005Natur.436..214Z . дои : 10.1038/nature03817 . ПМИД 15951802 . S2CID 4340449 .
- ^ Сяо Дж, Луо Икс, Линь Х, Чжан Ю, Лу Ю, Ван Н и др. (апрель 2007 г.). «МикроРНК миР-133 подавляет экспрессию канала HERG K+, способствуя удлинению интервала QT в диабетическом сердце» . Журнал биологической химии . 282 (17): 12363–7. дои : 10.1074/jbc.C700015200 . ПМК 3151106 . ПМИД 17344217 . (Отозвано, см. дои : 10.1074/jbc.A111.700015 , PMID 21961173 , Часы втягивания )
- ^ Ян Б., Линь Х., Сяо Дж., Лу Ю., Луо Икс., Ли Б. и др. (апрель 2007 г.). «Мышечная специфичная микроРНК миР-1 регулирует сердечный аритмогенный потенциал, воздействуя на GJA1 и KCNJ2». Природная медицина . 13 (4): 486–91. дои : 10.1038/nm1569 . ПМИД 17401374 . S2CID 1935811 .
- ^ Каре А., Каталуччи Д., Феличетти Ф., Бончи Д., Аддарио А., Галло П. и др. (май 2007 г.). «МикроРНК-133 контролирует гипертрофию сердца». Природная медицина . 13 (5): 613–8. дои : 10.1038/nm1582 . ПМИД 17468766 . S2CID 10097893 .
- ^ ван Рой Э., Сазерленд Л.Б., Ци Икс, Ричардсон Дж.А., Хилл Дж., Олсон Э.Н. (апрель 2007 г.). «Контроль стресс-зависимого роста сердца и экспрессии генов с помощью микроРНК». Наука . 316 (5824): 575–9. Бибкод : 2007Sci...316..575V . дои : 10.1126/science.1139089 . ПМИД 17379774 . S2CID 1927839 .
- ^ Келлер Т., Бекель Дж.Н., Гросс С., Клотше Дж., Палапис Л., Лейстнер Д. и др. (июль 2017 г.). «Улучшенная стратификация риска при профилактике за счет использования панели выбранных циркулирующих микроРНК» . Научные отчеты . 7 (1): 4511. Бибкод : 2017НатСР...7.4511К . дои : 10.1038/s41598-017-04040-w . ПМЦ 5495799 . ПМИД 28674420 .
- ^ Инсулл В. (январь 2009 г.). «Патология атеросклероза: развитие бляшек и реакция бляшек на лечение». Американский медицинский журнал . 122 (1 дополнение): S3 – S14. doi : 10.1016/j.amjmed.2008.10.013 . ПМИД 19110086 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Сон Д.Д., Кумар С., Такабэ В., Ким К.В., Ни К.В., Альбертс-Гриль Н. и др. (2013). «Атипичная механочувствительная микроРНК-712, полученная из прерибосомальной РНК, вызывает воспаление эндотелия и атеросклероз» . Природные коммуникации . 4 : 3000. Бибкод : 2013NatCo...4.3000S . дои : 10.1038/ncomms4000 . ПМЦ 3923891 . ПМИД 24346612 .
- ^ Jump up to: а б Басу Р., Фан Д., Кандалам В., Ли Дж., Дас С.К., Ван Х и др. (декабрь 2012 г.). «Потеря гена Timp3 приводит к образованию аневризмы брюшной аорты в ответ на ангиотензин II» . Журнал биологической химии . 287 (53): 44083–96. дои : 10.1074/jbc.M112.425652 . ПМЦ 3531724 . ПМИД 23144462 .
- ^ Либби П. (2002). «Воспаление при атеросклерозе». Природа . 420 (6917): 868–74. Бибкод : 2002Natur.420..868L . дои : 10.1038/nature01323 . ПМИД 12490960 . S2CID 407449 .
- ^ Jump up to: а б Фуа Ю.Л., Чу Дж.И., Марроне А.К., Боднар А.Дж., Симс-Лукас С., Хо Дж. (октябрь 2015 г.). «МиРНК почечной стромы необходимы для нормального нефрогенеза и выживания клубочковой мезангии» . Физиологические отчеты . 3 (10): е12537. дои : 10.14814/phy2.12537 . ПМЦ 4632944 . ПМИД 26438731 .
- ^ Цао Д.Д., Ли Л., Чан Вайоминг (май 2016 г.). «МикроРНК: ключевые регуляторы центральной нервной системы и их значение при неврологических заболеваниях» . Международный журнал молекулярных наук . 17 (6): 842. doi : 10.3390/ijms17060842 . ПМЦ 4926376 . ПМИД 27240359 .
- ^ Ланг М.Ф., Ши Ю (2012). «Динамическая роль микроРНК в нейрогенезе» . Границы в неврологии . 6 : 71. дои : 10.3389/fnins.2012.00071 . ПМК 3356852 . ПМИД 22661924 .
- ^ Шратт Дж. (декабрь 2009 г.). «микроРНК в синапсе». Обзоры природы. Нейронаука . 10 (12): 842–849. дои : 10.1038/nrn2763 . ПМИД 19888283 . S2CID 3507952 .
- ^ Луо М, Ли Л, Дин М, Ню Ю, Сюй Х, Ши Х и др. (19 января 2023 г.). «С помощью секвенирования микроРНК и биоинформатического анализа были выявлены микроРНК, регулирующие долговременное потенцирование и депрессию при нарушениях обучения и памяти, вызванных бисфенолом А» . ПЛОС ОДИН . 18 (1): e0279029. Бибкод : 2023PLoSO..1879029L . дои : 10.1371/journal.pone.0279029 . ПМЦ 9851566 . PMID 36656826 .
- ^ Куэльяр Т.Л., Дэвис Т.Х., Нельсон П.Т., Леб Г.Б., Харф Б.Д., Уллиан Э. и др. (апрель 2008 г.). «Потеря Dicer в нейронах полосатого тела приводит к поведенческим и нейроанатомическим фенотипам при отсутствии нейродегенерации» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (14): 5614–5619. Бибкод : 2008PNAS..105.5614C . дои : 10.1073/pnas.0801689105 . ПМК 2291142 . ПМИД 18385371 .
- ^ Эбер С.С., Пападопулу А.С., Смит П., Галас М.К., Планель Э., Силахтароглу А.Н. и др. (октябрь 2010 г.). «Генетическая абляция Dicer в нейронах переднего мозга взрослых приводит к аномальному гиперфосфорилированию тау и нейродегенерации» . Молекулярная генетика человека . 19 (20): 3959–3969. дои : 10.1093/hmg/ddq311 . ПМИД 20660113 .
- ^ Рой Б., Ли Э., Ли Т., Рамперсауд М. (февраль 2022 г.). «Роль микроРНК в нейродегенерации: от причины заболевания к инструментам открытия биомаркеров и терапии» . Гены . 13 (3): 425. doi : 10.3390/genes13030425 . ПМЦ 8951370 . ПМИД 35327979 .
- ^ Хеншалл Д.К., Хамер Х.М., Пастеркамп Р.Дж., Гольдштейн Д.Б., Кджемс Дж., Прен Дж.Х. и др. (декабрь 2016 г.). «МикроРНК при эпилепсии: патофизиология и клиническое применение» . «Ланцет». Неврология . 15 (13): 1368–1376. дои : 10.1016/S1474-4422(16)30246-0 . ПМИД 27839653 .
- ^ Хоммерс Л.Г., Домшке К., Декерт Дж. (январь 2015 г.). «Гетерогенность и индивидуальность: микроРНК при психических расстройствах» (PDF) . Журнал нейронной передачи . 122 (1): 79–97. дои : 10.1007/s00702-014-1338-4 . ПМИД 25395183 . S2CID 25088900 . Архивировано (PDF) из оригинала 23 мая 2022 года.
- ^ Фэн Дж., Сунь Г., Ян Дж., Нолтнер К., Ли В., Бузин CH. и др. (июль 2009 г.). Рейф А (ред.). «Доказательства Х-хромосомной шизофрении, связанной с изменениями микроРНК» . ПЛОС ОДИН . 4 (7): е6121. Бибкод : 2009PLoSO...4.6121F . дои : 10.1371/journal.pone.0006121 . ПМК 2699475 . ПМИД 19568434 .
- ^ Беверидж, Нью-Джерси, Гардинер Э., Кэрролл А.П., Туни П.А., Кэрнс М.Дж. (декабрь 2010 г.). «Шизофрения связана с увеличением биогенеза кортикальных микроРНК» . Молекулярная психиатрия . 15 (12): 1176–1189. дои : 10.1038/mp.2009.84 . ПМК 2990188 . ПМИД 19721432 .
- ^ «Факты об инсульте» . Центры по контролю и профилактике заболеваний . 15 марта 2019 года . Проверено 5 декабря 2019 г.
- ^ Ринк С., Ханна С. (май 2011 г.). «МикроРНК в этиологии и патологии ишемического инсульта» . Физиологическая геномика . 43 (10): 521–528. doi : 10.1152/физиологгеномика.00158.2010 . ПМК 3110894 . ПМИД 20841499 .
- ^ Оуян Ю.Б., Старый К.М., Ян Г.И., Гиффард Р. (январь 2013 г.). «микроРНК: инновационные мишени для лечения ишемии головного мозга и инсульта» . Текущие цели по борьбе с наркотиками . 14 (1): 90–101. дои : 10.2174/138945013804806424 . ПМЦ 3673881 . ПМИД 23170800 .
- ^ Jump up to: а б с Левол Дж. М., Нуньес Ю. О., Додд П. Р., Тивари Г. Р., Харрис Р. А., Мэйфилд Р. Д. (ноябрь 2011 г.). «Повышающая регуляция микроРНК в мозгу алкоголиков» . Алкоголизм: клинические и экспериментальные исследования . 35 (11): 1928–37. дои : 10.1111/j.1530-0277.2011.01544.x . ПМК 3170679 . ПМИД 21651580 .
- ^ Тапочик Дж.Д., Соломон М., Фланиган М., Мейнхардт М., Барбье Э., Шанк Дж.Р. и др. (июнь 2013 г.). «Координированная дисрегуляция мРНК и микроРНК в медиальной префронтальной коре крыс после алкогольной зависимости» . Журнал «Фармакогеномика» . 13 (3): 286–96. дои : 10.1038/tpj.2012.17 . ПМЦ 3546132 . ПМИД 22614244 .
- ^ Горини Дж., Нуньес Ю.О., Мэйфилд Р.Д. (2013). «Интеграция микроРНК и профилирование белков выявляет скоординированные нейроадаптации в мозге зависимых от алкоголя мышей» . ПЛОС ОДИН . 8 (12): е82565. Бибкод : 2013PLoSO...882565G . дои : 10.1371/journal.pone.0082565 . ПМЦ 3865091 . ПМИД 24358208 .
- ^ Тапокик Дж.Д., Барбье Э., Фланиган М., Соломон М., Пинкус А., Пиллинг А. и др. (март 2014 г.). «МикроРНК-206 в медиальной префронтальной коре крыс регулирует экспрессию BDNF и употребление алкоголя» . Журнал неврологии . 34 (13): 4581–8. doi : 10.1523/JNEUROSCI.0445-14.2014 . ПМЦ 3965783 . ПМИД 24672003 .
- ^ Липпай Д., Бала С., Чак Т., Курт-Джонс Э.А., Сабо Г. (2013). «Хроническая микроРНК-155, индуцированная алкоголем, способствует нейровоспалению TLR4-зависимым образом у мышей» . ПЛОС ОДИН . 8 (8): е70945. Бибкод : 2013PLoSO...870945L . дои : 10.1371/journal.pone.0070945 . ПМЦ 3739772 . ПМИД 23951048 .
- ^ Jump up to: а б Ли Дж., Ли Дж., Лю Икс, Цинь С., Гуань Ю., Лю Ю. и др. (сентябрь 2013 г.). «Профиль экспрессии микроРНК и функциональный анализ показывают, что миР-382 является важным новым геном алкогольной зависимости» . ЭМБО Молекулярная медицина . 5 (9): 1402–14. дои : 10.1002/emmm.201201900 . ПМК 3799494 . ПМИД 23873704 .
- ^ Ромао Дж. М., Джин В., Додсон М. В., Хаусман Г. Дж., Мур С. С., Гуан Л. Л. (сентябрь 2011 г.). «Регуляция микроРНК в адипогенезе млекопитающих». Экспериментальная биология и медицина . 236 (9): 997–1004. дои : 10.1258/ebm.2011.011101 . ПМИД 21844119 . S2CID 30646787 .
- ^ Скорн М., Намлёс Х.М., Нордхус П., Ван М.Ю., Меза-Сепеда Л.А., Миклебост О. (апрель 2012 г.). «Дифференцировка адипоцитов стромальных клеток костного мозга человека модулируется микроРНК-155, микроРНК-221 и микроРНК-222». Стволовые клетки и развитие . 21 (6): 873–83. дои : 10.1089/scd.2010.0503 . hdl : 10852/40423 . ПМИД 21756067 .
- ^ Цзо Ю, Цян Л, фермер-старший (март 2006 г.). «Активация экспрессии CCAAT/энхансер-связывающего белка (C/EBP) альфа с помощью C/EBP бета во время адипогенеза требует активируемой пролифератором пероксисомы гамма-ассоциированной репрессии HDAC1 на промоторе гена C/ebp альфа» . Журнал биологической химии . 281 (12): 7960–7. дои : 10.1074/jbc.M510682200 . ПМИД 16431920 .
- ^ Джун-Хао ET, Гупта Р.Р., Шых-Чанг Н. (март 2016 г.). «Lin28 и let-7 в метаболической физиологии старения». Тенденции в эндокринологии и обмене веществ . 27 (3): 132–141. дои : 10.1016/j.tem.2015.12.006 . ПМИД 26811207 . S2CID 3614126 .
- ^ Чжу Х., Шых-Чанг Н., Сегре А.В., Шинода Г., Шах С.П., Эйнхорн В.С. и др. (сентябрь 2011 г.). «Ось Lin28/let-7 регулирует метаболизм глюкозы» . Клетка . 147 (1): 81–94. дои : 10.1016/j.cell.2011.08.033 . ПМЦ 3353524 . ПМИД 21962509 .
- ^ Фрост Р.Дж., Олсон Э.Н. (декабрь 2011 г.). «Контроль гомеостаза глюкозы и чувствительности к инсулину с помощью семейства микроРНК Let-7» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (52): 21075–80. Бибкод : 2011PNAS..10821075F . дои : 10.1073/pnas.1118922109 . ПМЦ 3248488 . ПМИД 22160727 .
- ^ Теруэль-Монтойя Р., Розендал Ф.Р., Мартинес К. (февраль 2015 г.). «МикроРНК в гемостазе» . Журнал тромбозов и гемостаза . 13 (2): 170–181. дои : 10.1111/jth.12788 . ПМИД 25400249 .
- ^ Jump up to: а б Нурс Дж., Браун Дж., Лакнер К., Хюттельмайер С., Данквардт С. (ноябрь 2018 г.). «Крупномасштабная идентификация функционального нацеливания микроРНК выявляет совместную регуляцию системы гемостаза» . Журнал тромбозов и гемостаза . 16 (11): 2233–2245. дои : 10.1111/jth.14290 . ПМИД 30207063 .
- ^ Нурс Дж., Данквардт С. (февраль 2021 г.). «Новое обоснование нацеливания на FXI: данные о гемостатическом мишени микроРНК для новых стратегий антикоагулянтов» . Фармакология и терапия . 218 : 107676. doi : 10.1016/j.pharmthera.2020.107676 . ПМИД 32898547 .
- ^ Берарди Э., Пуэс М., Торрес Л., Сампаолеси М. (октябрь 2012 г.). «миРНК в дифференцировке ЭСК». Американский журнал физиологии. Физиология сердца и кровообращения . 303 (8): H931–H939. дои : 10.1152/ajpheart.00338.2012 . ПМИД 22886416 . S2CID 6402014 .
- ^ Jump up to: а б Борчерс А., Пилер Т. (ноябрь 2010 г.). «Программирование плюрипотентных клеток-предшественников, полученных из эмбрионов Xenopus, для создания специфических тканей и органов» . Гены . 1 (3): 413–426. дои : 10.3390/ijms232314755 . ПМК 9740008 . ПМИД 36499082 .
- ^ Кураба Дж., Сприггс А., Тейлор Дж., Ли З., Хелливелл С. (июль 2012 г.). «Регуляция микроРНК на раннем этапе развития семян ячменя» . Биология растений BMC . 12 (1): 120. дои : 10.1186/1471-2229-12-120 . ПМК 3443071 . ПМИД 22838835 .
- ^ Чоудхари А., Кумар А., Каур Х., Каур Н. (май 2021 г.). «МиРНК: надсмотрщик растительного мира». Биология . 76 (5): 1551–1567. Бибкод : 2021Биолг..76.1551С . дои : 10.1007/s11756-021-00720-1 . ISSN 1336-9563 . S2CID 233685660 .
- ^ Вахид С., Цзэн Л. (март 2020 г.). «Критическая роль микроРНК в регуляции времени цветения и развития цветов» . Гены . 11 (3): 319. doi : 10.3390/genes11030319 . ПМК 7140873 . ПМИД 32192095 .
- ^ Вонг CE, Чжао Ю.Т., Ван XJ, Крофт Л., Ван З.Х., Хаэризаде Ф. и др. (май 2011 г.). «МикроРНК в апикальной меристеме побега сои». Журнал экспериментальной ботаники . 62 (8): 2495–2506. дои : 10.1093/jxb/erq437 . hdl : 10536/DRO/DU:30106047 . ПМИД 21504877 .
- ^ Куреши А., Тхакур Н., Монга И., Тхакур А., Кумар М. (1 января 2014 г.). «VIRmiRNA: комплексный ресурс экспериментально подтвержденных вирусных микроРНК и их мишеней» . База данных . 2014 : бау103. дои : 10.1093/база данных/bau103 . ПМК 4224276 . ПМИД 25380780 .
- ^ Кумар М. «ВИРмиРНК» . Ресурс экспериментальных вирусных микроРНК и их мишеней . Центр биоинформатики, ЦНИР-ИМТЕХ.
- ^ Тадденхэм Л., Юнг Дж. С., Чейн-Вун-Минг Б., Долкен Л., Пфеффер С. (февраль 2012 г.). «Глубокое секвенирование малых РНК идентифицирует микроРНК и другие малые некодирующие РНК из вируса герпеса человека 6B» . Журнал вирусологии . 86 (3): 1638–49. дои : 10.1128/JVI.05911-11 . ПМК 3264354 . ПМИД 22114334 .
- ^ Чжэн Х., Фу Р., Ван Дж.Т., Лю Ц., Чэнь Х., Цзян С.В. (апрель 2013 г.). «Достижения в методах прогнозирования мишеней микроРНК» . Международный журнал молекулярных наук . 14 (4): 8179–87. дои : 10.3390/ijms14048179 . ПМЦ 3645737 . ПМИД 23591837 .
- ^ Агарвал В., Белл Г.В., Нам Дж.В., Бартель Д.П. (август 2015 г.). «Прогнозирование эффективных целевых участков микроРНК в мРНК млекопитающих» . электронная жизнь . 4 : e05005. doi : 10.7554/eLife.05005 . ПМЦ 4532895 . ПМИД 26267216 .
Дальнейшее чтение
[ редактировать ]- Определение и классификация микроРНК: Амброс В., Бартель Б., Бартель Д.П., Бердж С.Б., Кэррингтон Дж.К., Чен X и др. (март 2003 г.). «Единая система аннотации микроРНК» . РНК . 9 (3): 277–9. дои : 10.1261/rna.2183803 . ПМЦ 1370393 . ПМИД 12592000 .
- Научный обзор малых РНК: Баулкомб Д. (сентябрь 2002 г.). «ДНК-события. Микрокосм РНК». Наука . 297 (5589): 2002–3. дои : 10.1126/science.1077906 . ПМИД 12242426 . S2CID 82531727 .
- Открытие lin-4 , первой обнаруженной микроРНК: Ли Р.К., Фейнбаум Р.Л., Амброс В. (декабрь 1993 г.). «Гетерохронный ген lin-4 C. elegans кодирует малые РНК, антисмысловые, комплементарные lin-14» . Клетка . 75 (5): 843–54. дои : 10.1016/0092-8674(93)90529-Y . ПМИД 8252621 .
Внешние ссылки
[ редактировать ]- База данных miRBase
- miRTarBase , экспериментально проверенная база данных взаимодействий микроРНК-мишени.
- semirna , веб-приложение для поиска микроРНК в геноме растений.
- ONCO.IO : Интегративный ресурс для анализа микроРНК и факторов транскрипции при раке.
- MirOB Архивировано 4 марта 2014 г. в Wayback Machine : MicroRNA нацелена на базу данных, инструмент для анализа и визуализации данных.
- База данных ChIPBase : база данных с открытым доступом для расшифровки факторов транскрипции , которые участвовали в транскрипции микроРНК или влияли на нее, из данных ChIP-seq.
- Анимированное видео процесса биогенеза микроРНК .
- Реагенты для модуляции микроРНК , обеспечивающие усиление или подавление функции эндогенной зрелой микроРНК
Виды нуклеиновых кислот |
|---|