миР-155
| до меня-155 | |
|---|---|
 Вторичная структура и последовательность pre-mir-155.ard | |
| Идентификаторы | |
| Символ | миР-155 |
| Рфам | RF00731 |
| семейство miRBase | МИПФ0000157 |
| Другие данные | |
| РНК Тип | микроРНК |
| Домен(ы) | Эукариоты ; |
| PDB Структуры | ПДБе |
| МИР155 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MIR155 , MIRN155, миРНК155, мир-155, миР-155, микроРНК 155 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 609337 ; Генные карты : МИР155 ; ОМА : МИР155 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


МиР-155 представляет собой микроРНК , которая у человека кодируется MIR155 хозяином геном- или MIR155HG . [3] МиР-155 играет роль в различных физиологических и патологических процессах. [4] [5] [6] [7] [8] [9] Экзогенный молекулярный контроль in vivo экспрессии миР-155 может ингибировать рост злокачественных опухолей . [10] [11] вирусные инфекции, [12] и ускоряют прогрессирование сердечно-сосудистых заболеваний. [13]
Открытие
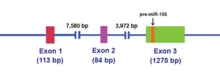
[ редактировать ]Первоначально MIR155HG , был идентифицирован как ген который транскрипционно активировался путем вставки промотора в общий сайт интеграции ретровирусов в В-клеточных лимфомах , и ранее назывался BIC (Кластер интеграции В-клеток). MIR155HG из транскрибируется РНК-полимеразой II , и полученная нуклеотидов РНК кэпируется примерно 1500 и полиаденилируется . Одноцепочечная миР-155 из 23 нуклеотидов, которая находится в экзоне 3, впоследствии процессируется из родительской молекулы РНК. [14]
Биогенез
[ редактировать ]Транскрипт РНК MIR155HG не содержит длинной открытой рамки считывания (ORF), однако он включает стволовую петлю с несовершенным спариванием оснований , которая консервативна у разных видов. [15] Эта некодирующая РНК ( нкРНК ) теперь определяется как первичная микроРНК (при-миРНК). [15] Как только pri-миРНК miR-155 транскрибируется, этот транскрипт расщепляется ядерным микропроцессорным комплексом , основными компонентами которого являются типа РНКазы III эндонуклеаза Drosha и белок критической области 8 DiGeorge ( DGCR8 ). [16] [17] для получения 65-нуклеотидной микроРНК -предшественника «стебель-петля» (pre-mir-155) (см. рисунок 2).

После экспорта из ядра экспортином-5 молекулы pre-mir-155 расщепляются Dicer возле терминальной петли , что приводит к образованию дуплексов РНК длиной ~22 нуклеотида. [16] [17] После расщепления Dicer белок Argonaute (Ago) связывается с короткими дуплексами РНК, образуя ядро многосубъединичного комплекса, называемого комплексом РНК-индуцированного молчания ( RISC ). [18] Аналогично дуплексам миРНК , одна из двух цепей, «миРНК-пассажир» (миР-155*), высвобождается и разрушается, в то время как другая цепь, называемая «направляющей цепью» или «зрелой микроРНК» (миР-155 ), сохраняется в RISC. [18]
Недавние данные подтверждают, что оба плеча шпильки пре-миРНК могут давать начало зрелым микроРНК. [19] [20] В связи с увеличением числа примеров, когда две функциональные зрелые микроРНК процессируются из противоположных плеч одной и той же пре-микроРНК, продукты пре-мир-155 теперь обозначаются суффиксом -5p (от 5'-плеча) (например, миР-155 -5p) и -3p (от 3'-плеча) (например, миР-155-3p) после их названия (см. Рисунок 3). [21]

Как только миР-155-5p/-3p собирается в RISC, эти молекулы впоследствии узнают свою целевую информационную РНК ( мРНК ) посредством взаимодействия пар оснований между нуклеотидами 2 и 8 миР-155-5p/-3p (затравочная область) и комплементарные нуклеотиды преимущественно в 3'-нетранслируемой области ( 3'-UTR ) мРНК (см. рисунки 4 и 5 ниже). [22] Наконец, когда миР-155-5p/-3p действует как адаптер для RISC, связанные с комплексом мРНК подвергаются репрессии трансляции (т.е. ингибированию инициации трансляции ) и/или деградации после деаденилирования . [18]
Эволюционное сохранение
[ редактировать ]Ранний филогенетический анализ показал, что последовательность пре-мир-155 и миР-155-5p консервативна у человека, мыши и курицы. [15] Недавние аннотированные данные секвенирования показали, что 22 различных организма, включая млекопитающих, земноводных, птиц, рептилий, асцидий и морских миног, экспрессируют консервативную миР-155-5p. [1] В настоящее время доступно гораздо меньше данных о последовательностях миР-155-3p, поэтому неясно, насколько консервативна эта миРНК у разных видов. [2]
Распределение тканей
[ редактировать ]Нозерн-блот -анализ показал, что при-миРНК миР-155 обильно экспрессируется в селезенке и тимусе человека и обнаруживается в печени, легких и почках. [15] Анализ последовательностей библиотек малых клонов РНК, сравнивающий экспрессию миРНК со всеми другими исследованными системами органов, установил, что миР-155-5p была одной из пяти миРНК (т.е. миР-142, миР-144 , миР-150 , миР-155 и миР-223). ), который был специфичен для гемопоэтических клеток, включая B-клетки , T-клетки , моноциты и гранулоциты . [23] В совокупности эти результаты позволяют предположить, что миР-155-5p экспрессируется во многих тканях и типах клеток и, следовательно, может играть решающую роль в широком спектре биологических процессов, включая гемопоэз. [4] [5] [6]
Хотя очень мало исследований изучали уровни экспрессии миР-155-3p, Landgraf et al. [23] установили, что уровни экспрессии этой микроРНК в гемопоэтических клетках были очень низкими. Кроме того, ПЦР-анализ показал, что, хотя миР-155-3p обнаруживалась в ряде тканей человека, уровни экспрессии этой миРНК были в 20–200 раз меньше по сравнению с уровнями миР-155-5p. [24] Несмотря на то, что функция миР-155-3p в значительной степени игнорировалась, в настоящее время некоторые исследования показывают, что в некоторых случаях ( астроциты и плазмоцитоидные дендритные клетки ) как миР-155-5p, так и -3p могут функционально созревать из пре-миР-155-3p. 155. [25] [26]
Цели
[ редактировать ]Биоинформатический анализ с использованием TargetScan 6.2 (дата выпуска — июнь 2012 г.) [3] выявил наличие по меньшей мере 4174 предполагаемых мишеней мРНК человеческой miR-155-5p, в общей сложности с 918 консервативными сайтами (т.е. между мышью и человеком) и 4249 плохо консервативными сайтами (т. т.е. только человек). [22] [27] Хотя алгоритм TargetScan 6.2 не может быть использован для определения предполагаемых мишеней miR-155-3p, можно предположить, что эта микроРНК также потенциально может регулировать экспрессию тысяч мишеней мРНК.
Недавно был собран полный список мишеней миР-155-5p/мРНК, которые были экспериментально подтверждены как демонстрацией регуляции эндогенных транскриптов с помощью миР-155-5p, так и проверкой затравочной последовательности миР-155-5p с помощью репортерного анализа. [28] В этот список вошли 140 генов и регуляторные белки миелопоэза и лейкемогенеза (например, SHIP-1 , AICDA , ETS1 , JARID2 , SPI1 и др.), воспаления (например, BACH1 , FADD , IKBKE , INPP5D , MYD88 , RIPK1 , SPI1 , SOCS , и т. д.) и известные супрессоры опухолей (например, CEBPβ , IL17RB , PCCD4 , TCF12 , ZNF652 и т. д.). [28] Подтвержденный сайт связывания миР-155-5p, содержащийся в мРНК SPI1. [29] и подтвержденный сайт связывания miR-155-3p, содержащийся в мРНК IRAK3. [26] показаны на рисунках 4 и 5 соответственно.


Физиологические роли
[ редактировать ]Кроветворение
[ редактировать ]Гематопоэз определяется как образование и развитие клеток крови, все из которых происходят из гемопоэтических стволовых клеток-предшественников (HSPC). [30] HSPC представляют собой примитивные клетки, способные к самообновлению и первоначально дифференцируются в клетки -общие миелоидные предшественники (CMP) или общие лимфоидные клетки-предшественники (CLP). [30] CMP представляют собой клеточную популяцию, которая стала миелоидной линией, и это момент, когда начинается миелопоэз . [30] В ходе миелопоэза происходит дальнейшая дифференцировка клеток, включая тромбопоэз , эритропоэз , гранулопоэз и моноцитопоэз . [30] CLP впоследствии дифференцируются в B-клетки и T-клетки в процессе, называемом лимфопоэзом . [30] Учитывая, что миР-155-5p экспрессируется в гемопоэтических клетках [23] была выдвинута гипотеза, что эта микроРНК играет критическую роль в этих процессах клеточной дифференцировки. В подтверждение этой предпосылки было обнаружено, что миР-155-5p экспрессируется в CD34(+) HSPC человека, и было высказано предположение, что эта миРНК может удерживать эти клетки на ранней стадии ствола-предшественника, ингибируя их дифференцировку в более зрелые клетки. клетка (т.е. мегакариоцитарная/эритроидная/гранулоцитарная/моноцитарная/В-лимфоидная/Т-лимфоидная). [31] Эта гипотеза была подтверждена, когда трансдуцированные pre-mir-155 HSPC генерировали в 5 раз меньше миелоидных и в 3 раза меньше эритроидных колоний. [31] Кроме того, Ху и др. [32] продемонстрировали, что гомеобоксный белок HOXA9 регулирует экспрессию MIR155HG в миелоидных клетках и что эта микроРНК играет функциональную роль в гемопоэзе. Эти исследователи обнаружили, что принудительная экспрессия миР-155-5p в клетках костного мозга приводила к снижению SPI1 (т.е. PU.1) примерно на 50%. [32] фактор транскрипции и регулятор миелопоэза, [33] и подтвержденная мишень этой микроРНК. [29] Установлено также, что дифференцировка in vitro очищенных эритроидных клеток-предшественников человека приводит к прогрессивному снижению экспрессии миР-155-5p в зрелых эритроцитах. [34] Кроме того, у мышей с дефицитом pre-mir-155 наблюдались явные дефекты развития лимфоцитов и генерации B- и T-клеточных ответов in vivo . [29] [35] [36] Наконец, было установлено, что для развития регуляторных Т-клеток ( Treg ) требуется миР-155-5p, и было показано, что эта миРНК играет роль в гомеостазе Treg и общей выживаемости путем прямого воздействия на SOCS1 , негативный регулятор передачи сигналов IL-2 . [37] [38] В совокупности эти результаты убедительно свидетельствуют о том, что миР-155-5p является важной молекулой в контроле нескольких аспектов гемопоэза, включая миелопоэз, эритропоэз и лимфопоэз.
Иммунная система
[ редактировать ]Врожденная иммунная система представляет собой первую линию защиты от проникновения патогенов и считается основным инициатором воспалительных реакций. [39] Его клеточный компонент включает в себя в первую очередь моноциты / макрофаги , гранулоциты и дендритные клетки (DC), которые активируются при распознавании консервативных структур патогена ( PAMP ) с помощью рецепторов распознавания образов, таких как Toll-подобные рецепторы ((TLR)). [40] Экспрессия MIR155HG (т.е. miR-155-5p) значительно усиливается за счет стимуляции агонистами TLR макрофагов и дендритных клеток. [41] [42] [43] [44] [45] [46] Поскольку микробный липополисахарид (агонист TLR4 ) активирует цепочку событий, которые приводят к стимуляции факторов транскрипции NF-κB и AP-1 , [40] было высказано предположение, что эндотоксина активация MIR155HG может быть опосредована этими факторами транскрипции. [41] Действительно, было обнаружено, что экспрессия MIR155HG активируется в обработанных LPS мышиных макрофагах (т.е. Raw264.7) по механизму, опосредованному NF-κB. [42] Кроме того, инфицирование H. pylori первичных мышиных макрофагов, полученных из костного мозга, приводило к NF-κB-зависимой активации MIR155HG . [47] Сообщалось , что в контексте вирусной инфекции вирус везикулярного стоматита (VSV) заражение мышиных перитонеальных макрофагов приводило к сверхэкспрессии миР-155-5p посредством индуцируемого ретиноевой кислотой гена I /JNK/NF-κB-зависимого пути. [48] Подтверждение роли AP-1 в активации MIR155HG получено в исследованиях с использованием стимулов, связанных с вирусной инфекцией, таких как поли(I:C) лиганда TLR3 или бета-интерферон (IFN-β). [43] Ниже этих стимулов AP-1, по-видимому, играет важную роль в активации MIR155HG . [43] [49] [50] [51]
При его инициировании путем активации, например, TLRs патогенными стимулами, миР-155-5p действует как посттранскрипционный регулятор сигнальных путей врожденного иммунитета. Важно отметить, что миР-155-5p демонстрирует такую же реакцию на патогенные стимулы (например, агонист TLR4 ЛПС), что и мРНК основного провоспалительного маркера. [52] После активации миР-155-5p подавляет негативные регуляторы воспаления. К ним относятся инозитолполифосфат-5-фосфатаза (INPP5D, также обозначаемая SHIP1) и супрессор передачи сигналов цитокина 1 (SOCS1), подавление которого способствует выживанию клеток, росту, миграции и антипатогенным реакциям. [48] [53] [54] [55] Помимо поддержки активации защитных путей, миР-155-5p может также ограничивать силу результирующей NF-κB-зависимой воспалительной реакции, [52] что позволяет предположить различные функции миР-155 на разных стадиях воспаления.
В совокупности эти наблюдения подразумевают, что активация MIR155HG может быть контекстно-зависимой, учитывая, что как AP-1-, так и NF-κB-опосредованные механизмы регулируют экспрессию этого гена. Эти исследования также предполагают, что широкий спектр вирусных и бактериальных медиаторов воспаления могут стимулировать экспрессию миР-155-5p и указывают на тесную связь между воспалением, врожденным иммунитетом и экспрессией MIR155HG .
Активность и фенотипы
[ редактировать ]Имеются доказательства того, что миР-155 участвует в каскадах, связанных с сердечно-сосудистыми заболеваниями и гипертонией, а также было обнаружено, что она участвует в иммунитете, нестабильности генома клеток , дифференцировке , воспалении, вирус-ассоциированных инфекциях, раке и сахарном диабете . [56]
Защитная роль миР-155 может возникать в ответ на ее действие на молчание генов, тем самым регулируя время их экспрессии; мутации в сайте-мишени миР-155 лишают ее оптимального доступа, необходимого для подавления генов, что приводит к чрезмерному количеству нарушений активности, которые могут например , роль миР-155 как защитного агента против предрасположенности к злокачественным новообразованиям , связанным с В-клетками, подчеркивается за счет поддержания баланса фермента индуцируемой активацией цитидиндезаминазы ( AID ). МиР-155 опосредует регуляцию численности AID и времени экспрессии по иммунологическим сигналам, однако мутации в мишени мРНК AID приводят к ее невосприимчивости к молчанию миР-155 и приводят к необузданной экспрессии ее белка, вызывая всплески диких незрелых B-лимфоцитов и AID. опосредованные хромосомные транслокации . [5] [6]
Клиническое значение
[ редактировать ]Сердечно-сосудистая система
[ редактировать ]Трансфекция миР-155 в первичные фибробласты легких человека снижает эндогенную экспрессию ангиотензина II рецептора белка AT1R . Кроме того, AT1R опосредует повышение артериального давления, связанное с ангиотензином II, и способствует патогенезу сердечной недостаточности. Дефектная функция миР-155 может быть связана с гипертонией и сердечно-сосудистыми заболеваниями, если цис-регуляторный сайт на 3'-UTR AT1R (мишень-сайт миР-155) был затронут из-за полиморфизма SNP в самом AT1R. Эта мутация нарушает нацеливание на миР-155 и, таким образом, предотвращает снижение экспрессии AT1R. [5] При низком артериальном давлении сверхэкспрессия миР-155 коррелирует с нарушением активности AT1R. [4]
Иммунитет
[ редактировать ]миР-155 участвует в иммунитете, играя ключевую роль в модуляции гуморальных и врожденных клеточно-опосредованных иммунных ответов, например. У мышей с дефицитом миР-155 нарушается иммунологическая память; делая его жертвой повторяющихся приступов инвазий одного и того же патогена (Rodriguez et al. 2007), созревание и специфичность В-лимфоцитов с дефицитом миР-155 нарушаются, поскольку этот процесс зависит от фермента AID, который имеет мишень миР-155 в его 3'-конец UTR. [5] [6] Фенотипические последствия, связанные с дефицитом миР-155 у мышей, проявляются в более позднем возрасте, когда у животных развиваются поражения легких и кишечника . [4]
Активированные В- и Т-клетки демонстрируют повышенную экспрессию миР-155, то же самое касается макрофагов и дендритных клеток иммунной системы . МиР-155 имеет решающее значение для правильного развития и созревания лимфоцитов. Детали различных проявлений уровней миР-155 и участия в деятельности, обеспечивающей оптимальные иммунные реакции, были предметом многих исследований:
Снижение IgG1
[ редактировать ]Дефектные Т- и В-клетки, а также заметно сниженные ответы IgG1 наблюдались у мышей с дефицитом miR-155, уровень IgG1 снижается, тогда как экспрессия иммуноглобулина IgM у этих мышей остается нормальной. Изменение уровней IgG1 можно объяснить тем фактом, что он является мишенью для миР-155 в В-клетках, белка, кодирующего мРНК регулятора транскрипции белка Pu.1 , повышение уровня белка Pu.1 предрасполагает к дефектной продукции IgG1. Помимо Pu.1, в В-клетках с дефицитом miR-155 имеется около 60 других генов с дифференциальным уровнем повышенного уровня, дальнейшее исследование выявило возможные сайты-мишени miR-155 в 3'-UTR-областях этих генов. [6]
Злокачественные новообразования лимфоцитов
[ редактировать ]зрелых рецепторов Сродство и специфичность лимфоцитов к патогенным агентам лежат в основе правильных иммунных ответов, оптимальная координация миР-155 необходима для производства нормальных В-лимфоцитов, продукции высокоаффинных антител и балансировки передачи сигналов BCR. Было продемонстрировано, что миР-155 может передаваться через щелевые контакты от лейкозных клеток к здоровым В-клеткам и способствовать их трансформации в онкогенноподобные клетки. [57]
Отбор компетентных В-клеток происходит в зародышевом центре , где они обучаются дифференцировать клетки организма от чужеродных антигенов, они конкурируют за распознавание антигена и за помощь Т-клеток, таким образом оказывая селективное давление на те В-клетки, которые продемонстрировали высокоаффинные рецепторы. и сотрудничество с Т-клетками ( созревание аффинности ) рекрутируются и развертываются в костном мозге или становятся В-клетками памяти, апоптозное прекращение происходит для тех В-клеток, которые терпят неудачу в конкуренции. Незрелые В-клетки с дефицитом миР-155 уклоняются от апоптоза в результате повышенных уровней белка Bcl-2 ; белок, который, как было обнаружено, участвует в злокачественных новообразованиях В-клеток и контролируется миР-155. [6]
Воспаление
[ редактировать ]Воспалительные реакции на триггеры, такие как TNF-α, включают макрофаги с компонентами, включающими миР-155. миР-155 сверхэкспрессируется при атопическом дерматите и способствует хроническому воспалению кожи за счет усиления пролиферативного ответа Т(Н)-клеток за счет подавления CTLA-4. [58] При аутоиммунных заболеваниях, таких как ревматоидный артрит, миР-155 демонстрирует более высокую экспрессию в тканях пациентов и синовиальных фибробластах. [4] При рассеянном склерозе повышенная экспрессия mir-155 также была измерена в периферических и резидентных миелоидных клетках ЦНС, включая циркулирующие моноциты крови и активированную микроглию. [59] Также было обнаружено, что мир-155 участвует в воспалении. Сверхэкспрессия мир-155 приведет к хроническому воспалительному состоянию у человека. [60]
ДНК-вирусы
[ редактировать ]У ДНК-вирусов микроРНК подтверждены экспериментально, микроРНК у вирусов кодируются дцДНК, [5] примеры таких вирусов включают герпесвирусы, такие как вирус человека-Эпштейна-Барра ( EBV ) и аденовирусы , [4] другим вирусом, экспрессирующим микроРНК, подобную миР-155, у кур, является онкогенный MDV-1, неонкогенный родственник которого MDV-2 этого не делает, это предполагает участие миР-155 в лимфомагенезе. [5] Вирусы могут использовать микроРНК хозяина в той степени, в которой они используют микроРНК хозяина для кодирования вирусных клонов, например:миР-K12-11 в герпесвирусе, ассоциированном с саркомой Капоши, имеет область целевой специфичности , ортологичную области миР-155; имитируя действие миР-155 [61] и, разделяя с ним мишени, можно считать, что он подавляет доступность миР-155 к своим мишеням путем конкуренции, и это фактически подавляет экспрессию генов, играющих роль в клеточном росте и апоптозе, таким образом, который игнорирует регулирование со стороны миР-155. [4] ВЭБ модулирует экспрессию миР-155 хозяина, которая необходима для роста В-клеток, инфицированных ВЭБ. [62] Клетки, инфицированные EBV, имеют повышенную экспрессию миР-155, тем самым нарушая равновесие экспрессии генов, регулирующих транскрипцию в этих клетках. [4] [5]
Рак
[ редактировать ]Чрезмерное подавление миР-155 может привести к запуску онкогенных каскадов, которые начинаются с апоптотической устойчивости, проапоптотический ядерный белок 1, индуцированный опухолевым белком-53 ( TP53INP1 ), подавляется миР-155, сверхэкспрессия миР-155 приводит к снижению уровней TP53INP1 при аденокарциномах протоков поджелудочной железы и, возможно, при других эпителиальных раковых заболеваниях, где активность TP53INP1 теряется, что приводит к уклонению от апоптоза и неконтролируемым приступам роста. [5]
Инактивация процесса восстановления несоответствия ДНК ( MMR ), выявленная по повышению частоты мутаций, является причиной синдрома Линча (LS), также известного как наследственный неполипозный колоректальный рак (HNPCC). Понижение регуляции белка, контролирующего MMR, осуществляется за счет сверхэкспрессии. В случае миР-155 MMR контролируется группой консервативных белков, снижение активности этих белков приводит к повышенному уровню мутаций в фенотипе, запуская путь к развитию этого типа рака. [63]
Другие типы опухолей, в которых сообщалось о сверхэкспрессии миР-155, включают: карциному щитовидной железы, рак молочной железы, рак толстой кишки, рак шейки матки и рак легких, где количественное определение различных профилей экспрессии миР-155 потенциально может служить сигналом для обнаружения и оценки опухоли. прогноза исхода. [4] Анализ показал, что экспрессия миР-155 связана с выживаемостью при трижды негативном раке молочной железы. [64]
Примечания
[ редактировать ]Версия этой статьи 2012 года была обновлена внешним экспертом в рамках модели двойной публикации. Соответствующая академическая рецензируемая статья была опубликована в журнале Gene и может цитироваться как: Терри С. Элтон, Хелина Селемон, Шейн М. Элтон, Нарасимхам Л. Паринанди (14 декабря 2012 г.). «Регуляция гена хозяина MIR155 в физиологических и патологических процессах». Джин . 532 (1): 1–12. дои : 10.1016/J.GENE.2012.12.009 . ISSN 0378-1119 . ПМИД 23246696 . Викиданные Q34318174 . |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000283904 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Энтрез Ген: MIR155HG» .
- ^ Jump up to: а б с д и ж г час я Фараони I, Антонетти ФР, Кардоне Дж, Бонмассар Э (июнь 2009 г.). «Ген миР-155: типичная многофункциональная микроРНК» (PDF) . Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1792 (6): 497–505. дои : 10.1016/j.bbadis.2009.02.013 . ПМИД 19268705 .
- ^ Jump up to: а б с д и ж г час я Тенг Дж., Папавасилиу Ф.Н. (март 2009 г.). «Шшш! Замалчивание микроРНК-155» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1517): 631–637. дои : 10.1098/rstb.2008.0209 . ПМК 2660923 . ПМИД 19008191 .
- ^ Jump up to: а б с д и ж Каламе К. (декабрь 2007 г.). «Функция микроРНК-155 в В-клетках» . Иммунитет . 27 (6): 825–827. doi : 10.1016/j.immuni.2007.11.010 . ПМИД 18093533 .
- ^ Тили Э, Кроче СМ, Мишель Дж.Дж. (2009). «МиР-155: о взаимосвязи между воспалением и раком». Международные обзоры иммунологии . 28 (5): 264–284. дои : 10.1080/08830180903093796 . ПМИД 19811312 . S2CID 205589961 .
- ^ О'Коннелл Р.М., Рао Д.С., Балтимор Д. (2012). «регуляция микроРНК воспалительных реакций». Ежегодный обзор иммунологии . 30 : 295–312. doi : 10.1146/annurev-immunol-020711-075013 . ПМИД 22224773 .
- ^ Элтон Т.С., Селемон Х, Элтон С.М., Паринанди Н.Л. (декабрь 2013 г.). «Регуляция гена хозяина MIR155 в физиологических и патологических процессах». Джин . 532 (1): 1–12. дои : 10.1016/j.gene.2012.12.009 . ПМИД 23246696 .
- ^ Маттиске С., Суэтани Р.Дж., Нильсен П.М., Каллен Д.Ф. (август 2012 г.). «Онкогенная роль миР-155 при раке молочной железы» . Эпидемиология рака, биомаркеры и профилактика . 21 (8): 1236–1243. doi : 10.1158/1055-9965.EPI-12-0173 . hdl : 2440/73553 . ПМИД 22736789 .
- ^ Бабар И.А., Ченг С.Дж., Бут С.Дж., Лян Х, Вайдхас Дж.Б., Зальцман В.М., Слэк Ф.Дж. (июнь 2012 г.). «Терапия на основе наночастиц в мышиной модели лимфомы, зависимой от микроРНК-155 (миР-155) in vivo» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (26): E1695–1704. дои : 10.1073/pnas.1201516109 . ПМК 3387084 . ПМИД 22685206 .
- ^ Ван Л., Туми Н.Л., Диас Л.А., Уокер Дж., Рамос Дж.К., Барбер Г.Н., Нин С. (август 2011 г.). «Онкогенные IRF обеспечивают преимущество в выживании клеток, трансформированных вирусом Эпштейна-Барра или вирусом Т-клеточного лейкоза человека типа 1, за счет индукции экспрессии BIC» . Журнал вирусологии . 85 (16): 8328–8337. дои : 10.1128/JVI.00570-11 . ПМК 3147954 . ПМИД 21680528 .
- ^ Корстен М.Ф., Папагеоргиу А., Верхесен В., Караи П., Линдоу М., Обад С., Саммер Дж., Коорт С.Л., Хазебрук М., ван Леувен Р., Гейбельс М.Дж., Вейнандс Е., Биссен Э.А., Де Винтер М.П., Стассен Ф.Р., Кармелиет П., Кауппинен С., Шроен Б., Хейманс С. (август 2012 г.). «Профилирование микроРНК идентифицирует микроРНК-155 как неблагоприятный медиатор сердечного повреждения и дисфункции во время острого вирусного миокардита» . Исследование кровообращения . 111 (4): 415–425. дои : 10.1161/CIRCRESAHA.112.267443 . ПМИД 22715471 .
- ^ Эйс П.С., Тэм В., Сан Л., Чадберн А., Ли З., Гомес М.Ф., Лунд Э., Дальберг Дж.Э. (март 2005 г.). «Накопление миР-155 и BIC РНК в В-клеточных лимфомах человека» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (10): 3627–3632. Бибкод : 2005PNAS..102.3627E . дои : 10.1073/pnas.0500613102 . ПМК 552785 . ПМИД 15738415 .
- ^ Jump up to: а б с д Тэм В. (август 2001 г.). «Идентификация и характеристика человеческого BIC, гена на хромосоме 21, который кодирует некодирующую РНК». Джин . 274 (1–2): 157–167. дои : 10.1016/S0378-1119(01)00612-6 . ПМИД 11675008 .
- ^ Jump up to: а б Ким В.Н., Хан Дж., Сиоми MC (февраль 2009 г.). «Биогенез малых РНК у животных». Nature Reviews Молекулярно-клеточная биология . 10 (2): 126–139. дои : 10.1038/nrm2632 . ПМИД 19165215 . S2CID 8360619 .
- ^ Jump up to: а б Крол Дж., Ледидж И., Филипович В. (сентябрь 2010 г.). «Широкая регуляция биогенеза, функции и распада микроРНК». Обзоры природы Генетика . 11 (9): 597–610. дои : 10.1038/nrg2843 . ПМИД 20661255 . S2CID 2619579 .
- ^ Jump up to: а б с Фабиан М.Р., Соненберг Н. (июнь 2012 г.). «Механика молчания генов, опосредованного микроРНК: взгляд под капот miRISC» . Структурная и молекулярная биология природы . 19 (6): 586–593. дои : 10.1038/nsmb.2296 . ПМИД 22664986 . S2CID 13176647 .
- ^ Бушати Н., Коэн С.М. (2007). «функции микроРНК». Ежегодный обзор клеточной биологии и биологии развития . 23 : 175–205. doi : 10.1146/annurev.cellbio.23.090506.123406 . ПМИД 17506695 .
- ^ Филипович В., Бхаттачарья С.Н., Соненберг Н. (февраль 2008 г.). «Механизмы посттранскрипционной регуляции микроРНК: ответы видны?» . Обзоры природы Генетика . 9 (2): 102–114. дои : 10.1038/nrg2290 . ПМИД 18197166 . S2CID 11824239 .
- ^ Гриффитс-Джонс С. (январь 2004 г.). «Реестр микроРНК» . Исследования нуклеиновых кислот . 32 (Проблема с базой данных): D109–11. дои : 10.1093/nar/gkh023 . ПМК 308757 . ПМИД 14681370 .
- ^ Jump up to: а б Фридман Р.К., Фарх К.К., Бердж CB, Бартель Д.П. (январь 2009 г.). «Большинство мРНК млекопитающих являются консервативными мишенями микроРНК» . Геномные исследования . 19 (1): 92–105. дои : 10.1101/гр.082701.108 . ПМЦ 2612969 . ПМИД 18955434 .
- ^ Jump up to: а б с Ландграф П., Русу М., Шеридан Р., Канализатор А., Иовино Н., Аравин А., Пфеффер С., Райс А., Камфорст А.О., Ландталер М., Лин С., Соччи Н.Д., Эрмида Л., Фульчи В., Кьяретти С., Фоа Р., Шливка Дж. , Фукс Ю, Новосел А, Мюллер Р.У., Шермер Б., Биссельс У., Инман Дж., Фан К., Чиен М., Вейр Д.Б., Чокси Р., Де Вита Г., Фреццетти Д., Тромпетер Х.И., Хорнунг В., Тенг Г., Хартманн Г., Палковиц М, Ди Лауро Р, Вернет П, Мачино Г, Роглер С.Э., Нэгл Дж.В., Джу Дж., Папавасилиу Ф.Н., Бенцинг Т., Лихтер П., Тэм В., Браунштейн М.Дж., Босио А., Боркхардт А., Руссо Дж.Дж., Сандер С., Заволан М., Тушл Т. (июнь 2007 г.). «Атлас экспрессии микроРНК млекопитающих, основанный на секвенировании небольшой библиотеки РНК» . Клетка . 129 (7): 1401–1414. дои : 10.1016/j.cell.2007.04.040 . ПМЦ 2681231 . ПМИД 17604727 .
- ^ Элтон Т.С., Сэнсом С.Э., Мартин М.М. (2010). «Сверхэкспрессия микроРНК в дозе гена трисомии-21 приводит к гаплонедостаточности специфических белков-мишеней» . Биология РНК . 7 (5): 540–547. дои : 10.4161/rna.7.5.12685 . ПМК 3073250 . ПМИД 21081842 .
- ^ Тарассишин Л., Лаудиг О., Бауман А., Шафит-Загардо Б., Су Х.С., Ли С.К. (декабрь 2011 г.). «Регуляторный фактор интерферона 3 ингибирует экспрессию воспалительных генов астроцитов посредством подавления провоспалительных миР-155 и миР-155*» . Глия . 59 (12): 1911–1922. дои : 10.1002/glia.21233 . ПМК 3241213 . ПМИД 22170100 .
- ^ Jump up to: а б Чжоу Х, Хуан Х, Цуй Х, Луо Х, Тан Ю, Чен С, Ву Л, Шен Н (декабрь 2010 г.). «МиР-155 и ее звездообразный партнер миР-155* совместно регулируют выработку интерферона типа I плазмоцитоидными дендритными клетками человека» . Кровь . 116 (26): 5885–5894. дои : 10.1182/кровь-2010-04-280156 . ПМИД 20852130 .
- ^ Льюис Б.П., Бердж CB, Бартель Д.П. (январь 2005 г.). «Консервативные пары семян, часто окруженные аденозинами, указывают на то, что тысячи человеческих генов являются мишенями микроРНК» . Клетка . 120 (1): 15–20. дои : 10.1016/j.cell.2004.12.035 . ПМИД 15652477 . S2CID 17316349 .
- ^ Jump up to: а б Нильсен П.М., Нолл Дж.Э., Маттиске С., Бракен С.П., Грегори П.А., Шульц Р.Б., Лим С.П., Кумар Р., Суэтани Р.Дж., Гудолл Г.Дж., Каллен Д.Ф. (июнь 2013 г.). «Мутантный р53 вызывает инвазию в опухоли молочной железы за счет повышения регуляции миР-155» . Онкоген . 32 (24): 2992–3000. дои : 10.1038/onc.2012.305 . hdl : 2440/76596 . ПМИД 22797073 .
- ^ Jump up to: а б с Вигорито Э., Перкс К.Л., Абреу-Гуджер С., Бантинг С., Сян З., Кольхас С., Дас П.П., Миска Э.А., Родригес А., Брэдли А., Смит К.Г., Рада С., Энрайт А.Дж., Тёлльнер К.М., Макленнан И.С., Тернер М. ( декабрь 2007 г.). «МикроРНК-155 регулирует образование плазматических клеток с переключенным классом иммуноглобулинов» . Иммунитет . 27 (6): 847–859. doi : 10.1016/j.immuni.2007.10.009 . ПМЦ 4135426 . ПМИД 18055230 .
- ^ Jump up to: а б с д и Маяни Х (март 2010 г.). «Биологические различия между неонатальными и взрослыми гемопоэтическими стволовыми клетками / клетками-предшественниками человека». Стволовые клетки и развитие . 19 (3): 285–298. дои : 10.1089/scd.2009.0327 . ПМИД 19778207 .
- ^ Jump up to: а б Георгантас Р.В., Хилдрет Р., Моризо С., Алдер Дж., Лю К.Г., Хаймфельд С., Калин Г.А., Кроче СМ, Сивин CI (февраль 2007 г.). «Экспрессия и функция микроРНК гемопоэтических стволовых клеток-предшественников CD34 +: принципиальная схема контроля дифференцировки» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (8): 2750–2755. Бибкод : 2007PNAS..104.2750G . дои : 10.1073/pnas.0610983104 . ПМЦ 1796783 . ПМИД 17293455 .
- ^ Jump up to: а б Ху Ю.Л., Фонг С., Ларгман С., Шен В.Ф. (сентябрь 2010 г.). «НОХА9 регулирует миР-155 в кроветворных клетках» . Исследования нуклеиновых кислот . 38 (16): 5472–5478. дои : 10.1093/нар/gkq337 . ПМЦ 2938212 . ПМИД 20444872 .
- ^ Кастнер П., Чан С. (2008). «PU.1: решающий и универсальный игрок в области кроветворения и лейкемии». Международный журнал биохимии и клеточной биологии . 40 (1): 22–27. doi : 10.1016/j.biocel.2007.01.026 . ПМИД 17374502 .
- ^ Масаки С., Оцука Р., Абэ Ю., Мута К., Умемура Т. (декабрь 2007 г.). «Схемы экспрессии микроРНК 155 и 451 во время нормального эритропоэза человека». Связь с биохимическими и биофизическими исследованиями . 364 (3): 509–514. дои : 10.1016/j.bbrc.2007.10.077 . ПМИД 17964546 .
- ^ Родригес А., Вигорито Э., Клэр С., Уоррен М.В., Кутте П., Сунд Д.Р., ван Донген С., Грокок Р.Дж., Дас П.П., Миска Э.А., Ветри Д., Оккенхауг К., Энрайт Эй.Дж., Дуган Г., Тернер М., Брэдли А. (апрель). 2007). «Потребность в bic/микроРНК-155 для нормальной иммунной функции» . Наука . 316 (5824): 608–611. Бибкод : 2007Sci...316..608R . дои : 10.1126/science.1139253 . ПМК 2610435 . ПМИД 17463290 .
- ^ Тай Т.Д., Каладо Д.П., Касола С., Ансель К.М., Сяо С., Сюэ Ю., Мерфи А., Френдьюи Д., Валенсуэла Д., Куток Дж.Л., Шмидт-Супприан М., Раевски Н., Янкопулос Г., Рао А., Раевски К. (апрель 2007 г.) . «Регуляция ответа зародышевого центра с помощью микроРНК-155». Наука . 316 (5824): 604–608. Бибкод : 2007Sci...316..604T . дои : 10.1126/science.1141229 . ПМИД 17463289 . S2CID 8174458 .
- ^ Кольхас С., Гарден О.А., Скадамор К., Тернер М., Оккенхауг К., Вигорито Э (март 2009 г.). «Авангард: миР-155-мишень Foxp3 способствует развитию регуляторных Т-клеток» . Журнал иммунологии . 182 (5): 2578–2582. doi : 10.4049/jimmunol.0803162 . ПМИД 19234151 .
- ^ Лу Л.Ф., Тай Т.Х., Каладо Д.П., Чаудри А., Кубо М., Танака К., Леб ГБ, Ли Х., Йошимура А., Раевски К., Руденский А.Ю. (январь 2009 г.). «Foxp3-зависимая микроРНК155 придает конкурентоспособность регуляторным Т-клеткам путем нацеливания на белок SOCS1» . Иммунитет . 30 (1): 80–91. doi : 10.1016/j.immuni.2008.11.010 . ПМЦ 2654249 . ПМИД 19144316 .
- ^ Меджитов Р. (март 2010 г.). «Воспаление 2010: новые приключения старого флейма» . Клетка . 140 (6): 771–776. дои : 10.1016/j.cell.2010.03.006 . ПМИД 20303867 . S2CID 10297400 .
- ^ Jump up to: а б Такеда К., Акира С. (январь 2005 г.). «Толл-подобные рецепторы врожденного иммунитета». Международная иммунология . 17 (1): 1–14. дои : 10.1093/intimm/dxh186 . ПМИД 15585605 .
- ^ Jump up to: а б Таганов К.Д., Болдин М.П., Чанг К.Дж., Балтимор Д. (август 2006 г.). «NF-kappaB-зависимая индукция микроРНК миР-146, ингибитора, направленного на сигнальные белки врожденного иммунного ответа» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12481–12486. Бибкод : 2006PNAS..10312481T . дои : 10.1073/pnas.0605298103 . ПМК 1567904 . ПМИД 16885212 .
- ^ Jump up to: а б Тили Э, Мишель Дж.Дж., Чимино А., Костинян С., Думитру К.Д., Адэр Б., Фаббри М., Алдер Х., Лю К.Г., Калин Г.А., Кроче СМ (октябрь 2007 г.). «Модуляция уровней миР-155 и миР-125b после стимуляции липополисахаридом/ФНО-альфа и их возможная роль в регуляции ответа на эндотоксиновый шок» . Журнал иммунологии . 179 (8): 5082–5089. дои : 10.4049/jimmunol.179.8.5082 . ПМИД 17911593 .
- ^ Jump up to: а б с О'Коннелл Р.М., Таганов К.Д., Болдин М.П., Ченг Г., Балтимор Д. (январь 2007 г.). «МикроРНК-155 индуцируется во время воспалительной реакции макрофагов» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (5): 1604–1609. Бибкод : 2007PNAS..104.1604O . дои : 10.1073/pnas.0610731104 . ПМК 1780072 . ПМИД 17242365 .
- ^ Чеппи М., Перейра П.М., Дюнан-Сотье I, Баррас Э., Рейт В., Сантос М.А., Пьер П. (февраль 2009 г.). «МикроРНК-155 модулирует сигнальный путь интерлейкина-1 в активированных дендритных клетках, происходящих из моноцитов человека» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (8): 2735–2740. Бибкод : 2009PNAS..106.2735C . дои : 10.1073/pnas.0811073106 . ПМК 2650335 . ПМИД 19193853 .
- ^ Кремер Т.Дж., Равнеберг Д.Х., Клэй CD, Пайпер-Хантер М.Г., Марш CB, Элтон Т.С., Ганн Дж.С., Амер А., Каннеганти Т.Д., Шлезингер Л.С., Бучар Дж.П., Тридандапани С. (2009). «Индукция МиР-155 F. novicida, но не вирулентным F. tularensis, приводит к снижению регуляции SHIP и усилению провоспалительной цитокиновой реакции» . ПЛОС ОДИН . 4 (12): е8508. Бибкод : 2009PLoSO...4.8508C . дои : 10.1371/journal.pone.0008508 . ПМЦ 2794384 . ПМИД 20041145 .
- ^ Мао КП, Хэ Л, Цай Ю.К., Пэн С., Кан Т.Х., Пан Х, Мони А., Хунг К.Ф., Ву TC (2011). «Экспрессия микроРНК-155 in vivo влияет на антигенспецифические Т-клеточные иммунные реакции, генерируемые ДНК-вакцинацией» . Клетка и биологические науки . 1 (1): 3. дои : 10.1186/2045-3701-1-3 . ПМК 3116247 . ПМИД 21711593 .
- ^ Кох М., Молленкопф Х.Дж., Клемм У., Мейер Т.Ф. (май 2012 г.). «Индукция микроРНК-155 зависит от системы секреции TLR и типа IV в макрофагах и ингибирует апоптоз, вызванный повреждением ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (19): E1153–1162. дои : 10.1073/pnas.1116125109 . ПМЦ 3358876 . ПМИД 22509021 .
- ^ Jump up to: а б Ван П, Хоу Дж, Линь Л, Ван С, Лю Х, Ли Д, Ма Ф, Ван З, Цао X (ноябрь 2010 г.). «Индуцируемая обратная связь микроРНК-155 способствует передаче сигналов IFN типа I при противовирусном врожденном иммунитете путем нацеливания на супрессор передачи сигналов цитокинов 1» . Журнал иммунологии . 185 (10): 6226–6233. doi : 10.4049/jimmunol.1000491 . ПМИД 20937844 .
- ^ Тили Э, Мишель Дж. Дж., Адэр Б., Алдер Х., Лимань Е., Таччоли С., Феррацин М., Дельмас Д., Латруфф Н., Кроче СМ (сентябрь 2010 г.). «Ресвератрол снижает уровень миР-155 за счет активации миР-663, микроРНК, нацеленной на JunB и JunD» . Канцерогенез . 31 (9): 1561–1566. дои : 10.1093/carcin/bgq143 . ПМЦ 4647642 . ПМИД 20622002 .
- ^ Маккой С.Э., Шиди Ф.Дж., Куаллс Дж.Э., Дойл С.Л., Куинн С.Р., Мюррей П.Дж., О'Нил Л.А. (июль 2010 г.). «IL-10 ингибирует индукцию миР-155 толл-подобными рецепторами» . Журнал биологической химии . 285 (27): 20492–20498. дои : 10.1074/jbc.M110.102111 . ПМЦ 2898307 . ПМИД 20435894 .
- ^ Руджеро Т., Трабукки М., Де Санта Ф, Зупо С., Харф Б.Д., Макманус М.Т., Розенфельд М.Г., Бриата П., Герци Р. (сентябрь 2009 г.). «ЛПС индуцирует зависимый от регуляторного белка сплайсинга KH-типа процессинг предшественников микроРНК-155 в макрофагах» . Журнал ФАСЭБ . 23 (9): 2898–2908. дои : 10.1096/fj.09-131342 . ПМИД 19423639 . S2CID 588328 .
- ^ Jump up to: а б Шульте Л.Н., Вестерманн А.Ю., Фогель Дж. (январь 2013 г.). «Дифференциальная активация и функциональная специализация миР-146 и миР-155 при чувствительности врожденного иммунитета» . Исследования нуклеиновых кислот . 41 (1): 542–553. дои : 10.1093/нар/gks1030 . ПМЦ 3592429 . ПМИД 23143100 .
- ^ Андроулидаки А, Илиопулос Д, Арранц А, Доксаки С, Шворер С, Захариудаки В, Маргиорис А.Н., Цихлис П.Н., Цацанис С (август 2009 г.). «Киназа Akt1 контролирует реакцию макрофагов на липополисахарид путем регулирования микроРНК» . Иммунитет . 31 (2): 220–231. doi : 10.1016/j.immuni.2009.06.024 . ПМЦ 2865583 . ПМИД 19699171 .
- ^ О'Коннелл Р.М., Чаудхури А.А., Рао Д.С., Балтимор Д. (апрель 2009 г.). «Инозитолфосфатаза SHIP1 является основной мишенью миР-155» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (17): 7113–7118. Бибкод : 2009PNAS..106.7113O . дои : 10.1073/pnas.0902636106 . ПМЦ 2678424 . ПМИД 19359473 .
- ^ Костинян С., Сандху С.К., Педерсен И.М., Тили Э., Тротта Р., Перротти Д., Чиарлариелло Д., Невиани П., Харб Дж., Кауфман Л.Р., Шидхам А., Кроче К.М. (август 2009 г.). «Домен Src-гомологии 2, содержащий инозитол-5-фосфатазу, и белок бета, связывающий энхансер CCAAT, является мишенью для миР-155 в В-клетках трансгенных мышей Emicro-MiR-155» . Кровь . 114 (7): 1374–1382. дои : 10.1182/blood-2009-05-220814 . ПМК 2727407 . ПМИД 19520806 .
- ^ Янкаускас СС, Гамбарделла Дж, Сарду К, Ломбарди А, Сантулли Г (июль 2021 г.). «Функциональная роль миР-155 в патогенезе сахарного диабета и его осложнений» . Некодирующая РНК . 7 (3): 39. дои : 10.3390/ncrna7030039 . ПМЦ 8293470 . ПМИД 34287359 .
- ^ Несмиянов П, Стрыгин А, Толкачев Б, Капланов К, Доценко А, Стрыгина А (2016). «Перемещение мИРНА-155 через щелевые соединения способствует прогрессированию ХЛЛ». 14-е ежегодное собрание CIMT «Механизмы эффективности иммунотерапии рака», Майнц, Германия, 2016 г.
- ^ Сонколи Э, Янсон П, Маджури МЛ, Савинко Т, Фирквист Н, Эйдсмо Л, Сюй Н, Мейсген Ф, Вэй Т, Брэдли М, Стенванг Дж, Кауппинен С, Алениус Х, Лауэрма А, Хоми Б, Винквист О, Столе М , Пиварчи А (сентябрь 2010 г.). «МиР-155 сверхэкспрессируется у пациентов с атопическим дерматитом и модулирует пролиферативные реакции Т-клеток путем нацеливания на антиген 4, ассоциированный с цитотоксическими Т-лимфоцитами». Журнал аллергии и клинической иммунологии . 126 (3): 581–589.e1–20. дои : 10.1016/j.jaci.2010.05.045 . ПМИД 20673989 .
- ^ Мур К.С., Рао В.Т., Дюрафур Б.А., Беделл Б.Дж., Людвин С.К., Бар-Ор А., Антел Дж.П. (ноябрь 2013 г.). «МиР-155 как регулятор поляризации миелоидных клеток, имеющий отношение к рассеянному склерозу» . Анналы неврологии . 74 (5): 709–720. дои : 10.1002/ana.23967 . ПМИД 23818336 . S2CID 205344718 .
- ^ О'Коннелл Р.М., Рао Д.С., Балтимор Д. (2012). «регуляция микроРНК воспалительных реакций». Ежегодный обзор иммунологии . 30 : 295–312. doi : 10.1146/annurev-immunol-020711-075013 . ПМИД 22224773 .
- ^ Скальский Р.Л., Самолс М.А., Плезанс К.Б., Босс И.В., Рива А., Лопес М.К., Бейкер Х.В., Ренне Р. (декабрь 2007 г.). «Герпесвирус, ассоциированный с саркомой Капоши, кодирует ортолог миР-155» . Журнал вирусологии . 81 (23): 12836–12845. дои : 10.1128/JVI.01804-07 . ПМК 2169101 . ПМИД 17881434 .
- ^ Линнштадт С.Д., Готтвейн Э., Скальски Р.Л., Люфтиг М.А., Каллен Б.Р. (ноябрь 2010 г.). «Вирусно-индуцированная клеточная микроРНК миР-155 играет ключевую роль в иммортализации В-клеток вирусом Эпштейна-Барр» . Журнал вирусологии . 84 (22): 11670–11678. дои : 10.1128/JVI.01248-10 . ПМЦ 2977875 . ПМИД 20844043 .
- ^ Валери Н, Гаспарини П, Фаббри М, Бракони С, Веронезе А, Ловат Ф, Адэр Б, Ваннини И, Фанини Ф, Боттони А, Костинян С, Сандху СК, Нуово ГДж, Алдер Х, Гафа Р, Калоре Ф, Феррацин М , Ланца Г., Волиния С., Негрини М., Макилхаттон М.А., Амадори Д., Фишел Р., Кроче СМ (апрель 2010 г.). «Модуляция репарации ошибочных спариваний и стабильности генома с помощью миР-155» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (15): 6982–6987. Бибкод : 2010PNAS..107.6982V . дои : 10.1073/pnas.1002472107 . ПМЦ 2872463 . ПМИД 20351277 .
- ^ Ланчки А, Надь А, Боттай Г, Мункачи Г, Сабо А, Сантарпиа Л, Дьерфи Б (01 декабря 2016 г.). «miRpower: веб-инструмент для проверки микроРНК, связанных с выживанием, с использованием данных об экспрессии 2178 пациентов с раком молочной железы». Исследование и лечение рака молочной железы . 160 (3): 439–446. дои : 10.1007/s10549-016-4013-7 . ISSN 1573-7217 . ПМИД 27744485 . S2CID 11165696 .
Дальнейшее чтение
[ редактировать ]- Бентвич И, Авниэль А, Каров Ю, Ааронов Р, Гилад С, Барад О, Барзилай А, Эйнат П, Эйнав Ю, Мейри Э, Шарон Э, Спектор Ю, Бентвич З (июль 2005 г.). «Идентификация сотен консервативных и неконсервативных микроРНК человека». Природная генетика . 37 (7): 766–770. дои : 10.1038/ng1590 . ПМИД 15965474 . S2CID 7746954 .
- Хуан Р.С., Ху GQ, Линь Б, Линь Цзы, Сунь CC (декабрь 2010 г.). «Замалчивание микроРНК-155 усиливает воспалительную реакцию и поглощение липидов в окисленных стимулированных липопротеинами низкой плотности макрофагах человека THP-1». Журнал исследовательской медицины . 58 (8): 961–967. дои : 10.2310/jim.0b013e3181ff46d7 . ПМИД 21030878 . S2CID 83123221 .
- Чеолотто Г., Паппарелла И., Бортолуцци А., Страпаццон Дж., Рагаццо Ф., Братти П., Фабрисио А.С., Скуарчина Е., Гион М., Палатини П., Семпличини А. (февраль 2011 г.). «Взаимодействие между полиморфизмом миР-155, AT1R A1166C и экспрессией AT1R у молодых нелеченых гипертоников» . Американский журнал гипертонии . 24 (2): 241–246. дои : 10.1038/ajh.2010.211 . ПМИД 20966899 .
- Ван Г, Там Л.С., Ли Е.К., Кван Б.С., Чоу К.М., Лук CC, Ли ПК, Сзето CC (декабрь 2010 г.). «Безклеточные МиР-146а и МиР-155 в сыворотке и моче у пациентов с системной красной волчанкой» . Журнал ревматологии . 37 (12): 2516–2522. дои : 10.3899/jrheum.100308 . ПМИД 20952466 . S2CID 23208771 .
- Катти Р.К., Нагинени К.Н., Сэмюэл В., Виджаясарати К., Хукс Дж.Дж., Редмонд Т.М. (ноябрь 2010 г.). «Воспалительные цитокины регулируют экспрессию микроРНК-155 в клетках пигментного эпителия сетчатки человека путем активации пути JAK/STAT» . Связь с биохимическими и биофизическими исследованиями . 402 (2): 390–395. дои : 10.1016/j.bbrc.2010.10.042 . ПМЦ 2992362 . ПМИД 20950585 .
- Имаидзуми Т., Танака Х., Тадзима А., Ёконо Ю., Мацумия Т., Ёсида Х., Цуруга К., Айзава-Яширо Т., Хаякари Р., Иноуэ И., Ито Э., Сато К. (2010). «IFN-γ и TNF-α синергически индуцируют микроРНК-155, которая регулирует экспрессию TAB2/IP-10 в мезангиальных клетках человека». Американский журнал нефрологии . 32 (5): 462–468. дои : 10.1159/000321365 . ПМИД 20948191 . S2CID 12859943 .
- Томпсон Р.К., Херскович М., Чжао И., Форд Т.Дж., Гилмор Т.Д. (январь 2011 г.). «NF-kappaB подавляет экспрессию маркера B-лимфомы CD10 через путь miR-155/PU.1» . Журнал биологической химии . 286 (3): 1675–1682. дои : 10.1074/jbc.M110.177063 . ПМК 3023462 . ПМИД 20947507 .
- Ван П, Хоу Дж, Линь Л, Ван С, Лю Х, Ли Д, Ма Ф, Ван З, Цао X (ноябрь 2010 г.). «Индуцируемая обратная связь микроРНК-155 способствует передаче сигналов IFN типа I при противовирусном врожденном иммунитете путем нацеливания на супрессор передачи сигналов цитокинов 1» . Журнал иммунологии . 185 (10): 6226–6233. doi : 10.4049/jimmunol.1000491 . ПМИД 20937844 .
- О'Коннелл Р.М., Кан Д., Гибсон В.С., Раунд Дж.Л., Шольц Р.Л., Чаудхури А.А., Кан М.Е., Рао Д.С., Балтимор Д. (октябрь 2010 г.). «МикроРНК-155 способствует аутоиммунному воспалению, усиливая развитие воспалительных Т-клеток» . Иммунитет . 33 (4): 607–619. doi : 10.1016/j.immuni.2010.09.009 . ПМЦ 2966521 . ПМИД 20888269 .
- Ся QS, Исигаки Ю, Чжао X, Симасаки Т, Накадзима Х, Накагава Х, Такегами Т, Чен Чж, Мотоо Ю (январь 2011 г.). «Человеческий SMG-1 участвует в индуцированной гемцитабином первичной активации микроРНК-155/BIC в клетках PANC-1 рака поджелудочной железы человека». Поджелудочная железа . 40 (1): 55–60. дои : 10.1097/MPA.0b013e3181e89f74 . ПМИД 20871480 . S2CID 39789659 .
- Чжоу Х, Хуан Х, Цуй Х, Луо Х, Тан Ю, Чен С, Ву Л, Шен Н (декабрь 2010 г.). «МиР-155 и ее звездообразный партнер миР-155* совместно регулируют выработку интерферона типа I плазмоцитоидными дендритными клетками человека» . Кровь . 116 (26): 5885–5894. дои : 10.1182/кровь-2010-04-280156 . ПМИД 20852130 .
- Линнштадт С.Д., Готтвейн Э., Скальски Р.Л., Люфтиг М.А., Каллен Б.Р. (ноябрь 2010 г.). «Вирусно-индуцированная клеточная микроРНК миР-155 играет ключевую роль в иммортализации В-клеток вирусом Эпштейна-Барр» . Журнал вирусологии . 84 (22): 11670–11678. дои : 10.1128/JVI.01248-10 . ПМЦ 2977875 . ПМИД 20844043 .
- Чжэн Л., Сюй CC, Чен В.Д., Шен В.Л., Жуань CC, Чжу Л.М., Чжу Д.Л., Гао П.Дж. (октябрь 2010 г.). «МикроРНК-155 регулирует экспрессию рецептора ангиотензина II типа 1 и фенотипическую дифференцировку в сосудистых адвентициальных фибробластах». Связь с биохимическими и биофизическими исследованиями . 400 (4): 483–488. дои : 10.1016/j.bbrc.2010.08.067 . ПМИД 20735984 .
- Тили Э, Мишель Дж. Дж., Адэр Б., Алдер Х., Лимань Е., Таччоли С., Феррацин М., Дельмас Д., Латруфф Н., Кроче СМ (сентябрь 2010 г.). «Ресвератрол снижает уровень миР-155 за счет активации миР-663, микроРНК, нацеленной на JunB и JunD» . Канцерогенез . 31 (9): 1561–1566. дои : 10.1093/carcin/bgq143 . ПМЦ 4647642 . ПМИД 20622002 .
- Бош-Саадатманди С, Лобода А, Вагнер А.Е., Стачурска А, Йозкович А, Дулак Дж, Дёринг Ф, Вольфрам С, Римбах Г (март 2011 г.). «Влияние кверцетина и его метаболитов изорамнетина и кверцетин-3-глюкуронида на экспрессию воспалительных генов: роль миР-155». Журнал пищевой биохимии . 22 (3): 293–299. дои : 10.1016/j.jnutbio.2010.02.008 . ПМИД 20579867 .
- Лю Дж., ван Мил А., Врейсен К., Чжао Дж., Гао Л., Мец Ч., Гуманс М.Дж., Довенданс П.А., Слюйтер Дж.П. (июль 2011 г.). «МикроРНК-155 предотвращает некротическую гибель клеток в клетках-предшественниках кардиомиоцитов человека посредством нацеливания на RIP1» . Журнал клеточной и молекулярной медицины . 15 (7): 1474–1482. дои : 10.1111/j.1582-4934.2010.01104.x . ПМЦ 3823192 . ПМИД 20550618 .
- Погрибный И.П., Старлард-Дэвенпорт А., Трындяк В.П., Хан Т., Росс С.А., Русин И., Беланд Ф.А. (октябрь 2010 г.). «Разница в экспрессии печеночных микроРНК миР-29c, миР-34а, миР-155 и миР-200b связана со штаммоспецифической восприимчивостью к пищевому неалкогольному стеатогепатиту у мышей» . Лабораторное исследование . 90 (10): 1437–1446. дои : 10.1038/labinvest.2010.113 . ПМК 4281935 . ПМИД 20548288 .
- Чжан Ю, Дяо З, Су Л, Сунь Х, Ли Р, Цуй Х, Ху Ю (май 2010 г.). «МикроРНК-155 способствует развитию преэклампсии за счет снижения уровня CYR61». Американский журнал акушерства и гинекологии . 202 (5): 466.e1–7. дои : 10.1016/j.ajog.2010.01.057 . ПМИД 20452491 .
- Ху Ю.Л., Фонг С., Ларгман С., Шен В.Ф. (сентябрь 2010 г.). «НОХА9 регулирует миР-155 в кроветворных клетках» . Исследования нуклеиновых кислот . 38 (16): 5472–5478. дои : 10.1093/нар/gkq337 . ПМЦ 2938212 . ПМИД 20444872 .
- Маккой С.Э., Шиди Ф.Дж., Куаллс Дж.Э., Дойл С.Л., Куинн С.Р., Мюррей П.Дж., О'Нил Л.А. (июль 2010 г.). «IL-10 ингибирует индукцию миР-155 толл-подобными рецепторами» . Журнал биологической химии . 285 (27): 20492–20498. дои : 10.1074/jbc.M110.102111 . ПМЦ 2898307 . ПМИД 20435894 .
- Инь Ц, Ван Х, Фьюэлл С, Кэмерон Дж, Чжу Х, Бадду М, Лин З, Флемингтон Е.К. (июль 2010 г.). «МикроРНК миР-155 ингибирует передачу сигналов костного морфогенетического белка (BMP) и опосредованную BMP реактивацию вируса Эпштейна-Барр» . Журнал вирусологии . 84 (13): 6318–6327. дои : 10.1128/JVI.00635-10 . ПМЦ 2903268 . ПМИД 20427544 .
- Чжу Дж, Ху XQ, Го ГЛ, Чжан Ю, Ван О.К., Ю Дж, Хуан QD, Чжан XH (февраль 2010 г.). «[Экспрессия и ее клиническое значение миР-155 при первичном раке молочной железы человека]». Чжунхуа Вай Кэ За Чжи [Китайский журнал хирургии] . 48 (3): 205–208. ПМИД 20388420 .
- Ся QS, Исигаки Ю, Сунь Л, Чен Р, Мотоо Ю (январь 2010 г.). «[Влияние противораковых препаратов на экспрессию BIC/миР-155 в клетках PANC-1 рака поджелудочной железы человека]». Чжунхуа И Сюэ Цза Чжи . 90 (2): 123–127. ПМИД 20356498 .
- Цзян С., Чжан Х.В., Лу М.Х., Хэ С.Х., Ли Ю, Гу Х, Лю М.Ф., Ван Э.Д. (апрель 2010 г.). «МикроРНК-155 действует как онкомиР при раке молочной железы, воздействуя на супрессор гена, передающего сигнал цитокина 1» . Исследования рака . 70 (8): 3119–3127. doi : 10.1158/0008-5472.CAN-09-4250 . ПМИД 20354188 .
- Рю Дж.К., Хонг С.М., Карикари К.А., Хрубан Р.Х., Гоггинс М.Г., Майтра А. (2010). «Аберрантная экспрессия микроРНК-155 является ранним событием многоступенчатого прогрессирования аденокарциномы поджелудочной железы» . Панкреатология . 10 (1): 66–73. дои : 10.1159/000231984 . ПМЦ 2865485 . ПМИД 20332664 .
- Фабани М.М., Абреу-Гуджер С., Уильямс Д., Лайонс П.А., Торрес А.Г., Смит К.Г., Энрайт А.Дж., Гейт М.Дж., Вигорито Э (июль 2010 г.). «Эффективное ингибирование функции миР-155 in vivo пептидно-нуклеиновыми кислотами» . Исследования нуклеиновых кислот . 38 (13): 4466–4475. дои : 10.1093/нар/gkq160 . ПМК 2910044 . ПМИД 20223773 .
- Тан Б, Сяо Б, Лю З, Ли Н, Чжу ЭД, Ли БС, Се QH, Чжуан Ю, Цзоу QM, Мао XH (апрель 2010 г.). «Идентификация MyD88 как новой мишени миР-155, участвующей в негативной регуляции воспаления, вызванного Helicobacter pylori». Письма ФЭБС . 584 (8): 1481–1486. дои : 10.1016/j.febslet.2010.02.063 . ПМИД 20219467 . S2CID 41984991 .
- Фасси Фери Л., Кох М., Белоголова Е., Халил Х., Больц С., Калали Б., Молленкопф Х.Дж., Бежье-Бомпадр М., Карлас А., Шнайдер Т., Чурин Ю., Герхард М., Мейер Т.Ф. (2010). «Helicobacter pylori индуцирует миР-155 в Т-клетках цАМФ-Foxp3-зависимым образом» . ПЛОС ОДИН . 5 (3): е9500. Бибкод : 2010PLoSO...5.9500F . дои : 10.1371/journal.pone.0009500 . ПМЦ 2830477 . ПМИД 20209161 .
- Сидоркевич М., Грек М., Йозвяк Б., Майда-Станиславска Е., Пекарска А., Бартковяк Дж. (2010). «Экспрессия предшественника микроРНК-155 в мононуклеарных клетках периферической крови пациентов с гепатитом С после противовирусного лечения». Акта вирусологика . 54 (1): 75–78. дои : 10.4149/av_2010_01_75 . ПМИД 20201617 .
- Гуэта К, Молоцкий Н, Герчиков Н, Мор Э, Савьон С, Фейн А, Тодер В, Шомрон Н, Торчинский А (2010). «Вызванные тератогеном изменения в экспрессии микроРНК-34, микроРНК-125b и микроРНК-155: корреляция с эмбриональным генотипом р53 и фенотипом конечностей» . Биология развития BMC . 10:20 . дои : 10.1186/1471-213X-10-20 . ПМЦ 2841584 . ПМИД 20170545 .
- Рай Д., Ким С.В., Маккеллер М.Р., Дахия П.Л., Агиар Р.К. (февраль 2010 г.). «Нацеливание на SMAD5 связывает микроРНК-155 с путем TGF-бета и лимфомогенезом» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (7): 3111–3116. Бибкод : 2010PNAS..107.3111R . дои : 10.1073/pnas.0910667107 . ПМЦ 2840369 . ПМИД 20133617 .
- Кремер Т.Дж., Равнеберг Д.Х., Клэй CD, Пайпер-Хантер М.Г., Марш CB, Элтон Т.С., Ганн Дж.С., Амер А., Каннеганти Т.Д., Шлезингер Л.С., Бучар Дж.П., Тридандапани С. (2009). «Индукция МиР-155 F. novicida, но не вирулентным F. tularensis, приводит к снижению регуляции SHIP и усилению провоспалительной цитокиновой реакции» . ПЛОС ОДИН . 4 (12): е8508. Бибкод : 2009PLoSO...4.8508C . дои : 10.1371/journal.pone.0008508 . ПМЦ 2794384 . ПМИД 20041145 .
- Форрест А.Р., Канамори-Катаяма М., Томару Ю., Лассманн Т., Ниномия Н., Такахаши Ю., де Хун М.Дж., Кубосаки А., Кайхо А., Сузуки М., Ясуда Дж., Каваи Дж., Хаяшизаки Ю., Хьюм Д.А., Сузуки Х. (февраль 2010 г.) ). «Индукция микроРНК, мир-155, мир-222, мир-424 и мир-503, способствует моноцитарной дифференцировке посредством комбинаторной регуляции». Лейкемия . 24 (2): 460–466. arXiv : 1007.2689 . дои : 10.1038/leu.2009.246 . ПМИД 19956200 . S2CID 2333658 .
- Педерсен И.М., Отеро Д., Као Э., Милетич А.В., Хотер С., Ральфкиер Э., Рикерт Р.К., Гронбек К., Дэвид М. (август 2009 г.). «Онко-миР-155 нацелена на SHIP1, чтобы стимулировать TNF-зависимый рост В-клеточных лимфом» . ЭМБО Молекулярная медицина . 1 (5): 288–295. дои : 10.1002/emmm.200900028 . ПМЦ 2771872 . ПМИД 19890474 .
- Хэ М, Сюй З, Дин Т, Куанг ДМ, Чжэн Л (октябрь 2009 г.). «МикроРНК-155 регулирует выработку воспалительных цитокинов в опухолеассоциированных макрофагах посредством нацеливания на C/EBPbeta» . Клеточная и молекулярная иммунология . 6 (5): 343–352. дои : 10.1038/cmi.2009.45 . ПМЦ 4003217 . ПМИД 19887047 .
- Тили Э, Кроче СМ, Мишель Дж.Дж. (2009). «МиР-155: о взаимосвязи между воспалением и раком». Международные обзоры иммунологии . 28 (5): 264–284. дои : 10.1080/08830180903093796 . ПМИД 19811312 . S2CID 205589961 .
- Шталь Х.Ф., Фаути Т., Ульрих Н., Бопп Т., Кубах Дж., Раст В., Лабхарт П., Алексиадис В., Беккер С., Хафнер М., Вейт А., Лентер М.К., Йонулейт Х., Шмитт Э., Меннерих Д. (2009). «Ингибирование миР-155 повышает чувствительность CD4+ Th-клеток к подавлению, опосредованному TREG» . ПЛОС ОДИН . 4 (9): е7158. Бибкод : 2009PLoSO...4.7158S . дои : 10.1371/journal.pone.0007158 . ПМК 2743997 . ПМИД 19777054 .
- Болисетти М.Т., Дай Дж., Тэм В., Бимон К.Л. (декабрь 2009 г.). «Штамт вируса ретикулоэндотелиоза Т индуцирует миР-155, которая нацелена на JARID2 и способствует выживанию клеток» . Журнал вирусологии . 83 (23): 12009–12017. дои : 10.1128/JVI.01182-09 . ПМЦ 2786729 . ПМИД 19759154 .
- Ван Б., Маджумдер С., Нуово Г., Кутай Х., Волиния С., Патель Т., Шмиттген Т.Д., Кроче С., Гошал К., Джейкоб С.Т. (октябрь 2009 г.). «Роль микроРНК-155 на ранних стадиях гепатокарциногенеза, индуцированного диетой с дефицитом холина и аминокислотами у мышей C57BL/6» . Гепатология . 50 (4): 1152–1161. дои : 10.1002/геп.23100 . ПМЦ 2757532 . ПМИД 19711427 .
- Потье Н., Морен Т., Шевалье Б., Пюиссегюр М.П., Лебриганд К., Робб-Сермесант К., Бертеро Т., Лино Карденас С.Л., Курко Е., Риос Ж., Фурр С., Ло-Гидис Ж.М., Марсет Б., Кардино Б., Барбри П., Мари Б (2009). «Идентификация фактора роста кератиноцитов как мишени микроРНК-155 в фибробластах легких: участие в эпителиально-мезенхимальных взаимодействиях» . ПЛОС ОДИН . 4 (8): е6718. Бибкод : 2009PLoSO...4.6718P . дои : 10.1371/journal.pone.0006718 . ПМК 2726943 . ПМИД 19701459 .
- Сяо Б, Лю З, Ли БС, Тан Б, Ли В, Го Г, Ши Ю, Ван Ф, Ву Ю, Тонг ВД, Го Х, Мао XH, Цзоу QM (сентябрь 2009 г.). «Индукция микроРНК-155 при инфекции Helicobacter pylori и ее негативная регуляторная роль в воспалительной реакции». Журнал инфекционных болезней . 200 (6): 916–925. дои : 10.1086/605443 . ПМИД 19650740 . S2CID 8719437 .
- Ворм Дж., Стенванг Дж., Петри А., Фредериксен К.С., Обад С., Эльмин Дж., Хедтьярн М., Страаруп Э.М., Хансен Дж.Б., Кауппинен С. (сентябрь 2009 г.). «Замалчивание микроРНК-155 у мышей во время острой воспалительной реакции приводит к дерепрессии бета-c/ebp и снижению регуляции G-CSF» . Исследования нуклеиновых кислот . 37 (17): 5784–5792. дои : 10.1093/nar/gkp577 . ПМЦ 2761263 . ПМИД 19596814 .
- Костинян С., Сандху С.К., Педерсен И.М., Тили Э., Тротта Р., Перротти Д., Чиарлариелло Д., Невиани П., Харб Дж., Кауфман Л.Р., Шидхам А., Кроче К.М. (август 2009 г.). «Домен Src-гомологии 2, содержащий инозитол-5-фосфатазу, и белок бета, связывающий энхансер CCAAT, является мишенью для миР-155 в В-клетках трансгенных мышей Emicro-MiR-155» . Кровь . 114 (7): 1374–1382. дои : 10.1182/blood-2009-05-220814 . ПМК 2727407 . ПМИД 19520806 .
- Руджеро Т., Трабукки М., Де Санта Ф, Зупо С., Харф Б.Д., Макманус М.Т., Розенфельд М.Г., Бриата П., Герци Р. (сентябрь 2009 г.). «ЛПС индуцирует зависимый от регуляторного белка сплайсинга KH-типа процессинг предшественников микроРНК-155 в макрофагах» . Журнал ФАСЭБ . 23 (9): 2898–2908. дои : 10.1096/fj.09-131342 . ПМИД 19423639 . S2CID 588328 .
- Мартинес-Нунес Р.Т., Луафи Ф., Фридманн П.С., Санчес-Эльснер Т. (июнь 2009 г.). «МикроРНК-155 модулирует способность дендритных клеток (ДК) связывать патогены путем подавления специфической для ДК молекулы межклеточной адгезии-3, захватывающей неинтегрин (DC-SIGN)» . Журнал биологической химии . 284 (24): 16334–16342. дои : 10.1074/jbc.M109.011601 . ПМЦ 2713543 . ПМИД 19386588 .
- О'Коннелл Р.М., Чаудхури А.А., Рао Д.С., Балтимор Д. (апрель 2009 г.). «Инозитолфосфатаза SHIP1 является основной мишенью миР-155» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (17): 7113–7118. Бибкод : 2009PNAS..106.7113O . дои : 10.1073/pnas.0902636106 . ПМЦ 2678424 . ПМИД 19359473 .
- Кольхас С., Гарден О.А., Скадамор К., Тернер М., Оккенхауг К., Вигорито Э (март 2009 г.). «Авангард: миР-155-мишень Foxp3 способствует развитию регуляторных Т-клеток» . Журнал иммунологии . 182 (5): 2578–2582. doi : 10.4049/jimmunol.0803162 . ПМИД 19234151 .
- Чеппи М., Перейра П.М., Дюнан-Сотье I, Баррас Э., Рейт В., Сантос М.А., Пьер П. (февраль 2009 г.). «МикроРНК-155 модулирует сигнальный путь интерлейкина-1 в активированных дендритных клетках, происходящих из моноцитов человека» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (8): 2735–2740. Бибкод : 2009PNAS..106.2735C . дои : 10.1073/pnas.0811073106 . ПМК 2650335 . ПМИД 19193853 .
- Хаббе Н., Курстра Дж.Б., Менделл Дж.Т., Офферхаус Г.Дж., Рю Дж.К., Фельдманн Г., Маллендор М.Е., Гоггинс М.Г., Хонг С.М., Майтра А (февраль 2009 г.). «МикроРНК миР-155 является биомаркером ранней неоплазии поджелудочной железы» . Биология и терапия рака . 8 (4): 340–346. дои : 10.4161/cbt.8.4.7338 . ПМК 2692997 . ПМИД 19106647 .
- Юнг И, Агиар RC (январь 2009 г.). «Экспрессия микроРНК-155 и исход при диффузной крупноклеточной В-клеточной лимфоме» . Британский журнал гематологии . 144 (1): 138–140. дои : 10.1111/j.1365-2141.2008.07424.x . ПМЦ 4654117 . ПМИД 19016736 .
- Румыния П., Лулли В., Пелоси Э., Биффони М., Пешле С., Марциали Г. (ноябрь 2008 г.). «МикроРНК 155 модулирует мегакариопоэз на уровне предшественников и предшественников, нацеливаясь на факторы транскрипции Ets-1 и Meis1» . Британский журнал гематологии . 143 (4): 570–580. дои : 10.1111/j.1365-2141.2008.07382.x . ПМИД 18950466 . S2CID 9025761 .
- Чжао Ю, Яо Ю, Сюй Х, Ламбет Л, Смит ЛП, Кгосана Л, Ван Х, Наир В (январь 2009 г.). «Функциональный ортолог микроРНК-155, кодируемый онкогенным вирусом болезни Марека» . Журнал вирусологии . 83 (1): 489–492. дои : 10.1128/JVI.01166-08 . ПМК 2612317 . ПМИД 18945769 .
- Гатто Дж., Росси А., Росси Д., Кренинг С., Бонатти С., Маллардо М. (ноябрь 2008 г.). «Латентный мембранный белок 1 вируса Эпштейна-Барра трансактивирует транскрипцию миР-155 через путь NF-kappaB» . Исследования нуклеиновых кислот . 36 (20): 6608–6619. дои : 10.1093/нар/gkn666 . ПМЦ 2582607 . ПМИД 18940871 .
- Рахадиани Н., Такакува Т., Треснасари К., Мории Э., Аозаса К. (декабрь 2008 г.). «Латентный мембранный белок-1 вируса Эпштейна-Барра индуцирует экспрессию кластера интеграции B-клеток, предшественника микроРНК-155, в клеточных линиях B-лимфомы». Связь с биохимическими и биофизическими исследованиями . 377 (2): 579–583. дои : 10.1016/j.bbrc.2008.10.007 . ПМИД 18926796 .
- Конг В., Ян Х., Хэ Л., Чжао Дж.Дж., Коппола Д., Далтон В.С., Ченг Дж.К. (ноябрь 2008 г.). «МикроРНК-155 регулируется путем трансформационного фактора роста бета/Smad и способствует пластичности эпителиальных клеток путем нацеливания на RhoA» . Молекулярная и клеточная биология . 28 (22): 6773–6784. дои : 10.1128/MCB.00941-08 . ПМЦ 2573297 . ПМИД 18794355 .
- Лу Ф., Вейдмер А., Лю К.Г., Волиния С., Кроче С.М., Либерман П.М. (ноябрь 2008 г.). «Индуцированная вирусом Эпштейна-Барра миР-155 ослабляет передачу сигналов NF-kappaB и стабилизирует латентную персистенцию вируса» . Журнал вирусологии . 82 (21): 10436–10443. дои : 10.1128/JVI.00752-08 . ПМЦ 2573162 . ПМИД 18753206 .
- Дорсетт Ю., МакБрайд К.М., Янкович М., Газумян А., Тай Т.Х., Роббиани Д.Ф., Ди Вирджилио М., Рейна Сан-Мартин Б., Хайдкамп Г., Швикерт Т.А., Эйзенрайх Т., Раевски К., Нуссенцвейг М.К. (май 2008 г.). «МикроРНК-155 подавляет индуцированную активацией транслокацию Myc-Igh, опосредованную цитидиндезаминазой» . Иммунитет . 28 (5): 630–638. doi : 10.1016/j.immuni.2008.04.002 . ПМЦ 2713656 . ПМИД 18455451 .
- Тенг Г., Хакимпур П., Ландграф П., Райс А., Тушл Т., Каселлас Р., Папавасилиу Ф.Н. (май 2008 г.). «МикроРНК-155 является негативным регулятором цитидиндезаминазы, индуцированной активацией» . Иммунитет . 28 (5): 621–629. doi : 10.1016/j.immuni.2008.03.015 . ПМК 2430982 . ПМИД 18450484 .
- Инь Кью, МакБрайд Дж., Фьюэлл С., Лейси М., Ван Х, Лин З., Кэмерон Дж., Флемингтон Э.К. (июнь 2008 г.). «МикроРНК-155 представляет собой ген, индуцируемый вирусом Эпштейна-Барра, который модулирует пути экспрессии генов, регулируемые вирусом Эпштейна-Барр» . Журнал вирусологии . 82 (11): 5295–5306. дои : 10.1128/JVI.02380-07 . ПМК 2395216 . ПМИД 18367535 .
- Чжан Т., Не К., Тэм В. (сентябрь 2008 г.). «BIC эффективно процессируется до микроРНК-155 в клетках лимфомы Беркитта». Лейкемия . 22 (9): 1795–1797. дои : 10.1038/leu.2008.62 . ПМИД 18354490 . S2CID 8783237 .
- Ван М., Тан Л.П., Дейкстра М.К., ван Лом К., Робертус Дж.Л., Хармс Г., Блокзейл Т., Кооистра К., ван Твеер М.Б., Розати С., Виссер Л., Йонген-Лавренчич М., Клюин П.М., ван ден Берг А. май 2008 г.). «Анализ микроРНК при В-клеточном хроническом лимфоцитарном лейкозе: центры пролиферации, характеризующиеся низкой экспрессией миР-150 и высокой экспрессией BIC/миР-155». Журнал патологии . 215 (1): 13–20. дои : 10.1002/путь.2333 . ПМИД 18348159 . S2CID 29470560 .
- О'Коннелл Р.М., Рао Д.С., Чаудхури А.А., Болдин М.П., Таганов К.Д., Николл Дж., Пакетт Р.Л., Балтимор Д. (март 2008 г.). «Устойчивая экспрессия микроРНК-155 в гемопоэтических стволовых клетках вызывает миелопролиферативное заболевание» . Журнал экспериментальной медицины . 205 (3): 585–594. дои : 10.1084/jem.20072108 . ПМК 2275382 . ПМИД 18299402 .
- Рай Д., Каранти С., Юнг И., Дахия П.Л., Агиар Р.К. (февраль 2008 г.). «Координированная экспрессия микроРНК-155 и предсказанные гены-мишени при диффузной крупноклеточной В-клеточной лимфоме» . Генетика рака и цитогенетика . 181 (1): 8–15. doi : 10.1016/j.cancergencyto.2007.10.008 . ПМК 2276854 . ПМИД 18262046 .
- Готтвейн Э., Мукерджи Н., Саксе С., Френцель С., Майорос В.Х., Чи Дж.Т., Брейч Р., Манохаран М., Соутшек Дж., Олер У., Каллен Б.Р. (декабрь 2007 г.). «Вирусная микроРНК действует как ортолог клеточной миР-155» . Природа . 450 (7172): 1096–1099. Бибкод : 2007Natur.450.1096G . дои : 10.1038/nature05992 . ПМК 2614920 . ПМИД 18075594 .
- Вигорито Э., Перкс К.Л., Абреу-Гуджер С., Бантинг С., Сян З., Кольхас С., Дас П.П., Миска Э.А., Родригес А., Брэдли А., Смит К.Г., Рада С., Энрайт А.Дж., Тёлльнер К.М., Макленнан И.С., Тернер М. ( декабрь 2007 г.). «МикроРНК-155 регулирует образование плазматических клеток с переключенным классом иммуноглобулинов» . Иммунитет . 27 (6): 847–859. doi : 10.1016/j.immuni.2007.10.009 . ПМЦ 4135426 . ПМИД 18055230 .
- Инь Q, Ван X, Макбрайд Дж, Фьюэлл С, Флемингтон Э (февраль 2008 г.). «Активация B-клеточного рецептора индуцирует экспрессию BIC/миР-155 через консервативный элемент AP-1» . Журнал биологической химии . 283 (5): 2654–2662. дои : 10.1074/jbc.M708218200 . ПМЦ 2810639 . ПМИД 18048365 .
- Тили Э, Мишель Дж.Дж., Чимино А., Костинян С., Думитру К.Д., Адэр Б., Фаббри М., Алдер Х., Лю К.Г., Калин Г.А., Кроче СМ (октябрь 2007 г.). «Модуляция уровней миР-155 и миР-125b после стимуляции липополисахаридом/ФНО-альфа и их возможная роль в регуляции ответа на эндотоксиновый шок» . Журнал иммунологии . 179 (8): 5082–5089. дои : 10.4049/jimmunol.179.8.5082 . ПМИД 17911593 .
- Жиронелла М, Се М, Се МДж, Кано С, Томасини Р, Гоммо Дж, Гарсиа С, Новак Дж, Юнг МЛ, Жанг К.Т., Чайкс А, Фазли Л, Мотоо Ю, Ван К, Рокки П, Руссо А, Глив М , Дагорн Дж.К., Йованна Дж.Л., Кэрриер А, Пебуск М.Дж., Дузетти, Нью-Джерси (октябрь 2007 г.). «Экспрессия ядерного белка 1, индуцированная опухолевым белком 53, подавляется миР-155, и ее восстановление ингибирует развитие опухоли поджелудочной железы» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (41): 16170–16175. Бибкод : 2007PNAS..10416170G . дои : 10.1073/pnas.0703942104 . ПМК 2042180 . ПМИД 17911264 .
- Скальский Р.Л., Самолс М.А., Плезанс К.Б., Босс И.В., Рива А., Лопес М.К., Бейкер Х.В., Ренне Р. (декабрь 2007 г.). «Герпесвирус, ассоциированный с саркомой Капоши, кодирует ортолог миР-155» . Журнал вирусологии . 81 (23): 12836–12845. дои : 10.1128/JVI.01804-07 . ПМК 2169101 . ПМИД 17881434 .
- Сетупати П., Борель С., Гагнебин М., Грант Г.Р., Дойч С., Элтон Т.С., Хацигеоргиу А.Г., Антонаракис С.Е. (август 2007 г.). «Человеческая микроРНК-155 на хромосоме 21 по-разному взаимодействует со своей полиморфной мишенью в 3'-нетранслируемой области AGTR1: механизм функционального однонуклеотидного полиморфизма, связанного с фенотипами» . Американский журнал генетики человека . 81 (2): 405–413. дои : 10.1086/519979 . ПМК 1950808 . PMID 17668390 .
- Родригес А., Вигорито Э., Клэр С., Уоррен М.В., Кутте П., Сунд Д.Р., ван Донген С., Грокок Р.Дж., Дас П.П., Миска Э.А., Ветри Д., Оккенхауг К., Энрайт Эй.Дж., Дуган Г., Тернер М., Брэдли А. (апрель). 2007). «Потребность в bic/микроРНК-155 для нормальной иммунной функции» . Наука . 316 (5824): 608–611. Бибкод : 2007Sci...316..608R . дои : 10.1126/science.1139253 . ПМК 2610435 . ПМИД 17463290 .
- Тай Т.Д., Каладо Д.П., Касола С., Ансель К.М., Сяо С., Сюэ Ю., Мерфи А., Френдьюи Д., Валенсуэла Д., Куток Дж.Л., Шмидт-Супприан М., Раевски Н., Янкопулос Г., Рао А., Раевски К. (апрель 2007 г.) . «Регуляция ответа зародышевого центра с помощью микроРНК-155». Наука . 316 (5824): 604–608. Бибкод : 2007Sci...316..604T . дои : 10.1126/science.1141229 . ПМИД 17463289 . S2CID 8174458 .
- О'Коннелл Р.М., Таганов К.Д., Болдин М.П., Ченг Г., Балтимор Д. (январь 2007 г.). «МикроРНК-155 индуцируется во время воспалительной реакции макрофагов» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (5): 1604–1609. Бибкод : 2007PNAS..104.1604O . дои : 10.1073/pnas.0610731104 . ПМК 1780072 . ПМИД 17242365 .
- Чунг К.Х., Харт CC, Аль-Бассам С., Эйвери А., Тейлор Дж., Патель П.Д., Войтек А.Б., Тернер Д.Л. (2006). «Векторы экспрессии полицистронной РНК-полимеразы II для РНК-интерференции на основе BIC/miR-155» . Исследования нуклеиновых кислот . 34 (7): е53. дои : 10.1093/нар/gkl143 . ПМЦ 1435982 . ПМИД 16614444 .
- Тэм В., Дальберг Дж. Э. (февраль 2006 г.). «МиР-155/BIC как онкогенная микроРНК». Гены, хромосомы и рак . 45 (2): 211–212. дои : 10.1002/gcc.20282 . ПМИД 16252262 . S2CID 8513721 .
- Клюйвер Дж., Хараламбиева Э., де Йонг Д., Блокзейл Т., Джейкобс С., Кроесен Б.Дж., Поппема С., ван ден Берг А. (февраль 2006 г.). «Отсутствие экспрессии BIC и микроРНК миР-155 в первичных случаях лимфомы Беркитта». Гены, хромосомы и рак . 45 (2): 147–153. дои : 10.1002/gcc.20273 . ПМИД 16235244 . S2CID 16897980 .
- Цзян Дж., Ли Э.Дж., Шмиттген Т.Д. (январь 2006 г.). «Повышенная экспрессия микроРНК-155 в линиях лимфобластоидных клеток, трансформированных вирусом Эпштейна-Барр». Гены, хромосомы и рак . 45 (1): 103–106. дои : 10.1002/gcc.20264 . ПМИД 16175574 . S2CID 19138209 .
- Клюйвер Дж., Поппема С., де Йонг Д., Блокзейл Т., Хармс Г., Джейкобс С., Кроесен Б.Дж., ван ден Берг А. (октябрь 2005 г.). «BIC и миР-155 высоко экспрессируются при лимфомах Ходжкина, первичных медиастинальных и диффузных крупноклеточных В-клеточных лимфомах». Журнал патологии . 207 (2): 243–249. дои : 10.1002/путь.1825 . ПМИД 16041695 . S2CID 14329069 .
- Эйс П.С., Тэм В., Сан Л., Чадберн А., Ли З., Гомес М.Ф., Лунд Э., Дальберг Дж.Э. (март 2005 г.). «Накопление миР-155 и BIC РНК в В-клеточных лимфомах человека» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (10): 3627–3632. Бибкод : 2005PNAS..102.3627E . дои : 10.1073/pnas.0500613102 . ПМК 552785 . ПМИД 15738415 .
- Мецлер М., Вильда М., Буш К., Виманн С., Боркхардт А. (февраль 2004 г.). «Высокая экспрессия предшественника микроРНК-155/BIC РНК у детей с лимфомой Беркитта». Гены, хромосомы и рак . 39 (2): 167–169. дои : 10.1002/gcc.10316 . ПМИД 14695998 . S2CID 10009892 .
- Сюй Ю, Ву Дж, Юань Икс, Лю В, Пань Дж, Сюй Б (2021). «МикроРНК-155 способствует иммунитету хозяина против Toxoplasma gondii » . Паразит . 28 : 83. doi : 10.1051/parasite/2021082 . ПМЦ 8672677 . ПМИД 34907898 .

Внешние ссылки
[ редактировать ]- miRBase
- Записи miRBase для всех известных mir-155
- Страница семейства предшественников микроРНК mir-155 в Rfam