Х-хромосома
| Х-хромосома человека | |
|---|---|
Х-хромосома человека (после G-бэндинга ) | |
 мужчины человека Х-хромосома в кариограмме | |
| Функции | |
| Длина ( б.п. ) | 154 259 566 п.н. (CHM13) |
| Количество генов | 804 ( ССДС ) |
| Тип | Аллосома |
| Положение центромеры | Субметацентрический [ 1 ] (61,0 Мбит/с) [ 2 ] |
| Полные списки генов | |
| CCDS | Список генов |
| HGNC | Список генов |
| ЮниПрот | Список генов |
| NCBI | Список генов |
| Внешние программы просмотра карт | |
| Вместе | Хромосома Х |
| Входить | Хромосома Х |
| NCBI | Хромосома Х |
| УКСК | Хромосома Х |
| Полные последовательности ДНК | |
| RefSeq | NC_000023 ( ИСПРАВЛЕНО ) |
| ГенБанк | CM000685 ( ИСПРАВЛЕНО ) |
является Х-хромосома одной из двух половых хромосом у многих организмов, включая млекопитающих, и встречается как у мужчин, так и у женщин. Это часть системы определения пола XY и системы определения пола XO . Х-хромосома была названа в честь своих уникальных свойств ранними исследователями, в результате чего ее коллега Y-хромосома была названа по следующей букве алфавита после ее последующего открытия. [ 3 ]
Открытие
[ редактировать ]Особость Х-хромосомы впервые было отмечено в 1890 году Германом Хенкингом в Лейпциге. Хенкинг изучал семенники пиррокориса и заметил, что одна хромосома не участвует в мейозе . Хромосомы названы так из-за их способности окрашиваться ( хрома по -гречески означает цвет ). Хотя Х-хромосому можно было окрасить так же хорошо, как и другие, Хенкинг не был уверен, относится ли это к другому классу объекта, и поэтому назвал его X-элементом . [ 4 ] которая позже стала Х-хромосомой после того, как было установлено, что это действительно хромосома. [ 5 ]
Представление о том, что Х-хромосома получила свое название из-за ее сходства с буквой «Х», ошибочно. Все хромосомы в норме выглядят под микроскопом как аморфная капля и принимают четко очерченную форму только во время митоза. Эта форма отдаленно напоминает Х-образную для всех хромосом. Совершенно случайно, что Y-хромосома во время митоза имеет две очень короткие ветви, которые под микроскопом могут выглядеть слившимися и выглядеть как потомок Y-образной ветви. [ 6 ]
Впервые предположение о том, что Х-хромосома участвует в определении пола, было высказано Кларенсом Эрвином МакКлюнгом в 1901 году. Сравнив свои работы по саранче с работами Хенкинга и других, МакКлунг отметил, что только половина сперматозоидов получила Х-хромосому. Он назвал эту хромосому дополнительной хромосомой и настаивал (правильно), что это правильная хромосома, и предположил (ошибочно), что это хромосома, определяющая мужского пола. [ 4 ]
Шаблон наследования
[ редактировать ]Этот раздел в значительной степени или полностью опирается на один источник . ( Август 2021 г. ) |

Люк Хатчисон заметил, что ряд возможных предков на линии наследования Х-хромосомы в данном наследственном поколении следует последовательности Фибоначчи . [ 7 ] Особь мужского пола имеет Х-хромосому, которую он получил от матери, и Y-хромосому , которую он получил от отца. Самец считается «происхождением» его собственной Х-хромосомы ( ), а в поколении его родителей его Х-хромосома происходила от одного родителя ( ). Мать мужчины получила одну Х-хромосому от своей матери (бабушки сына по материнской линии) и одну от отца (дедушки сына по материнской линии), поэтому два дедушки и бабушки внесли свой вклад в Х-хромосому потомка мужского пола ( ). Дедушка по материнской линии получил свою Х-хромосому от своей матери, а бабушка по материнской линии получила Х-хромосомы от обоих своих родителей, поэтому три прадеда и прадеда внесли свой вклад в Х-хромосому потомка мужского пола ( ). Пять прапрабабушек и дедушек внесли свой вклад в Х-хромосому потомка мужского пола ( ) и т. д. (Обратите внимание, что это предполагает, что все предки данного потомка независимы, но если какая-либо генеалогия прослеживается достаточно далеко назад во времени, предки начинают появляться в нескольких линиях генеалогии, пока в конечном итоге основатель популяции не появится в все линии родословной.)





Люди
[ редактировать ]Функция
[ редактировать ]
Х-хромосома человека насчитывает более 153 миллионов пар оснований (строительный материал ДНК ). Он представляет собой около 800 генов, кодирующих белок, по сравнению с Y-хромосомой, содержащей около 70 генов из 20 000–25 000 генов в геноме человека. У каждого человека в каждой клетке обычно имеется одна пара половых хромосом. Женщины обычно имеют две Х-хромосомы, тогда как мужчины обычно имеют одну Х-хромосому и одну Y-хромосому . И мужчины, и женщины сохраняют одну из Х-хромосом от матери, а женщины сохраняют вторую Х-хромосому от отца. Поскольку отец сохраняет свою Х-хромосому от матери, женщина имеет одну Х-хромосому от бабушки по отцовской линии (по отцовской линии) и одну Х-хромосому от матери. Этот шаблон наследования соответствует числам Фибоначчи на заданной наследственной глубине. [ нужна ссылка ]
Генетические нарушения , возникающие из-за мутаций в генах Х-хромосомы, описываются как Х-сцепленные . Если Х-хромосома содержит ген генетического заболевания, он всегда вызывает заболевание у пациентов мужского пола, поскольку у мужчин есть только одна Х-хромосома и, следовательно, только одна копия каждого гена. Вместо этого женщинам для заболевания необходимы обе Х-хромосомы, и в результате они потенциально могут быть только носителями генетического заболевания, поскольку их вторая Х-хромосома переопределяет первую. Например, гемофилия А и В и врожденная красно-зеленая дальтонизм таким образом в семьях передаются .
Х-хромосома несет в себе сотни генов, но немногие из них (если таковые имеются) имеют какое-либо отношение непосредственно к определению пола. На ранних стадиях эмбрионального развития у самок одна из двух Х-хромосом постоянно инактивируется почти во всех соматических клетках (клетках, кроме яйцеклеток и сперматозоидов ). Это явление называется Х-инактивацией или лионизацией и создает тельце Барра . Если бы Х-инактивация в соматической клетке означала полную дефункционализацию одной из Х-хромосом, это гарантировало бы, что у женщин, как и у мужчин, в каждой соматической клетке будет только одна функциональная копия Х-хромосомы. Ранее предполагалось, что это так. Однако недавние исследования показывают, что тельце Барра может быть более биологически активным, чем предполагалось ранее. [ 8 ]
Частичная инактивация Х-хромосомы обусловлена репрессивным гетерохроматином , который уплотняет ДНК и предотвращает экспрессию большинства генов. Уплотнение гетерохроматина регулируется репрессивным комплексом Polycomb 2 ( PRC2 ). [ 9 ]
Гены
[ редактировать ]Количество генов
[ редактировать ]Ниже приведены некоторые оценки количества генов Х-хромосомы человека. Поскольку исследователи используют разные подходы к аннотированию генома, их прогнозы количества генов в каждой хромосоме различаются (технические подробности см. в разделе «Прогнозирование генов» ). Среди различных проектов проект совместного консенсусного кодирования последовательностей ( CCDS ) использует чрезвычайно консервативную стратегию. Таким образом, прогноз числа генов CCDS представляет собой нижнюю границу общего количества генов, кодирующих человеческие белки. [ 10 ]
| По оценкам | Гены, кодирующие белки | Некодирующие гены РНК | Псевдогены | Источник | Дата выпуска |
|---|---|---|---|---|---|
| CCDS | 804 | — | — | [ 11 ] | 2016-09-08 |
| HGNC | 825 | 260 | 606 | [ 12 ] | 2017-05-12 |
| Вместе | 841 | 639 | 871 | [ 13 ] | 2017-03-29 |
| ЮниПрот | 839 | — | — | [ 14 ] | 2018-02-28 |
| NCBI | 874 | 494 | 879 | [ 15 ] [ 16 ] [ 17 ] | 2017-05-19 |
Список генов
[ редактировать ]Ниже приводится неполный список генов X-хромосомы человека. Полный список можно найти по ссылке в информационном окне справа.
- AD16 : кодирует белок 16 болезни Альцгеймера.
- AIC : кодирующий белок AIC
- APOO : кодирующий белок аполипопротеин O
- ARMCX6 : кодирующий повтор белка Armadillo, содержащий X-связанный 6.
- BEX1 : кодирующий белок, экспрессируемый в мозге Х-связанный белок 1.
- BEX2 : кодирующий белок, экспрессируемый в мозге Х-связанный белок 2.
- BEX4 : кодирующий белок, экспрессируемый мозгом, X-связанный 4.
- CCDC120 : кодирующий белок Домен спиральной спирали, содержащий белок 120.
- CCDC22 : кодирующий белок спиральный домен, содержащий 22
- CD99L2 : антигеноподобный белок 2 CD99.
- CDR1-AS : кодирующий антисмысловую РНК белка CDR1.
- CFAP47 : кодирующий белок 47, связанный с ресничками и жгутиками.
- CHRDL1 : кодирующий белок Chordin-подобный 1
- CMTX2, кодирующий белок невропатии Шарко-Мари-Тута, Х-сцепленный 2 (рецессивный)
- CMTX3, кодирующий белок невропатии Шарко-Мари-Тута, Х-связанный 3 (доминантный)
- CT45A5 : кодирующий белок семейства 45 антигенов рака/семенников, член A5.
- CT55 : кодирующий белок антиген рака/семенников 55.
- CXorf36 : кодирующий гипотетический белок LOC79742.
- CXorf57 : кодирующий белок открытая рамка считывания 57 хромосомы X.
- CXorf40A : открытая рамка считывания X-хромосомы 40.
- CXorf49 : открытая рамка считывания хромосомы X 49. кодирующий белок.
- CXorf66 : кодирующий белок Открытая рамка считывания 66 хромосомы X
- CXorf67 : кодирующий белок Неохарактеризованный белок CXorf67
- DACH2 : кодирующий белок гомолог 2 таксы
- EFHC2 : кодирующий домен белка EF-hand (С-конец), содержащий 2
- ERCC6L, кодирующий белок ERCC эксцизионной репарации 6, геликаза контрольной точки сборки веретена
- FAAH2 : гидролаза амидов жирных кислот 2.
- F8A1 : белок интрона 22 фактора VIII.
- FAM104B : кодирующее семейство белков со сходством последовательностей, член B 104.
- FAM120C : кодирующее семейство белков со сходством последовательностей 120C.
- FAM122B : Семейство со сходством последовательностей, 122 члена B.
- FAM122C : кодирующее семейство белков со сходством последовательностей 122C.
- FAM127A : белок 1 CAAX-бокса.
- FAM50A : Семейство со сходством последовательностей, 50 членов A.
- FATE1 : транскриптный белок, экспрессируемый в семенниках плода и взрослого человека.
- FMR1-AS1 : кодирует длинную некодирующую РНК FMR1, антисмысловую РНК 1.
- FRMPD3 : кодирующий белок FERM и домен PDZ, содержащий 3
- FRMPD4 : кодирующий белок FERM и домен PDZ, содержащий 4
- FUNDC1 : кодирующий домен белка FUN14, содержащий 1
- FUNDC2 : белок 2, содержащий домен FUN14.
- GAGE12F : кодирующий белок G-антигена 12F.
- GAGE2A : кодирующий белок G-антигена 2A.
- GATA1 : кодирует фактор транскрипции GATA1.
- GNL3L, кодирующий белок G, белок ядрышка 3, подобный
- GPRASP2 : сортирующий белок 2, связанный с G-белком и рецептором.
- GRIPAP1 : кодирующий белок GRIP1-ассоциированный белок 1
- GRDX : кодирующий белок болезни Грейвса, восприимчивость к Х-связанному
- HDHD1A : кодирующий ферменту галокислотной дегалогеназы. белок 1A, содержащий домен гидролазы, подобный
- HS6ST2 : кодирующий белок гепарансульфат-6-О-сульфотрансфераза 2.
- ITM2A : кодирующий белок Интегральный мембранный белок 2A
- LAS1L : кодирующий белок LAS1-подобный белок
- LOC101059915 : кодирующий белок *LOC101059915.
- MAGEA2 : кодирующий белок антиген 2, ассоциированный с меланомой.
- MAGEA5 : кодирующий белок семейства антигенов меланомы A, 5.
- MAGEA8 : кодирующий белок семейства антигенов меланомы A, 8.
- MAGED4B : кодирующий белок антиген D4, ассоциированный с меланомой.
- MAGT1 : кодирующий белок белок-переносчик магния 1.
- MAGED4 : кодирующий белок члена семейства MAGE D4.
- МАО-А : моноаминоксидаза А.
- МАО-B : моноаминоксидаза B.
- MAP3K15 : кодирующий белок митоген-активируемая протеинкиназная киназа 15.
- MBNL3 : кодирующий белок 3, подобный мышечному слепому белку.
- MBTPS2 : кодирующий фермент протеазу сайта 2 мембраносвязанного фактора транскрипции.
- MCT-1 : кодирующий белок MCTS1, фактор повторной инициации и высвобождения.
- MIR106A : кодирует микроРНК. МикроРНК 106.
- MIR222 : кодирует микроРНК. МикроРНК 222.
- MIR223 : кодирующий белок микроРНК 223.
- MIR361 : кодирует микроРНК. МикроРНК 361.
- MIR503 : кодирует микроРНК. МикроРНК 503.
- MIR6087 : кодирует микроРНК. МикроРНК 6087.
- MIR660 : кодирующая микроРНК МикроРНК 660
- MIRLET7F2 : кодирующий белок микроРНК let-7f-2.
- MORF4L2 : кодирующий белок 2, подобный фактору смертности 4.
- MOSPD1 : кодирующий белок. Домен подвижного сперматозоида, содержащий 1
- MOSPD2 : кодирующий белок. Домен подвижного сперматозоида, содержащий 2
- NAP1L3 : кодирующий белок сборки нуклеосомы 1, подобный 3.
- NBDY : кодирующий белок. Отрицательный регулятор ассоциации P-тел.
- NKRF : кодирующий белок NF-каппа-B-репрессирующий фактор.
- NRK : кодирующий фермент Nik-родственную протеинкиназу.
- OPN1LW : длинноволновой (красный конус) чувствительный опсин.
- OPN1MW : чувствительный к средней волне (зеленый конус) опсин.
- OTUD5 : кодирующий белок деубиквитиназу 5 OTU.
- PASD1 : кодирующий белок 1, содержащий домен PAS
- PAGE1 : кодирующий белок члена 1 семейства PAGE.
- PAGE2B : кодирует белок 2B члена семейства PAGE.
- PBDC1 : кодирует белок с неустановленной функцией.
- PCYT1B : кодирующий фермент холинфосфатцитидилилтрансферазу B.
- PIN4 : кодирующий фермент пептидил-пролил-цис-транс-изомеразу, взаимодействующую с NIMA 4
- PLAC1 : кодирующий белок Плацентарно-специфичный белок 1.
- PLP2 : кодирующий белок Протеолипидный белок 2
- PRR32 : кодирующий белок PRR32.
- RPA4 : кодирующий белок Репликационный белок A, субъединица 30 кДа.
- RPS6KA6 : кодирующий белок Рибосомальная протеинкиназа S6, 90 кДа, полипептид 6.
- RRAGB : кодирующий белок Ras-связывающий GTP-связывающий белок B.
- RTL3 : кодирующий белок ретротранспозон Gag, подобный 3.
- SFRS17A : кодирующий фактор сплайсинга белка, богатый аргинином/серином 17A.
- SLC38A5 : кодирующий белок, член 5 семейства растворенных носителей 38
- SLITRK2 : кодирующий белок SLIT и NTRK-подобный белок 2.
- SMARCA1 : кодирующий белок. Вероятный глобальный активатор транскрипции SNF2L1.
- SMS : кодирующий фермент сперминсинтазу.
- SPANXN1 : кодирующий белок N1 семейства SPANX.
- SPANXN5 : кодирующий белок N5 семейства SPANX.
- SPG16 : кодирующий белок Спастическая параплегия 16 (сложная, Х-сцепленная рецессивная форма)
- SSR4 : кодирующий белок субъединицу дельта белка, связанного с транслоконом.
- TAF7L : кодирующий белок, связывающий белок TATA-бокс, связанный с фактором 7, подобным
- TCEAL1 : кодирующий белок фактор элонгации транскрипции A, подобный белку 1.
- TCEAL4 : кодирующий белок фактор элонгации транскрипции A, подобный белку 4.
- TENT5D : кодирует белок-концевую нуклеотидилтрансферазу 5D.
- TEX11 : кодирующий белок Testis экспрессируется 11
- THOC2 : кодирует субъединицу 2 комплекса белка THO.
- TMEM29 : кодирующий белок FAM156A.
- TMEM47 : кодирующий белок Трансмембранный белок 47
- TMLHE : кодирующий фермент триметиллизиндиоксигеназу, митохондриальный.
- TNMD, кодирующий белок теномодулин (также называемый тендином, миодулин, Tnmd и TeM)
- TRAPPC2P1, кодирующий субъединицу 2 комплекса белковых частиц белка, осуществляющего транспортировку белка
- TREX2 : кодирующий фермент экзонуклеазу 2 трехосновной репарации.
- TRO : кодирующий белок тропинин.
- TSPYL2 : кодирующий белок, специфичный для семенников белок 2, кодируемый Y-подобным.
- TTC3P1 : кодирующий псевдоген 1 домена 3 повтора тетратрикопептида белка.
- USP51 : кодирующий фермент карбоксилконцевую гидролазу убиквитина 51.
- VSIG1 : кодирующий белок V-set и домен иммуноглобулина, содержащий 1
- YIPF6 : кодирующий белок. Белок YIPF6.
- ZC3H12B : кодирующий белок ZC3H12B
- ZC4H2 : кодирует дефицит белка ZC4H2.
- ZCCHC18 : кодирующий белок типа CCHC с цинковым пальцем, содержащий 18
- ZFP92 : кодирующий белок ZFP92, белок цинкового пальца.
- ZMYM3 : encoding protein Zinc finger MYM-type protein 3
- ZNF157 : кодирующий белок Белок 157 цинкового пальца
- ZNF182, кодирующий белок Белок 182 цинкового пальца
- ZNF275 : кодирующий белок Белок 275 цинкового пальца
- ZNF674 : кодирующий белок белок 674 цинкового пальца.
Структура
[ редактировать ]Это теоретизирует Росс и др. 2005 и Ohno 1967, что Х-хромосома, по крайней мере частично, произошла от аутосомного (не связанного с полом) генома других млекопитающих, о чем свидетельствуют межвидовые выравнивания геномных последовательностей.
Х-хромосома заметно крупнее и имеет более активную область эухроматина , чем ее аналог Y-хромосомы . Дальнейшее сравнение X и Y выявляет области гомологии между ними. Однако соответствующая область Y кажется намного короче и в ней отсутствуют области, которые консервативны в X у всех видов приматов, что подразумевает генетическую дегенерацию Y в этой области. Поскольку у мужчин есть только одна Х-хромосома, у них больше шансов заболеть заболеванием, связанным с Х-хромосомой.
Подсчитано, что около 10% генов, кодируемых Х-хромосомой, связаны с семейством генов «CT», названных так потому, что они кодируют маркеры, обнаруженные как в опухолевых клетках (у онкологических больных), так и в семенниках человека. (у здоровых пациентов). [ 18 ]
Роль в болезни
[ редактировать ]Числовые аномалии
[ редактировать ]- Синдром Клайнфельтера вызван наличием одной или нескольких дополнительных копий Х-хромосомы в клетках мужчины.
- Мужчины с синдромом Клайнфельтера обычно имеют одну дополнительную копию Х-хромосомы в каждой клетке, всего две Х-хромосомы и одна Y-хромосома (47,XXY). У больных мужчин реже встречаются две или три дополнительные Х-хромосомы (48,XXXY или 49,XXXXY) или дополнительные копии X- и Y-хромосом (48,XXYY) в каждой клетке. Дополнительный генетический материал может привести к высокому росту, неспособности к обучению и чтению, а также к другим медицинским проблемам. ребенка Каждая дополнительная Х-хромосома снижает IQ примерно на 15 пунктов. [ 19 ] [ 20 ] а это значит, что средний IQ при синдроме Клайнфельтера в целом находится в пределах нормы, хотя и ниже среднего. Когда дополнительные X- и/или Y-хромосомы присутствуют в 48,XXXY, 48,XXYY или 49,XXXXY, задержки развития и когнитивные трудности могут быть более серьезными и легкая умственная отсталость . может наблюдаться
- Синдром Клайнфельтера также может быть результатом наличия дополнительной Х-хромосомы только в некоторых клетках организма. Эти случаи называются мозаикой 46,XY/47,XXY.
- Этот синдром возникает из-за наличия дополнительной копии Х-хромосомы в каждой женской клетке. Женщины с трисомией Х имеют три Х-хромосомы, всего 47 хромосом на клетку. Средний IQ женщин с этим синдромом составляет 90, тогда как средний IQ здоровых братьев и сестер составляет 100. [ 21 ] Их рост в среднем выше, чем у обычных женщин. Они плодовиты, и их дети не наследуют это заболевание. [ 22 ]
- женщины с более чем одной дополнительной копией Х-хромосомы (48, тетрасомия X или 49, пентасомия X ), но эти состояния встречаются редко. Были идентифицированы
- Это происходит, когда каждая клетка женщины имеет одну нормальную Х-хромосому, а другая половая хромосома отсутствует или изменена. Недостающий генетический материал влияет на развитие и вызывает такие особенности заболевания, как низкий рост и бесплодие.
- Около половины людей с синдромом Тернера имеют моносомию Х (45,Х), что означает, что каждая клетка женского тела имеет только одну копию Х-хромосомы вместо обычных двух копий. Синдром Тернера также может возникнуть, если одна из половых хромосом частично отсутствует или перестроена, а не полностью отсутствует. У некоторых женщин с синдромом Тернера хромосомные изменения наблюдаются только в некоторых клетках. Эти случаи называются мозаикой синдрома Тернера (45,X/46,XX).
Х-сцепленные рецессивные заболевания
[ редактировать ]Сцепление с полом было впервые обнаружено у насекомых, например, в 1910 г., когда Т. Х. Морган обнаружил закономерность наследования мутации белых глаз у Drosophila melanogaster . [ 23 ] Такие открытия помогли объяснить Х-сцепленные заболевания у людей, например, гемофилию А и В, адренолейкодистрофию и красно-зеленую цветовую слепоту .
Другие расстройства
[ редактировать ]Мужской синдром ХХ — редкое заболевание, при котором область SRY Y-хромосомы рекомбинируется и располагается на одной из Х-хромосом. В результате комбинация XX после оплодотворения оказывает тот же эффект, что и комбинация XY, в результате чего образуется самец. Однако другие гены Х-хромосомы также вызывают феминизацию.
Х-сцепленная эндотелиальная дистрофия роговицы — чрезвычайно редкое заболевание роговицы, связанное с областью Xq25. Эпителиальная дистрофия роговицы Лиша связана с Xp22.3.
Мегалокорнеа 1 связана с Xq21.3-q22. [ нужна медицинская ссылка ]
Адренолейкодистрофия — редкое и смертельное заболевание, передающееся матерью через Х-клетку. Он поражает только мальчиков в возрасте от 5 до 10 лет и разрушает защитные клетки, окружающие нервы, миелин , в мозге. Женщина-носитель почти не проявляет никаких симптомов, поскольку у женщин есть копия Х-клетки. Это заболевание приводит к тому, что некогда здоровый мальчик теряет способность ходить, говорить, видеть, слышать и даже глотать. В течение 2 лет после постановки диагноза большинство мальчиков с адренолейкодистрофией умирают.
Цитогенетическая полоса
[ редактировать ]
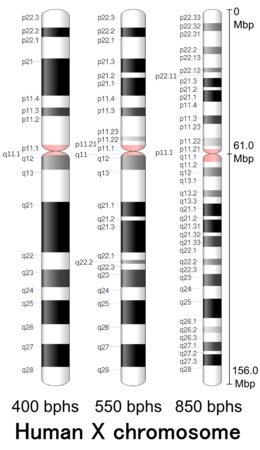
| Хр. | Рука [ 28 ] | Группа [ 29 ] | ISCN начинать [ 30 ] |
ISCN останавливаться [ 30 ] |
Базовая пара начинать |
Базовая пара останавливаться |
Пятно [ 31 ] | Плотность |
|---|---|---|---|---|---|---|---|---|
| Х | п | 22.33 | 0 | 323 | 1 | 4,400,000 | хороший | |
| Х | п | 22.32 | 323 | 504 | 4,400,001 | 6,100,000 | GPS | 50 |
| Х | п | 22.31 | 504 | 866 | 6,100,001 | 9,600,000 | хороший | |
| Х | п | 22.2 | 866 | 1034 | 9,600,001 | 17,400,000 | GPS | 50 |
| Х | п | 22.13 | 1034 | 1345 | 17,400,001 | 19,200,000 | хороший | |
| Х | п | 22.12 | 1345 | 1448 | 19,200,001 | 21,900,000 | GPS | 50 |
| Х | п | 22.11 | 1448 | 1577 | 21,900,001 | 24,900,000 | хороший | |
| Х | п | 21.3 | 1577 | 1784 | 24,900,001 | 29,300,000 | GPS | 100 |
| Х | п | 21.2 | 1784 | 1862 | 29,300,001 | 31,500,000 | хороший | |
| Х | п | 21.1 | 1862 | 2120 | 31,500,001 | 37,800,000 | GPS | 100 |
| Х | п | 11.4 | 2120 | 2430 | 37,800,001 | 42,500,000 | хороший | |
| Х | п | 11.3 | 2430 | 2624 | 42,500,001 | 47,600,000 | GPS | 75 |
| Х | п | 11.23 | 2624 | 2948 | 47,600,001 | 50,100,000 | хороший | |
| Х | п | 11.22 | 2948 | 3129 | 50,100,001 | 54,800,000 | GPS | 25 |
| Х | п | 11.21 | 3129 | 3206 | 54,800,001 | 58,100,000 | хороший | |
| Х | п | 11.1 | 3206 | 3297 | 58,100,001 | 61,000,000 | акцент | |
| Х | д | 11.1 | 3297 | 3491 | 61,000,001 | 63,800,000 | акцент | |
| Х | д | 11.2 | 3491 | 3620 | 63,800,001 | 65,400,000 | хороший | |
| Х | д | 12 | 3620 | 3827 | 65,400,001 | 68,500,000 | GPS | 50 |
| Х | д | 13.1 | 3827 | 4137 | 68,500,001 | 73,000,000 | хороший | |
| Х | д | 13.2 | 4137 | 4292 | 73,000,001 | 74,700,000 | GPS | 50 |
| Х | д | 13.3 | 4292 | 4447 | 74,700,001 | 76,800,000 | хороший | |
| Х | д | 21.1 | 4447 | 4732 | 76,800,001 | 85,400,000 | GPS | 100 |
| Х | д | 21.2 | 4732 | 4809 | 85,400,001 | 87,000,000 | хороший | |
| Х | д | 21.31 | 4809 | 5107 | 87,000,001 | 92,700,000 | GPS | 100 |
| Х | д | 21.32 | 5107 | 5184 | 92,700,001 | 94,300,000 | хороший | |
| Х | д | 21.33 | 5184 | 5430 | 94,300,001 | 99,100,000 | GPS | 75 |
| Х | д | 22.1 | 5430 | 5701 | 99,100,001 | 103,300,000 | хороший | |
| Х | д | 22.2 | 5701 | 5843 | 103,300,001 | 104,500,000 | GPS | 50 |
| Х | д | 22.3 | 5843 | 6050 | 104,500,001 | 109,400,000 | хороший | |
| Х | д | 23 | 6050 | 6322 | 109,400,001 | 117,400,000 | GPS | 75 |
| Х | д | 24 | 6322 | 6619 | 117,400,001 | 121,800,000 | хороший | |
| Х | д | 25 | 6619 | 7059 | 121,800,001 | 129,500,000 | GPS | 100 |
| Х | д | 26.1 | 7059 | 7253 | 129,500,001 | 131,300,000 | хороший | |
| Х | д | 26.2 | 7253 | 7395 | 131,300,001 | 134,500,000 | GPS | 25 |
| Х | д | 26.3 | 7395 | 7602 | 134,500,001 | 138,900,000 | хороший | |
| Х | д | 27.1 | 7602 | 7808 | 138,900,001 | 141,200,000 | GPS | 75 |
| Х | д | 27.2 | 7808 | 7886 | 141,200,001 | 143,000,000 | хороший | |
| Х | д | 27.3 | 7886 | 8145 | 143,000,001 | 148,000,000 | GPS | 100 |
| Х | д | 28 | 8145 | 8610 | 148,000,001 | 156,040,895 | хороший |

Исследовать
[ редактировать ]без пробелов В июле 2020 года ученые сообщили о первой полной сборке Х- хромосомы человека . [ 32 ] [ 33 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- Более ранние версии этой статьи содержат материалы из Национальной медицинской библиотеки , входящей в состав Национальных институтов здравоохранения (США), которые, как публикация правительства США, находятся в свободном доступе.
- ^ Том Страчан; Эндрю Рид (2 апреля 2010 г.). Молекулярная генетика человека . Гирляндная наука. п. 45. ИСБН 978-1-136-84407-2 .
- ^ Перейти обратно: а б с Страница оформления генома, NCBI. Данные идеограммы Homo sapience (850 ударов в час, сборка GRCh38.p3) . Последнее обновление 3 июня 2014 г. Проверено 26 апреля 2017 г.
- ^ Энджер, Натали (1 мая 2007 г.). «Для материнской Х-хромосомы пол — это только начало» . Нью-Йорк Таймс . Проверено 1 мая 2007 г.
- ^ Перейти обратно: а б Джеймс Шварц, В поисках гена: от Дарвина к ДНК , страницы 155–158, издательство Гарвардского университета, 2009 г. ISBN 0674034910
- ^ Дэвид Бейнбридж, «X в сексе: как X-хромосома контролирует нашу жизнь» , страницы 3–5, издательство Гарвардского университета, 2003 г. ISBN 0674016211 .
- ^ Бейнбридж, страницы 65-66.
- ^ Перейти обратно: а б Хатчисон, Люк (сентябрь 2004 г.). «Выращивание генеалогического древа: сила ДНК в восстановлении семейных отношений» (PDF) . Материалы Первого симпозиума по биоинформатике и биотехнологии (БИОТ-04) . Проверено 03 сентября 2016 г.
- ^ Каррел Л., Уиллард Х (2005). «Профиль Х-инактивации обнаруживает значительную вариабельность экспрессии Х-связанных генов у женщин». Природа . 434 (7031): 400–4. Бибкод : 2005Natur.434..400C . дои : 10.1038/nature03479 . ПМИД 15772666 . S2CID 4358447 .
- ^ Венети З., Гкуску К.К., Элиопулос А.Г. (июль 2017 г.). «Репрессорный комплекс Polycomb 2 при геномной нестабильности и раке» . Int J Mol Sci . 18 (8): 1657. doi : 10.3390/ijms18081657 . ПМЦ 5578047 . ПМИД 28758948 .
- ^ Пертеа М., Зальцберг С.Л. (2010). «Между курицей и виноградом: оценка количества генов человека» . Геном Биол . 11 (5): 206. doi : 10.1186/gb-2010-11-5-206 . ПМК 2898077 . ПМИД 20441615 .
- ^ «Результаты поиска — X[CHR] И «Homo sapiens»[Организм] И («имеет ccds»[Свойства] И живой[prop]) — Ген» . НКБИ . CCDS Release 20 для Homo Sapiens . 08.09.2016 . Проверено 28 мая 2017 г.
- ^ «Статистика и загрузки для хромосомы X» . Комитет по генной номенклатуре Хьюго . 12 мая 2017 г. Архивировано из оригинала 29 июня 2017 г. Проверено 19 мая 2017 г.
- ^ «Хромосома X: Краткое описание хромосом - Homo sapiens» . Ансамбль Выпуск 88 . 2017-03-29 . Проверено 19 мая 2017 г.
- ^ «Хромосома человека X: записи, названия генов и перекрестные ссылки на MIM» . ЮниПрот . 28 февраля 2018 г. Проверено 16 марта 2018 г.
- ^ «Результаты поиска — X[CHR] И «Homo sapiens»[Организм] И («кодирующий белок генотипа»[Свойства] И живой[prop]) — Ген» . НКБИ . 19 мая 2017 г. Проверено 20 мая 2017 г.
- ^ "Результаты поиска - X[CHR] И "Homo sapiens"[Организм] И ( ("genetype miscrna"[Свойства] OR "генотип ncrna"[Свойства] OR "генотип rrna"[Свойства] OR "генотип trna"[Свойства] ИЛИ "генотип scrna"[Свойства] ИЛИ "генотип snrna"[Свойства] ИЛИ "генотип snrna"[Свойства]) НЕ "генотип, кодирующий белок"[Свойства] И живой[prop]) - Ген" . НКБИ . 19 мая 2017 г. Проверено 20 мая 2017 г.
- ^ «Результаты поиска — X[CHR] И «Homo sapiens»[Организм] И («псевдогенный тип»[Свойства] И живой[prop]) — Ген» . НКБИ . 19 мая 2017 г. Проверено 20 мая 2017 г.
- ^ Росс М. и др. (2005). «Последовательность ДНК Х-хромосомы человека» . Природа . 434 (7031): 325–37. Бибкод : 2005Natur.434..325R . дои : 10.1038/nature03440 . ПМЦ 2665286 . ПМИД 15772651 .
- ^ Гарольд Чен; Ян Кранц; Мэри Л. Виндл; Маргарет М. Макговерн; Пол Д. Петри; Брюс Бюлер (22 февраля 2013 г.). «Патофизиология синдрома Клайнфельтера» . Медскейп . Проверено 18 июля 2014 г.
- ^ Висоотсак Дж., Грэм Дж.М. (2006). «Синдром Клайнфельтера и другие половые хромосомные анеуплоидии» . Orphanet J Редкий дис . 1:42 . дои : 10.1186/1750-1172-1-42 . ПМЦ 1634840 . ПМИД 17062147 .
- ^ Бендер Б., Пак М., Зальбенблатт Дж., Робинсон А. (1986). Смит С. (ред.). Когнитивное развитие детей с аномалиями половых хромосом . Сан-Диего: College Hill Press. стр. 175–201.
- ^ «Синдром тройного Х» . Домашний справочник по генетике . 14 июля 2014 г. Проверено 18 июля 2014 г.
- ^ Морган, TH (1910). «Наследование, ограниченное полом у дрозофилы» . Наука . 32 (812): 120–122. Бибкод : 1910Sci....32..120M . дои : 10.1126/science.32.812.120 . ПМИД 17759620 .
- ^ Страница оформления генома, NCBI. Данные идеограммы Homo sapience (400 ударов в час, сборка GRCh38.p3) . Последнее обновление 4 марта 2014 г. Проверено 26 апреля 2017 г.
- ^ Страница оформления генома, NCBI. Данные идеограммы Homo sapience (550 ударов в час, сборка GRCh38.p3) . Последнее обновление 11 августа 2015 г. Проверено 26 апреля 2017 г.
- ^ Международный постоянный комитет по цитогенетической номенклатуре человека (2013 г.). ISCN 2013: Международная система цитогенетической номенклатуры человека (2013) . Каргерское медицинское и научное издательство. ISBN 978-3-318-02253-7 .
- ^ Сетакулвичай, В.; Манитпорнсут, С.; Вибунрат, М.; Лилакиацакун, В.; Ассавамакин, А.; Тонгсима, С. (2012). «Оценка разрешения на уровне полос изображений хромосом человека». 2012 Девятая Международная конференция по информатике и программной инженерии (JCSSE) . стр. 276–282. дои : 10.1109/JCSSE.2012.6261965 . ISBN 978-1-4673-1921-8 . S2CID 16666470 .
- ^ " p ": Короткая рука; « q »: Длинная рука.
- ^ Номенклатуру цитогенетических полос см. в статье locus .
- ^ Перейти обратно: а б Эти значения (начало/конец ISCN) основаны на длине полос/идеограмм из книги ISCN «Международная система цитогенетической номенклатуры человека» (2013). Произвольная единица .
- ^ gpos : область, положительно окрашенная G-полосами , обычно богатая АТ и бедная генами; gneg : область, отрицательно окрашенная G-полосами, обычно богатая CG и богатая генами; acen Центромера . var : переменная область; стебель : Стебель.
- ^ «Ученые впервые достигли полной сборки Х-хромосомы человека» . физ.орг . Проверено 16 августа 2020 г. .
- ^ Мига, Карен Х .; Корень, Сергей; Ри, уголь; Фоллгер, Митчелл Р.; Гершман, Ариэль; Бзикадзе, Андрей; Брукс, Шелис; Хау, Эдмунд; Порубский, Дэвид; Логсдон, Гленнис А.; Шнайдер, Валери А.; Потапова Тамара; Вуд, Джонатан; Чоу, Уильям; Армстронг, Джоэл; Фредриксон, Жанна; Пак, Евгения; Тигби, Кристофер; Кремицкий, Милленн; Маркович, Кристофер; Мадуро, Валери; доктор Амалия; Буффар, Жерар Г.; Чанг, Александр М.; Хансен, Нэнси Ф.; Уилферт, Эми Б.; Тибо-Ниссен, Франсуаза; Шмитт, Энтони Д.; Белтон, Джон-Мэтью; Сельварадж, Сиддарт; Деннис, Меган Ю.; Сото, Даниэла С.; Сахасрабудхе, Рута; Итак, Гюльхан; Быстрее, Джош; Ломан, Николас Дж.; Холмс, Надин; Свободный, Мэтью; Сурти, Урваши; Риски, Роза Ана; Линдси, Тина А. Грейвс; Фултон, Роберт; Холл, Ира; Патен, Бенедикт; Хау, Керстин; Тимп, Уинстон; Янг, Алиса; Малликин, Джеймс С.; Певзнер, Пол А.; Гертон, Дженнифер Л.; Салливан, Бет А.; Эйхлер, Эван Э.; Филлиппи, Адам М. (14 июля 2020 г.). «Сборка теломер-теломеры полной Х-хромосомы человека» . Природа . 585 (7823): 79–84. Бибкод : 2020Природа.585...79М . дои : 10.1038/s41586-020-2547-7 . ISSN 1476-4687 . ПМЦ 7484160 . ПМИД 32663838 .
Внешние ссылки
[ редактировать ]- Национальные институты здравоохранения. «Х-хромосома» . Домашний справочник по генетике . Архивировано из оригинала 8 июля 2007 г. Проверено 6 мая 2017 г.
- «Х-хромосома» . Информационный архив проекта «Геном человека», 1990–2003 гг . Проверено 6 мая 2017 г.
| Базы данных органов управления : Национальные |
|---|