ноцицептор
| ноцицептор | |
|---|---|
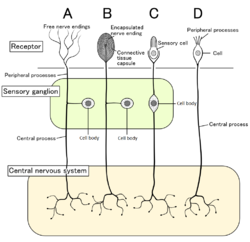 Четыре типа сенсорных нейронов и их рецепторных клеток. Ноцицепторы показаны как свободные нервные окончания типа А. | |
| Идентификаторы | |
| МеШ | D009619 |
| Анатомическая терминология | |
Ноцицептор , (от латинского nocere «причинять вред или ранить»; букв. « рецептор боли » ) — это сенсорный нейрон который реагирует на повреждающие или потенциально вредные стимулы, посылая сигналы «возможной угрозы». [ 1 ] [ 2 ] [ 3 ] в спинной и головной мозг. Мозг создает ощущение боли, чтобы направить внимание на ту или иную часть тела, чтобы можно было смягчить угрозу; этот процесс называется ноцицепцией .
Терминология
[ редактировать ]Ноцицепция и боль обычно вызываются только давлением и температурой, потенциально повреждающими ткани. Этот барьер или порог контрастирует с более чувствительными зрительными, слуховыми, обонятельными, вкусовыми и соматосенсорными реакциями на раздражители. Переживание боли индивидуально и может подавляться стрессом или обостряться ожиданием. Простая активация ноцицептора не всегда приводит к ощущению боли, поскольку последняя также зависит от частоты потенциалов действия , интеграции пре- и постсинаптических сигналов и влияний со стороны высших или центральных процессов. [ 4 ]
Научное исследование
[ редактировать ]Ноцицепторы были открыты Чарльзом Скоттом Шеррингтоном в 1906 году. Раньше учёные считали, что животные подобны механическим устройствам, преобразующим энергию сенсорных стимулов в двигательные реакции. Шеррингтон провел множество различных экспериментов, чтобы продемонстрировать, что разные типы стимуляции афферентного нервного волокна приводят рецептивного поля к разным реакциям. Некоторые интенсивные стимулы вызывают рефлекторное отстранение , определенные вегетативные реакции и боль . Специфические рецепторы для этих интенсивных стимулов были названы ноцицепторами. [ 5 ]
Исследования ноцицепторов проводились на людях, находящихся в сознании, а также на суррогатных моделях животных. Этот процесс сложен из-за инвазивных методов, которые могут изменить клеточную активность изучаемых ноцицепторов, невозможности записи с небольших нейрональных структур и неопределенности в модельных системах животных относительно того, следует ли объяснить реакцию болью или каким-либо другим фактором. [ 4 ]
Расположение
[ редактировать ]У млекопитающих ноцицепторы находятся в любой части тела, способной воспринимать вредные раздражители. Внешние ноцицепторы обнаруживаются в таких тканях , как кожа ( кожные ноцицепторы ), роговица и слизистая оболочка . Внутренние ноцицепторы обнаруживаются в различных органах, таких как мышцы , суставы , мочевой пузырь , внутренние органы и пищеварительный тракт. Тела клеток этих нейронов расположены либо в ганглиях дорсальных корешков , либо в ганглиях тройничного нерва . [ 6 ] Тройничные ганглии являются специализированными нервами лица, тогда как ганглии дорсальных корешков связаны с остальной частью тела. Аксоны проникают в периферическую нервную систему и оканчиваются ветвями, образуя рецептивные поля.
Типы и функции
[ редактировать ]Ноцицепторы обычно электрически молчат, когда их не стимулируют. [ 4 ] Периферическое окончание зрелого ноцицептора — это место, где вредные стимулы обнаруживаются и преобразуются в электрическую энергию. [ 7 ] Когда электрическая энергия достигает порогового значения, потенциал действия индуцируется и направляется к центральной нервной системе (ЦНС). Это приводит к череде событий, которая позволяет сознательно осознавать боль. Сенсорная специфичность ноцицепторов определяется высоким порогом только к отдельным особенностям раздражителей. Ноцицепторы срабатывают только тогда, когда достигается высокий порог под воздействием химической, термической или механической среды.
По скорости проведения ноцицепторы делятся на две группы. Аксоны волокон Aδ миелинизированы и позволяют потенциалу действия перемещаться к ЦНС со скоростью от 5 до 30 метров в секунду. Аксоны C-волокна проводят медленнее на скоростях от 0,4 до 2 метров в секунду из-за их меньшего диаметра и незначительной миелинизации аксона или ее отсутствия. [ 8 ] [ 4 ] В результате боль возникает в две фазы: первоначальная чрезвычайно острая боль, связанная с волокнами Аδ, и вторая, более продолжительная и несколько менее интенсивная боль, связанная с волокнами С. Массивное или продолжительное воздействие на С-волокно приводит к прогрессирующему нарастанию дорсального рога спинного мозга; это явление, называемое заводным, похоже на столбняк в мышцах. Заведение увеличивает вероятность большей чувствительности к боли. [ 9 ]
Термальный
[ редактировать ]Тепловые ноцицепторы активируются ядовитым теплом или холодом при различных температурах. Существуют специальные ноцицепторные преобразователи, которые отвечают за то, как и если определенное нервное окончание реагирует на тепловой стимул. Первым был обнаружен TRPV1 , порог которого совпадает с температурой жара и боли 43 °C. Другая температура в тепло-горячем диапазоне определяется более чем одним каналом TRP . Каждый из этих каналов экспрессирует определенный С-концевой домен, который соответствует чувствительности к теплу и горячему. Взаимодействие между всеми этими каналами и то, как уровень температуры определяется как превышающий болевой порог, в настоящее время неизвестны. Холодные стимулы воспринимаются каналами TRPM8 . Его С-концевой домен отличается от термочувствительных TRP. Хотя этот канал соответствует холодным стимулам, до сих пор неизвестно, способствует ли он обнаружению сильного холода. Интересный вывод, связанный с холодовыми раздражителями, заключается в том, что тактильная чувствительность и двигательная функция ухудшаются, в то время как восприятие боли сохраняется.
Механический
[ редактировать ]Механические ноцицепторы реагируют на избыточное давление или механическую деформацию. Они также реагируют на разрезы, которые повреждают поверхность кожи. Реакция на раздражитель обрабатывается корой головного мозга как боль, точно так же, как химические и термические реакции. Эти механические ноцицепторы часто имеют полимодальные характеристики. Поэтому вполне возможно, что некоторые преобразователи тепловых раздражителей одинаковы и для механических раздражителей. То же самое верно и для химических стимулов, поскольку TRPA1, по-видимому, обнаруживает как механические, так и химические изменения. Некоторые механические стимулы могут вызывать высвобождение промежуточных химических веществ, таких как АТФ , который может быть обнаружен пуринергическими рецепторами P2 , или фактор роста нервов , который может быть обнаружен киназой А рецептора тропомиозина (TrkA) . [ 10 ]
Химическая
[ редактировать ]Химические ноцицепторы имеют каналы TRP, которые реагируют на самые разные специи. Тот, который вызывает наибольший отклик и широко тестируется, — это капсаицин . Другими химическими стимуляторами являются раздражители окружающей среды, такие как акролеин , времен Первой мировой войны химическое оружие и компонент сигаретного дыма. Помимо этих внешних стимуляторов, химические ноцицепторы способны обнаруживать эндогенные лиганды и некоторые амины жирных кислот, которые возникают в результате изменений во внутренних тканях. Как и тепловые ноцицепторы, TRPV1 может обнаруживать такие химические вещества, как капсаицин, а также токсины и кислоты пауков. [ 11 ] [ 10 ] Ионные каналы, чувствительные к кислоте (ASIC), также определяют кислотность. [ 10 ]
Спящий/тихий
[ редактировать ]Хотя каждый ноцицептор может иметь множество возможных пороговых уровней, некоторые из них вообще не реагируют на химические, термические или механические раздражители, если только не произошло фактическое повреждение. Их обычно называют «тихими» или «спящими» ноцицепторами, поскольку их реакция возникает только при возникновении воспаления в окружающих тканях. [ 6 ] Их идентифицировали с помощью электрической стимуляции их рецептивного поля . [ 4 ]
Полимодальный
[ редактировать ]Ноцицепторы, реагирующие более чем на один тип раздражителей, называются полимодальными . [ 12 ] Они являются наиболее распространенным типом ноцицепторов С-волокон и экспрессируют богатый набор нейромедиаторов . [ 4 ]
Путь
[ редактировать ]
восходящий
[ редактировать ]Афферентные ноцицептивные волокна (те, которые передают информацию в мозг , а не из него) возвращаются к спинному мозгу , где образуют синапсы в его дорсальном роге . Это ноцицептивное волокно (расположенное на периферии) является нейроном первого порядка. Клетки . спинного рога разделены на физиологически отдельные слои, называемые пластинками Различные типы волокон образуют синапсы в разных слоях и используют либо глутамат , либо вещество Р. в качестве нейромедиатора Волокна Aδ образуют синапсы в пластинках I и V, волокна C соединяются с нейронами пластинок II, волокна Aβ соединяются с пластинками I, III и V. [ 6 ] Достигнув определенной пластинки спинного мозга, ноцицептивные нейроны первого порядка передаются на нейроны второго порядка, которые пересекают срединную линию в передней белой спайке. Нейроны второго порядка затем отправляют свою информацию по двум путям в таламус : медиально-лемнискальную систему дорсального столба и переднелатеральную систему . Первый предназначен больше для регулярных безболезненных ощущений, а второй — для болевых ощущений. Достигнув таламуса, информация обрабатывается в вентральном заднем ядре и отправляется в кору головного мозга через волокна задней ножки внутренней капсулы.
По убыванию
[ редактировать ]Поскольку существует восходящий путь к мозгу, который инициирует сознательное осознание боли, существует также нисходящий путь, который модулирует ощущение боли. Мозг может запросить высвобождение определенных гормонов или химических веществ, которые могут оказывать обезболивающее действие и уменьшать или подавлять болевые ощущения. Областью мозга, которая стимулирует выработку этих гормонов, является гипоталамус . [ 13 ] Этот эффект нисходящего торможения можно продемонстрировать путем электрической стимуляции околоводопроводной серой области среднего мозга или перивентрикулярного ядра . Оба они, в свою очередь, проецируются на другие области, участвующие в регуляции боли, такие как большое ядро шва , которое также получает аналогичные афференты от парагигантоцеллюлярного ретикулярного ядра (НПГ). В свою очередь, большое ядро шва проецируется в область желатинозной субстанции дорсального рога и обеспечивает ощущение спиноталамических входов. Сначала это делается с помощью большого ядра шва, посылающего серотонинергические нейроны к нейронам спинного мозга, которые, в свою очередь, секретируют энкефалин к промежуточным нейронам, ответственным за восприятие боли. [ 14 ] Энкефалин действует путем связывания опиоидных рецепторов , вызывая торможение постсинаптического нейрона, тем самым подавляя боль. [ 10 ] Периакведуктальный серый цвет также содержит опиоидные рецепторы , что объясняет один из механизмов, посредством которого опиоиды, такие как морфин и диацетилморфин, проявляют анальгетический эффект.
Чувствительность
[ редактировать ]Чувствительность ноцицепторов модулируется множеством медиаторов во внеклеточном пространстве, таких как токсичные и воспалительные молекулы. [ 15 ] [ 4 ] Периферическая сенсибилизация представляет собой форму функциональной пластичности ноцицептора. Ноцицептор может превратиться из простого детектора вредных стимулов в детектор безвредных стимулов. В результате раздражители низкой интенсивности, вызванные регулярной деятельностью, вызывают болезненные ощущения. Это широко известно как гипералгезия . Воспаление является одной из частых причин, приводящих к сенсибилизации ноцицепторов. Обычно гипералгезия прекращается, когда воспаление стихает, однако иногда генетические дефекты и/или повторные травмы могут привести к аллодинии : совершенно безвредный раздражитель, такой как легкое прикосновение, вызывает сильную боль. Аллодиния также может быть вызвана повреждением ноцицептора в периферических нервах. Это может привести к деафферентации, что означает развитие различных центральных отростков из выжившего афферентного нерва. В этой ситуации выжившие аксоны дорсальных корешков ноцицепторов могут вступить в контакт со спинным мозгом, изменяя тем самым нормальный входной сигнал. [ 9 ]
Нейронное развитие
[ редактировать ]Ноцицепторы развиваются из нервного гребня стволовых клеток во время эмбриогенеза . Нервный гребень отвечает за большую часть раннего развития позвоночных. Он конкретно отвечает за развитие периферической нервной системы (ПНС). Стволовые клетки нервного гребня отделяются от нервной трубки, когда она закрывается, и ноцицепторы растут из дорсальной части этой ткани нервного гребня. Они формируются на позднем этапе нейрогенеза. Ранее формирующиеся клетки этой области могут стать неболевыми рецепторами, либо проприорецепторами , либо низкопороговыми механорецепторами . Все нейроны, происходящие из нервного гребня, включая эмбриональные ноцицепторы, экспрессируют киназу А рецептора тропомиозина (TrkA), которая является рецептором фактора роста нервов (NGF). Однако факторы транскрипции, определяющие тип ноцицептора, остаются неясными. [ 11 ]
Вслед за сенсорным нейрогенезом происходит дифференцировка и формируются два типа ноцицепторов. Их классифицируют как пептидергические или непептидергические ноцицепторы, каждый из которых экспрессирует особый набор ионных каналов и рецепторов. Их специализация позволяет рецепторам иннервировать различные центральные и периферические мишени. Эта дифференциация происходит как в перинатальном, так и в постнатальном периоде. Непептидергические ноцицепторы отключают TrkA и начинают экспрессировать протоонкоген RET , который является трансмембранным сигнальным компонентом, который обеспечивает экспрессию нейротрофического фактора, происходящего из глиальных клеточных линий (GDNF). Этому переходу способствует фактор транскрипции 1, связанный с runt (RUNX1), который жизненно важен для развития непептидергических ноцицепторов. Напротив, пептидергические ноцицепторы продолжают использовать TrkA и экспрессируют фактор роста совершенно другого типа. В настоящее время проводится множество исследований различий между ноцицепторами. [ 11 ]
У других животных
[ редактировать ]Ноцицепция была зарегистрирована у животных, не являющихся млекопитающими. [ 16 ] включая рыбу [ 17 ] и широкий спектр беспозвоночных , включая пиявок , [ 18 ] черви -нематоды , [ 19 ] морские слизни , [ 20 ] и личинки плодовых мух . [ 21 ] Хотя эти нейроны могут иметь пути и связи с центральной нервной системой, которые отличаются от таковых у ноцицепторов млекопитающих, ноцицептивные нейроны немлекопитающих часто активируются в ответ на те же стимулы, что и у млекопитающих, такие как высокая температура (40 градусов Цельсия или выше), низкий pH , капсаицин и повреждение тканей.
Например, у плодовых мух в ноцицепции играют роль специфические мультидендритные сенсорные нейроны. [ 22 ] У моллюсков ноцицептивные реакции опосредуются педальными сенсорными нейронами. [ 23 ] [ 24 ] С другой стороны, ракообразные используют различные типы сенсорных клеток, включая хордотональные органы и механорецепторы, для обнаружения потенциально повреждающих стимулов (см. также Боль у ракообразных ).
См. также
[ редактировать ]- Капсаицин и механизм его действия на ноцицепторы
- Ноцицептин и ноцицептиновый рецептор
- Пиперин из черного перца
- TRPC Ионный канал
Ссылки
[ редактировать ]- ^ «НОИ – Нейроортопедический институт» . www.noigroup.com . Архивировано из оригинала 17 октября 2018 г. Проверено 13 октября 2017 г.
- ^ «Ноцицепция и боль: в чем разница и почему это важно? - Массаж Сент-Луис, Сент-Луис, Миссури» . www.massage-stlouis.com . Архивировано из оригинала 1 ноября 2018 г. Проверено 13 октября 2017 г.
- ^ Животные НР (8 декабря 2017 г.). Механизмы боли . National Academies Press (США) – через www.ncbi.nlm.nih.gov.
- ^ Jump up to: а б с д и ж г Дубин А.Е., Патапутян А. (ноябрь 2010 г.). «Ноцицепторы: сенсоры болевого пути» . Журнал клинических исследований . 120 (11): 3760–72. дои : 10.1172/JCI42843 . ПМЦ 2964977 . ПМИД 21041958 .
- ^ Шеррингтон К. Интегративное действие нервной системы. Оксфорд: Издательство Оксфордского университета; 1906.
- ^ Jump up to: а б с Джесселл, Томас М., Кандел, Эрик Р., Шварц, Джеймс Х. (1991). Принципы нейробиологии . Норуолк, Коннектикут: Appleton & Lange. стр. 100-1 472–79 . ISBN 978-0-8385-8034-9 .
- ^ Фейн, А. Ноцицепторы: клетки, ощущающие боль http://cell.uchc.edu/pdf/fein/nociceptors_fein_2012.pdf
- ^ Уильямс, С.Дж., Первс, Дейл (2001). Нейронаука . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-742-4 .
- ^ Jump up to: а б Филдс Х.Л., Роуботэм М., Барон Р. (октябрь 1998 г.). «Постгерпетическая невралгия: раздражительные ноцицепторы и деафферентация». Нейробиол. Дис . 5 (4): 209–27. дои : 10.1006/nbdi.1998.0204 . ПМИД 9848092 . S2CID 13217293 .
- ^ Jump up to: а б с д Юань Дж., Брукс Х.Л., Бармен С.М., Барретт К.Э. (2019). Обзор медицинской физиологии Ганонга . Макгроу-Хилл Образование. ISBN 978-1-260-12240-4 .
- ^ Jump up to: а б с Вульф CJ, Ма Кью (август 2007 г.). «Ноцицепторы — детекторы вредных раздражителей» . Нейрон . 55 (3): 353–64. дои : 10.1016/j.neuron.2007.07.016 . ПМИД 17678850 . S2CID 13576368 .
- ^ Фейн А. Ноцицепторы: клетки, ощущающие боль .
- ^ «Путь боли» . Проверено 2 июня 2008 г. [ мертвая ссылка ]
- ^ Зал ME, Зал JE (2021). Учебник Гайтона и Холла по медицинской физиологии (14-е изд.). Филадельфия, Пенсильвания: Сондерс/Эльзевир. ISBN 978-0-323-59712-8 .
- ^ Хучо Т., Левин JD (август 2007 г.). «Сигнальные пути при сенсибилизации: к биологии ноцицепторных клеток» . Нейрон . 55 (3): 365–76. дои : 10.1016/j.neuron.2007.07.008 . ПМИД 17678851 . S2CID 815135 .
- ^ Смит Э.С., Левин Г.Р. (1 декабря 2009 г.). «Ноцицепторы: филогенетический взгляд» . Журнал сравнительной физиологии А. 195 (12): 1089–1106. дои : 10.1007/s00359-009-0482-z . ISSN 1432-1351 . ПМК 2780683 . ПМИД 19830434 .
- ^ Снеддон Л.У., Брейтуэйт В.А., Джентл М.Дж. (2003). «Есть ли у рыб ноцицепторы? Доказательства эволюции сенсорной системы позвоночных» . Труды Лондонского королевского общества B: Биологические науки . 270 (1520): 1115–1121. дои : 10.1098/rspb.2003.2349 . ПМК 1691351 . ПМИД 12816648 .
- ^ Пастор Дж., Сория Б., Бельмонте К. (1996). «Свойства ноцицептивных нейронов сегментарного ганглия пиявки» . Журнал нейрофизиологии . 75 (6): 2268–2279. дои : 10.1152/jn.1996.75.6.2268 . ПМИД 8793740 .
- ^ Виттенбург Н., Баумайстер Р. (1999). «Термическое избегание Caenorhabditis elegans : подход к изучению ноцицепции» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (18): 10477–10482. Бибкод : 1999PNAS...9610477W . дои : 10.1073/pnas.96.18.10477 . ЧВК 17914 . ПМИД 10468634 .
- ^ Ильич П.А., Уолтерс Э.Т. (1997). «Механосенсорные нейроны, иннервирующие сифон Aplysia, кодируют вредные стимулы и проявляют ноцицептивную сенсибилизацию» . Журнал неврологии . 17 (1): 459–469. doi : 10.1523/JNEUROSCI.17-01-00459.1997 . ПМК 6793714 . ПМИД 8987770 .
- ^ Трейси Дж., Дэниел В., Уилсон Р.И., Лоран Г., Бензер С. (2003). « безболезненно , ген дрозофилы необходим для ноцицепции» . Клетка . 113 (2): 261–273. дои : 10.1016/S0092-8674(03)00272-1 . ПМИД 12705873 . S2CID 1424315 .
- ^ Симоно К., Фудзимото А., Цуяма Т., Ямамото-Кочи М., Сато М., Хаттори Ю., Сугимура К., Усуи Т., Кимура Ки, Уэмура Т. (02.10.2009). «Мультидендритные сенсорные нейроны в брюшке взрослой дрозофилы: происхождение, морфология дендритов, а также сегментированная и возрастная запрограммированная гибель клеток» . Нейронное развитие . 4 (1): 37. дои : 10.1186/1749-8104-4-37 . ISSN 1749-8104 . ПМЦ 2762467 . ПМИД 19799768 .
- ^ Эдгар Т.В. (1996-08-01), «Сравнительные и эволюционные аспекты функции ноцицепторов» , Нейробиология ноцицепторов , Oxford University Press, стр. 92–114, doi : 10.1093/acprof:oso/9780198523345.003.0004 , ISBN 978-0-19-852334-5 , получено 21 марта 2024 г.
- ^ Кадет П., Чжу В., Мантионе К.Дж., Баггерман Г., Стефано ГБ (28 февраля 2002 г.). «Холодовой стресс изменяет экспрессию транскриптов мю-опиатных рецепторов в педальных ганглиях Mytilus edulis, определяемую с помощью ОТ-ПЦР в реальном времени и уровней морфина» . Исследования мозга. Молекулярные исследования мозга . 99 (1): 26–33. дои : 10.1016/s0169-328x(01)00342-4 . ISSN 0169-328X . ПМИД 11869805 .