Сериновая протеаза
| Сериновая протеаза | |||
|---|---|---|---|
 Кристаллическая структура бычьего химотрипсина. Каталитические остатки показаны желтыми полосками. Визуализировано из PDB 1CBW . | |||
| Идентификаторы | |||
| Номер ЕС. | 3.4.21.- | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||
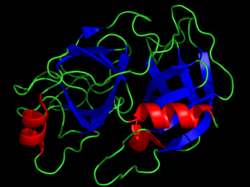
Сериновые протеазы (или сериновые эндопептидазы ) — ферменты , расщепляющие пептидные связи в белках . Серин служит нуклеофильной аминокислотой (фермента) в активном центре . [ 1 ] Они встречаются повсеместно как у эукариот , так и у прокариот . Сериновые протеазы делятся на две широкие категории в зависимости от их структуры: химотрипсиноподобные (трипсиноподобные) и субтилизинподобные . [ 2 ]
Классификация
[ редактировать ]Система классификации протеаз MEROPS насчитывает 16 суперсемейств (по состоянию на 2013 год), каждое из которых содержит множество семейств . Каждое суперсемейство использует каталитическую триаду или диаду в разных белковых складках и, таким образом, представляет собой конвергентную эволюцию каталитического механизма . Большинство из них принадлежит к семейству S1 клана PA (суперсемейства) протеаз .
Для суперсемейств P: суперсемейство, содержащее смесь семейств классов нуклеофилов , S: чисто сериновые протеазы. суперсемья. Внутри каждого суперсемейства семейства обозначаются каталитическим нуклеофилом (S: сериновые протеазы).

Специфичность субстрата
[ редактировать ]Сериновые протеазы характеризуются своеобразной структурой, состоящей из двух доменов бета-бочонка, сходящихся в каталитическом активном центре. Эти ферменты можно далее классифицировать в зависимости от их субстратной специфичности: трипсиноподобные, химотрипсиноподобные или эластазоподобные. [ 5 ]
Трипсиноподобный
[ редактировать ]Трипсиноподобные протеазы расщепляют пептидные связи после положительно заряженной аминокислоты ( лизина или аргинина ). [ 6 ] Эта специфичность обусловлена остатком, который лежит в основании кармана S1 фермента (обычно отрицательно заряженная аспарагиновая кислота или глутаминовая кислота ).
Химотрипсиноподобный
[ редактировать ]Карман S1 химотрипсиноподобных ферментов более гидрофобен, чем у трипсиноподобных протеаз. Это приводит к специфичности к гидрофобным остаткам среднего и большого размера, таким как тирозин , фенилаланин и триптофан .
Тромбиноподобный
[ редактировать ]К ним относятся тромбин , тканеактивирующий плазминоген и плазмин . Было обнаружено, что они играют роль в коагуляции и пищеварении, а также в патофизиологии нейродегенеративных заболеваний, таких как деменция, вызванная болезнью Альцгеймера и Паркинсона. В ядах змей обнаружено множество высокотоксичных изоформ тромбиноподобных сериновых протеаз. [ 7 ]
Эластазоподобный
[ редактировать ]Эластазоподобные протеазы имеют гораздо меньшую щель S1, чем трипсин- или химотрипсиноподобные протеазы. Следовательно, такие остатки, как аланин , глицин и валин, имеют тенденцию быть предпочтительными.
Субтилизиноподобный
[ редактировать ]Субтилизин — сериновая протеаза прокариот . Субтилизин эволюционно не связан с кланом химотрипсина, но имеет тот же каталитический механизм, использующий каталитическую триаду для создания нуклеофильного серина . Это классический пример, используемый для иллюстрации конвергентной эволюции , поскольку один и тот же механизм развивался дважды независимо в ходе эволюции .
Каталитический механизм
[ редактировать ]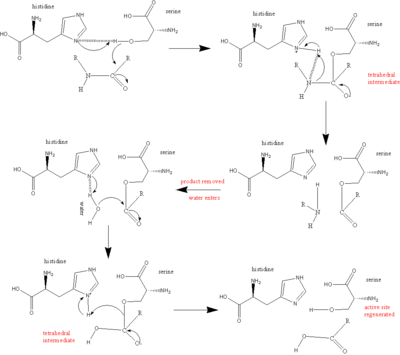
Основным участником каталитического механизма сериновых протеаз является каталитическая триада. Триада расположена в активном центре фермента, где происходит катализ, и сохраняется во всех суперсемействах ферментов сериновых протеаз. Триада представляет собой скоординированную структуру, состоящую из трех аминокислот : His 57, Ser 195 (отсюда и название «сериновая протеаза») и Asp 102. Каждая из этих трех ключевых аминокислот играет важную роль в расщепляющей способности протеаз. Хотя аминокислотные члены триады расположены далеко друг от друга в последовательности белка, из-за сворачивания они будут очень близки друг к другу в сердце фермента. Особая геометрия членов триады весьма характерна для их конкретной функции: было показано, что положение всего четырех точек триады характеризует функцию содержащего их фермента. [ 8 ]
В случае катализа возникает упорядоченный механизм, при котором образуется несколько промежуточных продуктов. Катализ расщепления пептида можно рассматривать как катализ «пинг-понга» , при котором связывается субстрат (в данном случае расщепляемый полипептид), высвобождается продукт (С-концевая «половина» пептида с аминогруппой видимый), связывается другой субстрат (в данном случае вода) и высвобождается другой продукт (видима N-концевая «половина» пептида с карбоксильной группой).
Каждая аминокислота в триаде выполняет в этом процессе определенную задачу:
- Серин карбонильный которая способна действовать как нуклеофил , атакуя углерод разрезаемой имеет группу -ОН , пептидной связи субстрата.
- Пара электронов на азоте гистидина обладает способностью принимать водород из группы серина -ОН, координируя таким образом атаку пептидной связи .
- Карбоксильная аспарагиновой группа кислоты, в свою очередь, образует водородные связи с гистидином , делая атом азота, упомянутый выше, гораздо более электроотрицательным .
Всю реакцию можно резюмировать следующим образом:
- Полипептидный субстрат связывается с поверхностью фермента сериновой протеазы таким образом, что разрезаемая связь вставляется в активный центр фермента, при этом карбонильный углерод этой связи располагается рядом с нуклеофильным серином .
- Серин водород от -ОН атакует карбонильный углерод, а азот гистидина принимает -ОН [серина], и пара электронов от двойной связи карбонильного кислорода переходит к кислороду. В результате образуется тетраэдрический интермедиат.
- Связь, соединяющая азот и углерод в пептидной связи, теперь разорвана. Ковалентные электроны, создающие эту связь, атакуют водород гистидина , разрывая связь. Электроны, которые ранее переместились из карбонильной двойной связи кислорода, возвращаются от отрицательного кислорода, чтобы воссоздать связь, образуя промежуточный ацил-фермент.
- Теперь в реакцию вступает вода. Вода заменяет N-конец расщепленного пептида и атакует карбонильный углерод. И снова электроны двойной связи переходят к кислороду, делая его отрицательным, поскольку образуется связь между кислородом воды и углеродом. Это координируется азотом гистидина , который принимает протон из воды. В целом это создает еще один тетраэдрический промежуточный продукт.
- В заключительной реакции связь, образовавшаяся на первом этапе между серином и карбонильным углеродом, движется, атакуя водород, который только что приобрел гистидин . углерод с дефицитом электронов Карбонильный повторно образует двойную связь с кислородом. В результате С-конец пептида теперь выбрасывается.
Дополнительные стабилизирующие эффекты
[ редактировать ]Было обнаружено, что дополнительные аминокислоты протеазы, Gly 193 и Ser 195 , участвуют в создании так называемой оксианионной дырки . И Gly 193 , и Ser 195 могут отдавать водороды основной цепи для образования водородных связей. Когда тетраэдрический интермедиат образуется на шаге 1 и шаге 3, отрицательный ион кислорода, приняв электроны от двойной карбонильной связи, идеально вписывается в оксианионную дырку. Фактически, сериновые протеазы преимущественно связывают переходное состояние , и общая структура благоприятствует, снижая энергию активации реакции. Это «предпочтительное связывание» отвечает за большую часть каталитической эффективности фермента.
Регуляция активности сериновой протеазы
[ редактировать ]Организмы-хозяева должны обеспечить адекватную регуляцию активности сериновых протеаз. Это достигается за счет необходимости начальной активации протеазы и секреции ингибиторов.
Активация зимогена
[ редактировать ]Зимогены обычно являются неактивными предшественниками фермента. Если бы пищеварительные ферменты были активны при синтезе, они немедленно начали бы пережевывать синтезирующие органы и ткани. Острый панкреатит – это состояние, при котором происходит преждевременная активация пищеварительных ферментов поджелудочной железы, что приводит к самоперевариванию (аутолизу). Это также усложняет посмертные исследования , поскольку поджелудочная железа часто переваривает себя прежде, чем ее можно будет оценить визуально.
Зимогены представляют собой крупные неактивные структуры, способные распадаться на части или превращаться в более мелкие активированные ферменты. Отличие зимогенов от активированных ферментов заключается в том, что активный центр катализа зимогенов искажен. В результате полипептид-субстрат не может эффективно связываться, и протеолиз не происходит. Только после активации, в ходе которой изменяются конформация и структура зимогена и открывается активный центр, может произойти протеолиз .
| Зимоген | Фермент | Примечания |
|---|---|---|
| Трипсиноген | трипсин | При попадании трипсиногена в тонкую кишку из поджелудочной железы секрет энтеропептидазы из слизистой двенадцатиперстной кишки расщепляет пептидную связь лизин 15 - изолейцин 16 зимогена. В результате зимоген трипсиноген распадается на трипсин. Напомним, что трипсин также отвечает за расщепление лизиновых пептидных связей, и, таким образом, как только образуется небольшое количество трипсина, он участвует в расщеплении собственного зимогена, генерируя еще больше трипсина. Таким образом, процесс активации трипсина можно назвать автокаталитическим . |
| Химотрипсиноген | химотрипсин | После того, как связь Arg 15 - Ile 16 в зимогене химотрипсиногена расщепляется трипсином, вновь созданная структура, называемая пи-химотрипсином, подвергается автолизу (самоперевариванию), образуя активный химотрипсин. |
| Проэластаза | эластаза | Он активируется расщеплением трипсином. |
Как видно, активация трипсиногена на трипсин имеет важное значение, поскольку он активирует собственную реакцию, а также реакцию как химотрипсина , так и эластазы . Поэтому очень важно, чтобы эта активация не происходила преждевременно. Организм предпринимает несколько защитных мер для предотвращения самопереваривания:
- Активация трипсиногена трипсином происходит относительно медленно.
- Зимогены хранятся в зимогенных гранулах — капсулах, стенки которых считаются устойчивыми к протеолизу.
Торможение
[ редактировать ]Существуют определенные ингибиторы , которые напоминают тетраэдрический промежуточный продукт и, таким образом, заполняют активный центр, препятствуя правильной работе фермента. Трипсин, мощный пищеварительный фермент, вырабатывается в поджелудочной железе. Ингибиторы предотвращают самопереваривание самой поджелудочной железы.
Сериновые протеазы сочетаются с ингибиторами сериновых протеаз , которые отключают свою активность, когда они больше не нужны. [ 9 ] [ самостоятельно опубликованный источник? ]
Сериновые протеазы ингибируются разнообразной группой ингибиторов , включая синтетические химические ингибиторы для исследовательских или терапевтических целей, а также природные белковые ингибиторы. Одно семейство природных ингибиторов, называемое «серпины» (сокращенно от «ингибиторы сериновой протеазы »), может образовывать ковалентную связь с сериновой протеазой, ингибируя ее функцию. Наиболее изученными серпинами являются антитромбин и альфа-1-антитрипсин , изученные на предмет их роли в процессах коагуляции / тромбоза и эмфиземы / А1АТ соответственно. Искусственные необратимые низкомолекулярные ингибиторы включают AEBSF и PMSF .
Семейство ингибиторов сериновой пептидазы членистоногих , называемое пацифастин , было обнаружено у саранчи и раков и может действовать в иммунной системе членистоногих . [ 10 ]
Роль в болезни
[ редактировать ]Мутации могут приводить к снижению или повышению активности ферментов. Это может иметь разные последствия в зависимости от нормальной функции сериновой протеазы. Например, мутации белка С могут привести к дефициту белка С и предрасполагать к тромбозу . Кроме того, некоторые протеазы играют жизненно важную роль в активации слияния клетки-хозяина и вируса, запуская белок Spike вируса, чтобы показать белок, называемый «слитый белок» ( TMPRSS2 активирует слияние SARS-CoV-2 ). Экзогенные сериновые протеазы змеиного яда вызывают широкий спектр коагулопатий при введении хозяину из-за отсутствия регуляции их активности. [ 7 ]
Диагностическое использование
[ редактировать ]Определение уровней сериновой протеазы может быть полезно в контексте конкретных заболеваний.
- Уровни фактора свертывания крови могут потребоваться при диагностике геморрагических или тромботических состояний.
- Фекальную эластазу используют для определения экзокринной активности поджелудочной железы, например, при муковисцидозе или хроническом панкреатите .
- Сывороточный простатоспецифический антиген используется при скрининге рака простаты , стратификации риска и мониторинге после лечения.
- Сериновая протеаза, выделяемая тучными клетками , является важным диагностическим маркером реакций гиперчувствительности 1 типа , например, анафилаксии . Более полезен, чем гистамин, из-за более длительного периода полувыведения , что означает, что он остается в организме в течение клинически полезного периода времени.
Антимикробный эффект
[ редактировать ]Благодаря своей каталитической активности некоторые сериновые протеазы обладают мощными противомикробными свойствами. Несколько исследований in vitro продемонстрировали эффективность некоторых протеаз в снижении вирулентности путем расщепления поверхностных белков вируса. Проникновение вируса в клетки-хозяева опосредовано взаимодействием этих поверхностных белков с клеткой-хозяином. Когда эти белки фрагментируются или инактивируются на поверхности вируса, проникновение вируса нарушается, что приводит к снижению инфекционности широкого спектра патологически значимых микроорганизмов, таких как грипп , hRSV и другие. [ 11 ] [ 12 ]
См. также
[ редактировать ]- Серингидролаза
- Протеаза
- Клан ПА
- Конвергентная эволюция
- Протеолиз
- Каталитическая триада
- Карта протеолиза
- Протеазы в ангиогенезе
- Внутримембранные протеазы
- Ингибитор протеазы (фармакология)
- Ингибитор протеазы (биология)
- TopFIND - база данных специфичности протеаз, субстратов, продуктов и ингибиторов
- MEROPS - База данных эволюционных групп протеаз
Ссылки
[ редактировать ]- ^ Хедстрем Л. (декабрь 2002 г.). «Механизм и специфичность сериновой протеазы». Химические обзоры . 102 (12): 4501–4524. дои : 10.1021/cr000033x . ПМИД 12475199 .
- ^ Мадала П.К., Тиндалл Дж.Д., Налл Т., Фэрли Д.П. (июнь 2010 г.). «Обновление 1: Протеазы универсально распознают бета-цепи в своих активных сайтах». Химические обзоры . 110 (6): ПР1–П31. дои : 10.1021/cr900368a . ПМИД 20377171 .
- ^ Хаде П.М., Кумар А., Джерниган Р.Л. (январь 2020 г.). «Описание и прогнозирование белковых шарниров для понимания механизмов» . Журнал молекулярной биологии . 432 (2): 508–522. дои : 10.1016/j.jmb.2019.11.018 . ПМК 7029793 . ПМИД 31786268 .
- ^ Хаде П.М., Скарамоццино Д., Кумар А., Ласидогна Г., Карпинтери А., Джерниган Р.Л. (ноябрь 2021 г.). «hdANM: новая комплексная модель динамики белковых шарниров» . Биофизический журнал . 120 (22): 4955–4965. Бибкод : 2021BpJ... 120.4955K дои : 10.1016/j.bpj.2021.10.017 . ПМЦ 8633836 . ПМИД 34687719 .
- ^ Оваре П., Липпенс С., Ванденабиле П., Деклерк В. (сентябрь 2009 г.). «Новая роль каскадов сериновых протеаз в эпидермисе». Тенденции биохимических наук . 34 (9): 453–463. дои : 10.1016/j.tibs.2009.08.001 . ПМИД 19726197 .
- ^ Эвнин Л.Б., Васкес-младший, Крейк К.С. (сентябрь 1990 г.). «Субстратная специфичность трипсина, исследованная с помощью генетической селекции» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (17): 6659–6663. Бибкод : 1990PNAS...87.6659E . дои : 10.1073/pnas.87.17.6659 . JSTOR 2355359 . ПМК 54596 . ПМИД 2204062 .
- ^ Jump up to: а б Оливейра А.Л., Виегас МФ, да Силва С.Л., Соареш А.М., Рамос М.Ю., Фернандес П.А. (10 июня 2022 г.). «Химия змеиного яда и его лечебный потенциал» . Обзоры природы. Химический . 6 (7): 451–469. дои : 10.1038/s41570-022-00393-7 . ПМЦ 9185726 . ПМИД 35702592 .
- ^ Иван Г., Сабадка З., Ордог Р., Гролмуш В., Нарай-Сабо Г. (июнь 2009 г.). «Четыре пространственных точки, определяющие семейства ферментов». Связь с биохимическими и биофизическими исследованиями . 383 (4): 417–420. CiteSeerX 10.1.1.150.1086 . дои : 10.1016/j.bbrc.2009.04.022 . ПМИД 19364497 .
- ^ «Страницы биологии Кимбалла, сериновые протеазы» . Архивировано из оригинала 13 декабря 2005 г. Проверено 2 июня 2008 г.
- ^ Брейгельманс Б., Симонет Г., ван Хоф В., Ван Соест С., Ванден Брук Дж. (март 2009 г.). «Пептиды, родственные пацифастину: структурные и функциональные характеристики семейства ингибиторов сериновой пептидазы». Пептиды . 30 (3): 622–632. doi : 10.1016/j.peptides.2008.07.026 . ПМИД 18775459 . S2CID 8797134 .
- ^ Лопес Б.Р., да Силва Г.С., де Лима Менезес Г., де Оливейра Х., Ватанабэ А.С., Порту Б.Н. и др. (май 2022 г.). «Сериновые протеазы во внеклеточных ловушках нейтрофилов проявляют активность против респираторного синцитиального вируса». Международная иммунофармакология . 106 : 108573. doi : 10.1016/j.intimp.2022.108573 . ПМИД 35183035 .
- ^ Сакаи К., Ами Ю., Тахара М., Кубота Т., Анраку М., Абэ М. и др. (май 2014 г.). Дермоди Т.С. (ред.). «Протеаза хозяина TMPRSS2 играет важную роль в репликации in vivo новых вирусов H7N9 и сезонного гриппа» . Журнал вирусологии . 88 (10): 5608–5616. дои : 10.1128/JVI.03677-13 . ПМК 4019123 . ПМИД 24600012 .
Внешние ссылки
[ редактировать ]- по Онлайн -база данных MEROPS пептидазам и их ингибиторам: Сериновая пептидаза . Архивировано 4 апреля 2017 г. на Wayback Machine.
- сериновых протеаз Сайт в Университете Сент-Луиса (SLU)
- Серин + протеазы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)