Лейцин
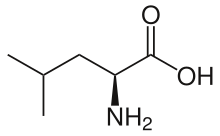 Скелетная формула L - лейцина | |||
| |||
| Имена | |||
|---|---|---|---|
| Название ИЮПАК Лейцин | |||
| Другие имена 2-амино-4-метилпентановая кислота | |||
| Идентификаторы | |||
3D model ( JSmol ) | |||
| ЧЭБИ | |||
| ЧЕМБЛ | |||
| ХимическийПаук | |||
| Лекарственный Банк | |||
| Информационная карта ECHA | 100.000.475 | ||
| КЕГГ | |||
ПабХим CID | |||
| НЕКОТОРЫЙ | |||
Панель управления CompTox ( EPA ) | |||
| Характеристики | |||
| C6H13NOC6H13NO2 | |||
| Молярная масса | 131.175 g·mol −1 | ||
| Кислотность ( pKa ) | 2,36 (карбоксил), 9,60 (амино) [2] | ||
| -84.9·10 −6 см 3 /моль | |||
| Страница дополнительных данных | |||
| Лейцин (страница данных) | |||
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |||

Лейцин (символ Leu или L ) [3] , незаменимая аминокислота используется в биосинтезе белков которая . Лейцин представляет собой α-аминокислоту, то есть содержит α- аминогруппу (которая находится в протонированной группе −NH 3 + образуются в биологических условиях), группа α-карбоновой кислоты (которая находится в депротонированном состоянии −COO − образуются в биологических условиях) и изобутильную группу боковой цепи , что делает ее неполярной алифатической аминокислотой. Он необходим человеку, то есть организм не может его синтезировать: его необходимо получать с пищей. Источниками питания человека являются продукты, содержащие белок, такие как мясо, молочные продукты, соевые продукты, а также фасоль и другие бобовые. Он кодируется кодонами . UUA, UUG, CUU, CUC, CUA и CUG Лейцин назван в честь греческого слова, означающего «белый»: λευκός ( leukós , «белый»), из-за его обычного внешнего вида в виде белого порошка, свойства, которое он разделяет со многими другими аминокислотами . [4]
Как валин и изолейцин , лейцин представляет собой аминокислоту с разветвленной цепью . Первичными конечными продуктами метаболизма лейцина являются ацетил-КоА и ацетоацетат ; следовательно, это одна из двух исключительно кетогенных аминокислот , лизин . второй – [5] Это самая важная кетогенная аминокислота для человека. [6]
Лейцин и β-гидрокси-β-метилмасляная кислота , второстепенный метаболит лейцина , проявляют фармакологическую активность у людей и, как было продемонстрировано, способствуют биосинтезу белка посредством фосфорилирования механистической мишени рапамицина (mTOR). [7] [8]
Диетический лейцин
[ редактировать ]В качестве пищевой добавки L-лейцин имеет номер E E641 и классифицируется как усилитель вкуса . [9]
Требования
[ редактировать ]Совет по продовольствию и питанию (FNB) Института медицины США установил рекомендуемые диетические нормы (RDA) для незаменимых аминокислот в 2002 году. Что касается лейцина, для взрослых 19 лет и старше — 42 мг/кг массы тела в день. [10]
Источники
[ редактировать ]| Еда | г/100г |
|---|---|
| Концентрат сывороточного белка , сухой порошок | 10.0-12.0 |
| Концентрат соевого белка , сухой порошок | 7.5-8.5 |
| Концентрат горохового протеина , сухой порошок | 6.6 |
| Соевые бобы , зрелые семена, жареные, соленые | 2.87 |
| Семя конопли , очищенное | 2.16 |
| Говядина , круглая, верхняя круглая, сырая | 1.76 |
| Арахис | 1.67 |
| Рыба , лосось, розовая, сырая | 1.62 |
| Зародыши пшеницы | 1.57 |
| Миндаль | 1.49 |
| Курица , бройлеры или фритюры, бедра, сырые | 1.48 |
| Яйцо куриное , желток сырой | 1.40 |
| Овес | 1.28 |
| Эдамаме (соевые бобы, зеленые, сырые) | 0.93 |
| Фасоль пинто, приготовленная | 0.78 |
| Чечевица , приготовленная | 0.65 |
| Нут , приготовленный | 0.63 |
| Кукуруза , желтая | 0.35 |
| Молоко коровье цельное, 3,25% молочного жира. | 0.27 |
| Рис коричневый, среднезернистый, приготовленный | 0.19 |
| Молоко , человеческое, зрелое, жидкое | 0.10 |
Влияние на здоровье
[ редактировать ]Было обнаружено, что в качестве пищевой добавки лейцин замедляет деградацию мышечной ткани за счет увеличения синтеза мышечных белков у старых крыс. [12] Однако результаты сравнительных исследований противоречивы. Длительный прием лейцина не увеличивает мышечную массу или силу у здоровых пожилых мужчин. [13] Необходимы дополнительные исследования, желательно основанные на объективной случайной выборке общества. В анализе необходимо учитывать такие факторы, как выбор образа жизни, возраст, пол, диета, физические упражнения и т. д., чтобы выделить эффекты дополнительного приема лейцина как отдельно, так и в сочетании с другими аминокислотами с разветвленной цепью (BCAA). До тех пор пищевая добавка лейцина не может считаться основной причиной мышечного роста или оптимального поддержания здоровья всего населения.
И L-лейцин, и D-лейцин защищают мышей от эпилептических припадков . [14] D-лейцин также прекращает судороги у мышей после начала судорожной активности, по крайней мере, так же эффективно, как диазепам, и без седативного эффекта. [14] Снижение потребления L-лейцина с пищей уменьшает ожирение у мышей. [15] Высокий уровень лейцина в крови связан с резистентностью к инсулину у людей, мышей и грызунов. [16] Это может быть связано с влиянием лейцина на стимуляцию передачи сигналов mTOR . [17] Диетическое ограничение лейцина и других BCAA может обратить вспять вызванное диетой ожирение у мышей дикого типа за счет увеличения затрат энергии и может ограничить набор жировой массы у гиперфагических крыс. [18] [19]
Безопасность
[ редактировать ]Токсичность лейцина, наблюдаемая при декомпенсированной болезни мочи кленового сиропа , вызывает делирий и неврологические расстройства и может быть опасной для жизни. [20]
Высокое потребление лейцина может вызвать или усугубить симптомы пеллагры у людей с низким статусом ниацина , поскольку оно препятствует превращению L-триптофана в ниацин. [21]
Лейцин в дозе, превышающей 500 мг/кг/сутки, наблюдался с гипераммониемией . [22] Таким образом, неофициально допустимый верхний уровень потребления (UL) лейцина для здоровых взрослых мужчин может быть предложен на уровне 500 мг/кг/день или 35 г/день при острых диетических состояниях. [22] [23]
Фармакология
[ редактировать ]Фармакодинамика
[ редактировать ]Лейцин – это пищевая аминокислота , способная напрямую стимулировать миофибриллярных мышц синтез белка . [24] Этот эффект лейцина обусловлен его ролью активатора механистической мишени рапамицина (mTOR). [8] серин -треониновая протеинкиназа , регулирующая биосинтез белка и рост клеток . Активация mTOR лейцином осуществляется через Rag GTPases . [25] [26] [27] лейцина связывание с лейцил-тРНК-синтетазой , [25] [26] связывание лейцина с сестрином 2 , [28] [29] [30] и, возможно, другие механизмы.
Метаболизм у человека
[ редактировать ]Метаболизм лейцина у человека |
Метаболизм лейцина происходит во многих тканях человеческого организма ; однако большая часть пищевого лейцина метаболизируется в печени , жировой и мышечной тканях . [36] Жировая и мышечная ткань используют лейцин для образования стеринов и других соединений. [36] Совместное использование лейцина в этих двух тканях в семь раз больше, чем в печени. [36]
У здоровых людей примерно 60% пищевого L -лейцина метаболизируется через несколько часов, при этом примерно 5% ( диапазон 2–10% ) пищевого L -лейцина превращается в β-гидрокси-β-метилмасляную кислоту (HMB). [37] [38] [35] Около 40% поступающего с пищей L -лейцина превращается в ацетил-КоА , который впоследствии используется в синтезе других соединений. [35]
Подавляющее большинство метаболизма L -лейцина первоначально катализируется ферментом аминотрансферазой аминокислот с разветвленной цепью , продуцируя α-кетоизокапроат (α-KIC). [37] [35] α-KIC в основном метаболизируется митохондриальным ферментом с разветвленной цепью α-кетокислот дегидрогеназой , который превращает его в изовалерил-КоА . [37] [35] Изовалерил-КоА впоследствии метаболизируется изовалерил-КоА-дегидрогеназой и превращается в MC-КоА , который используется в синтезе ацетил-КоА и других соединений. [35] При дефиците биотина HMB может синтезироваться из MC-CoA с помощью еноил-CoA-гидратазы и неизвестного фермента тиоэстеразы . [31] [32] [39] которые преобразуют MC-CoA в HMB-CoA и HMB-CoA в HMB соответственно. [32] количество α-KIC метаболизируется в печени ферментом цитозольным Относительно небольшое 4-гидроксифенилпируватдиоксигеназой (KIC-диоксигеназа), который превращает α-KIC в HMB. [37] [35] [40] У здоровых людей этот второстепенный путь, который включает превращение L -лейцина в α-KIC, а затем в HMB, является преобладающим путем синтеза HMB. [37] [35]
Небольшая часть метаболизма L -лейцина – менее 5% во всех тканях, кроме семенников , где на него приходится около 33% – первоначально катализируется лейцинаминомутазой , производя β-лейцин , который впоследствии метаболизируется в β-кетоизокапроат (β-лейцин). -KIC), β-кетоизокапроил-КоА , а затем ацетил-КоА с помощью ряда нехарактерных ферментов. [35] [41]
Метаболизм HMB катализируется неизвестным ферментом, который превращает его в β-гидрокси β-метилбутирил-КоА ( HMB-CoA ). [31] [35] HMB-CoA метаболизируется либо еноил-CoA-гидратазой , либо другим неизвестным ферментом, образуя β-метилкротонил-CoA ( MC-CoA ) или гидроксиметилглутарил-CoA ( HMG-CoA ) соответственно. [37] [35] MC-CoA затем превращается ферментом метилкротонил-CoA-карбоксилазой в метилглутаконил-CoA ( MG-CoA ), который впоследствии превращается в HMG-CoA с помощью метилглутаконил-CoA гидратазы . [37] [35] [41] Затем HMG-CoA расщепляется на ацетил-CoA и ацетоацетат с помощью HMG-CoA -лиазы или используется в производстве холестерина по мевалонатному пути . [37] [35]
Синтез в нечеловеческих организмах
[ редактировать ]Лейцин является незаменимой аминокислотой в рационе животных, поскольку у них отсутствует полный ферментный путь для синтеза ее de novo из потенциальных соединений-предшественников. Следовательно, они должны потреблять его, обычно как компонент белков. Растения и микроорганизмы синтезируют лейцин из пировиноградной кислоты с помощью ряда ферментов: [42]
- Ацетолактатсинтаза
- ацетогидроксикислоты Изомероредуктаза
- Дегидратаза дигидроксикислот
- α- изопропилмалатсинтаза
- α- изопропилмалатизомераза
- Лейцинаминотрансфераза
Синтез небольшой гидрофобной аминокислоты валина также включает начальную часть этого пути.
Химия
[ редактировать ]
Лейцин — это аминокислота с разветвленной цепью (BCAA), поскольку она имеет алифатическую нелинейную боковую цепь.
Рацемический лейцин был [ когда? ] подвергли с круговой поляризацией синхротронному излучению , чтобы лучше понять природу биомолекулярной асимметрии. биомолекул Было вызвано энантиомерное усиление на 2,6%, что указывает на возможную фотохимическую природу гомохиральности . [43]
См. также
[ редактировать ]- Лейцины , изомеры и производные лейцина.
- Лейциновая молния , распространенный мотив в белках транскрипционных факторов.
Примечания
[ редактировать ]- ^ Эта реакция катализируется неизвестным ферментом тиоэстеразой . [31] [32]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Биннс Дж., Парсонс С., Макинтайр Г.Дж. (декабрь 2016 г.). «Точные водородные параметры аминокислоты L-лейцина» (PDF) . Acta Crystallographica Раздел B. 72 (Часть 6): 885–892. дои : 10.1107/S2052520616015699 . hdl : 20.500.11820/c784fdaf-aa3a-48e4-86a2-d0a0bd7fdb7a . ПМИД 27910839 . S2CID 19288938 .
- ^ Доусон, RMC и др., Данные для биохимических исследований , Оксфорд, Clarendon Press, 1959.
- ^ «Номенклатура и символика аминокислот и пептидов» . Совместная комиссия IUPAC-IUB по биохимической номенклатуре. 1983. Архивировано из оригинала 9 октября 2008 года . Проверено 5 марта 2018 г.
- ^ Флек, Мишель; Петросян, Арам М. (2014). Соли аминокислот: кристаллизация, строение и свойства . Чам: Международное издательство Springer. дои : 10.1007/978-3-319-06299-0 . ISBN 978-3-319-06298-3 .
- ^ Ферье Д.Р. (24 мая 2013 г.). Биохимия . Липпинкотт Уильямс и Уилкинс. ISBN 9781451175622 .
- ^ Кинобер Л.А. (13 ноября 2003 г.). Метаболические и терапевтические аспекты аминокислот в клиническом питании (второе изд.). ЦРК Пресс. п. 101. ИСБН 9780203010266 .
- ^ Сильва В.Р., Белозо Ф.Л., Микелетти Т.О., Конрадо М., Стаут Дж.Р., Пиментел Г.Д., Гонсалес А.М. (сентябрь 2017 г.). «Добавка свободной кислоты β-гидрокси-β-метилбутирата может улучшить восстановление и адаптацию мышц после тренировок с отягощениями: систематический обзор». Исследования питания . 45 : 1–9. дои : 10.1016/j.nutres.2017.07.008 . hdl : 11449/170023 . ПМИД 29037326 .
Обычно считается, что механизмы действия HMB связаны с его влиянием как на синтез мышечного белка, так и на его распад (рис. 1) [2, 3]. HMB, по-видимому, стимулирует синтез мышечного белка посредством усиления регуляции млекопитающих/механической мишени рапамицинового комплекса 1 (mTORC1), сигнального каскада, участвующего в координации инициации трансляции синтеза мышечного белка [2, 4]. Кроме того, HMB может оказывать антагонистическое действие на путь убиквитин-протеасома, систему, которая деградирует внутриклеточные белки [5, 6]. Данные также свидетельствуют о том, что HMB способствует миогенной пролиферации, дифференцировке и слиянию клеток [7]. ... Было показано, что экзогенное введение HMB-FA увеличивает внутримышечную анаболическую передачу сигналов, стимулирует синтез мышечного белка и ослабляет распад мышечного белка у людей [2].
- ^ Перейти обратно: а б Уилкинсон Д.Д., Хоссейн Т., Хилл Д.С., Филлипс Б.Е., Кроссланд Х., Уильямс Дж. и др. (июнь 2013 г.). «Влияние лейцина и его метаболита β-гидрокси-β-метилбутирата на метаболизм белков скелетных мышц человека» . Журнал физиологии . 591 (11): 2911–23. дои : 10.1113/jphysicalol.2013.253203 . ПМК 3690694 . ПМИД 23551944 .
Стимуляция MPS посредством передачи сигналов mTORc1 после воздействия HMB согласуется с доклиническими исследованиями (Eley et al. 2008). ... Кроме того, наблюдалось явное расхождение в амплитуде фосфорилирования 4EBP1 (Thr37/46 и Ser65/Thr70) и p70S6K (Thr389) в ответ как на Leu, так и на HMB, причем последний демонстрировал более выраженное и устойчивое фосфорилирование. ... Тем не менее, поскольку общий ответ MPS был схожим, это различие клеточных сигналов не привело к статистически различимым анаболическим эффектам в нашем первичном показателе исхода MPS. ... Интересно, что, хотя пероральный прием HMB не вызывал увеличения уровня инсулина в плазме, он вызывал депрессию MPB (-57%). Обычно постпрандиальное снижение МПБ (около 50%) объясняется азотсберегающим действием инсулина, поскольку фиксация инсулина в постабсорбционных концентрациях (5 мкЕмл/мл) −1 ) при непрерывном введении АК (18 г в час −1 ) не подавлял МПБ (Greenhaff et al. 2008), поэтому мы решили не измерять МПБ в группе Leu из-за ожидаемой гиперинсулинемии (рис. 3C). Таким образом, HMB снижает MPB аналогично инсулину, но независимо от него. Эти данные согласуются с сообщениями об антикатаболических эффектах HMB, подавляющих MPB в доклинических моделях, посредством ослабления протеасомно-опосредованного протеолиза в ответ на LPS (Eley et al. 2008).
- ^ Зимний Р (2009). Потребительский словарь пищевых добавок (7-е изд.). Нью-Йорк: Три Риверс Пресс. ISBN 978-0307408921 .
- ^ Медицинский институт (2002). «Белки и аминокислоты» . Диетическая норма потребления энергии, углеводов, клетчатки, жиров, жирных кислот, холестерина, белков и аминокислот . Вашингтон, округ Колумбия: Издательство национальных академий. стр. 589–768. дои : 10.17226/10490 . ISBN 978-0-309-08525-0 .
- ^ Национальная база данных питательных веществ для стандартных справок . Министерство сельского хозяйства США. Архивировано из оригинала 3 марта 2015 года . Проверено 16 сентября 2009 г.
- ^ Комбаре Л., Дардеве Д., Рье И., Пуш М.Н., Беше Д., Тайландье Д. и др. (декабрь 2005 г.). «Диета с добавлением лейцина восстанавливает дефектное постпрандиальное ингибирование протеасомозависимого протеолиза в скелетных мышцах старых крыс» . Журнал физиологии . 569 (Часть 2): 489–99. дои : 10.1113/jphysicalol.2005.098004 . ПМЦ 1464228 . ПМИД 16195315 .
- ^ Верховен С., Ваншунбек К., Вердейк Л.Б., Купман Р., Водзиг В.К., Дендейл П., ван Лун Л.Дж. (май 2009 г.). «Длительный прием лейцина не увеличивает мышечную массу или силу у здоровых пожилых мужчин» . Американский журнал клинического питания . 89 (5): 1468–75. дои : 10.3945/ajcn.2008.26668 . ПМИД 19321567 .
- ^ Перейти обратно: а б Хартман А.Л., Сантос П., О'Риордан К.Дж., Стафстром К.Э., Мари Хардвик Дж. (октябрь 2015 г.). «Мощное противосудорожное действие D-лейцина» . Нейробиология болезней . 82 : 46–53. дои : 10.1016/j.nbd.2015.05.013 . ПМЦ 4640989 . ПМИД 26054437 .
- ^ Фонтана Л., Каммингс Н.Э., Арриола Апело С.И., Нойман Дж.К., Каса И., Шмидт Б.А. и др. (июль 2016 г.). «Снижение потребления аминокислот с разветвленной цепью улучшает метаболическое здоровье» . Отчеты по ячейкам . 16 (2): 520–530. дои : 10.1016/j.celrep.2016.05.092 . ПМЦ 4947548 . ПМИД 27346343 .
- ^ Линч CJ, Адамс С.Х. (декабрь 2014 г.). «Аминокислоты с разветвленной цепью в метаболической передаче сигналов и резистентности к инсулину» . Обзоры природы. Эндокринология . 10 (12): 723–36. дои : 10.1038/nrendo.2014.171 . ПМЦ 4424797 . ПМИД 25287287 .
- ^ Кэрон А., Ричард Д., Лапланте М. (2015). «Роль комплексов mTOR в метаболизме липидов». Ежегодный обзор питания . 35 : 321–48. doi : 10.1146/annurev-nutr-071714-034355 . ПМИД 26185979 .
- ^ Каммингс Н.Э., Уильямс Э.М., Каса И., Конон Э.Н., Шайд М.Д., Шмидт Б.А. и др. (февраль 2018 г.). «Восстановление метаболического здоровья за счет снижения потребления аминокислот с разветвленной цепью» . Журнал физиологии . 596 (4): 623–645. дои : 10.1113/JP275075 . ПМК 5813603 . ПМИД 29266268 .
- ^ Уайт П.Дж., Лэпворт А.Л., Ан Дж., Ван Л., МакГарра Р.В., Стивенс Р.Д. и др. (июль 2016 г.). «Ограничение аминокислот с разветвленной цепью у крыс с ожирением Цукера улучшает чувствительность мышц к инсулину за счет повышения эффективности окисления жирных кислот и экспорта ацил-глицина» . Молекулярный метаболизм . 5 (7): 538–551. doi : 10.1016/j.molmet.2016.04.006 . ПМЦ 4921791 . ПМИД 27408778 .
- ^ Юдкофф, Марк; Дайхин Евгений; Ниссим, Илана; Горынь Оксана; Луховой, Богдан; Лазаров, Адам; Ниссим, Ицхак (1 июня 2005 г.). «Потребность мозга в аминокислотах и токсичность: пример лейцина» . Журнал питания . 135 (6): 1531С–1538С. дои : 10.1093/jn/135.6.1531S . ISSN 0022-3166 . ПМИД 15930465 .
- ^ Бадави А.А., Лейк С.Л., Догерти Д.М. (2014). «Механизмы пеллаграгенного эффекта лейцина: стимуляция окисления триптофана в печени путем введения аминокислот с разветвленной цепью здоровым добровольцам и роль свободного триптофана в плазме и общих кинуренинов» . Международный журнал исследований триптофана . 7 : 23–32. дои : 10.4137/IJTR.S18231 . ПМЦ 4259507 . ПМИД 25520560 .
- ^ Перейти обратно: а б Эланго Р., Чепмен К., Рафии М., Болл Р.О., Пенчарз П.Б. (октябрь 2012 г.). «Определение допустимого верхнего уровня потребления лейцина в острых диетических исследованиях у молодых мужчин» . Американский журнал клинического питания . 96 (4): 759–67. дои : 10.3945/ajcn.111.024471 . ПМИД 22952178 .
Значительное увеличение концентрации аммиака в крови выше нормальных значений, концентрации лейцина в плазме и экскреции лейцина с мочой наблюдалось при приеме лейцина >500 мг · кг. −1 · д −1 . Окисление l-[1- 13 C]-лейцин выражается в окислении метки-индикатора при дыхании (F 13 CO 2 ), окисление лейцина и окисление α-кетоизокапроновой кислоты (KIC) привели к разным результатам: плато в F 13 CO 2 наблюдается после 500 мг · кг −1 · д −1 , при окислении лейцина не наблюдается четкого плато, а окисление KIC выходит на плато после 750 мг · кг. −1 · д −1 . На основании показателей плазмы и мочи UL для лейцина у здоровых взрослых мужчин можно предложить на уровне 500 мг · кг. −1 · д −1 или ~35 г/день (по осторожной оценке) при острых диетических состояниях.
- ^ Расмуссен Б., Гилберт Э., Турки А., Мэдден К., Эланго Р. (июль 2016 г.). «Определение безопасности приема лейцина у здоровых пожилых мужчин». Аминокислоты . 48 (7): 1707–16. дои : 10.1007/s00726-016-2241-0 . ПМИД 27138628 . S2CID 3708265 .
верхний предел потребления лейцина здоровыми пожилыми людьми может быть установлен аналогично молодым мужчинам на уровне 500 мг/кг-1 день-1 или ~35 г/день для человека весом 70 кг.
- ^ Эцель М.Р. (апрель 2004 г.). «Производство и применение молочных белковых фракций» . Журнал питания . 134 (4): 996С–1002С. дои : 10.1093/jn/134.4.996S . ПМИД 15051860 .
- ^ Перейти обратно: а б Ким Дж.Х., Ли С., Ли М., Ван Х., Ким К., Пак С.Дж. и др. (сентябрь 2017 г.). «Контроль лейцин-зависимого пути mTORC1 посредством химического вмешательства лейцил-тРНК-синтетазы и взаимодействия RagD» . Природные коммуникации . 8 (1): 732. Бибкод : 2017NatCo...8..732K . дои : 10.1038/s41467-017-00785-0 . ПМЦ 5622079 . ПМИД 28963468 .
- ^ Перейти обратно: а б Джуэлл Дж.Л., Рассел Р.К., Гуань К.Л. (март 2013 г.). «Передача сигналов аминокислот выше mTOR» . Обзоры природы. Молекулярно-клеточная биология . 14 (3): 133–9. дои : 10.1038/nrm3522 . ПМЦ 3988467 . ПМИД 23361334 .
- ^ Санджак Ю., Петерсон Т.Р., Шауль Ю.Д., Линдквист Р.А., Торин К.С., Бар-Пелед Л., Сабатини Д.М. (июнь 2008 г.). «Rag GTPases связывают хищника и опосредуют передачу сигналов аминокислот с mTORC1» . Наука . 320 (5882): 1496–501. Бибкод : 2008Sci...320.1496S . дои : 10.1126/science.1157535 . ПМЦ 2475333 . ПМИД 18497260 .
- ^ Вольфсон Р.Л., Чантранупонг Л., Сакстон Р.А., Шен К., Скариа С.М., Кантор Дж.Р., Сабатини Д.М. (январь 2016 г.). «Сестрин2 является сенсором лейцина для пути mTORC1» . Наука . 351 (6268): 43–8. Бибкод : 2016Sci...351...43W . дои : 10.1126/science.aab2674 . ПМК 4698017 . ПМИД 26449471 .
- ^ Сакстон Р.А., Нокенхауэр К.Е., Вольфсон Р.Л., Чантранупонг Л., Паколд М.Е., Ван Т. и др. (январь 2016 г.). «Структурная основа определения лейцина по пути Sestrin2-mTORC1» . Наука . 351 (6268): 53–8. Бибкод : 2016Sci...351...53S . дои : 10.1126/science.aad2087 . ПМЦ 4698039 . ПМИД 26586190 .
- ^ Чантранупонг Л., Вольфсон Р.Л., Ороско Дж.М., Сакстон Р.А., Скариа С.М., Бар-Пелед Л. и др. (октябрь 2014 г.). «Сестрины взаимодействуют с GATOR2, отрицательно регулируя путь восприятия аминокислот выше mTORC1» . Отчеты по ячейкам . 9 (1): 1–8. дои : 10.1016/j.celrep.2014.09.014 . ПМЦ 4223866 . ПМИД 25263562 .
- ^ Перейти обратно: а б с «Реакция KEGG: R10759» . Киотская энциклопедия генов и геномов . Лаборатории Канеиса. Архивировано из оригинала 1 июля 2016 года . Проверено 24 июня 2016 г.
- ^ Перейти обратно: а б с Мок Д.М., Страттон С.Л., Хорват Т.Д., Богусевич А., Мэтьюз Н.И., Хенрих К.Л., Доусон А.М., Спенсер Х.Дж., Оуэн С.Н., Бойсен Г., Моран Дж.Х. (ноябрь 2011 г.). «Выведение 3-гидроксиизовалериановой кислоты и 3-гидроксиизовалерилкарнитина с мочой увеличивается в ответ на введение лейцина у людей с незначительным дефицитом биотина» . первоисточник. Журнал питания . 141 (11): 1925–1930. дои : 10.3945/jn.111.146126 . ПМК 3192457 . ПМИД 21918059 .
Снижение активности MCC ухудшает катализ важнейшего этапа митохондриального катаболизма лейцина с разветвленной цепью (BCAA). Метаболические нарушения превращают метилкротонил-КоА в 3-гидроксиизовалерил-КоА в реакции, катализируемой еноил-КоА-гидратазой (22, 23). Накопление 3-гидроксиизовалерил-КоА может ингибировать клеточное дыхание либо напрямую, либо путем воздействия на соотношение ацил-КоА:свободный КоА, если не происходит дальнейшего метаболизма и детоксикации 3-гидроксиизовалерил-КоА (22). Перенос карнитина с помощью 4 карнитин-ацил-КоА-трансфераз, распределенных в субклеточных компартментах, вероятно, служит важным резервуаром для ацильных фрагментов (39–41). 3-Гидроксиизовалерил-КоА, вероятно, детоксицируется карнитин-ацетилтрансферазой с образованием 3HIA-карнитина, который транспортируется через внутреннюю митохондриальную мембрану (и, следовательно, эффективно выходит из митохондрий) посредством карнитин-ацилкарнитинтранслоказы (39). Считается, что 3HIA-карнитин либо напрямую деацилируется гидролазой до 3HIA, либо подвергается второму обмену КоА с образованием 3-гидроксиизовалерил-КоА с последующим высвобождением 3HIA и свободного КоА тиоэстеразой.
- ^ Перейти обратно: а б Уилсон Дж.М., Фитшен П.Дж., Кэмпбелл Б., Уилсон Дж.Дж., Занчи Н., Тейлор Л., Уилборн С., Калман Д.С., Стаут Дж.Р., Хоффман Дж.Р., Зигенфусс Т.Н., Лопес Х.Л., Крайдер Р.Б., Смит-Райан А.Е., Антонио Дж. (февраль 2013 г.) . «Позиция Международного общества спортивного питания: бета-гидрокси-бета-метилбутират (HMB)» . Журнал Международного общества спортивного питания . 10 (1): 6. дои : 10.1186/1550-2783-10-6 . ПМК 3568064 . ПМИД 23374455 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Кольмайер М. (май 2015 г.). «Лейцин» . Метаболизм питательных веществ: структуры, функции и гены (2-е изд.). Академическая пресса. стр. 385–388. ISBN 978-0-12-387784-0 . Архивировано из оригинала 22 марта 2018 года . Проверено 6 июня 2016 г.
Энергетическое топливо: со временем большая часть лея расщепляется, образуя около 6,0 ккал/г. Около 60% поступившего в организм Leu окисляется в течение нескольких часов... Кетогенез: значительная часть (40% поступившей в организм дозы) превращается в ацетил-КоА и тем самым способствует синтезу кетонов, стероидов, жирных кислот и других соединения
Рисунок 8.57: Метаболизм L -лейцина. Архивировано 22 марта 2018 г. на Wayback Machine. - ^ Перейти обратно: а б с Розенталь Дж., Анхель А., Фаркас Дж. (февраль 1974 г.). «Метаболическая судьба лейцина: важный предшественник стерола в жировой ткани и мышцах». Американский журнал физиологии . 226 (2): 411–8. дои : 10.1152/ajplegacy.1974.226.2.411 . ПМИД 4855772 .
- ^ Brioche T, Pagano AF, Py G, Chopard A (август 2016 г.). «Мышечное истощение и старение: экспериментальные модели, жировые инфильтраты и профилактика» (PDF) . Молекулярные аспекты медицины . 50 : 56–87. дои : 10.1016/j.mam.2016.04.006 . ПМИД 27106402 . S2CID 29717535 .
В заключение, лечение HMB, очевидно, является безопасной эффективной стратегией против саркопении и, в более общем плане, против атрофии мышц, поскольку HMB улучшает мышечную массу, мышечную силу и физическую работоспособность. Похоже, что HMB способен воздействовать на три из четырех основных механизмов, участвующих в деформировании мышц (обмен белка, апоптоз и регенеративный процесс), тогда как предположительно он сильно влияет на четвертый (динамика и функции митохондрий). Более того, HMB стоит недорого (~30–50 долларов США в месяц при дозе 3 г в день) и может предотвратить остеопению (Bruckbauer and Zemel, 2013; Татара, 2009; Татара и др., 2007, 2008, 2012) и снизить сердечно-сосудистые риски. (Ниссен и др., 2000). По всем этим причинам HMB следует регулярно использовать при атрофии мышц, особенно у пожилых людей. ... 3 г CaHMB, принимаемые три раза в день (по 1 г каждый раз), являются оптимальной дозировкой, обеспечивающей постоянную биодоступность HMB в организме (Wilson et al., 2013).
- ^ «Реакция KEGG: R04137» . Киотская энциклопедия генов и геномов . Лаборатории Канеиса. Архивировано из оригинала 1 июля 2016 года . Проверено 24 июня 2016 г.
- ^ «Homo sapiens: реакция 4-гидроксифенилпируватдиоксигеназы» . МетаЦик . НИИ Интернешнл. 20 августа 2012 года . Проверено 6 июня 2016 г.
- ^ Перейти обратно: а б «Метаболизм лейцина» . БРЕНДА . Технический университет Брауншвейга. Архивировано из оригинала 17 августа 2016 года . Проверено 12 августа 2016 г.
- ^ Ленинджер А.Л., Нельсон Д.Л., Кокс М.М. (2000). Ленингерские принципы биохимии (3-е изд.). Нью-Йорк: Издательство Worth. ISBN 978-1-57259-153-0 .
- ^ Мейеренрих : Аминокислоты и асимметрия жизни , Springer-Verlag, 2008, ISBN 978-3-540-76885-2 .
Внешние ссылки
[ редактировать ]- Биосинтез лейцина на Wayback Machine (архивировано 6 мая 2017 г.)
