Эволюция китообразных

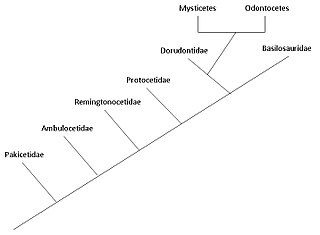
Считается, что эволюция китообразных началась на Индийском субконтиненте от парнокопытных (парнокопытных) 50 миллионов лет назад (млн лет назад) и продолжалась в течение как минимум 15 миллионов лет. [ 2 ] Китообразные — полностью водные морские млекопитающие , принадлежащие к отряду парнокопытных и ответвившиеся от других парнокопытных около 50 млн лет назад . Считается, что китообразные эволюционировали во время эоцена (56–34 млн лет назад), второй эпохи продолжающейся в настоящее время кайнозойской эры. Молекулярный и морфологический анализ позволяет предположить, что китообразные имеют относительно недавнего ближайшего общего предка с гиппопотамами и что они являются сестринскими группами. [ 3 ] Будучи млекопитающими, они выходят на поверхность, чтобы подышать воздухом; в плавниках у них пять пальцевых костей (парнокопытных); они выкармливают своих детенышей; и, несмотря на полностью водный образ жизни, они сохранили многие черты скелета своих наземных предков. [ 4 ] [ 5 ] Исследования, проведенные в конце 1970-х годов в Пакистане, выявили несколько этапов перехода китообразных с суши в море.
Считается, что два современных вида китообразных — Mysticeti (усатые киты) и Odontoceti (зубатые киты) — отделились друг от друга примерно 28–33 млн лет назад во время второй радиации китообразных , первая из которых произошла с археоцетами . [ 6 ] Адаптация животной эхолокации у зубатых китов отличает их от полностью водных археоцетов и ранних усатых китов . Присутствие усатых китов у усатых китов происходило постепенно, при этом более ранние разновидности имели очень мало усатых китов, а их размер связан с зависимостью от усатых китов (и последующим увеличением фильтрационного питания).
Ранняя эволюция
[ редактировать ]














Водный образ жизни китообразных впервые начался на Индийском субконтиненте от парнокопытных 50 миллионов лет назад, причем этот начальный этап длился примерно 4-15 миллионов лет. [ 8 ] Archaeoceti — вымерший членистоногий китообразных, содержащий древних китов. Традиционная гипотеза эволюции китообразных, впервые предложенная Ван Валеном в 1966 году, [ 9 ] Было то, что киты были связаны с мезонихиями , вымершим отрядом хищных копытных (копытных животных), которые напоминали волков с копытами и были сестринской группой парнокопытных ( парнокопытных ). Эта гипотеза была выдвинута из-за сходства необычных треугольных зубов мезонихий и ранних китов. Однако данные молекулярной филогении показывают, что киты очень тесно связаны с парнокопытными, а бегемоты являются их ближайшими живыми родственниками. Из-за этого наблюдения китообразные и гиппопотамы отнесены к подотряду одному Whippomorpha . [ 10 ] Cetartiodactyla (образовано от слов Cetacea и Artiodactyla) — предложенное название отряда , включающего как китообразных, так и парнокопытных. Однако самые ранние антракотеры , предки гиппопотамов, не появляются в летописи окаменелостей до среднего эоцена, через миллионы лет после Pakicetus , тогда как первый известный предок кита появился в раннем эоцене; эта разница во времени означает, что эти две группы разошлись задолго до эоцена. Молекулярный анализ идентифицирует парнокопытных как очень близких родственников китообразных, поэтому мезонихи, вероятно, являются ответвлением парнокопытных, а китообразные не произошли напрямую от мезонихий, но эти две группы могут иметь общего предка. [ 11 ]
Молекулярные данные подтверждаются открытием Pakicetus , самого раннего археоцита . Скелеты Pakicetus показывают, что киты не произошли напрямую от мезонихий. Вместо этого это парнокопытные, которые начали плавать в воде вскоре после того, как парнокопытные отделились от мезонихий. Археоцеты сохранили аспекты своего мезонихического происхождения (например, треугольные зубы), которые утратили современные парнокопытные и современные киты. Самые ранние предки всех копытных млекопитающих, вероятно, были, по крайней мере частично, плотоядными или падальщиками, а современные парнокопытные и непарнокопытные стали травоядными на более позднем этапе своей эволюции. Киты, однако, сохранили свою плотоядную диету, потому что добыча была более доступной, и им требовалась более высокая калорийность , чтобы жить как морские эндотермы (теплокровные). Мезонихи также стали специализированными хищниками, но это, вероятно, было недостатком, поскольку крупная добыча была редкостью. Возможно, именно поэтому их вытеснили более приспособленные животные, такие как гиенодонтиды. и позже Карнивора . [ 11 ]
Индохьюс
[ редактировать ]Индохиус был небольшим животным, похожим на шевротана , которое жило около 48 миллионов лет назад на территории современного Кашмира . [ 12 ] Он принадлежит к семейству парнокопытных Raoellidae , которое считается ближайшей сестринской группой китообразных . Indohyus идентифицируется как парнокопытное, потому что у него есть две шарнирные петли — особенность, уникальная для парнокопытных. [ 7 ] Это всеядное существо размером примерно с енота или домашнюю кошку имело некоторые черты современных китов, в первую очередь обертку , характер роста костей, который является диагностической характеристикой любого китообразного; этого нет ни у одного другого вида. [ 1 ] У него также были обнаружены признаки адаптации к водной жизни, в том числе плотные кости конечностей, которые уменьшают плавучесть, чтобы они могли оставаться под водой, что аналогично адаптации, обнаруженной у современных водных млекопитающих, таких как бегемот. [ 10 ] [ 13 ] Это предполагает схожую стратегию выживания с африканским шевротином или водным шевротином , который при угрозе со стороны хищной птицы ныряет в воду и прячется под поверхностью на срок до четырех минут. [ 14 ]
Первые окаменелости индохиусов обнаружил индийский геолог А. Ранга Рао. [ 15 ] Среди собранных им камней он обнаружил несколько зубов и челюстную кость. После его смерти его вдова Лилавати Рао подарила камни профессору Хансу Тьювиссену . Его техник случайно взломал пару подаренных камней и обнаружил дополнительные окаменелости Индохиуса.
Пакицетиды
[ редактировать ]Пакицетиды были пальцевидными копытными млекопитающими, которые считаются самыми ранними известными китообразными, а Indohyus - ближайшей сестринской группой. [ 12 ] [ 16 ] Они жили в раннем эоцене , около 50 миллионов лет назад. Их окаменелости были впервые обнаружены в Северном Пакистане в 1979 году, на реке недалеко от берегов бывшего моря Тетис . [ 17 ] После первоначального открытия было обнаружено больше окаменелостей, в основном в речных отложениях раннего эоцена на севере Пакистана и северо-западе Индии. жили в засушливой среде с эфемерными ручьями и умеренно развитыми поймами. Судя по этому открытию, пакицетиды, скорее всего , миллионы лет назад [ 1 ] С помощью анализа стабильных изотопов кислорода было показано, что они пьют пресную воду, а это означает, что они жили вблизи пресноводных водоемов. [ 18 ] В их рацион, вероятно, входили наземные животные, приближавшиеся к воде для питья, или некоторые пресноводные водные организмы, обитавшие в реке. [ 1 ] Удлиненные шейные позвонки и четыре сросшихся крестцовых позвонка соответствуют парнокопытным, что делает Pakicetus одним из самых ранних окаменелостей, обнаруженных в период после события расхождения китообразных и парнокопытных . [ 19 ]
Пакицетиды относят к китообразным главным образом из-за строения слуховой буллы (ушной кости), которая формируется только из эктотимпанальной кости . Форма ушной области у пакицетид весьма необычна, череп напоминает китообразное, хотя дыхало на этой стадии еще отсутствует. В челюстной кости пакицетид также отсутствует расширенное пространство ( нижнечелюстное отверстие ), заполненное жиром или маслом, которое используется для восприятия подводного звука у современных китообразных. [ 20 ] У них есть спинные орбиты (глазницы обращены вверх), похожие на крокодиловые. Такое расположение глаз помогает подводным хищникам наблюдать за потенциальной добычей над водой. [ 18 ] Согласно исследованию 2009 года, зубы пакицетид также напоминают зубы ископаемых китов, менее похожи на резцы собаки и имеют зазубренные треугольные зубы, что является еще одной связью с более современными китообразными. [ 21 ] Первоначально считалось, что уши пакицетид приспособлены для слуха под водой, но, как и следовало ожидать из анатомии остальных животных, уши пакицетид специализированы для слуха на суше. [ 22 ] Однако пакицетиды могли слушать под водой за счет усиленной костной проводимости, а не за счет барабанной перепонки, как другие наземные млекопитающие. Этот метод слуха не давал направленного слуха под водой. [ 20 ]
У пакицетид длинные тонкие ноги и относительно короткие руки и ноги, что позволяет предположить, что они были плохими пловцами. [ 1 ] Чтобы компенсировать это, их кости необычайно толстые ( остеосклеротические ), что, вероятно, является адаптацией, призванной утяжелить животное, чтобы противодействовать плавучести воды. [ 7 ] Согласно морфологическому анализу, проведенному Тевиссеном и др. в 2001 году, пакицетиды не демонстрируют адаптации водного скелета; вместо этого они демонстрируют приспособления для бега и прыжков. Следовательно, пакицетиды, скорее всего, были водными куликами. [ 23 ]
Амбулоцетиды
[ редактировать ]Ambulocetus , живший около 49 миллионов лет назад, был обнаружен в Пакистане в 1994 году. Они напоминали млекопитающих, отдаленно напоминающих крокодилов , с большими бревиростриновыми челюстями. В эоцене амбулоцетиды населяли заливы и устья моря Тетис на севере Пакистана. Окаменелости амбулоцетид всегда находят в прибрежных мелководных морских отложениях, связанных с обильными окаменелостями морских растений и прибрежных моллюсков . [ 1 ] Хотя они встречаются только в морских отложениях, значения их изотопов кислорода указывают на то, что они потребляли воду с разной степенью солености, при этом некоторые экземпляры не имели никаких признаков потребления морской воды, а другие не потребляли пресную воду в то время, когда их зубы окаменели. Очевидно, что амбулоцетиды переносили широкий диапазон концентраций соли. Следовательно, амбулоцетиды представляют собой переходную фазу предков китообразных между пресной водой и морской средой обитания. [ 18 ]
Нижнечелюстное отверстие у амбулоцетид увеличилось в размерах, что указывает на то, что в нижней челюсти, вероятно, располагался жировой слой. У современных зубатых китов этот жировой слой в отверстии нижней челюсти простирается назад до среднего уха. Это позволяет звукам восприниматься нижней челюстью, а затем передаваться через жировую подушечку в среднее ухо. Как и у пакицетид, орбиты амбулоцетид находятся на верхней части черепа, но обращены более латерально, чем у пакицетид. [ 18 ]
У амбулоцетид были относительно длинные конечности с особенно сильными задними ногами, а также у них сохранился хвост без признаков двуустки . [ 17 ] Строение задних конечностей амбулоцетид показывает, что их способность к наземному передвижению была значительно ограничена по сравнению с таковой у современных наземных млекопитающих и, вероятно, вообще не выходила на сушу. Скелетные структуры колена и лодыжки указывают на то, что движение задних конечностей было ограничено в одной плоскости. Это говорит о том, что на суше движение задних конечностей осуществлялось за счет растяжения спинных мышц. [ 24 ] Они, вероятно, плавали , используя тазовые движения (способ плавания, при котором в основном используются задние конечности для создания движения в воде) и хвостовые волны (способ плавания, при котором колебания позвоночного столба используются для создания силы для движений), как выдры и тюлени. и современные китообразные делают. [ 25 ] Это промежуточный этап в эволюции передвижения китообразных, поскольку современные китообразные плавают за счет каудальных колебаний (способ плавания, аналогичный каудальному волнообразному плаванию, но более энергоэффективный). [ 18 ]
Недавнее исследование показывает, что амбулоцетиды, как и современные китообразные, вели полностью водный образ жизни, обладали сходной морфологией грудной клетки и были неспособны выдерживать свой вес на суше. Это говорит о том, что полное оставление суши у китообразных произошло гораздо раньше, чем считалось ранее. Однако ученые, участвовавшие в исследовании, предупредили, что исследование было ограничено отсутствием информации о точной плотности кости, расположении центра массы и использовании ложных ребер для поддержки грудной клетки. [ 26 ]
Ремингтоноцетиды
[ редактировать ]Ремингтоноцетиды жили в среднем эоцене в Южной Азии , примерно 49–43 миллиона лет назад. [ 27 ] По сравнению с семействами Pakicetidae и Ambulocetidae, Remingtonocetidae представляли собой разнообразное семейство, встречающееся в северном и центральном Пакистане и западной Индии. Ремингтоноцетиды также были обнаружены в мелководных морских отложениях, но они были явно более водными, чем амбулоцетиды. Об этом свидетельствует обнаружение их окаменелостей в различных прибрежных морских средах, включая прибрежные и лагунные отложения. [ 1 ] Согласно анализу стабильных изотопов кислорода, большинство ремингтоноцитов не потребляли пресную воду и, следовательно, утратили зависимость от пресной воды относительно вскоре после своего происхождения. [ 18 ]
Орбиты ремингтоноцетид обращены латерально и имеют небольшие размеры. Это говорит о том, что зрение не было для них важным чувством. Носовое отверстие, которое у современных китообразных со временем становится дыхалом, располагалось недалеко от кончика морды . Положение носового отверстия осталось неизменным со времен пакицетид. [ 18 ] Одной из примечательных особенностей ремингтоноцетид является то, что полукружные каналы , которые важны для равновесия у наземных млекопитающих, уменьшились в размерах. [ 28 ] Это уменьшение размера тесно сопровождало проникновение китообразных в морскую среду. Согласно исследованию 2002 года, проведенному Спуром и др., эта модификация системы полукружных каналов может представлять собой решающее событие «точки невозврата» в ранней эволюции китообразных, которая исключала длительную полуводную фазу. [ 28 ]
По сравнению с амбулоцетидами ремингтоноцетиды имели относительно короткие конечности. [ 18 ] Судя по остаткам скелета, ремингтоноцетиды, вероятно, были китообразными-земноводными, которые были хорошо приспособлены к плаванию и, вероятно, плавали только за счет хвостовых движений. [ 1 ]
Протоцетиды
[ редактировать ]Протоцетиды образуют разнообразную и гетерогенную группу, известную из Азии, Европы, Африки и Северной Америки. Они жили в эоцене, примерно 48–35 миллионов лет назад. Ископаемые останки протоцетид были обнаружены в прибрежных и лагунных фациях Южной Азии; в отличие от предыдущих семейств китообразных, их окаменелости обнаружены в Африке (например, Phiomicetus , Protocetus , Aegyptocetus , Togocetus ) [ 29 ] и Северная Америка (например, Georgiacetus ) также включает открытые морские формы. [ 1 ] Вероятно, они были амфибиями, но более водными по сравнению с ремингтоноцетидами. [ 27 ] Протоцетиды были первыми китообразными, покинувшими Индийский субконтинент и расселившимися по всем мелководным субтропическим океанам мира. [ 18 ] В семействе Protocetidae было много родов. В этой группе существовала разная степень водной адаптации: некоторые могли выдерживать свой вес на суше, а другие — нет. [ 1 ] Открытие юго-восточной части Тихого океана Peregocetus указывает на то, что они пересекли Атлантику и достигли экваториального распространения к 40 млн лет назад. [ 30 ] Их амфибийная природа подтверждается открытием беременной Maiacetus , у которой окаменелый плод был помещен для родов головой вперед, что позволяет предположить, что Maiacetus родила на суше. Если бы они рожали в воде, плод располагали бы для родов хвостом вперед, чтобы не утонуть во время родов. [ 31 ]
В отличие от ремингтоноцетид и амбулоцетид, протоцетиды имеют большие орбиты, ориентированные латерально. Глаза, обращенные вбок, все чаще могут использоваться для наблюдения за подводной добычей, и они похожи на глаза современных китообразных. Кроме того, носовые отверстия были большими и находились на середине морды. Большое разнообразие зубов предполагает разнообразные способы питания протоцетид. [ 27 ] Как у ремингтоноцетид, так и у протоцетид размер нижнечелюстного отверстия увеличился. [ 18 ] Большое нижнечелюстное отверстие указывает на наличие нижнечелюстного жирового слоя. Однако заполненные воздухом пазухи, присутствующие у современных китообразных и выполняющие функцию акустической изоляции уха, позволяющей лучше слышать под водой, все еще отсутствовали. [ 20 ] Присутствовал также наружный слуховой проход ( ушной канал ), отсутствующий у современных китообразных. Следовательно, имевшийся у них способ передачи звука сочетает в себе черты пакицетид и современных зубатых китов (зубатых китов). [ 20 ] На этой промежуточной стадии развития слуха передача воздушного звука была плохой из-за модификации уха, приспособленной к подводному слуху, в то время как направленный подводный слух также был плохим по сравнению с современными китообразными. [ 20 ]
У некоторых протоцетид были короткие и широкие передние и задние конечности, которые, вероятно, использовались при плавании, но конечности обеспечивали медленное и тяжелое передвижение по суше. [ 18 ] Возможно, у некоторых протоцетид были сосальщики. Однако очевидно, что они были еще более приспособлены к водному образу жизни. У Rodhocetus , например, крестец (кость, которая у наземных млекопитающих представляет собой сращение пяти позвонков, соединяющее таз с остальной частью позвоночного столба ) был разделен на свободные позвонки. Однако таз все еще был соединен с одним из крестцовых позвонков. Копытное происхождение этих археоцетов до сих пор подчеркивается такими характеристиками, как наличие копыт на концах пальцев у Rodhocetus . [ 32 ]
Строение стопы Rodhocetus показывает, что протоцетиды вели преимущественно водный образ жизни. Исследование 2001 года, проведенное Gingerich et al. выдвинули гипотезу, что Rodhocetus передвигался в океанической среде аналогично тому, как гребли тазом амбулоцетид, что дополнялось волнистостью хвоста. Наземное передвижение Rodhocetus было очень ограничено из-за строения задних конечностей. Считается, что они двигались аналогично тому, как ушастые тюлени передвигаются по суше, вращая задними ластами вперед и под телом. [ 33 ]
Базилозавриды
[ редактировать ]
Базилозавриды и дорудонтины жили вместе в позднем эоцене примерно от 41 до 33,9 миллионов лет назад и являются старейшими известными облигатными водными китообразными. [ 22 ] Это были вполне узнаваемые киты, которые жили полностью в океане. Это подтверждается их окаменелостями, которые обычно обнаруживаются в отложениях, характерных для полностью морской среды, без притока пресной воды. [ 1 ] Вероятно, они были распространены во всех тропических и субтропических морях мира. Базилозавриды обычно встречаются вместе с дорудонтинами и были тесно связаны друг с другом. [ 18 ] Окаменевшее содержимое желудка одного базилозавра указывает на то, что он ел рыбу. [ 1 ]
Хотя они очень похожи на современных китообразных, у базилозавридов отсутствовал « дынный орган », который позволяет зубатым китам использовать эхолокацию . У них был маленький мозг; это говорит о том, что они были одинокими и не имели сложной социальной структуры некоторых современных китообразных. Нижнечелюстное отверстие у базилозавридов покрывало всю глубину нижней челюсти, как и у современных китообразных. Их орбиты были обращены вбок, а носовое отверстие переместилось еще выше по морде, ближе к положению дыхала у современных китообразных. [ 18 ] Более того, их ушные структуры были функционально современными, с наличием заполненных воздухом пазух между ухом и черепом. [ 20 ] В отличие от современных китообразных, базилозавриды сохранили большой наружный слуховой проход. [ 20 ]
У обоих базилозавридов есть скелеты, по которым сразу можно узнать китообразных. Базилозавриды были такими же большими, как и более крупные современные киты, а представители таких родов, как Basilosaurus, достигали длины до 60 футов (18 м); dorudontines были меньше, а такие виды, как Dorudon, достигали около 15 футов (4,6 м) в длину. Самый маленький кит-базилозаврид — Tutcetus , его длина составляет 8,2 фута (2,5 м). [ 34 ] Большой размер базилозавридов обусловлен чрезмерным удлинением их поясничных позвонков. У них была хвостовая двуустка, но пропорции их тела позволяют предположить, что они плавали за счет хвостовых волн и что двуустка не использовалась для движения. [ 1 ] [ 35 ] Напротив, у дорудонтинов был более короткий, но мощный позвоночник. У них тоже была двуустка, и, в отличие от базилозавридов, они, вероятно, плавали так же, как современные китообразные, используя хвостовые колебания. [ 18 ] Передние конечности базилозавридов, вероятно, имели форму ласт, а внешние задние конечности были крошечными и уж точно не участвовали в передвижении. [ 1 ] Однако их пальцы сохранили подвижные суставы своих родственников-амбулоцетид. Две крошечные, но хорошо сформированные задние ноги базилозавридов, вероятно, использовались в качестве застежек при спаривании. Кости таза, связанные с этими задними конечностями, не были соединены с позвоночником, как у протоцетид. По сути, крестцовые позвонки уже невозможно четко отличить от других позвонков. [ 1 ] [ 36 ]
И базилозавриды, и дорудонтины относительно тесно связаны с современными китообразными, принадлежащими к parvorders Odontoceti и Mysticeti . Однако, согласно исследованию 1994 года, проведенному Фордайсом и Барнсом, большой размер и удлиненное тело позвонка базилозавридов не позволяют им быть предками современных форм. Что касается дорудонтинов, то в этом семействе есть виды, не имеющие удлиненных тел позвонков, которые могут быть непосредственными предками Odontoceti и Mysticeti. Остальные базилозавриды вымерли. [ 27 ]
Эволюция современных китообразных
[ редактировать ]Усатые киты
[ редактировать ]
Все современные усатые киты или усатые киты являются фильтраторами, у которых вместо зубов есть ус , хотя точные способы использования уса различаются у разных видов (кормление залпом у балаеноптерид, обезжиренное питание у баленид и вспашка дна у эсхрихтиид). Первые представители обеих групп появились в среднем миоцене . Фильтрационное кормление очень полезно, поскольку позволяет усатым китам эффективно получать огромные энергетические ресурсы, что делает возможным большие размеры тела у современных разновидностей. [ 37 ] Развитие фильтрационного питания могло быть результатом глобальных изменений окружающей среды и физических изменений в океанах. Крупномасштабное изменение океанского течения и температуры могло способствовать излучению современных усатых китов. [ 38 ] Более ранние разновидности усатых китов, или «археомистицеты», такие как Janjucetus и Mammalodon, имели очень мало усатых китов и полагались в основном на свои зубы. [ 39 ]
Есть также свидетельства генетической составляющей эволюции беззубых китов. Множественные мутации были выявлены в генах, связанных с производством эмали у современных усатых китов. [ 40 ] В первую очередь это инсерционные/делеционные мутации, которые приводят к преждевременному образованию стоп-кодонов. [ 40 ] Предполагается, что эти мутации произошли у китообразных, уже обладающих предварительными структурами уса, что привело к псевдогенизации «генетического набора инструментов» для производства эмали. [ 41 ] Недавние исследования также показали, что развитие уса и потеря зубов с эмалевыми крышками произошли один раз, и оба произошли на стволовой ветви усатых китов.
Обычно предполагается, что четыре современных семейства усатых китов имеют отдельное происхождение среди цетотерий . Современные усатые киты, Balaenopteridae (роквалы и горбатые кит, Megaptera novaengliae ), Balaenidae (гладкие киты), Eschrichtiidae (серый кит, Eschrictiusrobustus ) и Neobalaenidae (карликовый кит, Caperea Marginata ) - все они имеют характеристики, неизвестные в настоящее время ни в одной цетотерие и наоборот (например, сагиттальный гребень [ 42 ] ). [ 43 ] Mysticetes также известны своим гигантизмом, поскольку усатые киты являются одними из крупнейших когда-либо живших организмов; они достигают длины более 20 м и весят более 100 000 кг. [ 44 ] Этот гигантизм напрямую связан с механизмом их питания: было обнаружено, что размер усатых кистей зависит от количества уса, который усатые киски могут использовать для фильтрации своей добычи. [ 45 ] Кроме того, размер — это позитивно выбранный признак, который повышает физическую форму усатых китов. [ 46 ] Таким образом, популяции Mysticete постепенно станут еще более гигантскими по мере того, как будут отбираться киты с большим количеством усатых китов.
Зубатые киты
[ редактировать ]Адаптация эхолокации произошла, когда зубатые киты (Odontoceti) отделились от усатых китов, и отличает современных зубатых китов от полностью водных археоцетов. Это произошло около 34 миллионов лет назад во время второй радиации китообразных . [ 47 ] [ 48 ] Современные зубатые киты при охоте на добычу полагаются не на зрение, а на гидролокатор . Эхолокация также позволила зубатым китам глубже нырять в поисках пищи, при этом свет больше не был необходим для навигации, что открыло новые источники пищи. [ 27 ] [ 49 ] Зубатые киты используют эхолокацию, создавая серию щелчков, издаваемых на разных частотах. Звуковые импульсы излучаются, отражаются от предметов и проходят через нижнюю челюсть. Черепа сквалодона свидетельствуют о первом гипотетическом появлении эхолокации. [ 50 ] Сквалодон жил с раннего и среднего олигоцена до среднего миоцена , около 33–14 миллионов лет назад. Сквалодон имел несколько общих черт с современными зубатыми китами: череп был хорошо сжат (чтобы освободить место для дыни, части носа ), рострум выдвигался наружу в клюв, что характерно для современных зубатых китов, что придавало Сквалодону сходный внешний вид. им. Однако маловероятно, что сквалодонтиды являются прямыми предками современных зубатых китов. [ 51 ]
Первые океанические дельфины, такие как кентриодонты , появились в позднем олигоцене и значительно разнообразились в середине миоцена. [ 52 ] Первые ископаемые китообразные вблизи мелководных морей (где обитают морские свиньи) были обнаружены в северной части Тихого океана; такие виды, как Semirostrum, были обнаружены вдоль Калифорнии (в тогдашних эстуариях). [ 53 ] К европейским побережьям и Южному полушарию эти животные распространились лишь значительно позже, в плиоцене . [ 54 ] Самый ранний известный предок арктических китов — Denebola brachycephala из позднего миоцена, около 9–10 миллионов лет назад. [ 55 ] Единственная окаменелость из Нижней Калифорнии указывает на то, что эта семья когда-то обитала в более теплых водах. [ 27 ] [ 56 ] [ 57 ]

Древние кашалоты отличаются от современных кашалотов количеством зубов, формой лица и челюстей. Например, у Scaldicetus был конический рострум . Роды олигоцена и миоцена имели зубы на верхних челюстях. Эти анатомические различия позволяют предположить, что эти древние виды, возможно, не обязательно были охотниками на глубоководных кальмаров, как современные кашалоты, но некоторые виды в основном питались рыбой. [ 27 ] [ 58 ] В отличие от современных кашалотов, самые древние кашалоты были созданы для охоты на китов. У Ливятана был короткий и широкий рострум размером 10 футов (3,0 м) в поперечнике, что давало киту возможность наносить серьезный урон крупной борющейся добыче, такой как другие ранние киты. Подобные виды известны под общим названием «кашалоты-убийцы» или « кашалоты-макрорапториалы» . [ 58 ] [ 59 ]
Клюворылые киты состоят из более чем 20 родов. [ 60 ] [ 61 ] На более ранние разновидности, вероятно, охотились косатки и крупные акулы, такие как мегалодон . В 2008 году у берегов Южной Африки было обнаружено большое количество ископаемых зифиид, что подтвердило, что оставшиеся виды зифиид могут быть всего лишь остатком более высокого разнообразия, которое с тех пор вымерло. Изучив многочисленные ископаемые черепа, исследователи обнаружили отсутствие функциональных верхнечелюстных зубов у всех южноафриканских зифиид, что является свидетельством того, что всасывающее питание уже развилось у нескольких линий клюворылых китов в миоцене. У вымерших зифиидов также были крепкие черепа, что позволяет предположить, что бивни использовались для взаимодействия самцов с самцами. [ 60 ]
Скелетная эволюция
[ редактировать ]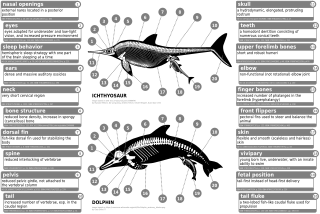
Современные китообразные имеют внутренние рудиментарные задние конечности, такие как редуцированные бедренные, малоберцовые и голени, а также тазовый пояс . У Индохиуса утолщена эктотимпанальная внутренняя губа ушной кости. Эта особенность напрямую сравнима с особенностью современных китообразных. Другой схожей особенностью был состав зубов, которые содержали в основном фосфат кальция , который необходим для еды и питья водным животным, хотя, в отличие от современных зубатых китов, у них был гетеродонтный (морфология более одного зуба) зубной ряд, в отличие от гомодонтный (имеется морфология одного зуба) зубной ряд. [ 62 ] Хотя они чем-то напоминали волков, окаменелости пакицетид показали, что их глазницы располагались гораздо ближе к макушке головы, чем у других наземных млекопитающих, но сходны со строением глаз китообразных. Их переход с суши в воду привел к изменению формы черепа и оборудования для обработки пищевых продуктов, поскольку изменились привычки питания. Изменение положения глаз и костей конечностей связано с превращением пакицетид в куликов. У амбулоцетид также начали развиваться длинные морды, что наблюдается у нынешних китообразных. Их конечности (и предполагаемые движения) были очень похожи на выдр. [ 63 ]
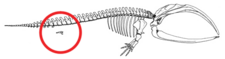
Отсутствие конечностей у китообразных не представляет собой регрессию полностью сформировавшихся конечностей или отсутствие зачатков конечностей, а скорее остановку развития зачатков конечностей. [ 64 ] Зачатки конечностей у эмбрионов китообразных развиваются нормально. [ 10 ] [ 65 ] Зачатки конечностей переходят к фазе конденсации раннего скелетогенеза, когда нервы врастают в зачатки конечностей, и апикальный эктодермальный гребень (AER), структура, обеспечивающая правильное развитие конечностей, становится функциональной. [ 64 ] [ 65 ] Иногда гены, отвечающие за более длинные конечности, приводят к развитию у современного кита миниатюрных ног ( атавизм ). [ 66 ]

У Pakicetus была тазовая кость, наиболее похожая на тазовую кость наземных млекопитающих. У более поздних видов, таких как Basilosaurus , тазовая кость, уже не прикрепленная к позвонкам и подвздошной кости , редуцировалась. [ 62 ] Считается, что за изменения, произошедшие в структуре таза китообразных, ответственны определенные гены, такие как BMP7 , PBX1 , PBX2 , PRRX1 и PRRX2 . [ 67 ] Когда-то считалось, что тазовый пояс современных китообразных представляет собой рудиментарную структуру , не служащую вообще никакой цели. Тазовый пояс самцов китообразных отличается по размеру от самок, и считается, что этот размер является результатом полового диморфизма. [ 68 ] Кости таза современных самцов китообразных массивнее, длиннее и крупнее, чем у самок. Из-за проявленного полового диморфизма они, скорее всего, участвовали в поддержании мужских гениталий, которые остаются скрытыми за стенками брюшной полости до тех пор, пока не произойдет половое размножение. [ 67 ] [ 69 ] [ 70 ]
У ранних археоцетов, таких как Pakicetus, носовые отверстия находились на конце морды , но у более поздних видов, таких как Rodhocetus , отверстия начали смещаться к верхней части черепа. Это известно как занос носа. [ 71 ] Ноздри современных китообразных превратились в дыхало , которые позволяют им с удобством вырываться на поверхность, вдыхать и погружаться под воду. Уши также начали двигаться внутрь, а в случае с базилозавром средние уши начали получать вибрации от нижней челюсти. Современные зубатые киты используют свой дынный орган — жировую подушечку — для эхолокации . [ 72 ]
Радиационные события
[ редактировать ]Есть три основных радиационных события, которые отмечают диверсификацию и видообразование в эволюционной истории китообразных. Первый произошел примерно в среднем эоцене (40 млн лет назад), когда эти ранние китообразные покинули речные и мелководные прибрежные места обитания, создав условия для появления Protocetidae – первых полностью морских китообразных. [ 1 ] Имея в своем распоряжении океаны и их питательные вещества, быстро диверсифицирующиеся протоцетиды также были ответственны за первую крупную географическую экспансию, распространившись по Северной Африке, Европе и Северной Америке. [ 19 ] Второе из трех крупных радиационных событий произошло в начале олигоцена (~ 34 млн лет назад), когда Neoceti отделились от Basilosauridae. [ 5 ] Это радиационное событие происходит одновременно с распадом Гондваны и открытием Южного океана, что приводит к резкому изменению океанских экосистем, продуктивности и температурных градиентов. [ 73 ] Время этого второго радиационного события не случайно, поскольку последующая диверсификация китообразных, вероятно, была связана с новыми экологическими возможностями, которые им предоставило изменение состояния океанов. [ 74 ] Последнее крупное радиационное событие, произошедшее в среднем миоцене и в плиоцене (от 12 до 2 млн лет назад), не было связано с каким-то конкретным событием, а связано с широким распространением зубатых и усатых китов. [ 19 ] [ 73 ] Стали появляться некоторые современные роды китообразных, в том числе Balaenoptera , род рорквалов, включающий синего кита. [ 19 ] [ 73 ] Delphinidae, океанские дельфины, также возникли во время этого радиационного события в позднем миоцене. [ 73 ]
Продолжающаяся эволюция
[ редактировать ]Культура
[ редактировать ]Культура — это групповое поведение, передаваемое посредством социального обучения. Одним из примеров является использование инструментов для поиска пищи. Использует ли дельфин инструмент или нет, это влияет на его пищевое поведение, что приводит к различиям в рационе. Кроме того, использование инструмента позволяет открыть новую нишу и новую добычу для этого конкретного дельфина. Из-за этих различий уровень приспособленности дельфинов в популяции меняется, что в конечном итоге приводит к эволюции. [ 76 ] Культура и социальные сети сыграли большую роль в эволюции современных китообразных, о чем свидетельствуют исследования, показывающие, что дельфины предпочитают партнеров с таким же социально усвоенным поведением, а горбатые киты используют песни между районами размножения. В частности, наибольшее негенетическое воздействие на эволюцию дельфинов обусловлено культурой и социальной структурой. [ 77 ]
По данным исследования 2014 года, популяцию афалин Индо-Тихоокеанского региона ( Tursiops sp.) в районе залива Шарк в Западной Австралии можно разделить на приживающихся и не приживающихся. Губки прикрепляют морские губки к морде для защиты от ссадин острыми предметами, зазубринами скатов или токсичными организмами. Губки также помогают дельфинам нацеливаться на рыбу без плавательного пузыря, поскольку эхолокация не может легко обнаружить эту рыбу на сложном фоне. Губки также добывают пищу в глубоких каналах, но негубцы добывают пищу как в глубоких, так и в мелких каналах. [ 78 ] Такое добывающее поведение в основном передается от матери к ребенку. Следовательно, поскольку это групповое поведение, передаваемое в результате социального обучения, использование этого инструмента считается культурной чертой. [ 76 ]
Исследователи в исследовании 2014 года в Шарк-Бэй обнаружили, что анализ жирных кислот между популяциями Западного и Восточного залива различается, что связано с тем, что в этих двух районах имеются разные источники пищи. Однако при сравнении данных из Западного залива у губачей и негубцев в глубоких каналах были очень разные результаты по жирным кислотам, даже если они находились в одной и той же среде обитания. Неспонги из глубоких и неглубоких каналов имели схожие данные. Это говорит о том, что причиной разных данных было губчатое воздействие, а не глубокие и неглубокие каналы. Обтирание открыло для дельфинов новую нишу и позволило им получить доступ к новой добыче, что привело к долгосрочным изменениям в рационе. Благодаря производству различных источников пищи внутри популяции снижается внутрипопуляционная конкуренция за ресурсы, что свидетельствует о смещении характера . В результате увеличивается пропускная способность, поскольку все население не зависит от одного источника питания. Уровень физической подготовки среди населения также меняется, что позволяет этой культуре развиваться. [ 78 ]
Социальная структура
[ редактировать ]Социальная структура образует группы, состоящие из людей, которые взаимодействуют друг с другом, и это позволяет культурным чертам возникать, обмениваться и развиваться. Эта связь особенно заметна в популяциях афалин на юго-западе Австралии, которые, как известно, выпрашивают еду у рыбаков. Такое поведение попрошайничества распространилось среди населения в результате индивидуального (дельфины проводят время возле лодок) и социального (дельфины проводят время с другими дельфинами, которые выражают поведение попрошайничества) обучения. [ 76 ]
Однако культура может влиять на социальную структуру, вызывая совпадение поведения и ассортативное спаривание. Люди внутри определенной культуры с большей вероятностью будут спариваться с людьми, использующими такое же поведение, а не со случайным человеком, тем самым влияя на социальные группы и структуру. Например, обитатели Шарк-Бэй предпочитают держаться вместе с другими приживальщиками. [ 76 ] Кроме того, некоторые афалины в заливе Мортон в Австралии следовали за с креветками траулерами , чтобы питаться их отходами, в то время как другие дельфины в той же популяции этого не делали. Дельфины преимущественно общались с особями с одинаковым поведением, хотя все они жили в одной среде обитания. Позже креветочных траулеров уже не было, а дельфины через пару лет объединились в одну социальную сеть. [ 76 ]
Социальные сети по-прежнему могут влиять и вызывать эволюцию сами по себе, вызывая у людей различия в приспособленности. [ 79 ] Согласно исследованию 2012 года, телята-самцы имели более низкую выживаемость, если у них были более прочные связи с молодыми самцами. Однако когда были протестированы другие возрастные и половые классы, их выживаемость существенно не изменилась. [ 80 ] Это говорит о том, что молодые самцы создают социальный стресс для своих более молодых сверстников. Фактически, было документально подтверждено, что молодые самцы обычно совершают акты агрессии, доминирования и запугивания в отношении телят-самцов. [ 80 ] Согласно исследованию 2010 года, некоторые популяции дельфинов залива Шарк имели разный уровень физической подготовки и успешности детенышей. Это происходит либо из-за социального обучения (независимо от того, передала ли мать свои знания о репродуктивной способности детенышам), либо из-за прочной связи между матерями-дельфинами в популяции; держась в группе, отдельной матери не нужно постоянно проявлять бдительность в отношении хищников. [ 79 ]
Генетические исследования, проведенные на дельфинах-клименах ( Stenella clymene ), были сосредоточены на их естественной истории, и результаты показывают, что происхождение вида на самом деле было результатом гибридного видообразования . [ 81 ] [ 82 ] Гибридизация дельфинов-спиннеров ( Stenella longirostris ) и полосатых дельфинов ( Stenella coeruleoalba ) в Северной Атлантике была вызвана постоянным разделением среды обитания этих двух видов. Отношения между этими тремя видами предполагались на основании заметного сходства между анатомией климены и дельфинов-прядильщиков, в результате чего первые до 1981 года считались подвидами вторых. [ 83 ] и возможность существования клименского дельфина как гибрида прядильщика и полосатых дельфинов оказалась под вопросом из-за анатомического и поведенческого сходства между этими двумя видами. [ 84 ]
Факторы окружающей среды
[ редактировать ]Секвенирование генома, проведенное в 2013 году, показало, что у дельфина реки Янцзы , или «байцзи» ( Lipotes vexillifer ), отсутствуют однонуклеотидные полиморфизмы в геноме . После реконструкции истории генома байджи этого вида дельфинов исследователи обнаружили, что значительное снижение генетического разнообразия произошло, скорее всего, из-за узкого места во время последнего явления дегляциации . В этот период уровень моря повышался, а глобальная температура повышалась. Другие исторические климатические события также можно соотнести и сопоставить с историей генома дельфина реки Янцзы. Это показывает, как глобальное и локальное изменение климата может радикально повлиять на геном, приводя к изменениям в приспособленности, выживании и эволюции вида. [ 85 ]
Европейская популяция обыкновенных дельфинов ( Delphinus delphis ) в Средиземноморье дифференцировалась на два типа: восточный и западный. Согласно исследованию 2012 года, это, по-видимому, также связано с недавним узким местом, которое резко сократило численность населения восточного Средиземноморья. Кроме того, отсутствие структуры популяций между западными и восточными регионами кажется противоречащим различиям в структурах популяций между другими регионами обитания дельфинов. [ 86 ] Несмотря на то, что у дельфинов Средиземноморья не было физического барьера между своими регионами, они все же разделились на два типа из-за экологии и биологии. Таким образом, различия между восточными и западными дельфинами, скорее всего, связаны с выбором узкоспециализированной ниши, а не просто с физическими барьерами. Благодаря этому окружающая среда играет большую роль в дифференциации и эволюции этого вида дельфинов. [ 87 ]
Дивергенция и видообразование афалин во многом были связаны с изменениями климата и окружающей среды на протяжении истории. Согласно исследованиям, подразделения внутри рода коррелируют с периодами быстрого изменения климата. Например, изменение температуры может привести к изменению прибрежного ландшафта, опустошению ниш и появлению возможностей для разделения. [ 88 ] В частности, генетические данные свидетельствуют о том, что в Северо-Восточной Атлантике дельфины-афалины дифференцировались на прибрежные и пелагические типы. Расхождение, скорее всего, связано с событием основания, когда большая группа отделилась. После этого события отдельные группы соответствующим образом адаптировались и сформировали свои собственные ниши специализации и социальные структуры. Эти различия привели к тому, что две группы разошлись и остались разделенными. [ 89 ]
Два эндемичных, характерных типа гринд с короткими плавниками : Таппанага (или Сиогонду ), более крупный северный тип, и Магондоу, меньший южный тип, можно встретить вдоль Японского архипелага, где распространение этих двух типов в основном не пересекается с океаническими водами. передняя граница вокруг самой восточной точки Хонсю . Считается, что локальное исчезновение длинноплавниковых гринд в северной части Тихого океана в XII веке могло спровоцировать появление таппанаги , в результате чего короткоплавниковые гринды колонизировали более холодные ареалы длинноплавникового варианта. [ 90 ] [ 91 ] Китов с характеристиками, похожими на Таппанагу, можно встретить вдоль острова Ванкувер и у северного побережья США. [ 92 ]
См. также
[ редактировать ]- Водная адаптация
- Эволюция млекопитающих
- Эволюция сирен
- Список вымерших китообразных
- Переходная форма
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Тьювиссен, JGM; Уильямс, EM (1 ноября 2002 г.). «Ранняя радиация китообразных (млекопитающих): закономерности эволюции и корреляции развития». Ежегодный обзор экологии и систематики . 33 (1): 73–90. doi : 10.1146/annurev.ecolsys.33.020602.095426 .
- ^ Дэвис, RW 2019. Возвращение в море: эволюция морских млекопитающих. Страницы 7–27 в книге RW Davis, изд. Морские млекопитающие: приспособления к водной жизни. Springer International Publishing, Нью-Йорк.
- ^ Гейслер, Дж. Х. и доктор медицинских наук Уэн. 2005. Филогенетические взаимоотношения вымерших цетартиодактилей: результаты одновременного анализа молекулярных, морфологических и стратиграфических данных. Журнал эволюции млекопитающих 12: 145–160.
- ^ Берта, А., Дж. Л. Сумич и К. М. Ковач. 2005. Морские млекопитающие: эволюционная биология. Эльзевир, Амстердам.
- ^ Перейти обратно: а б Тьювиссен, Дж.Г.М., Л.Н. Купер, Дж.К. Джордж и С. Баджпай. 2009. От суши к воде: происхождение китов, дельфинов и морских свиней. Эволюция: Образование и просвещение 2: 272–288.
- ^ Никайдо, М.; Мацуно, Ф. (2001). «Ретропозонный анализ основных линий китообразных: монофилия зубатых китов и парафилия речных дельфинов» . Труды Национальной академии наук . 98 (13): 7384–9. Бибкод : 2001PNAS...98.7384N . дои : 10.1073/pnas.121139198 . ПМК 34678 . ПМИД 11416211 .
- ^ Перейти обратно: а б с Тьювиссен , JGM; Купер, Лиза Ноэль; Клементц, Марк Т.; Баджпай, Сунил; Тивари, Б.Н. (20 декабря 2007 г.). «Киты произошли от водных парнокопытных в эпоху эоцена Индии». Природа . 450 (7173): 1190–1194. Бибкод : 2007Natur.450.1190T . дои : 10.1038/nature06343 . ПМИД 18097400 . S2CID 4416444 .
- ^ Роуч, Джон (2011). «Найден самый старый антарктический кит; демонстрирует быструю эволюцию» . Архивировано из оригинала 18 ноября 2011 года . Проверено 3 января 2018 г.
- ^ Хун-Янь, Гао; Си-Цзюнь, Ни (2015). «Разнообразные стволовые китообразные и их филогенетические связи с мезонихидами и парнокопытными» (PDF) . Позвоночные Палазиатские . 53 (2): 165.
- ^ Перейти обратно: а б с Калифорнийский университет, Беркли (7 февраля 2005 г.). «Калифорнийский университет в Беркли, французские ученые находят недостающее звено между китом и его ближайшим родственником, бегемотом» . Проверено 21 декабря 2007 г.
{{cite newsgroup}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Перейти обратно: а б Л. Фентон, Кэрролл; В. Рич, Пэт; А. Фентон, Милдред; Х.В. Рич, Томас (1996). «Ранние млекопитающие» . Книга окаменелостей: летопись доисторической жизни . Дуврское издательство. стр. 547–548 . ISBN 978-0-486-29371-4 .
- ^ Перейти обратно: а б Медицинские и фармацевтические колледжи северо-восточных университетов Огайо (2007 г.). «Киты произошли от крошечных предков, похожих на оленей» . Проверено 21 декабря 2007 г.
- ^ Мичиганский университет (2001). «Новые окаменелости позволяют предположить, что киты и бегемоты являются близкими родственниками» . Проверено 21 декабря 2007 г.
- ^ Ян Сэмпл (2007). «Киты могут произойти от небольшого животного, похожего на оленя» . Проверено 21 декабря 2007 г.
- ^ Образец, Ян; корреспондент, наука (20 декабря 2007). «От Бэмби до Моби-Дика: Как маленький олень превратился в кита» . Хранитель . ISSN 0261-3077 . Проверено 16 мая 2020 г.
{{cite news}}:|last2=имеет общее имя ( справка ) - ^ Джинджерич, Филип Д.; Рассел, Делавэр (1981). « Pakicetus inachus , новый археоцитов (Mammalia, Cetacea) из ранне-среднеэоценовой формации Кулдана Кохата (Пакистан)». Музей палеонтологии . 25 : 235–246.
- ^ Перейти обратно: а б Кастро, Питер; Хубер, Майкл Э. (2003). Морская биология (4-е изд.) . МакГроу-Хилл.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Тьювиссен, JGM ; Баджпай, Сунхил (2001). «Происхождение китов как образец макроэволюции» . Бионаука . 51 (12): 1037. doi : 10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2 . ISSN 0006-3568 .
- ^ Перейти обратно: а б с д Уэн, Марк (2010). «Происхождение китов». Ежегодный обзор наук о Земле и планетах . 38 (1): 189–219. Бибкод : 2010AREPS..38..189U . doi : 10.1146/annurev-earth-040809-152453 .
- ^ Перейти обратно: а б с д и ж г Нуммела, Сирпа; Тьювиссен, JGM; Баджпай, Сунил; Хусейн, С. Тасир; Кумар, Кишор (11 августа 2004 г.). «Эоценовая эволюция слуха китов». Природа . 430 (7001): 776–778. Бибкод : 2004Natur.430..776N . дои : 10.1038/nature02720 . ПМИД 15306808 . S2CID 4372872 .
- ^ Купер, Лиза Н.; Тьювиссен, JGM ; Хусейн, ST (2009). «Новые среднеэоценовые археоцеты (Cetacea: Mammalia) из формации Кулдана в Северном Пакистане». Журнал палеонтологии позвоночных . 29 (4): 1289–1299. Бибкод : 2009JVPal..29.1289C . дои : 10.1671/039.029.0423 . S2CID 84127292 .
- ^ Перейти обратно: а б JGM Тьювиссен ; Э.М. Уильямс; Эл Джей Роу; СТ Хусейн (2001). «Скелеты наземных китообразных и родство китов с парнокопытными». Природа . 413 (6853): 277–281. Бибкод : 2001Natur.413..277T . дои : 10.1038/35095005 . ПМИД 11565023 . S2CID 4416684 .
- ^ Тьювиссен , JGM; Уильямс, EM; Роу, Эл Джей; Хусейн, ST (2001). «Скелеты наземных китообразных и родство китов с парнокопытными». Природа . 413 (6853): 277–281. Бибкод : 2001Natur.413..277T . дои : 10.1038/35095005 . ПМИД 11565023 . S2CID 4416684 .
- ^ Тьювиссен, JGM ; Хуссейн, Южная Каролина; Алиф, М. (1994). «Ископаемые свидетельства происхождения водного передвижения археокитовых китов». Наука . 263 (5144): 210–212. Бибкод : 1994Sci...263..210T . дои : 10.1126/science.263.5144.210 . ПМИД 17839179 . S2CID 20604393 .
- ^ Тьювиссен, JG M; FEFish (август 1997 г.). «Локомоторная эволюция у древнейших китообразных: функциональная модель, современные аналоги и палеонтологические данные». Палеобиология . 23 (4): 482–490. Бибкод : 1997Pbio...23..482T . дои : 10.1017/S0094837300019850 . S2CID 51685113 .
- ^ Андо, Konami (2016). «Прощание с жизнью на суше – сила грудной клетки как новый индикатор для определения палеоэкологии вторичных водных млекопитающих» . Журнал анатомии . 229 (6): 768–777. дои : 10.1111/joa.12518 . ПМК 5108153 . ПМИД 27396988 .
- ^ Перейти обратно: а б с д и ж г Фордайс, RE; Барнс, LG (30 апреля 1994 г.). «Эволюционная история китов и дельфинов». Ежегодный обзор наук о Земле и планетах . 22 (1): 419–455. Бибкод : 1994AREPS..22..419F . doi : 10.1146/annurev.ea.22.050194.002223 .
- ^ Перейти обратно: а б Спур, Ф.; Баджпай, С.; Хуссейн, Южная Каролина; Кумар, К.; Тьювиссен , JGM (8 мая 2002 г.). «Вестибулярные данные эволюции водного поведения ранних китообразных». Природа . 417 (6885): 163–166. Бибкод : 2002Natur.417..163S . дои : 10.1038/417163а . ПМИД 12000957 . S2CID 4331789 .
- ^ Гоар, А.С.; Антар, М.С.; Бессенекер, RW; Сабри, Д.А.; Эль-Сайед, С; Зейферт, ER; Залмут, Исландия; Саллам, Ее Величество (25 августа 2021 г.). «Новый протоцетидный кит предлагает ключ к разгадке биогеографии и экологии питания на ранней стадии эволюции китообразных» . Слушания. Биологические науки . 288 (1957): 20211368. doi : 10.1098/rspb.2021.1368 . ПМЦ 8385364 . ПМИД 34428967 .
- ^ Ламберт, О.; Бьянуччи, Г.; Салас-Жисмонди, Р.; Ди Сельма, К.; Стербо, Э.; Урбина, М.; де Мюзон, М. (2019). «Кит-амфибия из среднего эоцена Перу обнаруживает раннее распространение четвероногих китообразных в южной части Тихого океана» . Современная биология . 29 (8): 1352–1359.e3. Бибкод : 2019CBio...29E1352L . дои : 10.1016/j.cub.2019.02.050 . hdl : 11581/425570 . ПМИД 30955933 .
- ^ Джинджерич, доктор медицинских наук; уль-Хак, М.; фон Кенигсвальд, Ж; Сандерс, WJ; Смит, Б.Х. (2009). «Новый протоцетидный кит из среднего эоцена Пакистана: рождение на суше, раннее развитие и половой диморфизм» . ПЛОС ОДИН . 4 (2): e4366. Бибкод : 2009PLoSO...4.4366G . дои : 10.1371/journal.pone.0004366 . ПМЦ 2629576 . ПМИД 19194487 .
- ^ Мадар, Сандра И. (1998). «Структурные адаптации длинных костей ранних археоцетов». В Тьювиссене , JGM (ред.). Появление китов . Достижения в палеобиологии позвоночных. Том. 1. С. 353–378. дои : 10.1007/978-1-4899-0159-0_12 . ISBN 978-1-4899-0161-3 .
- ^ Джинджерич, Филип Д.; Мунир уль Хак; Лиад С. Залмут; Интизар Хиссейн Хан; М. Садик Малкани (21 сентября 2001 г.). «Происхождение китов от ранних парнокопытных: руки и ноги эоценовых протоцетид из Пакистана» . Наука . 293 (5538): 2239–42. Бибкод : 2001Sci...293.2239G . дои : 10.1126/science.1063902 . ПМИД 11567134 . S2CID 21441797 .
- ^ Антар, Мохаммед С.; Гохар, Абдулла С.; Эль-Десуки, Хеба; Зайферт, Эрик Р.; Эль-Сайед, Сана; Клэкстон, Александр Г.; Саллам, Хешам М. (10 августа 2023 г.). «Миниатюрный новый кит-басилозавр раскрывает траекторию истории жизни китообразных в эоцене» . Коммуникационная биология . 6 (1): 707. doi : 10.1038/s42003-023-04986-w . ISSN 2399-3642 . ПМЦ 10415296 . ПМИД 37563270 .
- ^ Уссей, Александра; Таффоро, Пол; де Мюизон, Кристиан; Джинджерич, Филип Д. (2015). «Переход эоценовых китов с суши в море: данные по микроструктуре костей» . ПЛОС ОДИН . 10 (2): e0118409. Бибкод : 2015PLoSO..1018409H . дои : 10.1371/journal.pone.0118409 . ПМК 4340927 . ПМИД 25714394 .
- ^ Н. Уилфорд, Джон (1990). «Задние лапы китов обнаруживаются в окаменелостях» . Проверено 1 марта 2016 г.
- ^ Демере, Т.А.; Макгоуэн, MR; Берта, А.; Гейтси, Дж. (2008). «Морфологические и молекулярные доказательства поэтапного эволюционного перехода от зубов к усатым у усатых китов» . Систематическая биология . 57 (1): 15–37. дои : 10.1080/10635150701884632 . ПМИД 18266181 .
- ^ Демере, Томас; Майкл Р. Макгоуэн; Анналиса Берта; Джон Гейси (сентябрь 2007 г.). «Морфологические и молекулярные доказательства поэтапного эволюционного перехода от зубов к усатым у усатых китов» . Систематическая биология . 57 (1): 15–37. дои : 10.1080/10635150701884632 . ПМИД 18266181 .
- ^ М.Г. Фицджеральд, Эрих (2012). «Челюсти археокитообразного усатого кита» . Письма по биологии . 8 (1): 94–96. дои : 10.1098/rsbl.2011.0690 . ПМК 3259978 . ПМИД 21849306 .
- ^ Перейти обратно: а б Демере, Томас А.; Макгоуэн, Майкл Р.; Берта, Анналиса; Гейтси, Джон (1 февраля 2008 г.). «Морфологические и молекулярные доказательства поэтапного эволюционного перехода от зубов к усатым у усатых китов» . Систематическая биология . 57 (1): 15–37. дои : 10.1080/10635150701884632 . ISSN 1063-5157 . ПМИД 18266181 .
- ^ Мередит, RW; Гейтси, Дж. (2010). «Псевдогенизация зубного гена эмализина (MMP20) у общего предка современных усатых китов» . Труды Королевского общества Б. 278 (1708): 993–1002. дои : 10.1098/rspb.2010.1280 . ПМК 3049022 . ПМИД 20861053 .
- ^ Э. Фордайс, Р.; Г. Маркс, Феликс (2012). «Карликовый кит Caperea Marginata : последний из цетотериев» . Слушания. Биологические науки . 280 (1753): 20122645. doi : 10.1098/rspb.2012.2645 . ПМЦ 3574355 . ПМИД 23256199 .
- ^ Бисконти, Микеланджело; Ламберт, Оливье; Босселерс, Марк (2013). «Таксономический пересмотр Isocetus depauwi (Mammalia, Cetacea, Mysticeti) и филогенетические отношения архаичных китов-китотеров» . Палеонтология . 56 (1): 95–127. Бибкод : 2013Palgy..56...95B . дои : 10.1111/j.1475-4983.2012.01168.x .
- ^ Бисконти, Микеланджело; Пеллегрино, Лука; Карневале, Джорджио (18 июля 2021 г.). «Эволюция гигантизма у гладких и гренландских китов (Cetacea: Mysticeti: Balaenidae)» . Биологический журнал Линнеевского общества . 134 (2): 498–524. doi : 10.1093/biolinnean/blab086 . ISSN 0024-4066 .
- ^ Бьянуччи, Джованни; Маркс, Феликс Г.; Колларета, Альберто; Ди Стефано, Агата; Ландини, Уолтер; Мориги, Катерина; Варола, Анджело (май 2019 г.). «Восстание титанов: усатые киты стали гигантами раньше, чем считалось» . Письма по биологии . 15 (5): 20190175. doi : 10.1098/rsbl.2019.0175 . ISSN 1744-9561 . ПМК 6548731 . ПМИД 31039728 .
- ^ Слейтер, Грэм Дж.; Голдбоген, Джереми А.; Пьенсон, Николас Д. (24 мая 2017 г.). «Независимая эволюция гигантизма усатых китов, связанная с динамикой океана плио-плейстоцена» . Труды Королевского общества B: Биологические науки . 284 (1855): 20170546. doi : 10.1098/rspb.2017.0546 . ISSN 0962-8452 . ПМЦ 5454272 . ПМИД 28539520 .
- ^ Метте Э. Стиман; Мартин Б. Хебсгаард; Р. Юэн Фордайс; Саймон Ю.В. Хо; Дэниел Л. Рабоски; Расмус Нильсен; Карстен Рахбек; Хенрик Гленнер; Мартин В. Соренсен; Эске Виллерслев (24 августа 2009 г.). «Радиация современных китообразных, вызванная реструктуризацией океанов» . Систематическая биология . 58 (6): 573–585. дои : 10.1093/sysbio/syp060 . ПМЦ 2777972 . ПМИД 20525610 .
- ^ Александр Дж. П. Хубен1; Питер К. Бейл; Йорг Просс; Стивен М. Бохати; Сандра Пасшир; Кэтрин Э. Стикли; Урсула Рёль; Сайко Сугисаки; Лиза Токс; Тина ван де Флирдт; Мэтью Олни; Франческа Санджорджи; Аппи Слуйс; Карлота Эскутия; Хенк Бринкхейс (19 апреля 2013 г.). «Реорганизация планктонной экосистемы Южного океана с началом антарктического оледенения». Наука . 340 (6130): 341–344. Бибкод : 2013Sci...340..341H . дои : 10.1126/science.1223646 . hdl : 1874/385683 . ПМИД 23599491 . S2CID 30549019 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Нуммела1, Сирпа; Тьювиссен, JGM ; Баджпай, Сунил; Т. Хусейн, С.; Кумар, Кишор (2004). «Эоценовая эволюция слуха китов». Природа . 430 (7001): 776–778. Бибкод : 2004Natur.430..776N . дои : 10.1038/nature02720 . ПМИД 15306808 . S2CID 4372872 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Уитмор-младший, ФК; Сандерс, А.Е. (1976). «Обзор олигоценовых китообразных» . Систематическая зоология . 25 (4): 304–320. дои : 10.2307/2412507 . JSTOR 2412507 .
- ^ Каузак, Бруно; Бюиссон, Себастьен; Помье, Мишель; Роше, Филипп (2006). «Обнаружение двух зубов сквалодона (Cetacea Odontoceti) у бурдигалов на юго-западе Франции (Мартильяк, Леоньян). Соображения о сквалодоне Аквитании, палеоэкологии их гнезд и типовых видах рода». Новогодний альбом по геологии и палеонтологии (на французском языке). 238 (3): 413–451. дои : 10.1127/njgpa/238/2006/413 .
- ^ Маркс, Феликс Г.; Ламберт, Оливье; Уэн, Марк Д. (2016). Палеобиология китообразных . Джон Уайли и сыновья. стр. 242–243. ISBN 978-111-856127-0 .
- ^ Рэйчел А. Расикот; Томас А. Демере; Брайан Л. Битти; Роберт В. Бессенекер (2014). «Уникальная морфология питания новой вымершей прогнатной морской свиньи из плиоцена Калифорнии» . Современная биология . 24 (7): 774–779. Бибкод : 2014CBio...24..774R . дои : 10.1016/j.cub.2014.02.031 . ПМИД 24631245 .
- ^ Гаскин, Дэвид Э. (1984). Макдональд, Д. (ред.). Энциклопедия млекопитающих . Нью-Йорк: факты в архиве. стр. 196–199 . ISBN 978-0-87196-871-5 .
- ^ Барнс, Лоуренс Г. Ископаемые зубатые киты (Mammalia: Cetacea) из формации Альмехас, остров Седрос, Мексика . Калифорнийский университет, Музей палеонтологии. п. 46. СОЛЬ B0006YRTU4 .
- ^ Барнс, Л.Г. (1977). «Очерк ископаемых китообразных восточной части северной части Тихого океана». Систематическая зоология . 25 (4): 321–343. дои : 10.2307/2412508 . JSTOR 2412508 .
- ^ Перрин, Уильям Ф.; Вюрсиг, Бернд Г.; Тьювиссен , JGM (2009). Энциклопедия морских млекопитающих (2-е изд.). Академическая пресса. п. 214. ИСБН 978-0-12-373553-9 .
- ^ Перейти обратно: а б Бьянуччи, Г.; Ландини, В. (8 сентября 2006 г.). «Кашалот-убийца: новый базальный физетероид (Mammalia, Cetacea) из позднего миоцена Италии» . Зоологический журнал Линнеевского общества . 148 (1): 103–131. дои : 10.1111/j.1096-3642.2006.00228.x .
- ^ Брайан Свитек. «Кашалоты: долгая и порочная история» . Проверено 12 августа 2015 г.
- ^ Перейти обратно: а б Джованни Бьянуччи; Клаас Пост; Оливье Ламбер (2008). «Тайны клюворылых китов, раскрытые окаменелостями морского дна, выловленными тралом у берегов Южной Африки» . Пизанский университет, Музей естественной истории Роттердама, Королевский институт естественных наук Бельгии . 104 (3–4): 140–142.
- ^ Бьянуччи, Джованни; Михан, Исмаэль; Ламберт, Оливье; Пост, Клаас; Матеус, Октавио (2013). «Причудливые ископаемые клюворылы (Odontoceti, Ziphiidae), выловленные на дне Атлантического океана у Пиренейского полуострова» (PDF) . Геодиверситас . 35 (1): 105–153. дои : 10.5252/g2013n1a6 . S2CID 129787505 .
- ^ Перейти обратно: а б Тьювиссен, JGM ; Купер, Лиза Н.; Джордж, Джон К.; Баджпай, Сунил (2009). «От суши к воде: происхождение китов, дельфинов и морских свиней» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 272–288. дои : 10.1007/s12052-009-0135-2 .
- ^ Тьювиссен, JGM ; Уильямс, EM; Роу, Эл Джей; Хусейн, ST (2001). «Скелеты наземных китообразных и связь китов с парнокопытными» (PDF) . Природа . 413 (6853): 277–281. Бибкод : 2001Natur.413..277T . дои : 10.1038/35095005 . ПМИД 11565023 . S2CID 4416684 . Архивировано из оригинала (PDF) 4 марта 2016 года . Проверено 16 января 2016 г. .
- ^ Перейти обратно: а б Бейдер, Л. (2002). «Конечности у китов и отсутствие конечностей у других позвоночных: механизмы эволюционной и развивающей трансформации и утраты». Эволюция и развитие . 4 (6): 445–58. дои : 10.1046/j.1525-142X.2002.02033.x . ПМИД 12492145 . S2CID 8448387 .
- ^ Перейти обратно: а б Тьювиссен, JGM ; Кон, MJ; Стивенс, Л.С.; Баджпай, С.; Хейнинг, Дж.; Хортон-младший, МЫ (2006). «Основы развития потери задних конечностей у дельфинов и происхождение строения тела китообразных» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (22): 8414–8418. Бибкод : 2006PNAS..103.8414T . дои : 10.1073/pnas.0602920103 . ПМК 1482506 . ПМИД 16717186 .
- ^ Бейдер, Ларс; Холл, Брайан К. (2002). «Конечности у китов и отсутствие конечностей у других позвоночных: механизмы эволюционной и развивающей трансформации и утраты». Эволюция и развитие . 4 (6): 445–458. дои : 10.1046/j.1525-142X.2002.02033.x . ПМИД 12492145 . S2CID 8448387 .
- ^ Перейти обратно: а б Гольдин, Павел (2014). «Именование безымянного: таз и задние конечности миоценовых китов дают представление об эволюции и гомологии тазового пояса китообразных». Эволюционная биология . 41 (3): 473–479. Бибкод : 2014EvBio..41..473G . дои : 10.1007/s11692-014-9281-8 . S2CID 15055515 .
- ^ Дайнс, Джеймс П.; и др. (2014). «Половой отбор нацелен на тазовые кости китообразных» . Эволюция . 68 (11): 3296–3306. дои : 10.1111/evo.12516 . ПМК 4213350 . ПМИД 25186496 .
- ^ Каори, Онбе; Нисида, Шин; Соне, Эми; Канда, Наохиса; Гото, Муцуо; А. Пастене, Луис; Танабэ, Синсуке; Койке, Хироко (2007). «Вариация последовательности гена Tbx4 у морских млекопитающих». Зоологическая наука . 24 (5): 449–464. дои : 10.2108/zsj.24.449 . ПМИД 17867844 . S2CID 31047933 .
- ^ Тадзима, Юко; Ёсихиро, Хаяси; Тадасу, Ямада (2004). «Сравнительное анатомическое исследование взаимоотношений между рудиментарными тазовыми костями и окружающими структурами бесплавных морских свиней» . Журнал ветеринарной медицины . 66 (7): 761–766. дои : 10.1292/jvms.66.761 . ПМИД 15297745 .
- ^ Т. Джонсон, Артур (2011). «Принципы биологии» . Биология для инженеров . ЦРК Пресс. п. 227. ИСБН 978-1-4200-7763-6 .
- ^ Ямато, Майя; Пьенсон, Николас Д. (2015). «Раннее развитие и ориентация акустической воронки дает представление об эволюции путей восприятия звука у китообразных» . ПЛОС ОДИН . 10 (3): e0118582. Бибкод : 2015PLoSO..1018582Y . дои : 10.1371/journal.pone.0118582 . ПМК 4356564 . ПМИД 25760328 .
- ^ Перейти обратно: а б с д Фордайс, Р. Юэн (1 января 2018 г.), «Эволюция китообразных» , Вюрсиг, Бернд; Тьювиссен, JGM; Ковач, Кит М. (ред.), Энциклопедия морских млекопитающих (третье издание) , Academic Press, стр. 180–185, ISBN 978-0-12-804327-1 , получено 13 ноября 2022 г.
- ^ Фордайс, Р. Юэн; Маркс, Феликс Г. (21 мая 2018 г.). «Гигантизм предшествует фильтрованному питанию в эволюции усатых китов» . Современная биология . 28 (10): 1670–1676.e2. Бибкод : 2018CBio...28E1670F . дои : 10.1016/j.cub.2018.04.027 . ISSN 0960-9822 . ПМИД 29754903 . S2CID 21680283 .
- ^ Паттерсон, Э.М.; Манн, Дж. (2011). «Экологические условия, благоприятствующие использованию орудий и инновациям у диких афалин (Tursiops sp.)» . ПЛОС ОДИН . 6 (е22243): е22243. Бибкод : 2011PLoSO...622243P . дои : 10.1371/journal.pone.0022243 . ПМК 3140497 . ПМИД 21799801 .
- ^ Перейти обратно: а б с д и Кантор, М.; Уайтхед, Х. (2013). «Взаимодействие между социальными сетями и культурой: теоретически и среди китов и дельфинов» . Философские труды Королевского общества B: Биологические науки . 368 (1618): 20120340. doi : 10.1098/rstb.2012.0340 . ПМЦ 3638443 . ПМИД 23569288 .
- ^ Кантор, М.; Уайтхед, Х. (2013). «Взаимодействие между социальными сетями и культурой: теоретически и среди китов и дельфинов» . Философские труды Королевского общества B: Биологические науки . 368 (1618): 20120340. doi : 10.1098/rstb.2012.0340 . ПМЦ 3638443 . ПМИД 23569288 .
- ^ Перейти обратно: а б Круцен, М.; Крейкер, С.; Д. Маклауд, К.; Лермонт, Дж.; М. Коппс, А.; Уолшем, П.; Дж. Аллен, С. (2014). «Культурная передача использования орудий индо-тихоокеанскими афалинами ( Tursiops sp.) обеспечивает доступ к новой кормовой нише» . Труды Королевского общества B: Биологические науки . 281 (1784): 20140374. doi : 10.1098/rspb.2014.0374 . ПМК 4043097 . ПМИД 24759862 .
- ^ Перейти обратно: а б Фрер, Швейцария; Круцен, М.; Манн, Дж.; Коннор, RC; Бейдер, Л.; Шервин, ВБ (2010). «Социальные и генетические взаимодействия приводят к изменению приспособленности свободноживущей популяции дельфинов» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 19, 949–19, 954. Бибкод : 2010PNAS..10719949F . дои : 10.1073/pnas.1007997107 . ПМЦ 2993384 . ПМИД 21041638 .
- ^ Перейти обратно: а б Стэнтон, Массачусетс; Манн, Дж. (2012). «Ранние социальные сети предсказывают выживание диких афалин» . ПЛОС ОДИН . 7 (10): е47508. Бибкод : 2012PLoSO...747508S . дои : 10.1371/journal.pone.0047508 . ПМЦ 3471847 . ПМИД 23077627 .
- ^ Амарал, Арканзас; Лаввелл, Г.; Коэльо, ММ; Амато, Г.; Розенбаум, ХК (2014). Джонсон, Норман (ред.). «Гибридное видообразование морских млекопитающих: клименовый дельфин ( Stenella clymene )» . ПЛОС ОДИН . 9 (1): е83645. Бибкод : 2014PLoSO...983645A . дои : 10.1371/journal.pone.0083645 . ПМЦ 3885441 . ПМИД 24421898 .
- ^ Чой, Чарльз (2014). «Открытие ДНК раскрывает удивительное происхождение дельфинов» . Архивировано из оригинала 12 января 2014 года . Проверено 2 января 2016 г.
- ^ Перрин, ВФ; Митчелл, Эд; Мид, Дж. Г.; Колдуэлл, Дания; ван Бри, PJH (1981). « Стенелла Климена , вновь открытый тропический дельфин Атлантики». Журнал маммологии . 62 (3): 583–598. дои : 10.2307/1380405 . JSTOR 1380405 .
- ^ В. Чой, Чарльз (2014). «Открытие ДНК раскрывает удивительное происхождение дельфинов» . Нэшнл Географик . Архивировано из оригинала 12 января 2014 года . Проверено 20 января 2016 г.
- ^ Чжоу, X.; Сан, Ф.; Сюй, С.; Фан, Г.; Чжу, К.; Лю, X.; Ян, Г. (2013). «Геномы байджи демонстрируют низкую генетическую изменчивость и позволяют по-новому взглянуть на вторичные водные адаптации» . Природные коммуникации . 4 : 2708. Бибкод : 2013NatCo...4.2708Z . дои : 10.1038/ncomms3708 . ПМЦ 3826649 . ПМИД 24169659 .
- ^ Моура, А.Е.; Натоли, А.; Роган, Э.; Хельзель, Арканзас (2012). «Атипичная панмиксия у европейского вида дельфинов ( Delphinus delphis ): последствия для эволюции разнообразия за пределами границ океана» . Журнал эволюционной биологии . 26 (1): 63–75. дои : 10.1111/jeb.12032 . ПМИД 23205921 . S2CID 7205948 .
- ^ Беарзи, Г.; Фортуна, К.; Ривз, Р. (2012). « Средиземноморская субпопуляция Tursiops truncatus » . Красный список исчезающих видов МСОП . 2012 : e.T16369383A16369386. doi : 10.2305/IUCN.UK.2012-1.RLTS.T16369383A16369386.en . Проверено 19 ноября 2021 г.
- ^ Моура, А.Е.; Нильсен, SCA; Вилструп, Дж. Т.; Морено-Майар, СП; Гилберт, MTP; Грей, HWI; Хельзель, Арканзас (2013). «Недавняя диверсификация морского рода ( Tursiops spp.) Отслеживает предпочтения среды обитания и изменения окружающей среды» . Систематическая биология . 62 (6): 865–877. дои : 10.1093/sysbio/syt051 . ПМИД 23929779 .
- ^ Луи, М.; Вирисель, А.; Лукас, Т.; Пельтье, Х.; Альфонси, Э.; Берроу, С.; Симон-Буэ, Б. (2014). «Структура популяции афалин, Tursiops truncatus , обусловленная средой обитания, в Северо-Восточной Атлантике». Молекулярная экология . 23 (4): 857–874. Бибкод : 2014MolEc..23..857L . дои : 10.1111/mec.12653 . hdl : 10261/92313 . ПМИД 24383934 . S2CID 22266293 .
- ^ Л. Тейлор, Б.; Бэрд, Р.; Барлоу, Дж.; М. Доусон, С.; Форд, Дж.; Дж. Мид, Дж.; Нотарбартоло ди Скиара, Г.; Уэйд, П.; Л. Питман, Р. (2011). « Глобицефала макроринхус » . Красный список исчезающих видов МСОП . 2011 . doi : 10.2305/IUCN.UK.2011-2.RLTS.T9249A12972356.en .
- ^ Амано М. (2012). «Дельфины в море Мичиноку (Специальный материал: Морские млекопитающие в море и аквариуме Мичиноку)» . Исана 56 : 60–65 . Проверено 9 марта 2017 г.
- ^ Хидака Т. Касуя Т. Идзава К. Кавамичи Т. 1996. Энциклопедия животных Японии (2) - Млекопитающие ISBN 9784582545524 (9784582545517) (4582545521) .
Внешние ссылки
[ редактировать ]Обзор эволюции китов см. Уэн, доктор медицины (2010). «Происхождение китов». Ежегодный обзор наук о Земле и планетах . 38 (1): 189–219. Бибкод : 2010AREPS..38..189U . doi : 10.1146/annurev-earth-040809-152453 .
- Хронология эволюции китов - Смитсоновский океанический портал
- Палеобиология китообразных - Бристольский университет
- BBC: Эволюция китов
- BBC: Эволюция китов – ископаемые свидетельства
- Зацепить Левиафана его прошлым Стивен Джей Гулд
- Исследование происхождения и ранней эволюции китов (Cetacea) , Джинджерич, доктор медицинских наук, Мичиганский университет.
- Pakicetus inachus, новый археоцитов (Mammalia, Cetacea) из ранне-среднеэоценовой формации Кулдана Кохата (Пакистан) . Джинджерич, доктор медицинских наук, 1981, Музей палеонтологии, Мичиганский университет.
- Скелеты наземных китообразных и связь китов с парнокопытными , Nature 413, 277–281 (20 сентября 2001 г.), JGM Thewissen , EM Williams, LJ Roe и ST Hussain.
- Сегмент «Эволюция китов» из выставки «Киты Киты» Мини-сайт Музея Новой Зеландии Те Папа Тонгарева
