Пространственная память

В психологии и нейронауке когнитивной пространственная память — это форма памяти, отвечающая за запись и восстановление информации, необходимой для планирования курса к определенному месту и вспоминания местоположения объекта или возникновения события. [1] Пространственная память необходима для ориентации в пространстве. [2] [3] Пространственную память также можно разделить на эгоцентрическую и аллоцентрическую пространственную память. [4] Пространственная память человека необходима для того, чтобы ориентироваться в знакомом городе. Пространственная память крысы необходима, чтобы запомнить расположение еды в конце лабиринта . И у людей, и у животных пространственная память обобщена в виде когнитивной карты . [5]
Пространственная память имеет представления в рабочей, кратковременной и долговременной памяти . Исследования показывают, что существуют определенные области мозга, связанные с пространственной памятью. [6] Для измерения пространственной памяти у детей, взрослых и животных используется множество методов. [5]
Кратковременная пространственная память
[ редактировать ]Кратковременную память (КТМ) можно описать как систему, позволяющую временно хранить и управлять информацией, необходимой для выполнения сложных когнитивных задач. [7] Задачи, в которых используется кратковременная память, включают обучение , рассуждение и понимание. [7] Пространственная память — это когнитивный процесс, который позволяет человеку запоминать различные местоположения, а также пространственные отношения между объектами. [7] Это позволяет запомнить, где находится объект по отношению к другому объекту; [7] например, позволяя кому-то перемещаться по знакомому городу. Говорят, что пространственные воспоминания формируются после того, как человек уже собрал и обработал сенсорную информацию об окружающей среде. [7]
Пространственная рабочая память
[ редактировать ]Рабочую память (WM) можно охарактеризовать как систему ограниченной емкости, позволяющую временно хранить и обрабатывать информацию. [8] Это временное хранилище позволяет выполнять сложные задачи или работать над ними, сохраняя при этом информацию. [8] Например, способность работать над сложной математической задачей использует рабочую память.
Одной из влиятельных теорий WM является Бэддели и Хитча многокомпонентная модель рабочей памяти . [8] [9] Самая последняя версия этой модели предполагает, что WM состоит из четырех подкомпонентов: фонологического цикла , зрительно-пространственного блокнота , центрального исполнительного органа и эпизодического буфера . [8] Один из компонентов этой модели, зрительно-пространственный блокнот, вероятно, отвечает за временное хранение, обслуживание и манипулирование как визуальной, так и пространственной информацией. [8] [9]

В отличие от многокомпонентной модели, некоторые исследователи считают, что СТМ следует рассматривать как единую конструкцию. [9] В этом отношении считается, что визуальная, пространственная и вербальная информация организована по уровням репрезентации, а не по типу хранилища, к которому они принадлежат. [9] В литературе предлагается изучить дальнейшие исследования фракционирования СТМ и WM. [9] [10] Однако большая часть исследований конструкции зрительно-пространственной памяти проводилась в соответствии с парадигмой, предложенной Бэддели и Хитчем. [8] [9] [10] [11] [12]
Роль центрального исполнительного органа
[ редактировать ]Исследования точной функции зрительно-пространственного блокнота показали, что как пространственная кратковременная память , так и рабочая память зависят от исполнительных ресурсов и не полностью различны. [8] Например, на производительность рабочей памяти, но не на задачу кратковременной памяти, повлияло подавление артикуляции , что позволяет предположить, что ухудшение пространственной задачи было вызвано параллельным выполнением задачи, в которой активно использовались исполнительные ресурсы. [8] Результаты также показали, что производительность была снижена при выполнении задач STM и WM с подавлением исполнительной власти. [8] Это иллюстрирует, как в визуально-пространственной области и STM, и WM требуют одинаковой полезности центрального исполнительного органа. [8]
Кроме того, во время задачи пространственной визуализации (которая связана с исполнительным функционированием, а не с STM или WM) одновременное подавление исполнительной власти ухудшило производительность, что указывает на то, что эффекты были вызваны общими требованиями к центральному исполнительному органу, а не кратковременным хранением. [8] В заключение исследователи объяснили, что центральный исполнительный орган использует когнитивные стратегии, позволяющие участникам как кодировать, так и поддерживать мысленные представления во время задач на кратковременную память. [8]
Хотя исследования показывают, что центральный исполнительный орган тесно вовлечен в ряд пространственных задач, точный способ их связи еще предстоит выяснить. [13]
Долговременная пространственная память
[ редактировать ]Пространственная память построена на иерархической структуре . Люди запоминают общую планировку конкретного пространства, а затем «указывают целевые местоположения» внутри этого пространственного набора. [14] Эта парадигма включает в себя порядковую шкалу характеристик, на которые человек должен обратить внимание, чтобы составить свою когнитивную карту. [15] Воспоминание пространственных деталей - это нисходящая процедура, которая требует от человека вспомнить вышестоящие функции когнитивной карты, за которыми следуют порядковые и подчиненные функции. При навигации по маршруту важны две пространственные характеристики: общая планировка и ориентация на ориентиры (Kahana et al., 2006). Люди не только способны изучать пространственную структуру своего окружения, но они также могут собирать воедино новые маршруты и новые пространственные отношения посредством умозаключений.
Когнитивная карта — это «мысленная модель пространственной конфигурации объектов, которая позволяет перемещаться по оптимальному пути между произвольными парами точек». [16] Эта мысленная карта построена на двух фундаментальных основах: планировке, также известной как знание маршрута, и ориентации ориентиров. Макет потенциально является первым методом навигации, который люди учатся использовать; его работа отражает наше самое базовое понимание мира.
Гермер и Спелке (1994) определили, что когда малыши начинают ходить, примерно в восемнадцать месяцев, они ориентируются на основе своего чувства устройства мира. Макнамара, Харди и Хиртл определили членство в регионе как главный строительный блок чьей-либо когнитивной карты (1989). В частности, членство в регионе определяется любыми границами, будь то физические, перцептивные или субъективные (McNamara et al., 1989). Границы являются одними из самых основных и эндемичных качеств окружающего нас мира. Эти границы представляют собой не что иное, как осевые линии, которые являются особенностью, к которой люди склонны относиться к пространству; например, одним из факторов, определяющих осевую линию, является сила тяжести (McNamara & Shelton, 2001; Kim & Penn, 2004). Осевые линии помогают каждому распределить наше восприятие по регионам. Эта идея разделенного мира дополнительно подтверждается тем фактом, что элементы, которые вспоминаются вместе, с большей вероятностью будут сгруппированы в одной и той же области более крупной когнитивной карты. [15] Кластеризация показывает, что люди склонны группировать информацию в соответствии с меньшими структурами внутри более крупной когнитивной карты.
Границы не являются единственным фактором, определяющим планировку. Кластеризация также демонстрирует еще одно важное свойство отношения к пространственным представлениям: пространственное воспоминание представляет собой иерархический процесс. Когда кто-то вспоминает окружающую среду или перемещается по местности, он сначала неявно вспоминает общую планировку. Затем, благодаря «богатой корреляционной структуре» концепта, активируется ряд ассоциаций. [14] В конце концов, результирующий каскад активаций пробудит конкретные детали, соответствующие вызываемой области. Именно так люди кодируют множество сущностей разных онтологических уровней, например расположение степлера; в столе; который находится в офисе.
Одновременно можно отозвать только один регион (узкое место). Проблемой может стать узкое место в когнитивной навигационной системе человека. Например, если во время длительной поездки возникла необходимость внезапного объезда. Недостаток опыта работы в данном месте или просто его размер могут дезориентировать мышление, особенно в большом и незнакомом месте со множеством подавляющих стимулов. В такой среде люди по-прежнему могут ориентироваться и ориентироваться по ориентирам. Эта способность «расставлять приоритеты для объектов и регионов в сложных сценах для выбора (и) распознавания» была описана Чуном и Цзяном в 1998 году. Ориентиры дают людям указания, активируя «выученные ассоциации между глобальным контекстом и целевыми местоположениями». [14] Маллот и Гиллнер (2000) показали, что испытуемые усвоили связь между конкретным ориентиром и направлением поворота, тем самым укрепив взаимосвязь между ассоциациями и ориентирами. [17] Шелтон и Макнамара (2001) кратко резюмировали, почему ориентиры как маркеры так полезны: «Местоположение... невозможно описать без ссылки на ориентацию наблюдателя».
Для навигации люди используют как планировку конкретного пространства, так и наличие ориентировочных ориентиров. Психологи еще не объяснили, влияет ли планировка на ориентиры или ориентиры определяют границы планировки. Из-за этого концепция страдает от парадокса курицы и яйца . Макнамара обнаружила, что испытуемые используют «группы ориентиров как внутренние системы отсчета», что только еще больше запутывает проблему. [16]
Люди воспринимают объекты в своей среде относительно других объектов в той же среде. Ориентиры и планировка являются взаимодополняющими системами пространственной памяти, но неизвестно, как эти две системы взаимодействуют, когда доступны оба типа информации. В результате людям приходится делать определенные предположения о взаимодействии двух систем. Например, когнитивные карты не являются «абсолютными», а скорее, как может подтвердить каждый, «используются для обеспечения значений по умолчанию... (которые) модулируются в соответствии с... требованиями задачи». [14] Психологи также считают, что когнитивные карты основаны на конкретных примерах, что объясняет «дискриминационное сопоставление с прошлым опытом». [14]
Этой области традиционно мешали смешанные переменные, такие как стоимость и возможность предыдущего воздействия экспериментальной среды. Технологические достижения, в том числе в области технологий виртуальной реальности, сделали выводы более доступными. Виртуальная реальность предоставляет экспериментаторам роскошь полного контроля над своей тестовой средой. Любой переменной можно манипулировать, включая вещи, которые в реальности были бы невозможны.
Виртуальная реальность
[ редактировать ]В ходе исследования 2006 года исследователи спроектировали три разных виртуальных города, каждый из которых имел свою «уникальную схему дорог и уникальный набор из пяти магазинов». [16] Однако общая площадь разных карт была одинаковой — 80 кв. В этом эксперименте участникам пришлось принять участие в двух разных сериях испытаний.
Исследование, проведенное в Университете Мэриленда, сравнило влияние различных уровней погружения на пространственную память. [18] В исследовании 40 участников использовали как традиционный рабочий стол, так и головной дисплей для просмотра двух сред: средневекового города и богато украшенного дворца, где они запоминали два набора из 21 лица, представленных в виде 3D-портретов. После просмотра этого 21 лица в течение 5 минут с последующим коротким периодом отдыха лица в виртуальных средах заменялись числами, и участники вспоминали, какое лицо было в каждом месте. Исследование показало, что в среднем те, кто использовал наголовный дисплей, запоминали лица на 8,8% точнее и с большей уверенностью. Участники заявляют, что ключом к их успеху было использование их врожденных вестибулярных и проприоцептивных чувств с помощью дисплея на голове и картографирования аспектов окружающей среды относительно их тела, элементов, которые отсутствуют на настольном компьютере.
Пространственная экспертиза
[ редактировать ]В литературе есть свидетельства того, что специалисты в определенной области способны выполнять задачи по запоминанию в соответствии со своими навыками на исключительном уровне. [12] Уровень навыков, демонстрируемый экспертами, может превышать пределы обычных возможностей как STM, так и WM. [12] Поскольку эксперты обладают огромным объемом предварительно изученных и конкретных знаний, они могут быть способны кодировать информацию более эффективным способом. [12]
Интересное исследование по изучению памяти таксистов об улицах Хельсинки ( Финляндия ) изучило роль заранее приобретенных пространственных знаний. [12] В этом исследовании эксперты сравнивались с контрольной группой, чтобы определить, как эти предварительно полученные знания в их области навыков позволяют им преодолеть ограничения возможностей STM и WM. [12] В исследовании использовались четыре уровня пространственной случайности:
- Порядок маршрута – пространственно непрерывный маршрут [12]
- Route Random – пространственно непрерывный список, представленный случайным образом. [12]
- Порядок карты - названия улиц образуют на карте прямую линию, но без промежуточных улиц. [12]
- Map Random – улицы на карте представлены в случайном порядке. [12]

Результаты этого исследования показывают, что запоминание улиц таксистами (экспертами) было выше как в режиме порядка маршрута, так и в режиме порядка карты, чем в двух случайных условиях. [12] Это указывает на то, что эксперты смогли использовать свои заранее полученные пространственные знания для организации информации таким образом, что они превзошли ограничения возможностей STM и WM. [12] Стратегия организации, которую использовали водители, известна как фрагментирование . [12] Кроме того, комментарии, сделанные экспертами в ходе процедуры, указывают на использование ими знаний маршрута при выполнении задания. [12] Чтобы убедиться, что они действительно кодируют пространственную информацию, исследователи также представили списки в алфавитном порядке и семантических категориях. [12] Однако исследователи обнаружили, что на самом деле эксперты группировали именно пространственную информацию, что позволило им преодолеть ограничения как визуально-пространственной STM, так и WM. [12]
Исследования на животных
[ редактировать ]Некоторые виды париевых и врановых (например, черношапочная синица и кустарниковая сойка ) способны использовать пространственную память, чтобы запоминать, где, когда и какой тип пищи они спрятали. [19] Исследования на крысах и белках также показали, что они способны использовать пространственную память, чтобы находить ранее спрятанную еду. [19] Эксперименты с использованием радиального лабиринта позволили исследователям контролировать ряд переменных, таких как тип спрятанной еды, места, где она спрятана, интервал хранения, а также любые запахи, которые могут исказить результаты исследования памяти. [19] Исследования показали, что крысы помнят, где и какой тип еды они спрятали. [19] Это проявляется в поисковом поведении: крысы избирательно чаще идут к рукавам лабиринта, где они ранее спрятали предпочитаемую пищу, чем к рукавам с менее предпочтительной пищей или где еда не была спрятана. [19]
Данные о пространственной памяти некоторых видов животных, таких как крысы, указывают на то, что они действительно используют пространственную память для обнаружения и извлечения скрытых запасов пищи. [19]
Исследование с использованием GPS-слежения, чтобы увидеть, куда идут домашние кошки , когда хозяева выпускают их на улицу, показало, что кошки обладают хорошей пространственной памятью. Некоторые кошки, участвовавшие в исследовании, продемонстрировали исключительную долговременную пространственную память. Один из них, обычно путешествующий не дальше, чем от 200 м (660 футов) до 250 м (820 футов) от своего дома, неожиданно преодолел расстояние примерно в 1250 м (4100 футов) от своего дома. Исследователи сначала подумали, что это неисправность GPS, но вскоре обнаружили, что владельцы кошки уехали из города в те выходные, и что дом, в который ходил кот, был старым домом владельца. Хозяева и кот не жили в этом доме уже больше года. [20]
Визуально-пространственное различие
[ редактировать ]Логи (1995) предположил, что зрительно-пространственный блокнот разбит на два подкомпонента: визуальный и пространственный. [11] Это визуальный кэш и внутренний писец соответственно. [11] Визуальный кэш — это временное хранилище визуальных данных, включающее такие измерения, как цвет и форма. [11] И наоборот, внутренний писец представляет собой механизм репетиции визуальной информации и отвечает за информацию, касающуюся последовательностей движений. [11] Хотя в литературе отмечается общее отсутствие консенсуса относительно этого различия, [10] [21] [22] появляется все больше свидетельств того, что эти два компонента являются отдельными и выполняют разные функции. [ нужна ссылка ]
Зрительная память отвечает за сохранение визуальных форм и цветов (т. е. чего), тогда как пространственная память отвечает за информацию о местах и движении (т. е. где). Это различие не всегда однозначно, поскольку часть зрительной памяти включает в себя пространственную информацию и наоборот. Например, память о формах объектов обычно предполагает сохранение информации о пространственном расположении характеристик, определяющих рассматриваемый объект. [21]
На практике эти две системы в некоторой степени работают вместе, но были разработаны разные задачи, чтобы подчеркнуть уникальные способности, связанные как со зрительной, так и с пространственной памятью. Например, тест визуальных паттернов (VPT) измеряет зрительный охват, тогда как задача Corsi Blocks измеряет пространственный охват. Корреляционные исследования этих двух показателей предполагают разделение зрительных и пространственных способностей из-за отсутствия корреляции, обнаруженной между ними как у здоровых пациентов, так и у пациентов с повреждением головного мозга . [10]
Поддержка разделения компонентов зрительной и пространственной памяти обнаруживается посредством экспериментов с использованием парадигмы двойной задачи . Ряд исследований показал, что сохранение визуальных форм или цветов (т. е. визуальной информации) нарушается при предъявлении не относящихся к делу изображений или динамического визуального шума. И наоборот, сохранение местоположения (т. е. пространственной информации) нарушается только задачами пространственного отслеживания, задачами пространственного постукивания и движениями глаз. [21] [22] Например, участники выполнили как задачу VPT, так и задачу блоков Корси в эксперименте с выборочным вмешательством. В течение периода сохранения ВПТ субъект просматривал нерелевантные изображения (например, авангардные картины). Задача пространственной интерференции требовала, чтобы участники, прикасаясь к стимулам, следили за набором небольших деревянных колышков, спрятанных за ширмой. И зрительный, и пространственный диапазоны были сокращены из-за соответствующих задач интерференции, подтверждая, что задача блоков Корси относится в первую очередь к пространственной рабочей памяти. [10]
Измерение
[ редактировать ]Психологи используют множество задач для измерения пространственной памяти на моделях взрослых, детей и животных. Эти задачи позволяют профессионалам выявлять когнитивные нарушения у взрослых и детей, а также позволяют исследователям назначать участникам различные типы лекарств и/или поражений и измерять последующее воздействие на пространственную память.
Задача на постукивание блока Корси
[ редактировать ]Тест на постукивание по блоку Корси, также известный как отдых Корси, представляет собой психологический тест, обычно используемый для определения объема зрительно-пространственной памяти и неявных зрительно-пространственных способностей человека к обучению. [23] [24] Участники сидят с девятью деревянными кубиками размером 3х3 см, закрепленными перед ними на плинтусе размером 25 х 30 см в стандартном случайном порядке. В ходе эксперимента на блоках используется шаблон последовательности, который участники затем должны воспроизвести. Блоки пронумерованы на стороне экспериментаторов, чтобы обеспечить эффективную демонстрацию закономерностей. Длина последовательности увеличивается с каждым испытанием до тех пор, пока участник не перестанет правильно воспроизводить образец. Тест можно использовать для измерения как кратковременной, так и долговременной пространственной памяти, в зависимости от продолжительности времени между тестом и воспроизведением.
Тест был создан канадским нейропсихологом Филипом Корси, который смоделировал его по образцу Хебба задачи , заменив числовые элементы теста пространственными. В среднем большинство участников справляются с пятью заданиями в тесте Корси и семью с заданием на диапазон цифр.
Диапазон визуальных шаблонов
[ редактировать ]Анализ зрительных паттернов аналогичен тесту постукивания по блоку Корси, но считается более чистым тестом на кратковременное визуальное запоминание. [25] Участникам предлагается серия матричных шаблонов, половина ячеек которых окрашена, а другая половина пуста. Матричные шаблоны расположены таким образом, что их трудно закодировать вербально, что вынуждает участника полагаться на зрительную пространственную память. Начиная с небольшой матрицы 2х2, участники копируют шаблон матрицы из памяти в пустую матрицу. Матричные шаблоны увеличиваются в размере и сложности со скоростью две ячейки, пока способность участника воспроизводить их не нарушается. В среднем результативность участников падает на шестнадцати ячейках.
Задача промежутка пути
[ редактировать ]Это задание предназначено для измерения способностей пространственной памяти у детей. [23] Экспериментатор просит участника визуализировать пустую матрицу с маленьким человечком. Посредством серии инструкций по направлению, таких как вперед, назад, влево или вправо, экспериментатор направляет маленького человечка участника по пути через матрицу. В конце участника просят указать на реальной матрице, где закончился представленный им человечек. Длина пути варьируется в зависимости от уровня сложности (1-10), а сами матрицы могут иметь длину от 2 х 2 ячеек до 6 х 6.
Динамические лабиринты
[ редактировать ]Динамические лабиринты предназначены для измерения пространственных способностей детей. В этом тесте экспериментатор предъявляет участнику рисунок лабиринта с изображением человека в центре. [23] Пока участник наблюдает, экспериментатор пальцем прослеживает путь от входа в лабиринт до рисунка человека. Затем ожидается, что участник повторит продемонстрированный путь через лабиринт до рисунка человека. Лабиринты различаются по сложности по мере увеличения сложности.
Радиальный лабиринт
[ редактировать ]
Впервые предложенный Олтоном и Самуэльсоном в 1976 году. [26] Лабиринт с радиальными рукавами предназначен для проверки способности крыс к пространственной памяти. Лабиринты обычно имеют центральную платформу и различное количество рычагов. [27] разветвляющиеся с пищей, расположенной на концах. Руки обычно каким-то образом защищены друг от друга, но не до такой степени, чтобы внешние сигналы не могли использоваться в качестве ориентиров.
В большинстве случаев крысу помещают в центр лабиринта, и ей необходимо исследовать каждый рукав по отдельности, чтобы найти еду, одновременно запоминая, по каким рукам она уже преследовала. Лабиринт устроен так, что крыса вынуждена вернуться в центр лабиринта, прежде чем преследовать другую руку. Обычно принимаются меры, чтобы крыса не могла использовать свои обонятельные чувства для навигации, например, размещают дополнительную еду на дне лабиринта.
Задача Морриса по водной навигации
[ редактировать ]Задача Морриса по водной навигации — классический тест для изучения пространственного обучения и памяти у крыс. [28] и был впервые разработан в 1981 году Ричардом Г. Моррисом, в честь которого назван тест. Субъект помещается в круглый резервуар с прозрачной водой, стенки которого слишком высоки, чтобы он мог выбраться, а вода слишком глубока, чтобы он мог в нем стоять. Стены резервуара украшены визуальными подсказками, служащими ориентирами. Крыса должна плавать вокруг бассейна, пока случайно не обнаружит чуть ниже поверхности скрытую платформу, на которую она может забраться.
Как правило, крысы сначала плавают вокруг края бассейна, а затем выходят в центр извилистым узором, а затем натыкаются на скрытую платформу. Однако по мере того, как время, проведенное в бассейне, увеличивает опыт, количество времени, необходимое для поиска платформы, уменьшается: крысы-ветераны плывут прямо к платформе почти сразу после помещения в воду. Из-за характера задачи, связанной с плаванием крыс, большинство исследователей полагают, что привыкание необходимо для снижения уровня стресса у животного. Стресс животного может ухудшить результаты когнитивного тестирования. [29]
Физиология
[ редактировать ]Гиппокамп
[ редактировать ]
Гиппокамп . предоставляет животным пространственную карту окружающей их среды [30] Он хранит информацию о неэгоцентрическом пространстве (эгоцентрический означает положение тела в пространстве) и, следовательно, поддерживает независимость точки зрения в пространственной памяти. [31] Это означает, что он позволяет манипулировать точкой зрения по памяти. Это важно для долговременной пространственной памяти аллоцентрического пространства (ссылка на внешние сигналы в пространстве). [32] Таким образом, поддержание и извлечение воспоминаний зависят от отношений или контекста . [33] Гиппокамп использует справочную и рабочую память и играет важную роль в обработке информации о пространственных местоположениях. [34]
Блокирование пластичности в этой области приводит к проблемам с целенаправленной навигацией и ухудшает способность запоминать точные местоположения. [35] Пациенты с амнезией и повреждением гиппокампа не могут учиться или запоминать пространственные расположения, а у пациентов, перенесших удаление гиппокампа, серьезно нарушена пространственная навигация. [31] [36]
Обезьяны с поражениями этой области не могут научиться ассоциациям объект-место, а крысы также демонстрируют пространственный дефицит, не реагируя на пространственные изменения. [31] [37] Кроме того, было показано, что у крыс с поражениями гиппокампа наблюдается недифференцированная во времени (независимая от времени) ретроградная амнезия , которая устойчива к распознаванию выученной задачи на платформе только тогда, когда поражен весь гиппокамп, но не когда он поврежден частично. [38] Дефицит пространственной памяти также обнаруживается при выполнении задач на пространственную дискриминацию. [36]

Большие различия в пространственных нарушениях обнаружены в дорсальном и вентральном гиппокампе. Поражения вентрального гиппокампа не влияют на пространственную память, тогда как дорсальный гиппокамп необходим для извлечения информации, обработки кратковременной памяти и перевода памяти с краткосрочной на более длительную задержку. [39] [40] [41] введение амфетамина в дорсальный гиппокамп улучшает память на ранее изученные пространственные положения. Также было показано, что [42] Эти данные указывают на то, что существует функциональная диссоциация между дорсальным и вентральным гиппокампом.
Наблюдаются также полушарные различия внутри гиппокампа. В ходе исследования лондонских таксистов водителей просили вспомнить сложные маршруты по городу, а также известные достопримечательности , местонахождение которых водители не знали. Это привело к активации правого гиппокампа исключительно во время воспроизведения сложных маршрутов, что указывает на то, что правый гиппокамп используется для навигации в крупномасштабных пространственных средах. [43]
Известно, что гиппокамп содержит два отдельных контура памяти. Одна схема используется для памяти распознавания мест на основе воспоминаний и включает в себя систему энторинал-СА1 . [44] в то время как другая система, состоящая из трисинаптической петли гиппокампа (энтохинал-дентат-CA3-CA1), используется для запоминания места. [45] и облегчение пластичности энторинально-зубчатого синапса у мышей достаточно для улучшения запоминания мест. [46]
Клетки места также обнаружены в гиппокампе.
Задняя теменная кора
[ редактировать ]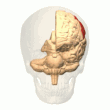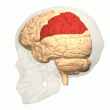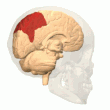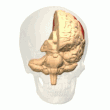
Теменная кора кодирует пространственную информацию, используя эгоцентрическую систему отсчета. Таким образом, он участвует в преобразовании координат сенсорной информации в координаты действия или эффекторные координаты путем обновления пространственного представления тела в окружающей среде. [47] В результате поражения теменной коры вызывают дефицит в освоении и сохранении эгоцентрических задач, тогда как незначительные нарушения наблюдаются среди аллоцентрических задач. [48]
Крысы с поражением передней области задней теменной коры повторно исследуют смещенные объекты, в то время как крысы с поражением задней области задней теменной коры не проявляют реакции на пространственные изменения. [37]
Известно также, что поражения теменной коры вызывают недифференцированную по времени ретроградную амнезию . [49]
Энторинальная кора
[ редактировать ]
Дорсально-каудальная медиальная энторинальная кора (dMEC) содержит топографически организованную карту пространственной среды, состоящую из ячеек сетки . [50] Таким образом, эта область мозга преобразует сенсорную информацию из окружающей среды и сохраняет ее как устойчивое аллоцентрическое представление в мозге, которое можно использовать для интеграции путей . [51]
Энторинальная кора способствует обработке и интеграции геометрических свойств и информации в окружающей среде. [52] Поражения этой области ухудшают использование дистальных , но не проксимальных ориентиров во время навигации и вызывают зависимый от задержки дефицит пространственной памяти, пропорциональный продолжительности задержки. [53] [54] Также известно, что поражения этой области приводят к дефициту запоминания задач, изученных за 4 недели, но не за 6 недель до поражения. [49]
Консолидация памяти в энторинальной коре достигается за счет киназной активности, регулируемой внеклеточными сигналами. [55]
Префронтальная кора
[ редактировать ]
Медиальная префронтальная кора обрабатывает эгоцентрическую пространственную информацию. Он участвует в обработке кратковременной пространственной памяти, используемой для управления запланированным поисковым поведением, и, как полагают, объединяет пространственную информацию с ее мотивационным значением. [41] [56] Идентификация нейронов, ожидающих ожидаемого вознаграждения в пространственной задаче, подтверждает эту гипотезу. Медиальная префронтальная кора также участвует во временной организации информации. [57]
Специализация полушарий находится в этой области мозга. Левая префронтальная кора преимущественно обрабатывает категориальную пространственную память, включая память источника (ссылка на пространственные отношения между местом или событием), тогда как правая префронтальная кора преимущественно обрабатывает координатную пространственную память, включая память предметов (ссылка на пространственные отношения между особенностями предмета). [58]
Поражения медиальной префронтальной коры ухудшают производительность крыс в ранее обученном лабиринте с радиальными рукавами, но крысы могут постепенно улучшаться до уровня контрольной группы в зависимости от опыта. [59] Поражения этой области также вызывают дефицит выполнения задач с задержкой, не связанных с сопоставлением позиций, и нарушения в освоении задач на пространственную память во время обучающих испытаний. [60] [61]
Ретросплениальная кора
[ редактировать ]Ретросплениальная кора участвует в обработке аллоцентрической памяти и геометрических свойств окружающей среды. [52] Инактивация этой области приводит к нарушению навигации в темноте и может быть вовлечена в процесс интеграции пути . [62]
Поражения ретросплениальной коры последовательно ухудшают тесты аллоцентрической памяти, сохраняя при этом эгоцентрическую память. [63] Животные с поражением каудальной ретросплениальной коры головного мозга демонстрируют ухудшение производительности в лабиринте с радиальными рукавами только тогда, когда лабиринт вращается, чтобы избавиться от зависимости от сигналов внутри лабиринта. [64]

У людей повреждение ретросплениальной коры приводит к топографической дезориентации. В большинстве случаев повреждается правая ретросплениальная кора, в том числе зона Бродмана 30. Пациенты часто испытывают затруднения в освоении новых маршрутов и навигации в знакомой обстановке. [65] Однако большинство пациентов обычно выздоравливают в течение 8 недель.
Ретросплениальная кора преимущественно обрабатывает пространственную информацию в правом полушарии. [65]
Периринальная кора
[ редактировать ]Периринальная кора связана как с пространственной ориентировкой, так и с пространственной рабочей памятью. [34] Он обрабатывает реляционную информацию об окружающих сигналах и местах.
Поражения периринальной коры вызывают дефицит опорной и рабочей памяти, а также увеличивают скорость забывания информации во время обучающих испытаний водного лабиринта Морриса. [66] Это объясняет ухудшение первоначального выполнения задачи. Поражения также ухудшают задачу определения местоположения объекта и снижают привыкание к новой среде. [34]
Нейропластичность
[ редактировать ]Пространственные воспоминания формируются после того, как животное собирает и обрабатывает сенсорную информацию об окружающем мире (особенно зрение и проприоцепцию ). В целом млекопитающим необходим функционирующий гиппокамп (особенно область CA1), чтобы формировать и обрабатывать воспоминания о космосе. Есть некоторые свидетельства того, что пространственная память человека прочно связана с правым полушарием мозга. [67] [68] [69]
Для пространственного обучения требуются как NMDA , так и AMPA- рецепторы, для консолидации требуются NMDA-рецепторы, а для извлечения пространственных воспоминаний требуются AMPA-рецепторы. [70] Было показано, что у грызунов пространственная память зависит от размера части проекции мшистых волокон гиппокампа . [71]
Функция NMDA-рецепторов варьируется в зависимости от субрегиона гиппокампа. Рецепторы NMDA необходимы в СА3 гиппокампа, когда необходимо реорганизовать пространственную информацию, тогда как рецепторы NMDA в СА1 необходимы для приобретения и восстановления памяти после задержки, а также для формирования полей места СА1. [72] Блокада рецепторов NMDA предотвращает индукцию долгосрочной потенциации и ухудшает пространственное обучение. [73]
CA3 гиппокампа играет особенно важную роль в кодировании и восстановлении пространственной памяти. CA3 иннервируется двумя афферентными путями, известными как перфорантный путь (PPCA3) и зубчатой извилиной (DG), опосредованными мшистыми волокнами (MF). Первый путь рассматривается как путь индекса поиска, а второй связан с кодированием. [74]
Расстройства/дефицит
[ редактировать ]Топографическая дезориентация
[ редактировать ]Топографическая дезориентация (ТД) — когнитивное расстройство, в результате которого человек не может ориентироваться в реальной или виртуальной среде. Пациенты также испытывают трудности с задачами, зависящими от пространственной информации. Эти проблемы могут быть результатом нарушения способности доступа к своей когнитивной карте, мысленного представления окружающей среды или неспособности судить о местонахождении объектов по отношению к самому себе. [75]
Топографическую дезориентацию развития (ТДД) диагностируют, когда пациенты с рождения демонстрируют неспособность ориентироваться даже в знакомой обстановке и не обнаруживают очевидных неврологических причин этого дефицита, таких как повреждения или повреждение головного мозга. DTD является относительно новым расстройством и может возникать в различной степени тяжести.
Было проведено исследование, чтобы выяснить, влияет ли топографическая дезориентация на людей с легкими когнитивными нарушениями (MCI). В исследование был включен 41 пациент с диагнозом MCI и 24 здоровых человека из контрольной группы. Стандарты, которые были установлены для этого эксперимента, были:
- Субъективные когнитивные жалобы со стороны пациента или лица, осуществляющего уход за ним.
- Нормальная общая когнитивная функция выше 16-го процентиля по корейской версии мини-психического государственного экзамена (K-MMSE).
- Нормальную повседневную активность (ADL) оценивали как клинически, так и по стандартизированной шкале (как описано ниже).
- Объективное снижение когнитивных функций ниже 16-го процентиля по нейропсихологическим тестам.
- Исключение деменции.
TD оценивалась клинически у всех участников. Неврологические и нейропсихологические оценки определялись с помощью магнитного сканирования, которое проводилось каждому участнику. Морфометрия на основе вокселей использовалась для сравнения паттернов атрофии серого вещества у пациентов с TD и без него, а также у группы нормальных людей из контрольной группы. В результате эксперимента ТД обнаружили у 17 из 41 пациента с МРП (41,4%). Функциональные способности были значительно нарушены у пациентов с MCI с TD по сравнению с пациентами с MCI без TD, и что наличие TD у пациентов с MCI связано с потерей серого вещества в медиальных височных областях, включая гиппокамп. [76]
Повреждение гиппокампа и шизофрения
[ редактировать ]повреждение гиппокампа может отрицательно влиять на пространственную память Исследования на крысах показывают, что неонатальное , что очень похоже на шизофрению . Считается, что шизофрения возникает из-за проблем нервного развития вскоре после рождения. [77]
Крыс обычно используют в качестве моделей пациентов с шизофренией. Экспериментаторы создают поражения в вентральной области гиппокампа вскоре после рождения — процедура, известная как неонатальное вентральное поражение гиппокампа (NVHL). У взрослых крыс с НВХЛ наблюдаются типичные признаки шизофрении, такие как гиперчувствительность к психостимуляторам , снижение социального взаимодействия и нарушение преимпульсного торможения , рабочей памяти и переключения установок. [78] [79] [80] [81] [82] Подобно шизофрении, крысы с нарушениями не могут использовать контекст окружающей среды в задачах пространственного обучения, таких как трудности с прохождением радиального лабиринта и водного лабиринта Мориса. [83] [84] [85]
НИЛ1
[ редактировать ]Эндонуклеаза VIII-подобная 1 ( NEIL1 ) представляет собой фермент репарации ДНК , который широко экспрессируется по всему мозгу . NEIL1 представляет собой ДНК-гликозилазу , которая инициирует первый этап репарации вырезаемых оснований , расщепляя основания, поврежденные активными формами кислорода, а затем вызывая разрыв цепи ДНК посредством связанной лиазной реакции. Этот фермент распознает и удаляет окисленные основания ДНК , включая формамидопиримидин , тимингликоль , 5-гидроксиурацил и 5-гидроксицитозин . NEIL1 способствует сохранению кратковременной пространственной памяти. [86] У мышей, лишенных NEIL1, нарушена кратковременная пространственная память в тесте на водный лабиринт. [86]

Технология глобальной системы позиционирования (GPS) произвела революцию в том, как мы ориентируемся и исследуем окружающую среду. GPS стал важным инструментом в нашей повседневной жизни, предоставляя в режиме реального времени информацию о нашем местоположении и направлениях, по которым нам нужно двигаться, чтобы добраться до пункта назначения. Однако некоторые исследователи выразили обеспокоенность по поводу влияния использования GPS на наше пространственное обучение и память. Пространственное обучение относится к нашей способности воспринимать, запоминать и использовать пространственную информацию, полученную в окружающей среде. Память , с другой стороны, включает в себя нашу способность хранить и извлекать информацию об окружающем нас мире. И пространственное обучение, и память имеют решающее значение для нашей способности эффективно ориентироваться и исследовать окружающую среду.Было показано, что использование GPS оказывает как положительное, так и отрицательное влияние на пространственное обучение и память. Исследования показали, что люди, которые полагаются на GPS для навигации, с меньшей вероятностью разрабатывают и используют мысленные карты и им труднее запоминать детали об окружающей среде, поскольку использование GPS может со временем привести к ухудшению этих навыков. [87] Более того, пользователи GPS склонны больше полагаться на технологию, чем на свои собственные когнитивные способности , что приводит к потере уверенности в своих навигационных навыках. [88]
Однако этой потере уверенности в своих силах противодействует осознание того, что заблудиться больше не проблема, благодаря GPS на наших телефонах, что, в свою очередь, восстанавливает нашу уверенность в своих способностях ориентироваться . Некоторые положительные результаты, связанные с помощью GPS, — это более эффективная и точная навигация в сочетании со значительным снижением когнитивной нагрузки , необходимой для навигации. Когда люди используют устройства GPS, им не нужно беспокоиться о запоминании маршрута, обращании внимания на ориентиры или постоянной проверке карт . Это может освободить когнитивные ресурсы для других задач, что приведет к повышению производительности при выполнении таких задач и более высокому уровню концентрации и сосредоточенности. Это позволяет высвободить когнитивные ресурсы для облегчения обработки информации и обучения. [89]
Чтобы компенсировать проблемы, возникающие при использовании GPS, были проведены обширные исследования, предлагающие альтернативные формы GPS-навигации или дополнения к существующим, которые, как было показано, улучшают пространственное обучение. В исследовании 2021 года была реализована трехмерная пространственная аудиосистема, похожая на слуховой компас, в которой пользователи направляются к месту назначения без явных указаний. Вместо того, чтобы пассивно вестись посредством словесных указаний, пользователям предлагается играть активную роль в собственной пространственной навигации. Это привело к созданию более точных когнитивных карт пространства, улучшение, которое было продемонстрировано, когда участники исследования нарисовали точные карты после выполнения задачи по поиску мусора . [90] Другое исследование предложило выделить местные особенности, такие как достопримечательности , вдоль маршрута и в точках принятия решения ; или выделение структурных особенностей, которые обеспечивают глобальную ориентацию (не детали маршрута, пройденного участниками исследования, а ориентиры на большей территории, окружающей его). Исследование показало, что выделение местных особенностей на навигационных картах (GPS) способствует приобретению знаний о маршруте, которые измерялись с помощью наведения и задачи по запоминанию глобальных объектов. [91]
Кроме того, у слепых и слабовидящих людей использование GPS дает преимущества в пространственном обучении и памяти. Слепым и слабовидящим людям часто необходимо заранее получить информацию о местах и потренироваться на определенном маршруте с помощью родственника, друга или специализированного инструктора, прежде чем самостоятельно путешествовать по маршруту к указанному пункту назначения. GPS предлагает полезную информацию, что позволяет им стать более независимыми и уверенными в путешествии к определенному пункту назначения. [92]
В другой исследовательской работе утверждается, что GPS можно использовать для пациентов, страдающих деменцией . В исследовании, проведенном в 2014 году, водителям с болезнью Альцгеймера (БА) от легкой до очень легкой степени было проведено 3 испытания вождения с различными настройками GPS (нормальные, только визуальные и только аудио). Участникам необходимо было выполнить различные задачи по вождению на симуляторе вождения, следуя инструкциям GPS. Это исследование показало, что использование одиночных простых слуховых инструкций при отсутствии визуального сигнала GPS потенциально может помочь людям с легкой формой AD улучшить свои навыки вождения и добраться до места назначения, тем самым подтверждая, что использование GPS действительно снижает когнитивную нагрузку. [93]
Поскольку использование GPS поможет пациентам сориентироваться, это позволит им оставаться в безопасности в общественных местах, вернуть себе чувство самодостаточности и препятствовать «бродяжничеству». В целом, наиболее убедительны доказательства использования технологий GPS для предотвращения вреда и улучшения благополучия . [94]
Влияние использования GPS на пространственное обучение и память еще не до конца изучено, и необходимы дальнейшие исследования для изучения долгосрочного воздействия использования GPS на эти когнитивные процессы. Однако очевидно, что технология GPS имеет как преимущества, так и недостатки, и пользователи должны осознавать потенциальное влияние их зависимости от GPS.В заключение отметим, что технология GPS произвела революцию в том, как мы ориентируемся и исследуем окружающую среду, но ее влияние на наше пространственное обучение и память до сих пор остается предметом споров. Хотя использование GPS может помочь людям более эффективно и уверенно ориентироваться, а также помочь группам населения, которые в противном случае столкнулись бы со значительными трудностями; его использование может со временем привести к снижению пространственных когнитивных навыков. Поэтому пользователям важно сбалансировать преимущества и недостатки использования GPS и осознавать его потенциальное влияние на их когнитивные способности.
Трудности в обучении
[ редактировать ]Невербальная неспособность к обучению (NVLD) характеризуется нормальными вербальными способностями, но нарушенными зрительно-пространственными способностями. Проблемные области для детей с невербальными нарушениями обучаемости включают арифметику, геометрию и естествознание. Нарушения пространственной памяти связаны с расстройствами невербального обучения и другими трудностями в обучении. [95]
Арифметические задачи со словами включают в себя письменный текст, содержащий набор данных, за которым следует один или несколько вопросов, и требуют использования четырех основных арифметических операций (сложение, вычитание, умножение или деление). [22] Исследователи предполагают, что успешное решение арифметических задач со словами задействует пространственную рабочую память (участвующую в построении схематических представлений), которая облегчает создание пространственных отношений между объектами. Создание пространственных отношений между объектами является важной частью решения текстовых задач, поскольку требуются мыслительные операции и преобразования. [22]
Исследователи исследовали роль пространственной и зрительной памяти в способности решать арифметические задачи со словами. Дети, участвовавшие в исследовании, выполнили задание на блок Корси (прямые и обратные серии) и задание на пространственную матрицу, а также задание на зрительную память, называемое тестом на узнавание дома. Плохо решавшие проблемы были хуже справлялись с задачами блока Корси и задачей пространственной матрицы, но нормально справлялись с тестом на узнавание дома по сравнению с детьми с нормальной успеваемостью. Эксперимент показал, что плохое решение задач связано именно с недостаточной обработкой пространственной информации. [22]
Спать
[ редактировать ]сон Было обнаружено, что благотворно влияет на пространственную память, усиливая консолидацию памяти , зависящую от гиппокампа . [96] повышение уровня различных путей, которые отвечают за силу синапсов, контролируют транскрипцию генов, связанных с пластичностью, и трансляцию белков (Доминик Пибер, 2021). [97] Области гиппокампа, активированные при обучении маршруту, повторно активируются во время последующего сна ( медленного сна в частности, ). Одно исследование показало, что фактическая степень реактивации во время сна коррелирует с улучшением воспроизведения маршрута и, следовательно, с улучшением памяти на следующий день. [98] Исследование установило идею о том, что сон усиливает процесс консолидации на системном уровне, что, следовательно, улучшает поведенческие показатели. Период бодрствования не влияет на стабилизацию следов памяти по сравнению с периодом сна. Сон после первой ночи после тренировки, т. е. на вторую ночь, не способствует дальнейшей консолидации пространственной памяти. Поэтому очень важно спать в первую ночь после тренировки, например, после изучения маршрута. [96]
Кроме того, было показано, что ранний и поздний ночной сон по-разному влияет на пространственную память. Предполагается, что N3 медленного сна, также называемый медленным сном (SWS), играет важную роль в зависимом от сна создании пространственной памяти у людей. В частности, в исследовании, проведенном Плихалом и Борном (1999), [99] Производительность при выполнении заданий на умственное вращение была выше среди участников, у которых были ранние интервалы сна (23:00–02:00) после изучения задания, по сравнению с теми, у кого были поздние интервалы сна (03:00–06:00). Эти результаты позволяют предположить, что ранний сон, богатый SWS, имеет определенные преимущества для формирования пространственной памяти. Когда исследователи исследовали, окажет ли ранний сон такое влияние на задание на закрепление основы слова (вербальное задание), результаты оказались противоположными. Это не стало сюрпризом для исследователей, поскольку задачи прайминга в основном полагаются на процедурную память , и, таким образом, она приносит больше пользы позднему удерживающему сну (где преобладает быстрый сон ), чем раннему. [99]
Лишение сна и сон также были исследованной связью. Депривация сна препятствует улучшению производительности памяти из-за активного нарушения консолидации пространственной памяти. [96] В результате пространственная память усиливается в период сна. Аналогичные результаты были подтверждены другим исследованием, посвященным влиянию полной депривации сна (TSD) на пространственную память крыс (Guan et al., 2004). [100] В первом проведенном эксперименте крыс обучали в водном лабиринте Морриса 12 попыткам в течение 6 часов, чтобы найти скрытую платформу (прозрачную и не видимую в воде), используя пространственные сигналы в окружающей среде. В каждом испытании они начинали с разных точек, и им разрешалось плыть максимум 120 секунд, чтобы достичь платформы. После фазы обучения они провели пробное испытание для проверки пространственной памяти (через 24 часа). В этом испытании скрытая платформа была удалена из лабиринта, а время, проведенное животными в целевой области (которая раньше была занята скрытой платформой), было мерой устойчивости пространственной памяти. Контрольные крысы, у которых был спонтанный сон, провели значительно больше времени в целевом квадранте по сравнению с крысами, у которых было полное лишение сна. С точки зрения пространственного обучения, о котором свидетельствует задержка поиска скрытой платформы, различий не было. Как для контрольных, так и для лишенных сна крыс время, необходимое для поиска платформы, уменьшалось с каждым новым испытанием. [100]
Во втором эксперименте крыс обучали плыть к видимой платформе, местоположение которой менялось в каждом испытании. Каждое новое испытание крысы начинали с противоположной стороны платформы. После обучения в одном испытании их память была проверена через 24 часа. Платформа все еще находилась в лабиринте. Расстояние и время, необходимое им, чтобы доплыть до видимой платформы, рассматривались как непространственные меры памяти. Никакой существенной разницы не было обнаружено между лишенными сна крысами и контрольными крысами. Аналогичным образом, с точки зрения пространственного обучения, о котором свидетельствует задержка достижения видимой платформы, не было существенных различий. ТСД не влияет на непространственное обучение и непространственную память. [100]
Что касается влияния лишения сна на человека, Доминик Пибер (2021 г.) [97] В своем обзоре литературы он представил клинические наблюдения, которые показывают, что люди с тяжелыми нарушениями сна часто имеют нарушения пространственной памяти. Как видно из исследований обоих, у пациентов с бессонницей, страдающих расстройством сна, характеризующимся прерывистым, невосстанавливающим сном и дефицитом когнитивных функций в течение дня, зафиксировано отрицательное выполнение пространственных задач по сравнению со здоровыми людьми. участники (Ли и др., 2016; [101] Чен и др., 2016; [102] Хассауна и др., 2018 г.; [103] Он и др., 2021 г. [104] ).
Точно так же сновидения играют важную роль в пространственной памяти. Исследование, проведенное Вамсли и Стикголдом (2019). [105] доказали, что участники, которые включают недавний опыт обучения в содержание своих ночных сновидений, демонстрируют повышенное улучшение производительности за ночь. Таким образом, это подтверждает гипотезу о том, что сновидения отражают обработку памяти в спящем мозге. Более того, по мнению авторов, одно из объяснений заключается в том, что сны, связанные с лабиринтом, являются индикатором того, что в спящем мозге реактивируются важные для производительности компоненты памяти о задачах. Кроме того, исследование подтверждает идею о том, что отчеты о сновидениях могут включать в себя экспериментальное обучающее задание на всех стадиях сна, включая фазу быстрого и медленного сна. [105]
Виртуальную реальность (VR) также использовали для изучения связи между сновидениями и пространственной памятью. Рибейро, Гунден и Кваглино (2021) [106] предложили пространственные элементы в контексте виртуальной реальности и обнаружили, что после полной ночи сна в домашней обстановке, когда изученный материал был включен в содержание сна, эффективность запоминания этих элементов была лучше, чем производительность, полученная после сопоставимого периода бодрствования. [106]
См. также
[ редактировать ]- Познание животных
- Когнитивная карта
- Диссоциация (нейропсихология)
- Метод локусов
- Пространственные способности
- Картографирование пространства
- Зрительная память
Ссылки
[ редактировать ]- ^ Берджесс, Нил (2021). Пространственная память . Британская энциклопедия.
- ^ Мехта, Митул А. (2010), «Пространственная память у людей» , в Столермане, Ян П. (редактор), Энциклопедия психофармакологии , Берлин, Гейдельберг: Springer, стр. 1262–1266, doi : 10.1007/978-3 -540-68706-1_355 , ISBN 978-3-540-68706-1 , получено 5 июня 2021 г.
- ^ «Пространственная память — обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 5 июня 2021 г.
- ^ Коларик, Б.; Экстром, А. (1 января 2015 г.). «Нейронные основы пространственной памяти и навигации» . Картирование мозга . стр. 507–514. дои : 10.1016/B978-0-12-397025-1.00277-3 . ISBN 9780123973160 .
- ^ Перейти обратно: а б Ван, Цзинь-Ху (2019). Ячейки ассоциативной памяти: основные единицы следа памяти . Спрингер Природа. п. 94. ИСБН 978-9811395017 .
- ^ Поппель, Дэвид; Мангун, Джордж Р.; Газзанига, Майкл С. (2020). Когнитивные нейронауки . Массачусетский технологический институт Пресс. п. 194. ИСБН 978-0262043250 .
- ^ Перейти обратно: а б с д и Джонсон, Э.; Адамо-Виллани, Н. (2010). «Исследование влияния погружения на кратковременную пространственную память». Инженерия и технологии . 71 : 582–587.
- ^ Перейти обратно: а б с д и ж г час я дж к л Анг, С.Ю.; Ли, К. (2008). «Центральное исполнительное участие в пространственной памяти детей». Память . 16 (8): 918–933. дои : 10.1080/09658210802365347 . ПМИД 18802804 . S2CID 42610047 .
- ^ Перейти обратно: а б с д и ж Джонс, Д.; Фарранд, П.; Стюарт, Г.; Моррис, Н.; и др. (1995). «Функциональная эквивалентность вербальной и пространственной информации в серийной кратковременной памяти». Журнал экспериментальной психологии: обучение, память и познание . 21 (4): 1008–1018. дои : 10.1037/0278-7393.21.4.1008 . ПМИД 7673864 .
- ^ Перейти обратно: а б с д и Делла Сала, С.; Грей, К.; Бэддели, А.; Алламано, Н.; Уилсон, Л.; и др. (1999). «Размах паттернов: инструмент для раскола зрительно-пространственной памяти». Нейропсихология . 37 (10): 1189–1199. дои : 10.1016/S0028-3932(98)00159-6 . ПМИД 10509840 . S2CID 1894333 .
- ^ Перейти обратно: а б с д и Маммарелла, IC; Паццалья, Ф.; Корнольди, К.; и др. (2008). «Доказательства существования различных компонентов зрительно-пространственной рабочей памяти детей» . Британский журнал психологии развития . 26 (3): 337–355. дои : 10.1348/026151007X236061 . hdl : 11577/2440989 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Калакоски, В.; Саарилуома, П. (2001). «Исключительная память таксистов на названия улиц» . Память и познание . 29 (4): 634–638. дои : 10.3758/BF03200464 . ПМИД 11504011 .
- ^ Фиск, Дж. Э.; Шарп, Калифорния; и др. (2003). «Роль исполнительной системы в функционировании зрительно-пространственной памяти». Мозг и познание . 52 (3): 364–381. дои : 10.1016/S0278-2626(03)00183-0 . ПМИД 12907181 . S2CID 37589101 .
- ^ Перейти обратно: а б с д и Чун, М.; Цзян, Ю. (1998). «Контекстная подсказка: неявное обучение и запоминание визуального контекста направляют пространственное внимание». Когнитивная психология . 36 (1): 28–71. CiteSeerX 10.1.1.25.5066 . дои : 10.1006/cogp.1998.0681 . ПМИД 9679076 . S2CID 1955059 .
- ^ Перейти обратно: а б Макнамара, Т.; Харди, Дж.; Хиртл, С.; и др. (1989). «Субъективные иерархии в пространственной памяти». Журнал экспериментальной психологии . 15 (2): 211–227. дои : 10.1037/0278-7393.15.2.211 . ПМИД 2522511 .
- ^ Перейти обратно: а б с Ньюман, Эл.; Каплан, Дж.Б.; Киршен, член парламента; Королев, И.О.; Секулер Р.; Кахана, MJ; и др. (2007). «Изучение передвижения по городу: как водители виртуальных такси учатся использовать информацию о планировке и ориентирах» (PDF) . Познание . 104 (2): 231–253. CiteSeerX 10.1.1.69.5387 . дои : 10.1016/j.cognition.2006.05.013 . ПМИД 16879816 . S2CID 267034 .
- ^ Гиллнер С., Маллот Х. (2000). «Роль глобальных и местных ориентиров в навигации по виртуальной среде». Восприятие . 9 (1): 69–83. CiteSeerX 10.1.1.138.8266 . дои : 10.1162/105474600566628 . S2CID 13062306 .
- ^ Крокос, Эрик; Плезант, Кэтрин; Варшней, Амитабх (16 мая 2018 г.). «Дворцы виртуальной памяти: вспоминание с помощью средств погружения» . Виртуальная реальность . 23 : 1–15. дои : 10.1007/s10055-018-0346-3 .
- ^ Перейти обратно: а б с д и ж Берд, LR; Робертс, Вашингтон; Абромс, Б.; Кит, Калифорния; Крупи, К. (2003). «Пространственная память на пищу, спрятанную крысами ( Rattus norvegicus ) в радиальном лабиринте: исследования памяти о том, где, что и когда». Журнал сравнительной психологии . 117 (2): 176–187. дои : 10.1037/0735-7036.117.2.176 . ПМИД 12856788 .
- ^ Дженнифер С. Холланд (8 августа 2014 г.). «Смотреть: как далеко бродят ваши кошки?» . Нэшнл Географик . Архивировано из оригинала 10 августа 2014 года . Проверено 23 августа 2016 г.
- ^ Перейти обратно: а б с Клауер, КЦ; Чжао, З.; и др. (2004). «Двойные диссоциации в зрительной и пространственной кратковременной памяти». Журнал экспериментальной психологии: Общие сведения . 133 (3): 355–381. дои : 10.1037/0096-3445.133.3.355 . ПМИД 15355144 . S2CID 7224613 .
- ^ Перейти обратно: а б с д и Пассолунги, MC; Маммарелла, IC; и др. (2010). «Пространственная и зрительная рабочая память у детей с трудностями в решении арифметических словесных задач». Европейский журнал когнитивной психологии . 22 (6): 944–963. дои : 10.1080/09541440903091127 . S2CID 143882629 .
- ^ Перейти обратно: а б с Маммарелла, IC; Паццалья, Ф.; Корнольди, К. (2008). «Свидетельства о различных компонентах зрительно-пространственной рабочей памяти детей» . Британский журнал психологии развития . 26 (3): 337–355. дои : 10.1348/026151007X236061 . hdl : 11577/2440989 .
- ^ Корси, премьер-министр (1972). «Человеческая память и медиальная височная область мозга». Авторефераты диссертаций . 34 (2): 891.
- ^ Делла Сала, С., Грей, К., Баддели, А., и Уилсон, Л. (1997). Тест визуальных паттернов: новый тест на кратковременное визуальное запоминание. Фелтэм, Саффолк: Испытательная компания Thames Valley.
- ^ Олтон, Д.С.; Самуэльсон, Р.Дж. (1976). «Память о прошлых местах: пространственная память у крыс». Журнал экспериментальной психологии: процессы поведения животных . 2 (2): 97–116. CiteSeerX 10.1.1.456.3110 . дои : 10.1037/0097-7403.2.2.97 .
- ^ Коул, MR; Чаппелл-Стефенсон, Робин (2003). «Исследование пределов пространственной памяти с помощью очень больших лабиринтов» . Обучение и поведение . 31 (4): 349–368. дои : 10.3758/BF03195996 . ПМИД 14733483 .
- ^ Моррис, Р.Г. (1981). «Пространственная локализация не требует присутствия локальных признаков». Обучение и мотивация . 12 (2): 239–260. дои : 10.1016/0023-9690(81)90020-5 .
- ^ Шарма, Сунита; Ракоци, Шарлин; Браун-Борг, Холли (23 октября 2010 г.). «Оценка пространственной памяти у мышей» . Науки о жизни . 87 (17): 521–536. дои : 10.1016/j.lfs.2010.09.004 . ISSN 0024-3205 . ПМК 6457258 . ПМИД 20837032 .
- ^ О'Киф, Дж.; Достровский, Ю. (1971). «Гиппокамп как пространственная карта. Предварительные данные об активности единиц у свободно передвигающейся крысы». Исследования мозга . 34 (1): 171–175. дои : 10.1016/0006-8993(71)90358-1 . ПМИД 5124915 .
- ^ Перейти обратно: а б с Сквайр, ЛР (1992). «Память и гиппокамп: синтез результатов исследований на крысах, обезьянах и людях». Психологический обзор . 99 (2): 195–231. дои : 10.1037/0033-295X.99.2.195 . ПМИД 1594723 . S2CID 14104324 .
- ^ Рамос, JMJ (2000). «Долговременная пространственная память у крыс с поражениями гиппокампа». Европейский журнал неврологии . 12 (9): 3375–3384. дои : 10.1046/j.1460-9568.2000.00206.x . ПМИД 10998120 . S2CID 18121369 .
- ^ Винокур, Г.; Москович, М.; Каруана, Д.А.; Биннс, Массачусетс (2005). «Ретроградная амнезия у крыс с поражением гиппокампа при тесте на пространственную память». Нейропсихология . 43 (11): 1580–1590. doi : 10.1016/j.neuropsychologia.2005.01.013 . ПМИД 16009240 . S2CID 9160452 .
- ^ Перейти обратно: а б с Лю, П.; Билки, ДК (2001). «Влияние эксайтотоксических поражений, сосредоточенных в гиппокампе или периринальной коре головного мозга, в задачах распознавания объектов и пространственной памяти». Поведенческая нейронаука . 115 (1): 94–111. дои : 10.1037/0735-7044.115.1.94 . ПМИД 11256456 .
- ^ Хеберт, А.Е.; Даш, ПК (2004). «Неизбыточные роли пластичности гиппокампа и энторинальной коры в хранении пространственной памяти». Фармакология Биохимия и поведение . 79 (1): 143–153. дои : 10.1016/j.pbb.2004.06.016 . ПМИД 15388294 . S2CID 25385758 .
- ^ Перейти обратно: а б Сейв, Э.; Пусет, Б.; Форман Н.; Бухот, М. (1992). «Исследование объектов и реакции на пространственные и непространственные изменения у крыс с капюшоном после повреждения теменной коры или образования гиппокампа». Поведенческая нейронаука . 106 (3): 447–456. дои : 10.1037/0735-7044.106.3.447 . ПМИД 1616611 .
- ^ Мартин, С.Дж.; де Озль, Л.; Моррис, RGM (2005). «Ретроградная амнезия: ни частичные, ни полные поражения гиппокампа у крыс не приводят к преимущественному сохранению удаленной пространственной памяти даже после напоминания». Нейропсихология . 43 (4): 609–624. doi : 10.1016/j.neuropsychologia.2004.07.007 . ПМИД 15716151 . S2CID 24894665 .
- ^ Баннерман, DM; Дьякон, RMJ; Оффен, С.; Фрисвелл, Дж.; Грабб, М.; Роулинз, JNP (2002). «Двойная диссоциация функций гиппокампа: пространственная память и гипонеофагия». Поведенческая нейронаука . 116 (5): 884–901. дои : 10.1037/0735-7044.116.5.884 . ПМИД 12369808 .
- ^ Мозер, М.; Мозер, Э.И. (1998). «Распределенное кодирование и восстановление пространственной памяти в гиппокампе» . Журнал неврологии . 18 (18): 7535–7542. doi : 10.1523/JNEUROSCI.18-18-07535.1998 . ПМК 6793256 . ПМИД 9736671 .
- ^ Перейти обратно: а б Ли, И.; Кеснер, Р.П. (2003). «Зависящие от времени взаимоотношения между дорсальным гиппокампом и префронтальной корой головного мозга в пространственной памяти» . Журнал неврологии . 23 (4): 1517–1523. doi : 10.1523/JNEUROSCI.23-04-01517.2003 . ПМЦ 6742248 . ПМИД 12598640 .
- ^ Макго, JL (2000). «Память — век консолидации». Наука . 287 (5451): 248–251. Бибкод : 2000Sci...287..248M . дои : 10.1126/science.287.5451.248 . ПМИД 10634773 . S2CID 40693856 .
- ^ Магуайр, Э.А.; Фраковяк, RSJ; Фрит, компакт-диск (1997). «Вспоминая маршруты по Лондону: Активация правого гиппокампа у таксистов» . Журнал неврологии . 17 (18): 7103–7110. doi : 10.1523/JNEUROSCI.17-18-07103.1997 . ПМК 6573257 . ПМИД 9278544 .
- ^ Брун, В.Х.; Отнаесс, МК; Молден, С.; Штеффенах, Х.; Виттер, член парламента; Мозер, М.; Мозер, Э.И. (2002). «Клетки места и распознавание места поддерживаются прямой энторинально-гиппокампальной схемой». Наука . 296 (5576): 2243–2246. Бибкод : 2002Sci...296.2243B . дои : 10.1126/science.1071089 . ПМИД 12077421 . S2CID 8458253 .
- ^ Гудрич-Хансакер, штат Нью-Джерси, Хансакер М.Р., Кеснер Р.П. (2008). «Взаимодействие и диссоциация субрегионов дорсального гиппокампа: как зубчатая извилина, CA3 и CA1 обрабатывают пространственную информацию». Поведение. Нейроски . 122 (1): 16–26. дои : 10.1037/0735-7044.122.1.16 . ПМИД 18298245 .
- ^ Сааб Б.Дж., Джорджиу Дж., Нат А., Ли Ф.Дж., Ван М., Михалон А., Лю Ф., Мансуи И.М., Родер Дж.К. (2009). «NCS-1 в зубчатой извилине способствует исследованию, синаптической пластичности и быстрому приобретению пространственной памяти» . Нейрон . 63 (5): 643–56. дои : 10.1016/j.neuron.2009.08.014 . ПМИД 19755107 . S2CID 5321020 .
- ^ Колби, CL; Голдберг, Мэн (1999). «Пространство и внимание в теменной коре». Ежегодный обзор неврологии . 22 : 319–349. дои : 10.1146/annurev.neuro.22.1.319 . ПМИД 10202542 . S2CID 14379470 .
- ^ Сейв, Э.; Могаддам, М. (1996). «Влияние поражений ассоциативной теменной коры на приобретение и использование пространственной памяти в задачах эгоцентрической и аллоцентрической навигации у крыс». Поведенческая нейронаука . 110 (1): 74–85. дои : 10.1037/0735-7044.110.1.74 . ПМИД 8652075 .
- ^ Перейти обратно: а б Чо, Ю.Х.; Кеснер, Р.П. (1996). «Вовлечение энторинальной коры или теменной коры в долговременную память о пространственной дискриминации у крыс: ретроградная амнезия». Поведенческая нейронаука . 110 (3): 436–442. дои : 10.1037/0735-7044.110.3.436 . ПМИД 8888988 .
- ^ Хафтинг, Т.; Фин, М.; Молден, С.; Мозер, М.; Мозер, Э.И. (2005). «Микроструктура пространственной карты энторинальной коры». Природа . 436 (7052): 801–806. Бибкод : 2005Natur.436..801H . дои : 10.1038/nature03721 . ПМИД 15965463 . S2CID 4405184 .
- ^ Фин, М.; Молден, С.; Виттер, член парламента; Мозер, Э.И.; Мозер, М. (2004). «Пространственное представление в энторинальной коре» . Наука . 305 (5688): 1258–1264. Бибкод : 2004Sci...305.1258F . дои : 10.1126/science.1099901 . ПМИД 15333832 .
- ^ Перейти обратно: а б Паррон, К.; Сейв, Э. (2004). «Сравнение влияния энторинальных и ретросплениальных поражений коры головного мозга на привыкание, реакцию на пространственные и непространственные изменения во время исследования объектов у крыс». Нейробиология обучения и памяти . 82 (1): 1–11. дои : 10.1016/j.nlm.2004.03.004 . ПМИД 15183166 . S2CID 36475737 .
- ^ Паррон, К.; Пусет, Б.; Сейв, Э. (2004). «Поражения энторинальной коры головного мозга ухудшают использование дистальных, но не проксимальных ориентиров во время навигации по месту у крысы». Поведенческие исследования мозга . 154 (2): 345–352. дои : 10.1016/j.bbr.2004.03.006 . ПМИД 15313022 . S2CID 919685 .
- ^ Нагахара, штат Ха; Отто, Т.; Галлахер, М. (1995). «Энторинально-периринальные поражения ухудшают работоспособность крыс в двух вариантах обучения месту в водном лабиринте Морриса». Поведенческая нейронаука . 109 (1): 3–9. дои : 10.1037/0735-7044.109.1.3 . ПМИД 7734077 .
- ^ Хеберт, А.Е.; Даш, ПК (2002). «Киназная активность, регулируемая внеклеточными сигналами, в энторинальной коре необходима для долговременной пространственной памяти» . Обучение и память . 9 (4): 156–166. дои : 10.1101/lm.48502 . ПМК 182586 . ПМИД 12177229 .
- ^ Пратт, МЫ; Мизумори, SJY (2001). «Нейроны в медиальной префронтальной коре крысы демонстрируют упреждающие изменения скорости в ответ на предсказуемое дифференциальное вознаграждение в задаче на пространственную память». Поведенческие исследования мозга . 123 (2): 165–183. дои : 10.1016/S0166-4328(01)00204-2 . ПМИД 11399329 . S2CID 3075976 .
- ^ Кеснер, Р.П.; Холбрук, Т. (1987). «Диссоциация пространственной памяти предметов и порядков у крыс после поражения медиальной префронтальной коры». Нейропсихология . 25 (4): 653–664. дои : 10.1016/0028-3932(87)90056-X . ПМИД 3658148 . S2CID 23596034 .
- ^ Слотник, С.Д.; Му, ЛР (2006). «Специализация полушарий префронтальной коры для категориальной и координационной зрительно-пространственной памяти». Нейропсихология . 44 (9): 1560–1568. doi : 10.1016/j.neuropsychologia.2006.01.018 . ПМИД 16516248 . S2CID 14396813 .
- ^ Беккер, Дж. Т.; Уокер, Дж.А.; Олтон, DS (1980). «Нейроанатомические основы пространственной памяти». Исследования мозга . 200 (2): 307–320. дои : 10.1016/0006-8993(80)90922-1 . ПМИД 7417818 . S2CID 1429885 .
- ^ Агглтон, JP; Нив, Н.; Нэгл, С.; Сахгал, А. (1995). «Сравнение влияния поражений медиальной префронтальной, поясной коры и поясных пучков на тесты пространственной памяти: свидетельства двойной диссоциации между вкладами лобных и поясных пучков» . Журнал неврологии . 15 (11): 7270–7281. doi : 10.1523/JNEUROSCI.15-11-07270.1995 . ПМК 6578066 . ПМИД 7472481 .
- ^ Лакруа, Л.; Белый, И.; Фелдон, Дж. (2002). «Влияние эксайтотоксических поражений медиальной префронтальной коры крыс на пространственную память». Поведенческие исследования мозга . 133 (1): 69–81. дои : 10.1016/S0166-4328(01)00442-9 . ПМИД 12048175 . S2CID 24280276 .
- ^ Купер, Б.Г.; Манка, ТФ; Мизумори, SJY (2001). «Найти дорогу в темноте: ретросплениальная кора способствует пространственной памяти и навигации без визуальных подсказок». Поведенческая нейронаука . 115 (5): 1012–1028. дои : 10.1037/0735-7044.115.5.1012 . ПМИД 11584914 .
- ^ Ванн, SD; Агглтон, JP (2002). «Обширные цитотоксические поражения ретросплениальной коры головного мозга крыс выявляют постоянный дефицит задач, требующих аллоцентрической пространственной памяти». Поведенческая нейронаука . 116 (1): 85–94. дои : 10.1037/0735-7044.116.1.85 . ПМИД 11895186 .
- ^ Ванн, SD; Уилтон, Луизиана; Мьюир, Дж.Л.; Агглтон, JP (2003). «Проверка важности каудальной ретросплениальной коры для пространственной памяти у крыс». Поведенческие исследования мозга . 140 (1–2): 107–118. дои : 10.1016/S0166-4328(02)00274-7 . ПМИД 12644284 . S2CID 10400806 .
- ^ Перейти обратно: а б Магуайр, Э.А. (2001). «Ретросплениальный вклад в навигацию человека: обзор поражений и результатов нейровизуализации». Скандинавский журнал психологии . 42 (3): 225–238. дои : 10.1111/1467-9450.00233 . ПМИД 11501737 .
- ^ Лю, П.; Билки, ДК (1998). «Вклад периринальной коры головного мозга в работу в водном лабиринте Морриса». Поведенческая нейронаука . 112 (2): 304–315. дои : 10.1037/0735-7044.112.2.304 . ПМИД 9588480 .
- ^ Гутброд, К; Коэн, Р; Майер, Т; Мейер, Э. (1987). «Память на пространственный и временной порядок у пациентов с афазией и поражением правого полушария» . Кортекс . 23 (3): 463–74. дои : 10.1016/s0010-9452(87)80007-2 . ПМИД 3677733 . S2CID 4489682 .
- ^ Нанн, Дж.А.; Грейдон, Ф.Дж.; Полки, CE; Моррис, Р.Г. (1999). «Дифференциальное нарушение пространственной памяти после правосторонней височной лобэктомии, продемонстрированное с помощью временного титрования» . Мозг . 122 (1): 47–59. дои : 10.1093/мозг/122.1.47 . ПМИД 10050894 .
- ^ Такер, DM; Хартри-Спейзер, А; Макдугал, Л; Луу, П; Дегранпре, Д. (1999). «Настроение и пространственная память: эмоции и вклад правого полушария в пространственное познание». Биол Психол . 50 (2): 103–25. дои : 10.1016/S0301-0511(99)00005-8 . ПМИД 10403200 . S2CID 42669695 .
- ^ Лян, КЦ; Достопочтенный, В; Тян, Ю.М.; Ляо, WL (1994). «Участие гиппокампальных рецепторов NMDA и AMPA в приобретении, формировании и восстановлении пространственной памяти в водном лабиринте Морриса». Чин Дж. Физиол . 37 (4): 201–12. ПМИД 7796636 .
- ^ Крузио, МЫ; Швеглер, Х. (2005). «Изучение задач пространственной ориентации в радиальном лабиринте и структурные изменения гиппокампа у инбредных мышей» . Поведенческие и функции мозга . 1 (1): 3. дои : 10.1186/1744-9081-1-3 . ПМЦ 1143776 . ПМИД 15916698 .
- ^ Ли, И.; Кеснер, Р.П. (2002). «Дифференциальный вклад рецепторов NMDA в субрегионах гиппокампа в пространственную рабочую память». Природная неврология . 5 (2): 162–168. дои : 10.1038/nn790 . ПМИД 11780144 . S2CID 17727921 .
- ^ Моррис, RGM; Андерсон, Э.; Линч, Г.С.; Бодри, М. (1986). «Селективное нарушение обучения и блокада долговременного потенциирования антагонистом рецептора N-метил-D-аспартата, AP5». Природа . 319 (6056): 774–776. Бибкод : 1986Natur.319..774M . дои : 10.1038/319774a0 . ПМИД 2869411 . S2CID 4356601 .
- ^ Ли, И.; Кеснер, Р.П. (2004). «Кодирование и восстановление пространственной памяти: двойная диссоциация между зубчатой извилиной и перфорантным путем, входящими в CA3 в дорсальном гиппокампе». Гиппокамп . 14 (1): 66–76. дои : 10.1002/hipo.10167 . ПМИД 15058484 . S2CID 21361912 .
- ^ Старк, М; Кослетт, HB; Шафран, Э.М. (1996). Нарушение эгоцентрической карты мест: последствия для восприятия и действий. 13. Когнитивная нейропсихология. стр. 481–523.
- ^ Тэ-Сон Лим, Джузеппе Иария, Со Молодая Мун. «Топографическая дезориентация при легких когнитивных нарушениях: исследование морфометрии на основе вокселей». 9 августа 2010 г. 16 апреля 2011 г. < http://www.neurolab.ca/2010(5)_Lim.pdf >.
- ^ Льюис, Д.А.; Левитт, П. (2002). «Шизофрения как нарушение нервного развития». Ежегодный обзор неврологии . 25 : 409–432. дои : 10.1146/annurev.neuro.25.112701.142754 . ПМИД 12052915 .
- ^ Липска, БК; Вайнбергер, Д.Р. (2000). «Моделировать психическое расстройство у животных: шизофрения как тест реальности» . Нейропсихофармакология . 23 (3): 223–239. дои : 10.1016/S0893-133X(00)00137-8 . ПМИД 10942847 .
- ^ Липска, БК; Олтман, Дж. М.; Верма, А.; Вайнбергер, доктор медицинских наук; Могаддам, Б. (2002). «Неонатальное повреждение вентрального гиппокампа ухудшает рабочую память у крыс» . Нейропсихофармакология . 27 (1): 47–54. дои : 10.1016/S0893-133X(02)00282-8 . ПМИД 12062906 .
- ^ Маркиз, JP; Гуле, С.; Доре, финансовый год (2008). «Диссоциируемое начало когнитивных и мотивационных дисфункций после неонатальных поражений вентрального гиппокампа у крыс». Поведенческая нейронаука . 122 (3): 629–642. дои : 10.1037/0735-7044.122.3.629 . ПМИД 18513133 .
- ^ Брэди, AM (2009). «Неонатальные поражения вентральной части гиппокампа нарушают способность к переключению установок у взрослых крыс». Поведенческие исследования мозга . 205 (1): 294–298. дои : 10.1016/j.bbr.2009.07.025 . ПМИД 19646488 . S2CID 29943421 .
- ^ Маркиз, JP; Гуле, С.; Доре, финансовый год (2008). «Поражения неонатального вентрального гиппокампа нарушают межпространственный сдвиг и изменяют плотность дендритных отростков в медиальной префронтальной коре молодых крыс». Нейробиология обучения и памяти . 90 (2): 339–346. дои : 10.1016/j.nlm.2008.04.005 . ПМИД 18490183 . S2CID 26066133 .
- ^ Винокур, Г. и Миллс, Дж. А. (1970). Переход между родственными и несвязанными проблемами после поражений гиппокампа у крыс. Журнал сравнительной и физиологической психологии
- ^ Левин, Эд; Кристофер, Северная Каролина (2006). «Влияние клозапина на функцию памяти на модели шизофрении при неонатальном гиппокампе крыс». Прогресс нейропсихофармакологии и биологической психиатрии . 30 (2): 223–229. дои : 10.1016/j.pnpbp.2005.10.018 . ПМИД 16356617 . S2CID 23594752 .
- ^ Сильва-Гомез, AB; Бермудес, М.; Кирион, Р.; Шривастава, ЛК; Пикасо, О.; Флорес, Г. (2003). «Сравнительные изменения в поведении самцов и самок крыс постпубертатного возраста после неонатальных эксайтотоксических поражений вентрального гиппокампа». Исследования мозга . 973 (2): 285–292. дои : 10.1016/S0006-8993(03)02537-X . ПМИД 12738072 . S2CID 17456907 .
- ^ Перейти обратно: а б Канугови С., Юн Дж.С., Фельдман Н.Х., Крото Д.Л., Мэттсон М.П., Бор В.А. (сентябрь 2012 г.). «Эндонуклеаза VIII-подобная 1 (NEIL1) способствует сохранению кратковременной пространственной памяти и защищает от дисфункции мозга и смерти, вызванной ишемическим инсультом» . Учеб. Натл. акад. наук. США . 109 (37): 14948–53. Бибкод : 2012PNAS..10914948C . дои : 10.1073/pnas.1204156109 . ПМЦ 3443144 . ПМИД 22927410 .
- ^ Ругински, Ян Т; Крим-Регер, Сара Х; Стефануччи, Жанин К; Кашдан, Элизабет (2019). «Использование GPS отрицательно влияет на обучение в окружающей среде из-за способности к пространственной трансформации» . Журнал экологической психологии . 64 : 12–20. дои : 10.1016/j.jenvp.2019.05.001 . S2CID 164329548 .
- ^ Монтелло, Д.Р.; Сасинка, Дж. М. (2006). «Человеческий фактор мореплавания» . В W. Карвовски, W (ред.). Международная энциклопедия эргономики и человеческого фактора . CRC Press/Taylor & Francisco, Ltd., стр. 2003–2008 гг.
- ^ Брюггер, А; Рихтер, К.Ф.; Фабрикант, С.И. (2019). «Как поведение навигационной системы влияет на поведение человека?» . Cogn Res Princ Implicity . 4 (1): 5. дои : 10.1186/s41235-019-0156-5 . ПМК 6374493 . ПМИД 30758681 .
- ^ Клеменсон, Грегори Д.; Маселли, Антонелла; Фианнака, Александр Дж.; Миллер, Амос; Гонсалес-Франко, март (01 января 2021 г.). «Переосмысление GPS-навигации: создание когнитивных карт с помощью слуховых подсказок» . Научные отчеты . 11 (1): 7764. Бибкод : 2021NatSR..11.7764C . дои : 10.1038/s41598-021-87148-4 . ISSN 2045-2322 . ПМЦ 8032695 . ПМИД 33833290 .
- ^ Лёвен, Генрих; Крукар, Якуб; Шверинг, Анжела (2019). «Пространственное обучение с помощью ориентационных карт: влияние различных особенностей окружающей среды на приобретение пространственных знаний» . Международный геоинформационный журнал ISPRS . 8 (3): 149. Бибкод : 2019IJGI....8..149L . дои : 10.3390/ijgi8030149 .
- ^ «Технология GPS увеличивает независимость» . www.alleganaesa.org . Проверено 8 мая 2023 г.
- ^ Йи, Джуэл; Ли, Хо Чунг-Юнг; Парсонс, Ричард; Фолкмер, Торбьорн (2014). «Влияние глобальной системы позиционирования на качество вождения людей с легкой болезнью Альцгеймера» . Геронтология . 61 (1): 79–88. дои : 10.1159/000365922 . ПМИД 25342271 . S2CID 27190155 .
- ^ Бартлетт, Рут; Браннелли, Тула; Топо, Пяйви (2019). «Использование технологий GPS для людей с деменцией» . Тидскрипт для Омсоргсфорскнинга . 5 (3): 84–98. doi : 10.18261/issn.2387-5984-2019-03-08 . S2CID 208989715 .
- ^ Маммарелла, IC; Луканджели, Д.; Корнольди, К. (2010). «Пространственная рабочая память и арифметические нарушения у детей с трудностями невербального обучения». Журнал неспособности к обучению . 43 (5): 455–468. дои : 10.1177/0022219409355482 . hdl : 11577/2453526 . ПМИД 20375290 . S2CID 2596429 .
- ^ Перейти обратно: а б с Феррара, Мишель; Иария, Джузеппе; Темпеста, Даниэла; Курчо, Джузеппе; Мороний, Фабио; Марцано, Кристина; Де Дженнаро, Луиджи; Пачитти, Клаудио (август 2008 г.). «Сон, чтобы найти свой путь: роль сна в консолидации памяти для навигации у людей» . Гиппокамп . 18 (8): 844–851. дои : 10.1002/hipo.20444 . ПМИД 18493970 . S2CID 25998045 .
- ^ Перейти обратно: а б Пибер, Доминик (01 ноября 2021 г.). «Роль нарушения сна и воспаления для пространственной памяти» . Мозг, поведение и иммунитет — Здоровье . 17 : 100333. дои : 10.1016/j.bbih.2021.100333 . ISSN 2666-3546 . ПМЦ 8474561 . ПМИД 34589818 .
- ^ Пенье, Филипп; Лорейс, Стивен; Фукс, Соня; Коллетт, Фабьен; Перрен, Фабьен; Реджерс, Джин; Филлипс, Кристоф; Дегельдре, Кристиан; Дель Фиоре, Гай; Аэртс, Жоэль; Люксен, Андре; Маке, Пьер (октябрь 2004 г.). «Усиливаются ли пространственные воспоминания в гиппокампе человека во время медленного сна?» . Нейрон . 44 (3): 535–545. дои : 10.1016/j.neuron.2004.10.007 . ПМИД 15504332 . S2CID 1424898 .
- ^ Перейти обратно: а б Плихал, Вернер; Родился Ян (сентябрь 1999 г.). «Влияние раннего и позднего ночного сна на прайминг и пространственную память» . Психофизиология . 36 (5): 571–582. дои : 10.1111/1469-8986.3650571 . ISSN 0048-5772 . ПМИД 10442025 .
- ^ Перейти обратно: а б с Гуань, Живэй; Пэн, Сюйвэнь; Фанг, Цзидун (20 августа 2004 г.). «Лишение сна ухудшает пространственную память и снижает фосфорилирование киназы, регулируемое внеклеточными сигналами, в гиппокампе» . Исследования мозга . 1018 (1): 38–47. дои : 10.1016/j.brainres.2004.05.032 . ISSN 0006-8993 . ПМИД 15262203 . S2CID 30718458 .
- ^ Ли, Юнли, Лия; Ван, Хунджу, Шевэй; Чэн, Чуаньлян, Дапэн (20 января 2016 г.) . : Данные исследования пространственной рабочей памяти фМРТ» . Европейская неврология . 75 (1–2): 48–57. : 10.1159 /000443372 . ISSN 0014-3022 . PMID 26784899. . S2CID 27938150 doi
- ^ Чен, Гуй-хай; Ся, Лан; Ван, Фанг; Ли, Сюэ-Вэй; Цзяо, Чуан-ань (октябрь 2016 г.). «Пациенты с хронической бессонницей имеют избирательные нарушения памяти, которые модулируются кортизолом: ухудшение памяти у больных хронической бессонницей» . Психофизиология . 53 (10): 1567–1576. дои : 10.1111/psyp.12700 . ПМИД 27412857 .
- ^ Хассауна, Башир Ю.; Батгейт, Кристина Дж.; Цай, Шейла С.; Эдингер, Джек Д. (декабрь 2018 г.). «Нейрокогнитивные функции при бессоннице: влияние повышенного возбуждения и короткой продолжительности сна» . Журнал исследований сна . 27 (6): e12747. дои : 10.1111/jsr.12747 . ПМИД 30069961 . S2CID 51892240 .
- ^ Он, Шуо; Чен, Си-Си; Ге, Вэй; Ян, Шуай; Чен, Цзюнь-Тао; Ню, Цзин-Вэнь; Ся, Лан; Чен, Гуй-Хай (29 июня 2021 г.). «Связаны ли противовоспалительные цитокины с когнитивными нарушениями у пациентов с бессонницей, коморбидной с депрессией? Пилотное исследование» . Природа и наука сна . 13 : 989–1000. дои : 10.2147/NSS.S312272 . ПМЦ 8254552 . ПМИД 34234602 .
- ^ Перейти обратно: а б Уомсли, Эрин Дж.; Стикголд, Роберт (февраль 2019 г.). «Сны об учебной задаче связаны с усилением консолидации памяти: повторение в исследовании ночного сна» . Журнал исследований сна . 28 (1): e12749. дои : 10.1111/jsr.12749 . ISSN 0962-1105 . ПМК 6338510 . ПМИД 30091247 .
- ^ Перейти обратно: а б Рибейро, Николас; Гунден, Янник; Кваглино, Вероник (15 апреля 2021 г.). «Усиление пространственной памяти на ассоциативном и реляционном уровнях после полной ночи сна и вероятность включения сновидений» . Международный журнал исследований сновидений . 14 : 67–79. дои : 10.11588/ijodr.2021.1.75882 . ISSN 1866-7953 . S2CID 234856576 .