Транскрипция (биология)
| Часть серии на |
| Генетика |
|---|
 |
|
|
Транскрипция - это процесс копирования сегмента ДНК в РНК. Сегменты ДНК, транскрибируемые в молекулы РНК, которые могут кодировать белки, продуцируют РНК мессенджера (мРНК). Другие сегменты ДНК транскрибируются в молекулы РНК, называемые некодирующими РНК (NCRNAS).
Как ДНК, так и РНК являются кислотами , которые используют пары оснований нуклеотидов нуклеиновыми в качестве дополнительного языка. Во время транскрипции последовательность ДНК считывается РНК -полимеразой , которая продуцирует комплементарную антипараллельную РНК -нить, называемую первичной транскрипцией .
В вирусологии термин транскрипция используется при ссылке на синтез мРНК из вирусной молекулы РНК. Геном РНК многих вирусов [ А ] состоит из РНК отрицательного смысла , которая действует как матрица для РНК с положительным смыслом вирусного мессенджера - необходимый шаг в синтезе вирусных белков, необходимых для репликации вируса . Этот процесс катализируется вирусной РНК -зависимой РНК -полимеразой . [ 1 ]
Фон
[ редактировать ]Блок транскрипции ДНК, кодирующий для белка, может содержать как кодирующую последовательность , которая будет транслироваться в белок, так и регуляторные последовательности , которые направляют и регулируют синтез этого белка. Регуляторная последовательность перед ( вверх по течению от) кодирующая последовательность называется пятью основными нетранслируемыми областями (5'UTR); Последовательность после ( вниз по течению) кодирующая последовательность называется тремя основными нетранслируемыми областями (3'UTR). [ 2 ]
В отличие от репликации ДНК , транскрипция приводит к комплементу РНК, который включает нуклеотид урацил (U) во всех случаях, когда тимин (T) произошел бы в комплементе ДНК. [ 3 ]
Только одна из двух прядей ДНК служит шаблоном для транскрипции. Антисмысленная . цепь ДНК считывается РНК -полимеразой с 3 'конца до 5' конца во время транскрипции (3 '→ 5') Дополнительная РНК создается в противоположном направлении, в направлении 5 '→ 3', что соответствует последовательности цепочки смысла, за исключением переключения урацила на тимин. Эта направленность связана с тем, что РНК -полимераза может добавлять только нуклеотиды в 3 'конце растущей цепи мРНК. Это использование только 3 '→ 5' Цепи ДНК устраняет необходимость в фрагментах Оказаки , которые наблюдаются при репликации ДНК. [ 2 ] Это также устраняет необходимость в праймере РНК для инициирования синтеза РНК, как в случае репликации ДНК.
Несоверная , потому что ее последовательность такая же , (смысл) цепь ДНК называется кодирующей цепью как вновь созданная транскрипт РНК (за исключением замены урацила на тимин). Это прядь, которая используется соглашением при представлении последовательности ДНК. [ 4 ]
Транскрипция имеет некоторые механизмы корректуры, но они меньше и менее эффективны, чем контроль для копирования ДНК. В результате транскрипция имеет более низкую копирую, чем репликация ДНК. [ 5 ]
Основные шаги
[ редактировать ]Транскрипция разделена на инициацию , выпуск промотора , удлинение и прекращение . [ 6 ]
Настройка для транскрипции
[ редактировать ]В этом разделе может потребоваться очистка Википедии для соответствия стандартам качества . Конкретная проблема: дублирование с регуляторной последовательности . Можем ли мы просто сделать каноническую «главную статью» и перенаправить там людей? ( Сентябрь 2021 г. ) |
Энхансеры, транскрипционные факторы, медиаторный комплекс и петли ДНК в транскрипции млекопитающих
[ редактировать ]Настройка для транскрипции у млекопитающих регулируется многими цис-регуляторными элементами , включая промотор-сердечник и промотор-проксимальные элементы , которые расположены вблизи участков начала транскрипции генов. Основные промоторы в сочетании с общими факторами транскрипции являются достаточными для прямого инициации транскрипции, но, как правило, имеют низкую базальную активность. [ 7 ] Другие важные цис-регуляторные модули локализованы в областях ДНК, которые отдали от участков начала транскрипции. К ним относятся усилители , глушители , изоляторы и привязанные элементы. [ 8 ] Среди этого созвездия элементов усилители и связанные с ними факторы транскрипции играют ведущую роль в инициации транскрипции генов. [ 9 ] Энхансер, локализованный в области ДНК, отдаленной от промотора гена, может оказать очень большое влияние на транскрипцию генов, при этом некоторые гены, подвергающиеся увеличению транскрипции до 100 раз из-за активированного энхансера. [ 10 ]
Усильники являются областями генома, которые являются основными ген-регуляторными элементами. Энхансеры контролируют программы транскрипции генов специфичных клеток, чаще всего путем зацикливания через большие расстояния, чтобы они находились в физической близости с промоторами их целевых генов. [ 11 ] В то время как существуют сотни тысяч областей ДНК энхансеры, [ 12 ] Для конкретного типа ткани только специфические усилители находятся в непосредственной близости от промоторов, которые они регулируют. В исследовании корковых нейронов головного мозга было обнаружено 24 937 петли, что привело к усилениям к их промоторам -мишеням. [ 10 ] Множественные усилители, каждый из которых часто на десятках или сотнях тысяч нуклеотидов, отдаленных от их генов -мишеней, перецироваться к их промоторам генов -мишени и могут координировать друг с другом для контроля транскрипции их общего гена -мишени. [ 11 ]
Схематическая иллюстрация в этом разделе показывает, как усилитель, зацикливаясь вокруг, чтобы прийти в тесную физическую близость с промотором целевого гена. Петля стабилизируется димером разъема белка (например, димер CTCF или YY1 ), причем один член димера прикреплен к его связывающему мотиву на энхансере, а другой, прикрепленный к его связывающему мотиву на промоторе (представленным Красные зигзаги на иллюстрации). [ 13 ] Несколько специфических факторов транскрипции функции клеток (в клетке человека насчитывается около 1600 факторов транскрипции. [ 14 ] ), как правило, связываются с конкретными мотивами на энхансере [ 15 ] и небольшая комбинация этих факторов транскрипции, связанных с энхансером, когда она приближается к промотору с помощью петли ДНК, управлять уровнем транскрипции гена-мишени. Медиатор (комплекс, обычно состоящий из примерно 26 белков в взаимодействующей структуре), передает регуляторные сигналы из энхансерных ДНК-связанных факторов транскрипции непосредственно с ферментом РНК-полимеразы II (POL II), связанного с промотором. [ 16 ]
Энхансеры, когда они активны, обычно транскрибируются из обеих целей ДНК с РНК -полимеразами, действующими в двух разных направлениях, продуцируя два РНК усилителя (ERNAS), как показано на рисунке. [ 17 ] Неактивный энхансер может быть связан неактивным фактором транскрипции. Фосфорилирование фактора транскрипции может активировать его и что активированный фактор транскрипции может затем активировать энхансер, с которым он связан (см. Маленькую красную звезду, представляющую фосфорилирование фактора транскрипции, связанного с энхансером на иллюстрации). [ 18 ] Активированный энхансер начинает транскрипцию своей РНК перед активацией транскрипции мессенджера РНК из его мишени. [ 19 ]
Метилирование и деметилирование острова CPG
[ редактировать ]
Регуляция транскрипции примерно в 60% промоторов также контролируется метилированием цитозинов в динуклеотидах CPG (где 5 'цитозин сопровождается 3' сайтами гуанина или CPG ). 5-метилцитозин (5-MC) представляет собой метилированную форму ДНК основания цитозина (см. Рисунок). 5-MC-это эпигенетический маркер, который можно было преимущественно в местах CPG. Около 28 миллионов динуклеотидов CPG встречаются в геноме человека. [ 20 ] В большинстве тканей млекопитающих в среднем от 70% до 80% цитозинов CPG метилируются (образуя 5-метилкпг или 5-MCPG). [ 21 ] Тем не менее, неметилированные цитозины в 5'ccytosine-guanine 3 'последовательностях часто встречаются в группах, называемых островами CPG , у активных промоторов. Около 60% промоторных последовательностей имеют остров CPG, в то время как только около 6% последовательностей энхансеров имеют остров CPG. [ 22 ] Острова CPG составляют регуляторные последовательности, поскольку, если острова CPG метилируются в промоторе гена, это может уменьшить или замолчать транскрипцию генов. [ 23 ]
Метилирование ДНК регулирует транскрипцию генов посредством взаимодействия с метил -связывающим доменом (MBD) белками, такими как MECP2, MBD1 и MBD2. Эти белки MBD наиболее сильно связываются с высокометилированными островами CPG . [ 24 ] Эти белки MBD имеют как метил-CPG-связывающий домен, так и домен репрессии транскрипции. [ 24 ] Они связываются с метилированной ДНК и направляют или прямые белковые комплексы с ремоделированием хроматина и/или модификацией гистонов на метилированные острова CPG. Белки MBD обычно репрессируют локальный хроматин, например, путем катализирования введения репрессивных марок гистонов или создания общей репрессивной хроматиновой среды посредством ремоделирования нуклеосом и реорганизации хроматина. [ 24 ]

Как отмечалось в предыдущем разделе, факторы транскрипции - это белки, которые связываются со специфическими последовательностями ДНК, чтобы регулировать экспрессию гена. Последовательность связывания для транскрипционного фактора в ДНК обычно составляет около 10 или 11 нуклеотидов длиной. Как обобщено в 2009 году, Vaquerizas et al. Указано, что в генах человека есть около 1400 различных транскрипционных факторов, кодируемых в геноме человека, которые составляют около 6% всех генов, кодирующих белок человека. [ 25 ] Около 94% сайтов связывания транскрипционных факторов (TFBS), которые связаны с генами, реагирующими на сигнал, встречаются у энхансеров, в то время как у промоторов встречается только около 6% таких TFBS. [ 15 ]
Белок EGR1 является конкретным транскрипционным фактором, который важен для регуляции метилирования островов CPG. Сайт связывания фактора транскрипции EGR1 часто расположен в энхансере или промоторных последовательностях. [ 26 ] В геноме млекопитающих насчитывается около 12 000 сайтов связывания EGR1, и около половины сайтов связывания EGR1 расположены у промоторов и половины усилителей. [ 26 ] Связывание EGR1 с сайтом связывания ДНК -мишени нечувствительно к метилированию цитозина в ДНК. [ 26 ]
В то время как только небольшие количества белка транскрипционного фактора EGR1 обнаруживаются в клетках, которые не стимулированы, трансляция гена EGR1 в белок через час после стимуляции резко повышается. [ 27 ] Производство белков транскрипционного фактора EGR1 в различных типах клеток может стимулировать факторами роста, нейротрансмиттеров, гормонами, стрессом и повреждением. [ 27 ] В мозге, когда нейроны активируются, белки EGR1 активируются, и они связываются с (рекрутирующими) ранее существовавшими ферментами TET1 , которые продуцируются в больших количествах в нейронах. Тет-ферменты могут катализировать деметилирование 5-метилцитозина. Когда факторы транскрипции EGR1 приносят ферменты TET1 в сайты связывания EGR1 у промоторов, ферменты TET могут деметилизировать метилированные острова CPG у этих промоторов. После деметилирования эти промоторы могут затем инициировать транскрипцию своих целевых генов. Сотни генов в нейронах дифференциально экспрессируются после активации нейронов посредством рекрутирования EGR1 TET1 для метилированных регуляторных последовательностей у их промоторов. [ 26 ]
Метилирование промоторов также изменяется в ответ на сигналы. Три ДНК -метилтрансферазы (DNMT1, DNMT3A и DNMT3B) катализируют добавление метильных групп в цитозины в ДНК. В то время как DNMT1 является поддерживающей метилтрансферазой, DNMT3A и DNMT3B могут выполнять новые метилирование. Существуют также две сплайсинга изоформы белка , продуцируемые из гена DNMT3A : белки ДНК метилтрансферазы DNMT3A1 и DNMT3A2. [ 28 ]
Изоформа сплайсинга DNMT3A2 ведет себя как продукт классического гена с непосредственным ранним и, например, он надежно и временно продуцируется после активации нейронов. [ 29 ] Где изоформа ДНК -метилтрансфераза DNMT3A2 связывается и добавляет метильные группы к цитозинам, по -видимому, определяется с помощью гистоновых пост трансляционных модификаций. [ 30 ] [ 31 ] [ 32 ]
С другой стороны, нейронная активация вызывает деградацию DNMT3A1, сопровождаемое снижением метилирования, по меньшей мере, одного оцениваемого целевого промотора. [ 33 ]
Инициация
[ редактировать ]Транскрипция начинается с РНК-полимеразы и одного или нескольких общих факторов транскрипции , связывающегося с последовательности промотора ДНК с образованием закрытого комплекса РНК-полимеразы. В замкнутом комплексе промоторная ДНК все еще полностью двухцепочечная. [ 6 ]
РНК-полимераза, помогая одним или несколькими общими факторами транскрипции, затем расстреляет приблизительно 14 пар оснований ДНК с образованием открытого комплекса РНК-полимеразы. В открытом комплексе промоторная ДНК частично разбита и одноцепочечная. Открытая одноцепочечная ДНК называется «пузырьком транскрипции». [ 6 ]
РНК -полимераза, помогая одним или несколькими общими факторами транскрипции, затем выбирает начальный сайт транскрипции в пузырьке транскрипции, связывается с инициативным NTP и расширяющимся NTP (или коротким РНК -праймером и расширяющим NTP), дополняющим последовательность начала транскрипции. и катализирует образование связей с учетом начального РНК -продукта. [ 6 ]
У бактерий РНК -полимеразы голоефермент состоит из пяти субъединиц: 2 α -субъединицы, 1 β -субъединицы, субъединицы 1 β 'и субъединицы 1 Ом. У бактерий есть один общий фактор транскрипции РНК, известный как сигма -фактор . РНК -полимеразное фермент сердечник связывается с бактериальным общим транскрипционным фактором (Sigma) с образованием голоефермента РНК -полимеразы, а затем связывается с промотором. [ 6 ] (РНК -полимераза называется голооферментом, когда субъединица Sigma прикреплена к основному ферменту, который состоят из 2 α -субъединицы, 1 β -субъединицы, только 1 β 'субъединицы). В отличие от эукариот, инициирующий нуклеотид зарождающейся бактериальной мРНК не ограничен модифицированным гуаниновым нуклеотидом. Инициирующий нуклеотид бактериальных транскриптов имеет 5'-трифосфат (5'-PPP), который может использоваться для картирования всего генома сайтов инициации транскрипции. [ 35 ]
У археи и эукариот РНК -полимераза содержит субъединицы, гомологичные каждой из пяти субъединиц РНК -полимеразы у бактерий, а также содержит дополнительные субъединицы. У археи и эукариот функции бактериального общего транскрипционного фактора Sigma выполняются множеством общих факторов транскрипции, которые работают вместе. [ 6 ] В археи есть три общих фактора транскрипции: TBP , TFB и TFE . У эукариот в РНК -полимеразе II транскрипции существует шесть общих факторов транскрипции: TFIIA , TFIIB ( ортолог архаального TFB), TFIID (мультисубонитный фактор, в котором ключевой субъединица, TBP, является ортологом архиального TBP),, ключевой субъединицы, TBP , является ортологом археального TBP), Tfiie ( ортолог архаального TFE), tfiif и tfiih Полем TFIID является первым компонентом, связывающим с ДНК из -за связывания TBP, в то время как TFIIH является последним компонентом, который должен быть набран. У археи и эукариот закрытый комплекс РНК-полимеразы-промоттер обычно называют « комплексом предварительного проживания ». [ 36 ]
Инициирование транскрипции регулируется дополнительными белками, известными как активаторы и репрессоры , и, в некоторых случаях, связанными коактиваторами или корепрессорами , которые модулируют образование и функцию комплекса инициации транскрипции. [ 6 ]
Промоутер побег
[ редактировать ]После того, как первая связь синтезируется, РНК -полимераза должна избежать промотора. В течение этого времени существует тенденция высвобождать транскрипт РНК и производить усеченные транскрипты. Это называется абортивным инициацией и распространено как для эукариот, так и для прокариот. [ 37 ] Абортивное инициация продолжает происходить до тех пор, пока не будет синтезируется РНК -продукт пороговой длины приблизительно 10 нуклеотидов, после чего возникает выход промотора и формируется комплекс удлинения транскрипции. [ Цитация необходима ]
Механистически, выход промотора происходит за счет Scrunging ДНК , обеспечивая энергию, необходимую для разбивания взаимодействия между голоуферментом РНК -полимеразы и промотором. [ 38 ]
В бактериях исторически считалось, что фактор сигма определенно выпущен после того, как промоторный очистка. Эта теория была известна как модель облигатного выпуска. Однако более поздние данные показали, что после и после очистки промотора фактор сигма высвобождается в соответствии со стохастической моделью, известной как модель стохастического высвобождения . [ 39 ]
У эукариот при РНК-полимеразе II-зависимого промотора при промоторном клиренсе TFIIH фосфорилирует серин 5 на карбокси-терминальном домене РНК-полимеразы II, что приводит к рекрутированию фермента с покрытием (CE). [ 40 ] [ 41 ] Точный механизм того, как CE индуцирует промоторный клиренс у эукариот, еще не известен.
Удлинение
[ редактировать ]Одна цепь ДНК, шаблона цепи (или некодирующая нить), используется в качестве матрицы для синтеза РНК. По мере прохождения транскрипции РНК -полимераза пересекает цепь матрицы и использует комплементарность спаривания основания с матрицей ДНК для создания копии РНК (которая удлиняется во время обхода). Хотя РНК-полимераза пересекает шаблонную цепь от 3 '→ 5', кодирующая (неэлемтная) цепь и вновь образованная РНК также могут использоваться в качестве контрольных точек, поэтому транскрипция может быть описана как происходящая 5 '→ 3'. Это производит молекулу РНК из 5 '→ 3', точную копию кодирующей нити (за исключением того, что тимины заменяются урацилами , а нуклеотиды состоят из рибозы (5-углеродого) сахара, тогда как ДНК имеет дезоксирибозу (один меньше кислорода. атом) в своей основе сахара-фосфата). [ 3 ]
Транскрипция мРНК может включать множественные РНК -полимеразы в одном матрице ДНК и множественные раунды транскрипции (амплификация определенной мРНК), так что многие молекулы мРНК могут быстро продуцироваться из одной копии гена. [ Цитация необходима ] Характерные показатели удлинения у прокариот и эукариот составляют около 10–100 нс/сек. [ 42 ] Однако у эукариот нуклеосомы действуют как основные барьеры для транскрибирования полимераз во время удлинения транскрипции. [ 43 ] [ 44 ] В этих организмах пауза, индуцированная нуклеосом, может регулироваться факторами удлинения транскрипции, такими как TFIIS. [ 44 ]
Удлинение также включает в себя механизм корректурирования, который может заменить неправильно включенные основания. У эукариот это может соответствовать коротким паузам во время транскрипции, которые позволяют связывать соответствующие факторы редактирования РНК. Эти паузы могут быть внутренними для РНК -полимеразы или из -за структуры хроматина. [ Цитация необходима ]
Двойные разрывы в активно транскрибируемых областях ДНК восстанавливаются гомологичной рекомбинацией во время фаз S и G2 клеточного цикла . [ 45 ] [ 46 ] Поскольку транскрипция усиливает доступность ДНК в экзогенные химические вещества и внутренние метаболиты, которые могут вызывать рекомбиногенные поражения, гомологичная рекомбинация конкретной последовательности ДНК может сильно стимулировать транскрипцией. [ 47 ]
Завершение
[ редактировать ]Бактерии используют две разные стратегии для прекращения транскрипции-Rh-независимое прекращение и Rho-зависимое прекращение. В Rh-независимом завершении транскрипции транскрипция РНК останавливается, когда вновь синтезированная молекула РНК образует богатую GC петлю шпильки с последующей пробежкой нас. Когда образуется шпилька, механическое напряжение разрушает слабые связи Ru-DA, теперь заполняя гибрид ДНК-РНК. Это вытаскивает транскрипт поли-U из активного сайта РНК-полимеразы, завершая транскрипцию. В Rho-зависимое завершение, Rho , белковой фактор, дестабилизирует взаимодействие между шаблоном и мРНК, что высвобождает вновь синтезированную мРНК из комплекса удлинения. [ 48 ]
Прекращение транскрипции у эукариот менее хорошо изучено, чем у бактерий, но включает в себя расщепление нового транскрипта с последующим шаблоном, независимым от добавления аденинов на его новом 3-дюймовом конце, в процессе, называемом полиаденилированием . [ 49 ]
Помимо прекращения последовательностями терминатора (которая является частью гена ) , транскрипция также может быть прекращена, когда она сталкивается с такими условиями, как повреждение ДНК или активная репликация вилка . У бактерий MFD АТФаза может удалять РНК -полимеразу, остановившуюся при поражении, открывая его зажим. Он также набирает механизм восстановления нуклеотидных ударов для ремонта поражения. Предлагается, что MFD также разрешает конфликты между репликацией ДНК и транскрипцией. [ 50 ] В Eukayrotes ATPase TTF2 помогает подавить действие RNAP I и II во время митоза , предотвращая ошибки в хромосомной сегрегации. [ 51 ] В Archaea предложена ATAPase ETA, чтобы играть аналогичную роль. [ 52 ]
Роль РНК-полимеразы в посттранскрипционных изменениях РНК
[ редактировать ]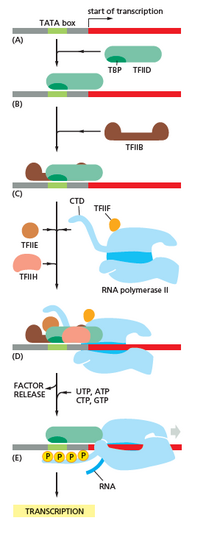
РНК-полимераза играет очень важную роль во всех этапах, включая посттранскрипционные изменения РНК.
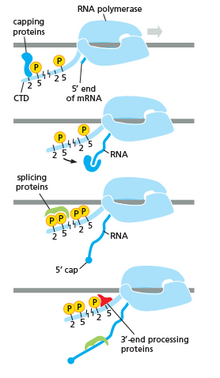
Как показано на изображении справа. Очевидно, что CTD (C -терминальный домен) - это хвост, который изменяет его форму; Этот хвост будет использоваться в качестве носителя сплайсинга, уплотнения и полиаденилирования , как показано на изображении слева. [ 53 ]
Ингибиторы
[ редактировать ]Ингибиторы транскрипции могут использоваться в качестве антибиотиков против, например, патогенных бактерий ( антибактерий ) и грибов ( антигрифганты ). Примером такого антибактериального является рифампицин , который ингибирует бактериальную транскрипцию ДНК в мРНК, ингибируя ДНК-зависимую РНК-полимеразу путем связывания его бета-субъединицы, в то время как 8-гидроксихинолин является ингибитором противогрибкового транскрипции. [ 54 ] Эффекты метилирования гистонов также могут работать, чтобы ингибировать действие транскрипции. Мощные биологически активные натуральные продукты, такие как триптолид, которые ингибируют транскрипцию млекопитающих посредством ингибирования субъединицы XPB общего фактора транскрипции TFIIH, недавно сообщалось в качестве конъюгата глюкозы для нацеливания гипоксических раковых клеток с увеличением продукции транспортера глюкозы. [ 55 ]
Эндогенные ингибиторы
[ редактировать ]У позвоночных большинство генных промоторов содержат остров CPG с многочисленными участками CPG . [ 56 ] Когда многие из участков промотора гена метилированы, ген ингибируется (молчали). [ 57 ] Колоректальный рак обычно имеет от 3 до 6 драйвера мутаций с автостопом или пассажирами. и от 33 до 66 мутаций [ 58 ] Тем не менее, ингибирование транскрипции (молчание) может иметь большее значение, чем мутация при выборе прогрессирования рака. Например, при колоректальном раке от 600 до 800 генов транскрипционно ингибируются метилированием острова CPG (см. Регуляцию транскрипции при раке ). Репрессия транскрипции при раке также может возникать другими эпигенетическими механизмами, такими как измененная продукция микроРНК . [ 59 ] При раке молочной железы транскрипционная репрессия BRCA1 может происходить чаще с помощью переоценки микроРНК-182, чем при гиперметилировании промотора BRCA1 (см. Низкую экспрессию BRCA1 при раке молочной железы и яичников ). [ Цитация необходима ]
Транскрипционные фабрики
[ редактировать ]Активные транскрипционные единицы кластерируются в ядре, в дискретных участках, называемых транскрипционными фабриками или эухроматином . Такие сайты могут быть визуализированы, позволяя заинтересованной полимеразам расширять свои транскрипты в предшественниках с меткой (BR-UTP или BR-U) и иммуно-маркировки помеченной зарождающейся РНК. Транскрипционные фабрики также могут быть локализованы с использованием гибридизации флуоресценции in situ или отмечены антителами, направленными против полимераз. В нуклеоплазме клетки HeLa есть ~ 10000 фабрик , среди которых представлены заводы ~ 8000 полимеразы II и ~ 2000 заводов полимеразы III. Каждая фабрика полимеразы II содержит ~ 8 полимеразы. Поскольку большинство активных транскрипционных единиц связаны только с одной полимеразой, каждая фабрика обычно содержит ~ 8 различных единиц транскрипции. Эти единицы могут быть связаны с промоторами и/или усилителями, причем петли формируют «облако» вокруг этого фактора. [ 60 ]
История
[ редактировать ]Молекула, которая позволяет реализовать генетический материал как белок, была сначала предполагалась монодом Франсуа Джейкоба и Жака . Северо Очоа получил Нобелевскую премию по физиологии или медицине в 1959 году за разработку процесса синтеза РНК in vitro с полинуклеотидфосфорилазой , который был полезен для взлома генетического кода . Синтез РНК с помощью РНК -полимеразы был установлен in vitro несколькими лабораториями к 1965 году; Однако РНК, синтезированная этими ферментами, имели свойства, которые предполагают существование дополнительного фактора, необходимого для правильного прекращения транскрипции. [ Цитация необходима ]
Роджер Д. Корнберг получил Нобелевскую премию 2006 года по химии «за исследования молекулярной основы эукариотической транскрипции ». [ 61 ]
Измерение и обнаружение
[ редактировать ]Транскрипция может быть измерена и обнаружена различными способами: [ Цитация необходима ]
- кассеты без G : Анализ транскрипции
- Анализ транскрипции стока : идентифицирует начальные сайты транскрипции (TSS)
- Анализ по ядерным забегам : измеряет относительное количество вновь сформированных транскриптов
- Kas-seq : измеряет одноцепочечную ДНК, генерируемую РНК-полимеразами; может работать с 1000 ячеек. [ 62 ]
- Анализ защиты РНКазы : обнаружение активных и чип-чип RNAP сайтов транскрипции
- RT-PCR : измеряет абсолютное количество уровней общего или ядерного РНК, что, однако, может отличаться от скорости транскрипции
- Микрочипы ДНК : измеряют относительную численность глобальных уровней общего или ядерного РНК; Однако они могут отличаться от скорости транскрипции
- Гибридизация in situ : обнаруживает наличие транскрипта
- Метка MS2 : Включив петли ствола РНК , такие как MS2, в ген, они становятся включенными в вновь синтезированную РНК. Затем петли ствола могут быть обнаружены с использованием слияния GFP и белка покрытия MS2, который имеет высокое аффинное, специфичное для последовательности взаимодействие с петлями ствола MS2. Рекрутирование GFP в место транскрипции визуализируется как одно флуоресцентное место. Этот новый подход показал, что транскрипция происходит в прерывистых всплесках или импульсах (см. Разрыв транскрипции ). За заметным исключением методов in situ, большинство других методов обеспечивают средние показатели сотовой популяции и не способны обнаружить это фундаментальное свойство генов. [ 63 ]
- Северный блот : традиционный метод, и до появления РНК-seq , самый количественный
- RNA-seq : применяет методы секвенирования следующего поколения для последовательности целых транскриптомов , что позволяет измерять относительную численность РНК, а также обнаружение дополнительных вариаций, таких как гены слияния, посттранскрипционные редакторы и новые сайты сплайсинга
- РНК-seq с одной клеток : усиливает и считывает частичные транскриптомы из изолированных клеток, что позволяет проводить подробный анализ РНК в тканях, эмбрионах и раке
Обратная транскрипция
[ редактировать ]Некоторые вирусы (такие как ВИЧ , причина СПИДа ), способны транскрибировать РНК в ДНК. ВИЧ имеет геном РНК, который транскрибируется в ДНК. Полученная ДНК может быть объединена с геномом ДНК клетки -хозяина. Основной фермент, ответственный за синтез ДНК из матрицы РНК, называется обратной транскриптазой . [ 64 ]
В случае ВИЧ обратная транскриптаза отвечает за синтезирование комплементарной ДНК -цепи (кДНК) к геному вирусной РНК. Фермент рибонуклеаза H затем переваривает РНК -цепи, а обратная транскриптаза синтезирует комплементарную цепь ДНК с образованием ДНК -структуры двойной спирали (кДНК). КДНК интегрируется в геном клетки хозяина ферментом интегразы , что заставляет клетку -хозяина генерировать вирусные белки, которые собирают новые вирусные частицы. При ВИЧ после этого клетка -хозяина подвергается запрограммированной гибели клеток или апоптозу -клеток Т . [ 65 ] Однако в других ретровирусах клетка -хозяина остается нетронутой в качестве вирусных почек из клетки. [ Цитация необходима ]
Некоторые эукариотические клетки содержат фермент с активностью обратной транскрипции, называемой теломеразой . Теломераза несет матрицу РНК, из которого он синтезирует теломер , повторяющуюся последовательность ДНК, до конца линейных хромосом. Это важно, потому что каждый раз, когда линейная хромосома дублируется, она сокращается. С теломерой на концах хромосом, укорочение устраняет некоторые из несущественных, повторяющихся последовательности, а не белок-кодирующей последовательности ДНК дальше от конца хромосомы.
Теломераза часто активируется в раковых клетках, чтобы позволить раковым клеткам дублировать свои геномы на неопределенный срок без потери важной последовательности ДНК, кодирующей белок. Активация теломеразы может быть частью процесса, который позволяет раковым клеткам стать бессмертными. Было доказано, что иммортизирующий фактор рака посредством удлинения теломер из -за теломеразы встречается у 90% всех канцерогенных опухолей in vivo с оставшимися 10% с использованием альтернативного пути поддержания теломер, называемого ALT или альтернативным удлинением теломер. [ 66 ]
Смотрите также
[ редактировать ]- Жизнь
- Клетка (биология)
- Клеточная деление
- DBTSS
- ген
- генная регуляция
- Экспрессия гена
- Эпигенетика
- Геном
- Генная регуляция
- Длинная некодирующая РНК
- Миссенс -мРНК
- Сплайсинг -процесс удаления интронов из РНК-предшественника Мессенджера ( пре-мРНК ) для создания РНК-посланника ( мРНК )
- Транскриптомика
- Перевод (биология)
Примечания
[ редактировать ]- ^ Примечательные вирусы позвоночных - Ссрна включают вирус Эбола, хантавирусы, вирусы гриппа, вирус ласс -лихорадки и вирус бешенства.
Ссылки
[ редактировать ]- ^ Koonin EV, Gorbalenya AE, Chumakov KM (июль 1989 г.). «Предварительная идентификация РНК-зависимых РНК-полимераз вирусов дцРНК и их связь с положительными вирусными вирусными полимеразами РНК на цепи» . Письма Febs . 252 (1–2): 42–6. Bibcode : 1989febsl.252 ... 42K . doi : 10.1016/0014-5793 (89) 80886-5 . PMID 2759231 . S2CID 36482110 .
- ^ Jump up to: а беременный Эльдра П. Соломон, Линда Р. Берг, Диана В. Мартин. Биология, 8 -е издание, международное студенческое издание . Томсон Брукс/Коул. ISBN 978-0495317142
- ^ Jump up to: а беременный Кларк, Дэвид П. (2005-06-24). Молекулярная биология . Elsevier . п. 134. ISBN 978-0-08-045421-4 .
- ^ «Стины ДНК» . www.sci.sdsu.edu . Архивировано с оригинала 27 октября 2017 года . Получено 1 мая 2018 года .
- ^ Berg J, The JL Tureer, Strier L (2006). Биохимики (6 -е изд.). Сан -Франциско: WH Freeman. Я 0-7167-8724-5 .
- ^ Jump up to: а беременный в дюймовый и фон глин Уотсон Д.Д., Бейкер Т.А., Белл С.П., Ганн А.А., Левин М., Лосик Р.М. (2013). Молекулярная биология гена (7 -е изд.). Пирсон. ISBN 978-0-321-76243-6 Полем OCLC 0321762436 .
- ^ Haberle V, Stark A (октябрь 2018 г.). «Эукариотические промоторы основного ядра и функциональная основа инициации транскрипции» . Nat Rev Mol Cell Biol . 19 (10): 621–637. doi : 10.1038/s41580-018-0028-8 . PMC 6205604 . PMID 29946135 .
- ^ Verheul TC, Van Hijfte L, Perenthaler E, Barakat TS (2020). «Почему YY1: механизмы регуляции транскрипции с Инь Ян 1» . Передняя клетка Dev Biol . 8 : 592164. DOI : 10.3389/fcell.2020.592164 . PMC 7554316 . PMID 33102493 .
- ^ Spitz F, Furlong EE (сентябрь 2012 г.). «Факторы транскрипции: от усиления энхансера с контролем развития». Nat Rev Genet . 13 (9): 613–26. doi : 10.1038/nrg3207 . PMID 22868264 . S2CID 205485256 .
- ^ Jump up to: а беременный Биган JA, Pastuzyn ED, Fernandez LR, Guo MH, Feng K, Titus KR, Chandrashekar H, Shepherd JD, Phillips-Cremins JE (июнь 2020 г.). «Трехмерная реструктуризация генома во времена экспрессии нейрональных генов, индуцированной активностью» . Nat Neurosci . 23 (6): 707–717. doi : 10.1038/s41593-020-0634-6 . PMC 7558717 . PMID 32451484 .
- ^ Jump up to: а беременный Schoenfelder S, Fraser P (август 2019 г.). «Контакты с энхансером дальнего действия в контроле экспрессии генов». Nat Rev Genet . 20 (8): 437–455. doi : 10.1038/s41576-019-0128-0 . PMID 31086298 . S2CID 152283312 .
- ^ Pennacchio LA, Bickmore W, Dean A, Nobrega MA, Bejerano G (апрель 2013 г.). «Улучшители: пять важных вопросов» . Nat Rev Genet . 14 (4): 288–95. doi : 10.1038/nrg3458 . PMC 4445073 . PMID 23503198 .
- ^ Weintraub AS, Li Ch, Zamudio AV, Sigova AA, Hannett NM, Day DS, Abraham BJ, Cohen MA, Nabet B, Buckley DL, Guo Ye, Hnisz D, Jaenisch R, Bradner Je, Grey NS, Young Ra (декабрь 2017 ) «YY1-структурный регулятор петлей-энхансер-промотера» . Клетка . 171 (7): 1573–88.e28. doi : 10.1016/j.cell.2017.11.008 . PMC 5785279 . PMID 29224777 .
- ^ Ламберт С.А., Джольма А., Кампителли Л.Ф., Дас П.К., Инь Й., Альбу М., Чен Х, Тайпале Дж., Хьюз Т.Р., Вейраух М.Т. (февраль 2018 г.). «Человеческие факторы транскрипции» . Клетка . 172 (4): 650–665. doi : 10.1016/j.cell.2018.01.029 . PMID 29425488 .
- ^ Jump up to: а беременный Гроссман С.Р., Энгрейц Дж., Рэй Дж.П., Нгуен Т. Т., Хакохен Н., Ландерс (июль 2018 г.). «Позиционная специфичность различных классов факторов транскрипции в усилителях» . Proc Natl Acad Sci USA . 115 (30): E7222–30. BIBCODE : 2018PNAS..115E7222G . doi : 10.1073/pnas.1804663115 . PMC 6065035 . PMID 29987030 .
- ^ Аллен Бл, Таатджес DJ (март 2015 г.). «Комплекс медиатора: центральный интегратор транскрипции» . Nat Rev Mol Cell Biol . 16 (3): 155–66. doi : 10.1038/nrm3951 . PMC 4963239 . PMID 25693131 .
- ^ Михайлихенко О., Бондаренко В., Харнетт Д., Шор И.Е., мужчины М., Виалес Р.Р., Фарлонг Е.Е. (январь 2018). «Степень энхансерной или промоторной активности отражается на уровнях и направленности транскрипции ERNA» . Гены Дев . 32 (1): 42–57. doi : 10.1101/gad.308619.117 . PMC 5828394 . PMID 29378788 .
- ^ Li QJ, Yang SH, Maeda Y, Sladek FM, Sharrocks AD, Martins-Green M (январь 2003 г.). «Зависимая от фосфорилирования MAP-киназа активация ELK-1 приводит к активации ко-активатора p300» . Embo j . 22 (2): 281–91. doi : 10.1093/emboj/cdg028 . PMC 140103 . PMID 12514134 .
- ^ Carullo NV, Phillips I RA, Simon RC, Soto SA, Hinds JE, Salisbury AJ, Revanna JS, Bunner KD, Ianov L, Sultan FA, Savell KE, Gersbach CA, Day JJ (сентябрь 2020). «Enhancer RNAs предсказывают регуляторные связи с энхансером-геном и имеют решающее значение для функции энхансер в нейрональных системах» . Нуклеиновые кислоты Res . 48 (17): 9550–70. doi : 10.1093/nar/gkaa671 . PMC 7515708 . PMID 32810208 .
- ^ Lövkvist C, Dodd IB, Sneppen K, Haerter Jo (июнь 2016 г.). «Метилирование ДНК в эпигеномах человека зависит от локальной топологии сайтов CPG» . Нуклеиновые кислоты Res . 44 (11): 5123–32. doi : 10.1093/nar/gkw124 . PMC 4914085 . PMID 26932361 .
- ^ Джаббари К., Бернарди Г (май 2004 г.). «Метилирование цитозина и CPG, TPG (CPA) и частоты TPA». Ген . 333 : 143–9. doi : 10.1016/j.gene.2004.02.043 . PMID 15177689 .
- ^ Steanhaus R, Gonzalez T, Seelow D, Robinson PN (июнь 2020 г.). «Распространенные и CPG-зависимые промоторные характеристики транскрибируемых энхансеров» . Нуклеиновые кислоты Res . 48 (10): 5306–17. doi : 10.1093/nar/gkaa223 . PMC 7261191 . PMID 32338759 .
- ^ Птица А (январь 2002 г.). «Паттерны метилирования ДНК и эпигенетическая память» . Гены Дев . 16 (1): 6–21. doi : 10.1101/gad.947102 . PMID 11782440 .
- ^ Jump up to: а беременный в Du Q, Luu PL, Stirzaker C, Clark SJ (2015). «Метил-CPG-связывающие доменные белки: читатели эпигенома» . Эпигеномика . 7 (6): 1051–73. doi : 10.2217/epi.15.39 . PMID 25927341 .
- ^ Vaquerizas JM, Kummerfeld SK, Teichmann SA, Luscombe NM (апрель 2009 г.). «Перепись факторов транскрипции человека: функция, экспрессия и эволюция». НАТ Преподобный Генет . 10 (4): 252–63. doi : 10.1038/nrg2538 . PMID 19274049 . S2CID 3207586 .
- ^ Jump up to: а беременный в дюймовый Sun Z, Xu X, He J, Murray A, Sun Ma, Wei X, Wang X, McCoig E, Xie E, Jiang X, Li L, Zhu J, Chen J, Morozov A, Pickrell AM, Theus MH, Xie H (Август 2019). «EGR1 рекрутирует TET1 для формирования метилома мозга во время развития и на нейрональной активности» . Nat Commun . 10 (1): 3892. Bibcode : 2019natco..10.3892S . doi : 10.1038/s41467-019-11905-3 . PMC 6715719 . PMID 31467272 .
- ^ Jump up to: а беременный Кубосаки А., Томару Ю., Тагами М., Арнер Е., Миура Х, Сузуки Т., Сузуки М., Сузуки Х, Хаясизаки Ю (2009). «Обще геном исследования сайтов связывания in vivo egr-1 в моноцитарной дифференцировке» . Геном биол . 10 (4): R41. doi : 10.1186/gb-2009-10-4-r41 . PMC 2688932 . PMID 19374776 .
- ^ Bayraktar G, Kreutz MR (апрель 2018 г.). «Нейрональная ДНК метилтрансферазы: эпигенетические медиаторы между синаптической активностью и экспрессией генов?» Полем Нейробиолог . 24 (2): 171–185. doi : 10.1177/10738584417707457 . PMC 5846851 . PMID 28513272 .
- ^ Oliveira AM, Hemstedt TJ, Bading H (июль 2012 г.). «Спасение снижения экспрессии DNMT3A2, связанного с старением, восстанавливает когнитивные способности». Nat Neurosci . 15 (8): 1111–3. doi : 10.1038/nn.3151 . PMID 22751036 . S2CID 10590208 .
- ^ Dhayalan A, Rajavelu A, Rathert P, Tamas R, Jurkowska RZ, Ragozin S, Jeltsch A (август 2010 г.). «Домен DNMT3A PWWP считывает гистон 3 лизин 36 триметилирование и направляет метилирование ДНК» . J Biol Chem . 285 (34): 26114–20. doi : 10.1074/jbc.m109.089433 . PMC 2924014 . PMID 20547484 .
- ^ Манзо М., Вирз Дж., Амбрози С., Вилласеньор Р., Рошицки Б., Баубек Т (декабрь 2017 г.). «Специфичная для изоформы локализация DNMT3A регулирует верность метилирования ДНК на двухвалентных островах CPG» . Embo j . 36 (23): 3421–34. doi : 10.15252/embj.201797038 . PMC 5709737 . PMID 29074627 .
- ^ Dukatz M, Holzer K, Choudalakis M, Imperle M, Lungu C, Bashtrykov P, Jeltsch A (декабрь 2019 г.). «Связывание H3K36ME2/3 и ДНК -связывание ДНК метилтрансферазы DNMT3A домен PWWP оба способствуют его взаимодействию хроматина». J Mol Biol . 431 (24): 5063–74. doi : 10.1016/j.jmb.2019.09.006 . PMID 31634469 . S2CID 204832601 .
- ^ Bayktar G, Yuanxang P, Confetture AD, Gomes GM, Raza SA, Stork O, Tajima S, Suetake I, Carpopova, Yildirim F, Kreutz MR (ноябрь 2020 г.). «Синаптический контроль ДНК-метилва-зависимой от деградации DNMT3A1 в ядре » Нейропсихофармакология 45 (12): 2120–3 Doi : 10.1038/s41386-020-0780-2 . PMC 7547096 . PMID 32726795
- ^ Jump up to: а беременный Пакай, Джулиан; Устройства, Хенрика; Шафи, Томас; Кларк, Кейтлин (2023). Трэш -концепции в биохимии . La Trobe Ebureau. doi : 10 26826/1017 . ISBN 978-0-6484681-9-6 Полем S2CID 258899183 .
- ^ Бутард, Магали (2016). «Глобальное перемещение участков начала транскрипции у бактерии для производства растений» . Природная связь . 7 : 13783. Bibcode : 2016natco ... 713783b . doi : 10.1038/ncomms13783 . PMC 5171806 . PMID 27982035 .
- ^ Редер, Роберт Дж. (1991). «Сложности инициации эукариотической транскрипции: регуляция сборки комплекса предварительного проживания». Тенденции в биохимических науках . 16 (11): 402–8. doi : 10.1016/0968-0004 (91) 90164-q . PMID 1776168 .
- ^ Goldman SR, Ebright RH , Nickels Be (май 2009 г.). «Прямое обнаружение абортивных транскриптов РНК in vivo» . Наука . 324 (5929): 927–8. Bibcode : 2009Sci ... 324..927G . doi : 10.1126/science.1169237 . PMC 2718712 . PMID 19443781 .
- ^ Revyakin A, Liu C, Ebright RH, Strick TR (ноябрь 2006 г.). «Абортивная инициация и продуктивная инициация РНК -полимеразой включают в себя заключение ДНК» . Наука . 314 (5802): 1139–43. Bibcode : 2006sci ... 314.1139R . doi : 10.1126/science.1131398 . PMC 2754787 . PMID 17110577 .
- ^ Raffaelle M, Kanin EI, Vogt J, Burgess RR, Ansari AZ (ноябрь 2005 г.). «Переключение голозгензимов и стохастическое высвобождение сигма -факторов из РНК -полимеразы in vivo» . Молекулярная клетка . 20 (3): 357–66. doi : 10.1016/j.molcel.2005.10.011 . PMID 16285918 .
- ^ Мандал С.С., Чу С, Вада Т., Ханда Х, Шаткин А.Дж., Рейнберг Д (май 2004 г.). «Функциональные взаимодействия фермента РНК-капитан с факторами, которые положительно и отрицательно регулируют выход промотора с помощью РНК-полимеразы II» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (20): 7572–7. Bibcode : 2004pnas..101.7572m . doi : 10.1073/pnas.0401493101 . PMC 419647 . PMID 15136722 .
- ^ Goodrich JA, Tjian R (апрель 1994 г.). «Факторы транскрипции IIE и IIH и АТФ гидролиз Прямой промотор с помощью РНК -полимеразы II». Клетка . 77 (1): 145–56. doi : 10.1016/0092-8674 (94) 90242-9 . PMID 8156590 . S2CID 24602504 .
- ^ Мило, Рон; Филипс, Роб (2015). «4. Показатели и продолжительность: Центральная догма: что быстрее: транскрипция или перевод?». Клеточная биология по числам . CRC Press. С. 231–6. ISBN 978-1-317-23069-4 Полем OCLC 1105558425 .
- ^ Ходжес С., Бинту Л., Любковская Л., Кашлев М., Бустаманте С (июль 2009 г.). «Нуклеосомные флуктуации управляют динамикой транскрипции РНК -полимеразы II» . Наука . 325 (5940): 626–8. Bibcode : 2009Sci ... 325..626H . doi : 10.1126/science.1172926 . PMC 2775800 . PMID 19644123 .
- ^ Jump up to: а беременный Fitz V, Shin J, Ehrlich C, Farnung L, Cramer P, Zaburdaev V, Grill SW (2016). «Нуклеосомное расположение влияет на динамику транскрипции с одной молекулой» . Труды Национальной академии наук . 113 (45): 12733–12738. BIBCODE : 2016PNAS..11312733F . doi : 10.1073/pnas.1602764113 . PMC 5111697 . PMID 27791062 .
- ^ Aymard F, Bugler B, Schmidt CK, Guillou E, Caron P, Briois S, Iacovoni JS, Daburon V, Miller KM, Jackson SP, Legube G (апрель 2014). «Транскрипционно активный хроматин рекрутирует гомологичную рекомбинацию при двойных разрывах ДНК» . Nat Struct Mol Biol . 21 (4): 366–74. doi : 10.1038/nsmb.2796 . PMC 4300393 . PMID 24658350 .
- ^ Ouyang J, Yadav T, Zhang JM, Yang H, Rheinbay E, Guo H, Haber Da, Lan L, Zou L (июнь 2021 г.). «РНК-транскрипты стимулируют гомологичную рекомбинацию путем образования петлей DR» . Природа . 594 (7862): 283–8. Bibcode : 2021natur.594..283o . doi : 10.1038/s41586-021-03538-8 . PMC 8855348 . PMID 33981036 .
- ^ García-Rubio M, Huertas P, González-Barrera S, Aguilera A (октябрь 2003 г.). «Рекомбиногенные эффекты агентов, повреждающих ДНК, синергически увеличиваются за счет транскрипции у Saccharomyces cerevisiae. Новое понимание рекомбинации, связанной с транскрипцией» . Генетика . 165 (2): 457–66. doi : 10.1093/Genetics/165.2.457 . PMC 1462770 . PMID 14573461 .
- ^ Ричардсон JP (сентябрь 2002 г.). «Rho-зависимое прекращение и АТФазы в прекращении транскрипта». Biochimica et Biophysica Acta (BBA) - структура и экспрессия гена . 1577 (2): 251–260. doi : 10.1016/s0167-4781 (02) 00456-6 . PMID 12213656 .
- ^ Lykke-Andersen S, Jensen TH (октябрь 2007 г.). «Перекрывающиеся пути определяют прекращение транскрипции РНК -полимеразы II». Биохими . 89 (10): 1177–82. doi : 10.1016/j.biochi.2007.05.007 . PMID 17629387 .
- ^ Ши, J; Вэнь, а; Чжао, м; Джин, с; Ты, л; Ши, y; Dong, s; Хуа, х; Чжан, y; Feng, Y (18 ноября 2020 года). «Структурная основа MFD-зависимого завершения транскрипции» . Исследование нуклеиновых кислот . 48 (20): 11762–11772. doi : 10.1093/nar/gkaa904 . PMC 7672476 . PMID 33068413 .
- ^ Цзян, y; Лю, м; Спенсер, Калифорния; Цена, DH (7 мая 2004 г.). «Вовлечение фактора завершения транскрипции 2 в митотической репрессии удлинения транскрипции» . Молекулярная клетка . 14 (3): 375–85. doi : 10.1016/s1097-2765 (04) 00234-5 . PMID 15125840 .
- ^ Маршалл, CJ; Qayyum, MZ; Уокер, JE; Мураками, KS; Santangelo, TJ (9 августа 2022 г.). «Структура и активность архейного фактора завершения транскрипции DETA детализируют уязвимости комплекса удлинения транскрипции» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (32): E22075881119. Bibcode : 2022pnas..11907581m . doi : 10.1073/pnas.2207581119 . PMC 9371683 . PMID 35917344 .
- ^ Cramer, P.; Armache, K.-J.; Baumli, S.; Benkert, S.; Брюкнер, Ф.; Бухен, C.; Дамсма, GE; Dengl, S.; Гейгер, ср; Jasiak, AJ; Jawhari, A. (июнь 2008 г.). «Структура эукариотических РНК -полимераз» . Ежегодный обзор биофизики . 37 (1): 337–352. doi : 10.1146/annurev.biophys.37.032807.130008 . PMID 18573085 .
- ^ http://www.sigmaaldrich.com/us/en/product/sial/h6878 8-гидроксихинолин от Sigma-Aldrich. Получено 2022-02-15
- ^ Datan E, Minn I, Peng X, He Ql, Ahn H, Yu B, Pomper Mg, Liu Jo (2020). «Конъюгат глюкозы-триптолида избирательно нацелен на раковые клетки при гипоксии» . ISCAING . 23 (9): 101536. Bibcode : 202020SICI ... 23J1536D . doi : 10.1016/j.isci.2020.101536 . PMC 7509213 . PMID 33083765 .
- ^ Саксонов С., Берг П., Брутлаг Д.Л. (январь 2006 г.). «Анализ общегенома CPG-динуклеотидов в человеческом геноме отличает два различных класса промоторов» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (5): 1412–7. Bibcode : 2006pnas..103.1412S . doi : 10.1073/pnas.0510310103 . PMC 1345710 . PMID 16432200 .
- ^ Птица А (январь 2002 г.). «Паттерны метилирования ДНК и эпигенетическая память» . Гены и развитие . 16 (1): 6–21. doi : 10.1101/gad.947102 . PMID 11782440 .
- ^ Vogelstein B, Papadopoulos N, Velculescu Ve, Zhou S, Diaz LA, Kinzler KW (март 2013 г.). «Ландшафты генома рака» . Наука . 339 (6127): 1546–58. BIBCODE : 2013SCI ... 339.1546V . doi : 10.1126/science.1235122 . PMC 3749880 . PMID 23539594 .
- ^ Tessitore A, Cicciarelli G, Del Vecchio F, Gaggiano A, Verzella D, Whistles M, Vecchotti D, Capece D, Zazzeroni F, Alese E (2014). «МикроРНК в сети повреждения/восстановления ДНК и рак» . Международный журнал геномики . 2014 : 820248. DOI : 10.1155/2014/820248 . PMC 3926391 . PMID 24616890 .
- ^ Я не уверен, что это Kohro S, Larkin JD, Deng B, Short P, Tsutsumi S, Taylor S, Kayki Y, Kayashi M, Li G, Poh HM, Ruan X, Aburatan H, Ruan Y, Kodama, Wada в, Cook PR (ноябрь 2012 г.). «TNFα сигнализирует через специализированные фабрики, где генов отзывчивого кодирования и miRNA подвергаются распространению » Embo Journal 31 (23): 4404–1 Citeseerx 10.1.1.1.919.1 Doi : 10.1038/ emboj.2012.2 PMC 3512387 PMID 23103767
- ^ «Химия 2006» . Нобелевский фонд . Архивировано из оригинала 15 марта 2007 года . Получено 29 марта 2007 г.
- ^ Wu, t (апрель 2020 г.). «Кетоксаль с помощью одноцепочечного секвенирования ДНК отражает глобальную динамику транскрипции и энхансерную активность in situ» . Природные методы . 17 (5): 515–523. doi : 10.1038/s41592-020-0797-9 . PMC 7205578 . PMID 32251394 . S2CID 214810294 .
- ^ Радж А., Ван Уденаарден А (октябрь 2008 г.). «Природа, воспитание или случайность: стохастическая экспрессия генов и его последствия» . Клетка . 135 (2): 216–26. doi : 10.1016/j.cell.2008.09.050 . PMC 3118044 . PMID 18957198 .
- ^ Кларк, Дэвид П. (2005-06-24). Молекулярная биология . Elsevier. п. 63. ISBN 978-0-08-045421-4 .
- ^ Колесникова в (2000). «Некоторые модели механизма апоптоза во время ВИЧ-инфекции» . Диссертация (на русском языке). Архивировано из оригинала 10 июля 2011 года . Получено 20 февраля 2011 года .
- ^ Cesare AJ, Reddel RR (май 2010 г.). «Альтернативное удлинение теломер: модели, механизмы и последствия». Nature Reviews Genetics . 11 (5): 319–30. doi : 10.1038/nrg2763 . PMID 20351727 . S2CID 19224032 .
Внешние ссылки
[ редактировать ]- Интерактивное моделирование Java инициации транскрипции. Архивировал 2011-07-22 в The Wayback Machine от Центра моделей жизни, архивировавших 2011-08-09 на машине Wayback в Институте Нильса Бора.
- Интерактивное моделирование Java по интерференции транскрипции - игра промотора доминирования в бактериальном вирусе. Архивированный 2011-08-26 в The Wayback Machine из Центра моделей жизни архивировал 2011-08-09 в The Wayback Machine в Институте Niels Bohr.
- Коллекция анимации виртуальных ячеек, представленная транскрипционная архивирована 2021-04-14 на машине Wayback