Т -клетка
| Т -клетка | |
|---|---|
 Анимация Т -клеток | |
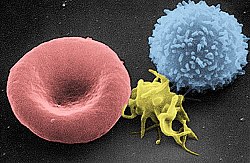 Сканирующая электронная микрофотография эритроцитов ( слева), тромбоцитов (центра) и Т -лимфоцита (справа); раскрашен | |
| Подробности | |
| Система | Иммунная система |
| Идентификаторы | |
| латинский | лимфоцит т |
| Сетка | D013601 |
| Тур | H2.00.04.1.02007 |
| FMA | 62870 |
| Анатомические термины микроанатомии | |
Т -клетки являются одним из важных типов лейкоцитов иммунной системы и играют центральную роль в адаптивном иммунном ответе . Т-клетки можно отличить от других лимфоцитов по присутствию Т-клеточного рецептора (TCR) на их клеточной поверхности .
Т -клетки рождаются из гематопоэтических стволовых клеток , [ 1 ] найдено в костном мозге . Развивающиеся Т -клетки затем мигрируют в тимусную железу для развития (или зрелого). Т -клетки получают свое название из тимуса . [ 2 ] [ 3 ] После миграции в тимус клетки -предшественники разразились в несколько различных типов Т -клеток. Дифференциация Т -клеток также продолжается после того, как они покинули тимус. Группы специфических, дифференцированных подтипов Т -клеток имеют множество важных функций в контроле и формировании иммунного ответа .
Одной из этих функций является иммун-опосредованная гибель клеток, и она осуществляется двумя основными подтипами: CD8+ «Killer» (Cytotoxic) и CD4+ «Helper» T-клетки. (Они названы в связи с присутствием белков клеточной поверхности CD8 или CD4 .) CD8+ T-клетки, также известные как «киллер-Т-клетки», являются цитотоксичными -это означает, что они способны непосредственно уничтожать инфицированные вирусом клетки, а также, а также раковые клетки. CD8+ T -клетки также способны использовать небольшие сигнальные белки, известные как цитокины , для рекрутирования других типов клеток при установке иммунного ответа. Другая популяция Т -клеток, CD4+ Т -клетки, функционируют как «помощники». В отличие от T -клеток CD8+ Killer T -клеток, клетки CD4+ Helper T (T T ) функционируют путем дальнейшего активации В -клеток памяти и цитотоксических Т -клеток, что приводит к большему иммунному ответу. Специфический адаптивный иммунный ответ, регулируемый T- C- клеткой, зависит от его подтипа (например, t-helper1, t-helper2, t-helper17, регуляторная Т-клетка), [ 4 ] который отличается типами цитокинов, которые они секретируют. [ 2 ]
Регуляторные Т -клетки являются еще одной отдельной популяцией Т -клеток, которые обеспечивают критический механизм толерантности , в результате чего иммунные клетки способны отличать вторжения в клетки от «самости». Это мешает иммунным клеткам ненадлежащего реагирования на свои собственные клетки, известные как « аутоиммунный » ответ. По этой причине эти регуляторные Т -клетки также были названы «супрессорами» Т -клетками. Эти же регуляторные Т-клетки также могут быть кооптированы раковыми клетками, чтобы предотвратить распознавание и иммунный ответ на опухолевые клетки.
Разработка
[ редактировать ]Происхождение, раннее развитие и миграция в тимус
[ редактировать ]Все Т-клетки происходят из c-kit + SCA1 + Гематопоэтические стволовые клетки (HSC), которые находятся в костном мозге. В некоторых случаях происхождение может быть печенью плода во время эмбрионального развития . Затем HSC дифференцируется в мультипотентные предшественники (MPP), которые сохраняют потенциал, чтобы стать как миелоидными , так и лимфоидными клетками . Затем процесс дифференцировки переходит к общему лимфоидному предшественнику (CLP), который может дифференцироваться только на T, B или NK -клетки. [ 5 ] Эти клетки CLP затем мигрируют через кровь в тимус, где они приживают : . Отныне они известны как тимоциты , незрелую стадию Т -клетки.
Самые ранние клетки, которые прибыли в тимус, обычно называются двойными отрицательными , поскольку они экспрессируют ни CD4 , ни CD8 . Недавно прибывшие клетки CLP являются CD4 − CD8 − CD44 + CD25 − CKIT + клетки и называются ранними клетками предшественника тимуса (ETP). [ 6 ] Затем эти клетки будут проходить раунд деления и понижать C-KIT и называются двойными отрицательными (DN1) клетками. Чтобы стать Т -клетками, тимоциты должны подвергаться множеству стадий DN, а также положительный отбор и отрицательный отбор.
Двойные отрицательные тимоциты могут быть идентифицированы с помощью поверхностной экспрессии CD2 , CD5 и CD7 . Тем не менее, на двойных отрицательных стадиях экспрессия CD34 останавливается и CD1 экспрессируется . Экспрессия как CD4, так и CD8 делает их двойными положительными , и превращается в CD4+ или CD8+ клетки.
Разработка TCR
[ редактировать ]Критическим этапом созревания Т -клеток является функциональный рецептор Т -клеток (TCR). Каждая зрелая Т -клетка в конечном итоге будет содержать уникальный TCR, который реагирует на случайную картину, позволяя иммунной системе распознавать множество различных типов патогенов . Этот процесс имеет важное значение для разработки иммунитета к угрозам, с которыми иммунная система не сталкивалась ранее, поскольку из -за случайных изменений всегда будет хотя бы один TCR, соответствующий любому новому патогену.
Тимоцит может стать активной Т -клетками только тогда, когда он переживает процесс разработки функционального TCR. TCR состоит из двух основных компонентов, альфа -и бета -цепей. Они оба содержат случайные элементы, предназначенные для создания широкого спектра различных TCR, но из -за этого огромного разнообразия они должны быть протестированы, чтобы убедиться, что они вообще работают. Во -первых, тимоциты пытаются создать функциональную бета -цепь, проверяя ее против «макетной» альфа -цепи. Затем они пытаются создать функциональную альфа -цепь. После того, как рабочий TCR был произведен, клетки должны проверить, правильно ли их TCR определит угрозы, и для этого необходимо распознать основной комплекс гистосовместимости организма (MHC) в процессе, известного как положительный отбор. Тимоцит также должен гарантировать, что он не реагирует негативно на «самог» , называемые негативным отбором. Если как положительный, так и отрицательный отбор успешны, TCR становится полностью эксплуатационным, а тимоцит становится Т -клеткой.
TCR β-цепь отбор
[ редактировать ]На стадии DN2 (CD44 + CD25 + ), клетки активируют гены рекомбинации RAG1 и RAG2 и переоценивают локус TCRβ , объединяя гены рекомбинации VDJ и постоянную область в попытке создать функциональную цепь TCRβ. По мере продвижения развивающихся тимоцитов на стадию DN3 (CD44 − CD25 + ), тимоцит экспрессирует инвариантную α-цепь, называемую Pre-Tα вместе с геном TCRβ. Если перестроенная β-цепь успешно сочетается с инвариантной α-цепи, образуются сигналы, которые перестают переставить β-цепь (и замолчать альтернативный аллель). [ 7 ] Хотя эти сигналы требуют предварительного TCR на клеточной поверхности, они не зависят от связывания лиганда с Pre-TCR. Если цепи успешно соединяют формы до TCR, а ячейка подавляет CD25 и называется ячейкой DN4 (CD25 − CD44 − ) Затем эти клетки подвергаются раунду пролиферации и начинают переоценивать локус TCRα на двойной положительной стадии.
Положительный выбор
[ редактировать ]Процесс положительного отбора занимает от 3 до 4 дней и происходит в коре тимуса. [ 8 ] Двойные тимоциты (CD4 + /CD8 + ) мигрируйте глубоко в кору тимуса , где они представлены с самогенами . Эти самоантигены экспрессируются с помощью эпителиальных клеток тимуса на молекулах MHC, которые находятся на поверхности корковых эпителиальных клеток. Только тимоциты, которые хорошо взаимодействуют с MHC-I или MHC-II, получат жизненно важный «сигнал выживания», в то время как те, которые не могут взаимодействовать достаточно сильно, не получат сигнал и умрут от пренебрежения. Этот процесс гарантирует, что выжившие тимоциты будут иметь «аффинность MHC», что означает, что они могут выполнять полезные функции в организме, отвечая на молекулы MHC, чтобы помочь иммунным ответам. Подавляющее большинство развивающихся тимоцитов не пройдут положительный отбор и умрет во время этого процесса. [ 9 ]
Судьба тимоцитов определяется во время положительного отбора. Двойные положительные клетки (CD4 + /CD8 + ) Это хорошо взаимодействует с молекулами MHC класса II, в конечном итоге станет CD4 + «помощники» клетки, тогда как тимоциты, которые хорошо взаимодействуют с молекулами MHC класса I, созревают в CD8 + «Убийца» клетки. Тимоцит становится CD4 + клетка путем подавляющей экспрессии рецепторов клеточной поверхности CD8. Если ячейка не потеряет свой сигнал, она будет продолжать понижать CD8 и стать CD4 + , оба CD8 + и CD4 + Клетки теперь являются одноположительными клетками. [ 10 ]
Этот процесс не фильтрует для тимоцитов, которые могут вызвать аутоиммунитет . Потенциально аутоиммунные клетки удаляются следующим процессом негативного отбора, который происходит в мозговом веществе тимуса.
Отрицательный отбор
[ редактировать ]Отрицательный отбор удаляет тимоциты, которые способны сильно связывать с «самостоятельными» молекулами MHC. Тимоциты, которые переживают положительный отбор, мигрируют в сторону границы коры и мозгового мозга в тимусе. Находясь в мозговом мозге, они снова представлены с самоантигеном, представленным на комплексе MHC медуллярных эпителиальных клеток тимуса (MTECS). [ 11 ] MTEC должны быть аутоиммунными регуляторами положительным (Aire + ) правильно экспрессировать самоантигены из всех тканей тела на их пептидах класса I MHC . Некоторые MTEC фагоцитозированы дендритными клетками тимуса ; Это делает их айре − Антиген, представляющие клетки (APC), позволяя представить самоантигены на молекулах MHC класса II (положительно выбранные CD4 + Клетки должны взаимодействовать с этими молекулами MHC класса II, таким образом, APC, которые обладают MHC Class II, должны присутствовать для CD4 + Т-клеточный отрицательный отбор). Тимоциты, которые слишком сильно взаимодействуют с самоантигеном, получают апоптотический сигнал, который приводит к гибели клеток. Однако некоторые из этих клеток выбираются, чтобы стать Treg клетками . Оставшиеся клетки выходят из тимуса в качестве зрелых наивных Т -клеток , также известных как недавние эмигранты тимуса. [ 12 ] Этот процесс является важным компонентом центральной толерантности и служит для предотвращения образования самореактивных Т-клеток, которые способны вызывать аутоиммунные заболевания у хозяина.
Сводка разработки TCR
[ редактировать ]β-выбор является первой контрольной точкой, где тимоциты, которые способны сформировать функциональную предварительную TCR (с инвариантной альфа-цепью и функциональной бета-цепью), позволяют продолжить развитие в тимусе. Затем, положительный отбор проверяет, что тимоциты успешно переставили свой локус TCRα и способны распознавать молекулы MHC с соответствующей сродством. Отрицательный отбор в мозге затем устраняет тимоциты, которые слишком сильно связываются с самоантигенами, экспрессируемыми на молекулах MHC. Эти процессы отбора позволяют иммунной системе устойчивость к себе. Типичные наивные Т-клетки, которые покидают тимус (через кортикомедлллярное соединение), являются самостоятельными, самоутоляющими и одинокими положительными.
Выход тимуса
[ редактировать ]Около 98% тимоцитов умирают во время процессов развития в тимусе, провалив либо положительный отбор, либо отрицательный отбор, тогда как остальные 2% выживают и оставляют тимус стать зрелыми иммунокомпетентными Т -клетками. [ 13 ] Тимус вносит меньше клеток в возрасте возраста. Поскольку тимус сокращается примерно на 3% [ 14 ] Год на протяжении всего среднего возраста происходит соответствующее падение при производстве тимусов наивных Т -клеток, оставляя периферические Т -клеточные расширения и регенерацию, чтобы играть большую роль в защите пожилых людей.
Типы Т -клеток
[ редактировать ]Т -клетки сгруппированы в серию подмножества на основе их функции. CD4 и CD8 T -клетки выбираются в тимусе, но подвергаются дальнейшей дифференцировке в периферии к специализированным клеткам, которые имеют разные функции. Подмножества Т -клеток первоначально определялись функцией, но также имеют связанные паттерны экспрессии генов или белка.
Обычные адаптивные Т -клетки
[ редактировать ]Helper CD4+ T -клетки
[ редактировать ]
Т -хелперские клетки (Т -клетки ) помогают другим лимфоцитам, включая созревание В -клеток в плазматические клетки и В -клетки памяти , а также активацию цитотоксических Т -клеток и макрофагов . Эти клетки также известны как CD4 + Т -клетки , когда они экспрессируют гликопротеин CD4 на своих поверхностях. Помощные Т-клетки активируются, когда они представлены пептидными антигенами молекулами MHC Class II , которые экспрессируются на поверхности антиген-презентативных клеток (APC). После активации они разделяют быстро и секретируют цитокины , которые регулируют или помогают иммунному ответу. Эти клетки могут дифференцироваться в один из нескольких подтипов, которые имеют разные роли. Цитокины направляют Т -клетки в конкретные подтипы. [ 15 ]
| Тип ячейки | Цитокины продуцируются | Ключевой фактор транскрипции | Роль в иммунной защите | Связанные заболевания |
|---|---|---|---|---|
| Th1 | IFNγ , IL-2 | TBET | Продуцируют воспалительный ответ, ключ к защите от внутриклеточных бактерий, вирусов и рака. | MS, диабет 1 типа |
| Th2 | IL-4, IL-5, IL-13 | Змея-3 | Иммунологически важен против внеклеточных патогенов, таких как инфекции червей | Астма и другие аллергические заболевания |
| Th17 | IL-17F, IL-17A, IL-22 | RORγt | Защита от кишечных патогенов и слизистых барьеров | MS, ревматоидный артрит, псориаз |
| Th9 [ 16 ] [ 17 ] | IL-9 | IRF4, PU.1 | Защита от гельминтов (паразитические черви) и клеточное аллергическое воспаление. | Рассеянный склероз |
| TFH | IL-21, IL-4 | BCL-6 | Помощь В -клетки продуцируют антитела | Астма и другие аллергические заболевания |
| Th22 [ 18 ] [ 17 ] | IL-22 | Ах | Патогенез аллергических заболеваний дыхательных путей и преимущественно противовоспалительных | Болезнь Крона, ревматоидный артрит, опухоли |
Цитотоксические CD8+ Т -клетки
[ редактировать ]
Цитотоксические Т-клетки (клетки T C , CTL, клетки T-Killer, Killer T-клетки) разрушают вирусные клетки и опухолевые клетки, а также участвуют в отторжении трансплантации . Эти клетки определяются экспрессией белка CD8 на их клеточной поверхности. Цитотоксические Т-клетки распознают их мишени путем связывания с короткими пептидами (8-11 аминокислот по длине), связанных с молекулами MHC класса I , присутствующими на поверхности всех зарожденных клеток. Цитотоксические Т-клетки также продуцируют ключевые цитокины IL-2 и IFNγ. Эти цитокины влияют на эффекторные функции других клеток, в частности макрофаги и NK -клетки.
Память Т -клетки
[ редактировать ]Т-клетки антигена расширяются и дифференцируются в память и эффекторные Т-клетки после того, как они сталкиваются с родственным антигеном в контексте молекулы MHC на поверхности профессионального антигена, представляющей клетку (например, дендритная клетка). Соответствующая костимуляция должна присутствовать во время встречи антигена для этого процесса. Исторически считалось, что Т -клетки памяти принадлежат либо подтипам эффекторной, либо центральной памяти, каждая из которых имеет свой собственный отличительный набор маркеров клеточной поверхности (см. Ниже). [ 19 ] Впоследствии были обнаружены многочисленные новые популяции T-клеток памяти, включая тканевые память T (TRM) памяти, клетки TSCM стволовой памяти и Т-клетки виртуальной памяти. Единственная объединяющая тема для всех подтипов Т-клеток памяти заключается в том, что они долгоживущие и могут быстро расширяться до большого количества эффекторных Т-клеток при повторном воздействии на их родственный антиген. По этому механизму они предоставляют иммунную систему «память» против ранее встречающихся патогенов. Т -клетки памяти могут быть или CD4 + или CD8 + и обычно выражают CD45RO . [ 20 ]
Память Т -ячейки подтипы:
- Центральные Т-клетки памяти (клетки T CM ) экспрессируют CD45RO, CC-хемокинового рецептора типа 7 (CCR7) и L-селектин (CD62L). Т -клетки центральной памяти также имеют промежуточную до высокой экспрессии CD44 . Эта субпопуляция памяти обычно встречается в лимфатических узлах и в периферической циркуляции. (Примечание- экспрессия CD44 обычно используется для отличия мышиной наивы от Т-клеток памяти).
- Эффекторные Т-клетки памяти (клетки T em и клетки T EMRA ) экспрессируют CD45RO, но не имеют экспрессии CCR7 и L-селектина . Они также имеют промежуточную или высокую экспрессию CD44 . У этих Т-клеток памяти не хватает рецепторов-лимфатических узлов и, таким образом, обнаружены в периферической циркуляции и тканях. [ 21 ] T EMRA означает терминально дифференцированные клетки эффекторной памяти, повторно экспрессирующие CD45RA, который является маркером, обычно обнаруживаемым на наивных Т-клетках. [ 22 ]
- Ткань-резидентные Т-клетки памяти (T RM ) занимают ткани (кожа, легкие и т. Д.) Без рециркуляции. Одним из маркеров клеточной поверхности, который был связан с TRM , является стажер αEβ7, также известный как CD103. [ 23 ]
- Т -клетки виртуальной памяти (T VM ) отличаются от других подмножеств памяти тем, что они не происходят после сильного клонального события расширения. Таким образом, хотя эта популяция в целом в изобилии в рамках периферической циркуляции, отдельные клоны Т -клеток виртуальной памяти находятся на относительно низких частотах. Одна теория заключается в том, что гомеостатическая пролиферация приводит к этой популяции Т -клеток. Хотя T -клетки CD8 виртуальной памяти были первыми, которые были описаны,, [ 24 ] Теперь известно, что CD4 виртуальной памяти также существуют. [ 25 ]
Регуляторные CD4+ Т -клетки
[ редактировать ]Регуляторные Т -клетки имеют решающее значение для поддержания иммунологической толерантности . Их основная роль заключается в том, чтобы закрыть, связанный с Т -клетками, иммунитет к концу иммунной реакции и подавлять аутореактивные Т -клетки , которые избежали процесса негативного отбора в тимусе.
Два основных класса CD4 + t -reg - Foxp3 Были описаны клетки + T -reg клетки и Foxp3 − Т -рег клетки.
Регуляторные Т -клетки могут развиваться либо во время нормального развития в тимусе, а затем известны как клетки Thymic TREG, либо могут быть индуцированы периферически и называются периферическими клетками Treg. Эти два подмножества ранее назывались «природными» и «адаптивными» (или «индуцированными») соответственно. [ 26 ] Оба подмножества требуют экспрессии транскрипционного фактора Foxp3 , который можно использовать для идентификации ячеек. Мутации гена FOXP3 могут предотвратить регуляторное развитие Т -клеток, вызывая фатальное аутоиммунное заболевание IPEX .
Несколько других типов Т -клеток обладают подавляющей активностью, но не экспрессируют конститутивно FOXP3. К ним относятся клетки TR1 и TH3 , которые, как считается, возникают во время иммунного ответа и действуют путем создания подавляющих молекул. Клетки TR1 связаны с IL-10, а клетки TH3 связаны с TGF-бета . В последнее время клетки Th17 были добавлены в этот список. [ 27 ]
Врожденные Т-клетки
[ редактировать ]Врожденные Т-клетки или нетрадиционные Т-клетки представляют некоторые подмножества Т-клеток, которые ведут себя по-разному при иммунитете. Они вызывают быстрые иммунные ответы, независимо от основной экспрессии комплекса гистосовместимости (MHC), в отличие от их обычных аналогов (CD4 T Helper Cltres и CD8 цитотоксических Т -клеток), которые зависят от распознавания пептидных антигенов в контексте молекулы MHC. В целом, существует три больших популяции нетрадиционных Т -клеток: клетки NKT, клетки MAIT и T -клетки гаммаделты. Теперь их функциональные роли уже хорошо создаются в контексте инфекций и рака. [ 28 ] Кроме того, эти подмножества Т -клеток переводятся во многие методы лечения против злокачественных новообразований, например, лейкемия. [ 29 ]
Т -клетка естественного убийцы
[ редактировать ]Естественные убийственные Т -клетки (клетки NKT - не следует путать с естественными клетками -убийцами врожденной иммунной системы) соединяют адаптивную иммунную систему с врожденной иммунной системой . В отличие от обычных Т -клеток, которые распознают белковые пептидные антигены, представленные основными молекулами комплекса гистосовместимости (MHC), клетки NKT распознают гликолипидные антигены, представленные CD1D . После активации эти клетки могут выполнять функции, приписываемые как вспомогательным, так и цитотоксическим Т -клеткам: продуцирование цитокинов и высвобождение цитолитических/клеточных молекул. Они также способны распознавать и устранять некоторые опухолевые клетки и клетки, инфицированные вирусами герпеса. [ 30 ]
Связанные слизистые инвариантные Т -клетки
[ редактировать ]Клетки, связанные с слизистой оболочкой (MAIT), демонстрируют врожденные , эффекторные качества. [ 31 ] [ 32 ] У людей клетки MAIT обнаруживаются в крови, печени, легких и слизистой оболочке , защищающих от микробной активности и инфекции. [ 31 ] Белок MHC -класса I, похожий на I, MR1 , отвечает за представление метаболитов витамина В бактериально продуцированных витамином В для клеток MAIT. [ 33 ] [ 34 ] [ 35 ] После представления иностранного антигена MR1 клетки MAIT секретируют провоспалительные цитокины и способны лизировать бактериально инфицированные клетки. [ 31 ] [ 35 ] Клетки MAIT также могут быть активированы с помощью MR1-независимой передачи сигналов. [ 35 ] В дополнение к областям врожденных функций, этот подмножество Т-клеток подтверждает адаптивный иммунный ответ и имеет фенотип, подобный памяти. [ 31 ] Кроме того, считается, что клетки MAIT играют роль в аутоиммунных заболеваниях , таких как рассеянный склероз , артрит и воспалительное заболевание кишечника , [ 36 ] [ 37 ] Хотя окончательные доказательства еще не опубликованы. [ 38 ] [ 39 ] [ 40 ] [ 41 ]
Гамма -дельта Т -клетки
[ редактировать ]Гамма -дельта -Т -клетки (γδ T -клетки) представляют собой небольшое подмножество Т -клеток, которое обладает γδ TCR, а не TCR αβ на клеточной поверхности. Большинство Т -клеток экспрессируют αβ -цепи TCR. Эта группа Т -клеток гораздо реже у людей и мышей (около 2% от общих Т -клеток) и встречается в основном в слизистой оболочке кишечника , в популяции внутриэпителиальных лимфоцитов . У кроликов, овец и цыплят количество γδ T -клеток может составлять до 60% от общих Т -клеток. Антигенные молекулы, которые активируют γδ T -клетки, все еще в основном неизвестны. Однако γδ T-клетки не ограничены MHC и, по-видимому, способны распознавать целые белки, а не требуют, чтобы пептиды были представлены молекулами MHC на APC . Некоторые мышиные γδ T -клетки распознают молекулы IB класса MHC. Человеческие γδ T -клетки, которые используют фрагменты гена Vγ9 и VΔ2, представляют собой основную популяцию γδ T -клеток в периферической крови. Эти клетки уникальны тем, что они специально и быстро реагируют на набор непептидических фосфорилированных предшественников изопреноидов , в совокупности названные фосфоантигены , которые продуцируются практически всеми живыми клетками. Наиболее распространенными фосфоантигенами из клеток животных и человека (включая раковые клетки) являются изопентенил пирофосфат (IPP) и его изомер -диметилалли -пирофосфат (DMPP). Многие микробы продуцируют активные соединения гидрокси-дмейп ( HMB-PP ) и соответствующие мононуклеотидные конъюгаты, в дополнение к IPP и DMAPP. Растительные клетки продуцируют оба типа фосфоантигенов. Препараты, активирующие человеческие Vγ9/VΔ2 T -клетки, составляют синтетические фосфоантигены и аминобисфосфонаты , которые активируют эндогенный IPP/DMAPP.
Активация
[ редактировать ]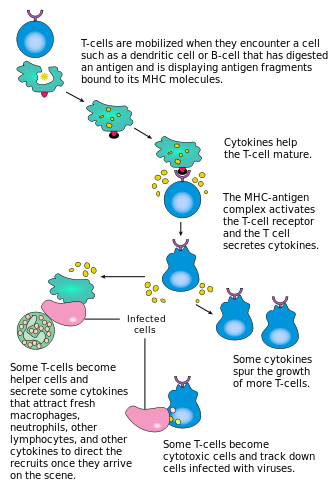
Активация CD4 + Т-клетки возникают благодаря одновременному взаимодействию Т-клеточного рецептора и костимулирующей молекулы (например, CD28 или ICO ) на Т-клетках с помощью основного комплекса гистосовместимости (MHCII) пептид и костимуляторных молекул на APC . Оба необходимы для производства эффективного иммунного ответа; В отсутствие костимуляции только передача сигналов рецептора Т-клеток приводит к анергии . Сигнальные пути вниз по течению от костимуляторных молекул обычно привлекают путь PI3K , генерирующий PIP3 в плазматической мембране и рекрутирующий домен pH, содержащий сигнальные молекулы, такие как PDK1 , которые необходимы для активации PKC-θ , и возможного IL-2 производства . Оптимальный CD8 + Ответ Т -клеток зависит от CD4 + сигнализация. [ 43 ] CD4 + Клетки полезны в начальной антигенной активации наивных CD8 T -клеток и поддерживающей CD8 памяти + Т -клетки после острой инфекции. Следовательно, активация CD4 + Т -клетки могут быть полезны для действия CD8 + Т -клетки. [ 44 ] [ 45 ] [ 46 ]
Первый сигнал предоставляется путем связывания рецептора Т -клеток с его родственным пептидом, представленным на MHCII на APC. MHCII ограничен так называемыми профессиональными антиген-презентативными клетками , такими как дендритные клетки, В-клетки и макрофаги, чтобы назвать некоторые из них. Пептиды представлены CD8 + Т -клетки по молекулам класса I MHC имеют длину 8–13 аминокислот; Пептиды представлены CD4 + клетки молекул MHC класса II длиннее, обычно 12–25 аминокислот в длину, [ 47 ] В качестве концов связывания расщелины молекулы MHC класса II открыты.
Второй сигнал исходит от костимуляции, в которой поверхностные рецепторы на APC индуцируются относительно небольшим количеством стимулов, обычно продуктов патогенов, но иногда прорывные продукты клеток, такие как некротические белки или белки теплового шока . Единственным костимуляторным рецептором, экспрессируемым конститутивно наивными Т-клетками, является CD28, поэтому костимуляция этих клеток поступает из белков CD80 и CD86 , которые вместе составляют белок B7 (B7.1 и B7.2, соответственно) на Апк Другие рецепторы экспрессируются при активации Т -клеток, таких как OX40 и ICO, но они в значительной степени зависят от CD28 для их экспрессии. Второй сигнал лицензирует Т Т -клетку для реагирования на антиген. Без этого Т -клетки становится аноргическим , и в будущем становится все труднее активировать. Этот механизм предотвращает неуместные реакции на себя, поскольку самопептиды обычно не представляются подходящей костимуляцией. После того, как Т -клетка была надлежащим образом активирована (т.е. получил сигнал один и сигнал два), она изменяет экспрессию клеточной поверхности различных белков. Маркеры активации Т-клеток включают CD69, CD71 и CD25 (также маркер для клеток TREG) и HLA-DR (маркер активации Т-клеток человека). Экспрессия CTLA-4 также активируется на активированных Т-клетках, что, как выясняется, OutCompetes CD28 для связывания с белками B7. Это механизм контрольно -пропускного пункта для предотвращения активации Т -клеток. Активированные Т -клетки также изменяют их профиль гликозилирования клеточной поверхности. [ 48 ]
Рецептор Т -клеток существует как комплекс из нескольких белков. Фактический рецептор Т -клеток состоит из двух отдельных пептидных цепей, которые продуцируются из независимых генов альфа -рецептора Т -клеток и бета ( TCRα и TCRβ ). Другими белками в комплексе являются белки CD3 : гетеродимеры CD3εγ и CD3εΔ и, что наиболее важное, гомодимер CD3ζ, который имеет в общей сложности шесть мотивов ITAM . Мотивы ITAM на CD3ζ могут быть фосфорилированы LCK и, в свою очередь, рекрутируют ZAP-70 . LCK и/или ZAP-70 также могут фосфорилировать тирозины во многих других молекулах, не менее важных CD28, LAT и SLP-76 , что позволяет агрегации сигнальных комплексов вокруг этих белков.
Фосфорилированный LAT- рекрутирует SLP-76 в мембрану, где он может затем внести PLC-γ , VAV1 , ITK и потенциально PI3K . PLC-γ расщепляет PI (4,5) P2 на внутренней листовке мембраны, чтобы создать активные посредники диацилглицерина ( DAG ), инозитол-1,4,5-трипласфат ( IP3 ); PI3K также действует на PIP2, фосфорилируя его, чтобы получить фосфатидинозитол-3,4,5-трисфосфат (PIP3). DAG связывает и активирует некоторые PKC. Наиболее важным в Т-клетках является PKC-θ, критический для активации факторов транскрипции NF-κB и AP-1. IP3 высвобождается из мембраны с помощью PLC-γ и быстро диффундирует для активации рецепторов кальциевых каналов на ER , что индуцирует высвобождение кальция в цитозоль. Низкий кальций в эндоплазматической ретикулуме вызывает кластеризацию STIM1 на мембране ER и приводит к активации каналов CRAC клеточной мембраны, которые позволяют дополнительному кальция течь в цитозоль из внеклеточного пространства. Этот агрегированный цитозольный кальций связывает кальмодулин, который затем может активировать кальциневрин . Кальциневрин, в свою очередь, активирует NFAT , который затем транслоцируется в ядро. NFAT является фактором транскрипции , который активирует транскрипцию плейотропного набора генов, наиболее заметных, IL-2, цитокина, который способствует долгосрочной пролиферации активированных Т-клеток.
PLC-γ также может инициировать путь NF-κB . DAG активирует PKC-θ, который затем фосфорилирует CARMA1, заставляя его разворачиваться и функционировать как каркас. Цитозольные домены связывают адаптер BCL10 через домены карты (активация и домены каспазы); Это затем связывает TRAF6, который убиквитинируется в K63. : 513–523 [ 49 ] Эта форма убиквитинирования не приводит к деградации целевых белков. Скорее, это служит для привлечения Немо, IKKα и -β и Tab1-2/ Tak1. [ 50 ] TAK 1 Фосфорилирует IKK-β, который затем фосфорилирует IκB, позволяя убиквитинировать K48: приводит к протеасомной деградации. REL A и P50 могут затем войти в ядро и связывать элемент отклика NF-κB. Это в сочетании с передачей сигналов NFAT допускает полную активацию гена IL-2. [ 49 ]
Хотя в большинстве случаев активация зависит от распознавания антигена TCR, были описаны альтернативные пути активации. Например, было показано, что цитотоксические Т -клетки активируются, когда нацелены на другие CD8 T -клетки, ведущие к толеризации последних. [ 51 ]
Весной 2014 года на Международной космической станции была запущена эксперимент по активации Т-клеток в космосе (TCAS) на международной космической станции на миссии SpaceX CRS-3, чтобы изучить, как «дефицит в иммунной системе человека влияет на среду микрогравитации». [ 52 ]
Активация Т -клеток модулируется реактивными формами кислорода . [ 53 ]
Дискриминация антигена
[ редактировать ]Уникальной особенностью Т -клеток является их способность различать здоровые и аномальные (например, инфицированные или раковые) клетки в организме. [ 54 ] Здоровые клетки обычно экспрессируют большое количество самого полученного PMHC на их клеточной поверхности, и, хотя рецептор антигена Т -клеток может взаимодействовать, по крайней мере, с подмножеством этих самостоятельных PMHC, Т -клетка обычно игнорирует эти здоровые клетки. Однако, когда эти те же клетки содержат даже минутные величины PMHC, полученного патогеном, Т -клетки способны активироваться и инициировать иммунные ответы. Способность Т -клеток игнорировать здоровые клетки, но реагируют, когда эти же клетки содержат патоген (или рак), полученный PMHC, известен как дискриминация антигена. Молекулярные механизмы, лежащие в основе этого процесса, противоречивы. [ 54 ] [ 55 ]
Клиническое значение
[ редактировать ]Дефицит
[ редактировать ]
Причины дефицита Т -клеток включают лимфоцитопению Т -клеток и/или дефекты на функции отдельных Т -клеток. Полная недостаточность функции Т -клеток может быть результатом наследственных условий, таких как тяжелый комбинированный иммунодефицит (SCID), синдром Омня и гипоплазию хряща - хартиаж . [ 56 ] Причины частичной недостатки функции Т-клеток включают синдром приобретенного иммунного дефицита (СПИД) и наследственные условия, такие как синдром Дигжорджа (DGS), синдромы хромосомального разрушения В-клеток и Т-клеточные комбинированные расстройства (CBS), а также В-клеточные комбинированные расстройства, такие как атаксия-телагиэктозия (AT и ) и синдром Вискотта -Альдрича (был). [ 56 ]
Основными патогенными микроорганизмами в дефиците Т -клеток являются внутриклеточные патогенные микроорганизмы , включая вирус простого герпеса , микобактерии и листерию . [ 57 ] Кроме того, грибковые инфекции также более распространены и тяжелые по дефициту Т -клеток. [ 57 ]
Рак
[ редактировать ]Рак Т-клеток называется Т-клеточной лимфомой и объясняет, возможно, каждый из десяти случаев неходжкинской лимфомы . [ 58 ] Основными формами Т -клеточной лимфомы являются:
- Экстранодальная Т -клеточная лимфома
- Кожные Т -клеточные лимфомы : синдром Сезари и грибов микоза
- Анапластическая большая клеточная лимфома
- Ангиоиммунобластическая Т -клеточная лимфома
Истощение
[ редактировать ]Было высказано предположение, что этот раздел будет разделен на другую статью под названием «Истощение Т -клеток» . ( Обсудить ) (май 2023 г.) |
Истощение Т -клеток является плохо определенным или неоднозначным термином. [ 59 ] [ 60 ] Есть три подхода к его определению. [ 59 ] "Первый подход в первую очередь определяет как истощенные клетки, которые представляют ту же клеточную дисфункцию (как правило, отсутствие ожидаемого эффекторного отклика). Второй подход в первую очередь определяет как истощенные клетки, которые вызываются данной причиной (обычно, но не обязательно не обязательно. , хроническое воздействие антигена). [ 59 ]
Дисфункциональные Т -клетки характеризуются прогрессивной потерей функции, изменениями в профилях транскрипции и устойчивой экспрессией ингибирующих рецепторов. Сначала клетки теряют свою способность продуцировать IL-2 и TNFα , после чего следует потеря высокой пролиферативной способности и цитотоксический потенциал, и в конечном итоге приводит к их делеции. Источенные Т -клетки обычно указывают на более высокие уровни CD43 , CD69 и ингибирующих рецепторов в сочетании с более низкой экспрессией CD62L и CD127 . Истощение может развиваться во время хронических инфекций, сепсиса и рака. [ 61 ] Источенные Т -клетки сохраняют их функциональное истощение даже после повторного воздействия антигена. [ 62 ]
Во время хронической инфекции и сепсиса
[ редактировать ]Истощение Т -клеток может быть вызвано несколькими факторами, такими как постоянное воздействие антигена и отсутствие помощи CD4 Т -клеток. [ 63 ] Воздействие антигена также влияет на ход истощения, потому что более длительное время воздействия и более высокая вирусная нагрузка увеличивает тяжесть истощения Т -клеток. Для установления истощения требуется не менее 2–4 недель. [ 64 ] Другим фактором, способным индуцировать истощение, являются ингибирующие рецепторы, включая запрограммированный белок гибели клеток (PD1), белок-3 CTLA-4 , T-клеточный мембранный белок-3 (TIM3) и белок активации лимфоцитов (LAG3). [ 65 ] [ 66 ] Растворимые молекулы, такие как цитокины IL-10 или TGF-β, также способны вызывать истощение. [ 67 ] [ 68 ] Последние известные факторы, которые могут играть роль в истощении Т -клеток, являются регуляторными клетками. Клетки Treg могут быть источником IL-10 и TGF-β, и поэтому они могут играть роль в истощении Т-клеток. [ 69 ] Кроме того, истощение Т -клеток возвращается после истощения клеток Treg и блокады PD1. [ 70 ] Истощение Т -клеток также может происходить во время сепсиса в результате шторма цитокинов. Позже после того, как начальная септическая встреча противовоспалительных цитокинов и проапоптотических белков вступают во владение для защиты организма от повреждения. Сепсис также несет высокую нагрузку на антиген и воспаление. На этой стадии исчерпания Т -клеток увеличивается истощение Т -клеток. [ 71 ] [ 72 ] В настоящее время есть исследования, направленные на использование ингибирующих рецепторных блокад при лечении сепсиса. [ 73 ] [ 74 ] [ 75 ]
Во время трансплантации
[ редактировать ]В то время как во время инфекционного истощения Т -клеток может развиться после постоянного воздействия антигена после трансплантата трансплантата, схожая ситуация возникает при наличии аллоантигена. [ 76 ] Было показано, что ответ Т -клеток уменьшается с течением времени после пересадки почки. [ 77 ] Эти данные предполагают, что истощение Т -клеток играет важную роль в толерантности к трансплантату, главным образом, путем истощения аллореактивных CD8 T -клеток. [ 72 ] [ 78 ] Несколько исследований показали положительное влияние хронической инфекции на принятие трансплантата и его долгосрочную выживаемость, частично опосредованную источником Т-клеток. [ 79 ] [ 80 ] [ 81 ] Также было показано, что истощение реципиента Т -клеток обеспечивает достаточные условия для переноса NK -клеток . [ 82 ] Хотя есть данные, показывающие, что индукция истощения Т -клеток может быть полезной для трансплантации, это также несет недостатки, среди которых можно подсчитать увеличение числа инфекций и риск развития опухоли. [ 83 ]
Во время рака
[ редактировать ]Во время рака Т -клеток истощение играет роль в защите опухоли. Согласно исследованию некоторые связанные с раком клеток, а также сами опухолевые клетки могут активно индуцировать истощение Т-клеток в месте опухоли. [ 84 ] [ 85 ] [ 86 ] Истощение Т -клеток также может играть роль в рецидиве рака, как было показано на лейкемии. [ 87 ] Некоторые исследования показали, что можно предсказать рецидив лейкоза на основе экспрессии ингибирующих рецепторов PD-1 и TIM-3 с помощью Т-клеток. [ 88 ] Многие эксперименты и клинические испытания были сосредоточены на блокаторах иммунных контрольно -пропускных пунктов при терапии рака, причем некоторые из них одобрены как действительные методы лечения, которые в настоящее время находятся в клиническом использовании. [ 89 ] Ингибирующие рецепторы, нацеленные на эти медицинские процедуры, жизненно важны при истощении Т -клеток, и блокировка их может изменить эти изменения. [ 90 ]
Смотрите также
[ редактировать ]- Т -клетка химерного антигена рецептор
- Специфичный для кишечника
- Иммунобласт
- Иммунозвука
- Парафолликулярная ячейка, также называемая C -клеткой
Ссылки
[ редактировать ]- ^ «5. гемопоэтические стволовые клетки» . Информация о стволовых клетках . Bethesda, MD: Национальные институты здравоохранения, Министерство здравоохранения и социальных служб США. 17 июня 2001 года. Архивировано с оригинала 29 октября 2016 года . Получено 21 декабря 2021 года .
- ^ Jump up to: а беременный Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Хелпер -Т -клетки и активация лимфоцитов» . Молекулярная биология клетки (4 -е изд.). Гарлендская наука.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Хелпер -Т -клетки и активация лимфоцитов» . Молекулярная биология клетки (4 -е изд.). Гарлендская наука. п. 1367.
Т -клетки ... Выводят их [имя] из органов, в которых они развиваются. Т -клетки развиваются [зрелый] в тимусе
- ^ Luckheeram RV, Zhou R, Verma AD, Xia B (2012). «CD4⁺T -клетки: дифференциация и функции» . Клиническая иммунология и иммунология развития . 2012 : 925135. DOI : 10.1155/2012/925135 . PMC 3312336 . PMID 22474485 .
- ^ Кондо М (декабрь 2016 г.). «Одна ниша, чтобы управлять как обслуживанием, так и потерей стебля в HSC» . Иммунитет . 45 (6): 1177–1179. doi : 10.1016/j.immuni.2016.12.003 . PMID 28002722 .
- ^ Osborne LC, Dhanji S, Snow JW, Priatel JJ, MA MC, Miners MJ, et al. (Март 2007 г.). «Нарушение памяти CD8 T-клеток и первичные ответы CD4 T-клеток у Alpha Mite IL-7R» . Журнал экспериментальной медицины . 204 (3): 619–631. doi : 10.1084/jem.20061871 . PMC 2137912 . PMID 17325202 .
- ^ Мерфи, Кеннет (2011). Иммунобиология Джейнвея (8 -е изд.). Гарлендская наука. С. 301–305. ISBN 9780815342434 .
- ^ Росс Джо, Меличар Х.Дж., Ау-Юн Б.Б., Герцмарк П., Вайс А., Роби Э.А. (июнь 2014 г.). «Отдельные фазы в положительном отборе CD8+ T-клеток, отличающиеся от интратимической миграции и рецептора T-клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (25): E2550 - E2558. BIBCODE : 2014PNAS..111E2550R . doi : 10.1073/pnas.1408482111 . PMC 4078834 . PMID 24927565 .
- ^ Starr TK, Jameson SC, Hogquist KA (2003-01-01). «Положительный и отрицательный отбор Т -клеток». Ежегодный обзор иммунологии . 21 (1): 139–176. doi : 10.1146/annurev.immunol.21.120601.141107 . PMID 12414722 .
- ^ Zerrahn J, Held W, Raulet DH (март 1997 г.). «Реакционная способность MHC репертуара Т -клеток перед положительным и отрицательным отбором» . Клетка . 88 (5): 627–636. doi : 10.1016/s0092-8674 (00) 81905-4 . PMID 9054502 . S2CID 15983629 .
- ^ Hinterberger M, Aichinger M, Prazeres Da Costa O, Voehringer D, Hoffmann R, Klein L (июнь 2010 г.). «Автономная роль медуллярных эпителиальных клеток тимуса в центральной толерантности к Т -клеткам CD4 (+)» (PDF) . Природа иммунология . 11 (6): 512–519. doi : 10.1038/ni.1874 . PMID 20431619 . S2CID 33154019 .
- ^ Pekalski ML, García AR, Ferreira RC, Rainbow DB, Smyth DJ, Mashar M, Brady J, Savinykh N, Dopico XC, Mahmood S, Duley S, Stevens HE, Walker NM, Cutler AJ, Waldron-Lynch F, Dunger DB, Shannon-Lowe C, Coles AJ, Jones JL, Wallace C, Todd JA, Wicker LS (август 2017 г.). «Недавние эмигранты из неонатальных и взрослых производят IL-8 и экспрессируют рецепторы комплемента CR1 и CR2» . JCI Insight . 2 (16). doi : 10.1172/jci.insight.93739 . PMC 5621870 . PMID 28814669 .
- ^ Мерфи, Кеннет (2011). Иммунобиология Джейнвея (8 -е изд.). Гарлендская наука. п. 297. ISBN 9780815342434 .
- ^ Haynes BF, Markert ML, Sempowski GD, Patel DD, Hale LP (2000). «Роль тимуса в иммунном восстановлении в старении, трансплантации костного мозга и инфекции ВИЧ-1». Анну. Преподобный иммунол . 18 : 529–560. doi : 10.1146/annurev.immunol.18.1.529 . PMID 10837068 .
- ^ Gutcher I, Becher B (2007). «Цитокины, полученные из APC и поляризация Т-клеток при аутоиммунном воспалении» . J. Clin. Инвестировать 117 (5): 1119–27. doi : 10.1172/jci31720 . PMC 1857272 . PMID 17476341 .
- ^ Wang W, Sung N, Gilman-Sachs A, Kwak-Kim J (18 августа 2020 г.). «Th Helper (TH) профили клеток при беременности и рецидивирующих потери беременности: TH1/TH2/TH9/TH17/TH22/TFH» . Границы в иммунологии . 11 : 2025. DOI : 10.3389/fimmu.2020.02025 . PMC 7461801 . PMID 32973809 .
- ^ Jump up to: а беременный Саравия Дж., Чепмен Н.М., Чи Х (июль 2019 г.). «Помощная дифференциация Т -клеток» . Клеточная и молекулярная иммунология . 16 (7): 634–643. doi : 10.1038/s41423-019-0220-6 . PMC 6804569 . PMID 30867582 .
- ^ Jia L, Wu C (2014). «Биология и функции клеток Th22». T Helper Cell Дифференциация и их функция . Достижения в области экспериментальной медицины и биологии. Тол. 841. С. 209–230. doi : 10.1007/978-94-017-9487-9_8 . ISBN 978-94-017-9486-2 Полем PMID 25261209 .
- ^ Саллусто Ф., Лениг Д., Фёрстер Р., Липп М., Ланзавеккия А (1999). «Два подмножества Т -лимфоцитов памяти с различными потенциалами самонинга и эффекторными функциями». Природа . 401 (6754): 708–712. Bibcode : 1999natur.401..708s . doi : 10.1038/44385 . PMID 10537110 . S2CID 4378970 .
- ^ Акбар А.Н., Терри Л., Тиммс А., Беверли ПК, Яносси Г. (апрель 1988 г.). «Потеря CD45R и усиление реакционной способности UCHL1 являются особенностью первичных Т -клеток» . J. Immunol . 140 (7): 2171–8. doi : 10.4049/jimmunol.140.7.2171 . PMID 2965180 . S2CID 22340282 .
- ^ Willinger T, Freeman T, Hasegawa H, McMichael AJ, Callan MF (2005). «Молекулярные сигнатуры отличают центральную память человека от подмножеств с эффекторной памятью CD8 T -клеток» (PDF) . Журнал иммунологии . 175 (9): 5895–903. doi : 10.4049/jimmunol.175.9.5895 . PMID 16237082 . S2CID 16412760 .
- ^ Koch S, Larbi A, Derhovanessian E, Ozcelik D, Naumova E, Pawelec G (2008). «Многопараметрический проточный цитометрический анализ подмножества CD4 и CD8 T -клеток у молодых и пожилых людей» . Иммунитет и старение . 5 (6): 6. doi : 10.1186/1742-4933-5-6 . PMC 2515281 . PMID 18657274 .
- ^ Shin H, Iwasaki A (сентябрь 2013 г.). «Ткань-резидентные Т-клетки памяти» . Иммунологические обзоры . 255 (1): 165–81. doi : 10.1111/imr.12087 . PMC 3748618 . PMID 23947354 .
- ^ Lee YJ, Jameson SC, Hogquist Ka (2011). «Альтернативная память в линии CD8 T -клеток» . Тенденции в иммунологии . 32 (2): 50–56. doi : 10.1016/j.it.2010.12.004 . PMC 3039080 . PMID 21288770 .
- ^ Marusina Ai, Ono Y, Merleev AA, Shimoda M, Ogawa H, Wang EA, Kondo K, Olney L, Luxardi G, Miyamura Y, Yilma TD, Villalobos IB, Bergstrom JW, Kronenberg DG, Soulika Am, Adamopoulos IE, Maverakis e E E E. (2017). «CD4+ виртуальная память: Т-клетки с антигеном, привязанными к наивным, регуляторным и памятникам Т-клеток на аналогичных частотах, последствия для аутоиммунитета» . Журнал аутоиммунитета . 77 : 76–88. doi : 10.1016/j.jaut.2016.11.001 . PMC 6066671 . PMID 27894837 .
- ^ Я Аббас, Бенуист С., Блюстоун Дж.А., Кэмпбелл Д.Дж., Гош С., Хори С., Джи С., Кубо В.К., Матис Д., Ронкороло М.Г., Руденский А., Сагагучи С., Шевач Э.М., Вигнали Д.А., Зеглер С.Ф. (2013). «Регуляторные Т -клетки: рекомендации по упрощению номенклатуры » НАТ Немедленно 14 (4): 307–8 Doi : 10.1038/ ni.2 PMID 23507634 . S2CID 11294516
- ^ Singh B, Schwartz JA, Sandrock C, Bellemore SM, Nikoopour E (2013). «Модуляция аутоиммунных заболеваний с помощью интерлейкина (IL) -17, продуцирующих регуляторные клетки T-Helper (TH17)» . Индиан Дж. Мед. Резерв 138 (5): 591–4. PMC 3928692 . PMID 24434314 .
- ^ Годфри Ди, Ульдрих А.П., МакКласки Дж., Россджон Дж., Муди Д.Б. (ноябрь 2015). «Растущее семейство нетрадиционных Т -клеток». Природа иммунология . 16 (11): 1114–1123. doi : 10.1038/ni.3298 . PMID 26482978 . S2CID 30992456 .
- ^ De Araújo ND, Gama FM, De Souza Barros M, Ribeiro TL, Alves FS, Xabregas LA, et al. (2021). «Перевод нетрадиционных Т -клеток и их роль в противоопухолевом иммунитете лейкемии» . Журнал иммунологических исследований . 2021 : 6633824. DOI : 10.1155/2021/6633824 . PMC 7808823 . PMID 33506055 .
- ^ Mallevaey T, Fontaine J, Breuilh L, Paget C, Castro-Keller A, Vendeville C, Capron M, Leite-De-Moraes M, Trottein F, Faveeuw C (май 2007 г.). «Инвариантные и неинвариантные Т -клетки с естественным убийцей оказывают противоположные регуляторные функции на иммунном ответе во время мышиного шистосомоза» . Инфекция и иммунитет . 75 (5): 2171–80. doi : 10.1128/iai.01178-06 . PMC 1865739 . PMID 17353286 .
- ^ Jump up to: а беременный в дюймовый Napier RJ, Adams EJ, Gold MC, Lewinsohn DM (2015-07-06). «Роль инвариантных Т -клеток, связанных с слизистой оболочкой в антимикробном иммунитете» . Границы в иммунологии . 6 : 344. DOI : 10.3389/fimmu.2015.00344 . PMC 4492155 . PMID 26217338 .
- ^ Gold MC, Lewinsohn DM (август 2011 г.). «Связанные слизистые инвариантные Т -клетки и иммунный ответ на инфекцию» . Микробы и инфекция . 13 (8–9): 742–8. doi : 10.1016/j.micinf.2011.03.007 . PMC 3130845 . PMID 21458588 .
- ^ Экл С.Б., Корбетт А.Дж., Келлер А.Н., Чен З., Годфри Ди, Лю Л., Мак Дж.Ю., Фэйрли Д.П., Россджон Дж., МакКласки Дж. (Декабрь 2015). «Распознавание предшественников витамина B и побочных продуктов с помощью инвариантных Т -клеток, связанных с слизистой оболочкой» . Журнал биологической химии . 290 (51): 30204–11. doi : 10.1074/jbc.r115.685990 . PMC 4683245 . PMID 26468291 .
- ^ Ussher JE, Klenerman P, Willberg CB (2014-10-08). «Инвариантные Т-клетки, связанные с слизистой оболочкой: новые игроки в антибактериальном иммунитете» . Границы в иммунологии . 5 : 450. DOI : 10.3389/fimmu.2014.00450 . PMC 4189401 . PMID 25339949 .
- ^ Jump up to: а беременный в Howson LJ, Salio M, Cerundolo V (2015-06-16). «Инвариантные Т-клетки, ограниченные слизистой оболочкой, ограниченные MR1, и их активация во время инфекционных заболеваний» . Границы в иммунологии . 6 : 303. DOI : 10.3389/fimmu.2015.00303 . PMC 4468870 . PMID 26136743 .
- ^ Hinks TS (май 2016 г.). «Инвариантные Т-клетки, связанные с слизистой оболочкой, в аутоиммунитете, иммунно-опосредованных заболеваниях и болезнях дыхательных путей» . Иммунология . 148 (1): 1–12. doi : 10.1111/imm.12582 . PMC 4819138 . PMID 26778581 .
- ^ Bianchini E, De Biasi S, Simone AM, Ferraro D, Sola P, Cossarizza A, Pinti M (март 2017 г.). «Инвариантные естественные убийственные Т-клетки и инвариантные Т-клетки, связанные с слизистой оболочкой, при рассеянном склерозе». Иммунологические письма . 183 : 1–7. doi : 10.1016/j.imlet.2017.01.009 . PMID 28119072 .
- ^ Serriari NE, Eoche M, Lamotte L, Lion J, Fumery M, Marcelo P, Chalatin D, Bar A, Nguyen-Khac E, Lantz OR, Dupas JL, Treiner и (May 2014). «Врожденные клетки, связанные с слизистой оболочкой (MAIT), активируются при воспаленных заболеваниях кишечника» . Клиническая и экспериментальная иммунология . 176 (2): 266–74. Doi : 10.1111/cei.12277 . PMC 3992039 . PMID 24450998 .
- ^ Huang S, Martin E, Kim S, Yu L, Soudais C, Fremont DH, Lantz O, Hansen TH (май 2009 г.). «Презентация антигена MR1 на инвариантные Т-клетки, ассоциированные с слизистой оболочкой, была высоко консервативна в эволюции» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (20): 8290–5. Bibcode : 2009pnas..106.8290h . doi : 10.1073/pnas.0903196106 . PMC 2688861 . PMID 19416870 .
- ^ Chua WJ, Hansen TH (ноябрь 2010 г.). «Бактерии, ассоциированные с слизистой оболочкой инвариантные Т-клетки и MR1» . Иммунология и клеточная биология . 88 (8): 767–9. doi : 10.1038/icb.2010.104 . PMID 20733595 . S2CID 27717815 .
- ^ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Хайраллах Г.Н., Годфри Д.П., Фэйрли Д.П., Россджон Дж., МакКласки Дж. (Ноябрь 2012 г.). «MR1 представляет микробные метаболиты витамина В в клетки MAIT» (PDF) . Природа . 491 (7426): 717–23. Bibcode : 2012natur.491..717k . doi : 10.1038/nature11605 . PMID 23051753 . S2CID 4419703 .
- ^ Бланша NIAID для ресурсов «Понимание иммунной системы (PDF)» .
- ^ Уильямс М.А., Беван М.Дж. (2007-01-01). «Эффектор и память CTL дифференциация». Ежегодный обзор иммунологии . 25 (1): 171–92. doi : 10.1146/annurev.immunol.25.022106.141548 . PMID 17129182 .
- ^ Janssen EM, Lemmens EE, Wolfe T, Christen U, Von Herrath MG, Schoenberger SP (февраль 2003 г.). «CD4+ T -клетки необходимы для вторичного расширения и памяти в CD8+ T -лимфоцитах». Природа . 421 (6925): 852–6. Bibcode : 2003natur.421..852j . doi : 10.1038/nature01441 . PMID 12594515 . S2CID 574770 .
- ^ Shedlock DJ, Shen H (апрель 2003 г.). «Требование для CD4 T -клеток помогает в создании функциональной памяти CD8 T -клеток». Наука . 300 (5617): 337–9. Bibcode : 2003sci ... 300..337S . doi : 10.1126/science.1082305 . PMID 12690201 . S2CID 38040377 .
- ^ Sun JC, Williams MA, Bevan MJ (сентябрь 2004 г.). «CD4+ T -клетки необходимы для поддержания, а не для программирования, памяти CD8+ T -клеток после острой инфекции» . Природа иммунология . 5 (9): 927–33. doi : 10.1038/ni1105 . PMC 2776074 . PMID 15300249 .
- ^ Дженнифер Ролланд и Робин О'Хехир, «Отключение Т -клеток: пептиды для лечения аллергических заболеваний», Todayship Life Science Publishing, 1999, стр. 32
- ^ Maverakis E, Kim K, Shimoda M, Gershwin M, Patel F, Wilken R, Raychaudhuri S, Ruhaak LR, Lebilla CB (2015). «Гликаны в иммунной системе и измененная теория аутоиммунитета гликана» . J Autoimmun . 57 (6): 1–13. doi : 10.1016/j.jaut.2014.12.002 . PMC 4340844 . PMID 25578468 .
- ^ Jump up to: а беременный Tatham P, Gomperts BD, Kramer IM (2003). Сигнальная трансдукция . Амстердам: Elsevier Academic Press. ISBN 978-0-12-289632-3 .
- ^ Wu H, Arron Jr (ноябрь 2003 г.). «TRAF6, молекулярный мост, охватывающий адаптивный иммунитет, врожденный иммунитет и остеоиммунология». Биологии . 25 (11): 1096–105. doi : 10.1002/bies.10352 . PMID 14579250 . S2CID 28521713 .
- ^ Милштейн О., Хагин Д., Ласк А., Рейх-Зелигер С., Шезен Е., Офир Е., Эйдельштейн Ю., Афик Р., Антеби Ю, Дастин М.Л., Рейснер Ю (январь 2011 г.). «CTL реагируют с активацией и секрецией гранул при служении в качестве мишеней для распознавания Т -клеток» . Кровь . 117 (3): 1042–52. doi : 10.1182/blood-2010-05-283770 . PMC 3035066 . PMID 21045195 .
- ^ Грэм В. (2014-04-14). «SpaceX, готов к запуску Dragon CRS-3 и новых вех» . Nasaspaceflight.com . Получено 2014-04-14 .
- ^ Беликов А.В., Шравен Б., Симеони Л (октябрь 2015 г.). «Т -клетки и реактивные формы кислорода» . Журнал биомедицинской науки . 22 : 85. DOI : 10.1186/S12929-015-0194-3 . PMC 4608155 . PMID 26471060 .
- ^ Jump up to: а беременный Feinerman O, Germain RN, Altan-Bonnet G (2008). «Количественные проблемы в понимании дискриминации лиганда алфавитными Т -клетками» . Мол Иммунол . 45 (3): 619–31. doi : 10.1016/j.molimm.2007.03.028 . PMC 2131735 . PMID 17825415 .
- ^ Душек О., Ван дер Мерве Па (2014). «Индуцированная модель антигена дискриминации антигена» . Тенденции Иммунол . 35 (4): 153–8. doi : 10.1016/j.it.2014.02.002 . PMC 3989030 . PMID 24636916 .
- ^ Jump up to: а беременный Т-клеточные расстройства в эмедицине
- ^ Jump up to: а беременный Джонс Дж., Баннистер Б.А., Гиллеспи Ш., ред. (2006). Инфекция: микробиология и управление . Wiley-Blackwell. п. 435. ISBN 978-1-4051-2665-6 .
- ^ «Лимфомы» (PDF) . Общество лейкемии и лимфомы. Май 2006 г. с. 2 Получено 2008-04-07 .
- ^ Jump up to: а беременный в Kaminski H, Lemoine M, Pradeu T (сентябрь 2021 г.). «Иммунологическое истощение: как сделать разрозненную концепцию в эксплуатации?» Полем PLO -патогены . 17 (9): E1009892. doi : 10.1371/journal.ppat.1009892 . PMC 8460019 . PMID 345555119 .
- ^ Blank Cu, Haining WN, Held W, Hogan PG, Kallies A, Lugli E, et al. (Ноябрь 2019). «Определение« истощения Т -клеток » » . Природные обзоры. Иммунология . 19 (11): 665–674. doi : 10.1038/s41577-019-0221-9 . PMC 7286441 . PMID 31570879 .
- ^ Yi JS, Cox MA, Zajac AJ (апрель 2010 г.). «Истощение Т-клеток: характеристики, причины и конверсия» . Иммунология . 129 (4): 474–81. doi : 10.1111/j.1365-2567.2010.03255.x . PMC 2842494 . PMID 20201977 .
- ^ Wang Q, Pan W, Liu Y, Luo J, Zhu D, Lu Y, Feng X, Yang X, Dittmer U, Lu M, Yang D, Liu J (2018). «Специфичные для вируса гепатита В CD8+ Т-клетки поддерживают функциональное истощение после повторного воздействия антигена в острого иммунной среды активации» . Передний иммунол . 9 : 219. doi : 10.3389/fimmu.2018.00219 . PMC 5816053 . PMID 29483916 .
- ^ Matloubian M, Concepcion RJ, Ahmed R (декабрь 1994 г.). «CD4+ T-клетки необходимы для поддержания CD8+ цитотоксических Т-клеточных реакций во время хронической вирусной инфекции» . Журнал вирусологии . 68 (12): 8056–63. doi : 10.1128/JVI.68.12.8056-8063.1994 . PMC 237269 . PMID 7966595 .
- ^ Angelosanto JM, Blackburn SD, Crawford A, Wherry EJ (август 2012 г.). «Прогрессирующая потеря памяти Т -клеток и приверженность истощению во время хронической вирусной инфекции» . Журнал вирусологии . 86 (15): 8161–70. doi : 10.1128/jvi.00889-12 . PMC 3421680 . PMID 22623779 .
- ^ Wherry EJ (июнь 2011 г.). «Истощение Т -клеток». Природа иммунология . 12 (6): 492–9. doi : 10.1038/ni.2035 . PMID 21739672 . S2CID 11052693 .
- ^ Окгава Т., Коннай С., Нишимори А., Майкева Н., Гото С., Икебучи Р., Кохара Дж., Сузуки Ю., Ямада С., Като Ю., Мурата С., Охаси К (июнь 2018 г.). «+ Т -клетки во время вирусной инфекции бычьего лейкоза » Ветеринарное исследование 49 (1): 50. doi : 10.1186/ s13567-018-018-0543-9 6006750PMC PMID 29914540
- ^ Брукс Д.Г., Трифило М.Дж., Эдельманн К.Х., Тейтон Л., МакГаверн Д.Б., Олдстоун М.Б. (ноябрь 2006 г.). «Интерлейкин-10 определяет вирусное разрешение или упорство в vivo» . Природная медицина . 12 (11): 1301–9. doi : 10.1038/nm1492 . PMC 2535582 . PMID 17041596 .
- ^ Tinoco R, Alcalde V, Yang Y, Sauer K, Zunga EI (июль 2009 г.). Специфический CD8 + Иммунитет 31 (1): 145–57. doi : 10.1016/j.mmuni . PMC 3039716 . PMID 19604493 .
- ^ Вейга-Парга Т., Сехрават С., Рауз Б.Т. (сентябрь 2013 г.). «Роль регуляторных Т -клеток во время вирусной инфекции» . Иммунологические обзоры . 255 (1): 182–96. doi : 10.1111/imr.12085 . PMC 3748387 . PMID 23947355 .
- ^ Penaloza-Macmaster P, Kamphorst AO, Wieland A, Araki K, Iyer SS, West EE, O'Mara L, Yang S, Konieczny BT, Sharpe AH, Freeman GJ, Rudensky AY, Ahmed R (август 2014). «Взаимодействие между регуляторными Т-клетками и PD-1 в модулировании истощения Т-клеток и вирусного контроля во время хронической инфекции LCMV» . Журнал экспериментальной медицины . 211 (9): 1905–18. doi : 10.1084/jem.20132577 . PMC 4144726 . PMID 25113973 .
- ^ Otto GP, Sossdorf M, Claus RA, Rödel J, Menge K, Reinhart K, Bauer M, Riedemann NC (июль 2011 г.). «Поздняя фаза сепсиса характеризуется повышенной микробиологической бремя и смертностью» . Интенсивная помощь . 15 (4): R183. doi : 10.1186/cc10332 . PMC 3387626 . PMID 21798063 .
- ^ Jump up to: а беременный Boomer JS, K, Chang KC, Takasu O, Osborne DF, Walton AH, Bricker TL, Jarman SD, Kreisel D, Krupnick AS, Srivastava A, Swanson PE, Green JM, Hotchkiss RS (декабрь 2011). «Иммуносупрессия у пациентов, которые умирают от сепсиса и множественной органной недостаточности» . Джама . 306 (23): 2594–605. doi : 10.1001/Jama.2011.1829 . PMC 3361243 . PMID 22187279 .
- ^ Shindo Y, McDonough JS, Chang KC, Ramachandra M, Sasikumar PG, Hotchkiss RS (февраль 2017 г.). «Пептид анти-PD-L1 улучшает выживаемость при сепсисе» . Журнал хирургических исследований . 208 : 33–39. doi : 10.1016/j.jss.2016.08.099 . PMC 5535083 . PMID 27993215 .
- ^ Patera AC, Drewry AM, Chang K, Beiter ER, Osborne D, Hotchkiss RS (декабрь 2016 г.). «Фронтальная наука: дефекты иммунной функции у пациентов с сепсисом связаны с экспрессией PD-1 или PD-L1 и могут быть восстановлены антителами, нацеленными на PD-1 или PD-L1» . Журнал биологии лейкоцитов . 100 (6): 1239–1254. doi : 10.1189/jlb.4hi0616-255r . PMC 5110001 . PMID 27671246 .
- ^ Wei Z, Li P, Yao Y, Deng H, Yi S, Zhang C, Wu H, Xie X, Xia M, He R, Yang XP, Tang ZH (июль 2018). «Альфа-лактоза меняет повреждение печени через блокаду TIM-3-опосредованного апоптоза CD8 при сепсисе». Клиническая иммунология . 192 : 78–84. doi : 10.1016/j.clim.2018.04.010 . PMID 29689313 . S2CID 21657071 .
- ^ Уэллс AD, Li XC, Strom TB, Turka LA (май 2001 г.). «Роль периферической делеции Т-клеток в толерантности к трансплантации» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 356 (1409): 617–23. doi : 10.1098/rstb.2001.0845 . PMC 1088449 . PMID 11375065 .
- ^ Halloran PF, Chang J, Famulski K, Hidalgo LG, Salazar ID, Merino Lopez M, Matas A, Picton M, De Freitas D, Bromberg J, Serón D, Sellarés J, Einecke G, Reeve J (июль 2015 г.). «Исчезновение, опосредованное Т-клетками, несмотря на продолжающееся антитело-опосредованное отторжение у реципиентов поздней трансплантации почки» . Журнал Американского общества нефрологии . 26 (7): 1711–20. doi : 10.1681/asn.2014060588 . PMC 4483591 . PMID 25377077 .
- ^ Steger U, Denecke C, Sawitzki B, Karim M, Jones ND, Wood KJ (май 2008 г.). «Исчерпывающая дифференциация аллореактивных CD8+ T -клеток: критическая для определения принятия трансплантата или отказа» (PDF) . Трансплантация . 85 (9): 1339–47. doi : 10.1097/tp.0b013e31816dd64a . PMID 18475193 . S2CID 33409478 .
- ^ Marre-Brede Il, Shi Xll, Mancham S, of r, heide-muser M, пердун, небесный MH, молодой J, двадцати LJ, Consure of LJ, Check J-j (август 2015 г.). «Цитом-индуцированная галовирусом экспрессия или CD244 от транс-плантации печени связаны с белыми CD8+ T-клетками гипозивируют Allares» . Журнал или иммунология . 195 (4): 1838–48. doi : 10 4049/jimmunol.1500440 . PMID 26170387 .
- ^ Гасса А., Цзянь Ф., Калкаван Х., Дюхан В., Хонке Н., Шаабани Н., Фридрих С.К., Дольф С., Валерс Т., Криббен А., Хардт С, Ланг П.А., Витцке О., Ланг К.С. (2016). «IL-10 индуцирует истощение Т-клеток во время трансплантации вирусов, инфицированных сердец» . Клеточная физиология и биохимия . 38 (3): 1171–81. doi : 10.1159/000443067 . PMID 26963287 .
- ^ Shi XL, de Mare-Bredemeijer EL, Tapirdamaz ö, Hansen Be, Van Gent R, Van Campenhout MJ, Mancham S, Litjens NH, Betjes MG, Van der Eijk AA, Xia Q, Van der Laan LJ, De Jonge J, Metselaar HJ, Kwekkeboom J (сентябрь 2015 г.). «Первичная инфекция ЦМВ связана с донорской гипореактивностью Т-клеток и поздними острыми отклонением после трансплантации печени» . Американский журнал трансплантации . 15 (9): 2431–42. Doi : 10.1111/ajt.13288 . PMID 25943855 . S2CID 5348557 .
- ^ Уильямс Р.Л., Кули С., Баханова В., Блазар Б.Р., Вайсдорф DJ, Миллер Дж. С., Вернерис г -н (март 2018 г.). «Истощение реципиента Т -клеток и успешный приемный перенос гаплоидноиментальных природных клеток -убийц» . Биология крови и трансплантации костного мозга . 24 (3): 618–622. doi : 10.1016/j.bbmt.2017.11.022 . PMC 5826878 . PMID 29197679 .
- ^ Woo Sr, Turnis ME, Goldberg MV, Bankoti J, Selby M, Nurschl CJ, Bettini ML, Gravano DM, Volu CL, Tangsombatvisit S, Grosso JF, Netto G, Smeltzer MP, Chaux A, Utz PJ, Workman CJ, Pardoll DM, Smeltzer , Korman AJ, Drake CG, Vignali DA (февраль 2012 г.). «Иммунные ингибирующие молекулы LAG-3 и PD-1 Синергически регулируют функцию Т-клеток, чтобы способствовать выходу опухолевого иммунного иммунитета » РАНКА 72 (4): 917–2 Doi : 10.1158/0008-5472.can- 11-1 3288154PMC 22186141PMID
- ^ Zelle-Rieser C, Thangavadivel S, Biedermann R, Brunner A, Stoitzner P, Willenbacher E, et al. (Ноябрь 2016). «Т -клетки при множественной миеломе демонстрируют особенности истощения и старения в месте опухоли» . Журнал гематологии и онкологии . 9 (1): 116. doi : 10.1186/s13045-016-0345-3 . PMC 5093947 . PMID 27809856 .
- ^ Лакинс М.А., Горани Е., Мунир Х., Мартинс К.П., Шилдс Д.Д. (март 2018 г.). «Фибробласты, связанные с раком, индуцируют антиген-специфическую делецию CD8 + Т-клетки для защиты опухолевых клеток » . Природная связь . 9 (1): 948. Bibcode : ... 9..948L . DOI : 10.1038/S41467-018-03347-0 . PMC 5838096. 2018natco PMID 29507342 .
- ^ Concorti L (февраль 2012 г.). «Сеть ионных каналов в Т -лимфоцитах, цель иммунотерапии». Клиническая иммунология . 142 (2): 105–106. doi : 10.1016/j.clim.2011.11.009 . PMID 22189042 .
- ^ Лю Л., Чанг Ю.Дж., Сюй Л.П., Чжан Х.Х., Ван Ю., Лю К.Ю., Хуан XJ (май 2018). «Истощение Т -клеток, характеризующееся скомпрометированной MHC класса I и II, ограниченной цитотоксической активности, связанной с острым B -лимфобластным рецидивом лейкоза после аллогенной гематопоэтической трансплантации стволовых клеток». Клиническая иммунология . 190 : 32–40. doi : 10.1016/j.clim.2018.02.009 . PMID 29477343 .
- ^ Kong Y, Zhang J, Claxton DF, Ehmann WC, Rybka WB, Zhu L, et al. (Июль 2015). «ПД-1 (HI) TIM-3 (+) Т-клетки ассоциируются с рецидивом лейкемии у пациентов с ОМЛ после аллогенной трансплантации стволовых клеток» . Журнал «Рак крови» . 5 (7): E330. doi : 10.1038/bcj.2015.58 . PMC 4526784 . PMID 26230954 .
- ^ «US FDA одобрило ингибиторы иммунной проверки и иммунотерапию» . Агентство медицинских писателей | 香港醫學作家 | Medipr | Медипапер Гонконг . 2018-08-21 . Получено 2018-09-22 .
- ^ Bhadra R, Gigley JP, Weiss LM, Khan IA (май 2011 г.). «Контроль реактивации токсоплазмы путем спасения дисфункционального ответа CD8+ T-клеток посредством блокады PD-1-PDL-1» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (22): 9196–9201. Bibcode : 2011pnas..108.9196b . doi : 10.1073/pnas.1015298108 . PMC 3107287 . PMID 21576466 .
Дальнейшее чтение
[ редактировать ]- Janeway Jr CA, Travers P, Walport M, Shlomchik MJ (2001). Иммунобиология 5: иммунная система в отношении здоровья и заболеваний (5 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-3642-6 .
- «Иммунная система» (PDF) . Национальный институт аллергии и инфекционных заболеваний . Сентябрь 2003 года. Архивировано из оригинала (PDF) 25 июня 2009 года.