Рибосома
| Клеточная биология | |
|---|---|
| Диаграмма животных | |
 Компоненты типичной животной клетки:
|
Рибосомы ( / ˈ r aɪ b ə z oʊ m , - s oʊ m / ) - это макромолекулярные машины , обнаруженные во всех клетках , которые выполняют синтез биологического белка ( Мессенджера РНК трансляция ). Рибосомы связывают аминокислоты вместе в порядке, указанном кодонами молекул РНК мессенджера с образованием полипептидных цепей. Рибосомы состоят из двух основных компонентов: маленьких и крупных рибосомных субъединиц. Каждая субъединица состоит из одной или нескольких рибосомных молекул РНК и многих рибосомных белков ( R-белков ). [ 1 ] [ 2 ] [ 3 ] Рибосомы и связанные с ними молекулы также известны как трансляционный аппарат .
Обзор
[ редактировать ]Последовательность ДНК , которая кодирует последовательность аминокислот в белке, транскрибируется в цепь РНК (мРНК) мессенджера (мРНК). Рибосомы связываются с РНКсенджером и используют их последовательности [ нужно разъяснения ] Чтобы определить правильную последовательность аминокислот для генерации данного белка. Аминокислоты выбираются и переносятся в рибосому с помощью молекул переноса РНК (тРНК), которые попадают в рибосому и связываются с цепью РНК мессенджера через петлю антикодонного стебля. Для каждой кодирующей триплета ( кодона ) в РНК мессенджера существует уникальная трансферная РНК, которая должна иметь точное антикодонское совпадение и несет правильную аминокислоту для включения в растущую полипептидную цепь. Как только белок получен, он может затем сложить , чтобы получить функциональную трехмерную структуру.
Рибосома сделана из комплексов РНК и белков и, следовательно, является комплексом рибонуклеопротеина . В прокариотах каждая рибосома состоит из небольших (30 с ) и больших (50 с ) компонентов, называемых субъединицами, которые связаны друг с другом:
- (30S) имеет в основном функцию декодирования и также связана с мРНК
- (50S) имеет в основном каталитическую функцию и также связана с аминоацилированными тРНК.
Синтез белков из их строительных блоков происходит в четырех этапах: инициация, удлинение, прекращение и переработка. Начальный кодон во всех молекулах мРНК имеет последовательность AUG. Стоп -кодон - один из UAA, UAG или UGA; Поскольку нет молекул тРНК, которые распознают эти кодоны, рибосома распознает, что трансляция завершена. [ 4 ] Когда рибосома заканчивает чтение молекулы мРНК, две субъединицы отделяются и обычно разбиты, но могут быть использованы повторно. Рибосомы представляют собой своего рода фермент , называемый рибозимами , потому что активность каталитической пептидилтрансферазы , которая связывает аминокислоты вместе, выполняется рибосомальной РНК. [ 5 ]
В эукариотических клетках рибосомы часто ассоциируются с внутриклеточными мембранами, которые составляют грубую эндоплазматическую ретикулуму .
Рибосомы из бактерий , археи и эукариот (в системе трех доменов ) напоминают друг друга в замечательной степени, свидетельством общего происхождения. Они различаются по размеру, последовательности, структуре и соотношению белка к РНК. Различия в структуре позволяют некоторым антибиотикам убивать бактерии, ингибируя их рибосомы, оставляя человеческие рибосомы незатронутыми. У всех видов более одной рибосомы может двигаться по одной цепи мРНК за один раз (как полисома ), каждый «чтение» специфическую последовательность и создавая соответствующую молекулу белка.
Митохондриальные рибосомы эукариотических клеток отличаются от других их рибосом. Они функционально напоминают бактерии, отражая эволюционное происхождение митохондрий как эндосимбиотические бактерии. [ 6 ] [ 7 ]
Открытие
[ редактировать ]Рибосомы впервые наблюдались в середине 1950-х годов румын-американским биологом клеток Джорджа Эмиля , с использованием электронного микроскопа в качестве плотных частиц или гранул. [ 8 ] Первоначально их называли паладными гранулами из -за их гранулированной структуры. Термин «рибосома» был предложен в 1958 году Говардом М. Динцисом: [ 9 ]
В течение симпозиума стала очевидной семантическая сложность. Для некоторых участников «микросомы» означают частицы рибонуклеопротеина микросом -фракции, загрязненной другим белком и липидным материалом; Для других микросомы состоят из белка и липидов, загрязненных частицами. Фраза «микросомальные частицы» не кажутся адекватными, и «частицы рибонуклеопротеинов фракции микросом» слишком неловко. Во время встречи было предложено слово «рибосома», которое имеет очень удовлетворительное имя и приятный звук. Нынешняя путаница была бы устранена, если бы «рибосома» была принята для обозначения частиц рибонуклеопротеина в размерах в диапазоне от 35 до 100 лет.
- Альберт Клод, микросомальные частицы и синтез белка [ 10 ]
Альберт Клод , Кристиан де Дув и Палада Джорджа Эмиля были совместно награждены Нобелевской премией по физиологии или медицине в 1974 году за открытие рибосомы. [ 11 ] Нобелевская премия по химии 2009 была присуждена Венкатраману Рамакришнану , Томасу А. Стейцу и Аде Э. Йонату за определение детальной структуры и механизма рибосомы. [ 12 ]
Структура
[ редактировать ]

Рибосома - это сложная клеточная машина. Он в основном состоит из специализированной РНК, известной как рибосомная РНК (рРНК), а также десятки различных белков (точное число немного варьируется между видами). Рибосомные белки и rRNAs расположены в два различных рибосомальных кусочков разных размеров, известных, как правило, как крупные и маленькие субъединицы рибосомы. Рибосомы состоят из двух субъединиц, которые соединяются друг с другом и работают как одна, чтобы транслировать мРНК в полипептидную цепь во время синтеза белка. Поскольку они образуются из двух субъединиц неравственного размера, они немного длиннее на оси, чем в диаметре.
Прокариотические рибосомы
[ редактировать ]Прокариотические рибосомы имеют диаметр около 20 нм (200 Å ) в диаметре и состоят из 65% рРНК и 35% рибосомных белков . [ 13 ] Эукариотические рибосомы имеют диаметр от 25 до 30 нм (250–300 Å) с соотношением рРНК к белкам, которое близко к 1. [ 14 ] Кристаллографическая работа [ 15 ] показал, что нет рибосомных белков, близких к месту реакции для синтеза полипептида. Это говорит о том, что белковые компоненты рибосом непосредственно не участвуют в катализе формирования пептидных связей, а скорее, что эти белки действуют как каркас, который может повысить способность рРНК синтезировать белок (см. Рибозим ).
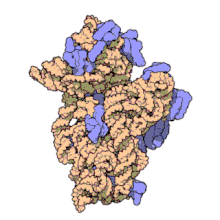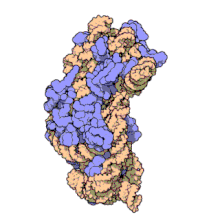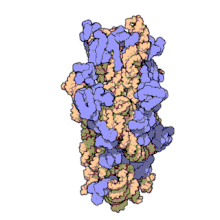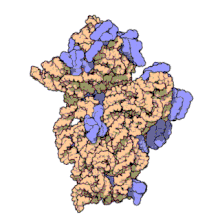
Рибосомные субъединицы прокариот и эукариот очень похожи. [ 17 ]
Единица измерения, используемая для описания рибосомных субъединиц и фрагментов рРНК, является единицей Сведберга , мерой скорости седиментации в центрифугировании , а не размере. Это объясняет, почему имена фрагментов не складываются: например, рибосомы бактериальных 70 -х годов сделаны из субъединиц 50 и 30 -х годов.
Прокариоты имеют 70 с рибосом, каждая из которых состоят из маленькой ( 30 -е ) и большой ( 50 -е ) субъединицы. E. coli , например, имеет субъединицу 16S РНК (состоящую из 1540 нуклеотидов), которая связана с 21 белком. Большая субъединица состоит из субъединицы РНК 5S (120 нуклеотидов), субъединицы РНК A 23S (2900 нуклеотидов) и 31 белка . [ 17 ]
Рибосома кишечной палочки (бактерия) [ 18 ] : 962 рибосома субъединица RRNAS R-белки 70 -е годы 50 -е годы 23S (2904 нт ) 31 5 с (120 нт) 30 -е годы 16S (1542 NT) 21
Аффинная метка для сайтов связывания тРНК на рибосоме E. coli позволила идентифицировать белки A и P -сайта, скорее всего, связанные с активностью пептидилтрансферазы; [ 5 ] Мечкованные белки - L27, L14, L15, L16, L2; По крайней мере, L27 расположен на донорском участке, как показано Э. Коллац и AP Czernilofsky. [ 19 ] [ 20 ] Дополнительные исследования показали, что белки S1 и S21 в связи с 3'-эндом рибосомальной РНК 16S участвуют в инициации трансляции. [ 21 ]
Архаальные рибосомы
[ редактировать ]Архаальные рибосомы имеют те же общие измерения бактерий, являющихся рибосомой 70 -х годов, состоящей из большой субъединицы 50 -х годов, небольшой субъединицы 30 -х годов, и содержащей три цепи рРНК. Однако на уровне последовательности они намного ближе к эукариотическим, чем к бактериальным. Каждый лишний рибосомный белок археи сравнивал с бактериями, имеет эукариотический аналог, в то время как такая связь не применяется между археей и бактериями. [ 22 ] [ 23 ] [ 24 ]
Эукариотические рибосомы
[ редактировать ]У эукариот есть рибосомы 80 -х годов, расположенные в их цитозоле, каждый из которых состоят из небольшой (40 -х) и большой (60 -х) субъединицы . Их субъединица 40 -х годов имеет 18S РНК (нуклеотиды 1900 года) и 33 белка. [ 25 ] [ 26 ] Большая субъединица состоит из РНК 5S (120 нуклеотидов), 28S РНК (4700 нуклеотидов), субъединиц РНК 5,8S (160 нуклеотидов) и 49 белков. [ 17 ] [ 25 ] [ 27 ]
Эукариотические цитозольные рибосомы ( R. norvegicus ) [ 18 ] : 65 рибосома субъединица RRNAS R-белки 80 -е годы 60 -е годы 28 с (4718 нт) 49 5,8 с (160 нт) 5 с (120 нт) 40 -е годы 18S (1874 NT) 33
В течение 1977 года Czernilofsky опубликовал исследование, в котором использовалась аффинная маркировка для выявления сайтов связывания тРНК на рибосомах печени крысы. Несколько белков, в том числе L32/33, L36, L21, L23, L28/29 и L13, были вовлечены как в центре пептидил -трансферазы . [ 28 ]
Пласторибосомы и миторибосомы
[ редактировать ]У эукариот рибосомы присутствуют в митохондриях (иногда называемых миторибосомами ) и в пластидах , таких как хлоропласты (также называемые пласторибосомами). Они также состоят из больших и маленьких субъединиц, связанных с белками в одну частицу 70 -х годов. [ 17 ] Эти рибосомы похожи на бактерии, и, как полагают, эти органеллы возникли как симбиотические бактерии . [ 17 ] Из двух хлоропластических рибосомов ближе к бактериальным, чем митохондриальные. Многие части рибосомальной РНК в митохондриях сокращаются, а в случае 5S рРНК заменены другими структурами у животных и грибов. [ 29 ] В частности, Leishmania tarentolae имеет минимализированный набор митохондриальной рРНК. [ 30 ] Напротив, растительные mitoribosomes имеют как расширенную рРНК, так и дополнительные белки по сравнению с бактериями, в частности, многие белки повторяющихся пентатропетид. [ 31 ]
Криптомонад , и хлорарачниофита водоросли могут содержать нуклеоморф который напоминает рудиментальное эукариотическое ядро. [ 32 ] Эукариотические рибосомы 80S могут присутствовать в компартменте, содержащем нуклеоморф. [ 33 ]
Использование различий
[ редактировать ]Различия между бактериальными и эукариотическими рибосомами эксплуатируются фармацевтическими химиками для создания антибиотиков , которые могут разрушить бактериальную инфекцию, не нанося вреда клеткам инфицированного человека. Из -за различий в их структурах рибосомы бактерий 70 -х годов уязвимы для этих антибиотиков, в то время как эукариотические рибосомы 80 -х не являются. [ 34 ] Несмотря на то, что митохондрии обладают рибосомами, сходными с бактериальными, эти антибиотики не влияют на митохондрии, потому что они окружены двойной мембраной, которая не легко допускает эти антибиотики в органелле . [ 35 ] Примечательным контрзаметом является противоопухолевый хлорампеникол антибиотиков , который ингибирует бактериальные 50 -х и эукариотические митохондриальные рибосомы 50S. [ 36 ] Однако рибосомы в хлоропластах различны: устойчивость к антибиотикам у хлоропластных рибосомных белков является признаком, которая должна быть представлена в качестве маркера, с генетической инженерией. [ 37 ]
Общие свойства
[ редактировать ]Различные рибосомы имеют основную структуру, которая очень похожа, несмотря на большие различия в размерах. Большая часть РНК очень организована в различные третичные структурные мотивы , например, псевдокноты , которые демонстрируют коаксиальную укладку . Дополнительная РНК в больших рибосомах находится в нескольких длинных непрерывных вставках, [ 38 ] так, что они образуют петли из основной структуры, не нарушая и не изменяя ее. [ 17 ] Вся каталитическая активность рибосомы осуществляется РНК ; Белки находятся на поверхности и, по -видимому, стабилизируют структуру. [ 17 ]
Структура высокого разрешения
[ редактировать ]
Общая молекулярная структура рибосомы была известна с начала 1970 -х годов. В начале 2000 -х годов структура была достигнута при высоких разрешениях по порядку нескольких Ångströms .
Первые статьи, дающие структуру рибосомы при атомной разрешении, были опубликованы почти одновременно в конце 2000 года. Субъединица 50 -х годов (большая прокариотическая) была определена из археонов Haloarcula Marishortui [ 39 ] и бактерия Deinococcus radiodurans , и структура субъединицы 30S определяли из Thermophilus бактерии Thermophilus . [ 16 ] [ 40 ] Эти структурные исследования были удостоены Нобелевской премии по химии в 2009 году. В мае 2001 года эти координаты были использованы для реконструкции всей частицы T. Thermophilus 70S с разрешением 5,5 Å . [ 41 ]
Две документы были опубликованы в ноябре 2005 года со структурами рибосомы Escherichia coli 70 -х годов. Структуры вакантной рибосомы определяли при 3,5 Å разрешении с использованием рентгеновской кристаллографии . [ 42 ] структура, основанная на крио-электронной микроскопии , Затем, две недели спустя, была опубликована [ 43 ] который изображает рибосому с разрешением 11–15 Å в акте передачи недавно синтезированной белковой цепи в канал, проводящий белок.
Первые атомные структуры рибосомы, комплексных с молекулами тРНК и мРНК , решали с использованием рентгеновской кристаллографии двумя группами независимо, при 2,8 Å [ 44 ] и в 3,7 Å . [ 45 ] Эти структуры позволяют увидеть детали взаимодействия рибосомы термофилуса с мРНК и с тРНК, связанными с классическими рибосомальными сайтами. Взаимодействие рибосомы с длинными мРНК, содержащими последовательности Shine-Dalgarno, были визуализированы вскоре после этого с разрешением 4,5–5,5 Å . [ 46 ]
В 2011 году первая атомная структура эукариотической рибосомы 80 -х годов из дрожжевых Saccharomyces cerevisiae была получена с помощью кристаллографии. [ 25 ] Модель раскрывает архитектуру специфичных для эукариота элементов и их взаимодействие с универсально консервативным ядром. В то же время была опубликована полная модель эукариотической рибосомальной структуры 40 -х годов в термофиле тетрагименной, и описала структуру субъединицы 40 -х годов , а также многое о взаимодействии субъединицы 40 -х годов с EIF1 во время инициации перевода . [ 26 ] Аналогичным образом, эукариотическая структура субъединицы 60 -х также была определена из термофилы тетрагименной в комплексе с EIF6 . [ 27 ]
Функция
[ редактировать ]Рибосомы - это мельчайшие частицы, состоящие из РНК, и связанных белков, которые функционируют для синтеза белков. Белки необходимы для многих клеточных функций, таких как восстановление повреждения или направление химических процессов. Рибосомы могут быть найдены плавающими в цитоплазме или прикреплены к эндоплазматической ретикулуме . Их основная функция заключается в преобразовании генетического кода в аминокислотную последовательность и создание белковых полимеров из аминокислотных мономеров.
Рибосомы действуют как катализаторы в двух чрезвычайно важных биологических процессах, называемых пептидил -переносом и пептидил -гидролизом. [ 5 ] [ 47 ] «Центр PT отвечает за продуцирование белковых связей во время удлинения белка». [ 47 ]
Таким образом, рибосомы имеют две основные функции: декодирование сообщения и образование пептидных связей. Эти две функции находятся в рибосомных субъединицах. Каждая субъединица изготовлена из одной или нескольких RRNAS и многих R-белков. Небольшая субъединица (30-е годы в бактериях и археи, 40-е годы у эукариот) имеет функцию декодирования, тогда как большая субъединица (50-е годы в бактериях и археи, 60S у эукариотов) катализирует образование пептидных связей, называемых как пептидил-трансфераза. Бактериальная (и архиальная) небольшая субъединица содержит 16S рРНК и 21 R-белок ( Escherichia coli ), тогда как малая субъединица эукариотической малой содержит 18S рРНК и 32 R-белковые (Saccharomyces cerevisiae, хотя число различается между видами). Бактериальная большая субъединица содержит 5S и 23S RRNAs и 34 R-белкотеины ( E. coli ), причем эукариотическая большая субъединица, содержащая 5S, 5,8 и 25 с/28S RRNAs и 46 R-протеинов ( S. cerevisiae ; снова,, Точные числа варьируются между видами). [ 48 ]
Перевод
[ редактировать ]Рибосомы - это рабочие места биосинтеза белка , процесс преобразования мРНК в белок . МРНК содержит серию кодонов , которые декодированы рибосомой для изготовления белка. Используя мРНК в качестве матрицы, рибосома пересекает каждый кодон (3 нуклеотида ) мРНК, сочетая ее с соответствующей аминокислотой, обеспечиваемой аминоацил-трНК . Аминоацил-трна содержит комплементарный антикодон на одном конце и соответствующую аминокислоту с другой. Для быстрого и точного распознавания соответствующей тРНК рибосома использует большие конформационные изменения ( конформационная корректура ). [ 49 ] Маленькая рибосомная субъединица, обычно связанная с аминоацил-тРНК, содержащей первый аминокислотный метионин , связывается с кодоном AUG на мРНК и рекрутирует большую рибосомальную субъединицу. Рибосома содержит три сайта связывания РНК, обозначенные A, P и E. A-site связывает коэффициенты аминоацил-тРНК или высвобождения завершения; [ 50 ] [ 51 ] P -сайт связывает пептидил-тРНК (тРНК, связанную с полипептидной цепью); и E-сайт (выход) связывает свободную тРНК. Синтез белка начинается в стартовом кодоне, около 5 'конца мРНК. МРНК сначала связывается с P -сайтом рибосомы. Рибосома распознает начальный кодон, используя последовательность Shine-Dalgarno мРНК в прокариотах и коробке Kozak у эукариот.
Хотя катализ пептидной связи C2 включает в себя гидроксил P-сайта РНК аденозина в механизме протонного челнока, другие этапы синтеза белка (такие как транслокация) вызваны изменениями в конформациях белка. Поскольку их каталитическое ядро сделано из РНК, рибосомы классифицируются как « рибозимы ». [ 52 ] И считается, что они могут быть остатками мира РНК . [ 53 ]

На рисунке 5 обе рибосомные субъединицы ( малые и большие ) собираются на стартовом кодоне (к 5 -'конец мРНК ) . Рибосома использует тРНК , которая соответствует текущему кодону (триплету) на мРНК, чтобы добавить аминокислоту в полипептидную цепь. Это делается для каждого триплета на мРНК, в то время как рибосома движется к 3 'конец мРНК. Обычно в бактериальных клетках несколько рибосомов работают параллельно одной мРНК, образуя так называемую полирибосому или полисомы .
Котранслациональное складывание
[ редактировать ]Известно, что рибосома активно участвует в складывании белка . [ 54 ] [ 55 ] Структуры, полученные таким образом, обычно идентичны таковым, полученным во время белкового химического прародинга; Однако пути, ведущие к конечному продукту, могут быть разными. [ 56 ] [ 57 ] В некоторых случаях рибосома имеет решающее значение для получения функциональной формы белка. Например, один из возможных механизмов складывания глубоко завязанных белков зависит от рибосомы, проталкивающей цепь через прикрепленную петлю. [ 58 ]
Добавление аминокислот-независимых от перевода
[ редактировать ]Присутствие контрольного белка качества рибосомы RQC2 связано с удлинением белка, независимого от мРНК. [ 59 ] [ 60 ] Это удлинение является результатом рибосомального добавления (через тРНК, принесенные RQC2) : кошек рибосомы расширяют C -терминус застопорившего белка со случайными, независимыми от трансляционными ланинов последовательностями и Threonines хвостов . [ 61 ] [ 62 ]
Расположение рибосом
[ редактировать ]Рибосомы классифицируются как «свободные» или «мембрану».
Свободные и мембранные рибосомы различаются только по их пространственному распределению; Они идентичны по структуре. Существует ли рибосома в свободном или мембрановом состоянии, зависит от наличия последовательности сигнала, нацеленного на ER на синтезируемом белке, поэтому отдельная рибосома может быть связана с мембраной, когда она делает один белок, но свободный в цитозоле Когда это делает другой белок.
Рибосомы иногда называют органеллами , но использование термина органелл часто ограничивается описанием субклеточных компонентов, которые включают фосфолипидную мембрану, которую рибосомы, являющиеся совершенно твердыми частицами, не делают. По этой причине рибосомы иногда могут быть описаны как «немамбранные органеллы».
Бесплатные рибосомы
[ редактировать ]Бесплатные рибосомы могут перемещаться в любом месте цитозоля , но исключаются из ядра клеток и других органеллов. Белки, которые образуются из свободных рибосом, высвобождаются в цитозоль и используются в клетке. Поскольку цитозоль содержит высокие концентрации глутатиона и, следовательно, является восстановительной средой , белки, содержащие дисульфидные связи , которые образуются из окисленных остатков цистеина, не могут быть получены внутри него.
Мембраносвязанные рибосомы
[ редактировать ]Когда рибосома начинает синтезировать белки, которые необходимы в некоторых органеллах, рибосома, создавая этот белок, может стать «мембраной». В эукариотических клетках это происходит в области эндоплазматического ретикулума (ER), называемой «грубой ER». Недавно продуцированные полипептидные цепи вставляются непосредственно в ER с помощью рибосомы, предпринятого вектора -синтеза , а затем транспортируются в свои пункты назначения через секреторный путь . Связанные рибосомы обычно продуцируют белки, которые используются в плазматической мембране или вытесняются из клетки посредством экзоцитоза . [ 63 ]
Биогенез
[ редактировать ]В бактериальных клетках рибосомы синтезируются в цитоплазме посредством транскрипции множественных рибосом оперонов . У эукариот процесс происходит как в клеточной цитоплазме, так и в ядреве , которая является областью в ядре клеток . Процесс сборки включает в себя скоординированную функцию более 200 белков в синтезе и обработке четырех рРНК, а также сборку этих РРНК с рибосомными белками. [ 64 ]
Источник
[ редактировать ]Рибосома, возможно, впервые возникла как проторибосома, [ 65 ] Возможно, содержащий центр пептидилтрансферазы (PTC) в мире РНК , который появляется как самореплицирующий комплекс, который только позже развил способность синтезировать белки, когда аминокислоты . начали появляться [ 66 ] Исследования показывают, что древние рибосомы, построенные исключительно из рРНК , могли развивать способность синтезировать пептидные связи . [ 67 ] [ 68 ] [ 69 ] [ 70 ] [ 71 ] Кроме того, доказательства сильно указывают на древние рибосомы как самореплицирующие комплексы, где рРНК в рибосомах имела информационные, структурные и каталитические цели, потому что они могли кодировать для тРНК и белков, необходимых для рибосомной саморепликации. [ 72 ] Гипотетические клеточные организмы с самореплицирующейся РНК, но без ДНК называются рибоцитами (или рибокеллами). [ 73 ] [ 74 ]
Поскольку аминокислоты постепенно появлялись в мире РНК в пребиотических условиях, [ 75 ] [ 76 ] Их взаимодействие с каталитической РНК увеличит как диапазон, так и эффективность функции каталитических молекул РНК. [ 66 ] Таким образом, движущая сила эволюции рибосомы из древней самореплицирующейся машины в его текущую форму, поскольку трансляционная машина, возможно, было селективным давлением для включения белков в самоотдавляемые механизмы рибосомы, чтобы увеличить его способность саморепликация. [ 72 ] [ 77 ] [ 78 ]
Гетерогенные рибосомы
[ редактировать ]Рибосомы являются композиционно гетерогенными между видами и даже внутри одной и той же клетки, о чем свидетельствует существование цитоплазматических и митохондрий рибосомов в одних и тех же эукариотических клетках. Некоторые исследователи предположили, что гетерогенность в составе рибосомных белков у млекопитающих важна для регуляции генов, то есть специализированной гипотезы рибосом. [ 79 ] [ 80 ] Однако эта гипотеза является противоречивой и темой текущих исследований. [ 81 ] [ 82 ]
Сначала было предложено, что гетерогенность в композиции рибосом участвует в трансляционном контроле синтеза белка Винсом Мауро и Джеральдом Эдельманом . [ 83 ] Они предложили гипотезу рибосомного фильтра, чтобы объяснить регуляторные функции рибосомов. Доказательства показали, что специализированные рибосомы, специфичные для различных клеточных популяций, могут влиять на то, как транслируются гены. [ 84 ] Некоторые рибосомные белки обмениваются из собранного комплекса с цитозольными копиями [ 85 ] предполагая, что структура рибосомы in vivo может быть модифицирована без синтеза всей новой рибосомы.
Некоторые рибосомные белки абсолютно критически важны для клеточной жизни, а другие - нет. У почкующихся дрожжей 14/78 рибосомных белков являются несущественными для роста, в то время как у людей это зависит от клетки исследования. [ 86 ] Другие формы гетерогенности включают посттрансляционные модификации рибосомных белков, таких как ацетилирование, метилирование и фосфорилирование. [ 87 ] Арабидопсис , [ 88 ] [ 89 ] [ 90 ] [ 91 ] Сайты вирусной внутренней рибосомы (iress) могут опосредовать переводы композиционно различными рибосомами. Например, рибосомные единицы 40 -х годов без ES25 в клетках дрожжей и млекопитающих не могут рекрутировать IGRE IGRE CRPV . [ 92 ]
Гетерогенность рибосомных модификаций РНК играет значительную роль в структурном поддержании и/или функции, а большинство модификаций мРНК обнаружены в высоко консервативных регионах. [ 93 ] [ 94 ] Наиболее распространенными модификациями рРНК являются псевдоуридилирование и 2'-О-метилирование рибозы. [ 95 ]
Смотрите также
[ редактировать ]- Аминогликозиды
- Биологические машины
- Посттрансляционная модификация
- Протеиновая динамика
- Связанная с рибосомой везикул
- РНК третичная структура
- Перевод (генетика)
- Пара базы колеблется
- Ада Йонат - израильский кристаллограф, известный своей новаторской работой над структурой рибосомы, за которую она получила Нобелевскую премию .
Ссылки
[ редактировать ]- ^ Konikkat S (февраль 2016 г.). Динамические события ремоделирования стимулируют удаление последовательности спейсеров ITS2 во время сборки рибосомальных субъединиц 60S у S. cerevisiae (Ph.D. Тезис). Университет Карнеги Меллона. Архивировано из оригинала 3 августа 2017 года.
- ^ Вейлер Е.В., Новер Л. (2008). Общая и молекулярная ботаника (на немецком языке). Штутгарт: Георг Тим Верлаг. п. 532. ISBN 9783131527912 .
- ^ De La Cruz J, Karbstein K, Woolford JL (2015). «Функции рибосомных белков в сборке эукариотических рибосомов in vivo» . Ежегодный обзор биохимии . 84 : 93–129. doi : 10.1146/annurev-biochem-060614-033917 . PMC 4772166 . PMID 25706898 .
- ^ «Scautbited по переводу на природе / РНК -перевод» .
- ^ Jump up to: а беременный в Tirumalai MR, Rivas M, Tran Q, Fox GE (ноябрь 2021 г.). «Центр пептидилтрансферазы: окно до прошлого» . Microbiol Mol Biol Rev. 85 (4): E0010421. doi : 10.1128/mmbr.00104-21 . PMC 8579967 . PMID 34756086 .
- ^ Benne R, Sloof P (1987). «Эволюция синтетического механизма митохондриального белка». Биосистемы . 21 (1): 51–68. Bibcode : 1987bisys..21 ... 51b . doi : 10.1016/0303-2647 (87) 90006-2 . PMID 2446672 .
- ^ «Рибосомы» . Архивировано с оригинала на 2009-03-20 . Получено 2011-04-28 .
- ^ Palade GE (январь 1955 г.). «Маленький компонент частиц цитоплазмы» . Журнал биофизической и биохимической цитологии . 1 (1): 59–68. doi : 10.1083/jcb.1.1.59 . PMC 2223592 . PMID 14381428 .
- ^ Rheinberger, Hans-Jörg (сентябрь 2022 г.). «Краткая история белкового биосинтеза и исследований рибосом» . Линдау Нобелевский лауреат собраний . Получено 2023-08-16 .
- ^ Робертс Р.Б., изд. (1958). "Введение". Микросомальные частицы и синтез белка . Нью -Йорк: Pergamon Press, Inc.
- ^ «Нобелевская премия по физиологии или медицине 1974» . Nobelprize.org . Нобелевский фонд. Архивировано с оригинала 26 января 2013 года . Получено 10 декабря 2012 года .
- ^ «Нобелевская премия 2009 года по химии» . Нобелевский фонд. Архивировано из оригинала 28 апреля 2012 года . Получено 10 декабря 2012 года .
- ^ Курланд К.Г. (1960). «Молекулярная характеристика рибонуклеиновой кислоты из рибосомов Escherichia coli». Журнал молекулярной биологии . 2 (2): 83–91. doi : 10.1016/s0022-2836 (60) 80029-0 .
- ^ Wilson DN, Doudna Cate JH (май 2012 г.). «Структура и функция эукариотической рибосомы» . Перспективы Cold Spring Harbor в биологии . 4 (5): A011536. doi : 10.1101/cshperspect.a011536 . PMC 3331703 . PMID 22550233 .
- ^ Nissen P, Hansen J, Ban N, Moore PB, Steitz TA (август 2000 г.). «Структурная основа активности рибосом в синтезе пептидной связи» (PDF) . Наука . 289 (5481): 920–30. Bibcode : 2000sci ... 289..920N . doi : 10.1126/science.289.5481.920 . PMID 10937990 . S2CID 8370119 . Архивировано из оригинала (PDF) 2020-11-30.
- ^ Jump up to: а беременный Wimberly BT, Brodersen DE, Clemons WM, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T, Ramakrishnan V (сентябрь 2000 г.). «Структура рибосомной субъединицы 30S». Природа . 407 (6802): 327–39. Bibcode : 2000natur.407..327W . doi : 10.1038/35030006 . PMID 11014182 . S2CID 4419944 .
- ^ Jump up to: а беременный в дюймовый и фон глин Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Мембраносвязанные рибосомы определяют грубую ER» . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. п. 342. ISBN 978-0-8153-4072-0 .
- ^ Jump up to: а беременный Гарретт Р., Гришам С.М. (2009). Биохимия (4 -е изд.). Cengage Learning Services. ISBN 978-0-495-11464-2 .
- ^ Collatz E, Küchler E, Stöffler G, Czernilofsky AP (апрель 1976 г.). «Сайт реакции на рибосомном белке L27 с производным с аффинной меткой тРНК Met F» . Письма Febs . 63 (2): 283–6. doi : 10.1016/0014-5793 (76) 80112-3 . PMID 770196 .
- ^ Czernilofsky AP, Collatz EE, Stöffler G, Kuechler E (январь 1974 г.). «Белки в сайтах связывания тРНК в рибосомах эшералихии» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (1): 230–4. Bibcode : 1974pnas ... 71..230c . doi : 10.1073/pnas.71.1.230 . PMC 387971 . PMID 4589893 .
- ^ Czernilofsky AP, Kurland CG, Stöffler G (октябрь 1975 г.). «Рибосомальные белки 30S, связанные с 3'-концевым концом 16S РНК» . Письма Febs . 58 (1): 281–4. doi : 10.1016/0014-5793 (75) 80279-1 . PMID 1225593 .
- ^ Cullen KE (2009). «Архаальные рибосомы». Энциклопедия науки о жизни . Нью -Йорк: Факты в файле. С. 1–5. doi : 10.1002/9780470015902.a0000293.pub3 . ISBN 9780470015902 Полем S2CID 243730576 .
- ^ Tirumalai MR, Anane-Bediakoh D, Rajesh R, Fox GE (ноябрь 2021 г.). «Чистые заряды рибосомных белков кластеров S10 и SPC галофилов обратно связаны со степенью галотолерантности» . Микробиол. Спектр 9 (3): E0178221. doi : 10.1128/spectrum.01782-21 . PMC 8672879 . PMID 34908470 .
- ^ Wang J, Dasgupta I, Fox GE (28 апреля 2009 г.). «Многие неравертные архаальные рибосомные белки обнаружены в консервативных генах кластеров» . Археи . 2 (4): 241–51. doi : 10.1155/2009/971494 . PMC 2686390 . PMID 19478915 .
- ^ Jump up to: а беременный в Бен-Шем А., Гарро де Луршес Н., Мельников С., Дженнер Л., Юсупова Г., Юсупов М (декабрь 2011 г.). «Структура эукариотической рибосомы при разрешении 3,0 Å » Наука . 334 (6062): 1524–9 Bibcode : 2011sci ... 334.1524b Doi : 10.1126/ science.1 22096102PMID 9099683S2CID
- ^ Jump up to: а беременный Rabl J, Leibundgut M, Ataide SF, Haag A, Ban N (февраль 2011 г.). «Кристаллическая структура рибосомной субъединицы эукариотической 40S в комплексе с фактором инициации 1» (PDF) . Наука . 331 (6018): 730–6. Bibcode : 2011sci ... 331..730R . doi : 10.1126/science.1198308 . HDL : 20.500.11850/153130 . PMID 21205638 . S2CID 24771575 .
- ^ Jump up to: а беременный Klinge S, Voigts-Hoffmann F, Leibundgut M, Arpagaus S, Ban N (ноябрь 2011 г.). «Кристаллическая структура рибосомной субъединицы эукариотической 60S в комплексе с фактором инициации 6». Наука . 334 (6058): 941–8. Bibcode : 2011sci ... 334..941K . doi : 10.1126/science.1211204 . PMID 22052974 . S2CID 206536444 .
- ^ Фабиджански С., Пеллегрини М. (1977). «Идентификация белков в сайте связывания пептидил-тРНК рибосомов печени крысы». Молекулярная и общая генетика . 184 (3): 551–6. doi : 10.1007/bf00431588 . PMID 6950200 . S2CID 9751945 .
- ^ Agrawal RK, Sharma MR (декабрь 2012 г.). «Структурные аспекты митохондриального трансляционного аппарата» . Современное мнение в структурной биологии . 22 (6): 797–803. doi : 10.1016/j.sbi.2012.08.003 . PMC 3513651 . PMID 22959417 .
- ^ Шарма М.Р., Бут Т.М., Симпсон Л., Маслов Д.А., Агравал Р.К. (июнь 2009 г.). «Структура митохондриальной рибосомы с минимальной РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (24): 9637–42. Bibcode : 2009pnas..106.9637S . doi : 10.1073/pnas.0901631106 . PMC 2700991 . PMID 19497863 .
- ^ Waltz F, Nguyen TT, Raviré M, Bochler A, Chicher J, Hammann P, Kuhn L, Quadrado M, Mireau H, Hashem Y, Giegé P (январь 2019). «Маленький большой в митохондриальной рибосоме арабидопсиса». Природные растения . 5 (1): 106–117. doi : 10.1038/s41477-018-0339-y . PMID 30626926 . S2CID 58004990 .
- ^ Archibald JM, Lane CE (2009). «Уйти, уйти, не совсем исчезнет: нуклеоморфы как тематическое исследование по снижению ядерного генома» . Журнал наследственности . 100 (5): 582–90. doi : 10.1093/jhered/esp055 . PMID 19617523 .
- ^ «Специализированные внутренние структуры прокариот | безграничная микробиология» . courses.lumenlearning.com . Получено 2021-09-24 .
- ^ Recht Mi, Douthwaite S, Puglisi JD (июнь 1999). «Основа для прокариотической специфичности действия аминогликозидных антибиотиков» . Embo Journal . 18 (11): 3133–8. doi : 10.1093/emboj/18.11.3133 . PMC 1171394 . PMID 10357824 .
- ^ О'Брайен Т.В. (май 1971). «Общее появление 55 с рибосом в митохондриях печени млекопитающих» . Журнал биологической химии . 246 (10): 3409–17. doi : 10.1016/s0021-9258 (18) 62239-2 . PMID 4930061 .
- ^ «Хлорамэмпеникол-сжиссер-подавление костного мозга» . Джама . 213 (7): 1183–1184. 1970-08-17. doi : 10.1001/Jama.1970.03170330063011 . ISSN 0098-7484 . PMID 5468266 .
- ^ Ньюман С.М., Бойнтон Дж., Гиллхэм Н.В., Рэндольф-Андерсон Б.Л., Джонсон А.М., Харрис Э.Х. (декабрь 1990 г.). «Трансформация генов рибосомальной РНК хлоропластов в Chlamydomonas: молекулярная и генетическая характеристика событий интеграции» . Генетика . 126 (4): 875–88. doi : 10.1093/Genetics/126.4.875 . PMC 1204285 . PMID 1981764 .
- ^ Пенев П.И., Фахретха-Авал С., Патель В.Дж., Кэннон Дж.Дж., Гутелл Р.Р., Петров А.С., Уильямс Л.Д., Гласс Дж.Б. (август 2020 г.). «Супер -рибосомальные сегменты расширения РНК в Asgard Archaea» . Биология и эволюция генома . 12 (10): 1694–1710. doi : 10.1093/gbe/evaa170 . PMC 7594248 . PMID 32785681 .
- ^ Jump up to: а беременный Бан Н., Ниссен П., Хансен Дж., Мур П.Б., Стейц Та (август 2000). «Полная атомная структура большой рибосомальной субъединицы при 2,4 в разрешении». Наука . 289 (5481): 905–20. Bibcode : 2000sci ... 289..905b . Citeseerx 10.1.1.58.2271 . doi : 10.1126/science.289.5481.905 . PMID 10937989 .
- ^ Schluenzen F, Tocolj A, Zarivach R, Harms J, Gluehmann M, Janell D, Bashan A, Bartels H, Agmon I, Franceschi F, Yonath A (сентябрь 2000 г.). «Структура функционально активированной маленькой рибосомной субъединицы при разрешении 3,3 ангстрем» . Клетка . 102 (5): 615–23. doi : 10.1016/s0092-8674 (00) 00084-2 . PMID 11007480 . S2CID 1024446 .
- ^ Юсупов М.М., Юсупова Г.З., Бауком А., Либерман К., Эрнест Т.Н., Кейт Дж. Х., Ноллер Х.Ф. (май 2001). «Кристаллическая структура рибосомы при 5,5 разрешении» . Наука . 292 (5518): 883–96. Bibcode : 2001sci ... 292..883y . doi : 10.1126/science.1060089 . PMID 11283358 . S2CID 39505192 .
- ^ Schuwirth BS, Borovinskaya MA, Hau CW, Zhang W, Vila-Sanjurjo A, Holton JM, Cate JH (ноябрь 2005 г.). «Структуры бактериальной рибосомы при 3,5 резолюции». Наука . 310 (5749): 827–34. Bibcode : 2005sci ... 310..827s . doi : 10.1126/science.1117230 . PMID 16272117 . S2CID 37382005 .
- ^ Митра К., Шаффицель С., Шейх Т., Тама Ф., Дженни С., Брукс К.Л., Бан Н., Фрэнк Дж. (Ноябрь 2005 г.). «Структура проводящего белка канала E. coli, связанной с трансляционной рибосомой» . Природа . 438 (7066): 318–24. Bibcode : 2005natur.438..318m . doi : 10.1038/nature04133 . PMC 1351281 . PMID 16292303 .
- ^ Selmer M, Dunham CM, Murphy FV, Weixlbaumer A, Petry S, Kelley AC, Weir JR, Ramakrishnan V (сентябрь 2006 г.). «Структура рибосомы 70 -х годов в комплексе с мРНК и тРНК». Наука . 313 (5795): 1935–42. Bibcode : 2006sci ... 313.1935s . doi : 10.1126/science.1131127 . PMID 16959973 . S2CID 9737925 .
- ^ Коростолев А., Траханов С., Лорберг М., Ноллер Х.Ф. (сентябрь 2006 г.). «Кристаллическая структура комплекса рибосом-тРНК 70-х годов выявляет функциональные взаимодействия и перестройки» . Клетка . 126 (6): 1065–77. doi : 10.1016/j.cell.2006.08.032 . PMID 16962654 . S2CID 13452915 .
- ^ Юсупова Г., Дженнер Л., Рис Б., Морас Д., Юсупов М (ноябрь 2006 г.). «Структурная основа для движения РНК мессенджера на рибосоме». Природа . 444 (7117): 391–4. Bibcode : 2006natur.444..391y . doi : 10.1038/nature05281 . PMID 17051149 . S2CID 4419198 .
- ^ Jump up to: а беременный «Специализированные внутренние структуры прокариот» . courses.lumenlearning.com . Безграничная микробиология . Получено 2018-09-27 .
- ^ LaFontaine, D.; Tollervey, D. (2001). «Функция и синтез рибосом». Nat Rev Mol Cell Biol . 2 (7): 514–520. doi : 10.1038/35080045 . HDL : 1842/729 . PMID 11433365 . S2CID 2637106 .
- ^ Савир Y, Tlusty T (апрель 2013 г.). «Рибосома как оптимальный декодер: урок в молекулярном распознавании» . Клетка . 153 (2): 471–479. BIBCODE : 2013APS..MARY46006T . doi : 10.1016/j.cell.2013.03.032 . PMID 23582332 .
- ^ Korkmaz G, Sanyal S (сентябрь 2017 г.). « Эшерехия Коли » . Журнал биологической химии . 292 (36): 15134–15142. doi : 10.1074/jbc.m117.785238 . PMC 5592688 . PMID 28743745 .
- ^ Konevega AL, Soboleva NG, Mahhno VI, Semenkov YP, Wintermeyer W, Rodnina MV, Katunin VI (январь 2004 г.). «Пуриновые основания в положении 37 тРНК стабилизируют кодон-и-атмодонское взаимодействие в рибосоме А. 2+ -зависимые взаимодействия » . РНК . 10 (1): 90–101. : 10.1261 /RNA.5142404 . PMC 1370521. 14681588 PMID DOI .
- ^ Rodnina MV, Beringer M, Wintermeyer W (январь 2007 г.). «Как рибосомы делают пептидные связи». Тенденции в биохимических науках . 32 (1): 20–26. doi : 10.1016/j.tibs.2006.11.007 . PMID 17157507 .
- ^ Чех, ТР (август 2000 г.). «Структурная биология. Рибосома - это рибозим». Наука . 289 (5481): 878–879. doi : 10.1126/science.289.5481.878 . PMID 10960319 . S2CID 24172338 .
- ^ Banerjee D, Sanyal S (октябрь 2014 г.). «Активность складывания белка рибосомы (PFAR) - мишень для антиприоновых соединений» . Вирусы . 6 (10): 3907–3924. doi : 10.3390/v6103907 . PMC 4213570 . PMID 25341659 .
- ^ Федоров Ан, Болдуин до (декабрь 1997 г.). «Котранслациональное складывание белка» . Журнал биологической химии . 272 (52): 32715–32718. doi : 10.1074/jbc.272.52.32715 . PMID 9407040 .
- ^ Болдуин Р.Л. (июнь 1975 г.). «Промежуточные продукты в реакциях складывания белка и механизм складывания белка». Ежегодный обзор биохимии . 44 (1): 453–475. doi : 10.1146/annurev.bi.44.070175.002321 . PMID 1094916 .
- ^ Das D, Das A, Samanta D, Ghosh J, Dasgupta S, Bhattacharya A, Basu A, Sanyal S, Das Gupta C (август 2008 г.). «Роль рибосомы в складывании белка» (PDF) . Биотехнологический журнал . 3 (8): 999–1009. doi : 10.1002/biot.200800098 . PMID 18702035 .
- ^ Dabrowski-Tumanski P, Piejko M, Niewieczerzal S, Stasiak A, Sulkowska Ji (декабрь 2018). «Белковое завязки с помощью активной резьбы зарождающейся полипептидной цепи, выходящей из канала выхода рибосомы». Журнал физической химии б . 122 (49): 11616–11625. doi : 10.1021/acs.jpcb.8b07634 . PMID 30198720 . S2CID 52176392 .
- ^ Брандман О., Стюарт-Эрнштейн Дж., Вонг Д., Ларсон А., Уильямс С.К., Ли Г.В., Чжоу С., Кинг Д., Шен П.С., Вейбезан Дж., Данн Дж.Г., Роски С., Инада Т., Фрост А., Вайсман Дж. (Ноябрь 2012 г.) Полем «Комплекс контроля качества, связанный с рибосомой, запускает деградацию зарождающихся пептидов и сигналов напряжения трансляции» . Клетка . 151 (5): 1042–1054. doi : 10.1016/j.cell.2012.10.044 . PMC 3534965 . PMID 23178123 .
- ^ Defenouillère Q, Yao Y, Mouaikel J, Namane A, Galopier A, Decourty L, Doyen A, Malabat C, Saveanu C, Jacquier A, Fromont-Racine M (март 2013 г.). «Комплекс, связанный с CDC48, связан с частицами 60-х годов, необходим для очистки аберрантных продуктов трансляции» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (13): 5046–5051. Bibcode : 2013pnas..110.5046d . doi : 10.1073/pnas.1221724110 . PMC 3612664 . PMID 23479637 .
- ^ Shen PS, Park J, Qin Y, Li X, Parsawar K, Larson MH, Cox J, Cheng Y, Lambowitz AM, Weissman JS, Brandman O, Frost A (январь 2015). «Синтез белка. Рибосомальные субъединицы RQC2P и 60S опосредуют мРНК-независимое удлинение зарождающихся цепей» . Наука . 347 (6217): 75–78. Bibcode : 2015sci ... 347 ... 75 с . doi : 10.1126/science.1259724 . PMC 4451101 . PMID 25554787 .
- ^ Кили, Дж.; Gutnikoff, R. (2 января 2015 г.). «Исследования рибосом выявляют новый механизм синтеза белка» (пресс -релиз). Медицинский институт Говарда Хьюза . Архивировано с оригинала 12 января 2015 года . Получено 16 января 2015 года .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Мембраносвязанные рибосомы определяют грубую ER» . Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарлендская наука. ISBN 978-0-8153-4072-0 .
- ^ Кресслер, Дитер; Боль, Эд; Babler, Jochen (2009). «Вождение в сборе рибосомы» (PDF) . Biochimica et Biophysica Acta (BBA) - исследования молекулярных клеток . 1803 (6): 673–683. doi : 10.1016/j.bbamcr.2009.10.009 . PMID 19879902 .
- ^ Танец, Эмбер (28 февраля 2023 г.). «Как началась жизнь? Один ключевой ингредиент появляется в поле зрения-команда ученых-нобелевского премия, привившая, сделала большой шаг вперед в своем стремлении реконструировать раннюю РНК, способную создавать белки» . Природа . 615 (7950): 22–25. doi : 10.1038/d41586-023-00574-4 . PMID 36854922 .
- ^ Jump up to: а беременный Noller HF (апрель 2012 г.). «Эволюция синтеза белка из мира РНК» . Перспективы Cold Spring Harbor в биологии . 4 (4): A003681. doi : 10.1101/cshperspect.a003681 . PMC 3312679 . PMID 20610545 .
- ^ Dabbs ER (1986). Мутантные исследования прокариотической рибосомы . Нью-Йорк: Springer-Verlag.
- ^ Noller HF, Hoffarth V, Zimniak L (июнь 1992 г.). «Необычная устойчивость пептидилтрансферазы к процедурам экстракции белка». Наука . 256 (5062): 1416–9. Bibcode : 1992sci ... 256.1416n . doi : 10.1126/science.1604315 . PMID 1604315 .
- ^ Nomura M, Mizushima S, Ozaki M, Traub P, Lowry CV (1969). «Структура и функция рибосом и их молекулярных компонентов». Симпозии Cold Spring Harbor по количественной биологии . 34 : 49–61. doi : 10.1101/sqb.1969.034.01.009 . PMID 4909519 .
- ^ Krupkin M, Matzov D, Tang H, Metz M, Kalaora R, Belousoff MJ, Zimmerman E, Bashan A, Yonath A (октябрь 2011 г.). «Взади, пережитая пребиотической связи, функционирует в современной рибосоме» . Фил. Транс. R. Soc. Беременный 366 (1580): 2972–8. doi : 10.1098/rstb.2011.0146 . PMC 3158926 . PMID 21930590 .
- ^ Bose T, Fridkin G, Davidovich C, Krupkin M, Dinger N, Falkovich AH, Peleg Y, Agmon I, Bashan A, Yonat A (февраль 2022 г.). «Происхождение жизни: проторибосома образует пептидные связи и связывает миры РНК и белок» . Нуклеиновые кислоты Res . 50 (4): 1815–1828. doi : 10.1093/nar/gkac052 . PMC 8886871 . PMID 35137169 .
- ^ Jump up to: а беременный Root-Bernstein M, Root-Bernstein R (февраль 2015 г.). «Рибосома как недостающая связь в эволюции жизни» . Журнал теоретической биологии . 367 : 130–158. doi : 10.1016/j.jtbi.2014.11.025 . PMID 25500179 .
- ^ Ярус М. (2002). «Изначальная генетика: фенотип рибоцита». Ежегодный обзор генетики . 36 : 125–51. doi : 10.1146/annurev.genet.36.031902.105056 . PMID 12429689 .
- ^ Forterre P, Krupovic M (2012). «Происхождение вирионов и виросел: повторная гипотеза о побеге». Вирусы: основные агенты жизни . С. 43–60. doi : 10.1007/978-94-007-4899-6_3 . ISBN 978-94-007-4898-9 .
- ^ Caetano-Anollés G, SeufferHeld MJ (2013). «Коэволюционные корни биохимии и клеточной организации бросают вызов мировой парадигме РНК». Журнал молекулярной микробиологии и биотехнологии . 23 (1–2): 152–77. doi : 10.1159/000346551 . PMID 23615203 . S2CID 41725226 .
- ^ Саладино Р., Ботта Г., Пино С., Костанцо Г., Ди Мауро Е (август 2012 г.). «Сначала генетика или метаболизм? Формамидная подсказка». Обзоры химического общества . 41 (16): 5526–65. doi : 10.1039/c2cs35066a . HDL : 11573/494138 . PMID 22684046 .
- ^ Fox GE (сентябрь 2010 г.). «Происхождение и эволюция рибосомы» . Холодный весна Harb Perspect Biol . 2 (9): A003483. doi : 10.1101/cshperspect.a003483 . PMC 2926754 . PMID 20534711 .
- ^ Fox GE (2016). «Происхождение и ранняя эволюция рибосомы». В Эрнандес Г., Ягус Р. (ред.). Эволюция механизма синтеза белка и его регуляция . Швейцария: Springer, Cham. С. 31–60. doi : 10.1007/978-3-319-39468-8 . ISBN 978-3-319-39468-8 Полем S2CID 27493054 .
- ^ Shi Z, Fujii K, Kovary KM, Genuth NR, Röst HL, Teruel MN, Barna M (июль 2017 г.). «Гетерогенные рибосомы преимущественно транслируют отдельные подборы мРНК по всему геному» . Молекулярная клетка . 67 (1). Elsevier BV: 71–83.e7. doi : 10.1016/j.molcel.2017.05.021 . PMC 5548184 . PMID 28625553 .
- ^ XUE S, Barna M (май 2012 г.). «Специализированные рибосомы: новая граница в области регуляции генов и биологии организма» . Природные обзоры. Молекулярная клеточная биология . 13 (6). Springer Science and Business Media LLC: 355–369. doi : 10.1038/nrm3359 . PMC 4039366 . PMID 22617470 .
- ^ Ферретти М.Б., Карбштейн К (май 2019). «Действительно ли функциональная специализация рибосом существует?» Полем РНК . 25 (5). Лаборатория Cold Spring Harbour: 521–538. doi : 10.1261/rna.069823.118 . PMC 6467006 . PMID 30733326 .
- ^ Farley-Barnes Ki, Ogawa LM, Baserga SJ (октябрь 2019). «Рибосомопатии: старые концепции, новые противоречия» . Тенденции в генетике . 35 (10). Elsevier BV: 754–767. doi : 10.1016/j.tig.2019.07.004 . PMC 6852887 . PMID 31376929 .
- ^ Mauro VP, Edelman GM (сентябрь 2002 г.). «Гипотеза рибосомного фильтра» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (19): 12031–6. Bibcode : 2002pnas ... 9912031M . doi : 10.1073/pnas.192442499 . PMC 129393 . PMID 12221294 .
- ^ XUE S, Barna M (май 2012 г.). «Специализированные рибосомы: новая граница в области регуляции генов и биологии организма» . Природные обзоры. Молекулярная клеточная биология . 13 (6): 355–69. doi : 10.1038/nrm3359 . PMC 4039366 . PMID 22617470 .
- ^ Матис А.Д., Нейлор Б.К., Карсон Р.Х., Эванс Э., Харвелл Дж., Кнехт Дж., Хексем Е., Пилор Ф.Ф., Миллер Б.Ф., Гамильтон К.Л., Транстрам М.К., Бикман Б.Т., Прайс Дж.С. (февраль 2017 г.). «Механизмы поддержания рибосомы in vivo в ответ на сигналы питательных веществ» . Молекулярная и клеточная протеомика . 16 (2): 243–254. doi : 10.1074/mcp.m116.063255 . PMC 5294211 . PMID 27932527 .
- ^ Штеффен К.К., Маккормик М.А., Фам К.М., Маккей В.Л., Делани Дж.Р., Мураками С.Дж. и др. (Май 2012 г.). «Дефицит рибосом защищает от стресса ER у Saccharomyces cerevisiae» . Генетика . 191 (1). Генетическое общество Америки: 107–118. doi : 10.1534/Genetics.111.136549 . PMC 3338253 . PMID 22377630 .
- ^ Ли С.В., Бергер С.Дж., Мартович С., Паса-Толич Л., Андерсон Г.А., Шен Ю. и др. (Апрель 2002 г.). «Прямой масс -спектрометрический анализ неповрежденных белков дрожжей крупной рибосомной субъединицы с использованием капиллярного LC/FTICR» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (9): 5942–5947. Bibcode : 2002pnas ... 99.5942L . doi : 10.1073/pnas.082119899 . PMC 122881 . PMID 11983894 .
- ^ Кэрролл AJ, Heazlewood JL, Ito J, Millar AH (февраль 2008 г.). «Анализ протеома цитозольной рибосомы арабидопсиса дает подробную информацию о своих компонентах и их посттрансляционной модификации» . Молекулярная и клеточная протеомика . 7 (2): 347–369. doi : 10.1074/mcp.m700052-mcp200 . PMID 17934214 .
- ^ Odintsova Ti, Müller EC, Ivanov AV, Egorov TA, Bienert R, Vladimirov SN, et al. (Апрель 2003 г.). «Характеристика и анализ посттрансляционных модификаций человеческих крупных цитоплазматических рибосомальных белков субъединицы с помощью масс -спектрометрии и секвенирования Эдмана». Журнал белковой химии . 22 (3): 249–258. doi : 10.1023/a: 1025068419698 . PMID 12962325 . S2CID 10710245 .
- ^ Yu Y, Ji H, Doudna JA, Leary JA (июнь 2005 г.). «Масс-спектрометрический анализ рибосомальной субъединицы 40-х годов: нативные и HCV-связанные комплексы» . Белковая наука . 14 (6): 1438–1446. doi : 10.1110/ps.041293005 . PMC 2253395 . PMID 15883184 .
- ^ Zeidan Q, Wang Z, De Maio A, Hart GW (июнь 2010 г.). «Циклоновые ферменты O-GLCNAC связываются с трансляционным механизмом и модифицируют сердечные рибосомные белки» . Молекулярная биология клетки . 21 (12): 1922–1936. doi : 10.1091/mbc.e09-11-0941 . PMC 2883937 . PMID 20410138 .
- ^ Ландри Д.М., Герц Ми, Томпсон -старший (декабрь 2009 г.). «RPS25 имеет важное значение для инициации перевода Dicistroviridae и Hepatite C Viral Iress» . Гены и развитие . 23 (23): 2753–2764. doi : 10.1101/gad.1832209 . PMC 2788332 . PMID 19952110 .
- ^ Decatur WA, Fournier MJ (июль 2002 г.). «Модификации рРНК и рибосома». Тенденции в биохимических науках . 27 (7): 344–51. doi : 10.1016/s0968-0004 (02) 02109-6 . PMID 12114023 .
- ^ Natchiar SK, Myasnikov AG, Kratzat H, Hazemann I, Klaholz BP (ноябрь 2017 г.). «Визуализация химических модификаций в структуре рибосомы человека 80 -х». Природа . 551 (7681): 472–477. Bibcode : 2017natur.551..472n . doi : 10.1038/nature24482 . PMID 29143818 . S2CID 4465175 .
- ^ Го Х (август 2018 г.). «Специализированные рибосомы и контроль перевода». Биохимическое общество транзакций . 46 (4): 855–869. doi : 10.1042/bst20160426 . PMID 29986937 . S2CID 51609077 .
Внешние ссылки
[ редактировать ]- Лабораторный компьютер имитирует рибосому в движении
- Роль рибосомы , Гвен В. Чайлдс, скопированная здесь
- Рибосома в протеопедии - свободная, совместная 3D -энциклопедия белков и других молекул
- Семейства рибосомальных белков в эмпизийном архивировании 2011-04-30 в The Wayback Machine
- Молекула месяца Архивировал 2009-10-27 в The Wayback Machine © RCSB Bloter Bank :
- Рибосома архивировал 2010-11-14 на The Wayback Machine
- Факторы удлинения архивировали 2011-03-16 на машине Wayback
- Палада
- 3D -электронная микроскопическая структура рибосом на банке данных EM (EMDB)
 Эта статья включает в себя материал общественного достояния из Научный учебник . NCBI . Архивировано из оригинала на 2009-12-08.
Эта статья включает в себя материал общественного достояния из Научный учебник . NCBI . Архивировано из оригинала на 2009-12-08.