Азиатский слон
| Азиатский слон Временной диапазон: | |
|---|---|
 | |
| Клыкастый бык Национальный парк Бандипур , Карнатака , Индия | |
 | |
| A cow and juveniles Yala National Park, Sri Lanka | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Proboscidea |
| Family: | Elephantidae |
| Genus: | Elephas |
| Species: | E. maximus[1] |
| Binomial name | |
| Elephas maximus[1] | |
| Subspecies[1] | |
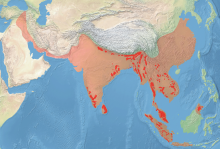 | |
historical range range in early 21st century | |

Азиатский слон ( Elephas maximus ), также известный как азиатский слон , — вид слонов , распространенный на Индийском субконтиненте и в Юго-Восточной Азии , от Индии на западе до Борнео на востоке и от Непала на севере до Суматры на юге. . три подвида Выделены — E. m. Максимус , Э.М. indicus и Э. м. суматранус . Азиатский слон отличается длинным хоботом с единственной пальцевой отделкой; большие бивни у самцов; сложенные по бокам большие уши, но меньшего размера в отличие от африканских слонов; и морщинистая серая кожа. Кожа более гладкая, чем у африканских слонов, и может быть депигментирована на туловище, ушах или шее. Взрослые самцы весят в среднем 4 т (4,4 коротких тонны), а самки - 2,7 т (3,0 коротких тонны).
Это один из трех ныне живущих видов слонов или слонов в мире, остальные — африканский кустарниковый слон и африканский лесной слон . Далее, азиатский слон — единственный ныне живущий вид рода Elephas . Это второй по величине вид слонов после африканского кустарникового слона. Он часто населяет луга , тропические вечнозеленые леса , полувечнозеленые леса, влажные лиственные леса , сухие лиственные леса и сухие терновые леса. Они травоядны и съедают около 150 кг (330 фунтов) растительности в день. Коровы и телята образуют группы, а самцы остаются поодиночке или образуют «холостяцкие группы» с другими самцами. В период размножения самцы временно присоединяются к женским группам для спаривания. Азиатские слоны имеют большой и хорошо развитый неокортекс головного мозга, они очень умны и самосознательны, способны демонстрировать поведение, связанное с горем, обучением, приветствием и т. д.
The Asian elephant is the largest living land animal in Asia. Since 1986, the Asian elephant has been listed as Endangered on the IUCN Red List, as the population has declined by at least 50 per cent over the last three elephant generations, which is about 60–75 years. It is primarily threatened by loss of habitat, habitat degradation, fragmentation and poaching. Wild Asian elephants live to be about 60 years old. While female captive elephants are recorded to have lived beyond 60 years when kept in semi-natural surroundings, Asian elephants die at a much younger age in captivity; captive populations are declining due to a low birth and high death rate. The earliest indications of captive use of Asian elephants are engravings on seals of the Indus Valley civilisation dated to the 3rd millennium BC.
Taxonomy
Carl Linnaeus proposed the scientific name Elephas maximus in 1758 for an elephant from Ceylon.[4] Elephas indicus was proposed by Georges Cuvier in 1798, who described an elephant from India.[5] Coenraad Jacob Temminck named an elephant from Sumatra Elephas sumatranus in 1847.[6] Frederick Nutter Chasen classified all three as subspecies of the Asian elephant in 1940.[7] These three subspecies are currently recognised as valid taxa.[8][9] Results of phylogeographic and morphological analyses indicate that the Sri Lankan and Indian elephants are not distinct enough to warrant classification as separate subspecies.[10]
Three subspecies are recognised:[8][9]
- Sri Lankan elephant (E. maximus maximus Linnaeus, 1758)

- Indian elephant (E. maximus indicus Cuvier, 1798)

- Sumatran elephant (E. maximus sumatranus Temminck, 1847)

Sri Lankan elephants are the largest subspecies. Their skin colour is darker than of E. m. indicus and of E. m. sumatranus with larger and more distinct patches of depigmentation on ears, face, trunk and belly.[11] The skin color of the Indian elephant is generally grey and lighter than that of E. m. maximus but darker than that of E. m. sumatranus.[9]
A potential fourth subspecies, the Borneo elephant (Elephas maximus borneensis), occurs in Borneo's northeastern parts, primarily in Sabah (Malaysia), and sometimes in Kalimantan (Indonesia).[12] It was proposed by Paules Deraniyagala in 1950, who described an elephant in an illustration published in the National Geographic magazine, but not as a living elephant in accordance with the rules of the International Code of Zoological Nomenclature.[13][14] These elephants living in northern Borneo are smaller than all the other subspecies, but had larger ears, a longer tail, and straight tusks.[15] Results of genetic analysis indicate that their ancestors separated from the mainland population about 300,000 years ago.[16] A study in 2003, using mitochondrial DNA analysis and microsatellite data, indicated that the Borneo elephant population is derived from stock that originated in the region of the Sunda Islands, and suggests that the Borneo population has been separated from the other elephant populations of southeast Asia since the Pleistocene.[17]
The following Asian elephants were proposed as extinct subspecies, but are now considered synonymous with the Indian elephant:[9]
- Syrian elephant (E. m. asurus), proposed by Deraniyagala, based on fossil remains and Bronze Age illustrations.[18][19]
- Chinese elephant (E. m. rubridens), also proposed by Deraniyagala, based on a Chinese bronze statuette.[20]
- Javan elephant (E. m. sondaicus), also proposed by Deraniyagala, based on an illustration of a carving on the Buddhist monument of Borobudur.[21][14]
Evolution
The genus Elephas, of which the Asian elephant is the only living member, is the closest relative of the extinct mammoths. The two groups are estimated to have split from each other around 7 million years ago.[22] Elephas originated in Sub-Saharan Africa during the Pliocene and spread throughout Africa before expanding into the southern half of Asia.[23] The earliest Elephas species, Elephas ekorensis, is known from the Early Pliocene of East Africa, around 5-4.2 million years ago.[24] The oldest remains of the genus in Asia are known from the Siwalik Hills in the Indian subcontinent, dating to the late Pliocene, around 3.6-3.2 million years ago, assigned to the species Elephas planifrons.[25] The modern Asian elephant is suggested to have evolved from the species Elephas hysudricus, which first appeared at the beginning of the Early Pleistocene around 2.6 million years ago, and is primarily known from remains of Early-Middle Pleistocene age found on the Indian subcontinent.[26] Skeletal remains of E. m. asurus have been recorded from the Middle East: Iran, Iraq, Syria, and Turkey from periods dating between at least 1800 BC and likely 700 BC.[27]
Description

In general, the Asian elephant is smaller than the African bush elephant and has the highest body point on the head. The back is convex or level. The ears are small with dorsal borders folded laterally. It has up to 20 pairs of ribs and 34 caudal vertebrae. The feet have five nail-like structures on each forefoot, and four on each hind foot.[9] The forehead has two hemispherical bulges, unlike the flat front of the African elephants.[28] Its long trunk or proboscis has only one fingerlike tip, in contrast to the African elephants which have two.[9] Hence, the Asian species relies more on wrapping around a food item and squeezing it into its mouth, rather than grasping with the tip. Asian elephants have more muscle coordination and can perform more complex tasks.[29]

Cows usually lack tusks; if tusks—in that case, called "tushes"—are present, they are barely visible and only seen when the mouth is open.[30] The enamel plates of the molars are greater in number and closer together in Asian elephants.[31] Some bulls may also lack tusks; these individuals are called "makhnas", and are especially common among the Sri Lankan elephant population.[32] A tusk from an 11 ft (3.4 m) tall elephant killed by Sir Victor Brooke measured 8 ft (2.4 m) in length, and nearly 17 in (43 cm) in circumference, and weighed 90 lb (41 kg). This tusk's weight is, however, exceeded by the weight of a shorter tusk of about 6 ft (1.8 m) in length which weighed 100 lb (45 kg), and there have reportedly been tusks weighing over 150 lb (68 kg).[33]

Skin colour is usually grey, and may be masked by soil because of dusting and wallowing. Their wrinkled skin is movable and contains many nerve centres. It is smoother than that of African elephants and may be depigmented on the trunk, ears, or neck. The epidermis and dermis of the body average 18 mm (0.71 in) thick; skin on the dorsum is 30 mm (1.2 in) thick providing protection against bites, bumps, and adverse weather. Its folds increase surface area for heat dissipation. They can tolerate cold better than excessive heat. Skin temperature varies from 24 to 32.9 °C (75.2 to 91.2 °F). Body temperature averages 35.9 °C (96.6 °F).[9]
Size
On average, when fully-grown, bulls are about 2.75 m (9.0 ft) tall at the shoulder and 4.0 t (4.4 short tons) in weight, while cows are smaller at about 2.40 m (7.9 ft) at the shoulder and 2.7 t (3.0 short tons) in weight.[34][35][36] Sexual dimorphism in body size is relatively less pronounced in Asian elephants than in African bush elephants; with bulls averaging 15% and 23% taller in the former and latter respectively.[34] Length of body and head including trunk is 5.5–6.5 m (18–21 ft) with the tail being 1.2–1.5 m (3.9–4.9 ft) long.[9] The largest bull elephant ever recorded was shot by the Maharajah of Susang in the Garo Hills of Assam, India, in 1924, it weighed an estimated 7 t (7.7 short tons), stood 3.43 m (11.3 ft) tall at the shoulder and was 8.06 m (26.4 ft) long from head to tail.[34][37][38] There are reports of larger individuals as tall as 3.7 m (12 ft).[33]
Distribution and habitat

Asian elephants are distributed throughout the Indian subcontinent and Southeast Asia, from India in the west, to Borneo in the east, and Nepal in the north, to Sumatra in the south.[2] They inhabit grasslands, tropical evergreen forests, semi-evergreen forests, moist deciduous forests, dry deciduous forests and dry thorn forests, in addition to cultivated and secondary forests and scrublands. Over this range of habitat types elephants occur from sea level to over 3,000 m (9,800 ft). In the eastern Himalaya in northeast India, they regularly move up above 3,000 m (9,800 ft) in summer at a few sites.[39]
In Bangladesh, some isolated populations survived in the south-east Chittagong Hills in the early 1990s.[40] In Malaysia's northern Johor and Terengganu National Park, two Asian elephants tracked using satellite tracking technology spent most of their time in secondary or "logged-over forest"; they travelled 75% of their time in an area less than 1.5 km (0.93 mi) away from a water source.[41] In China, the Asian elephant survives only in the prefectures of Xishuangbanna, Simao and Lincang of southern Yunnan. As of 2020[update], the estimated population was around 300 individuals.[42]
As of 2017, the estimated wild population in India account for nearly three-fourths of the extant population, at 27,312 individuals.[43] In 2019, the Asian elephant population in India increased to an estimated 27,000–29,000 individuals.[44][45] As of 2019[update], the global wild population was estimated at 48,323–51,680 individuals.[46]
Ecology and behaviour

Asian elephants are crepuscular.[9] They are classified as megaherbivores and consume up to 150 kg (330 lb) of plant matter per day.[47] Around 50 to 75% of the day is devoted to eating.[48] They are generalist feeders, and are both grazers and browsers. They are known to feed on at least 112 different plant species, most commonly of the order Malvales, as well as the legume, palm, sedge and true grass families.[49] They browse more in the dry season with bark constituting a major part of their diet in the cool part of that season.[50] They drink at least once a day and are never far from a permanent source of fresh water.[9] They need 80–200 litres of water a day and use even more for bathing. At times, they scrape the soil for clay or minerals.[51][52]

Cows and calves move about together as groups, while bulls disperse from their mothers upon reaching adolescence. Bulls are solitary or form temporary "bachelor groups".[53] Cow-calf units generally tend to be small, typically consisting of three adults (most likely related females) and their offspring.[54] Larger groups of as many as 15 adult females have also been recorded.[55] Seasonal aggregations of 17 individuals including calves and young adults have been observed in Sri Lanka's Uda Walawe National Park. Until recently, Asian elephants, like African elephants, were thought to be under the leadership of older adult females, or matriarchs. It is now recognized that cows form extensive and very fluid social networks, with varying degrees of associations between individuals.[56] Social ties generally tend to be weaker than in African bush elephants.[55] Unlike African elephants, which rarely use their forefeet for anything other than digging or scraping soil, Asian elephants are more agile at using their feet in conjunction with the trunk for manipulating objects. They can sometimes be known for their violent behavior.[28]

Asian elephants are recorded to make three basic sounds: growls, squeaks and snorts. Growls in their basic form are used for short distance communication. During mild arousal, growls resonate in the trunk and become rumbles while for long-distance communication, they escalate into roars. Low-frequency growls are infrasonic and made in many contexts. Squeaks come in two forms: chirpings and trumpets. Chirping consists of multiple short squeaks and signals conflict and nervousness. Trumpets are lengthened squeaks with increased loudness and are produced during extreme arousal. Snorts signal changes in activity and increase in loudness during mild or strong arousal. During the latter case, when an elephant bounces the tip of the trunk, it creates booms which serve as threat displays.[57] Elephants can distinguish low-amplitude sounds.[58]
Rarely, tigers have been recorded attacking and killing calves, especially if the calves become separated from their mothers, stranded from their herd, or orphaned. Adults are largely invulnerable to natural predation. There is a singular anecdotal case of a mother Asian elephant allegedly being killed alongside her calf; however, this account is contestable.[59][60] In 2011 and 2014, two instances were recorded of tigers successfully killing adult elephants; one by a single tiger in Jim Corbett National Park on a 20-year-old young adult elephant cow, and another on a 28-year-old sick adult bull in Kaziranga National Park further east, which was taken down and eaten by several tigers hunting cooperatively.[61][62] Elephants appear to distinguish between the growls of larger predators like tigers and smaller predators like leopards; they react to leopards less fearfully and more aggressively.[63]
Reproduction

Reproduction in Asian elephants can be attributed to the production and perception of signaling compounds called pheromones. These signals are transmitted through various bodily fluids. They are commonly released in urine but in males they are also found in special secretions from the temporal glands.[64] Once integrated and perceived, these signals provide the receiver with information about the reproductive status of the sender. If both parties are ready to breed, reproductive ritualic behavior occurs and the process of sexual reproduction proceeds.[65]

Bulls will fight one another to get access to oestrus cows. Strong fights over access to females are extremely rare. Bulls reach sexual maturity around the age of 12–15. Between the ages of 10 and 20 years, bulls undergo an annual phenomenon known as "musth". This is a period where the testosterone level is up to 100 times greater than non-musth periods, and they become aggressive. Secretions containing pheromones occur during this period, from the paired temporal glands located on the head between the lateral edge of the eye and the base of the ear.[66] The aggressive behaviors observed during musth can be attributed to varying amounts of frontalin (1,5-dimethyl-6,8-dioxabicyclo[3.2.1]octane) throughout the maturation process of bulls. Frontalin is a pheromone that was first isolated in bark beetles but can also be produced in the bulls of both Asian and African Elephants. The compound can be excreted through urine as well as through the temporal glands of the bull, allowing signaling to occur. During musth, increased concentrations of frontalin in the bull's urine communicate the reproductive status of the bull to female elephants.[67]
Similar to other mammals, hormone secretion in female elephants is regulated by an estrous cycle. This cycle is regulated by surges in Luteinizing hormone that are observed three weeks from each other. This type of estrous cycle has also been observed in African Elephants but is not known to affect other mammals. The first surge in Luteinizing hormone is not followed by the release of an egg from the ovaries.[68] However, some female elephants still exhibit the expected mating protocols during this surge. Female elephants give ovulatory cues by utilizing sex pheromones. A principal component thereof, (Z)-7-dodecen-1-yl acetate, has also been found to be a sex pheromone in numerous species of insects.[69][70] In both insects and elephants, this chemical compound is used as an attractant to assist the mating process. In elephants, the chemical is secreted through urination and this aids in the attraction of bulls to mate. Once detected, the chemical stimulates the vomeronasal organ of the bull, thus providing information on the maturity of the female.[71]
Репродуктивный обмен сигналами между слонами-самцами и самками передается через обонятельные сигналы в жидкостях организма. [65] У самцов повышение уровня фронталина во время сусла повышает их чувствительность к (Z)-7-додецен-1-илацетату, вырабатываемому самками слонов. [67] После восприятия рецепторами туловища следует последовательность ритуальных действий. Реакция самцов варьируется в зависимости от стадии развития и темперамента слона. [65] Этот процесс приема и обработки сигналов через хобот называется флеменом. Разница в движениях тела дает понять, заинтересован ли самец в спаривании с самкой, производящей секрет. [72] Бык, готовый к размножению, приближается к моче, и в некоторых случаях у него возникает эрекция. Бык, не готовый к размножению, будет робок и попытается дистанцироваться от сигнала. [65] Помимо репродуктивной коммуникации, хемосенсорная передача сигналов используется для облегчения однополых взаимодействий. Когда менее развитые самцы обнаруживают феромоны самца в сусле, они часто отступают, чтобы избежать контакта с агрессивным поведением. Также было замечено, что самки слонов общаются друг с другом посредством феромонов в моче. [65] Цель этого типа интерсексуального общения все еще исследуется. Однако существуют явные различия в силе сигнала и реакции приемника на разных стадиях эстрального цикла. [72]

Период беременности составляет 18–22 месяца, корова рожает одного теленка , лишь изредка двойню . Теленок полностью развивается к 19-му месяцу, но остается в утробе матери , чтобы вырасти и добраться до матери для кормления. При рождении теленок весит около 100 кг (220 фунтов) и находится на вскармливании до трех лет. После того, как самка рожает, она обычно не размножается снова, пока первый теленок не будет отнят от груди, в результате чего интервал между родами составляет четыре-пять лет. [73] [74] В этот период общение матери с теленком происходит преимущественно временными средствами. Однако известно, что у телят-самцов в молодом возрасте развиваются органы, вырабатывающие половые феромоны. Ранняя зрелость сошниково-носового органа позволяет неполовозрелым слонам производить и получать феромоны . [75] Маловероятно, что интеграция этих феромонов приведет к реакции Флемена у теленка. [72] Самки остаются в стаде, а половозрелых самцов прогоняют. [76]
Самки азиатских слонов достигают половой зрелости примерно в возрасте 10–15 лет и продолжают расти до 30 лет, тогда как самцы полностью созревают в возрасте старше 25 лет и постоянно растут на протяжении всей своей жизни. [77] [78] Средняя продолжительность жизни слонов составляет 60 лет в дикой природе и 80 лет в неволе, хотя в прошлом эта цифра преувеличивалась. [9] Продолжительность поколения азиатского слона составляет 22 года. [79]
Интеллект
У азиатских слонов очень большой и высокоразвитый неокортекс — черта, присущая людям , обезьянам и некоторым видам дельфинов . У них больший объем коры головного мозга, доступный для когнитивной обработки, чем у всех других существующих наземных животных. Результаты исследований показывают, что азиатские слоны обладают когнитивными способностями к использованию и изготовлению орудий, сходными с человекообразными обезьянами . [80] Они демонстрируют широкое разнообразие поведения, в том числе связанное с горем , обучением, алломотерингом , мимикрией , игрой, альтруизмом , использованием инструментов , состраданием , сотрудничеством , самосознанием , памятью и языком . [81] Сообщается, что слоны направляются в более безопасные места во время стихийных бедствий, таких как цунами и землетрясения, но данные двух шри-ланкийских слонов со спутниковыми ошейниками показывают, что это может быть неправдой. [82] Несколько студентов, изучающих когнитивные способности слонов и нейроанатомию, убеждены, что азиатские слоны очень умны и обладают самосознанием. [83] [84] [85] Другие оспаривают эту точку зрения. [86] [87]
Угрозы
Главной угрозой для азиатского слона сегодня являются потеря, деградация и фрагментация среды его обитания, что приводит к увеличению конфликтов между людьми и слонами. Азиатских слонов охотятся ради слоновой кости и множества других продуктов, включая мясо и кожу . [8] Спрос на кожу слона возрос в связи с тем, что она становится все более распространенным ингредиентом в традиционной китайской медицине. [88] [89]
Конфликт человека и слона


В некоторых частях Азии люди и слоны сосуществовали на протяжении тысячелетий. [90] В других районах люди и слоны вступают в конфликт, что приводит к насилию и, в конечном итоге, к перемещению слонов. [91] Основными причинами конфликта между людьми и слонами являются растущая численность населения, крупномасштабные проекты развития и плохое управление сверху вниз . Непосредственные причины включают утрату среды обитания из-за вырубки лесов , нарушение миграционных маршрутов слонов, расширение сельского хозяйства и незаконное вторжение на охраняемые территории. [92]
Уничтожение лесов в результате вырубки , вторжения, подсечек , сменного земледелия и монокультурных плантаций деревьев представляют собой серьезную угрозу выживанию слонов. Конфликты между человеком и слонами происходят, когда слоны совершают набеги на посевы вахтовых земледельцев на полях, которые разбросаны по большой территории, перемежающейся лесами. Разграбление населенных пунктов — еще одна серьезная область конфликтов между людьми и слонами, происходящая в небольших лесных массивах, вторжения в среду обитания слонов и на пути миграции слонов. [93] Однако исследования, проведенные в Шри-Ланке, показывают, что традиционное подсечно-огневое земледелие может создать оптимальную среду обитания для слонов, создавая мозаику сукцессионной растительности. Популяции, населяющие небольшие фрагменты среды обитания, гораздо более склонны к конфликту с человеком. [94]
Такие разработки, как ограждение границы вдоль границы Индии и Бангладеш, стали серьезным препятствием для свободного передвижения слонов. [95] В Ассаме более 1150 человек и 370 слонов погибли в результате конфликта между людьми и слонами в период с 1980 по 2003 год. [93] По оценкам исследования 2010 года, только в Индии слоны ежегодно убивают более 400 человек и повреждают от 0,8 до 1 миллиона гектаров, что затрагивает не менее 500 000 семей по всей стране. [96] [97] [98] Более того, известно, что слоны уничтожают посевы на сумму до 2–3 миллионов долларов США в год. [99] Это оказывает серьезное влияние на благосостояние и средства к существованию местных сообществ, а также на будущее сохранение этого вида. [92] В таких странах, как Бангладеш и Шри-Ланка , азиатский слон является одним из самых страшных диких животных, хотя он менее смертоносен, чем другие местные животные, такие как ядовитые змеи (которые, по оценкам, унесли в Шри-Ланке в 30 раз больше жизней, чем слоны). [100] [101]
В целом азиатские слоны демонстрируют очень сложное, а иногда и непредсказуемое поведение. Большинство диких слонов стараются избегать людей, но если они будут застигнуты врасплох какой-либо предполагаемой физической угрозой, включая людей, они, скорее всего, бросятся в атаку. Особенно это касается самцов в сусле и самок с молодняком. Стрельба и другие подобные методы отпугивания, которые, как известно, эффективны против многих видов диких животных, включая тигров, могут сработать, а могут и не сработать со слонами, и могут даже ухудшить ситуацию. Слоны, которые в прошлом подвергались насилию со стороны людей, часто становятся «слонами-изгоями», которые регулярно нападают на людей без каких-либо провокаций. [102] [103] [104]
Браконьерство
Для слоновой кости
Спрос на слоновую кость в 1970-е и 1980-е годы, особенно в Восточной Азии , привел к безудержному браконьерству и серьезному сокращению поголовья слонов как в Африке, так и в Азии. В Таиланде до сих пор процветает нелегальная торговля живыми слонами и слоновой костью. Хотя количество открыто продаваемой слоновой кости существенно сократилось с 2001 года, в Таиланде по-прежнему существует один из крупнейших и наиболее активных черных рынков слоновой кости в мире. На рынок также поступают бивни слонов, отловленных в тайском браконьерстве; в период с 1992 по 1997 год по меньшей мере 24 слона-самца были убиты из-за бивней. [105]
До начала 1990-х годов вьетнамские мастера по изготовлению изделий из слоновой кости использовали исключительно кость азиатского слона из Вьетнама и соседних Лаоса и Камбоджи. До 1990 года туристов было мало, а низкий спрос на обработанную слоновую кость мог удовлетворяться за счет домашних слонов. Экономическая либерализация и рост туризма повысили спрос как у местных жителей, так и у посетителей на обработанную слоновую кость, что привело к масштабному браконьерству. [106]
Для кожи
Кожа азиатского слона используется как ингредиент в китайской медицине , а также при изготовлении декоративных бус. Этой практике способствовала Государственная администрация лесного хозяйства Китая (SFA), которая выдала лицензии на производство и продажу фармацевтических продуктов, содержащих кожу слона, тем самым сделав торговлю законной. В 2010 году в лесу Мьянмы были найдены четыре слона со шкурой; В 2013 году браконьеры убили 26 слонов, а в 2016 году — 61. По данным неправительственной организации Elephant Family , Мьянма является основным источником слоновьих шкур, где с 2010 года быстро развивается кризис браконьерства. [107]
Болезнь
( Эндотелиотропный герпесвирус слона EEHV) является представителем рода Proboscivirus , новой клады, наиболее близкой к бетагерпевирусам млекопитающих . По состоянию на 2011 год он стал причиной смерти 70 слонов в зоопарках и диких азиатских слонов во всем мире, особенно молодых телят. [108] [109] несколько случаев гибели телят от эндотелиотропного герпевируса слонов . В частности, в Мьянме зафиксировано [110] Слоновья шистосома — паразитическая трематода , которая использует азиатского слона в качестве окончательного хозяина. Двумя другими хозяевами могут быть индийский слон и большой однорогий носорог. [111]
Сохранение


Азиатский слон внесен в Приложение I СИТЕС . [8] Это типичный флагманский вид , используемый для стимулирования ряда природоохранных целей, включая сохранение среды обитания в ландшафтных масштабах, повышение осведомленности общественности о вопросах сохранения и мобилизацию в качестве популярного культурного символа как в Индии, так и на Западе. [112] [113] [92] Ключевым аспектом сохранения является соединение предпочтительных маршрутов передвижения азиатских слонов через территории с высоким растительным покровом и низкой плотностью населения. [114]
Всемирный день слона отмечается ежегодно 12 августа с 2012 года. Мероприятия организуются с целью распространения информации и привлечения людей к проблемам, с которыми сталкивается азиатский слон. [115] Зоопарки и партнеры по охране природы в Соединенных Штатах объявили август Месяцем осведомленности об азиатских слонах. [116]
В штате Карнатака в Индии обитает больше всего азиатских слонов из всех известных территорий, и он составляет 20% от общей численности населения страны. Распространение слонов в штате, по одной из оценок, составляет примерно 38 310 км2. 2 (14 790 квадратных миль). [117] По данным исследования 2013 года, около 10 000 слонов населяли Западные Гаты , и им в первую очередь угрожало браконьерство и фрагментация среды обитания. В качестве вероятной проблемы также было названо увеличение числа конфликтов с людьми. Планы сохранения направлены на создание коридоров для дикой природы, прекращение браконьерства на быков, а также на защиту земельных территорий или управление ими. [118] Проект «Слон» был инициирован в 1992 году как программа централизованного спонсирования (CSS) Министерством окружающей среды, лесов и изменения климата правительства Индии . Проект был инициирован с целью защиты индийского слона и среды его обитания, а также создания специальных заповедников для поддержания популяции слонов. [119]
Распределение слонов на Шри-Ланке составляет лишь две пятых того, что было в конце 19 - начале 20 веков. Таким образом, взаимодействия с людьми происходят гораздо чаще. Во время опроса 2003 года местные жители выразили некоторую форму неодобрения по поводу сохранения азиатских слонов, поскольку фермеры считали их вредителями, однако большинство участников поддержали эту идею. [120]
В Китае азиатские слоны находятся под охраной первого уровня. В провинции Юньнань имеется 11 национальных и региональных заповедников. В общей сложности крытая охраняемая территория в Китае составляет около 510 000 га (1 300 000 акров). В 2020 году популяция азиатских слонов в Юньнани оценивалась примерно в 300 особей. Поскольку в последние годы вокруг охраняемых территорий возникли конфликты между людьми и дикими слонами, префектура Сишуанбаньна построила продовольственные базы и посадила бананы и бамбук, чтобы создать лучшую среду обитания. [42]
В Таиланде заповедник дикой природы Салакпра и национальный парк Тхам Тан Лот являются охраняемыми территориями, на которых, по данным за 2013 год, обитает около 250–300 слонов. [update]. [121] В последние годы Национальный парк столкнулся с проблемами из-за посягательств и чрезмерной эксплуатации. [122] В Индии Национальный совет дикой природы рекомендовал разрешить добычу угля в национальном парке Дехинг Паткай в апреле 2020 года. Это решение вызвало обеспокоенность среди студентов и активистов-экологов, которые начали онлайн-кампанию с целью остановить проект. [123]
В плену

Около половины мировой популяции слонов в зоопарках содержится в европейских зоопарках, где их продолжительность жизни составляет менее половины (18,9 года) средней продолжительности жизни сородичей (41,6 года) в охраняемых популяциях в странах ареала. Это несоответствие наиболее очевидно у азиатских слонов: детская смертность более чем в два-три раза выше, чем в бирманских лесных лагерях, а выживаемость взрослых особей в зоопарках за последние годы существенно не улучшилась. Одним из факторов риска для азиатских слонов в зоопарках является их перемещение из одного учреждения в другое, при этом раннее отделение от матери обычно приводит к дополнительным неблагоприятным последствиям. Еще одним фактором риска является рождение в зоопарке, а не завоз из дикой природы: плохая выживаемость взрослых азиатов, рожденных в зоопарке, очевидно, передается пренатально или в раннем детстве. Вероятными причинами снижения выживаемости являются стресс и/или ожирение . [124] Проблемы со стопами обычно наблюдаются у содержащихся в неволе слонов. Это связано с отсутствием физических упражнений, долгим стоянием на твердом грунте и заражением в результате стояния в навозе. Многие из этих проблем поддаются лечению. Однако неправильное обращение может привести к серьезной инвалидности или смерти. [125]
Демографический анализ содержащихся в неволе азиатских слонов в Северной Америке показывает, что популяция не является самодостаточной. Смертность в первый год жизни составляет почти 30 процентов, а плодовитость в первые репродуктивные годы крайне низка. [126] Данные региональных племенных книг Северной Америки и Европы за период с 1962 по 2006 год были проанализированы на наличие отклонений в соотношении полов при рождении и смертности молодняка. Из 349 рожденных в неволе телят 142 умерли преждевременно. Они умерли в течение одного месяца после рождения, основными причинами были мертворождение и детоубийство, совершенное матерью теленка или одним из товарищей по выставке. Установлено, что соотношение полов среди мертворожденных в Европе имеет тенденцию к избытку мужского пола. [127]
Методы обработки
Молодых слонов отлавливают в дикой природе и незаконно ввозят в Таиланд из Мьянмы для использования в туристической индустрии; телят используют в основном в парках развлечений и обучают выполнять различные трюки для туристов. [105] Телят часто подвергают процессу «взлома», который может включать в себя связывание, запирание, морение голодом, избиение и пытки; в результате две трети могут погибнуть. [128] Дрессировщики используют технику, известную как тренировочное давление , при которой «хендлеры используют лишение сна, голод и жажду, чтобы «сломать» дух слонов и сделать их покорными своим владельцам»; кроме того, проводники забивают слонам гвозди в уши и ноги. [129]
В культуре

Азиатский слон – национальное животное Таиланда и Лаоса. [130] [131] Он также был объявлен животным национального наследия Индии. [132] Кости азиатских слонов, раскопанные в Мохенджо-Даро в долине Инда, указывают на то, что они были приручены в цивилизации долины Инда и использовались для работы. Украшенные слоны также изображены на печатях и вылеплены из глины. [133] азиатский слон стал осадной машиной , скакуном на войне, символом статуса , вьючным животным и возвышенной площадкой для охоты В исторические времена в Южной Азии . [134]

Азиатских слонов поймали в дикой природе и приручили для использования людьми. Их способность работать под руководством делает их особенно полезными для переноски тяжелых предметов. Их использовали, в частности, для перевозки леса в джунглях. Помимо рабочего использования, их использовали на войне, на церемониях и для перевозки. [135] Сообщается, что Боевые слоны до сих пор используются Армией независимости Качина (KIA) в штате Качин на севере Мьянмы против вооруженных сил Мьянмы. Для перевозки грузов КИА использует около четырех десятков слонов. [136]
Азиатский слон играет важную роль в культуре субконтинента и за его пределами, он занимает видное место в баснях Панчатантры и буддийских сказках Джатака . Они играют важную роль в индуизме : голова бога Ганеши — голова слона, а «благословения» храмового слона высоко ценятся. Слонов часто используют в процессиях , где животных украшают праздничными нарядами. [137]
Азиатский слон изображен в нескольких индийских рукописях и трактатах, среди которых выделяется Матанга Лила (слоновий спорт) Нилаканты. [137] Рукопись Хастивидьярнава родом из Ассама на северо-востоке Индии. [138] В бирманском, тайском и сингальском животном и планетарном зодиаке азиатский слон, как с клыками, так и без клыков, является четвертым и пятым животным зодиаком бирманцев , четвертым животным зодиаком тайцев и вторым животным зодиаком сингальского народа . Шри-Ланка. [139] [140] Точно так же слон - двенадцатое животное зодиака в зодиаке животных Дай народа Дай на юге Китая. [141]
См. также
Ссылки
- ^ Перейти обратно: а б Шошани, Дж. (2005). «Вид Elephas maximus » . В Уилсоне, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. п. 90. ИСБН 978-0-8018-8221-0 . OCLC 62265494 .
- ^ Перейти обратно: а б Уильямс, К.; Тивари, СК; Госвами, В.Р.; де Сильва, С.; Кумар, А.; Баскаран, Н. ; Йогананд, К. и Менон, В. (2020). « Большой слон » Красный список исчезающих видов МСОП . 2020 : e.T7140A4 doi : 10.2305/IUCN.UK.2020-3.RLTS.T7140A45818198.en . Получено 15 января.
- ^ «Приложения» . Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения ( СИТЕС ). Архивировано из оригинала 5 декабря 2017 года . Проверено 6 июня 2022 г.
- ^ Линней, К. (1758). « Брута » . Карл Линней Система природы через три царства природы по классам, отрядам, родам, видам, с признаками, различиями, синонимами, местами . Том. Томус И. Холмиа: Impensis Direct. Лоран Сальвиус п. 33.
- ^ Кювье, Г. (1798). « Элефас индийский » . Элементарная таблица естественной истории животных . Париж: Бодуэн. стр. 148–149.
- ^ Темминк, CJ (1847). « Суматранский слон » . Общий обзор голландских владений в архипелаге Индия . Полет. Второй том. Лейде: А. Арнц и Комп. п. 91.
- ^ Чейзен, ФН (1940). «Список млекопитающих Малайзии, систематический список млекопитающих Малайского полуострова, Суматры, Борнео и Явы, включая прилегающие небольшие острова» (PDF) . Бюллетень музея Раффлз . 15 : 1–209. Архивировано из оригинала (PDF) 12 февраля 2020 года.
- ^ Перейти обратно: а б с д Чоудри, А.; Лахири Чоудхури, Дания; Десаи, А.; Дакворт, JW; Иса, PS; Джонсингх, AJT; Фернандо, П.; Хеджес, С.; Гунавардена, М.; Курт, Ф.; Карант, Ю; Листер, А.; Менон, В.; Риддл, Х.; Рубель, А.; Викраманаяке, Э.; Группа специалистов по азиатским слонам МСОП SSC (2008 г.). « Большой слон » Красный список исчезающих видов МСОП . 2008 : e.T7140A12828813.
- ^ Перейти обратно: а б с д и ж г час я дж к Шошани, Дж.; Айзенберг, Дж. Ф. (1982). « Большой слон » (PDF) . Виды млекопитающих (182): 1–8. дои : 10.2307/3504045 . JSTOR 3504045 . Архивировано из оригинала (PDF) 30 апреля 2013 года.
- ^ Флейшер, Р.К.; Перри, Э.А.; Муралидхаран, К.; Стивенс, Э.Э. и Веммер, К.М. (2001). «Филогеография азиатского слона ( Elephas maximus ) на основе митохондриальной ДНК» . Эволюция . 55 (9): 1882–1892. дои : 10.1111/j.0014-3820.2001.tb00837.x . JSTOR 2680432 . ПМИД 11681743 .
- ^ Шошани, Дж . (2006). «Таксономия, классификация и эволюция слонов» . В Фаулере, Мэн; Микота, СК (ред.). Биология, медицина и хирургия слонов . Уайли-Блэквелл. стр. 3–14. ISBN 0813806763 . Архивировано из оригинала 20 октября 2022 года . Проверено 4 июня 2020 г.
- ^ Фэрроуз. «Борнейский слон: виды в заповедниках Всемирного земельного фонда» . Всемирный земельный фонд . Архивировано из оригинала 14 мая 2021 года . Проверено 26 января 2021 г.
- ^ Дераниягала 1955 , с. 123-124.
- ^ Перейти обратно: а б Крэнбрук, Э.; Пейн, Дж. и Лех, CMU (2007). «Происхождение слонов Elephas maximus L. с Борнео» (PDF) . Журнал музея Саравака . 63 : 95–125. Архивировано (PDF) из оригинала 27 сентября 2012 года . Проверено 13 марта 2011 г.
- ^ Нуржафарина О.; Марьяти, М.; Ахмад, Ах; Натан, С.; Пирсон, ХТ; Гусенс, Б. (2008). «Предварительное исследование морфометрии борнейского слона» (PDF) . Журнал тропической биологии и охраны природы . 4 (1): 109–113. Архивировано из оригинала (PDF) 7 марта 2012 года.
- ^ Фернандо, П.; Видья, ТНК; Пейн, Дж.; Стьюве, М.; Дэвисон, Г.; Альфред, Р.Дж.; Андау, П.; Боси Э. и Килборн А. (2003). «Анализ ДНК показывает, что азиатские слоны являются коренными жителями Борнео и поэтому являются приоритетной задачей для сохранения» . ПЛОС Биология . 1 (1): e6. doi : 10.1371/journal.pbio.0000006 . ПМК 176546 . ПМИД 12929206 .
- ^ Фернандо, П.; Видья ТНК; Пейн Дж.; Стьюве М.; Дэвисон Г.; Альфред, Р.Дж.; Андау, П.; Боси, Э.; Килборн, А.; Мельник, диджей (2003). «Анализ ДНК показывает, что азиатские слоны являются родными для Борнео и поэтому их сохранение является приоритетной задачей» . ПЛОС Биология . 1 (1): e6. doi : 10.1371/journal.pbio.0000006 . ПМК 176546 . ПМИД 12929206 .
- ^ Дераниягала 1955 , с. 116.
- ^ Гирдланд Флинк, Эль; Албайрак Э. и Листер А. (2018). «Генетический взгляд на вымершую популяцию азиатских слонов ( Elephas maximus ) на Ближнем Востоке» . Открытый четвертичный период . 4 (1): 1–9. дои : 10.5334/ок.36 . hdl : 2164/13525 .
- ^ Дераниягала 1955 , с. 125.
- ^ Дераниягала 1955 , с. 124.
- ^ Роланд, Надин; Райх, Дэвид; Маллик, Свапан; Мейер, Матиас; Грин, Ричард Э.; Георгиадис, Николас Дж.; Рока, Альфред Л.; Хофрейтер, Михаэль (21 декабря 2010 г.). «Последовательности геномной ДНК мастодонта и шерстистого мамонта раскрывают глубокое видообразование лесных слонов и слонов саванны» . ПЛОС Биология . 8 (12): e1000564. дои : 10.1371/journal.pbio.1000564 . ISSN 1545-7885 . ПМК 3006346 . ПМИД 21203580 .
- ^ Хейнс, Г. (1993). Мамонты, мастодонты и слоны: биология, поведение и летопись окаменелостей . Издательство Кембриджского университета. п. 8. ISBN 978-0521456913 . Архивировано из оригинала 17 марта 2024 года . Проверено 19 марта 2018 г.
- ^ Сандерс, Уильям Дж.; Хайле-Селассие, Йоханнес (июнь 2012 г.). «Новое собрание хоботных среднего плиоцена из района Ворансо-Милле, регион Афар, Эфиопия: таксономические, эволюционные и палеоэкологические соображения» . Журнал эволюции млекопитающих . 19 (2): 105–128. дои : 10.1007/s10914-011-9181-y . ISSN 1064-7554 . S2CID 254703858 . Архивировано из оригинала 17 марта 2024 года . Проверено 12 августа 2023 г.
- ^ Яннуччи, Алессио; Сарделла, Рафаэле (28 февраля 2023 г.). «Что означает событие «Слон-Эквус» сегодня? Размышления о событиях расселения млекопитающих вокруг границы плиоцена и плейстоцена и гибкой двусмысленности биохронологии» . Четвертичный период . 6 (1): 16. дои : 10,3390/quat6010016 . hdl : 11573/1680082 . ISSN 2571-550X .
- ^ Листер, Адриан М.; Диркс, Венди; Ассаф, Амнон; Чазан, Майкл; Гольдберг, Пол; Аппльбаум, Яаков Х.; Гринбаум, Натали; Хорвиц, Лиора Кольска (сентябрь 2013 г.). «Новые ископаемые останки слонов из южного Леванта: значение для эволюционной истории азиатского слона» . Палеогеография, Палеоклиматология, Палеоэкология . 386 : 119–130. Бибкод : 2013PPP...386..119L . дои : 10.1016/j.palaeo.2013.05.013 . Архивировано из оригинала 2 января 2023 года . Проверено 12 августа 2023 г.
- ^ Чакырлар, Джанан; Икрам, Салима (3 мая 2016 г.). « Когда слоны дерутся, страдает трава». Власть, слоновая кость и сирийский слон» . Левант . 48 (2): 167–183. дои : 10.1080/00758914.2016.1198068 . ISSN 0075-8914 .
- ^ Перейти обратно: а б Клаттон-Брок, Джульетта (1987). Естественная история одомашненных млекопитающих . Издательство Кембриджского университета. п. 141. ИСБН 978-0-292-71532-5 .
- ^ Шошани, Дж., изд. (2000). Слоны: величественные создания дикой природы . Книги с галочками. стр. 38–41, 74–77. ISBN 978-0-87596-143-9 . OCLC 475147472 .
- ^ Челия, Карпагам; Сукумар, Раман (1 декабря 2013 г.). «Роль бивней, муста и размера тела в соревнованиях самцов и самцов среди азиатских слонов Elephas maximus» . Поведение животных . 86 (6): 1207–1214. дои : 10.1016/j.anbehav.2013.09.022 . ISSN 0003-3472 . Архивировано из оригинала 2 декабря 2018 года . Проверено 17 марта 2024 г.
- ^ Нэнси Э. Тодд (январь 2010 г.). «Качественное сравнение кранио-дентальной остеологии современных слонов Elephas Maximus (азиатский слон) и Loxodonta africana (африканский слон)» . Анатомическая запись . 293 (1): 62–73. дои : 10.1002/ar.21011 . ПМИД 19937635 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ «Махна» . База данных слонов . Архивировано из оригинала 21 мая 2024 года . Проверено 1 декабря 2023 г.
- ^ Перейти обратно: а б Лидеккер, Р. (1894). Королевская естественная история . Том. 2. Лондон: Фредерик Уорн и компания, стр. 522–523.
- ^ Перейти обратно: а б с Ларраменди, А. (2016). «Высота плеч, масса тела и форма хоботков» . Acta Palaeontologica Polonica . 61 (3): 537–574. дои : 10.4202/app.00136.2014 .
- ^ Сукумар, Р.; Джоши, Невада; Кришнамурти, В. (1988). «Рост азиатского слона». Труды: Науки о животных . 97 (6): 561–571. дои : 10.1007/BF03179558 . S2CID 84871322 .
- ^ Курт, Ф.; Кумарасингхе, Дж. К. (1998). «Замечания о росте тела и фенотипах азиатского слона Elephas maximus » . Акта Териологика . 5 (Приложение): 135–153. дои : 10.4098/AT.arch.98-39 .
- ^ Пиллаи, Н.Г. (1941). «О росте и возрасте слона». Журнал Бомбейского общества естествознания . 42 : 927–928.
- ^ Вуд, Г. (1983). Книга рекордов Гиннеса по фактам и подвигам животных . Энфилд, Миддлсекс: превосходная степень Книги Гиннеса. п. 17. ISBN 978-0-85112-235-9 .
- ^ Чоудри, Австралия (1999). «Статус и охрана азиатского слона Elephas maximus на северо-востоке Индии». Обзор млекопитающих . 29 (3): 141–173. дои : 10.1046/j.1365-2907.1999.00045.x .
- ^ Сукумар, Р. (1993). Азиатский слон: экология и управление (второе изд.). Кембридж: Издательство Кембриджского университета. п. 18. ISBN 978-0-521-43758-5 . Архивировано из оригинала 17 марта 2024 года . Проверено 15 декабря 2015 г.
- ^ Айни, С.; Суд, AM; Саабан, С. «Анализ параметров среды обитания слонов с использованием ГИС, дистанционного зондирования и процесса анализа иерархии на полуострове Малайзия» . Пертаника Журнал науки и технологий . 23 (1): 37–50. Архивировано из оригинала 17 мая 2023 года . Проверено 17 мая 2023 г.
- ^ Перейти обратно: а б Шуанг, Ф. (2020). «Решение дилеммы, вызванной слонами» . ЭКНС . Архивировано из оригинала 13 января 2021 года . Проверено 6 сентября 2020 г.
- ^ «По данным переписи, в Индии 27 312 слонов» . Индус . 16 августа 2017 года. Архивировано из оригинала 3 декабря 2023 года . Проверено 1 декабря 2023 г.
- ^ Шринивасайя, Нишант М.; Вайдьянатан, Шринивас; Сукумар, Раман; Синха, Аниндья (2019). «Слоны в движении: последствия для взаимодействия человека и слона» . Ежеквартальный журнал Индийского международного центра . 46 (3/4): 100–113. ISSN 0376-9771 . JSTOR 26946286 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ Баскаран, Н.; Варма, С.; Сар, СК; Сукумар, Р. (2011). «Современное состояние слонов в Индии» . Гаджа . 35 : 47–54. Архивировано из оригинала 21 мая 2024 года . Проверено 29 марта 2024 г.
- ^ Менон, В.; Тивари, С. (2019). «Состояние популяции азиатских слонов Elephas maximus и ключевые угрозы». Международный зоопаркский ежегодник . 53 (1): 17–30. дои : 10.1111/izy.12247 . S2CID 209571148 .
- ^ Самансири, КАП; Виракун, ДК (2007). «Пищевое поведение азиатских слонов в северо-западном регионе Шри-Ланки» (PDF) . Гаджа . 2 : 27–34. Архивировано (PDF) из оригинала 18 ноября 2016 года . Проверено 18 октября 2011 г.
- ^ Сукумар, Р. (июль 2006 г.). «Краткий обзор состояния, распространения и биологии диких азиатских слонов Elephas maximus» . Международный зоопаркский ежегодник . 40 (1): 1–8. дои : 10.1111/j.1748-1090.2006.00001.x . ISSN 0074-9664 . Архивировано из оригинала 18 марта 2024 года . Проверено 18 марта 2024 г.
- ^ Сукумар, Р. (1990). «Экология азиатского слона на юге Индии. II. Привычки питания и модели набегов на урожай» (PDF) . Журнал тропической экологии . 6 : 33–53. дои : 10.1017/S0266467400004004 . S2CID 85129439 . Архивировано из оригинала (PDF) 12 октября 2007 года.
- ^ Прадхан, НМБ; Вегге, П.; Мо, СР; Шреста, АК (2008). «Экология питания двух находящихся под угрозой исчезновения симпатрических мегатравоядных животных: азиатского слона Elephas maximus и большого однорогого носорога Rhinoceros unicornis в низменном Непале» (PDF) . Биология дикой природы . 14 : 147–154. doi : 10.2981/0909-6396(2008)14[147:FEOTES]2.0.CO;2 . S2CID 85964847 . Архивировано (PDF) из оригинала 13 марта 2023 года . Проверено 3 января 2023 г.
- ^ Сак, Фиона; Диренфельд, Эллен С.; Лэнгли-Эванс, Саймон К.; Гамильтон, Эллиотт; Мюррей Ларк, Р.; Йон, Лиза; Уоттс, Майкл Дж. (15 мая 2020 г.). «Потенциальные биоиндикаторы для оценки минерального статуса слонов» . Научные отчеты . 10 (1): 8032. Бибкод : 2020NatSR..10.8032S . дои : 10.1038/s41598-020-64780-0 . ISSN 2045-2322 . ПМК 7229182 . ПМИД 32415129 .
- ^ Сингх, АП; Шарма, Рамеш К. (24 сентября 2001 г.). «Конфликты между линейными застройками и азиатскими слонами в субгималайской зоне Уттранчала» . Дорожный экологический центр . Архивировано из оригинала 18 марта 2024 года . Проверено 18 марта 2024 г.
- ^ Маккей, генеральный менеджер (1973). «Поведение и экология азиатского слона на юго-востоке Цейлона». Смитсоновский вклад в зоологию . 125 (125): 1–113. дои : 10.5479/si.00810282.125 . S2CID 128585445 .
- ^ Фернандо, П.; Ланде, Р. (2000). «Молекулярно-генетический и поведенческий анализ социальной организации азиатского слона ( Elephas maximus )». Поведение Экол Социобиол . 48 (1): 84–91. дои : 10.1007/s002650000218 . S2CID 33707844 .
- ^ Перейти обратно: а б де Сильва, С.; Виттемайер, Г. (2012). «Сравнение социальной организации азиатских слонов и африканских слонов саванны». Международный журнал приматологии . 33 (5): 1125–1141. дои : 10.1007/s10764-011-9564-1 . S2CID 17209753 .
- ^ де Сильва, С.; Ранджиева, заместитель генерального директора; Кряжимский, С. (2011). «Динамика социальных сетей среди самок азиатских слонов» . БМК Экология . 11 (1): 17. Бибкод : 2011BMCE...11...17D . дои : 10.1186/1472-6785-11-17 . ПМК 3199741 . ПМИД 21794147 .
- ^ Сукумар, Р. (2003). Живые слоны: эволюционная экология, поведение и охрана . Оксфорд: Издательство Оксфордского университета. п. 142. ИСБН 978-0-19-510778-4 .
- ^ Хеффнер, Р.; Хеффнер, Х. (1980). «Слух у слона ( Elephas maximus )» . Наука . 208 (4443): 518–520. Бибкод : 1980Sci...208..518H . дои : 10.1126/science.7367876 . PMID 7367876 . Архивировано из оригинала 17 марта 2024 года . Проверено 27 сентября 2013 г.
- ^ Карант, К. К. и Николс, JD (1998). «Оценка плотности тигров в Индии с использованием фотографических отловов и повторных поимок» (PDF) . Экология . 79 (8): 2852–2862. doi : 10.1890/0012-9658(1998)079[2852:EOTDII]2.0.CO;2 . JSTOR 176521 . Архивировано из оригинала (PDF) 9 августа 2017 года . Проверено 7 декабря 2012 г.
- ^ «Тигр убивает мать слонёнка» . Новости о слонах . 2006. Архивировано из оригинала 14 июля 2014 года.
- ^ «Тигр убивает слона в заповеднике Корбетт» . Индус . 29 января 2011 г. Архивировано из оригинала 8 февраля 2024 г. Проверено 8 февраля 2024 г.
- ^ «Слон Казиранга убит при нападении тигра» . Таймс оф Индия . 22 октября 2014 г. Архивировано из оригинала 8 февраля 2024 г. Проверено 8 февраля 2024 г.
- ^ Туппил, В.; Косс, Р.Г. (2013). «Дикие азиатские слоны различают агрессивное рычание тигра и леопарда в зависимости от предполагаемой опасности» . Письма по биологии . 9 (5): 20130518. doi : 10.1098/rsbl.2013.0518 . ПМЦ 3971691 . ПМИД 24026347 . Архивировано из оригинала 14 июля 2014 года.
- ^ Расмуссен, ЛЕЛ (1 июня 1999 г.). «Эволюция химических сигналов у азиатского слона Elephas maximus: поведенческие и экологические влияния» . Журнал биологических наук . 24 (2): 241–251. дои : 10.1007/BF02941206 . ISSN 0973-7138 . S2CID 37477741 . Архивировано из оригинала 17 марта 2024 года . Проверено 27 ноября 2022 г.
- ^ Перейти обратно: а б с д и Шульте, Брюс А.; Бэгли, Кэтрин; Коррелл, Морин; Грей, Эми; Хейнеман, Сара М.; Лоизи, Хелен; Маламент, Мишель; Скотт, Нэнси Л.; Слэйд, Барбара Э.; Стэнли, Лорен; Гудвин, Томас Э.; Расмуссен, ЛЕЛ (2005). Мейсон, Роберт Т.; ЛеМастер, Майкл П.; Мюллер-Шварце, Дитланд (ред.). «Оценка химической коммуникации у слонов» . Химические сигналы у позвоночных 10 . Бостон, Массачусетс: Springer США: 140–151. дои : 10.1007/0-387-25160-X_18 . ISBN 978-0-387-25160-8 . Архивировано из оригинала 22 ноября 2022 года . Проверено 27 ноября 2022 г.
- ^ Джайнудин, MR; Маккей, генеральный менеджер; Айзенберг, Дж. Ф. (1972). «Наблюдение за суслом у домашнего азиатского слона ( Elephas maximus )». Млекопитающие . 36 (2): 247–261. дои : 10.1515/мамм.1972.36.2.247 . S2CID 84275661 .
- ^ Перейти обратно: а б Расмуссен, ЛЕЛ (1 июня 2003 г.). «Фронталин: химическое значение муста у азиатских слонов (elephas maximus)» . Химические чувства . 28 (5): 433–446. дои : 10.1093/chemse/28.5.433 . ПМИД 12826539 . Архивировано из оригинала 3 ноября 2022 года . Проверено 27 ноября 2022 г.
- ^ Браун, Дж.Л.; Шмитт, Д.Л.; Беллем, А.; Грэм, Л.Х.; Ленхардт, Дж. (ноябрь 1999 г.). «Секреция гормонов у азиатского слона (Elephas maximus): характеристика овуляторных и ановуляторных всплесков лютеинизирующего гормона» . Биология размножения . 61 (5): 1294–1299. дои : 10.1095/biolreprod61.5.1294 . ISSN 0006-3363 . ПМИД 10529277 .
- ^ Расмуссен, ЛЕЛ; Ли, ТД; Чжан, Эй Джей; Рулофс, В.Л.; Дэйвс, Джорджия (1997). «Очистка, идентификация, концентрация и биологическая активность (Z)-7-додецен-1-илацетата: полового феромона самки азиатского слона Elephas maximus » . Химические чувства . 22 (4): 417–437. дои : 10.1093/chemse/22.4.417 . ПМИД 9279465 .
- ^ Расмуссен, ЛЕЛ; Ли, ТД; Рулофс, В.Л.; Чжан, Эй Джей; Дэйвс, Джорджия (1996). «Феромоны насекомых у слонов» . Природа . 379 (6567): 684. Бибкод : 1996Natur.379..684R . дои : 10.1038/379684a0 . ПМИД 8602213 . S2CID 4330432 .
- ^ Расмуссен, ЛЕЛ (22 февраля 1996 г.). «Феромоны насекомых у слонов» (PDF) . Природа . 379 (6567): 684. Бибкод : 1996Natur.379..684R . дои : 10.1038/379684a0 . ПМИД 8602213 . S2CID 4330432 . Архивировано (PDF) из оригинала 3 ноября 2022 года . Проверено 27 ноября 2022 г.
- ^ Перейти обратно: а б с Шульте, Брюс А.; ЛаДью, Чейз А. (30 сентября 2021 г.). «Химическая экология слонов: дополнения 21 века к нашему пониманию и прогнозам на будущее» . Животные . 11 (10): 2860. дои : 10.3390/ani11102860 . ISSN 2076-2615 . ПМЦ 8532676 . ПМИД 34679881 .
- ^ Кроули, JAH; Мамби, HS; Чепмен, С.Н.; Лахденпера, М.; Мар, КУ; Хтут, В.; Тура Со, А.; Аунг, Х.Х.; Луммаа, В. (5 августа 2017 г.). «Чем больше, тем лучше? Взаимосвязь между размером и воспроизводством самок азиатских слонов» . Журнал эволюционной биологии . 30 (10): 1836–1845. дои : 10.1111/jeb.13143 . ISSN 1010-061X . ПМИД 28703384 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ Оуэн-Смит, Р. Норман (1988). Мегатравоядные животные: влияние очень крупных размеров тела на экологию . Издательство Кембриджского университета. стр. 145–150. ISBN 978-0-521-42637-4 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ Джонсон, Эдвард В.; Расмуссен, Лель (1 июля 2002 г.). «Морфологическая характеристика сошниково-носового органа новорожденного азиатского слона (Elephas maximus)» . Анатомическая запись . 267 (3): 252–259. дои : 10.1002/ar.10112 . ISSN 0003-276X . ПМИД 12115276 . S2CID 30345793 .
- ^ «Слоновья общественная организация» . ccrsl . Архивировано из оригинала 4 марта 2016 года . Проверено 29 августа 2015 г.
- ^ Фернандо, П.; Виджаякришнан, С.; Ранджива, AD; Пасторини, Дж. (2022). «Шкала размерно-возрастных классов азиатских слонов». Гаджа 55 :30-39.
- ^ Мамби, HS; Чепмен, С.Н.; Кроули, Дж.А.; Мар, КУ; Хтут, В.; Тура Со, А.; Луммаа, В. (2015). «Различие детерминантного и неопределенного роста у млекопитающих-долгожителей» . Эволюционная биология BMC . 15 (1): 1-9. Бибкод : 2015BMCEE..15..214M . дои : 10.1186/s12862-015-0487-x . ПМЦ 4604763 . ПМИД 26464339 .
- ^ Пасифичи, М.; Сантини, Л.; Ди Марко, М.; Байсеро, Д.; Франкуччи, Л.; Гроттоло Маразини, Г.; Висконти, П.; Рондинини, К. (2013). «Продолжительность поколений млекопитающих» . Охрана природы . 5 :87–94. doi : 10.3897/natureconservation.5.5734 . Архивировано из оригинала 26 января 2021 года . Проверено 20 ноября 2020 г.
- ^ Харт, Бенджамин Л.; Харт, Линетт А.; Маккой, Майкл; Сарат, Чехия (ноябрь 2001 г.). «Когнитивное поведение азиатских слонов: использование и модификация ветвей для переключения мух» . Поведение животных . 62 (5): 839–847. дои : 10.1006/anbe.2001.1815 . ISSN 0003-3472 . Архивировано из оригинала 5 февраля 2017 года . Проверено 18 марта 2024 г.
- ^ Барретт, Лиза П.; Бенсон-Амрам, Сара (август 2021 г.). «Множественные оценки личности и способности решать проблемы у содержащихся в неволе азиатских слонов (Elephas maximus) и африканских слонов саванны (Loxodonta africana)» . Журнал сравнительной психологии . 135 (3): 406–419. дои : 10.1037/com0000281 . ISSN 1939-2087 . ПМИД 34166041 .
- ^ Викраманаяке, Э.; Фернандо, П.; Леймгрубер, П. (2008). «Поведенческая реакция слонов со спутниковыми ошейниками на цунами на юге Шри-Ланки». Биотропика . 38 (6): 775–777. дои : 10.1111/j.1744-7429.2006.00199.x . S2CID 85358582 .
- ^ С., Дж.К.; Уильямс, Дж. Х. (октябрь 1950 г.). «Слон Билл» . Географический журнал . 116 (4/6): 229. Бибкод : 1950GeogJ.116..229S . дои : 10.2307/1789395 . ISSN 0016-7398 . JSTOR 1789395 . Архивировано из оригинала 21 мая 2024 года . Проверено 15 мая 2024 г.
- ^ Олдос, П. (2006). «Слоны видят себя в зеркале» . Новый учёный. Архивировано из оригинала 11 мая 2015 года . Проверено 24 августа 2017 г.
- ^ Пул, Дж. (1997). Слоны . Библиотека мировой жизни. п. 61. ИСБН 978-0-896-58357-3 .
- ^ Сандерсон, врач общей практики (1879 г.). Тринадцать лет среди диких зверей Индии . Лондон: WH Allen and Co., с. 80.
- ^ Ниссани, М. (2006). «Применяют ли азиатские слоны причинно-следственные связи к задачам, связанным с использованием инструментов?». Журнал экспериментальной психологии: процессы поведения животных . 31 (1): 91–96. дои : 10.1037/0097-7403.32.1.91 . ПМИД 16435969 .
- ^ Ван, Дэвид QH; Кэри, Мартин С. (7 августа 2014 г.). «Терапевтическое использование желчи животных в традиционной китайской медицине: этнофармакологический, биофизический, химический и медицинский обзор» . Всемирный журнал гастроэнтерологии . 20 (29): 9952–9975. дои : 10.3748/wjg.v20.i29.9952 . ISSN 1007-9327 . ПМЦ 4123376 . ПМИД 25110425 .
- ^ Нейман, Винсент; Шепард, Крис Р. (12 июля 2017 г.). «Этнозоологическая оценка животных, используемых торговцами традиционной медицины Мон в Кьяиктио, Мьянма» . Журнал этнофармакологии . 206 : 101–106. дои : 10.1016/j.jep.2017.05.010 . ISSN 0378-8741 . ПМИД 28506903 . Архивировано из оригинала 21 мая 2024 года . Проверено 18 марта 2024 г.
- ^ Лим, Теквин; Кампос-Арсеис, Ахимса (2022 г.). «Обзор экологических отношений человека и слона на Малайском полуострове: адаптация к сосуществованию» . Разнообразие . 14 (1): 36. дои : 10.3390/d14010036 . ISSN 1424-2818 .
- ^ де ла Торре, JA; Вонг, EP; Лехнер, AM; Зулайха, Н.; Завави, А.; Абдул-Патах, П.; Саабан, С.; Гуссенс, Б.; Кампос-Арсеис, А. (октябрь 2021 г.). «Конфликт будет – сельскохозяйственные ландшафты являются основной, а не маргинальной средой обитания азиатских слонов» . Охрана животных . 24 (5): 720–732. Бибкод : 2021AnCon..24..720D . дои : 10.1111/acv.12668 . ISSN 1367-9430 . Архивировано из оригинала 18 марта 2024 года . Проверено 18 марта 2024 г.
- ^ Перейти обратно: а б с Баруа, М. (2010). «Чья проблема? Репрезентации конфликта между человеком и слоном в индийских и международных СМИ» . Научная коммуникация . 32 : 55–75. дои : 10.1177/1075547009353177 . S2CID 143335411 . Архивировано из оригинала 8 октября 2013 года . Проверено 29 января 2012 г.
- ^ Перейти обратно: а б Чоудри, Австралия (2004). «Конфликты между людьми и слонами в Северо-Восточной Индии» (PDF) . Человеческое измерение дикой природы . 9 (4): 261–270. Бибкод : 2004HDW.....9..261C . дои : 10.1080/10871200490505693 . S2CID 145776270 . Архивировано (PDF) из оригинала 14 января 2014 года . Проверено 27 сентября 2013 г.
- ^ Фернандо, П. (2000). «Слоны в Шри-Ланке: прошлое, настоящее и будущее» (PDF) . Лорис . 22 (2): 38–44. Архивировано (PDF) из оригинала 14 января 2014 года . Проверено 8 декабря 2012 г.
- ^ Чоудри, Австралия (2007). «Влияние пограничного забора вдоль границы между Индией и Бангладеш на передвижение слонов» (PDF) . Гаджа . 26 : 27–30. Архивировано (PDF) из оригинала 25 апреля 2012 года . Проверено 18 октября 2011 г.
- ^ Рангараджан, М., Десаи, А., Сукумар, Р., Иса, П.С., Менон, В., Винсент, С., Гангули, С., Талукдар, Б.К., Сингх, Б., Мудаппа, Д., Чоудхари , С., Прасад, АН (2010). Гаджа: Обеспечение будущего слонов в Индии. Архивировано 19 июня 2013 года в Wayback Machine . Отчет Оперативной группы по слонам. Министерство окружающей среды и лесов, Нью-Дели.
- ^ Чоудхури, А. (2004). Конфликты между человеком и слонами на северо-востоке Индии . Человеческие измерения дикой природы, 9 (4), 261–270. дои : 10.1080/10871200490505693
- ^ Вудрофф, Рози; Тергуд, Саймон; Рабиновиц, Алан (2005), Рабиновиц, Алан; Вудрофф, Рози; Тергуд, Саймон (ред.), «Влияние конфликта между человеком и дикой природой на природные системы» , Люди и дикая природа, конфликт или сосуществование? , Биология сохранения, Кембридж: Издательство Кембриджского университета, стр. 1–12, ISBN. 978-0-521-82505-4 , заархивировано из оригинала 18 ноября 2019 года , получено 19 мая 2024 года.
- ^ Бист, СС (2006). «Охрана слонов в Индии – обзор» (PDF) . Гаджа . 25 : 27–35. Архивировано (PDF) из оригинала 3 июня 2013 года . Проверено 30 января 2012 г.
- ^ Райхан Саркер, AHM, и Рёскафт, Э. (2010). Конфликты между человеком и дикой природой и варианты управления в Бангладеш, с особым упором на азиатских слонов (Elephas maximus) . Международный журнал науки о биоразнообразии, экосистемных услугах и управлении, 6 (3–4), 164–175. дои : 10.1080/21513732.2011.554867
- ^ Сантьяпиллай, Чарльз; Виджеймохан, С.; Бандара, Ганга; Атурупана, Рукмали; Диссанаяке, Навин; Прочтите, Брюс (12 октября 2010 г.). «Оценка конфликта человека и слона в Шри-Ланке» . Джей. Дж. Наук. (Био. Науки) . 39 (1): 21. doi : 10.4038/cjsbs.v39i1.2350 .
- ^ Хоар, RE (1999). Детерминанты конфликта между человеком и слонами в мозаике землепользования . Журнал прикладной экологии, 36 (5), 689–700. два : 10.1046/j.1365-2664.1999.00437.x
- ^ Расмуссен, ЛЕЛ (1999). Эволюция химических сигналов у азиатского слона Elephas maximus: поведенческие и экологические влияния . Журнал биологических наук, 24 (2), 241–251. два : 10.1007/BF02941206
- ^ Сукумар, Р. (1995). Слоны-налетчики и мошенники . Естественная история, 104 (7), 52–61.
- ^ Перейти обратно: а б Стайлз, Д. (2009). Торговля слонами и слоновой костью в Таиланде. Архивировано 4 марта 2012 года в Wayback Machine TRAFFIC Southeast Asia, Петалинг-Джая, Селангор, Малайзия.
- ^ Стайлз, Д. (2009). «Состояние торговли слоновой костью в Таиланде и Вьетнаме» (PDF) . Бюллетень ДОРОЖНОГО ДВИЖЕНИЯ . 22 (2): 83–91. Архивировано из оригинала 17 апреля 2019 года . Проверено 8 декабря 2012 г.
- ^ Флинн, В.; Стюарт-Кокс, Б. и Мелидонис, К. (2018). со шкурой; Растущий аппетит к азиатским слонам (PDF) (Отчет). Лондон: Семья слонов. Архивировано (PDF) из оригинала 26 апреля 2018 года . Проверено 26 апреля 2018 г.
- ^ Латимер Э; Цзун Дж.К.; Хигганс С.Ю.; Ричман Л.К.; Хейворд Г.С. (2011). «Обнаружение и оценка новых герпесвирусов в обычных и патологических образцах азиатских и африканских слонов: идентификация двух новых пробоскивирусов (EEHV5 и EEHV6) и двух новых гаммагерпесвирусов (EGHV3B и EGHV5)» . Ветеринарная микробиология . 147 (1–2): 28–41. дои : 10.1016/j.vetmic.2010.05.042 . ПМЦ 2976818 . ПМИД 20579821 .
- ^ Рид С.Э., Хильдебрандт Т.Б., Маркс Н., Хант М., Тай Н., Рейнс Дж.М., Шафтенаар В., Фикель Дж. (2006). «Инфекция, вызванная эндотелиотропным вирусом герпеса слона (EEHV). Первый подтвержденный ПЦР смертельный случай в Азии». Ветеринарный ежеквартальный журнал . 28 (2): 61–64. дои : 10.1080/01652176.2006.9695209 . ПМИД 16841568 . S2CID 25537428 .
- ^ Оо, Зау Мин; Аунг, Йе Хтут; Аунг, Тин Тун; Сан, Ньо; Тун, Зау Мин; Хейворд, Гэри С.; Захария, Арун (январь 2020 г.). «Слоновья эндотелиотропная герпесвирусная геморрагическая болезнь у азиатских слонят в лесозаготовительных лагерях, Мьянма» . Новые инфекционные заболевания . 26 (1): 63–69. дои : 10.3201/eid2601.190159 . ISSN 1080-6040 . ПМК 6924905 . ПМИД 31855135 .
- ^ Девкота, Р.; Брант, СВ; Тапа, А.; Локер, Э.С. (2014) [Опубликовано в Интернете в 2012 г.]. «Обмен шистосомами: шистосома слона Bivitellobilharzia nairi также заражает большого однорогого носорога (Rhinoceros unicornis) в национальном парке Читван, Непал» . Журнал гельминтологии . 88 (1): 32–40. дои : 10.1017/S0022149X12000697 . ПМИД 23113960 . S2CID 7039264 . Архивировано из оригинала 6 января 2018 года . Проверено 18 марта 2024 г.
- ^ Боуэн-Джонс Э. и Энтвистл А. (2002). «Определение подходящих флагманских видов: важность культуры и местного контекста» . Орикс . 36 (2): 189–195. дои : 10.1017/S0030605302000261 .
- ^ Баруа, М.; Тамули Дж. и Ахмед Р.А. (2010). «Мятеж или ясное плавание? Изучение роли азиатского слона как флагманского вида» . Человеческое измерение дикой природы . 15 (2): 145–160. Бибкод : 2010HDW....15..145B . дои : 10.1080/10871200903536176 . S2CID 143021377 . Архивировано из оригинала 8 октября 2013 года . Проверено 6 мая 2012 г.
- ^ Васудев, Д.; Госвами, В.; Шринивас, Н.; Ла Нам Сием, Б.; Сарма, А. (2021). «Определение важных областей связи для широко распространенного азиатского слона в заповедных ландшафтах Северо-Восточной Индии» . Разнообразие и распространение . 27 (12): 2510–2523. Бибкод : 2021DivDi..27.2510V . дои : 10.1111/ddi.13419 . JSTOR 48632844 . S2CID 244245085 .
- ^ Шелден, Д. (2020). «Всемирный день слона 2020» . Атлантика . Архивировано из оригинала 7 сентября 2020 года . Проверено 6 сентября 2020 г.
- ^ Шелден, Д. (2020). «Зоопарк OKC проведет второй ежегодный Месяц осведомленности об азиатских слонах» . Городской страж . Архивировано из оригинала 26 сентября 2020 года . Проверено 6 сентября 2020 г.
- ^ Мадхусудан, доктор медицины; Шарма, Нараян; Рагхунатх, Р.; Баскаран, Н.; Бипин, CM; Губби, Санджай; Джонсингх, AJT; Кулкарни, Джаянт; Кумара, Хоннавалли Н.; Мехта, Прачи; Пиллэй, Раджив; Сукумар, Раман (июль 2015 г.). «Распространение, относительная численность и статус сохранения азиатских слонов в Карнатаке, южная Индия» . Биологическая консервация . 187 : 34–40. Бибкод : 2015BCons.187...34M . doi : 10.1016/j.biocon.2015.04.003 . ISSN 0006-3207 . Архивировано из оригинала 21 мая 2024 года . Проверено 20 мая 2024 г.
- ^ Баскаран, Нагараджан (26 октября 2013 г.). «Обзор азиатских слонов в Западных Гатах на юге Индии: значение для сохранения экологии Западных Гат» . Журнал таксонов, находящихся под угрозой исчезновения . 5 (14): 4854–4870. дои : 10.11609/jott.o3634.4854-70 . ISSN 0974-7893 . Архивировано из оригинала 21 мая 2024 года . Проверено 20 мая 2024 г.
- ^ «Проект Слон» . Правительство Индии. Архивировано из оригинала 21 мая 2024 года . Проверено 1 апреля 2024 г.
- ^ Бандара, Ранджит; Тисделл, Клем (1 апреля 2003 г.). «Сравнение отношения сельских и городских жителей к сохранению азиатских слонов в Шри-Ланке: эмпирические данные» . Биологическая консервация . 110 (3): 327–342. дои : 10.1016/S0006-3207(02)00241-0 . ISSN 0006-3207 . Архивировано из оригинала 21 мая 2024 года . Проверено 20 мая 2024 г.
- ^ Джирават на Таланг (2020). «Это рискованно, но слоны чувствуют себя резвыми» . Почта Бангкока . Архивировано из оригинала 2 октября 2021 года . Проверено 6 сентября 2020 г.
- ^ Сеть охраны слонов. Предлагаемое восточное расширение заповедника Салакпра (PDF) . Сеть охраны слонов (отчет). Архивировано (PDF) из оригинала 21 сентября 2020 г. Проверено 6 сентября 2020 г.
- ^ Парашар, У. (2020). «Студенты Ассама и активисты по защите окружающей среды запускают онлайн-кампанию, чтобы остановить добычу угля в слоновьем заповеднике» . Индостан Таймс . Архивировано из оригинала 29 июня 2020 года . Проверено 6 сентября 2020 г.
- ^ Клубб, Р.; Роуклифф, М.; Ли, П.; Мар, КУ; Мосс, К. и Мейсон, Дж.Дж. (2008). «Компрометированная выживаемость слонов в зоопарках» (PDF) . Наука . 322 (5908): 1649. Бибкод : 2008Sci...322.1649C . дои : 10.1126/science.1164298 . HDL : 1893/974 . ПМИД 19074339 . S2CID 30756352 . Архивировано (PDF) из оригинала 14 мая 2014 года . Проверено 9 декабря 2012 года .
- ^ Рукрофт, Алан; Остерхейс, Джеймс (декабрь 2000 г.). «Уход за ногами слонов в неволе» . Слоновья нога : 1-5. дои : 10.1002/9780470292150.ch5 . ISBN 978-0-8138-2820-6 . Архивировано из оригинала 21 мая 2024 года . Проверено 19 мая 2024 г.
- ^ Визе, Р.Дж. (2000). «Азиатские слоны не являются самодостаточными в Северной Америке» . Зоопарковая биология . 19 (5): 299–309. doi : 10.1002/1098-2361(2000)19:5<299::AID-ZOO2>3.0.CO;2-Z .
- ^ Сарагусти, Дж.; Гермес, Р.; Гориц, Ф.; Шмитт, Д.Л. и Хильдебрандт, ТБ (2009). «Искажение соотношения полов при рождении и преждевременная смертность слонов» . Наука о воспроизводстве животных . 115 (1–4): 247–254. дои : 10.1016/j.anireprosci.2008.10.019 . ПМИД 19058933 . Архивировано из оригинала 21 июня 2013 года . Проверено 9 декабря 2012 года .
- ^ «Туризм стимулирует незаконную торговлю слонами в Бирме и Таиланде – видео» . Хранитель . 24 июля 2012 года. Архивировано из оригинала 2 декабря 2016 года . Проверено 11 декабря 2016 г.
- ^ Хайл, Дж. (2002). «Активисты осуждают таиландский ритуал «раздавливания» слонов» . Национальное географическое общество . Архивировано из оригинала 18 октября 2002 года . Проверено 1 октября 2014 г.
Незадолго до рассвета в отдаленных высокогорьях северного Таиланда, к западу от деревни Мае Джаем, ревет четырехлетняя слониха, когда семь деревенских мужчин вонзают ей гвозди в уши и ноги. Ее связали и обездвижили в маленькой деревянной клетке. Ее крики — единственные звуки, которые нарушают тишину сельской местности. Клетка называется «тренировочная давка». Это центральный элемент многовекового ритуала на севере Таиланда, направленного на приручение молодых слонов. Помимо избиений, дрессировщики используют лишение сна, голод и жажду, чтобы «сломить» дух слонов и заставить их покориться хозяевам.
- ^ Хан, Цзянхуа (3 июля 2019 г.). «Изучение культуры тайских слонов на основе «слоновьих метафор» тайских идиом» . Сравнительная литература: Восток и Запад . 3 (2): 148–162. дои : 10.1080/25723618.2019.1701306 . ISSN 2572-3618 .
- ^ «Таиландско-лаосский слон» . Глобус Юго-Восточной Азии . 28 февраля 2022 года. Архивировано из оригинала 1 октября 2023 года . Проверено 1 декабря 2023 г.
- ^ Сукумар, Раман (26 июля 2016 г.). «Знаковая фауна, имеющая значение наследия в Индии» (PDF) . Индийский журнал истории науки . 51 (2,2). дои : 10.16943/ijhs/2016/v51i2.2/48450 . ISSN 0019-5235 . Архивировано (PDF) из оригинала 4 февраля 2023 года . Проверено 21 мая 2024 г.
- ^ Бейкер, Ильяс; Касио, Масакадзу (2002). Гиганты в наших руках: материалы международного семинара по домашним азиатским слонам, Бангкок, Таиланд, 5–10 февраля 2001 г. Региональное отделение ФАО для Азии и Тихого океана. ISBN 978-974-90757-1-5 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ Рангараджан, М. (2001). «Лес и поле в Древней Индии» . История дикой природы Индии . Дели: Перманентный черный. стр. 1–10. ISBN 978-8-178-24140-1 . Архивировано из оригинала 17 марта 2024 года . Проверено 19 марта 2018 г.
- ^ Лирманн, Океан; Кроули, Дженни А.Х.; Зельтманн, Мартин В.; Фейе, Шерин; Нью, У. Уайт; Аунг, Хту Хту; Хут, Вин; Ладенфер, Мирка; Острие, Лия; Луммаа, Вирпи (29 июля 2021 г.). «Знакомство с хэндлером помогает улучшить рабочие характеристики в новых ситуациях у азиатских слонов, находящихся в полуневоле» . Научные отчеты . 11 (1): 15480. doi : 10.1038/s41598-021-95048-w . ISSN 2045-2322 . ПМЦ 8322261 . ПМИД 34326446 .
- ^ Винн, П. (2017). «Боевые слоны все еще существуют. Но только в одном запретном месте» . Международное общественное радио (PRI) . Архивировано из оригинала 1 марта 2017 года . Проверено 1 марта 2017 г.
- ^ Перейти обратно: а б Раджамангалам), Нилакантха (1985). Индусские предания о слонах: Слоновьи игры (Матанга-лила) Нилакантхи . Публикации Мотилала Банарсидасса. стр. 1-10 . ISBN 978-8-12080-005-2 .
- ^ Шарма, Джаита (август 2011 г.). Сад Империи: Ассам и создание Индии . Издательство Университета Дьюка. п. 25. ISBN 978-0-82235-049-1 . Архивировано из оригинала 24 мая 2022 года . Проверено 15 мая 2024 г.
- ^ Апхэм, Э. (1829). История и доктрина буддизма: популярные иллюстрации: с уведомлениями о каппуизме, или поклонении демонам, и о Бали, или планетарных заклинаниях, на Цейлоне . Лондон: Р. Акерманн.
- ^ Линг, Тревор (март 1971 г.). «Сингальский буддизм в регентских антропологических трудах: некоторые последствия» . Религия . 1 (1): 49–59. дои : 10.1016/0048-721X(71)90007-8 . ISSN 0048-721X . Архивировано из оригинала 15 мая 2024 года . Проверено 15 мая 2024 г.
- ^ «Китайский Зодиак» . Воинтурс . Архивировано из оригинала 15 сентября 2018 года . Проверено 12 июня 2017 г.
Библиография
- Дераниягала, Паулюс Эдвард Пиерис (1955). Некоторые вымершие слоны, их родственники и два ныне живущих вида . Департамент национальных музеев, Цейлон.
Дальнейшее чтение
- Бандара, Ранджит и Клем Тисделл (2004). «Чистая выгода от спасения азиатского слона: исследование политики и условной оценки» (PDF) . Экологическая экономика . 48 (1): 93–107. Бибкод : 2004EcoEc..48...93B . doi : 10.1016/j.ecolecon.2003.01.001 .
- Миалл, ЖК; Гринвуд, Ф. (1878). Анатомия индийского слона . Лондон: Макмиллан и Ко.
Внешние ссылки
- Фонд спасения слонов
- Международный фонд слонов
- ElefantAsia: Защита азиатского слона
- Азиатские слоны в зоологических садах мира
- Хранилище информации о слонах. Архивировано 11 июля 2017 г. на Wayback Machine.
- WWF — Профиль видов азиатских слонов
- Факты об азиатских слонах из национального зоопарка и веб-камера выставки азиатских слонов
- Агентство экологических расследований: незаконная торговля дикими животными : слоны

- Виды, находящиеся под угрозой исчезновения из Красного списка МСОП
- Слоны
- Виды EDGE
- Животные, находящиеся под угрозой исчезновения
- Исчезающая биота Азии
- Первые появления дошедших до нас плиоцена
- Фауна Южной Азии
- Фауна Юго-Восточной Азии
- Травоядные млекопитающие
- Млекопитающие, описанные в 1758 г.
- Млекопитающие Азии
- Национальные символы Индии
- Национальные символы Лаоса
- Национальные символы Таиланда
- Плиоценовые млекопитающие Азии
- Четвертичные животные Азии
- Виды, находящиеся под угрозой исчезновения из-за фрагментации среды обитания
- Виды, находящиеся под угрозой исчезновения из-за утраты среды обитания
- Таксоны, названные Карлом Линнеем