РНК

| Часть серии на |
| Генетика |
|---|
 |
|
|
Рибонуклеиновая кислота ( РНК )-это полимерная молекула, которая необходима для большинства биологических функций, либо путем выполнения самой функции ( некодирующей РНК ), либо путем образования матрицы для производства белков ( Мессенджер РНК ). РНК и дезоксирибонуклеиновая кислота (ДНК) являются нуклеиновыми кислотами . Нуклеиновые кислоты представляют собой одну из четырех основных макромолекул, необходимых для всех известных форм жизни . РНК собирается как цепь нуклеотидов . Клеточные организмы используют Мессенджер РНК ( мРНК ) для передачи генетической информации (с использованием азотных оснований гуанина аденина , урацила , . и цитозина , обозначенных буквами g, u, a и c), которые направляют синтез специфических белков Многие вирусы РНК кодируют их генетическую информацию, используя геном .
Некоторые молекулы РНК играют активную роль в клетках, катализируя биологические реакции, контролируя экспрессию генов или восприятие и передавая реакции на клеточные сигналы. Одним из этих активных процессов является синтез белка , универсальная функция, в которой молекулы РНК направляют синтез белков на рибосомах . В этом процессе используются молекулы переноса РНК ( тРНК ) для доставки аминокислот в рибосому , где рибосомальная РНК ( рРНК ) затем связывает аминокислоты вместе с образованием кодированных белков.
Он стал широко принятым в науке [ 1 ] То, что в начале истории жизни на Земле , до эволюции ДНК и, возможно, белковых ферментов , существовал « мир РНК », в котором РНК служила как метод хранения живых организмов для генетической информации -роль выполненной роли, выполненной Сегодня ДНК, за исключением случаев РНК -вирусов - и потенциально выполненных каталитических функций в клетках - функция, выполняемая сегодня белковыми ферментами, за заметным и важным исключением из рибосомы, которая является рибозимом .
Сравнение с ДНК
[ редактировать ]
Как ДНК, большинство биологически активных РНК, включая мРНК , тРНК , рРНК , SNRNAS и другие некодирующие РНК , содержат самообслуживаемые последовательности, которые позволяют сложить части РНК [ 6 ] и соединиться с самим собой, чтобы сформировать двойные спирали. Анализ этих РНК показал, что они сильно структурированы. В отличие от ДНК, их структуры состоят не из длинных двойных спиралей, а скорее коллекций коротких спиралей, упакованных в структуры, сродни белкам.
Таким образом, РНК могут достичь химического катализа (например, ферментов). [ 7 ] Например, определение структуры рибосомы- комплекса РНК-белка , который катализирует сборку белков,-раскрывает, что его активный сайт полностью состоит из РНК. [ 8 ]
Структура
[ редактировать ]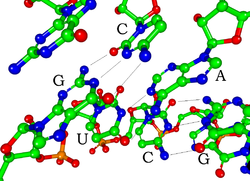
Каждый нуклеотид в РНК содержит рибозовый сахар, с углеродами, пронумерованными с 1 до 5 '. Основание прикрепляется к 1 'позиции, в целом, аденин (а), цитозин (с), гуанин (г) или урацил (u). Аденин и гуанин являются пуринами , а цитозин и урацил - пиримидины . Фосфатная группа прикреплена к 3 -дюймовому положению одной рибозы и 5 -дюймовым положением следующего. Фосфатные группы имеют отрицательный заряд, что делает РНК заряженной молекулой (Polyanion). Основы образуют водородные связи между цитозином и гуанином, между аденином и урацилом, а также между гуанином и урацилом. [ 9 ] Однако возможны другие взаимодействия, такие как группа адениновых оснований, связывающих друг друга в выпуклости, [ 10 ] или GNRA Tetraloop , который имеет базовую паре гуанина-аденина. [ 9 ]

Важным структурным компонентом РНК, который отличает ее от ДНК, является присутствие гидроксильной группы в 2 -'положении рибозного сахара . Присутствие этой функциональной группы заставляет спираль в основном принять геометрию А-формы , [ 11 ] Хотя в контекстах динуклеотидов с одной цепью РНК может редко также принимать B-форма, чаще всего наблюдаемая в ДНК. [ 12 ] Геометрия А-формы приводит к очень глубокой и узкой крупной канавке и неглубокой и широкой незначительной канавке. [ 13 ] Второе следствие присутствия 2'-гидроксильной группы состоит в том, что в конформационно гибких областях молекулы РНК (то есть не участвует в образовании двойной спирали), он может химически атаковать соседнюю фосфодиэфирную связь, чтобы расщеплять заднюю цепь. [ 14 ]
РНК транскрибируется только четырьмя основаниями (аденин, цитозин, гуанин и урацил), [ 15 ] Но эти основания и прикрепленные сахары могут быть изменены многочисленными способами, когда РНК созревают. Псевдоуридин (ψ), в котором связь между урацилом и рибозой изменяется от связи C - N на связь C - C, а риботимидин (T) обнаруживается в различных местах (наиболее заметные из них находятся в петле Tψc тРНК ) [ 16 ] Другим заметным модифицированным основанием является гипоксантин , деаминизированное адениновое основание, нуклеозид которого называется Инозин (I). Инозин играет ключевую роль в гипотезе колебания генетического кода . [ 17 ]
Существует более 100 других природных модифицированных нуклеозидов. [ 18 ] Наибольшее структурное разнообразие модификаций можно найти в тРНК , [ 19 ] в то время как псевдоуридин и нуклеозиды с 2'-О-метилрибозой, часто присутствующими в рРНК, являются наиболее распространенными. [ 20 ] Конкретные роли многих из этих модификаций в РНК не до конца понятны. Однако примечательно, что в рибосомной РНК многие посттранскрипционные модификации встречаются в высокофункциональных областях, таких как центр пептидилтрансферазы [ 21 ] и интерфейс субъединицы, подразумевая, что они важны для нормальной функции. [ 22 ]
Функциональная форма одноцепочечных молекул РНК, как и белки, часто требует определенной пространственной третичной структуры . Скафолт для этой структуры обеспечивается вторичными структурными элементами, которые являются водородными связями в молекуле. Это приводит к нескольким узнаваемым «доменам» вторичной структуры, таких как петли шпильки , выпуклости и внутренние петли . [ 23 ] Чтобы создать, то есть дизайн, РНК для любой данной вторичной структуры, двух или трех оснований было бы недостаточно, но четыре база достаточно. [ 24 ] Вероятно, именно поэтому природа «выбрала» четыре базового алфавита: менее четырех четырех не позволило бы создавать все структуры, в то время как для этого не нужно более четырех оснований. Поскольку РНК заряжена, ионы металлов, такие как Mg 2+ необходимы для стабилизации многих вторичных и третичных структур . [ 25 ]
Природным энантиомером РНК представляет собой d -РНК, состоящую из d -рибонуклеотидов. Все центры хиральности расположены в D -рибозе. Используя L -рибозу или, скорее, L -рибонуклеотиды, L -РНК может быть синтезирована. L -RNA гораздо более стабильна против деградации с помощью РНКазы . [ 26 ]
Как и другие структурированные биополимеры , такие как белки, можно определить топологию свернутой молекулы РНК. Это часто делается на основе расположения внутрицепочечных контактов в сложенной РНК, называемой топологией схемы .
Синтез
[ редактировать ]Синтез РНК обычно происходит в клеточном ядре и обычно катализируется ферментом - РНК -полимеразой - используя ДНК в качестве матрицы, процесс, известный как транскрипция . Инициирование транскрипции начинается с связывания фермента с промоторной последовательности в ДНК (обычно обнаруживается «вверх по течению» гена). Двойная спираль ДНК разматывается геликазной активностью фермента. Затем фермент прогрессирует вдоль шаблонной цепи в направлении от 3 до 5 ', синтезируя комплементарную молекулу РНК с удлинением, происходящим в направлении от 5 до 3'. Последовательность ДНК также определяет, где будет происходить прекращение синтеза РНК. [ 27 ]
Первичная транскрипта РНК часто модифицируются ферментами после транскрипции. Например, поли (а) хвост и 5-'колпачка добавляются к эукариотической пре-мРНК , а интроны удаляются сплайсосомой .
Существует также ряд РНК-зависимых РНК-полимераз , которые используют РНК в качестве их шаблона для синтеза новой цепи РНК. Например, ряд вирусов РНК (таких как полиовирус) использует этот тип фермента для воспроизведения их генетического материала. [ 28 ] Кроме того, РНК-зависимая РНК-полимераза является частью пути интерференции РНК во многих организмах. [ 29 ]
Типы РНК
[ редактировать ]Обзор
[ редактировать ]
Мессенджер РНК (мРНК) - это тип РНК, которая несет информацию от ДНК в рибосому , сайты синтеза белка ( трансляция ) в клеточной цитоплазме. Кодирующая последовательность мРНК определяет аминокислотную последовательность в белке . продуцируемом [ 30 ] Тем не менее, многие РНК не кодируют для белка (около 97% транскрипционного выхода на кодирование белка у эукариот [ 31 ] [ 32 ] [ 33 ] [ 34 ] ).
Эти так называемые не кодирующие РНК («NCRNA») могут кодироваться их собственными генами (гены РНК), но также могут вытекаться из интронов мРНК . [ 35 ] Наиболее заметными примерами некодирующих РНК являются переносная РНК (тРНК) и рибосомальная РНК (рРНК), обе из которых участвуют в процессе трансляции. [ 5 ] Существуют также некодирующие РНК, участвующие в регуляции генов, обработку РНК и другие роли. Некоторые РНК способны катализировать химические реакции, такие как резка и лигирование других молекул РНК, [ 36 ] и катализ образования пептидной связи в рибосоме ; [ 8 ] Они известны как рибозимы .
По длине
[ редактировать ]Согласно длине РНК -цепи, РНК включает в себя небольшую РНК и длинную РНК. [ 37 ] Обычно небольшие РНК имеют длину более 200 нт , а длинные РНК имеют длину более 200 нт . [ 38 ] Длинные РНК, также называемые большими РНК, в основном включают длинную некодирующую РНК (LNCRNA) и мРНК . (рРНК) 5,8S Небольшие РНК в основном включают в себя рибосомную РНК , 5S рРНК , переносную РНК (тРНК), микроРНК (miRNA), небольшую интерферирующую РНК (siRNA), небольшая нуклеолярная РНК (Snornas), Piwi-Interact RNA (PIRNA), тРНК- Полученная небольшая РНК (тсдра) [ 39 ] и небольшая РДНК-полученная РНК (SRRNA). [ 40 ] Существуют определенные исключения, как в случае 5S рРНК членов рода Halococcus ( Archaea ), которые имеют вставку, увеличивая его размер. [ 41 ] [ 42 ] [ 43 ]
В переводе
[ редактировать ]Мессенджер РНК (мРНК) несет информацию о белковой последовательности в рибосомы , фабрики синтеза белка в клетке. Он кодируется так, что каждые три нуклеотида ( кодон ) соответствуют одной аминокислоте. В эукариотических клетках, когда-то предшественника мРНК (пре-мРНК) транскрибируется из ДНК, она обрабатывается в зрелую мРНК. Это удаляет его интроны -некодирующие секции пре-мРНК. Затем мРНК экспортируется из ядра в цитоплазму , где она связана с рибосомами и транслируется в соответствующую форму белка с помощью тРНК . В прокариотических клетках, которые не имеют ядра и компартментов цитоплазмы, мРНК может связываться с рибосомами, в то время как она транскрибируется из ДНК. Через определенное количество времени сообщение разлагается в свои компонентные нуклеотиды с помощью рибонуклеаз . [ 30 ]
Передача РНК (тРНК) представляет собой небольшую РНК -цепь около 80 нуклеотидов , которая переносит специфическую аминокислоту в растущую полипептидную цепь в рибосомном сайте синтеза белка во время трансляции. Он имеет сайты для аминокислотной прикрепления и антикодонской области для распознавания кодонов , которая связывается со специфической последовательности на цепи РНК посланника посредством водородной связи. [ 35 ]
Рибосомальная РНК (рРНК) является каталитическим компонентом рибосом. РРНК является компонентом рибосомы, которая проводит перевод. Эукариотические рибосомы содержат четыре различных молекула рРНК: 18S, 5,8S, 28S и 5S рРНК. Три из молекул рРНК синтезируются в ядреве , а одна синтезируется в другом месте. В цитоплазме рибосомная РНК и белок объединяются с образованием нуклеопротеина, называемого рибосомой. Рибосома связывает мРНК и проводит синтез белка. Несколько рибосомов могут быть прикреплены к одной мРНК в любое время. [ 30 ] Почти вся РНК, обнаруженная в типичной эукариотической клетке, представляет собой рРНК.
РНК переноса-мссенджера (TMRNA) обнаружена во многих бактериях и пластидах . Он помечает белки, кодируемые мРНК, в которых отсутствуют стоп -кодоны для деградации и предотвращает остановку рибосомы. [ 44 ]
Регуляторная РНК
[ редактировать ]Самыми ранними известными регуляторами экспрессии генов были белки, известные как репрессоры и активаторы - регуляторы со специфическими короткометражными сайтами связывания в областях энхансера вблизи регулируемых генов. [ 45 ] Более поздние исследования показали, что РНК также регулируют гены. Существует несколько видов РНК-зависимых процессов у эукариот, регулирующих экспрессию генов в различных точках, таких как РНКи, гены, репрессирующие посттранскрипционно , длинные некодирующие РНК, закрывающие блоки , эпигенетически повышают усиливания РНК экспрессию генов. [ 46 ] бактерии и археи Также было показано, что используют регуляторные системы РНК, такие как бактериальные малые РНК и CRISPR . [ 47 ] Fire и Mello были награждены Нобелевской премией 2006 года по физиологии или медицине за обнаружение микроРНК (miRNAs), специфические короткие молекулы РНК, которые могут основываться на паре с мРНК. [ 48 ]
Вмешательство РНК с помощью miRNAs
[ редактировать ]Посттранскрипционные уровни экспрессии многих генов могут контролироваться РНК-интерференциями , в которых miRNAs , специфические молекулы коротких РНК, сочетаются с областями мРНК и нацелены на их для деградации. [ 49 ] Этот процесс на основе антисмысловой основы включает в себя шаги, которые сначала обрабатывают РНК, так что он может основывать паре с областью целевых мРНК. Как только пары оснований происходит, другие белки направляют мРНК быть разрушенной нуклеазами . [ 46 ]
Длинные некодирующие РНК
[ редактировать ]Затем, чтобы быть связанными с регулированием, были XIST и другие длинные некодирующие РНК, связанные с инактивацией Х -хромосомы . Их роли, поначалу таинственными, были показаны Джинни Т. Ли и другими, что является молчанием блоков хроматина посредством рекрутирования комплекса поликомба , чтобы Мессенджер РНК не могла быть транскрибирована из них. [ 50 ] Дополнительные LNCRNAS, в настоящее время определяемые как РНК из более чем 200 пар оснований, которые, по -видимому, не имеют потенциала кодирования, [ 51 ] были обнаружены связаны с регуляцией стволовых клеток плюрипотентности и деления клеток . [ 51 ]
Enhancer RNAS
[ редактировать ]Третья крупная группа регулирующих РНК называется Enhancer RNA . [ 51 ] В настоящее время неясно, являются ли они уникальной категорией РНК различной длины или представляют собой отдельную подмножество LNCRNAS. В любом случае, они транскрибируются из усилителей , которые известны регуляторные сайты в ДНК вблизи генов, которые они регулируют. [ 51 ] [ 52 ] Они активируют транскрипцию генов (и) под контролем усилителя, из которого они транскрибируются. [ 51 ] [ 53 ]
Регуляторная РНК у прокариот
[ редактировать ]Сначала считалось, что регуляторная РНК является эукариотическим явлением, частью объяснения того, почему было замечено гораздо больше транскрипции у более высоких организмов, чем было предсказано. Но как только исследователи начали искать возможные регуляторы РНК в бактериях, они также появились там, называемые маленькой РНК (SRNA). [ 54 ] [ 47 ] В настоящее время повсеместная природа систем регуляции РНК -регуляции генов обсуждалась как поддержка мировой теории РНК . [ 46 ] [ 55 ] Существуют признаки того, что энтеробактериальные SRNAs участвуют в различных клеточных процессах и, по -видимому, играют значительную роль в стрессовых реакциях, таких как мембранная стресс, стресс голода, фосфосугарный стресс и повреждение ДНК. Кроме того, было высказано предположение, что SRNAs были разработаны, чтобы играть важную роль в стрессовых реакциях из -за их кинетических свойств, которые позволяют быстро реагировать и стабилизацию физиологического состояния. [ 2 ] Бактериальные малые РНК обычно действуют посредством антисмыслового спаривания с мРНК для подавления его трансляции, либо путем влияния на стабильность, либо влияя на цис-связывающую способность. [ 46 ] Riboswitches также были обнаружены. Они представляют собой цис-действий регуляторных последовательностей РНК, действующих аллостерически . Они меняют форму, когда они связывают метаболиты, так что они получают или теряют способность связывать хроматин, чтобы регулировать экспрессию генов. [ 56 ] [ 57 ]
Археа также имеет системы регуляторной РНК. [ 58 ] Система CRISPR, недавно используемая для редактирования ДНК in situ , действует посредством регулирующих РНК в археи и бактериях для обеспечения защиты от вирусных захватчиков. [ 46 ] [ 59 ]
В обработке РНК
[ редактировать ]
Многие РНК участвуют в изменении других РНК. Интроны сплайсируются небольших ядерных из пре-мРНК сплайсосомами , , которые содержат несколько РНК (SNRNA) [ 5 ] Или интроны могут быть рибозимами, которые сами по себе сплачиваются. [ 60 ] РНК также может быть изменена путем изменения его нуклеотидов на нуклеотиды, отличные A , C , G и U. от У эукариот модификации РНК -нуклеотидов в целом направлены небольшими нуклеолярными РНК (Snorna; 60–300 нт), [ 35 ] найдено в телах ядрышко и каджала . Снорны ассоциируются с ферментами и направляют их к месту на РНК, базовой, к этой РНК. Эти ферменты затем выполняют модификацию нуклеотидов. РРНК и тРНК широко модифицированы, но SNRNAS и мРНК также могут быть целью модификации базы. [ 61 ] [ 62 ] РНК также может быть метилирована. [ 63 ] [ 64 ]
РНК геномы
[ редактировать ]Как ДНК, РНК может нести генетическую информацию. РНК -вирусы имеют геномы, состоящие из РНК, которая кодирует ряд белков. Вирусный геном реплицируется некоторыми из этих белков, в то время как другие белки защищают геном, когда вирусная частица перемещается к новой клетке -хозяину. Вироиды являются еще одной группой патогенных микроорганизмов, но они состоят только из РНК, не кодируют ни одного белка и воспроизводятся полимеразой растений -хозяины. [ 65 ]
В обратной транскрипции
[ редактировать ]Обратная транскрибирующая вирусы воспроизводит свои геномы путем обратной транскрибирования копий ДНК из их РНК; Эти копии ДНК затем транскрибируются в новую РНК. Ретротранспозоны также распространяются путем копирования ДНК и РНК друг с другом, [ 66 ] и теломераза содержит РНК, которая используется в качестве шаблона для построения концов эукариотических хромосом . [ 67 ]
Двухцепочечная РНК
[ редактировать ]
Двойная полоса РНК (дцРНК) представляет собой РНК с двумя комплементарными цепями, сходными с ДНК, обнаруженной во всех клетках, но с заменой тимина урацилом и добавлением одного атома кислорода. ДсРНК образует генетический материал некоторых вирусов ( двухцепочечные РНК-вирусы ). Двухцепочечная РНК, такая как вирусная РНК или миРНК , может запускать интерференцию РНК у эукариот , а также интерферона реакцию у позвоночных . [ 68 ] [ 69 ] [ 70 ] [ 71 ] У эукариот двухцепочечная РНК (дцРНК) играет роль в активации врожденной иммунной системы против вирусных инфекций. [ 72 ]
Круглая РНК
[ редактировать ]В конце 1970 -х годов было показано, что существует одноцепочечная ковалентно закрытая, то есть круговая форма РНК, экспрессируемой во всем царстве животных и растений (см. Circrna ). [ 73 ] Считается, что Circrnas возникают с помощью реакции «обратной сферы», когда сплайсосома соединяет акцептор вверх по течению 3 'к месту сплайсинга вниз по течению 5'. До сих пор функция Circrnas в значительной степени неизвестна, хотя для немногих примеров была продемонстрирована активность губки микроРНК.
Ключевые открытия в биологии РНК
[ редактировать ]
Исследования по РНК привели ко многим важным биологическим открытиям и многочисленным Нобелевским призам . Нуклеиновые кислоты были обнаружены в 1868 году Фридрихом Мишером , который назвал материал «ядром» с тех пор, как он был обнаружен в ядре . [ 74 ] Позже было обнаружено, что прокариотические клетки, которые не имеют ядра, также содержат нуклеиновые кислоты. Роль РНК в синтезе белка была подозревана уже в 1939 году. [ 75 ] Северо Очоа выиграл Нобелевскую премию 1959 года по медицине (общаясь с Артуром Корнбергом ) после того, как он обнаружил фермент, который может синтезировать РНК в лаборатории. [ 76 ] Тем не менее, было показано, что фермент, обнаруженный OCHOA ( полинуклеотидфосфорилаза ), является ответственным за деградацию РНК, а не синтез РНК. В 1956 году Алекс Рич и Дэвид Дэвис гибридировали две отдельные нити РНК, чтобы сформировать первый кристалл РНК, структуру которой можно определить с помощью рентгеновской кристаллографии. [ 77 ]
Последовательность 77 нуклеотидов дрожжевой тРНК была обнаружена Робертом В. Холли в 1965 году, [ 78 ] Победа Холли Нобелевская премия 1968 года в области медицины (поделилась Хар Гобинд Хорана и Маршалл Ниренберг ).
В начале 1970 -х годов были обнаружены ретровирусы и обратная транскриптаза , впервые показывая, что ферменты могут копировать РНК в ДНК (противоположность обычному пути для передачи генетической информации). За эту работу Дэвид Балтимор , Ренато Дульбекко и Говард Тейн были удостоены Нобелевской премии в 1975 году. В 1976 году Уолтер Фирс и его команда определили первую полную нуклеотидную последовательность генома РНК -вируса, бактериофага MS2 . [ 79 ]
В 1977 году интроны и сплайсинг РНК были обнаружены как у вирусов млекопитающих, так и в клеточных генах, что привело к тому, что Нобелевский Нобель в 1993 году Филипп Шарп и Ричард Робертс . Каталитические молекулы РНК ( рибозимы ) были обнаружены в начале 1980 -х годов, что привело к Нобелевской премии 1989 года Томасу Чеху и Сидни Альтману . было обнаружено В 1990 году в Petunia , что введенные гены могут замолчать сходные гены собственных растений, которые теперь известны как результат интерференции РНК . [ 80 ] [ 81 ]
Примерно в то же время было обнаружено, что 22 НТ -РНК, теперь называемые микроРНК , играют роль в развитии C. elegans . [ 82 ] Исследования по вмешательству РНК получили Нобелевскую премию за Эндрю Фьер и Крейг Мелло в 2006 году, а также еще одну Нобелевскую Нобелевскую работу по транскрипции РНК для Роджера Корнберга в том же году. Обнаружение регуляторных РНК генов привело к попыткам разработать лекарства из РНК, таких как siRNA , чтобы заставить замолчать гены. [ 83 ] В дополнение к Нобелевским призам для исследований по РНК, в 2009 году он был присужден за выяснение атомной структуры рибосомы Венки Рамакришнану , Томасу А. Стейцу и Ада Йонату . В 2023 году Нобелевская премия по физиологии или медицине была присуждена Каталин Карико и Дрю Вайсман за их открытия, касающиеся модифицированных нуклеозидов , которые позволили разработать эффективные вакцины мРНК против Covid-19. [ 84 ] [ 85 ] [ 86 ]
Актуальность для химии пребиотики и абиогенеза
[ редактировать ]В 1968 году Карл Воэз предположил, что РНК может быть каталитической, и предположила, что самые ранние формы жизни (самореплицирующиеся молекулы) могли опираться на РНК как для переноса генетической информации, так и для катализирования биохимических реакций- мира РНК . [ 87 ] [ 88 ] В мае 2022 года ученые обнаружили, что РНК может спонтанно формироваться на пребиотическом базальтовом стекле лавы , предположительно, была в изобилии на ранней земле . [ 89 ] [ 90 ]
В марте 2015 года ДНК и РНК -нуклеобазы , включая урацил , цитозин и тимин лаборатории были образованы в лаборатории в лаборатории в с использованием стартовых химических веществ, таких как пиримидин , органическое соединение, обычно встречающее в метеоритах . Пиримидин , как и полициклические ароматические углеводороды (ПАУ), является одним из самых богатых углеродами соединений, обнаруженными во вселенной , и, возможно, были образованы в красных гигантах или в межзвездных пыли и газовых облаках. [ 91 ] В июле 2022 года астрономы сообщили о огромных количествах пребиотических молекул , включая возможные предшественники РНК, в галактическом центре Галактики Млечного Пути . [ 92 ] [ 93 ]
Медицинские заявки
[ редактировать ]РНК, первоначально считающаяся неподходящей для терапевтических средств из-за его короткого периода полураспада, стала полезной посредством достижений в стабилизации. Терапевтические применения возникают как РНК складывается в сложные конформации и связывает белки, нуклеиновые кислоты и мелкие молекулы с образованием каталитических центров. [ 94 ] Считается, что вакцины на основе РНК легче производить, чем традиционные вакцины, полученные из убитых или измененных патогенных микроорганизмов, потому что для выращивания и изучения патогена могут потребоваться месяцы или годы. Маленькие молекулы с обычными терапевтическими свойствами могут нацелиться на РНК и структуры ДНК, тем самым обрабатывая новые заболевания. Тем не менее, исследования мало на мелких молекулах, нацеленных на РНК и одобренные лекарства для заболеваний человека. Рибавирин, Бранаплам и Аталурен в настоящее время являются доступными лекарствами, которые стабилизируют двухцепочечные структуры РНК и контролируют сплайсинг при различных расстройствах. [ 95 ] [ 96 ]
МРНК, кодирующие белок, стали новыми терапевтическими кандидатами, причем замена РНК особенно полезна для краткой, но проливного белка. [ 97 ] Странскрибированные мРНК in vitro (IVT-MRNA) использовались для доставки белков для регенерации костей, плюрипотентности и функции сердца на животных моделях. [ 98 ] [ 99 ] [ 100 ] [ 101 ] [ 102 ] МиРНК, короткие молекулы РНК, играют решающую роль в врожденной защите от вирусов и структуры хроматина. Они могут быть искусственно введены для тишины специфических генов, что делает их ценными для исследований функции генов, терапевтической проверки целей и разработке лекарств. [ 97 ]
Вакцины мРНК стали важным новым классом вакцин, используя мРНК для производства белков, которые вызывают иммунный ответ. Их первое успешное крупномасштабное применение было в форме вакцин Covid-19 во время пандемии Covid-19 .
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Copley SD, Smith E, Markowitz HJ (декабрь 2007 г.). «Происхождение мира РНК: коэволюция генов и метаболизма». Биоорганическая химия . 35 (6): 430–443. doi : 10.1016/j.bioorg.2007.08.001 . PMID 17897696 .
Предложение о том, что жизнь на земле возникло из мира РНК, широко распространено.
- ^ Jump up to: а беременный «РНК: универсальная молекула» . Университет Юты . 2015.
- ^ «Нуклеотиды и нуклеиновые кислоты» (PDF) . Калифорнийский университет, Лос -Анджелес . Архивировано из оригинала (PDF) 2015-09-23 . Получено 2015-08-26 .
- ^ Shukla RN (2014). Анализ хромосом . Agrotech Press. ISBN 978-93-84568-17-7 . [ Постоянная мертвая ссылка ]
- ^ Jump up to: а беременный в Berg JM, Tymoczko JL, Stryer L (2002). Биохимия (5 -е изд.). WH Freeman and Company. С. 118–19, 781–808. ISBN 978-0-7167-4684-3 Полем OCLC 179705944 .
- ^ Tinoco I, Bustamante C (октябрь 1999 г.). «Как складывается РНК». Журнал молекулярной биологии . 293 (2): 271–81. doi : 10.1006/jmbi.1999.3001 . PMID 10550208 .
- ^ Хиггс П.Г. (август 2000). «РНК вторичная структура: физические и вычислительные аспекты». Ежеквартальные обзоры биофизики . 33 (3): 199–253. doi : 10.1017/s0033583500003620 . PMID 11191843 . S2CID 37230785 .
- ^ Jump up to: а беременный Nissen P, Hansen J, Ban N, Moore PB, Steitz TA (август 2000 г.). «Структурная основа активности рибосом в синтезе пептидной связи». Наука . 289 (5481): 920–30. Bibcode : 2000sci ... 289..920N . doi : 10.1126/science.289.5481.920 . PMID 10937990 .
- ^ Jump up to: а беременный Ли JC, Gutell RR (декабрь 2004 г.). «Разнообразие конформаций базовой пары и их возникновение в структуре рРНК и структурных мотивах РНК». Журнал молекулярной биологии . 344 (5): 1225–49. doi : 10.1016/j.jmb.2004.09.072 . PMID 15561141 .
- ^ Барсишевский Дж., Фредерик Б., Кларк С. (1999). РНК биохимия и биотехнология . Спрингер. С. 73–87. ISBN 978-0-7923-5862-6 Полем OCLC 52403776 .
- ^ Salazar M, Fedoroff Oy, Miller JM, Ribeiro NS, Reid BR (апрель 1993 г.). «ДНК цепь в гибридных дуплексах ДНК не является ни B-формой, ни формой в растворе». Биохимия . 32 (16): 4207–15. doi : 10.1021/bi00067a007 . PMID 7682844 .
- ^ Седова А., Банавали Н.К. (февраль 2016 г.). «РНК приближается к B-форме в сложенных контекстах динуклеотидов с одной нити». Биополимеры . 105 (2): 65–82. doi : 10.1002/bip.22750 . PMID 26443416 . S2CID 35949700 .
- ^ Hermann T, Patel DJ (март 2000 г.). «РНК выпуклости как архитектурные мотивы и мотивы признания» . Структура 8 (3): R47–54. doi : 10.1016/s0969-2126 (00) 00110-6 . PMID 10745015 .
- ^ Миккола С., Стенман Е., Нурми К., Юсефи-Салакде Е., Стрёмберг Р., Лоннберг Х (1999). «Механизм иона металла способствовал расщеплению РНК -фосфодиэфирных связей включает в себя общий кислотный катализ со стороны металла -иона после ухода уходящей группы». Журнал Химического общества, Печкин Транзакции 2 (8): 1619–26. doi : 10.1039/a903691a .
- ^ Jankowski JA, Polak JM (1996). Клинический анализ генов и манипуляции: инструменты, методы и устранение неполадок . Издательство Кембриджского университета. п. 14 ISBN 978-0-521-47896-0 Полем OCLC 33838261 .
- ^ Yu Q, Morrow CD (май 2001 г.). «Идентификация критических элементов в стволе акцептора тРНК и петлей T (PSI) C, необходимой для инфекционности вируса иммунодефицита человека типа 1» . Журнал вирусологии . 75 (10): 4902–6. doi : 10.1128/JVI.75.10.4902-4906.2001 . PMC 114245 . PMID 11312362 .
- ^ Elliott MS, Trewyn RW (февраль 1984 г.). «Инозиновый биосинтез в РНК переноса путем ферментативной вставки гипоксантина» . Журнал биологической химии . 259 (4): 2407–10. doi : 10.1016/s0021-9258 (17) 43367-9 . PMID 6365911 .
- ^ Cantara WA, Crain PF, Rozenski J, McCloskey JA, Harris KA, Zhang X, Vendeix FA, Fabris D, Agris PF (январь 2011 г.). «База данных модификации РНК, RNAMDB: 2011 Обновление» . Исследование нуклеиновых кислот . 39 (выпуск базы данных): D195-201. doi : 10.1093/nar/gkq1028 . PMC 3013656 . PMID 21071406 .
- ^ Söll D, Rajbhandy U (1995). ТРНК: структура, биосинтез и функция ASM Press. П. 165. ISBN 978-1-55581-073-3 Полем OCLC 183036381 .
- ^ KISS T (июль 2001 г.). «Небольшая нуклеолярная РНК-подведенная посттранскрипционная модификация клеточных РНК» . Embo Journal . 20 (14): 3617–22. doi : 10.1093/emboj/20.14.3617 . PMC 125535 . PMID 11447102 .
- ^ Tirumalai MR, Rivas M, Tran Q, Fox GE (ноябрь 2021 г.). «Центр пептидилтрансферазы: окно до прошлого» . Microbiol Mol Biol Rev. 85 (4): E0010421. doi : 10.1128/mmbr.00104-21 . PMC 8579967 . PMID 34756086 .
- ^ King TH, Liu B, McCully RR, Fournier MJ (февраль 2003 г.). «Структура и активность рибосом изменяются в клетках, в которых отсутствуют Snornps, которые образуют псевдоридины в центре пептидилтрансферазы» . Молекулярная клетка . 11 (2): 425–35. doi : 10.1016/s1097-2765 (03) 00040-6 . PMID 12620230 .
- ^ Мэтьюз Д.Х., Дисней М.Д., Чайлдс Дж.Л., Шредер С.Дж., Зукер М., Тернер Д.Х. (май 2004 г.). «Включение ограничений химической модификации в алгоритм динамического программирования для прогнозирования РНК -вторичной структуры» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (19): 7287–92. Bibcode : 2004pnas..101.7287m . doi : 10.1073/pnas.0401799101 . PMC 409911 . PMID 15123812 .
- ^ Burghardt B, Hartmann AK (февраль 2007 г.). «РНК вторичная конструкция» . Физический обзор e . 75 (2): 021920. ARXIV : Физика/0609135 . BIBCODE : 2007FRVE..75B1920B . doi : 10.1103/physreve.75.021920 . PMID 17358380 . S2CID 17574854 .
- ^ Tan ZJ, Chen SJ (июль 2008 г.). «Солевая зависимость от стабильности шпильки нуклеиновой кислоты» . Биофизический журнал . 95 (2): 738–52. Bibcode : 2008bpj .... 95..738t . doi : 10.1529/biophysj.108.131524 . PMC 2440479 . PMID 18424500 .
- ^ Vater A, Klussmann S (январь 2015 г.). «Поворачивая зеркало-изображение олигонуклеотиды в лекарства: эволюция терапии Spiegelmer (®)» . Drug Discovery сегодня . 20 (1): 147–55. doi : 10.1016/j.drudis.2014.09.004 . PMID 25236655 .
- ^ Nudler E, Gottesman Me (август 2002 г.). «Транскрипционное прекращение и анти-концевые в E. coli». Гены к клеткам . 7 (8): 755–68. doi : 10.1046/j.1365-2443.2002.00563.x . PMID 12167155 . S2CID 23191624 .
- ^ Hansen JL, Long Am, Schultz SC (август 1997 г.). «Структура РНК-зависимой РНК-полимеразы полиовируса» . Структура 5 (8): 1109–22. doi : 10.1016/s0969-2126 (97) 00261-x . PMID 9309225 .
- ^ Ahlquist P (май 2002). «РНК-зависимые РНК-полимеразы, вирусы и молчание РНК». Наука . 296 (5571): 1270–73. Bibcode : 2002sci ... 296.1270a . doi : 10.1126/science.1069132 . PMID 12016304 . S2CID 42526536 .
- ^ Jump up to: а беременный в Купер GC, Hausman RE (2004). Клетка: молекулярный подход (3 -е изд.). Синауэр. С. 261–76, 297, 339–44. ISBN 978-0-87893-214-6 Полем OCLC 174924833 .
- ^ Mattick JS, Gagen MJ (сентябрь 2001 г.). «Эволюция контролируемых многозадачных генных сетей: роль интронов и других некодирующих РНК в развитии сложных организмов» . Молекулярная биология и эволюция . 18 (9): 1611–30. doi : 10.1093/oxfordjournals.molbev.a003951 . PMID 11504843 .
- ^ Mattick JS (ноябрь 2001 г.). «Некодирующие РНК: архитекторы эукариотической сложности» . Embo сообщает . 2 (11): 986–91. doi : 10.1093/embo-reports/kve230 . PMC 1084129 . PMID 11713189 .
- ^ Mattick JS (октябрь 2003 г.). «Вызов догме: скрытый слой непротеиновых РНК в сложных организмах» (PDF) . Биологии . 25 (10): 930–39. Citeseerx 10.1.1.476.7561 . doi : 10.1002/bies.10332 . PMID 14505360 . Архивировано из оригинала (PDF) на 2009-03-06.
- ^ Mattick JS (октябрь 2004 г.). «Скрытая генетическая программа сложных организмов». Scientific American . 291 (4): 60–67. Bibcode : 2004sciam.291d..60m . doi : 10.1038/Scientificamerican1004-60 . PMID 15487671 . [ мертвая ссылка ]
- ^ Jump up to: а беременный в Wirta W (2006). Добыча транскриптома - методы и приложения . Стокгольм: Школа биотехнологии, Королевский технологический институт. ISBN 978-91-7178-436-0 Полем OCLC 185406288 .
- ^ Росси JJ (июль 2004 г.). «Диагностика рибозима достигает совершеннолетия» . Химия и биология . 11 (7): 894–95. doi : 10.1016/j.chembiol.2004.07.002 . PMID 15271347 .
- ^ Storz G (май 2002). «Расширяющаяся вселенная некодирующих РНК». Наука . 296 (5571): 1260–63. Bibcode : 2002sci ... 296.1260s . doi : 10.1126/science.1072249 . PMID 12016301 . S2CID 35295924 .
- ^ Fatica A, Bozzoni I (январь 2014 г.). «Длинные некодирующие РНК: новые игроки в дифференциации и развитии клеток» . Nature Reviews Genetics . 15 (1): 7–21. doi : 10.1038/nrg3606 . PMID 24296535 . S2CID 12295847 . [ Постоянная мертвая ссылка ]
- ^ Chen Q, Yan M, Cao Z, Li X, Zhang Y, Shi J, et al. (Январь 2016). «Сперматозоиды способствуют наследству между поколениями приобретенного метаболического расстройства» (PDF) . Наука . 351 (6271): 397–400. Bibcode : 2016sci ... 351..397c . doi : 10.1126/science.aad7977 . PMID 26721680 . S2CID 21738301 .
- ^ Вей Х, Чжоу Б., Чжан Ф., Ту Й, Ху Й, Чжан Б., Чжай Q (2013). «Профилирование и идентификация мелких РНК, полученных из RDNA, и их потенциальные биологические функции» . Plos один . 8 (2): E56842. BIBCODE : 2013PLOSO ... 856842W . doi : 10.1371/journal.pone.0056842 . PMC 3572043 . PMID 23418607 .
- ^ Luehrsen KR, Nicholson DE, Eubanks DC, Fox GE (1981). «Archaebacterial 5s рРНК содержит длинную последовательность вставки». Природа . 293 (5835): 755–756. Bibcode : 1981natur.293..755L . doi : 10.1038/2937555A0 . PMID 6169998 . S2CID 4341755 .
- ^ Stan-Lotter H, McGenity TJ, Legat A, Denner EB, Glaser K, Stetter KO, Wanner G (1999). «Очень похожие штаммы Halococcus salifodinae обнаружены в географически разделенных пермо-трисосических отложениях соли» . Микробиология . 145 (Pt 12): 3565–3574. doi : 10.1099/00221287-145-12-3565 . PMID 10627054 .
- ^ Тирумалай М.Р., Келбер Дж. Т., Парк Д.Р., Тран К., Фокс Г.Е. (август 2020 г.). «Криоэлектронная микроскопия визуализация большой вставки в рибосомальную РНК 5S чрезвычайно галофильной археоны Halococcus morrhuae » . Febs Open Bio . 10 (10): 1938–1946. doi : 10.1002/2211-5463.12962 . PMC 7530397 . PMID 32865340 .
- ^ Gueneau de Novoa P, Williams KP (январь 2004 г.). «Веб -сайт TMRNA: восстановительная эволюция тмРНК в пластидах и других эндосимбионтах» . Исследование нуклеиновых кислот . 32 (проблема базы данных): D104–08. doi : 10.1093/nar/gkh102 . PMC 308836 . PMID 14681369 .
- ^ Джейкоб Ф., Монод Дж. (1961). «Генетические регуляторные механизмы в синтезе белков». Журнал молекулярной биологии . 3 (3): 318–56. doi : 10.1016/s0022-2836 (61) 80072-7 . PMID 13718526 . S2CID 19804795 .
- ^ Jump up to: а беременный в дюймовый и Моррис К., Маттик Дж. (2014). «Рост регуляторной РНК» . Nature Reviews Genetics . 15 (6): 423–37. doi : 10.1038/nrg3722 . PMC 4314111 . PMID 24776770 .
- ^ Jump up to: а беременный Готтесман С. (2005). «Микробы для микробов: некодирующие регуляторные РНК у бактерий». Тенденции в генетике . 21 (7): 399–404. doi : 10.1016/j.tig.2005.05.008 . PMID 15913835 .
- ^ «Нобелевская премия по физиологии или медицине 2006». Nobelprize.org. Nobel Media AB 2014. Веб. 6 августа 2018 года .
- ^ Огонь и др. (1998). «Мощное и специфическое генетическое вмешательство путем двойного мельчайшего РНК у Ceanorhabditis elegans» . Природа . 391 (6669): 806–11. Bibcode : 1998natur.391..806f . doi : 10.1038/35888 . PMID 9486653 . S2CID 4355692 .
- ^ Zhao J, Sun BK, Erwin JA, Song JJ, Lee JT (2008). «Поликомбные белки, нацеленные на короткую повторную РНК на хромосому мыши» . Наука . 322 (5902): 750–56. Bibcode : 2008Sci ... 322..750Z . doi : 10.1126/science.1163045 . PMC 2748911 . PMID 18974356 .
- ^ Jump up to: а беременный в дюймовый и Ринн Дж.Л., Чанг Хи (2012). «Регуляция генома длинными некодирующими РНК» . Анну. Преподобный Биохим . 81 : 1–25. doi : 10.1146/annurev-biochem-051410-092902 . PMC 3858397 . PMID 22663078 .
- ^ Taft RJ, Kaplan CD, Simons C, Mattick JS (2009). «Эволюция, биогенез и функция РНК, связанных с промотором» . Клеточный цикл . 8 (15): 2332–38. doi : 10.4161/cc.8.15.9154 . PMID 19597344 .
- ^ Orom UA, Derrien T, Beringer M, Gumireddy K, Gardini A, et al. (2010). « Длинные некодирующие РНК с энхансерной функцией в клетках человека» . Клетка . 143 (1): 46–58. doi : 10.1016/j.cell.2010.09.001 . PMC 4108080 . PMID 20887892 .
- ^ Egh Wagner, P Romby. (2015). «Маленькие РНК в бактериях и археи: кто они, что они делают и как они это делают». Достижения в области генетики (том 90, с. 133–208).
- ^ JW Nelson, RR Breaker (2017) «Потерянный язык мира РНК». Наука Сигнал 10 , eaam8812 1–11.
- ^ Winklef WC (2005). «Рибосвитчи и роль некодирующих РНК в бактериальном метаболическом контроле». Карт Мнение Химический Биол . 9 (6): 594–602. doi : 10.1016/j.cbpa.2005.09.016 . PMID 16226486 .
- ^ Такер BJ, Breaker RR (2005). «Riboswitchs как универсальные элементы контроля генов». Карт Мнение Структура Биол . 15 (3): 342–48. doi : 10.1016/j.sbi.2005.05.003 . PMID 15919195 .
- ^ Mojica FJ, Diez-Villasenor C, Soria E, Juez G (2000). " " Биологическое значение семейства регулярно распределенных повторений в геномах археи, бактерий и митохондрий » . Мол Микробиол . 36 (1): 244–46. doi : 10.1046/j.1365-2958.2000.01838.x . PMID 10760181 . S2CID 22216574 .
- ^ Brouns S, Joruns MM, Landgren M, Wesa E, Section R, Spartiary A, Dickman M, Makarova K, Change E, Der East JV (2008). «Убьем Crisr RNAS Guide Antiviral определяет в прокаритах» . Наука . 321 (5891): 960–64. Код BIB : 2008Sci… 321..960b . doi : 10,1126/science.1159689 . PMC 5898235 . PMID 187039 .
- ^ Steitz Ta, Steitz Ja (июль 1993 г.). «Общий двухметальный механизм для каталитической РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (14): 6498–502. Bibcode : 1993pnas ... 90.6498s . doi : 10.1073/pnas.90.14.6498 . PMC 46959 . PMID 8341661 .
- ^ Xie J, Zhang M, Zhou T, Hua X, Tang L, Wu W (январь 2007 г.). «Sno/Scarnabase: кураторская база данных для небольших нуклеолярных РНК и специфичных для тела Cajal RNA» . Исследование нуклеиновых кислот . 35 (проблема базы данных): D183–87. doi : 10.1093/nar/gkl873 . PMC 1669756 . PMID 17099227 .
- ^ Omer AD, Ziesche S, Decatur WA, Fournier MJ, Dennis PP (май 2003 г.). «Модифицирующие РНК машины в археи». Молекулярная микробиология . 48 (3): 617–29. doi : 10.1046/j.1365-2958.2003.03483.x . PMID 12694609 . S2CID 20326977 .
- ^ Cavaillé J, Nicoloso M, Bachellerie JP (октябрь 1996 г.). «Целевое метилирование рибозы РНК in vivo, направленное на индивидуальные антисмысловые руководства РНК» . Природа . 383 (6602): 732–35. Bibcode : 1996natur.383..732c . doi : 10.1038/383732A0 . PMID 8878486 . S2CID 4334683 .
- ^ Kiss-lászló Z, Henry Y, Bachellerie JP, Caizergues-Ferrer M, Kiss T (июнь 1996 г.). «Сайт-специфическое метилирование рибозы прерибосомной РНК: новая функция для небольших нуклеолярных РНК» . Клетка . 85 (7): 1077–88. doi : 10.1016/s0092-8674 (00) 81308-2 . PMID 8674114 . S2CID 10418885 .
- ^ Daròs JA, Elena SF, Flores R (июнь 2006 г.). «Вироиды: нить Ариадны в лабиринт РНК» . Embo сообщает . 7 (6): 593–98. doi : 10.1038/sj.embor.7400706 . PMC 1479586 . PMID 16741503 .
- ^ Kalendar R, Vaut CM, Peleg O, Anamthawat-Jonsson K, Bolshoy A, Schulman AH (март 2004 г.). «Большие производные ретротранспозона: обильные, консервативные, но неавтономные ретроэлементы ячменя и родственных геномов» . Генетика . 166 (3): 1437–50. doi : 10.1534/Genetics.166.3.1437 . PMC 1470764 . PMID 15082561 .
- ^ Podlevsky JD, Bley CJ, Omana RV, Qi X, Chen JJ (январь 2008 г.). «База данных теломеразы» . Исследование нуклеиновых кислот . 36 (проблема базы данных): D339–43. doi : 10.1093/nar/gkm700 . PMC 2238860 . PMID 18073191 .
- ^ Блевинс Т., Раджесваран Р., Шивапрасад П.В., Беназаряны Д., Си-Аммур А., Парк Х.С., Васкес Ф., Робертсон Д., Мейнс Ф., Хон Т., Пуггин М.М. (2006). «Четыре изделия из растений опосредуют вирусный малый РНК -биогенез, а ДНК -вирус индуцировал молчание» . Исследование нуклеиновых кислот . 34 (21): 6233–46. doi : 10.1093/nar/gkl886 . PMC 1669714 . PMID 17090584 .
- ^ Jana S, Chakraborty C, Nandi S, Deb JK (ноябрь 2004 г.). «Вмешательство РНК: потенциальные терапевтические мишени». Прикладная микробиология и биотехнология . 65 (6): 649–57. doi : 10.1007/s00253-004-1732-1 . PMID 15372214 . S2CID 20963666 .
- ^ Virol, J (май 2006 г.). «Двойная цепочка РНК продуцируется вирусами РНК с положительной цепью и вирусами ДНК, но не в обнаруживаемых количествах с помощью вирусов РНК с отрицательной цепью» . Журнал вирусологии . 80 (10): 5059–5064. doi : 10.1128/jvi.80.10.5059-5064.2006 . PMC 1472073 . PMID 16641297 .
- ^ Schultz U, Kaspers B, Staeheli P (май 2004). «Система интерферонов не млекопитающих позвоночных». Развитие и сравнительная иммунология . 28 (5): 499–508. doi : 10.1016/j.dci.2003.09.009 . PMID 15062646 .
- ^ Уайтхед К.А., Далман Дж., Лангер Р.С., Андерсон Д.Г. (2011). «Сильком или стимуляция? Доставка миРНК и иммунная система». Ежегодный обзор химической и биомолекулярной инженерии . 2 : 77–96. doi : 10.1146/annurev-chembioeng-061010-114133 . PMID 22432611 .
- ^ HSU MT, Coca-Prados M (июль 1979 г.). «Электронные микроскопические доказательства круглой формы РНК в цитоплазме эукариотических клеток». Природа . 280 (5720): 339–40. Bibcode : 1979natur.280..339H . doi : 10.1038/280339a0 . PMID 460409 . S2CID 19968869 .
- ^ Дам Р (февраль 2005 г.). «Фридрих Мишер и открытие ДНК». Биология развития . 278 (2): 274–88. doi : 10.1016/j.ydbio.2004.11.028 . PMID 15680349 .
- ^ Caspersson T, Schultz J (1939). «Пентозные нуклеотиды в цитоплазме растущих тканей». Природа . 143 (3623): 602–03. Bibcode : 1939natur.143..602c . doi : 10.1038/143602c0 . S2CID 4140563 .
- ^ Ochoa S (1959). «Ферментативный синтез рибонуклеиновой кислоты» (PDF) . Нобелевская лекция .
- ^ Рич А, Дэвис Д. (1956). «Новая двухцепочечная спиральная структура: полиадениловая кислота и полиуридиловая кислота». Журнал Американского химического общества . 78 (14): 3548–49. doi : 10.1021/ja01595a086 .
- ^ Holley RW, et al. (Март 1965 г.). «Структура рибонуклеиновой кислоты». Наука . 147 (3664): 1462–65. Bibcode : 1965sci ... 147.1462H . doi : 10.1126/science.147.3664.1462 . PMID 14263761 . S2CID 40989800 .
- ^ Fiers W, et al. (Апрель 1976 г.). «Полная нуклеотидная последовательность бактериофага РНК MS2: первичная и вторичная структура гена репликазы». Природа . 260 (5551): 500–07. Bibcode : 1976natur.260..500f . doi : 10.1038/260500A0 . PMID 1264203 . S2CID 4289674 .
- ^ Наполи С., Лемье С., Йоргенсен Р. (апрель 1990 г.). «Введение гена химерного синтазы халкона в петунии приводит к обратимой совместном положении гомологичных генов в транс» . Растительная ячейка . 2 (4): 279–89. doi : 10.1105/tpc.2.4.279 . PMC 159885 . PMID 12354959 .
- ^ Дафни-Йелин М., Чунг С.М., Франкман Эль, Цфира Т (декабрь 2007 г.). «Псат-РНК-интерференционные векторы: модульная серия для нескольких понижающих генов у растений» . Физиология растений . 145 (4): 1272–81. doi : 10.1104/pp.107.106062 . PMC 2151715 . PMID 17766396 .
- ^ Ruvkun G (октябрь 2001 г.). «Молекулярная биология. Проблемы крошечного мира РНК». Наука . 294 (5543): 797–99. doi : 10.1126/science.1066315 . PMID 11679654 . S2CID 83506718 .
- ^ Fichou Y, Férec C (декабрь 2006 г.). «Потенциал олигонуклеотидов для терапевтических применений». Тенденции в биотехнологии . 24 (12): 563–70. doi : 10.1016/j.tibtech.2006.10.003 . PMID 17045686 .
- ^ «Нобелевская премия по физиологии или медицине 2023» . Nobelprize.org . Получено 2023-10-03 .
- ^ «Венгерские и американские ученые выигрывают Нобель за открытия вакцины Covid-19» . Рейтер . 2023-10-02 . Получено 2023-10-03 .
- ^ «Нобелевская премия по физиологии или медицине 2023» . Nobelprize.org . Получено 2023-10-03 .
- ^ Зиберт С. (2006). «Общие свойства структуры последовательности и стабильные области во вторичных структурах РНК» (PDF) . Диссертация, Альберт-Людвигс-Универсат, Фрейбург Им Брейсгау . п. 1. Архивировано из оригинала (PDF) 9 марта 2012 года.
- ^ Szathmáry E (июнь 1999 г.). «Происхождение генетического кода: аминокислоты как кофакторы в мире РНК». Тенденции в генетике . 15 (6): 223–29. doi : 10.1016/s0168-9525 (99) 01730-8 . PMID 10354582 .
- ^ Джером, Крейг А.; и др. (19 мая 2022 г.). «Каталитический синтез полирибонуклеиновой кислоты на пребиотических горных очках» . Астробиология . 22 (6): 629–636. Bibcode : 2022sbio..22..629j . doi : 10.1089/ast.2022.0027 . PMC 9233534 . PMID 35588195 . S2CID 248917871 .
- ^ Основа для прикладной молекулярной эволюции (3 июня 2022 года). «Ученые объявляют о прорыве, определяя происхождение жизни на земле - и, возможно, Марс» . Phys.org . Получено 3 июня 2022 года .
- ^ Marlaire R (3 марта 2015 г.). «НАСА Эймс воспроизводит строительные блоки жизни в лаборатории» . НАСА . Архивировано с оригинала 5 марта 2015 года . Получено 5 марта 2015 года .
- ^ Старр, Мишель (8 июля 2022 года). «Нагрузки предшественников для РНК были обнаружены в центре нашей галактики» . Sciencealert . Получено 9 июля 2022 года .
- ^ Ривила, Виктор М.; и др. (8 июля 2022 г.). «Молекулярные предшественники РНК-мира в пространстве: новые нитрилы в молекулярном облаке G+0,693–0,027» . Границы в астрономии и космических науках . 9 : 876870. Arxiv : 2206.01053 . Bibcode : 2022frass ... 9.6870r . doi : 10.3389/fspas.2022.876870 .
- ^ Чех, Томас Р.; Стейц, Джоан А. (март 2014 г.). «Некодирующая РНК -революция - отталкивающая старые правила, чтобы создать новые» . Клетка . 157 (1): 77–94. doi : 10.1016/j.cell.2014.03.008 . ISSN 0092-8674 . PMID 24679528 . S2CID 14852160 .
- ^ Палачино, Джеймс; Свалли, Сюзанна Е; Песня, Ченг; Cheung, Atwood K; Шу, Лей; Чжан, Сяолу; Ван Гусеар, Мейлин; Шин, Юнах; Подбородок, Донован Н; Келлер, Кэролайн Губсер; Бейбел, Мартин; Рено, Николь А; Смит, Томас М; Салсиус, Майкл; Shi, Xiaoying (2015-06-01). «Модуляторы сплайсинга SMN2 усиливают ассоциацию U1-PRE-MRNA и спасающие мыши SMA». Природная химическая биология . 11 (7): 511–517. doi : 10.1038/nchembio.1837 . ISSN 1552-4450 . PMID 26030728 .
- ^ Рой, Биджойта; Фризен, Уэстли Дж.; Томизава, Юки; Лесжик, Джон Д.; Чжуо, Джин; Джонсон, Бриана; Дакка, Джумана; Тротта, Кристофер Р.; Сюэ, Сяодзиао; Mutyam, Venkatesshwar; Килинг, Ким М.; Мобли, Джеймс А.; Роу, Стивен М.; Бедвелл, Дэвид М.; Уэлч, Эллен М. (2016-10-04). «Аталурен стимулирует рибосомальный отбор почтикознанных тРНК для способности подать бессмысленную подавление» . Труды Национальной академии наук . 113 (44): 12508–12513. BIBCODE : 2016PNAS..11312508R . doi : 10.1073/pnas.1605336113 . ISSN 0027-8424 . PMC 5098639 . PMID 27702906 .
- ^ Jump up to: а беременный Кадир, Мухаммед Имран; Бухат, Шериен; Расул, Сумайра; Манзур, Хамид; Манзур, Маджид (2019-09-03). «РНК -терапия: выявление новых целей, ведущих к открытию лекарств». Журнал сотовой биохимии . 121 (2): 898–929. doi : 10.1002/jcb.29364 . ISSN 0730-2312 . PMID 31478252 . S2CID 201806158 .
- ^ Balmayor, Elizabeth R.; Гейгер, Йоханнес П.; Aneja, Manish K.; Бережански, Тарас; Утцингер, Максимилиан; Михайлик, Ольга; Рудольф, Карстен; Планка, Кристиан (май 2016 г.). «Химически модифицированная РНК индуцирует остеогенез стволовых клеток и эксплантов тканей человека, а также ускоряет заживление кости у крыс». Биоматериалы . 87 : 131–146. doi : 10.1016/j.biomaterial.2016.02.018 . ISSN 0142-9612 . PMID 26923361 .
- ^ Plews, Jordan R.; Ли, Цзяньлян; Джонс, Марк; Мур, Гарри Д.; Мейсон, Крис; Эндрюс, Питер В.; NA, Jie (2010-12-30). «Активация генов плюрипотентности в клетках фибробластов человека с помощью нового подхода на основе мРНК» . Plos один . 5 (12): E14397. BIBCODE : 2010PLOSO ... 514397P . doi : 10.1371/journal.pone.0014397 . ISSN 1932-6203 . PMC 3012685 . PMID 21209933 .
- ^ Прески, Дэвид; Эллисон, Томас Ф.; Джонс, Марк; Мамчауи, Камель; Унгер, Кристиан (май 2016 г.). «Синтетически модифицированная мРНК для эффективной и быстрой генерации клеток IPS человека и прямой трансдифференцировки к миобластам». Биохимическая и биофизическая исследовательская коммуникация . 473 (3): 743–751. doi : 10.1016/j.bbrc.2015.09.102 . ISSN 0006-291X . PMID 26449459 .
- ^ Уоррен, Луиджи; Манос, Филипп Д.; Ахфельдт, Тим; Ло, Юин-Хан; Ли, Ху; Лау, Фрэнк; Эбина, Ватару; Мандал, Панкадж К.; Смит, Захари Д.; Мейснер, Александр; Дейли, Джордж Q.; Брэк, Эндрю С.; Коллинз, Джеймс Дж.; Коуэн, Чад; Schlaeger, Thorsten M. (ноябрь 2010 г.). «Высокоэффективное перепрограммирование в плюрипотентность и направленная дифференцировка клеток человека с синтетической модифицированной мРНК» . Клеточная стволовая клетка . 7 (5): 618–630. doi : 10.1016/j.stem.2010.08.012 . ISSN 1934-5909 . PMC 3656821 . PMID 20888316 .
- ^ Elangovan, Satheesh; Хорсанд, Бехнуш; Делать, ANH-VU; Хонг, Лю; Дьюерт, Александр; Корманн, Майкл; Росс, Райан Д.; Рик Самнер, Д.; Алламарго, Шанталь; Салем, Aliasger K. (ноябрь 2015). «Химически модифицированные РНК -активированные матрицы усиливают регенерацию кости» . Журнал контролируемого выпуска . 218 : 22–28. doi : 10.1016/j.jconrel.2015.09.050 . ISSN 0168-3659 . PMC 4631704 . PMID 26415855 .
Внешние ссылки
[ редактировать ]- Сбор ссылок на веб -сайт RNA World (структуры, последовательности, инструменты, журналы)
- Изображения базы данных нуклеиновой кислоты ДНК, РНК и комплексов.
- Семинар Анны Мари Пайл: РНК -структура, функция и распознавание
Типы нуклеиновых кислот |
|---|