РНК -интерференция
РНК-интерференция ( RNAi )-это биологический процесс, в котором молекулы РНК участвуют в специфической последовательности подавления экспрессии генов с помощью двухцепочечной РНК посредством трансляционной или транскрипционной репрессии. Исторически, RNAi был известен под другими именами, включая со-супрессию , посттранскрипционное молчание генов (PTGS) и подавление . В детальном исследовании каждого из этих, казалось бы, разных процессов выяснилось, что идентичность этих явлений была на самом деле РНКи. Эндрю Фир и Крейг С. Мелло разделил Нобелевскую премию в области физиологии или медицины 2006 года за свою работу над РНКи в нематодном черве Каенорхабдит Элеганс , которую они опубликовали в 1998 году. С момента открытия РНКи и его регуляторных потенциалов стало очевидно, что РНКи. имеет огромный потенциал в подавлении желаемых генов. RNAi в настоящее время известен как точная, эффективная, стабильная и лучше, чем антисмысловая терапия для подавления генов. [ 1 ] Антисмысловая РНК, продуцируемая внутриклеточно вектором экспрессии, может быть разработана и обнаруживает полезность в качестве новых терапевтических агентов. [ 2 ]
Два типа молекул мелкой рибонуклеиновой кислоты (РНК), микроРНК (miRNA) и небольшую интерферирующую РНК ( siRNA ), являются центральными для компонентов для пути RNAi. После того, как мРНК разлагается, посттранскрипционное молчание происходит по мере предотвращения трансляции белка. Транскрипция может быть ингибирована с помощью предварительного транскрипционного механизма молчания РНКи, посредством которого ферментный комплекс катализирует метилирование ДНК в геномных положениях, комплементарных к комплексной миРНК или miRNA. RNAi играет важную роль в защите клеток от паразитных нуклеотидных последовательностей (например, вирусов или транспозонов ), а также влияет на развитие организмов.
Путь РНКи является естественным процессом, встречающимся во многих эукариотах и клетках животных. Он инициируется ферментом Dicer , который расщепляет длинные двухцепочечные РНК (DSRNA) молекулы в короткие двухцепочечные фрагменты приблизительно от 21 до 23 нуклеотидных миРНК. Каждая миРНК разматывается на два одноцепочечных РНК (SSRNAS), пассажирский (смысл) и направляющая (антисмысловая) прядь. Затем пассажирская прядь расщепляется белком Argonaute 2 (AGO2). Пассажирская цепь разлагается, и направляющая цепь включена в РНК-индуцированный комплекс молчания (RISC). Затем сборка RISC связывает и разрушает мРНК -мишени. В частности, это достигается, когда направляющие пряди пары с комплементарной последовательности в молекуле мРНК и вызывает расщепление AGO2, каталитическим компонентом RISC. В некоторых организмах этот процесс распространяется системно, несмотря на первоначально ограниченные молярные концентрации миРНК. [ 3 ]
RNAi является ценным инструментом исследования, как в клеточной культуре , так и в живых организмах , потому что синтетическая дцРНК, введенная в клетки, может избирательно и надежно вызывать подавление специфических генов, представляющих интерес. RNAi может использоваться для крупномасштабных экранов, которые систематически закрывают каждый ген (и последующие белки, которые он кодирует) в клетке, что может помочь идентифицировать компоненты, необходимые для конкретного клеточного процесса или события, такого как деление клеток . Путь также используется в качестве практического инструмента для еды, медицины и инсектицидов . [ 4 ]
Клеточный механизм
[ редактировать ]
РНКи является РНК-зависимым процессом молчания генов , который контролируется RISC и инициируется короткими двухцепочечными молекулами РНК в цитоплазме клетки, где они взаимодействуют с компонентом каталитического RISC Argonaute . [ 6 ] Когда дцРНК является экзогенной (исходя из инфекции вирусом с геномом РНК или лабораторными манипуляциями), РНК импортируется непосредственно в цитоплазму и расщепляется в короткие фрагменты Dicer. Инициирующая дцРНК также может быть эндогенной (происходящей в клетке), как в пре-микрона, экспрессируемых из генов кодирования РНК в геноме. Первичные транскрипты из таких генов сначала обрабатываются для формирования характерной структуры стволовой петли пре-мирны в ядре , а затем экспортируются в цитоплазму. Таким образом, два пути дцРНК, экзогенные и эндогенные, сходятся в RISC. [ 7 ]
Экзогенная дцРНК инициирует РНКи, активируя рибонуклеазовый белок Dicer, [ 8 ] который связывает и расщепляет дсрНК в растениях или РНК с короткими шпильками (SHRNAS) у людей, чтобы получить двухцепочечные фрагменты из 20–25 пар оснований с 2-нуклеотидным свесом на 3'-конце. [ 9 ] Исследования биоинформатики по геномам множества организмов показывают, что эта длина максимизирует специфичность целевого гена и сводит к минимуму неспецифические эффекты. [ 10 ] Эти короткие двойные фрагменты называются миРНК . Эти миРНК затем разделяются на отдельные цепи и интегрируются в активное RISC, комплексом загрузки RISC (RLC). RLC включает Dicer-2 и R2D2, и имеет решающее значение для объединения AGO2 и RISC. [ 11 ] TATA-связывающий белок-ассоциированный фактор 11 (TAF11) собирает RLC, облегчая тетрамеризацию DCR-2-R2D2, что увеличивает аффинность связывания с миРНК в 10 раз. Ассоциация с TAF11 будет преобразовать комплекс R2-D2-инициатора (RDI) в RLC. [ 12 ] R2D2 несет тандемные двународные РНК-связывающие домены, чтобы распознавать термодинамически стабильный концерн дуплексов миРНК , тогда как Dicer-2-другая менее стабильная конечность. Нагрузка асимметрична: средний домен AGO2 распознает термодинамически стабильный конец siRNA. Следовательно, «пассажир» (смысл) Strand, чья 5 'конец отбрасывается до середины, выброшен, в то время как сохраненный «путеводитель» (антисмысловая) Strand сотрудничает, чтобы сформировать RISC. [ 11 ]
После интеграции в RISC, базовая папа миРНК к их мРНК-мишени и расщепляйте ее, тем самым предотвращая его использование в качестве шаблона перевода . [ 13 ] В отличие от SiRNA , нагруженные miRNA-нагруженными комплексом RISC сканирует цитоплазматические мРНК для потенциальной взаимодополняемости. Вместо разрушительного расщепления (по AOGO2) miRNAs скорее нацелены на 3' -нетрансляционную область (UTR) мРНК, где они обычно связываются с несовершенной комплементарностью, тем самым блокируя доступ рибосомов для трансляции. [ 14 ]
Экзогенная дцРНК обнаруживается и связана эффекторным белком, известным как RDE-4 у C. elegans и R2D2 у дрозофилы , которые стимулируют активность DICER. [ 15 ] Механизм, продуцирующий эту специфичность длины, неизвестен, и этот белок связывает только длинные дцРНК. [ 15 ]
У C. elegans этот ответ инициации усиливается посредством синтеза популяции «вторичных» миРНК, в ходе которых инициирующие или «первичные» миРНК используются в качестве шаблонов. [ 16 ] Эти «вторичные» миРНК структурно отличаются от дицеровских миРНК и, по-видимому, продуцируются РНК-зависимой РНК-полимеразой (RDRP). [ 17 ] [ 18 ]
Микрорну
[ редактировать ]
МикроРНК (miRNAs) являются геномически кодируемыми некодирующими РНК , которые помогают регулировать экспрессию генов , особенно во время развития . [ 19 ] Феномен РНКи, широко определенного, включает в себя эндогенно индуцированные эффекты молчания генов, а также молчание, вызванные инородной дсРНК. Зрелые miRNAS структурно аналогичны миРНК, продуцируемым из экзогенной дсРНК, но до достижения зрелости miRNAs должны сначала пройти обширную посттранскрипционную модификацию . MiRNA экспрессируется из гораздо более длинного РНК-кодирующего гена в качестве первичного транскрипта, известного как PRI-MIRNA , которая обрабатывается в клеточном ядре 70 нуклеотидов, , в структуру стволовой ствола называемой домирной микропроцессорным комплексом . Этот комплекс состоит из фермента РНКазы III, называемого Drosha и DSRNA-связывающего белка DGCR8 . ДсРНК-часть этой пре-мирны связана и расщепляется DICER, чтобы получить зрелую молекулу miRNA, которая может быть интегрирована в комплекс RISC; Таким образом, miRNA и siRNA имеют одинаковую нижестоящую клеточную механизм. [ 20 ] Во -первых, вирусная кодированная miRNA была описана у вируса Эпштейна -Барра (EBV). [ 21 ] После этого все большее число микроРНК было описано в вирусах. Virmirna-это комплексный каталог, покрывающий вирусную микроРНК, их мишени и антивирусные miRNAs [ 22 ] (См. Также ресурс Virmirna: http://crdd.osdd.net/servers/virmirna/).
МиРНК, полученные из длинных предшественников дсРНК, отличаются от miRNAs в том, что miRNAs, особенно у животных, обычно имеют неполное сочетание оснований к мишени и ингибируют трансляцию многих различных мРНК с похожими последовательностями. Напротив, миРНК обычно идеально подходят и вызывают расщепление мРНК только в одной конкретной цели. [ 23 ] У Drosophila и C. elegans miRNA и siRNA обрабатываются различными аргонатными белками и ферментами Dicer. [ 24 ] [ 25 ]
Три первичных нетранслируемых регионов и микроРНК
[ редактировать ]Три основных нетранслируемых областей (3'UTR) мРНК часто содержат регуляторные последовательности, которые посттранскрипционно вызывают РНКи. Такие 3'-UTR часто содержат как сайты связывания, так и для регуляторных белков. Связываясь со специфическими сайтами в пределах 3'-UTR, miRNAs могут снижать экспрессию генов различных мРНК, либо ингибируя трансляцию, либо вызывая непосредственную деградацию транскрипта. 3'-UTR также могут иметь области глушителя, которые связывают репрессорные белки, которые ингибируют экспрессию мРНК.
3'-UTR часто содержит элементы отклика микроРНК (MRES) . MRE - это последовательности, с которыми связывают miRNAs. Это распространенные мотивы в пределах 3'-UTR. Среди всех регуляторных мотивов в пределах 3'-UTR (например, включая области глушителя) MRE составляют около половины мотивов.
По состоянию на 2023 год, веб -сайт mirbase , [ 26 ] Архив и аннотаций miRNA последовательностей , перечисленный 28 645 записей у 271 биологических видов. Из них 1917 miRNAs были в аннотированных локусах miRNA человека. Предполагалось, что miRNAs будут иметь в среднем около четырехсот мРНК мишени (влияя на экспрессию нескольких сотен генов). [ 27 ] Friedman et al. [ 27 ] Оценить, что> 45 000 сайтов мишеней miRNA в пределах 3'UTR для мРНК человека сохраняются выше фоновых уровней, а> 60% генов, кодирующих белок человека, находились под селективным давлением, чтобы поддерживать сопряжение с miRNAs.
Прямые эксперименты показывают, что одна miRNA может снизить стабильность сотен уникальных мРНК. [ 28 ] Другие эксперименты показывают, что одна miRNA может подавлять производство сотен белков, но эта репрессия часто является относительно легкой (менее чем в 2 раза). [ 29 ] [ 30 ]
Эффекты дисрегуляции miRNA экспрессии генов, по -видимому, важны при раке. [ 31 ] Например, при раке желудочно -кишечного тракта девять miRNAs были идентифицированы как эпигенетически измененные и эффективные при регулировании ферментов репарации ДНК. [ 32 ]
Эффекты дисрегуляции miRNA экспрессии генов также, по -видимому, также являются важными при нейропсихиатрических расстройствах, таких как шизофрения, биполярное расстройство, серьезная депрессия, болезнь Паркинсона, болезнь Альцгеймера и расстройства спектра аутизма. [ 33 ] [ 34 ] [ 35 ]
Активация RISC и катализ
[ редактировать ]Экзогенная дцРНК обнаруживается и связана эффекторным белком, известным как RDE-4 у C. elegans и R2D2 у дрозофилы , которые стимулируют активность DICER. [ 15 ] Этот белок связывает только длинные дцРНК, но механизм, продуцирующий эту специфичность длины, неизвестен. [ 15 ] Этот РНК-связывающий белок затем облегчает перенос расщепленных миРНК в комплекс RISC. [ 36 ]
У C. elegans этот ответ инициации усиливается посредством синтеза популяции «вторичных» миРНК, в ходе которых инициирующие или «первичные» миРНК используются в качестве шаблонов. [ 16 ] Эти «вторичные» миРНК структурно отличаются от дицеровских миРНК и, по-видимому, продуцируются РНК-зависимой РНК-полимеразой (RDRP). [ 17 ] [ 18 ]


Активными компонентами РНК-индуцированного молчанного комплекса (RISC) являются эндонуклеазы, называемые аргонатными белками, которые расщепляют целевую цепь, дополненную их связанной миРНК . [ 6 ] Поскольку фрагменты, произведенные Dicer, являются двународными, они могли бы создавать функциональную миРНК . Однако только одна из двух нитей, которая известна как направляющая нить , связывает аргонаут и направляет молчание генов. Другая противотуродейская нить или пассажирская нить разлагается во время активации RISC. [ 37 ] Хотя сначала считалось, что АТФ -зависимая геликаза отделяла эти две пряди, [ 38 ] Процесс оказался АТФ-независимым и выполняется непосредственно белковыми компонентами RISC. [ 3 ] [ 39 ] Однако кинетический анализ in vitro RNAi в присутствии и отсутствии АТФ показал, что АТФ может потребоваться для расслабления и удаления цепи мРНК из комплекса RISC после катализа. [ 40 ] Грубальная пряди имеет тенденцию быть теми, чей 5 'конец менее стабильно связан с его комплемент [ 41 ] Но на выбор пряди не влияет направление, в котором Dicer расщепляет дцРНК до включения RISC. [ 42 ] Вместо этого белок R2D2 может служить дифференцирующим фактором, связывая более стабильный 5'-конец пассажирской цепи. [ 43 ]
Структурная основа для связывания РНК с аргонатным белком была исследована с помощью рентгеновской кристаллографии связывающего домена аргонаута, связанного с РНК. Здесь фосфорилированный 5' -конец РНК цепь попадает в консервативный базовый поверхности карман и устанавливает контакты через дивалентный катион (атом с двумя положительными зарядами), такого как магний и ароматическая укладка (процесс, который позволяет более одного атома. электрон путем прохождения его вперед и назад) между 5' -нуклеотидом в миРНК и консервативным остатком тирозина . Считается, что этот сайт образует сайт зарождения для связывания миРНК с ее мРНК -мишенью. [ 44 ] Анализ ингибирующего эффекта несоответствий в 5 'или 3' конец направляющей нити продемонстрировал, что 5 'конец направляющей нити, вероятно, отвечает за сопоставление и связывание мРНК -мишени, в то время как 3' конец ответственен за Физическое расположение мРНК-мишени в область RISC с благоустройством расщепления. [ 40 ]
Не понятно, как активированный комплекс RISC обнаруживает комплементарные мРНК в клетке. Несмотря на то, что процесс расщепления был предложен для связи с трансляцией , трансляция мишени мРНК не является необходимым для RNAi-опосредованной деградации. [ 45 ] Действительно, RNAi может быть более эффективным против мРНК -мишеней, которые не переведены. [ 46 ] Аргонатные белки локализуются в специфических областях в цитоплазме, называемой P-боди (также цитоплазматические тела или тела GW), которые представляют собой области с высокой скоростью распада мРНК; [ 47 ] Активность miRNA также кластеризована в P-корзи. [ 48 ] Разрушение P-болей снижает эффективность RNAI, что позволяет предположить, что они являются критическим местом в процессе RNAI. [ 49 ]
Транскрипционное молчание
[ редактировать ]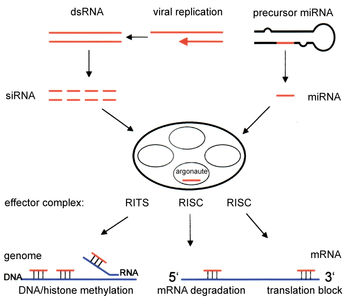
Компоненты пути RNAi используются во многих эукариотах при поддержании организации и структуры их геномов . Модификация гистонов и связанная с этим индукция образования гетерохроматина служит для подавления генов до транскрипционно ; [ 51 ] Этот процесс называется РНК-индуцированным транскрипционным молчанием (RIT) и осуществляется комплексом белков, называемых комплексом RITS. У делящихся дрожжей этот комплекс содержит Argonaute, хромодомен белок CHP1 и белок, называемый TAS3 неизвестной функции. [ 52 ] Как следствие, индукция и распространение гетерохроматических областей требуют белков аргонат и RDRP. [ 53 ] Действительно, делеция этих генов в делящихся дрожжах S. pombe нарушает метилирование гистонов и образование центромеры , [ 54 ] вызывая медленную или остановленную анафазу во время деления клеток . [ 55 ] В некоторых случаях были обнаружены аналогичные процессы, связанные с модификацией гистонов, транскрипционно активируют гены. [ 56 ]
Механизм, с помощью которого комплекс RITS индуцирует образование гетерохроматина и организацию, не совсем понятен. Большинство исследований были сосредоточены на области спариваемого типа в дрожжах делящихся, которые не могут быть репрезентативными для активности в других геномных регионах/организмах. При поддержании существующих гетерохроматиновых областей RITS образует комплекс с миРНК, комплементарными локальным генам и стабильно связывает локальные метилированные гистоны, действуя совместно транскрипционно, чтобы разрушить любые зарождающиеся транскрипты пре-мРНК, которые инициируются РНК-полимеразой . Образование такой области гетерохроматина, хотя и не ее обслуживание, зависит от DICER, предположительно, потому что Dicer необходим для создания начального комплемента миРНК , которые нацелены на последующие транскрипты. [ 57 ] Предполагается, что поддержание гетерохроматина функционирует как контур обратной связи самостоятельно, поскольку новые миРНК образуются из случайных зарождающихся транскриптов RDRP для включения в локальные комплексы RITS. [ 58 ] Актуальность наблюдений из областей делящихся дрожжевых типов и центромеров для млекопитающих неясно, поскольку поддержание гетерохроматина в клетках млекопитающих может быть независимым от компонентов пути RNAi. [ 59 ]
Перевод с редактированием РНК
[ редактировать ]Тип редактирования РНК , который наиболее распространен у более высоких эукариот, превращает аденозин -нуклеотиды в инозин в дцРНК через фермент аденозин -деаминазу (ADAR). [ 60 ] Первоначально было предложено в 2000 году, что пути редактирования РНК и → I могут конкурировать за общий субстрат дцРНК. [ 61 ] Некоторые пре-мирНК подвергаются редактированию РНК A → I [ 62 ] [ 63 ] и этот механизм может регулировать обработку и экспрессию зрелых miRNAs. [ 63 ] Кроме того, по крайней мере один адар млекопитающего может секвестировать миРНК из компонентов пути RNAI. [ 64 ] Дополнительная поддержка этой модели поступает из исследований на штаммах Adar-Null C. elegans, указывающих на то, что редактирование РНК A → I может противодействовать молчанию РНКи эндогенных генов и трансгенов. [ 65 ]

Различия среди организмов
[ редактировать ]Организмы различаются по своей способности принимать зарубежную дцРНК и использовать ее в пути РНКи. Эффекты RNAi могут быть как системными, так и наследственными у растений и C. elegans , хотя и не у дрозофилы или млекопитающих. Считается, что у растений RNAi распространяется путем переноса миРНК между клетками через Plasmodesmata (каналы в клеточных стенках, которые обеспечивают связь и транспорт). [ 38 ] Наследственность происходит от метилирования промоторов, нацеленных на RNAi; Новая картина метилирования копируется в каждом новом поколении клетки. [ 67 ] Широкое общее различие между растениями и животными заключается в нацеливании эндогенно продуцированных miRNAs; У растений miRNAs обычно идеально или почти идеально дополняют их гены -мишени и вызывают прямое расщепление мРНК с помощью RISC, в то время как микроРНК животных, как правило, более расходятся в последовательности и вызывают трансляционную репрессию. [ 66 ] Этот трансляционный эффект может быть получен путем ингибирования взаимодействия факторов инициации мРНК трансляции с помощью полиаденинового хвоста . [ 68 ]
Некоторые эукариотические простейшие, такие как Leishmania Major и Trypanosoma Cruzi, не хватает пути RNAi полностью. [ 69 ] [ 70 ] Большинство или все компоненты также отсутствуют в некоторых грибах , особенно в модельном организме Saccharomyces cerevisiae . [ 71 ] Присутствие РНКи у других почкующихся видов дрожжей, таких как Saccharomyces castellii и Candida albicans , также демонстрирует, что индуцирование двух связанных с РНК белка из S. castellii облегчает RNAi у S. cerevisiae . [ 72 ] То, что в определенных аскомицетах и базидиомицетах отсутствуют пути RNAi, указывает на то, что белки, необходимые для молчания РНК, были потеряны независимо от многих грибковых линий , возможно, из -за эволюции нового пути с аналогичной функцией или отсутствия селективного преимущества в определенных нишах . [ 73 ]
Связанные прокариотические системы
[ редактировать ]На экспрессию генов у прокариот влияет система РНК, сходная в некоторых отношениях с РНКи. Здесь гены, кодирующие РНК, контролируют содержание или трансляцию мРНК, продуцируя комплементарную РНК, которая отжигает мРНК. Однако эти регуляторные РНК, как правило, не считаются аналогичными miRNAS, потому что фермент Dicer не участвует. [ 74 ] Было высказано предположение, что системы интерференции CRISPR в прокариотах аналогичны эукариотическим системам РНКи, хотя ни один из компонентов белка не является ортологичным . [ 75 ]
Биологические функции
[ редактировать ]Этот раздел должен быть обновлен . ( Май 2020 г. ) |
Иммунитет
[ редактировать ]RNAi является жизненно важной частью иммунного ответа на вирусы и другого иностранного генетического материала , особенно в растениях, где он также может предотвратить самопроводство транспозонов. [ 76 ] Растения, такие как Arabidopsis thaliana, экспрессируют несколько гомологов Dicer , которые специализируются на по -разному, когда растение подвергается воздействию различных вирусов. [ 77 ] Еще до того, как путь RNAi был полностью понят, было известно, что индуцированное молчание генов у растений может распространяться по всему растению при системном эффекте и может быть перенесено из запаса в Scion растения через прививку . [ 78 ] Это явление с тех пор было признано особенностью иммунной системы растения, которая позволяет всем растению реагировать на вирус после первоначальной локализованной встречи. [ 79 ] В ответ многие вирусы растений эволюционировали сложные механизмы для подавления ответа РНКи. [ 80 ] К ним относятся вирусные белки, которые связывают короткие двухцепочечные фрагменты РНК с одноцепочечными концами выступления, например, продуцируемые Dicer. [ 81 ] Некоторые геномы растений также экспрессируют эндогенные миРНК в ответ на инфекцию специфическими типами бактерий . [ 82 ] Эти эффекты могут быть частью генерализованного ответа на патогены, которые подавляют любой метаболический процесс у хозяина, который помогает процессу инфекции. [ 83 ]
Хотя животные обычно экспрессируют меньше вариантов фермента Dicer, чем растения, RNAi у некоторых животных вызывает противовирусную реакцию. Как у ювенильной, так и у взрослой дрозофилы RNAi важен при противовирусном врожденном иммунитете и активен против патогенных микроорганизмов, таких как вирус Drosophila x . [ 84 ] [ 85 ] Аналогичная роль в иммунитете может работать у C. elegans , поскольку аргонатные белки активируются в ответ на вирусы и черви, которые сверхэкспрессируют компоненты пути RNAi, устойчивы к вирусной инфекции. [ 86 ] [ 87 ]
Роль RNAi в врожденном иммунитете млекопитающих плохо изучена, и доступно относительно мало данных. Однако существование вирусов, которые кодируют гены, способные подавлять ответ РНКи в клетках млекопитающих, может быть доказательством в пользу РНКи-зависимого иммунного ответа млекопитающих, [ 88 ] [ 89 ] Хотя эта гипотеза была оспорена как плохо обоснованная. [ 90 ] Были представлены данные о существовании функционального противовирусного пути РНКи в клетках млекопитающих. [ 91 ] [ 92 ]
Также существуют другие функции для RNAi у вирусов млекопитающих, таких как miRNAs, экспрессируемые вирусом герпеса , которые могут действовать как гетерохроматина для опосредования латентности вируса. триггеры [ 93 ]
Подавление генов
[ редактировать ]Эндогенно экспрессируемые miRNAs, включая как интронные , так и межгенные miRNAs, наиболее важны в трансляционных репрессиях [ 66 ] и в регуляции развития, особенно в сроках морфогенеза и поддержании недифференцированных или не полностью дифференцированных типов клеток, таких как стволовые клетки . [ 94 ] Роль эндогенно экспрессируемой miRNA в понижении экспрессии генов была впервые описана у C. elegans в 1993 году. [ 95 ] У растений эта функция была обнаружена, когда было показано, что «челюстная микроРНа» арабидопсиса участвует в регуляции нескольких генов, которые контролируют форму растения. [ 96 ] У растений большинство генов, регулируемых miRNAS, являются факторами транскрипции ; [ 97 ] Таким образом, активность miRNA особенно широко распространена и регулирует целые генные сети во время развития, модулируя экспрессию ключевых регуляторных генов, включая факторы транскрипции, а также белки F-бокса . [ 98 ] Во многих организмах, включая людей, miRNAs связаны с образованием опухолей и дисрегуляцией клеточного цикла . Здесь miRNAs могут функционировать как онкогены , так и опухолевые супрессоры . [ 99 ]
Эволюция
[ редактировать ]Основываясь на на основе скупости филогенетическом анализе , самый последний общий предок всех эукариот, скорее всего, уже имел ранний путь RNAi; Считается, что отсутствие пути у некоторых эукариот является производной. [ 100 ] Эта наследственная система РНКи, вероятно, содержала по крайней мере один дицероподобный белок, один аргонат, один белок пиви и РНК-зависимую РНК-полимеразу , которая также может сыграть другие клеточные роли. Крупномасштабное исследование сравнительной геномики также указывает на то, что эукариотическая группа коронов уже обладала этими компонентами, что затем может иметь более тесные функциональные ассоциации с обобщенными системами деградации РНК, такими как экзосома . [ 101 ] Это исследование также предполагает, что РНК-связывающее семейство аргонатных белков, которое разделяется среди эукариот, большинство археи, и, по крайней мере, некоторые бактерии (такие как Aquifex Aeolicus ), гомологичен и первоначально развивается из компонентов системы инициации трансляции .
Приложения
[ редактировать ]Путь RNAi для нокдауна гена
[ редактировать ]Нокдаун гена - это метод, используемый для снижения экспрессии специфических генов организма. Это достигается с использованием естественного процесса RNAI. [ 6 ] В этом методе нокдауна гена используется двухцепочечная молекула миРНК, которая синтезируется с последовательности, дополняющим интересующему гену. Каскад РНКи начинается, как только фермент Dicer начинает обрабатывать siRNA. Конечный результат процесса приводит к деградации мРНК и разрушает любые инструкции, необходимые для создания определенных белков. Используя этот метод, исследователи способны уменьшить (но не полностью устранить) экспрессию целевого гена. Изучение эффектов этого снижения экспрессии может показать физиологическую роль или влияние целевых генных продуктов. [ 102 ] [ 103 ]
Нецелевые эффекты нокдауна гена
[ редактировать ]Обширные усилия в области вычислительной биологии были направлены на разработку успешных реагентов дцРНК, которые максимизируют нокдаун генов, но минимизируют эффекты «нецелевого». Эффекты вне цели возникают, когда введенная РНК имеет базовую последовательность, которая может сочетаться с и, таким образом, снижать экспрессию нескольких генов. Такие проблемы возникают чаще, когда дцРНК содержит повторяющиеся последовательности. оценивалось По изучению геномов людей, C. elegans и S. Pombe , что около 10% возможных миРНК оказывают существенные эффекты, не связанные с целью. [ 10 ] Было разработано множество программных инструментов внедрение алгоритмов для проектирования общего [ 104 ] [ 105 ] специфичный для млекопитающих, [ 106 ] и вирус-специфический [ 107 ] миРНК , которые автоматически проверяются на возможную перекрестную реактивность.
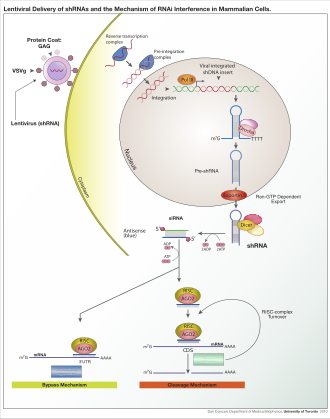
В зависимости от организма и экспериментальной системы, экзогенная РНК может быть длинной цепью, предназначенной для расщепления Dicer или коротких РНК, предназначенных для служащих в качестве миРНК субстратов . В большинстве клеток млекопитающих используются более короткие РНК, потому что длинные двухцепочечные молекулы РНК индуцируют реакцию интерферона млекопитающих , форму врожденного иммунитета , которая неспецифично реагирует на иностранный генетический материал. [ 108 ] Мышиные ооциты и клетки из ранних эмбрионов мыши не имеют этой реакции на экзогенную дцРНК и, следовательно, являются общей модельной системой для изучения эффектов снока генов млекопитающих. [ 109 ] Специализированные лабораторные методы также были разработаны для улучшения полезности RNAI в системах млекопитающих, избегая прямого введения siRNA , например, путем стабильной трансфекции плазмидой, , стабильная плазмида кодирующей соответствующую последовательность, из которой могут быть транскрибированы миРНК , из которой могут быть транскрибированы миРНК . [ 110 ] или более сложными лентивирусными векторными системами, позволяющими индуцибельной активации или дезактивации транскрипции, известной как условная RNAI . [ 111 ] [ 112 ]
Лекарства
[ редактировать ]

Техника сбивания генов с использованием RNAi Therapeutics продемонстрировала успех в рандомизированных контролируемых клинических исследованиях. Эти лекарства представляют собой растущий класс лекарств на основе siRNA, которые снижают экспрессию белков, кодируемых определенными генами. На сегодняшний день регулирующие органы в США и Европе одобрены пять лекарств РНКи: Патисиран (2018), Givosiran (2019), Лумасиран (2020), Накирран (2020 в Европе с ожидаемым одобрением США в 2021 году) и Vutrisiran (2022 ) [ 113 ] [ 114 ] [ 115 ] [ 116 ]
В то время как все нынешние регуляторные органы, одобренные RNAI Therapeutics, фокусируются на заболеваниях, которые возникают в печени, дополнительные лекарства, исследуемые, нацелены Полем
Patisiran -это первое лекарство на основе двойного мельницы, одобренное в 2018 году и разработанное Alnylam Pharmaceuticals . Patisiran использует каскад RNAI, чтобы подавить ген, который кодирует TTR (трансритин). Мутации в этом гене могут вызвать неправильное сворачивание белка, ответственного за наследственный амилоидоз ATTR . Для достижения терапевтического ответа патисиран заключается в мембране липидной наночастицы , которая облегчает кроссовер в цитоплазму. Оказавшись внутри клетки, миРНК начинает обрабатывать ферментом дицером. Patisiran управляется медицинским работником через внутривенную инфузию с дозированием в зависимости от массы тела. Предупреждения и меры предосторожности включают риск реакций, связанных с инфузией, и снижение уровней витамина А (сыворотка). [ 117 ]
В 2019 году FDA и EMA одобрили Givosiran для лечения взрослых с острой печеночной порфирией (AHP). [ 118 ] FDA также предоставило Givosiran обозначение прорывной терапии , обозначение приоритетного обзора и обозначение лекарств для сирот для лечения острого печени Порфирии (AHP) в ноябре 2019 года. [ 119 ] К 2020 году Гивосиран получил одобрение EMA. [ 120 ] Givosiran - это миРНК, которая разрушает мРНК аминолевулиновой кислоты синтазы 1 (ALAS1) в печени. Разрушение мРНК ALAS1 предотвращает токсины (ответственные за нейровискорильные атаки и болезнь AHP), такие как аминолевулиновая кислота (ALA) и порфобилиноген (PBG). [ 121 ] [ 122 ] [ 123 ] [ 124 ] Чтобы облегчить проникновение в цитоплазму, Givosiran использует лиганды Galnac и входит в клетки печени. Препарат вводится подкожно медицинским работником с дозированием в зависимости от массы тела. Предупреждения и меры предосторожности включают риск анафилактических реакций, печеночной токсичности, почечной токсичности и реакций места инъекции. [ 125 ]
Lumasiran был одобрен в качестве лекарства на основе SIRNA в 2020 году для использования как в Европейском союзе, так и в Соединенных Штатах. [ 126 ] [ 127 ] Это лекарство используется для лечения первичной гипероксалюрии типа 1 (PH1) в педиатрической и взрослой популяции. Препарат предназначен для снижения выработки оксалата печени и уровня оксалата мочи посредством RNAi путем нацеливания мРНК гидроксикидаксидазы 1 (HAO1) для разрушения. Понижение уровней фермента HAO1 снижает окисление гликолата до глиоксилата (который является субстратом для оксалата). Lumasiran вводится подкожно медицинским работником с дозированием в зависимости от массы тела. [ 128 ] Данные рандомизированных контролируемых клинических испытаний показывают, что наиболее распространенной побочной реакцией, которая была сообщена, были реакции участка инъекции. Эти реакции были мягкими и присутствовали у 38 процентов пациентов, получавших люмасиран. [ 129 ]
В 2022 году FDA и EMA одобрили Vutrisiran для лечения взрослых с наследственным транстиретином, опосредованным амилоидозом, с стадии 1 или 2 полинейропатии. [ 130 ] [ 131 ] Vutrisiran предназначен для разрушения мРНК, которая кодирует транстиретин .
Другие исследовательские препараты, использующие RNAI, которые разрабатываются фармацевтическими компаниями, такими как фармацевтические препараты Arrowhead , Dicerna, Alnylam Pharmaceuticals , Amgen и Sylentis. Эти лекарства охватывают различные мишени с помощью RNAI и болезней.
Исследовательская терапия РНКи в разработке:
| Лекарство | Цель | Система доставки | Болезнь | Фаза | Статус | Компания | Идентификатор |
|---|---|---|---|---|---|---|---|
| Aln - vsp02 | KSP и VEGF | LNP | Солидные опухоли | я | Завершенный | Alnylam Pharmaceuticals | NCT01158079 |
| siRNA - EPHA2 - DOPC | Epha2 | LNP | Продвинутый рак | я | Рекрутинг | М.Д. Андерсонский онкологический центр | NCT01591356 |
| ATU027 | PKN3 | LNP | Солидные опухоли | я | Завершенный | Молчание терапии | NCT00938574 |
| TKM -080301 | PLK1 | LNP | Рак | я | Рекрутинг | Tekmira Pharmaceutical | NCT01262235 |
| TKM -100201 | VP24, VP35, Zaire Ebola L-полимераза | LNP | Эбола-вирусная инфекция | я | Рекрутинг | Tekmira Pharmaceutical | NCT01518881 |
| Aln - RSV01 | RSV NucleocapsId | Голая миРНК | Респираторные синцитиальные вирусные инфекции | II | Завершенный | Alnylam Pharmaceuticals | NCT00658086 |
| Pro-040201 | Апоб | LNP | Гиперхолестеринемия | я | Прекращен | Tekmira Pharmaceutical | NCT00927459 |
| Aln - PCS02 | PCSK9 | LNP | Гиперхолестеринемия | я | Завершенный | Alnylam Pharmaceuticals | NCT01437059 |
| Aln - ttr02 | Тттров | LNP | Транстиретин-опосредованный амилоидоз | II | Рекрутинг | Alnylam Pharmaceuticals | NCT01617967 |
| Кусок 01 | RRM2 | Циклодекстрин NP | Солидные опухоли | я | Активный | Низкие фармацевтические препараты | NCT00689065 |
| TD101 | K6A (мутация N171K) | Голая миРНК | Врожденная пахионихия | я | Завершенный | Pachyonychia врожденный проект | NCT00716014 |
| Agn211745 | VEGFR1 | Голая миРНК | Возрастная макулярная дегенерация, хориоидальная неоваскуляризация | II | Прекращен | Аллерган | NCT00395057 |
| QPI-1007 | CASP2 | Голая миРНК | Зрительная атрофия, невозможная передняя ишемическая оптическая невропатия | я | Завершенный | Quark Pharmaceuticals | NCT01064505 |
| I5np | P53 | Голая миРНК | Повреждение почек, острая почечная недостаточность | я | Завершенный | Quark Pharmaceuticals | NCT00554359 |
| Задержка функции трансплантата, осложнения пересадки почки | Я, ii | Рекрутинг | Quark Pharmaceuticals | NCT00802347 | |||
| PF-655 (PF-04523655) | RTP801 (собственная цель) | Голая миРНК | Хориоидальная неоваскуляризация, диабетическая ретинопатия, диабетический отек желтого пятна | II | Активный | Quark Pharmaceuticals | NCT01445899 |
| SIG12D LODER | Крас | Много полимер | Рак поджелудочной железы | II | Рекрутинг | Силузед | NCT01676259 |
| Яростный | Овощ | Голая миРНК | Диабетический макулярный отек, макулярная дегенерация | II | Завершенный | Опокико здоровье | NCT00306904 |
| SYL1001 | TRPV1 | Голая миРНК | Глазовая боль, синдром сухого глаза | Я, ii | Рекрутинг | Топоры | NCT01776658 |
| SYL040012 | ADRB2 | Голая миРНК | Глазовая гипертония, открытая глаукома | II | Рекрутинг | Топоры | NCT01739244 |
| CEQ508 | Ctnnb1 | Escherichia coli-Carrying Shrna | Семейный аденоматозный полипоз | Я, ii | Рекрутинг | Марина биотехнология | Неизвестный |
| RXI-109 | Ctgf | Самоавторанное соединение RNAi | Профилактика шрамов Cicatrix | я | Рекрутинг | RXI Pharmaceuticals | NCT01780077 |
| Aln - ttrsc | Тттров | МиРНК -галнак конъюгат | Транстиретин-опосредованный амилоидоз | я | Рекрутинг | Alnylam Pharmaceuticals | NCT01814839 |
| ARC-520 | Консервативные районы HBV | DPC | HBV | я | Рекрутинг | Исследование стрел | NCT01872065 |
Правовая категоризация и юридические вопросы в ближайшем будущем
[ редактировать ]В настоящее время как miRNA, так и миРНК в настоящее время химически синтезируются и поэтому юридически классифицируются внутри ЕС и в США как «простые» лекарственные продукты. Но поскольку биоинженерная миРНК (BERAS) находится в разработке, они будут классифицированы как биологические лекарственные продукты, по крайней мере, в ЕС. Разработка технологии BERAS поднимает вопрос о категоризации лекарств, имеющих тот же механизм действия, но производится химически или биологически. Это отсутствие последовательности должно быть решено. [ 132 ]
Механизмы доставки
[ редактировать ]Для достижения клинического потенциала РНКи миРНК должна эффективно транспортироваться в клетки тканей -мишеней. Тем не менее, существуют различные барьеры, которые необходимо исправить, прежде чем их можно будет использовать клинически. Например, «голая» миРНК подвержена нескольким препятствиям, которые снижают ее терапевтическую эффективность. [ 133 ] Кроме того, после того, как siRNA вошла в кровоток, обнаженная РНК может быть разлагалась на нуклеазах сыворотки и может стимулировать врожденную иммунную систему. [ 133 ] Из -за своего размера и высокополианионной (содержащей негативные заряды в нескольких местах) природа, немодифицированные молекулы siRNA не могут легко попасть в клетки через клеточную мембрану. искусственную или наночастицу инкапсулированную миРНК Следовательно, необходимо использовать . Если миРНК переносится по всей клеточной мембране, непреднамеренная токсичность может возникнуть, если терапевтические дозы не оптимизированы, а миРНК могут оказывать нецелевое воздействие (например, непреднамеренное подавление генов с комплементатарной комплементарностью частичной последовательности ). [ 134 ] Даже после входа в ячейки требуется повторное дозирование, поскольку их эффекты разбавляются в каждом делении клеток. В ответ на эти потенциальные проблемы и барьеры два подхода помогают облегчить доставку миРНК к клеткам -мишеням: наночастицы липидов и конъюгаты. [ 135 ]
Липидные наночастицы
[ редактировать ]Липидные наночастицы (LNP) основаны на липосомоподобных структурах, которые обычно изготовлены из водного центра, окруженного липидной оболочкой. [ 136 ] Подмножество липосомальных структур, используемых для доставки лекарств в ткани, отдыхают в больших одноламеллярных везикулах (LUV), которые могут быть размером 100 нм. Механизмы доставки LNP стали растущим источником интуитивных нуклеиновых кислот и могут включать плазмиды , CRISPR и мРНК . [ 137 ]
Первое утвержденное использование липидных наночастиц в качестве механизма доставки лекарств началось в 2018 году с препаратом для препарата для миРНК, разработанного Alnylam Pharmaceuticals. Dicerna Pharmaceuticals, Persomics , Sanofi и SiRNA Therapeutics также работали над тем, чтобы вывести терапию RNAI на рынок. [ 138 ] [ 139 ]
Другие недавние заявки включают две одобренные FDA вакцины COVID-19: мРНК-1273, разработанные Moderna . и BNT162B , разработанный в результате сотрудничества между Pfizer и Biontech . [ 140 ] Эти две вакцины используют липидные наночастицы для доставки антигена мРНК. Инкапсуляция молекулы мРНК в наночастицах липидов была критическим прорывом для продуцирования жизнеспособных вакцинов мРНК, решающих ряд ключевых технических барьеров при доставке молекулы мРНК в клетку-хозяина, распределенные по аполипопротеину E (APOE) в рецепторе липопротеина низкой плотности (LDLR ) В декабре 2020 года Novartis объявил, что положительные результаты исследований эффективности фазы III считают, что наклонность является лечением гетерозиготной семейной гиперхолестеринемии (HEFH) и атеросклеротических сердечно -сосудистых заболеваний (ASCVD). [ 141 ]
Конъюгаты
[ редактировать ]В дополнение к LNP, терапевтические средства RNAI имеют нацеленную на доставку через конъюгаты siRNA (например, Galnac, углеводы, пептиды, аптамеры, антитела). [ 142 ] Терапии с использованием конъюгатов миРНК были разработаны для редких или генетических заболеваний, таких как острые печеночные порфирии (AHP), гемофилия , первичная гипероксалурия (PH) и наследственная амилоидоз ATTR , а также другие кардиометаболические заболевания, такие как гипертония и несущественный стеатогепатит (NASH). [ 143 ]
Биотехнология
[ редактировать ]RNAi использовался для множества других применений, включая продукты питания, сельскохозяйственные культуры и инсектициды. Использование пути RNAi разработало множество продуктов, таких как продукты, такие как арктические яблоки , без никотина табак, кофе без кофеина, растительность питательных веществ и гипоаллергенные культуры. [ 144 ] [ 145 ] [ 146 ] Новое использование RNAi может разработать множество других продуктов для будущего использования.
Вирусная инфекция
[ редактировать ]Антивирусное лечение является одним из самых ранних предлагаемых медицинских применений на основе РНКи, и были разработаны два разных типа. Первый тип - нацеливаться на вирусные РНК. Многие исследования показали, что нацеливание на вирусные РНК может подавлять репликацию многочисленных вирусов, включая ВИЧ , [ 147 ] HPV , [ 148 ] гепатит А , [ 149 ] гепатит В , [ 150 ] вирус гриппа , [ 151 ] [ 152 ] [ 153 ] [ 154 ] Респираторный синцитиальный вирус (RSV), [ 154 ] Коронавирус (Офис-Сарс), [ 154 ] Аденовирус [ 154 ] и вирус кори . [ 155 ] Другая стратегия состоит в том, чтобы заблокировать начальные вирусные записи, нацеливаясь на гены клеток -хозяев. [ 156 ] Например, подавление хемокиновых рецепторов ( CXCR4 и CCR5 ) на клетках -хозяева может предотвратить проникновение вируса ВИЧ. [ 157 ]
Рак
[ редактировать ]В то время как традиционная химиотерапия может эффективно убивать раковые клетки, отсутствие специфичности для различения нормальных клеток и раковых клеток в этих методах лечения обычно вызывает тяжелые побочные эффекты. Многочисленные исследования показали, что RNAi может обеспечить более специфический подход к ингибированию роста опухоли, нацеливаясь на гены, связанные с раком (то есть онкоген ). [ 158 ] Также было предложено, что RNAi может повысить чувствительность раковых клеток к химиотерапевтическим агентам , обеспечивая комбинаторный терапевтический подход с химиотерапией. [ 159 ] Другое потенциальное лечение на основе RNAi заключается в ингибировании инвазии и миграции клеток . [ 160 ]
По сравнению с химиотерапией или другими противораковыми препаратами, существует множество преимуществ препарата siRNA. [ 161 ] МиРНК действует на посттранскрипционную стадию экспрессии генов, поэтому она не модифицирует и не изменяет ДНК в вредном эффекте. [ 161 ] МиРНК также можно использовать для получения конкретного ответа определенным типом способа, например, путем понижения подавления экспрессии генов. [ 161 ] В одной раковой клетке миРНК может вызвать драматическое подавление экспрессии генов только с несколькими копиями. [ 161 ] Это происходит путем молчания генов, способствующих развитию рака, с RNAi, а также нацеливалось на последовательность мРНК. [ 161 ]
Препараты РНКи лечат рак, молчав определенные гены, способствующие развитию рака. [ 161 ] Это делается путем дополнения генов рака с помощью RNAi, таких как сохранение последовательностей мРНК в соответствии с лекарственным средством RNAi. [ 161 ] В идеале, RNAi должен быть введен и/или химически модифицирован, чтобы RNAi мог более эффективно достигать раковых клеток. [ 161 ] Поглощение и регуляция РНКи контролируются почками. [ 161 ]
Неврологические заболевания
[ редактировать ]Стратегии RNAi также демонстрируют потенциал для лечения нейродегенеративных заболеваний . Исследования в клетках и у мыши показали, что специально нацеливающиеся на амилоидные бета -продуцирующие гены (например, BACE1 и APP) с помощью RNAi могут значительно уменьшить количество Aβ -пептида, которое коррелирует с причиной болезни Альцгеймера . [ 162 ] [ 163 ] [ 164 ] Кроме того, эти подходы, основанные на молчане, также обеспечивают многообещающие результаты в лечении болезни Паркинсона и заболевания полиглутамина . [ 165 ] [ 166 ] [ 167 ]
Стимуляция иммунного ответа
[ редактировать ]Иммунная система человека разделена на две отдельные ветви: врожденная иммунная система и адаптивная иммунная система. [ 168 ] Врожденная иммунная система является первой защитой от инфекции и реагирует на патогенные микроорганизмы. [ 168 ] С другой стороны, адаптивная иммунная система, система, которая развивалась позже врожденной, состоит в основном из высокоспециализированных В и Т -клеток, которые обучены реагировать на определенные части патогенных молекул. [ 168 ]
Задача между старыми патогенами и новыми, помогла создать систему охраняемых ячеек и частиц, которые называются безопасной структурой. [ 168 ] Эта структура дала людям армию систем, которые ищут и разрушают частицы захватчиков, такие как патогены, микроскопические организмы, паразиты и инфекции. [ 168 ] Безопасная структура млекопитающих разработала для включения siRNA в качестве инструмента для обозначения вирусного загрязнения, что позволило siRNA создать интенсивный врожденный иммунный ответ. [ 168 ]
МиРНК контролируется врожденной иммунной системой, которая может быть разделена на острые воспалительные реакции и противовирусные реакции. [ 168 ] Воспалительный ответ создается сигналами из малых сигнальных молекул или цитокинов. [ 168 ] К ним относятся интерлейкин-1 (IL-1), интерлейкин-6 (IL-6), интерлейкин-12 (IL-12) и фактор некроза опухоли α (TNF-α). [ 168 ] Врожденная иммунная система генерирует воспаление и противовирусные реакции, которые вызывают рецепторы распознавания схемы высвобождения (PRR). [ 168 ] Эти рецепторы помогают маркировать, какие патогены представляют собой вирусы, грибы или бактерии. [ 168 ] Более того, важность миРНК и врожденной иммунной системы состоит в том, чтобы включить больше PRR, чтобы помочь распознать различные структуры РНК. [ 168 ] Это повышает вероятность того, что миРНК приводит к тому, что иммуностимуляторный ответ в случае патогена. [ 168 ]
Еда
[ редактировать ]RNAi использовался для генетически инженера растений для получения более низких уровней природных растительных токсинов. Такие методы используют преимущества стабильного и наследственного фенотипа РНКи в растительных запасах. Семена хлопка богаты диетическим белком , но естественным образом содержат токсичный тексичный продукт , что делает их непригодными для потребления человека. RNAi использовался для производства хлопковых запасов, семенам которых содержат снижение уровня дельта-кадиненсинтазы , ключевого фермента в производстве госсипола, не влияя на производство фермента в других частях растения, где сама госсипол сам важен для предотвращения повреждений от вредителей растений. [ 169 ]
Усилия по развитию успешно снизили уровень аллергенов у томата растений [ 170 ] и укрепление растений, таких как помидоры с диетическими антиоксидантами . [ 171 ] Глушение RNAi альфа-амилазы также использовалось для снижения грибкового роста Aspergillus flavus в кукурузе, которая в противном случае загрязняла бы ядра опасными афлатоксинами . [ 172 ] С молчанием синтазы лахримативного фактора у лука продуцировала безрелый лук, а RNAi использовался в генах BP1 у Rabeseeds для улучшения фотосинтеза. [ 173 ] Гены SBEIIA и SBEIIB в пшенице были нацелены на пшеницу для получения более высоких уровней амилозы , чтобы улучшить функцию кишечника, [ 174 ] и Travella et al. В 2006 году использовался RNAI для функциональной геномики. Исследование гексаплоидных хлебных гонок , в то время как Scofield et al. 2005 для изучения механизма сопротивления, обеспеченного LR21 против ржавчины листьев пшеницы в гексаплоидной пшенице . [ 175 ]
Инсектициды
[ редактировать ]RNAi находится в стадии разработки в качестве инсектицида , используя несколько подходов, включая ген -инженерию и актуальное применение. [ 4 ] Клетки в средней кишке некоторых насекомых занимают молекулы дцРНК в процессе, называемом РНК -экологией. [ 176 ] У некоторых насекомых эффект является системным, поскольку сигнал распространяется по всему телу насекомых (называется системным РНКи). [ 177 ]
Животные, подвергшиеся воздействию РНКи в дозах в миллионы раз выше, чем предполагаемые уровни воздействия на человека, не показывают побочных эффектов. [ 178 ] RNAi обладает различным эффектом у разных видов чешуекрылых (бабочек и мотыльков). [ 179 ]
Доктор Dr Diosophila spp., Bombyx mori , locua spp., Spodopters spp., Tribolium castaneum , Nilaparpata Lugens , Helicavarpa armigera и Apis mellifera - это модели, которые широко используются для изучения того, как работает RNAI, в частности, налога инсектив. У Musca Domestica есть два гена Rogy2 и Glossina Morsitans Three, найденные Lewis et al. 2016 и Hain et al. 2010 год. [ 180 ] [ 181 ] В случае miRNA пути Diuraphis noxia имеет два назад1 с, M. Domestica Two DCR1 S, Acyrthosiphon Pisum два каждого из AGE1 и LOQS и DCR1 и четыре паши . В то время как в Пирне и Г. Морситанец А. Писум имеют по два или три назад 3 с каждая. [ 181 ] Это привело к выявлению будущих целей развития инсектицидов , а также способам действия и причины устойчивости к инсектицидам других инсектицидов. [ 181 ]
Трансгенные растения
[ редактировать ]Трансгенные культуры были сделаны для экспрессии дцРНК, тщательно выбравшей для замораживания важных генов в вредителях -мишенях. Эти дцРНК предназначены для влияния только на насекомых, которые экспрессируют специфические генные последовательности. В качестве доказательства принципа , в 2009 году исследование показало РНК, которые могли бы убить любого из четырех видов фруктовых мух, не причиняя вреда другим трем. [ 4 ]
Актуально
[ редактировать ]В качестве альтернативы дцРНК может быть поставлена без генотической инженерии. Одним из подходов является добавление их в орошательную воду. систему растений Молекулы поглощаются в сосудистую и ядовитые насекомые, питающиеся им. Другой подход включает в себя опрыскивание дцРНК, как обычный пестицид. Это позволило бы более быстро адаптировать к сопротивлению. Такие подходы потребуют недорогих источников DSRNAS, которых в настоящее время не существует. [ 4 ]
Функциональная геномика
[ редактировать ]Подходы к дизайну библиотек RNAi по всему геному могут потребовать большей изощренности, чем конструкция одной миРНК для определенного набора экспериментальных условий. Искусственные нейронные сети часто используются для проектирования SIRNA библиотек [ 182 ] и предсказать их вероятной эффективности при нокдауне гена. [ 183 ] Массовый геномный скрининг широко рассматривается как многообещающий метод аннотации генома и вызвал разработку высокопроизводительных методов скрининга на основе микрочипов . [ 184 ] [ 185 ]
Скрининг генома
[ редактировать ]Исследование RNAI в масштабе генома зависит от технологии высокопроизводительного скрининга (HTS). Технология RNAI HTS позволяет скринингу потерь по потере функции по всему геному и широко используется при идентификации генов, связанных со специфическими фенотипами. Эта технология была провозглашена как потенциальная вторая волна геномики, после первой волны геномики экспрессии генов микрочипов и полиморфизма с одним нуклеотидным полиморфизмом . платформ обнаружения [ 186 ] Одним из основных преимуществ скрининга RNAi в масштабе генома является его способность одновременно опросить тысячи генов. С возможностью генерирования большого количества данных на эксперимент, скрининг RNAI в масштабе генома привел к взрыву скоростей генерации данных. Использование таких крупных наборов данных является фундаментальной проблемой, требующей подходящей статистики/биоинформатики. Основной процесс скрининга RNAI на основе клеток включает в себя выбор библиотеки RNAI, надежных и стабильных типов клеток, трансфекцию с помощью агентов RNAI, лечения/инкубации, обнаружения сигналов, анализа и идентификации важных генов или терапевтических мишеней. [ 187 ]
История
[ редактировать ]РНКИ открытие
[ редактировать ]
Процесс РНКи был назван «совместной категорией» и «подача», когда наблюдалось до знания механизма, связанного с РНК. Открытию РНКи предшествовали сначала наблюдения за ингибированием транскрипции антисмысловой РНК, экспрессируемой в трансгенных растениях, [ 189 ] и более прямо по сообщениям о неожиданных результатах в экспериментах, выполняемых учеными растения в Соединенных Штатах и Нидерландах в начале 1990 -х годов. [ 190 ] В попытке изменить цветы цветов в петуниях , исследователи ввели дополнительные копии гена, кодирующей халконесинтазу , ключевой фермент для пигментации цветов в растения петунии обычно розового или фиолетового цветового цвета. Ожидалось, что ген из сверхэкспрессии приведет к более темным цветам, но вместо этого заставил некоторые цветы иметь менее заметный фиолетовый пигмент, иногда в разнообразных узорах, что указывает на то, что активность халконе синтазы была существенно снижена или подавлялась специфичным для контекста. Позже это будет объяснено как результат того, что трансген вставлен рядом с промоторами в противоположном направлении в различных положениях по всему геномам некоторых трансформантов, что приводит к экспрессии антисмысловых транскриптов и молчания генов, когда эти промоторы активны. [ Цитация необходима ] Еще одно раннее наблюдение за РНКИ произошло из исследования грибной нейроспоры Crassa , [ 191 ] Хотя это не было сразу признано связанным. Дальнейшее исследование явления у растений показало, что подавление было связано с посттранскрипционным ингибированием экспрессии генов посредством повышенной скорости деградации мРНК. [ 192 ] Это явление называлось совместно-супрессией экспрессии генов , но молекулярный механизм оставался неизвестным. [ 193 ]
Вскоре после этого вирологи растений , работающие над повышением устойчивости растений к вирусным заболеваниям, наблюдали за аналогичным неожиданным явлением. Хотя было известно, что растения, экспрессирующие вирус-специфические белки, демонстрировали повышенную толерантность или устойчивость к вирусной инфекции, не ожидалось, что растения, несущие только короткие, некодирующие области последовательностей вирусной РНК, будут иметь сходные уровни защиты. Исследователи полагали, что вирусная РНК, продуцируемая трансгенами, также может ингибировать репликацию вируса. [ 194 ] Обратный эксперимент, в котором короткие последовательности генов растений были введены в вирусы, показал, что целевой ген был подавлен в инфицированном растении. [ 195 ] Это явление было помечено «индуцированным вирусом генов» (VIGS), [ 175 ] и набор таких явлений был коллективно называется пост -транскрипционным молчанием генов. [ 196 ]
После этих первоначальных наблюдений в растениях лаборатории искали это явление в других организмах. [ 197 ] [ 198 ] Первый случай молчания РНК у животных была задокументирована в 1996 году, когда Го и Кемфу заметили, что, введя смысл и антисмысловую РНК в мРНК PAR-1 у Caenorhabditis elegans , вызвало деградацию сообщения PAR-1. [ 199 ] Считалось, что эта деградация была вызвана одноцепочечной РНК (SSRNA), но два года спустя, в 1998 году, Fire и Mello обнаружили, что эта способность молчать экспрессию гена PAR-1 была фактически вызвана двумя цельной РНК (дцРНК ) [ 199 ] Крейг С. Мелло и Эндрю Фьер 1998 года Nature Paper сообщил о мощном эффекте молчания генов после введения двойной страной РНК в C. elegans . [ 200 ] При исследовании регуляции выработки мышечного белка они наблюдали, что ни мРНК, ни антисмысловая инъекции РНК не оказывали влияния на продукцию белка, но двухцепочечная РНК успешно замолчала целевой ген. В результате этой работы они придумали термин RNAI . Это открытие представляло собой первую идентификацию причинного агента для этого явления. Fire и Mello были удостоены Нобелевской премии 2006 года по физиологии или медицине . [ 6 ] [ 201 ]
RNAI Therapeutics
[ редактировать ]Сразу после пожара и новаторского открытия Мелло, Elbashir et al. Обнаружено, используя синтетически сделанную небольшую интерферирующую РНК (миРНК), можно было нацелить молчание специфических последовательностей в гене, а не молчать весь ген. [ 202 ] Только год спустя МакКаффри и его коллеги продемонстрировали, что это специфическое из последовательности молчание имело терапевтические применения, нацеленные на последовательность вируса гепатита С у трансгенных мышей. [ 203 ] С тех пор несколько исследователей пытались расширить терапевтические применения RNAI, в частности, стремятся нацелить гены, которые вызывают различные виды рака . [ 204 ] [ 205 ] К 2006 году первое применение для достижения клинических испытаний проводилось в лечении дегенерации желтого пятна и респираторного синцитиального вируса . [ 206 ] Четыре года спустя было начато первое клиническое исследование фазы I фазы с использованием системы доставки наночастиц для нацеливания на твердые опухоли . [ 207 ]
FDA одобрило первое лекарство на основе SIRNA ( Patisiran ) в 2018 году. Givosiran и Lumasiran впоследствии получили одобрение FDA для лечения AHP и PH1 в 2019 и 2020 годах соответственно. [ 113 ] Inclisiran получил одобрение EMA в 2020 году на лечение высокого уровня холестерина и в настоящее время находится в рассмотрении FDA. [ 208 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Saurabh S, Vidyarthi AS, Prasad D (март 2014 г.). «Вмешательство РНК: концепция реальности в улучшении урожая» . Планта 239 (3): 543–64. BIBCODE : 2014Plant.239..543S . doi : 10.1007/s00425-013-2019-5 . PMID 24402564 .
- ^ Вайс Б., Дэвидкова Г., Чжоу Л.В. (март 1999 г.). «Антисмысленная генная терапия РНК для изучения и модуляции биологических процессов» . Клеточные и молекулярные науки о жизни . 55 (3): 334–58. doi : 10.1007/s000180050296 . PMC 11146801 . PMID 10228554 . S2CID 9448271 .
- ^ Jump up to: а беременный Matranga C, Tomari Y, Shin C, Bartel DP, Zamore PD (ноябрь 2005 г.). «Расщепление пассажирских цепей облегчает сборку миРНК в EAGY2-содержащие ферментные комплексы RNAI» . Клетка . 123 (4): 607–20. doi : 10.1016/j.cell.2005.08.044 . PMID 16271386 .
- ^ Jump up to: а беременный в дюймовый Kupferschmidt K (август 2013 г.). «Смертельная доза РНК» . Наука . 341 (6147): 732–3. Bibcode : 2013sci ... 341..732K . doi : 10.1126/science.341.6147.732 . PMID 23950525 .
- ^ Macrae IJ, Zhou K, Li F, Repic A, Brooks AN, Cande WZ, et al. (Январь 2006 г.). «Структурная основа для двухцепочечной обработки РНК Dicer». Наука . 311 (5758): 195–8. Bibcode : 2006sci ... 311..195m . doi : 10.1126/science.1121638 . PMID 16410517 . S2CID 23785494 .
- ^ Jump up to: а беременный в дюймовый Данехольт Б. "Расширенная информация: РНК -интерференция" . Нобелевская премия по физиологии или медицине 2006 . Архивировано из оригинала 20 января 2007 года . Получено 25 января 2007 года .
- ^ Багасра О, Приллиман К.Р. (август 2004 г.). «Интерференция РНК: молекулярная иммунная система». Журнал молекулярной гистологии . 35 (6): 545–53. Citeseerx 10.1.1.456.1701 . doi : 10.1007/s10735-004-2192-8 . PMID 15614608 . S2CID 2966105 .
- ^ Бернштейн Е., Кауди А.А., Хаммонд С.М., Хэннон Г.Дж. (январь 2001 г.). «Роль бинтатной рибонуклеазы на стадии инициации помехи РНК». Природа . 409 (6818): 363–6. Bibcode : 2001natur.409..363b . doi : 10.1038/35053110 . PMID 11201747 . S2CID 4371481 .

- ^ Siomi H, Siomi MC (январь 2009 г.). «На пути к чтению кода РНК-интерференции». Природа . 457 (7228): 396–404. Bibcode : 2009natur.457..396s . doi : 10.1038/nature07754 . PMID 19158785 . S2CID 205215974 .
Zamore PD, Tuschl T, Sharp PA, Bartel DP (март 2000 г.). «РНКи: двухцепочечная РНК направляет АТФ-зависимое расщепление мРНК с 21-23 интервалами нуклеотидов» . Клетка . 101 (1): 25–33. doi : 10.1016/s0092-8674 (00) 80620-0 . PMID 10778853 .
Вермейлен А., Бехлен Л., Рейнольдс А., Вольфсон А., Маршалл В.С., Карпилоу Дж. И др. (Май 2005 г.). «Вклад структуры дцРНК в специфичность и эффективность DICER» . РНК . 11 (5): 674–82. doi : 10.1261/rna.7272305 . PMC 1370754 . PMID 15811921 .
Кастанотто Д., Росси Дж.Дж. (январь 2009 г.). «Обещания и подводные камни терапии на основе РНК-интерференции» . Природа . 457 (7228): 426–33. Bibcode : 2009natur.457..426c . doi : 10.1038/nature07758 . PMC 2702667 . PMID 19158789 . - ^ Jump up to: а беременный Qiu S, Adema CM, Lane T (2005). «Вычислительное исследование нецелевых эффектов интерференции РНК» . Исследование нуклеиновых кислот . 33 (6): 1834–47. doi : 10.1093/nar/gki324 . PMC 1072799 . PMID 15800213 .
- ^ Jump up to: а беременный Наканиши К (сентябрь 2016 г.). «Анатомия RISC: как небольшие РНК и шапероны активируют аргонатные белки?» Полем Wiley Междисциплинарные обзоры: РНК . 7 (5): 637–60. doi : 10.1002/wrna.1356 . PMC 5084781 . PMID 27184117 .
- ^ Liang C, Wang Y, Murota Y, Liu X, Smith D, Siomi MC, et al. (Сентябрь 2015). «TAF11 собирает комплекс нагрузки RISC для повышения эффективности RNAi» . Молекулярная клетка . 59 (5): 807–18. doi : 10.1016/j.molcel.2015.07.006 . PMC 4560963 . PMID 26257286 .
- ^ Ahlquist P (май 2002). «РНК-зависимые РНК-полимеразы, вирусы и молчание РНК». Наука . 296 (5571): 1270–3. Bibcode : 2002sci ... 296.1270a . doi : 10.1126/science.1069132 . PMID 12016304 . S2CID 42526536 .
- ^ Робертс Т.С. (2015). «Машина для микроРНК». МикроРНА: Основная наука . Достижения в области экспериментальной медицины и биологии. Тол. 887. С. 15–30. doi : 10.1007/978-3-319-22380-3_2 . ISBN 978-3-319-22379-7 Полем PMID 26662984 .
- ^ Jump up to: а беременный в дюймовый Паркер Г., Экерт Д., Басс Б. (2006). «RDE-4 преимущественно связывает длинную дцРНК, и ее димеризация необходима для расщепления дсРНК к миРНК» . РНК . 12 (5): 807–18. doi : 10.1261/rna.2338706 . PMC 1440910 . PMID 16603715 .
- ^ Jump up to: а беременный Baulcombe DC (январь 2007 г.). «Молекулярная биология. Усиленное молчание». Наука . 315 (5809): 199–200. doi : 10.1126/science.1138030 . PMID 17218517 . S2CID 46285020 .
- ^ Jump up to: а беременный Pak J, Fire A (январь 2007 г.). «Отдельные популяции первичных и вторичных эффекторов во время RNAi у C. elegans». Наука . 315 (5809): 241–4. Bibcode : 2007sci ... 315..241p . doi : 10.1126/science.1132839 . PMID 17124291 . S2CID 46620298 .
- ^ Jump up to: а беременный Sijen T, Steiner FA, Thijssen KL, Plasterk RH (январь 2007 г.). «Вторичные миРНК возникают из -за бесвязного синтеза РНК и образуют отдельный класс». Наука . 315 (5809): 244–7. Bibcode : 2007sci ... 315..244S . doi : 10.1126/science.1136699 . PMID 17158288 . S2CID 9483460 . (Втянут, см doi : 10.1126/science.abf4837 , PMID 33214285 , Retraction Watch )
- ^ Wang Ql, Li ZH (май 2007 г.). «Функции микроРНК в растениях» . Границы в биологии . 12 : 3975–82. doi : 10.2741/2364 . PMC 2851543 . PMID 17485351 . S2CID 23014413 .
Zhao Y, Srivastava D (апрель 2007 г.). «Просмотр функции микроРНК». Тенденции в биохимических науках . 32 (4): 189–97. doi : 10.1016/j.tibs.2007.02.006 . PMID 17350266 . - ^ Грегори Р.И., Чендримада Т.П., Шихаттар Р. (2006). «Биогенез микроРНК: выделение и характеристика микропроцессорного комплекса». Протоколы микроРНК . Методы в молекулярной биологии. Тол. 342. С. 33–47. doi : 10.1385/1-59745-123-1: 33 . ISBN 978-1-59745-123-9 Полем PMID 16957365 .
- ^ Pfeffer S, Zavolan M, Grässer FA, Chien M, Russo JJ, Ju J, et al. (Апрель 2004 г.). «Идентификация кодируемых вирусом микроРНК». Наука . 304 (5671): 734–6. Bibcode : 2004sci ... 304..734p . doi : 10.1126/science.1096781 . PMID 15118162 . S2CID 25287167 .
- ^ Qureshi A, Thakur N, Monga I, Thakur A, Kumar M (1 января 2014 года). «Virmirna: комплексный ресурс для экспериментально проверенных вирусных микроРНК и их мишеней» . База данных . 2014 : BAU103. doi : 10.1093/база данных/BAU103 . PMC 4224276 . PMID 25380780 .
- ^ Pillai RS, Bhattacharyya SN, Filipowicz W (2007). «Репрессия синтеза белка с помощью miRNAS: сколько механизмов?». Тенденции Cell Biol . 17 (3): 118–26. doi : 10.1016/j.tcb.2006.12.007 . PMID 17197185 .
- ^ Окамура К., Ишизука А., Сиоми Х., Сиоми М. (2004). «Отличительные роли для аргонатных белков в небольших путях расщепления РНК, направленных на РНК» . Гены Дев . 18 (14): 1655–66. doi : 10.1101/gad.1210204 . PMC 478188 . PMID 15231716 .
- ^ Ли Y, Накахара К., Фам Дж, Ким К, Хе З, Сонтхеймер Е. и др. (2004). «Отдельные роли для Drosophila Dicer-1 и Dicer-2 в путях молчания siRNA/miRNA» . Клетка . 117 (1): 69–81. doi : 10.1016/s0092-8674 (04) 00261-2 . PMID 15066283 .
- ^ mirbase.org
- ^ Jump up to: а беременный Friedman RC, Farh KK, Burge CB, Bartel DP (2009). «Большинство мРНК млекопитающих являются консервативными целями микроРНК» . Геном Res . 19 (1): 92–105. doi : 10.1101/gr.082701.108 . PMC 2612969 . PMID 18955434 .
- ^ Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, et al. (Февраль 2005 г.). «Анализ микрочипов показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа . 433 (7027): 769–73. Bibcode : 2005natur.433..769L . doi : 10.1038/nature03315 . PMID 15685193 . S2CID 4430576 .
- ^ Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N (сентябрь 2008 г.). «Широко распространенные изменения в синтезе белка, вызванные микроРНК». Природа . 455 (7209): 58–63. Bibcode : 2008natur.455 ... 58 с . doi : 10.1038/nature07228 . PMID 18668040 . S2CID 4429008 .
- ^ Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP (сентябрь 2008 г.). «Влияние микроРНК на выход белка» . Природа . 455 (7209): 64–71. Bibcode : 2008natur.455 ... 64b . doi : 10.1038/nature07242 . PMC 2745094 . PMID 18668037 .
- ^ Palmer EI, из Campos SG, Campos M, De Souza NC, Warrior ID, Carvalho AL, et al. (Июль 2011). «Механизмы и роль микронной дерегуляции в начале и прогрессировании рака» . Биология генетика и молекулярная . 34 (3): 363–70. Doi : 10.1590/s1415-47572011000300001 . PMC 3168173 . PMID 21931505 .
- ^ Бернштейн С, Бернштейн Х (май 2015). «Эпигенетическое восстановление репарации ДНК при прогрессировании на желудочно -кишечный рак» . Всемирный журнал желудочно -кишечной онкологии . 7 (5): 30–46. doi : 10.4251/wjgo.v7.i5.30 . PMC 4434036 . PMID 25987950 .
- ^ Maffioletti E, Tardito D, Gennarelli M, Bocchio-Chiavetto L (2014). «Микро -шпионы от мозга до периферии: новые подсказки из исследований микроРНК при нейропсихиатрических расстройствах» . Границы в клеточной нейробиологии . 8 : 75. DOI : 10.3389/fncel.2014.00075 . PMC 3949217 . PMID 24653674 .
- ^ Mellios N, Sur M (2012). «Новая роль микроРНК в расстройствах шизофрении и аутистического спектра» . Границы в психиатрии . 3 : 39. doi : 10.3389/fpsyt.2012.00039 . PMC 3336189 . PMID 22539927 .
- ^ Geaghan M, Cairns MJ (август 2015 г.). «МикроРНК и посттранскрипционная дисрегуляция в психиатрии» . Биологическая психиатрия . 78 (4): 231–9. doi : 10.1016/j.biopsych.2014.12.009 . HDL : 1959.13/1335073 . PMID 25636176 .
- ^ Liu Q, Rand TA, Kalidas S, Du F, Kim HE, Smith DP, et al. (Сентябрь 2003 г.). «R2D2, мост между инициацией и эффекторными этапами пути Drosophila RNAI». Наука . 301 (5641): 1921–5. Bibcode : 2003sci ... 301.1921L . doi : 10.1126/science.1088710 . PMID 14512631 . S2CID 41436233 .
- ^ Грегори Р.И., Чендримада Т.П., Куч Н., Шихаттар Р (ноябрь 2005 г.). «Человеческий RISC объединяет биогенез микроРНК и посттранскрипционное молчание генов» . Клетка . 123 (4): 631–40. doi : 10.1016/j.cell.2005.10.022 . PMID 16271387 .
- ^ Jump up to: а беременный Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, et al. (2004). Молекулярная клеточная биология (5 -е изд.). WH Freeman: Нью -Йорк, Нью -Йорк. ISBN 978-0-7167-4366-8 .
- ^ Leuschner PJ, Ameres SL, Kueng S, Martinez J (март 2006 г.). «Расщепление пассажирской нити миРНК во время сборки RISC в клетках человека» . Embo сообщает . 7 (3): 314–20. doi : 10.1038/sj.embor.7400637 . PMC 1456892 . PMID 16439995 .
- ^ Jump up to: а беременный Хейли Б., Замор П.Д. (июль 2004 г.). «Кинетический анализ ферментного комплекса РНКи». Природа структурная и молекулярная биология . 11 (7): 599–606. doi : 10.1038/nsmb780 . PMID 15170178 . S2CID 12400060 .
- ^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (октябрь 2003 г.). «Асимметрия в сборке ферментного комплекса РНКи» . Клетка . 115 (2): 199–208. doi : 10.1016/s0092-8674 (03) 00759-1 . PMID 14567917 .
- ^ Preall JB, He Z, Gorra JM, Sontheimer EJ (март 2006 г.). «Короткий мешающий отбор РНК не зависит от полярности обработки дцРНК во время РНКи у дрозофилы» . Текущая биология . 16 (5): 530–5. Bibcode : 2006cbio ... 16..530p . doi : 10.1016/j.cub.2006.01.061 . PMID 16527750 .
- ^ Tomari Y, Matranga C, Haley B, Martinez N, Zamore PD (ноябрь 2004 г.). «Белковый датчик для асимметрии миРНК». Наука . 306 (5700): 1377–80. Bibcode : 2004sci ... 306.1377t . doi : 10.1126/science.1102755 . PMID 15550672 . S2CID 31558409 .
- ^ MA JB, Yuan YR, Meister G, Pei Y, Tuschl T, Patel DJ (март 2005 г.). «Структурная основа для 5'-эндов-специфического распознавания направляющей РНК белком A. fulgidus piwi» . Природа . 434 (7033): 666–70. Bibcode : 2005natur.434..666m . doi : 10.1038/nature03514 . PMC 4694588 . PMID 15800629 .
- ^ Sen G, Wehrman T, Blau H (2005). «Трансляция мРНК не является предпосылкой для небольшого мешающего РНК-опосредованного расщепления мРНК». Дифференциация . 73 (6): 287–93. doi : 10.1111/j.1432-0436.2005.00029.x . PMID 16138829 . S2CID 41117614 .
- ^ Гу С., Росси Дж. (2005). «Необеспечение РНКи от активной трансляции в клетках млекопитающих» . РНК . 11 (1): 38–44. doi : 10.1261/rna.7158605 . PMC 1370689 . PMID 15574516 .
- ^ Sen G, Blau H (2005). «Аргонат 2/RISC находится на местах распада мРНК млекопитающих, известного как цитоплазматические тела». Nat Cell Biol . 7 (6): 633–6. doi : 10.1038/ncb1265 . PMID 15908945 . S2CID 6085169 .
- ^ Лиан С., Джакимиу А., Эйстатхии Т., Хэмел Дж., Фрицлер М., Чан Е. (2006). «Тела GW, микроРНК и клеточный цикл» . Клеточный цикл . 5 (3): 242–5. doi : 10.4161/cc.5.3.2410 . PMID 16418578 .
- ^ Jakymiw A, Lian S, Eystathioy T, Li S, Satoh M, Hamel J, et al. (2005). «Разрушение P тел ухудшает вмешательство РНК млекопитающих». Nat Cell Biol . 7 (12): 1267–74. doi : 10.1038/ncb1334 . PMID 16284622 . S2CID 36630239 .
- ^ Hammond S, Bernstein E, Beach D, Hannon G (2000). «РНК-направленная нуклеаза опосредует посттранскрипционное молчание генов в клетках дрозофилы». Природа . 404 (6775): 293–6. Bibcode : 2000natur.404..293H . doi : 10.1038/35005107 . PMID 10749213 . S2CID 9091863 .
- ^ Холмквист Г.П., Эшли Т. (2006). «Организация хромосом и модификация хроматина: влияние на функцию генома и эволюцию». Цитогенетические и геномные исследования . 114 (2): 96–125. doi : 10.1159/000093326 . PMID 16825762 . S2CID 29910065 .
- ^ Вердель А., Цзя С., Гербер С., Сугияма Т., Гиги С., Гревал С.И. и др. (Январь 2004 г.). «RNAI-опосредованное нацеливание гетерохроматина комплексом RITS» . Наука . 303 (5658): 672–6. Bibcode : 2004sci ... 303..672V . doi : 10.1126/science.1093686 . PMC 3244756 . PMID 14704433 .
- ^ Ирвин Д.В., Заратиги М., Толиа Н.Х., Гото Д.Б., Читвуд Д.Х., Вон М.В. и др. (Август 2006 г.). «Аргонаутная нарезка требуется для гетерохроматического молчания и распространения». Наука . 313 (5790): 1134–7. Bibcode : 2006sci ... 313.1134i . doi : 10.1126/science.1128813 . PMID 16931764 . S2CID 42997104 .
- ^ Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI, Martienssen RA (сентябрь 2002 г.). «Регуляция гетерохроматического молчания и метилирования лизина-9 гистона H3 с помощью РНКи» . Наука . 297 (5588): 1833–7. Bibcode : 2002sci ... 297.1833v . doi : 10.1126/science.1074973 . PMID 12193640 . S2CID 2613813 .
- ^ Volpe T, Schramke V, Hamilton GL, White SA, Teng G, Martienssen RA, et al. (2003). «РНК -интерференция требуется для нормальной функции центромеры у делящихся дрожжей». Исследование хромосом . 11 (2): 137–46. doi : 10.1023/a: 1022815931524 . PMID 12733640 . S2CID 23813417 .
- ^ Li LC, Okino St, Zhao H, Pookot D, Place RF, Urakami S, et al. (Ноябрь 2006 г.). «Маленькие дцРНК индуцируют активацию транскрипции в клетках человека» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (46): 17337–42. Bibcode : 2006pnas..10317337L . doi : 10.1073/pnas.0607015103 . PMC 1859931 . PMID 17085592 .
- ^ Noma K, Sugiyama T, Cam H, Verdel A, Zofall M, Jia S, et al. (Ноябрь 2004 г.). «RITS действует в CIS, чтобы способствовать опосредованному интерферентному интерферентному интерференции транскрипции и посттранскрипционного молчания» . Природа генетика . 36 (11): 1174–80. doi : 10.1038/ng1452 . PMID 15475954 .
- ^ Sugiyama T, Cam H, Verdel A, Moazed D, Grewal Si (январь 2005 г.). «РНК-зависимая РНК-полимераза является важным компонентом самосогловой сцепления сцепления гетерохроматина с выработкой миРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (1): 152–7. Bibcode : 2005pnas..102..152S . doi : 10.1073/pnas.0407641102 . PMC 544066 . PMID 15615848 .
- ^ Wang F, Koyama N, Nishida H, Haraguchi T, Reith W, Tsukamoto T (июнь 2006 г.). «Сборка и поддержание гетерохроматина, инициируемого трансгеновыми повторами, не зависят от пути интерференции РНК в клетках млекопитающих» . Молекулярная и клеточная биология . 26 (11): 4028–40. doi : 10.1128/mcb.02189-05 . PMC 1489094 . PMID 16705157 .
- ^ Басс Бл (2002). «Редактирование РНК с помощью аденозин деминаз, которые действуют на РНК» . Ежегодный обзор биохимии . 71 : 817–46. doi : 10.1146/annurev.biochem.71.110601.135501 . PMC 1823043 . PMID 12045112 .
- ^ Басс BL (апрель 2000 г.). «Двухцепочечная РНК как матрица для молчания генов» . Клетка . 101 (3): 235–8. doi : 10.1016/s0092-8674 (02) 71133-1 . PMID 10847677 .
- ^ Luciano DJ, Mirsky H, Vendetti NJ, Maas S (август 2004 г.). «РНК -редактирование предшественника miRNA» . РНК . 10 (8): 1174–7. doi : 10.1261/rna.7350304 . PMC 1370607 . PMID 15272117 .
- ^ Jump up to: а беременный Ян В., Чендримада Т.П., Ван К., Хигучи М., Сибург П.П., Шихаттар Р. и др. (Январь 2006 г.). «Модуляция обработки и экспрессии микроРНК посредством редактирования РНК адаром деминаз» . Природа структурная и молекулярная биология . 13 (1): 13–21. doi : 10.1038/nsmb1041 . PMC 2950615 . PMID 16369484 .
- ^ Yang W, Wang Q, Howell KL, Lee JT, Cho DS, Murray JM, et al. (Февраль 2005 г.). «Адар1 РНК -деаминаза ограничивает короткую мешающую эффективность РНК в клетках млекопитающих» . Журнал биологической химии . 280 (5): 3946–53. doi : 10.1074/jbc.m407876200 . PMC 2947832 . PMID 15556947 .
- ^ Нишикура К (декабрь 2006 г.). «Редактор встречает глушитель: перекрестные помехи между редактированием РНК и интерференцией РНК» . Природа обзор молекулярной клеточной биологии . 7 (12): 919–31. doi : 10.1038/nrm2061 . PMC 2953463 . PMID 17139332 .
- ^ Jump up to: а беременный в Saumet A, Lecellier CH (2006). "Антивирусное молчание РНК: мы похожи на растения?" Полем Ретровирология . 3 (1): 3. DOI : 10.1186/1742-4690-3-3 . PMC 1363733 . PMID 16409629 .
- ^ Jones L, Ratcliff F, Baulcombe DC (май 2001 г.). «РНК-направленное молчание транскрипционного гена в растениях может быть унаследовано независимо от триггера РНК и требует MET1 для поддержания» . Текущая биология . 11 (10): 747–57. Bibcode : 2001cbio ... 11..747j . doi : 10.1016/s0960-9822 (01) 00226-3 . PMID 11378384 . S2CID 16789197 .
- ^ Humphreys DT, Westman BJ, Martin Di, Preiss T (ноябрь 2005 г.). «МикроРНК контролирует инициацию трансляции путем ингибирования эукариотического фактора инициации 4E/CAP и поли (A) хвоста» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (47): 16961–6. Bibcode : 2005pnas..10216961H . doi : 10.1073/pnas.0506482102 . PMC 1287990 . PMID 16287976 .
- ^ Darocha WD, Otsu K, Teixeira SM, Donelson Je (февраль 2004 г.). «Тесты цитоплазматического интерференции РНК (RNAi) и построение промоторной системы TTRACECLINE-индуцируемой T7 в Trypanosoma Cruzi». Молекулярная и биохимическая паразитология . 133 (2): 175–86. doi : 10.1016/j.molbiopara.2003.10.005 . PMID 14698430 .
- ^ Робинсон К.А., Беверли С.М. (май 2003 г.). «Улучшения в эффективности трансфекции и тестов подходов РНК -интерференции (RNAi) в простейском паразите Leishmania». Молекулярная и биохимическая паразитология . 128 (2): 217–28. doi : 10.1016/s0166-6851 (03) 00079-3 . PMID 12742588 .
- ^ Aravind L, Watanabe H, Lipman DJ, Koonin EV (октябрь 2000 г.). «Специфичная для линии потеря и дивергенция функционально связанных генов у эукариот» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (21): 11319–24. Bibcode : 2000pnas ... 9711319a . doi : 10.1073/pnas.200346997 . PMC 17198 . PMID 11016957 .
- ^ Дринненберг И.А., Вайнберг Д.Е., Се К.Т., Мауэр Дж.П., Вулф К.Х., Финк Г.Р. и др. (Октябрь 2009 г.). «РНКИ в начинающих дрожжах» . Наука . 326 (5952): 544–550. Bibcode : 2009sci ... 326..544d . doi : 10.1126/science.1176945 . PMC 3786161 . PMID 19745116 .
- ^ Nakayashiki H, Kadotani N, Mayama S (июль 2006 г.). «Эволюция и диверсификация белков с молчанием РНК в грибах» (PDF) . Журнал молекулярной эволюции . 63 (1): 127–35. Bibcode : 2006jmole..63..127n . doi : 10.1007/s00239-005-0257-2 . PMID 16786437 . S2CID 22639035 . Архивировано из оригинала (PDF) 6 августа 2020 года . Получено 4 декабря 2019 года .
- ^ Морита Т., Мочизуки Ю., Айба Х (март 2006 г.). «Трансляционная репрессия достаточна для молчания генов бактериальными небольшими некодирующими РНК в отсутствие разрушения мРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (13): 4858–63. Bibcode : 2006pnas..103.4858m . doi : 10.1073/pnas.0509638103 . PMC 1458760 . PMID 16549791 .
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Йи, Кунин Э.В. (март 2006 г.). «Предполагаемая иммунная система на основе РНК в прокариотах: вычислительный анализ прогнозируемого ферментативного механизма, функциональные аналогии с эукариотической РНКи и гипотетические механизмы действия» . Биология прямой . 1 : 7. doi : 10.1186/1745-6150-1-7 . PMC 1462988 . PMID 16545108 .
- ^ Stram Y, Kuzntzova L (июнь 2006 г.). «Ингибирование вирусов с помощью РНК -интерференции» . Вирусные гены . 32 (3): 299–306. doi : 10.1007/s11262-005-6914-0 . PMC 7088519 . PMID 16732482 .
- ^ Блевинс Т., Раджесваран Р., Шивапрасад П.В., Беназаряны Д., Си-Аммур А., Парк Х.С. и др. (2006). «Четыре изделия из растений опосредуют вирусный малый РНК -биогенез, а ДНК -вирус индуцировал молчание» . Исследование нуклеиновых кислот . 34 (21): 6233–46. doi : 10.1093/nar/gkl886 . PMC 1669714 . PMID 17090584 .
- ^ Palauqui JC, Elmayan T, Pollien JM, Vaucheret H (август 1997 г.). «Системное приобретенное молчание: трансгене-специфическое посттранскрипционное молчание передается путем прививки от молчаливных запасов к не приличным Scions» . Embo Journal . 16 (15): 4738–45. doi : 10.1093/emboj/16.15.4738 . PMC 1170100 . PMID 9303318 .
- ^ Voinnet O (август 2001 г.). «Глушение РНК как иммунная система растений против вирусов». Тенденции в генетике . 17 (8): 449–59. doi : 10.1016/s0168-9525 (01) 02367-8 . PMID 11485817 .
- ^ Люси AP, Guo HS, Li WX, Ding SW (апрель 2000 г.). «Подавление посттранскрипционного молчания гена вирусным белком растения, локализованное в ядре» . Embo Journal . 19 (7): 1672–80. doi : 10.1093/emboj/19.7.1672 . PMC 310235 . PMID 10747034 .
- ^ Mérai Z, Kerényi Z, Kertész S, Magna M, Lakatos L, Silhavy D (июнь 2006 г.). «Двойное связывание РНК может быть общей вирусной стратегией РНК растительной РНК для подавления молчания РНК» . Журнал вирусологии . 80 (12): 5747–56. doi : 10.1128/jvi.01963-05 . PMC 1472586 . PMID 16731914 .
- ^ Katiyar-Agarwal S, Morgan R, Dahlbeck D, Borsani O, Villegas A, Zhu JK, et al. (Ноябрь 2006 г.). «Индуцируемая патогеном эндогенная миРНК при иммунитете растений» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (47): 18002–7. Bibcode : 2006pnas..10318002K . doi : 10.1073/pnas.0608258103 . PMC 1693862 . PMID 17071740 .
- ^ Фриц Дж. Х., Жирардин С.Е., Филпотт DJ (июнь 2006 г.). «Врожденная иммунная защита через РНК -вмешательство». Science's Stke . 2006 (339): PE27. doi : 10.1126/stke.3392006pe27 . PMID 16772641 . S2CID 33972766 .
- ^ Замбон Р.А., Вахария Вн, Ву Л.П. (май 2006 г.). «РНКи является противовирусным иммунным ответом на вирус дцРНК у дрозофилы меланогастер» . Клеточная микробиология . 8 (5): 880–9. doi : 10.1111/j.1462-5822.2006.00688.x . PMID 16611236 . S2CID 32439482 .
- ^ Wang XH, Aliyari R, Li WX, Li HW, Kim K, Carthew R, et al. (Апрель 2006 г.). «РНК -интерференция направляет врожденный иммунитет против вирусов у взрослых дрозофилы» . Наука . 312 (5772): 452–4. Bibcode : 2006sci ... 312..452W . doi : 10.1126/science.1125694 . PMC 1509097 . PMID 16556799 .
- ^ Lu R, Maduro M, Li F, Li HW, Broitman-Maduro G, Li WX, et al. (Август 2005 г.). «Репликация вируса животных и RNAi-опосредованное антивирусное молчание у Caenorhabditis elegans» . Природа . 436 (7053): 1040–1043. Bibcode : 2005natur.436.1040L . doi : 10.1038/nature03870 . PMC 1388260 . PMID 16107851 .
- ^ Wilkins C, Dishongh R, Moore SC, Whitt MA, Chow M, Machaca K (август 2005 г.). «Вмешательство РНК является механизм антивирусной защиты у Caenorhabditis elegans». Природа . 436 (7053): 1044–7. Bibcode : 2005natur.436.1044W . doi : 10.1038/nature03957 . PMID 16107852 . S2CID 4431035 .
- ^ Berkhout B, Haasnoot J (май 2006 г.). «Взаимодействие между вирусной инфекцией и механизмом клеточной РНК -интерференции» . Письма Febs . 580 (12): 2896–902. BIBCODE : 2006FEBSL.580.2896B . doi : 10.1016/j.febslet.2006.02.070 . PMC 7094296 . PMID 16563388 .
- ^ Schütz S, Sarnow P (январь 2006 г.). «Взаимодействие вирусов с путем помехи РНК млекопитающих» . Вирусология . 344 (1): 151–7. doi : 10.1016/j.virol.2005.09.034 . PMID 16364746 .
- ^ Cullen BR (июнь 2006 г.). «Является ли РНК -интерференция вовлечена в внутренний противовирусенный иммунитет у млекопитающих?». Природа иммунология . 7 (6): 563–7. doi : 10.1038/ni1352 . PMID 16715068 . S2CID 23467688 .
- ^ Maillard PV, Ciaudo C, Marchais A, Li Y, Jay F, Ding SW, et al. (Октябрь 2013). «Антивирусное интерференцию РНК в клетках млекопитающих» . Наука . 342 (6155): 235–8. Bibcode : 2013sci ... 342..235M . doi : 10.1126/science.1241930 . PMC 3853215 . PMID 24115438 .
- ^ Li Y, Lu J, Han Y, Fan X, Ding SW (октябрь 2013). «РНК -интерференция функционирует как механизм противовирусного иммунитета у млекопитающих» . Наука . 342 (6155): 231–4. Bibcode : 2013sci ... 342..231L . doi : 10.1126/science.1241911 . PMC 3875315 . PMID 24115437 .
- ^ Li HW, Ding SW (октябрь 2005 г.). «Противовирусное молчание у животных» . Письма Febs . 579 (26): 5965–73. Bibcode : 2005 Febsl.579.5965L . doi : 10.1016/j.febslet.2005.08.034 . PMC 1350842 . PMID 16154568 .
- ^ Carrington JC, Ambros V (июль 2003 г.). «Роль микроРНК в развитии растений и животных» . Наука . 301 (5631): 336–8. Bibcode : 2003sci ... 301..336c . doi : 10.1126/science.1085242 . PMID 12869753 . S2CID 43395657 .
- ^ Lee RC, Feinbaum RL, Ambros V (декабрь 1993 г.). «Гетерохронный ген C. elegans ген Lin-4 кодирует небольшие РНК с антисмысловой комплементарностью LIN-14» . Клетка . 75 (5): 843–54. doi : 10.1016/0092-8674 (93) 90529-y . PMID 8252621 .
- ^ Palatnik JF, Allen E, Wu X, Schommer C, Schwab R, Carrington JC, et al. (Сентябрь 2003 г.). «Контроль морфогенеза листьев микроРНК» . Природа . 425 (6955): 257–63. Bibcode : 2003natur.425..257p . doi : 10.1038/nature01958 . PMID 12931144 . S2CID 992057 .
- ^ Zhang B, Pan X, Cobb GP, Anderson TA (январь 2006 г.). «МИККОРНА ПАНД: небольшая регуляторная молекула с большим ударом» . Биология развития . 289 (1): 3–16. doi : 10.1016/j.ydbio.2005.10.036 . PMID 16325172 .
- ^ Jones-Rhoades MW, Bartel DP, Bartel B (2006). «МикроРНК и их регулирующие роли в растениях». Ежегодный обзор биологии растений . 57 : 19–53. doi : 10.1146/annurev.arplant.57.032905.105218 . PMID 16669754 . S2CID 13010154 .
- ^ Zhang B, Pan X, Cobb GP, Anderson TA (февраль 2007 г.). «МикроРНК в качестве онкогена и опухолевых супрессоров» . Биология развития . 302 (1): 1–12. doi : 10.1016/j.ydbio.2006.08.028 . PMID 16989803 .
- ^ Cerutti H, Casas-Mollano JA (август 2006 г.). «О происхождении и функциях РНК-опосредованного молчания: от протистов к человеку» . Текущая генетика . 50 (2): 81–99. doi : 10.1007/s00294-006-0078-x . PMC 2583075 . PMID 16691418 .
- ^ Anantharaman V, Koonin EV, Aravind L (апрель 2002 г.). «Сравнительная геномика и эволюция белков, участвующих в метаболизме РНК» . Исследование нуклеиновых кислот . 30 (7): 1427–64. doi : 10.1093/nar/30.7.1427 . PMC 101826 . PMID 11917006 .
- ^ Voorhoeve PM, Agami R (январь 2003 г.). «Нокдаун встает». Тенденции в биотехнологии . 21 (1): 2–4. doi : 10.1016/s0167-7799 (02) 00002-1 . PMID 12480342 .
- ^ Munkácsy G, Sztupinszki Z, Herman P, Bán B, Monetary Z, Szarvas N, et al. (Сентябрь 2016 г.). «Валидация эффективности молчания РНКи с использованием данных генов показывает 18,5% частоту отказов в 429 независимых экспериментах» . Молекулярная терапия: нуклеиновые кислоты . 5 (9): E366. Doi : 10.1038/mtna.2016.66 . PMC 5056990 . PMID 27673562 .
- ^ Найто Ю., Ямада Т., Мацумия Т., Уи-Тей К, Сайго К., Моришита С. (июль 2005 г.). «DSCHECK: высокочувствительное программное обеспечение для поиска вне цели для двухцепочечных РНК-опосредованных интерференций РНК» . Исследование нуклеиновых кислот . 33 (Проблема веб -сервера): W589–91. doi : 10.1093/nar/gki419 . PMC 1160180 . PMID 15980542 .
- ^ Хеншель А., Буххольц Ф., Хаберманн Б (июль 2004 г.). «Deqor: веб-инструмент для проектирования и контроля качества SIRNAS» . Исследование нуклеиновых кислот . 32 (Проблема веб -сервера): W113–20. doi : 10.1093/nar/gkh408 . PMC 441546 . PMID 15215362 .
- ^ Найто Ю., Ямада Т., Уи-Тей К., Моришита С., Сайго К (июль 2004 г.). «Sidirect: высокоэффективное программное обеспечение для дизайна SIRNA для мировой миРНК для интерференции РНК млекопитающих» . Исследование нуклеиновых кислот . 32 (Проблема веб -сервера): W124–9. doi : 10.1093/nar/gkh442 . PMC 441580 . PMID 15215364 .
- ^ Naito Y, Ui-Tei K, Nishikawa T, Takebe Y, Saigo K (июль 2006 г.). «Sivirus: веб-программное обеспечение для дизайна антивирусной миРНК для очень расходящихся вирусных последовательностей» . Исследование нуклеиновых кислот . 34 (Проблема веб -сервера): W448–50. doi : 10.1093/nar/gkl214 . PMC 1538817 . PMID 16845046 .
- ^ Рейнольдс А., Андерсон Э.М., Вермейлен А., Федоров Ю., Робинсон К., Лике Д. и др. (Июнь 2006 г.). «Индукция ответа интерферона миРНК зависит от длины типа и дуплекса» . РНК . 12 (6): 988–93. doi : 10.1261/rna.2340906 . PMC 1464853 . PMID 16611941 .
- ^ Stein P, Zeng F, Pan H, Schultz RM (октябрь 2005 г.). «Отсутствие неспецифических эффектов РНК интерференции, вызванных длинной двухцепочечной РНК в ооцитах мыши» . Биология развития . 286 (2): 464–71. doi : 10.1016/j.ydbio.2005.08.015 . PMID 16154556 .
- ^ Brummelkamp TR, Bernards R , Agami R (апрель 2002 г.). «Система для стабильной экспрессии коротких мешающих РНК в клетках млекопитающих». Наука . 296 (5567): 550–3. Bibcode : 2002sci ... 296..550B . doi : 10.1126/science.1068999 . HDL : 1874/15573 . PMID 11910072 . S2CID 18460980 .
- ^ Tiscornia G, Tergaonkar V, Galimi F, Verma IM (май 2004 г.). «Индуцируемое рекомбиназой РНК-интерференция, опосредованная лентивирусными векторами» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (19): 7347–51. Bibcode : 2004pnas..101.7347t . doi : 10.1073/pnas.0402107101 . PMC 409921 . PMID 15123829 .
- ^ Вентура А., Мейснер А., Диллон К.П., Макманус М., Шарп П.А., Ван Паридж Л. и др. (Июль 2004 г.). «Условное условное интерференцию РНК от трансгенов» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (28): 10380–5. Bibcode : 2004pnas..10110380V . doi : 10.1073/pnas.0403954101 . PMC 478580 . PMID 15240889 .
- ^ Jump up to: а беременный Адамс Д. (5 июля 2018 г.). Полем Журнал медицины Новой Англии 379 (1): 11–2 doi : 10.1056/nejmo 1716153 HDL : 2445/138257 . 29972753PMID S2CID 205102839 .
- ^ Balwani M, Sardh E, Ventura P, Peiró PA, Rees DC, Stölzel U, et al. (11 июня 2020 г.). «Фаза 3 Исследования RNAI Therapeutic Givosiran для острого прерывистого порфирии» . Новая Англия Журнал медицины . 382 (24): 2289–2301. doi : 10.1056/nejmoa1913147 . ISSN 0028-4793 . PMID 32521132 .
- ^ SF SF, Hullon MJ, Cocha P, Al. (1 апреля 2021 г.). Полем Новый английский журнал медицины 384 (13): 1216–1 doi : 10.1056/ nejmo2 ISSN 1533-4 PMID 33789010 .
- ^ Adams D, Tournev IL, Taylor MS, Coelho T, Planté-Bordeneuve V, Berk JL, et al. (Март 2023 г.). «Эффективность и безопасность Vutrisiran для пациентов с наследственным транстиретино-опосредованным амилоидозом с полинейропатией: рандомизированное клиническое исследование» . Амилоид . 30 (1): 18–26. doi : 10.1080/13506129.2022.2091985 . HDL : 1959,4/UNSWORKS_80922 . ISSN 1744-2818 . PMID 35875890 .
- ^ «Метка: опатро-говорящая инъекция, ликерный комплекс » Daily Med 10 Там 2021.
- ^ «Пакет одобрения наркотиков: givlaari (Givosiran) инъекция» . FDA .
- ^ Gonzalez-Aseguinolaza G (11 июня 2020 года). «Givosiran - управление РНК -вмешательством для борьбы с атаками Порфирии». Новая Англия Журнал медицины . 382 (28–4793): 2366–2367. doi : 10.1056/nejme2010986 . PMID 32521139 . S2CID 219592948 .
- ^ «Гивлаари» . Европейское агентство лекарств .
- ^ Puy H, Gouya L, Deybach JC (2010). «Порфирия». Лансет . 375 (9718): 924–937. doi : 10.1016/s0140-6736 (09) 61925-5 . PMID 20226990 . S2CID 208791867 .
- ^ Simon A, Pompilus F, Querbes W, Wei A, Strzok S, Penz C, et al. (2018). «Перспектива пациента на острую прерывистую порфирию с частыми атаками: заболевание с прерывистыми и хроническими проявлениями» . Пациент . 11 (5): 527–537. doi : 10.1007/s40271-018-0319-3 . PMC 6132435 . PMID 29915990 .
- ^ Пишик Э., Кауппинен Р. (2015). «Обновление клинического лечения острого прерывистого порфирии» . Применение клинической генетики . 8 : 201–214. doi : 10.2147/tacg.s48605 . PMC 4562648 . PMID 26366103 .
- ^ Бисселл Д.М., Лай Дж.С., Мейстер Р.К., Блан П.Д. (2015). «Роль дельта-аминолевулиновой кислоты в симптомах острой порфирии» . Американский журнал медицины . 128 (3): 313–317. doi : 10.1016/j.amjmed.2014.10.026 . PMC 4339446 . PMID 25446301 .
- ^ «Метка: givlaarivisiran впрыскивание натрия, раствор» . Daily Med . 5 апреля 2022 года.
- ^ «Пакет одобрения лекарств: oxlumo» . FDA .
- ^ "Oxlumo" . Европейское агентство лекарств .
- ^ «Метка: Oxlumo-Lumasiran внедрение, раствор» . Daily Med . 2 декабря 2021 года.
- ^ SF SF, Hullon MJ, Cocha P, Al. (1 апреля 2021 г.). Полем Журнал медицины Новой Англии 384 (13): 1216–1 doi : 10.1056/ nejmo2 PMID 33789010 . S2CID 232482623 .
- ^ Исследование CF (27 ноября 2023 г.). «Снимки для испытаний наркотиков: Amvuttra» . FDA .
- ^ «Амвуттра | Европейское агентство лекарственных средств» . www.ema.europa.eu . Получено 11 января 2024 года .
- ^ Guerriaud M, Kohli E (2022). «Препараты и регулирование на основе РНК: к необходимой эволюции определений, выпущенных из законодательства Европейского Союза» . Границы в медицине . 9 doi : 10.3389/fmed.2022.1012497 . ISSN 2296-858X . PMC 9618588 . PMID 36325384 .
- ^ Jump up to: а беременный Kanasty R, Dorkin Jr, Vegas A, Anderson D (ноябрь 2013). «Материалы доставки для терапии миРНК». Природные материалы . 12 (11): 967–77. Bibcode : 2013natma..12..967k . doi : 10.1038/nmat3765 . PMID 24150415 .
- ^ Wittrup A, Lieberman J (сентябрь 2015 г.). «Обивает болезнь: отчет о прогрессе о терапии миРНК» . Nature Reviews Genetics . 16 (9): 543–52. doi : 10.1038/nrg3978 . PMC 4756474 . PMID 26281785 .
- ^ Saluzaro EA, Salueiro WG, Moro RP, TL, AL. (Там 2019). «Interperence Rena имеет результат продолжающихся элеганов » нуклеиновых кислот Исследование 47 (8): 3957–3 doi : 10.1093/nar/gkz154 . PMC 6486631 . PMID 30838421 .
- ^ Li W, Szoka FC (24 марта 2007 г.). «Наночастицы на основе липидов для доставки нуклеиновых кислот». Pharm Res . 24 (3): 438–449. doi : 10.1007/s11095-006-9180-5 . PMID 17252188 . S2CID 9995555 .
- ^ Leung AK, Tam Yy, Cullis PR (2014). «Липидные наночастицы для короткой мешающей доставки РНК». Невирусные векторы для генной терапии- перенос генов на основе липидов и полимеров . Достижения в области генетики. Тол. 88. С. 71–110. doi : 10.1016/b978-0-12-800148-6.00004-3 . ISBN 978-0-12-800148-6 Полем PMC 7149983 . PMID 25409604 .
- ^ Джон Ламаттина (15 апреля 2014 г.). «Окрашивание Big Pharma на RNAi показывает, что новые технологии не гарантируют успех в исследованиях и разработках» . Форбс .
- ^ Эрик Бендер (1 сентября 2014 г.). «Второе пришествие РНКИ» . Ученый .
- ^ Баден Лр (2021). «Эффективность и безопасность вакцины мРНК-1273 SARS-COV-2» . Новая Англия Журнал медицины . 384 (5): 403–416. doi : 10.1056/nejmoa2035389 . PMC 7787219 . PMID 33378609 .
- ^ Фицджеральд К., Уайт С., Бородовский А., Беттенкур Б.Р., Страйс А., Клаузен В. (январь 2017 г.). «Очень прочный терапевтический ингибитор РНКи PCSK9» . Новая Англия Журнал медицины . 376 (1): 41–51. doi : 10.1056/nejmoa1609243 . PMC 5778873 . PMID 27959715 .
- ^ Kruspe S, Giangrande P (2017). «Аптамер-сирна химеры: открытие, прогресс и будущие перспективы» . Биомедицины . 5 (4): 45. doi : 10.3390/Biomedicines5030045 . PMC 5618303 . PMID 28792479 .
- ^ Balwani M, Sardh E, Ventura P, Peiró PA, Rees DC, Stölzel U, et al. (11 июня 2020 г.). «Фаза 3 Исследования RNAI Therapeutic Givosiran для острого прерывистого порфирии» . Новая Англия Журнал медицины . 382 (24): 2289–2301. doi : 10.1056/nejmoa1913147 . PMID 32521132 . S2CID 219586624 .
- ^ Andersson M, Melander M, Pojmark P, Larsson H, Bülow L, Hofvander P (2006). «Целевое подавление генов с помощью РНК-интерференции: эффективный метод производства линий с высоким амилозовым картофелем» . Журнал биотехнологии . 123 (2): 137–148. doi : 10.1016/j.jbiotec.2005.11.001 . PMID 16466822 .
- ^ Фриззи А., Хуан С. (2010). «Постукивание путей молчания РНК для биотехнологии растений» . Plant Biotechnology Journal . 8 (6): 655–677. doi : 10.1111/j.1467-7652.2010.00505.x . PMID 20331529 .
- ^ Parrott W, Chassy B, Ligon J, Meyer L, Patrick J, Zhou J, et al. (2010). «Применение принципов оценки безопасности продуктов питания и корм для оценки трансгенных подходов к модуляции генов в сельскохозяйственных культурах» . Британская ассоциация промышленных биологических исследований . 48 (7): 1773–1790. doi : 10.1016/j.fct.2010.04.017 . PMID 20399824 .
- ^ Berkhout B (апрель 2004 г.). «Вмешательство РНК как антивирусное подход: нацеливание на ВИЧ-1». Современное мнение о молекулярной терапии . 6 (2): 141–5. PMID 15195925 .
- ^ Цзян М., Милнер Дж. (Сентябрь 2002 г.). «Селективное молчание экспрессии вирусных генов в HPV-позитивных клетках карциномы шейки матки человека, обработанных миРНК, праймер интерференции РНК» . Онкоген . 21 (39): 6041–8. doi : 10.1038/sj.onc.1205878 . PMID 12203116 .
- ^ Кусов Ю., Канда Т., Палменберг А., Сгро Дж.Ю., Гаусс-Мюллер В. (июнь 2006 г.). «Сильком вирусной инфекции гепатита А небольшими мешающими РНК» . Журнал вирусологии . 80 (11): 5599–610. doi : 10.1128/jvi.01773-05 . PMC 1472172 . PMID 16699041 .
- ^ Цзя Ф., Чжан Юз, Лю С.М. (октябрь 2006 г.). «Система на основе ретровируса для стабильной тишины генов вируса гепатита В путем РНК-интерференции» . Биотехнологические письма . 28 (20): 1679–85. doi : 10.1007/s10529-006-9138-z . PMID 16900331 . S2CID 34511611 .
- ^ Li YC, Kong LH, Cheng BZ, Li KS (декабрь 2005 г.). «Строительство векторов экспрессии миРНК вируса гриппа и их ингибирующее влияние на умножение вируса гриппа». Птиевые заболевания . 49 (4): 562–73. doi : 10.1637/7365-041205R2.1 . PMID 16405000 . S2CID 86214047 .
- ^ Ханна М., Саксена Л., Раджпут Р., Кумар Б., Прасад Р. (2015). «Генсинг генов: терапевтический подход к борьбе с вирусными инфекциями гриппа». Будущая микробиология . 10 (1): 131–40. doi : 10.2217/fmb.14.94 . PMID 25598342 .
- ^ Раджпут Р., Ханна М., Кумар П., Кумар Б., Шарма С., Гупта Н. и др. (Декабрь 2012 г.). «Небольшая мешающая РНК, нацеленная на транскрипт Нонтруктурного гена 1, ингибирует репликацию вируса гриппа А у экспериментальных мышей». Нуклеиновая терапия . 22 (6): 414–22. doi : 10.1089/nat.2012.0359 . PMID 23062009 .
- ^ Jump up to: а беременный в дюймовый Asha K, Kumar P, Sanicas M, Meseko CA, Khanna M, Kumar B (декабрь 2018 г.). «Достижения в терапии на основе нуклеиновых кислот против респираторных вирусных инфекций» . Журнал клинической медицины . 8 (1): 6. doi : 10.3390/jcm8010006 . PMC 6351902 . PMID 30577479 .
- ^ Hu L, Wang Z, Hu C, Liu X, Yao L, Li W, et al. (2005). «Ингибирование умножения вируса кори в клеточной культуре на РНК -интерференцию». Acta virologica . 49 (4): 227–34. PMID 16402679 .
- ^ Куреши А., Такур Н., Монга И., Такур А., Кумар М. (2014). «Virmirna: комплексный ресурс для экспериментально проверенных вирусных микроРНК и их мишеней» . База данных . 2014 . doi : 10.1093/база данных/BAU103 . PMC 4224276 . PMID 25380780 .
- ^ Кроу С. (2003). «Подавление экспрессии рецептора хемокинов с помощью РНК-интерференции позволяет ингибировать репликацию ВИЧ-1, Martínez et al.» . СПИД . 17 (Suppl 4): S103–5. doi : 10.1097/00002030-200317004-00014 . PMID 15080188 .
- ^ Fuchs U, Damm-Welk C, Borkhardt A (август 2004 г.). «Сильком генов, связанных с болезнью, небольшими мешающими РНК». Современное молекулярное лекарство . 4 (5): 507–17. doi : 10.2174/1566524043360492 . PMID 15267222 .
- ^ Cioca DP, Aoki Y, Kiyosawa K (февраль 2003 г.). «Интерференция РНК является функциональным путем с терапевтическим потенциалом в клеточных линиях миелоидного лейкоза человека» . Генная терапия рака . 10 (2): 125–33. doi : 10.1038/sj.cgt.7700544 . PMID 12536201 .
- ^ Lapteva N, Yang AG, Sanders DE, Strube RW, Chen Sy (январь 2005 г.). «Нокдаун CXCR4 с помощью небольшой мешающей РНК отменяет рост опухоли молочной железы in vivo» . Генная терапия рака . 12 (1): 84–9. doi : 10.1038/sj.cgt.7700770 . PMID 15472715 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Xu CF, Wang J (1 февраля 2015 г.). «Системы доставки для разработки лекарственных средств SIRNA при терапии рака» . Азиатский журнал фармацевтических наук . 10 (1): 1–12. doi : 10.1016/j.ajps.2014.08.011 .
- ^ Singer O, Marr RA, Rockenstein E, Crews L, Coufal Ng, Gage FH, et al. (Октябрь 2005 г.). «Нацеливание на BACE1 с миРНК ослабевает невропатологию болезни Альцгеймера в трансгенной модели». Nature Neuroscience . 8 (10): 1343–9. doi : 10.1038/nn1531 . PMID 16136043 . S2CID 6978101 .
- ^ Rodríguez-Lebron E, Gouvion CM, Moore SA, Davidson BL, Paulson HL (сентябрь 2009 г.). «Аллель-специфическая RNAi смягчает фенотипическое прогрессирование в трансгенной модели болезни Альцгеймера» . Молекулярная терапия . 17 (9): 1563–73. doi : 10.1038/mt.2009.123 . PMC 2835271 . PMID 19532137 .
- ^ Piedrahita D, Hernández I, López-Tobón A, Fedorov D, Obara B, Manjunath BS, et al. (Октябрь 2010). «Связывание CDK5 уменьшает нейрофибриллярные клубки у мышей трансгенных болезней Альцгеймера» . Журнал нейробиологии . 30 (42): 13966–76. doi : 10.1523/jneurosci.3637-10.2010 . PMC 3003593 . PMID 20962218 .
- ^ Рауль С., Баркер С.Д., Эбишер П (март 2006 г.). «Вирусное моделирование и коррекция нейродегенеративных заболеваний с помощью РНК-интерференции» . Генная терапия . 13 (6): 487–95. doi : 10.1038/sj.gt.3302690 . PMID 16319945 .
- ^ Harper SQ, Steper PD, He X, Eliason SL, Martins IH, Mao Q, et al. (Апрель 2005 г.). «РНК -интерференция улучшает моторные и невропатологические нарушения в модели мыши болезни Хантингтона» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (16): 5820–5. Bibcode : 2005pnas..102.5820h . doi : 10.1073/pnas.0501507102 . PMC 556303 . PMID 15811941 .
- ^ Будро Р.Л., Родригес-Леброн Э., Дэвидсон Б.Л. (апрель 2011 г.). «Медицина RNAi для мозга: прогрессирует и вызовы» . Молекулярная генетика человека . 20 (R1): R21–7. doi : 10.1093/hmg/ddr137 . PMC 3095054 . PMID 21459775 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Уайтхед К.А., Далман Дж., Лангер Р.С., Андерсон Д.Г. (2011). «Сильком или стимуляция? Доставка миРНК и иммунная система». Ежегодный обзор химической и биомолекулярной инженерии . 2 : 77–96. doi : 10.1146/annurev-chembioeng-061010-114133 . PMID 22432611 . S2CID 28803811 .
- ^ Sunilkumar G, Campbell LM, Puckhaber L, Stypanovic Rd, Rathore KS (ноябрь 2006 г.). «Инженерное хлопок для использования в питании человека путем тканевого специфического восстановления токсичного госсипола» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (48): 18054–9. doi : 10.1073/pnas.0605389103 . PMC 1838705 . PMID 17110445 .
- ^ Le LQ, Lorenz Y, Scheurer S, Fötisch K, Enrique E, Bartra J, et al. (Март 2006 г.). «Дизайн томатных фруктов с уменьшенной аллергенности путем DSRNAi-опосредованной ингибирования экспрессии NS-LTP (LYC E 3)» . Plant Biotechnology Journal . 4 (2): 231–42. doi : 10.1111/j.1467-7652.2005.00175.x . PMID 17177799 .
- ^ Niggeweg R, Michael AJ, Martin C (июнь 2004 г.). «Инженерные установки с повышенным уровнем антиоксидантной хлорогеновой кислоты». Nature Biotechnology . 22 (6): 746–54. doi : 10.1038/nbt966 . PMID 15107863 . S2CID 21588259 .
- ^ Гилберт М.К., Маджамдар Р., Раджасекаран К., Чен Зи, Вэй К., Сиплер С.М. и др. (Июнь 2018 г.). «РНК-интерференционное молчание гена альфа-амилазы (AMY1) у Aspergillus flavus снижает рост грибов и выработку афлатоксина в ядрах кукурузы». Планта 247 (6): 1465–1473. Bibcode : 2018plant.247.1465g . doi : 10.1007/s00425-018-2875-0 . PMID 29541880 . S2CID 3918937 .
- ^ Katoch R, Thakur N (март 2013 г.). «Вмешательство РНК: многообещающая техника улучшения традиционных культур». Международный журнал пищевых наук и питания . 64 (2): 248–59. doi : 10.3109/09637486.2012.713918 . PMID 22861122 . S2CID 45212581 .
- ^ Katoch R, Thakur N (март 2013 г.). «Достижения в области технологии интерференции РНК и ее влияния на улучшение питания, заболевание и контроль насекомых у растений». Прикладная биохимия и биотехнология . 169 (5): 1579–605. doi : 10.1007/s12010-012-0046-5 . PMID 23322250 . S2CID 23733295 .
- ^ Jump up to: а беременный Дауни Р.К., Лин М., Корси Б., Фику А., Лиллемо М., Оливер Р.П. и др. (27 июля 2021 г.). «Septoria nodorum пятно пшеницы: лечение заболеваний и размножение устойчивости перед лицом смещения динамики заболеваний и меняющейся среды». Фитопатология . 111 (6). Американское фитопатологическое общество : фито - 07–20–028. doi : 10.1094/phyto-07-20-0280-rvw . HDL : 20.500.11937/83208 . ISSN 0031-949X . PMID 33245254 . S2CID 227181536 .
- ^ Ivashuta S, Zhang Y, Wiggins BE, Ramaseshadri P, Segers GC, Johnson S, et al. (Май 2015). «Экологическая РНКи у травоядных насекомых» . РНК . 21 (5): 840–50. doi : 10.1261/rna.048116.114 . PMC 4408792 . PMID 25802407 .
- ^ Miller SC, Miyata K, Brown SJ, Tomoyasu Y (2012). «Расширение системных интерференций РНК в The Castaneum Tribolium Tribolium Tribolium : параметры, влияющие на эффективность RNAi» . Plos один . 7 (10): E47431. Bibcode : 2012ploso ... 747431M . doi : 10.1371/journal.pone.0047431 . PMC 3484993 . PMID 23133513 .
- ^ Petrick JS, Friedrich GE, Carleton SM, Kessenich CR, Silvanovich A, Zhang Y, et al. (Ноябрь 2016). «Корневой червя, активный РНК DVSNF7: Повторная доза пероральная токсикологическая оценка в поддержку безопасности человека и млекопитающих» . Регуляторная токсикология и фармакология . 81 : 57–68. doi : 10.1016/j.yrtph.2016.07.009 . PMID 27436086 .
- ^ Terenius O, Papanicolaou A, Garbutt JS, Eleftherianos I, Huvenne H, Kanginakudru S, et al. (Февраль 2011 г.). «Вмешательство РНК в Lepidoptera: обзор успешных и неудачных исследований и последствий для экспериментального дизайна» . Журнал физиологии насекомых . 57 (2): 231–45. Bibcode : 2011jinsp..57..231t . doi : 10.1016/j.jinsphys.2010.11.006 . HDL : 1854/LU-11101411 . PMID 21078327 .
- ^ Mongelli V, Saleh MC (29 сентября 2016 г.). «Ошибки не должны молчать: небольшие пути РНК и противовирусные реакции у насекомых» (PDF) . Ежегодный обзор вирусологии . 3 (1). Ежегодные обзоры : 573–589. doi : 10.1146/annurev-virology-1110615-042447 . ISSN 2327-056X . PMID 27741406 . S2CID 38499958 .
- ^ Jump up to: а беременный в Чжу К.Ю., Палли -С.Р. (7 января 2020 года). «Механизмы, приложения и проблемы РНК -интерференции насекомых» . Ежегодный обзор энтомологии . 65 (1). Ежегодные обзоры : 293–311. doi : 10.1146/annurev-ento-011019-025224 . ISSN 0066-4170 . PMC 9939233 . PMID 31610134 . S2CID 204702574 .
- ^ Huesken D, Lange J, Mickanin C, Weiler J, Asselbergs F, Warner J, et al. (Август 2005 г.). «Дизайн библиотеки миРНК в геноме с использованием искусственной нейронной сети». Nature Biotechnology . 23 (8): 995–1001. doi : 10.1038/nbt1118 . PMID 16025102 . S2CID 11030533 .
- ^ Ge G, Wong GW, Luo B (октябрь 2005 г.). «Прогнозирование эффективности нокдауна siRNA с использованием моделей искусственной нейронной сети». Биохимическая и биофизическая исследовательская коммуникация . 336 (2): 723–8. doi : 10.1016/j.bbrc.2005.08.147 . PMID 16153609 .
- ^ Джаниц М., Ванке Д., Лерач Х. (2006). «Высокопроизводительные интерференции РНК в функциональную геномику». РНК к медицину . Справочник по экспериментальной фармакологии. Тол. 173. С. 97–104. doi : 10.1007/3-540-27262-3_5 . ISBN 978-3-540-27261-8 Полем PMID 16594612 .
- ^ Vanhecke D, Janitz M (февраль 2005 г.). «Функциональная геномика с использованием высокопроизводительных интерференций РНК» . Drug Discovery сегодня . 10 (3): 205–12. doi : 10.1016/s1359-6446 (04) 03352-5 . HDL : 11858/00-001M-0000-0010-86E7-8 . PMID 15708535 . S2CID 9934291 .
- ^ Мэтсон Р.С. (2005). Применение технологии геномного и протеомного микрочипа при обнаружении лекарств . CRC Press. п. 6 ISBN 978-0-8493-1469-8 .
- ^ Чжан XHD (2011). Оптимальный высокопроизводительный скрининг: практический экспериментальный дизайн и анализ данных для исследования RNAI в масштабе генома . Издательство Кембриджского университета. с. IX - XIII. ISBN 978-0-521-73444-8 .
- ^ Мацке М.А., Мацке А.Дж. (2004). «Посадка семян новой парадигмы» . PLOS BIOL . 2 (5): E133. doi : 10.1371/journal.pbio.0020133 . PMC 406394 . PMID 15138502 .
- ^ Экер -младший, Дэвис Р.В. (август 1986 г.). «Ингибирование экспрессии генов в клетках растений путем экспрессии антисмысловой РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 83 (15): 5372–6. Bibcode : 1986pnas ... 83,5372E . doi : 10.1073/pnas.83.15.5372 . PMC 386288 . PMID 16593734 .
- ^ Наполи С., Лемье С., Йоргенсен Р. (апрель 1990 г.). «Введение гена химерного синтазы халкона в петунии приводит к обратимой совместном положении гомологичных генов в транс» . Растительная ячейка . 2 (4): 279–289. doi : 10.1105/tpc.2.4.279 . PMC 159885 . PMID 12354959 .
- ^ Романо Н., Макино Дж (ноябрь 1992). «Окружение: временная инактивация экспрессии генов в нейроспоре Crassa путем трансформации с гомологичными последовательностями». Молекулярная микробиология . 6 (22): 3343–53. doi : 10.1111/j.1365-2958.1992.tb02202.x . PMID 1484489 . S2CID 31234985 .
- ^ Van Blokland R, Van Der Geest N, Mol JN, Koter JM (1994). «Трансгеновая опосредованная подавление экспрессии халконасинтазы в Petunia Hybrida является результатом увеличения оборота РНК» . Растение J. 6 (6): 861–77. doi : 10.1046/j.1365-313X.1994.6060861.x .
- ^ Mol Jn, Van der Krol AR (1991). Антисмысловые нуклеиновые кислоты и белки: основы и применения . М. Деккер. с. 4, 136. ISBN 978-0-8247-8516-1 .
- ^ Кови С., Аль-Кафф Н., Лайнгара А., Тернер Д. (1997). «Растения борьбы с инфекцией путем молчания генов». Природа . 385 (6619): 781–2. Bibcode : 1997natur.385..781c . doi : 10.1038/385781a0 . S2CID 43229760 .
- ^ Kumagai MH, Donson J, Della-Cioppa G, Harvey D, Hanley K, Grill LK (февраль 1995 г.). «Цитоплазматическое ингибирование биосинтеза каротиноидов с РНК, полученной из вируса» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (5): 1679–83. Bibcode : 1995pnas ... 92.1679K . doi : 10.1073/pnas.92.5.1679 . PMC 42583 . PMID 7878039 .
- ^ Ratcliff F, Harrison BD, Baulcombe DC (июнь 1997 г.). «Сходство между вирусной защитой и молчанием генов у растений». Наука . 276 (5318): 1558–60. doi : 10.1126/science.276.5318.1558 . PMID 18610513 .
- ^ Guo S, Kemphues KJ (май 1995). «Par-1, ген, необходимый для установления полярности у эмбрионов C. elegans, кодирует предполагаемую киназу Ser/thr, которая асимметрично распределена» . Клетка . 81 (4): 611–20. doi : 10.1016/0092-8674 (95) 90082-9 . PMID 7758115 .
- ^ Pal-Bhadra M, Bhadra U, Birchler JA (август 1997 г.). «Космосенсия у дрозофилы: молчание генов алкогольной дегидрогеназы трансгенами белого ADH зависит от поликомба» . Клетка . 90 (3): 479–90. doi : 10.1016/s0092-8674 (00) 80508-5 . PMID 9267028 .
- ^ Jump up to: а беременный Sen GL, Blau HM (июль 2006 г.). «Краткая история РНКИ: молчание генов» . FASEB Journal . 20 (9): 1293–9. doi : 10.1096/fj.06-6014rev . PMID 16816104 . S2CID 12917676 .
- ^ Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC (февраль 1998 г.). «Мощное и специфическое генетическое помехи с помощью двухцепочечного РНК у Caenorhabditis elegans» . Природа . 391 (6669): 806–11. Bibcode : 1998natur.391..806f . doi : 10.1038/35888 . PMID 9486653 . S2CID 4355692 .
- ^ Daneholt B (2 октября 2006 г.). «Нобелевская премия по физиологии или медицине 2006» . nobelprize.org . Получено 30 октября 2017 года .
- ^ Elbashir S, Harborth J, Lendeckel W, et al. (2001). «Дуплексы 21-нуклеотидных РНК опосредуют интерференцию РНК в культивируемых клетках млекопитающих». Природа . 411 (6836): 494–498. Bibcode : 2001natur.411..494e . doi : 10.1038/35078107 . PMID 11373684 . S2CID 710341 .
- ^ McCaffrey AP, Meuse L, Pham TT, Conklin DS, Hannon GJ , Kay Ma (июль 2002 г.). «Вмешательство РНК у взрослых мышей». Природа . 418 (6893): 38–9. Bibcode : 2002natur.418 ... 38M . doi : 10.1038/418038a . PMID 12097900 . S2CID 4361399 .
- ^ Деви Гр (сентябрь 2006 г.). «Подходы на основе миРНК в терапии рака» . Генная терапия рака . 13 (9): 819–29. doi : 10.1038/sj.cgt.7700931 . PMID 16424918 .
- ^ Стена Н.Р., Ши (октябрь 2003 г.). «Маленькая РНК: можно ли использовать интерференцию РНК для терапии?». Лансет . 362 (9393): 1401–3. doi : 10.1016/s0140-6736 (03) 14637-5 . PMID 14585643 . S2CID 25034627 .
- ^ SAH D (2006). «Терапевтический потенциал РНК -интерференции на неврологические расстройства». Life Sci . 79 (19): 1773–80. doi : 10.1016/j.lfs.2006.06.011 . PMID 16815477 .
- ^ Davis ME , Zuckerman JE, Choi CH, Seligson D, Tolcher A, Alabi CA , et al. (Апрель 2010). «Свидетельство РНКи у людей от системно вводимой миРНК через целевые наночастицы» . Природа . 464 (7291): 1067–70. Bibcode : 2010natur.464.1067d . doi : 10.1038/nature08956 . PMC 2855406 . PMID 20305636 .
- ^ «Novartis получает одобрение ЕС для Leqvio (Inclisiran), первой в классе миРНК для более низкого уровня холестерина с двумя дозами в год» . Novartis . 11 декабря 2020 года.
Внешние ссылки
[ редактировать ]- Обзор процесса RNAI , от Кембриджского университета «Обнаженные ученые»
- Анимация процесса RNAi , от природы
- Nova Sciencenow объясняет RNAI -15-минутное видео о трансляции Nova , которое транслировалось на PBS , 26 июля 2005 г.
- Геномы молчания архивировали 10 августа 2019 года в экспериментах по интерференции RNA Machine Machine (RNAI) и биоинформатике в C. elegans для образования. Из Центра обучения ДНК Долана в лаборатории Холодного Спринг -Харбора.
- Экраны RNAI в C. elegans в 96-луночном жидком формате и их применение к систематической идентификации генетических взаимодействий (протокол)
- 2 американских «червя» выигрывают Нобелевскую работу за РНК , от Нью -Йорка Таймс
- молекулярной терапии Степень .
- Genomernai : база данных фенотипов из экспериментов по скринингу РНК в Drosophila melanogaster и Homo sapiens
- RNAI Архивировали 19 июня 2018 года в The Wayback Machine Инструменты