РНК

| Часть серии о |
| Генетика |
|---|
 |
|
|
Рибонуклеиновая кислота ( РНК ) — полимерная молекула, необходимая для большинства биологических функций, либо выполняя саму функцию ( некодирующая РНК ), либо образуя матрицу для производства белков ( информационная РНК ). РНК и дезоксирибонуклеиновая кислота (ДНК) являются нуклеиновыми кислотами . Нуклеиновые кислоты составляют одну из четырех основных макромолекул, необходимых для всех известных форм жизни . РНК собирается в виде цепочки нуклеотидов . Клеточные организмы используют информационную РНК ( мРНК для передачи генетической информации (с использованием азотистых оснований гуанина аденина , урацила , ) и цитозина , обозначаемых буквами G, U, A и C), которая направляет синтез определенных белков. Многие вирусы РНК кодируют свою генетическую информацию с помощью генома .
Некоторые молекулы РНК играют активную роль внутри клеток, катализируя биологические реакции, контролируя экспрессию генов или воспринимая и передавая ответы на клеточные сигналы. Одним из таких активных процессов является синтез белка — универсальная функция, при которой молекулы РНК направляют синтез белков на рибосомах . В этом процессе используются молекулы транспортной РНК ( тРНК ) для доставки аминокислот к рибосоме , где рибосомальная РНК ( рРНК ) затем связывает аминокислоты вместе с образованием закодированных белков.
Оно получило широкое признание в науке. [ 1 ] что на ранних этапах истории жизни на Земле белковых ферментов , до эволюции ДНК и, возможно , также , существовал « мир РНК », в котором РНК служила в обоих живых организмах способом хранения генетической информации — роль, выполненная Сегодня ДНК, за исключением РНК-вирусов , — и потенциально выполняющих каталитические функции в клетках — функцию, выполняемую сегодня белковыми ферментами, за примечательным и важным исключением рибосомы, которая является рибозимом .
Сравнение с ДНК
[ редактировать ]
Как и ДНК, большинство биологически активных РНК, включая мРНК , тРНК , рРНК , мяРНК и другие некодирующие РНК , содержат самокомплементарные последовательности, которые позволяют частям РНК сворачиваться. [ 6 ] и соединяются сами с собой, образуя двойные спирали. Анализ этих РНК показал, что они высокоструктурированы. В отличие от ДНК, их структуры состоят не из длинных двойных спиралей, а скорее из набора коротких спиралей, упакованных вместе в структуры, подобные белкам.
Таким образом, РНК могут осуществлять химический катализ (как ферменты). [ 7 ] Например, определение структуры рибосомы — комплекса РНК-белок, катализирующего сборку белков — показало, что ее активный центр полностью состоит из РНК. [ 8 ]
Структура
[ редактировать ]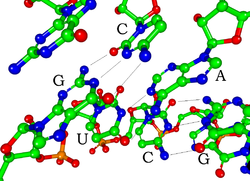
Каждый нуклеотид РНК содержит сахар рибозу с номерами атомов углерода от 1 до 5. К положению 1' присоединяется основание, обычно это аденин (А), цитозин (С), гуанин (G) или урацил (U). Аденин и гуанин являются пуринами , а цитозин и урацил — пиримидинами . Фосфатная группа присоединена к 3'-положению одной рибозы и 5' - положению следующей. Каждая из фосфатных групп имеет отрицательный заряд, что делает РНК заряженной молекулой (полианионом). Основания образуют водородные связи между цитозином и гуанином, между аденином и урацилом и между гуанином и урацилом. [ 9 ] Однако возможны и другие взаимодействия, например, связывание группы адениновых оснований друг с другом в выпуклости, [ 10 ] GNRA или тетрапетля , имеющая пару оснований гуанин-аденин. [ 9 ]

Важным структурным компонентом РНК, отличающим ее от ДНК, является наличие гидроксильной группы в 2'-положении сахара рибозы . Наличие этой функциональной группы приводит к тому, что спираль в основном принимает геометрию А-формы . [ 11 ] хотя в контексте одноцепочечных динуклеотидов РНК редко может также принимать B-форму, наиболее часто наблюдаемую в ДНК. [ 12 ] Геометрия А-образной формы обеспечивает очень глубокую и узкую основную канавку и неглубокую и широкую второстепенную канавку. [ 13 ] Вторым следствием присутствия 2'-гидроксильной группы является то, что в конформационно гибких участках молекулы РНК (т. е. не участвующих в образовании двойной спирали) она может химически атаковать соседнюю фосфодиэфирную связь, расщепляя основную цепь. [ 14 ]
РНК транскрибируется всего четырьмя основаниями (аденином, цитозином, гуанином и урацилом). [ 15 ] но эти основания и присоединенные сахара могут модифицироваться множеством способов по мере созревания РНК. Псевдоуридин (Ψ), в котором связь между урацилом и рибозой изменена со связи C–N на связь C–C, и риботимидин (T) обнаруживаются в различных местах (наиболее заметные из них находятся в петле TΨC тРНК). ). [ 16 ] Еще одним известным модифицированным основанием является гипоксантин , дезаминированное адениновое основание, нуклеозид которого называется инозином (I). Инозин играет ключевую роль в гипотезе колебания генетического кода . [ 17 ]
Существует более 100 других модифицированных нуклеозидов природного происхождения. [ 18 ] Наибольшее структурное разнообразие модификаций можно обнаружить у тРНК . [ 19 ] псевдоуридин и нуклеозиды с 2'-O-метилрибозой, часто присутствующие в рРНК. тогда как наиболее распространены [ 20 ] Конкретная роль многих из этих модификаций РНК до конца не изучена. Однако примечательно, что в рибосомальной РНК многие посттранскрипционные модификации происходят в высокофункциональных областях, таких как пептидилтрансферазный центр. [ 21 ] и интерфейс субблока, что означает, что они важны для нормального функционирования. [ 22 ]
Функциональная форма одноцепочечных молекул РНК, как и белков, часто требует определенной пространственной третичной структуры . Каркасом этой структуры служат вторичные структурные элементы, представляющие собой водородные связи внутри молекулы. Это приводит к появлению нескольких узнаваемых «доменов» вторичной структуры, таких как петли-шпильки , выпуклости и внутренние петли . [ 23 ] Чтобы создать, т. е. спроектировать, РНК для любой данной вторичной структуры, двух или трех оснований будет недостаточно, но достаточно четырех оснований. [ 24 ] Вероятно, поэтому природа «выбрала» четырехосновной алфавит: меньше четырех оснований не позволит создать все структуры, а более четырех оснований для этого не нужны. Поскольку РНК заряжена, ионы металлов, таких как Mg 2+ необходимы для стабилизации многих вторичных и третичных структур . [ 25 ]
Встречающийся в природе энантиомер РНК представляет собой D -РНК, состоящую из D -рибонуклеотидов. Все центры хиральности расположены в D -рибозе. Используя L -рибозу или, скорее , L -рибонуклеотиды, L можно синтезировать -РНК. L -РНК гораздо более устойчива к деградации под действием РНКазы . [ 26 ]
Как и в случае других структурированных биополимеров , таких как белки, можно определить топологию свернутой молекулы РНК. Это часто делается на основе расположения внутрицепочечных контактов внутри свернутой РНК, называемого топологией цепи .
Синтез
[ редактировать ]Синтез РНК обычно катализируется ферментом — РНК-полимеразой — с использованием ДНК в качестве матрицы. Этот процесс известен как транскрипция . Инициация транскрипции начинается со связывания фермента с последовательностью промотора в ДНК (обычно расположенной «выше» гена). Двойная спираль ДНК раскручивается за счет геликазной активности фермента. Затем фермент продвигается вдоль цепи матрицы в направлении от 3’ к 5’, синтезируя комплементарную молекулу РНК с удлинением, происходящим в направлении от 5’ к 3’. Последовательность ДНК также определяет, где произойдет прекращение синтеза РНК. [ 27 ]
Первичные транскрипты РНК часто модифицируются ферментами после транскрипции. Например, поли(А)-хвост и 5'-кэп добавляются к эукариотической пре-мРНК , а интроны удаляет сплайсосома .
Существует также ряд РНК-зависимых РНК-полимераз , которые используют РНК в качестве матрицы для синтеза новой цепи РНК. Например, ряд РНК-вирусов (таких как полиовирус) используют этот тип фермента для репликации своего генетического материала. [ 28 ] Кроме того, РНК-зависимая РНК-полимераза является частью пути РНК-интерференции у многих организмов. [ 29 ]
Типы РНК
[ редактировать ]Обзор
[ редактировать ]
Информационная РНК (мРНК) — это РНК, которая переносит информацию от ДНК к рибосоме , месту синтеза ( трансляции ) белка в клетке. мРНК является копией ДНК. Кодирующая последовательность мРНК определяет аминокислотную последовательность белка . образующегося [ 30 ] Однако многие РНК не кодируют белки (около 97% результатов транскрипции у эукариот не кодируют белки). [ 31 ] [ 32 ] [ 33 ] [ 34 ] ).
Эти так называемые некодирующие РНК («нкРНК») могут кодироваться собственными генами (генами РНК), но также могут происходить из интронов мРНК . [ 35 ] Наиболее яркими примерами некодирующих РНК являются транспортные РНК (тРНК) и рибосомальные РНК (рРНК), обе из которых участвуют в процессе трансляции. [ 5 ] Существуют также некодирующие РНК, участвующие в регуляции генов, процессинге РНК и других функциях. Некоторые РНК способны катализировать химические реакции, такие как разрезание и лигирование других молекул РНК. [ 36 ] и катализ образования пептидных связей в рибосоме ; [ 8 ] они известны как рибозимы .
В длину
[ редактировать ]По длине цепи РНК РНК включает малые РНК и длинные РНК. [ 37 ] Обычно малые РНК имеют длину менее 200 нт , а длинные РНК имеют длину более 200 нт . [ 38 ] Длинные РНК, также называемые большими РНК, в основном включают длинные некодирующие РНК (днРНК) и мРНК . Малые РНК в основном включают 5.8S рибосомальную РНК (рРНК), 5S рРНК , транспортную РНК (тРНК), микроРНК (миРНК), малую интерферирующую РНК (миРНК), малую ядрышковую РНК (мяРНК), Piwi-взаимодействующую РНК (пиРНК), тРНК- производная малая РНК (цРНК) [ 39 ] и малая РНК, полученная из рДНК (срРНК). [ 40 ] Есть определенные исключения, как в случае с 5S рРНК представителей рода Halococcus ( Archaea ), которые имеют вставку, увеличивающую таким образом ее размер. [ 41 ] [ 42 ] [ 43 ]
В переводе
[ редактировать ]Информационная РНК (мРНК) несет информацию о последовательности белка в рибосомы , фабрики по синтезу белка в клетке. Он закодирован так, что каждые три нуклеотида ( кодон ) соответствуют одной аминокислоте. В эукариотических клетках, как только мРНК-предшественник (пре-мРНК) транскрибируется с ДНК, она подвергается процессингу с образованием зрелой мРНК. При этом удаляются интроны — некодирующие участки пре-мРНК. Затем мРНК экспортируется из ядра в цитоплазму , где она связывается с рибосомами и транслируется в соответствующую белковую форму с помощью тРНК . В прокариотических клетках, не имеющих ядра и цитоплазмы, мРНК может связываться с рибосомами во время транскрипции с ДНК. Через определенное время сообщение распадается на составляющие его нуклеотиды с помощью рибонуклеаз . [ 30 ]
Транспортная РНК (тРНК) представляет собой небольшую цепочку РНК длиной около 80 нуклеотидов , которая переносит определенную аминокислоту на растущую полипептидную цепь в рибосомальном участке синтеза белка во время трансляции. Он имеет сайты для прикрепления аминокислот и антикодоновую область для распознавания кодонов , которая связывается со специфической последовательностью в цепи информационной РНК посредством водородных связей. [ 35 ]
Рибосомальная РНК (рРНК) является каталитическим компонентом рибосом. РРНК — это компонент рибосомы, обеспечивающий трансляцию. Рибосомы эукариот содержат четыре разные молекулы рРНК: 18S, 5,8S, 28S и 5S рРНК. Три молекулы рРНК синтезируются в ядрышке , а одна — в другом месте. В цитоплазме рибосомальная РНК и белок объединяются, образуя нуклеопротеин, называемый рибосомой. Рибосома связывает мРНК и осуществляет синтез белка. К одной мРНК в любое время могут быть присоединены несколько рибосом. [ 30 ] Почти вся РНК, обнаруженная в типичной эукариотической клетке, представляет собой рРНК.
Транспортная РНК (тмРНК) обнаружена во многих бактериях и пластидах . Он маркирует белки, кодируемые мРНК, у которых нет стоп-кодонов, для деградации и предотвращает остановку рибосомы. [ 44 ]
Регуляторная РНК
[ редактировать ]Самыми ранними известными регуляторами экспрессии генов были белки, известные как репрессоры и активаторы – регуляторы со специфическими короткими сайтами связывания в энхансерных областях рядом с регулируемыми генами. [ 45 ] Более поздние исследования показали, что РНК также регулируют гены. У эукариот существует несколько видов РНК-зависимых процессов, регулирующих экспрессию генов на различных этапах, таких как РНКи, репрессирующие гены посттранскрипционно , длинные некодирующие РНК, отключающие блоки хроматина эпигенетически , и РНК-энхансеры, индуцирующие повышенную экспрессию генов. [ 46 ] бактерии и археи Также было показано, что используют системы регуляторных РНК, такие как бактериальные малые РНК и CRISPR . [ 47 ] Файер и Мелло были удостоены Нобелевской премии по физиологии и медицине 2006 года за открытие микроРНК (миРНК), специфических коротких молекул РНК, которые могут образовывать пары оснований с мРНК. [ 48 ]
Интерференция РНК с помощью микроРНК
[ редактировать ]Уровни посттранскрипционной экспрессии многих генов можно контролировать с помощью РНК-интерференции , при которой микроРНК , специфические короткие молекулы РНК, соединяются с областями мРНК и направляют их на деградацию. [ 49 ] Этот основанный на антисмысловом процессе процесс включает в себя этапы, на которых сначала обрабатывается РНК, чтобы она могла соединить основания с областью целевой мРНК. Как только происходит спаривание оснований, другие белки направляют мРНК на разрушение нуклеазами . [ 46 ]
Длинные некодирующие РНК
[ редактировать ]Следующими были связаны с регуляцией Xist и другие длинные некодирующие РНК, связанные с инактивацией Х-хромосомы . Их роль, поначалу загадочная, как показали Джинни Т. Ли и другие, заключается в подавлении блоков хроматина посредством рекрутирования комплекса Polycomb , чтобы с них не могла транскрибироваться информационная РНК. [ 50 ] Дополнительные днРНК, в настоящее время определяемые как РНК, состоящие из более чем 200 пар оснований, которые, по-видимому, не обладают кодирующим потенциалом. [ 51 ] Было обнаружено, что они связаны с регуляцией стволовых клеток плюрипотентности и клеточного деления . [ 51 ]
Энхансерные РНК
[ редактировать ]Третья основная группа регуляторных РНК называется энхансерными РНК . [ 51 ] В настоящее время неясно, являются ли они уникальной категорией РНК различной длины или представляют собой отдельное подмножество днРНК. В любом случае они транскрибируются с энхансеров — известных регуляторных участков ДНК рядом с генами, которые они регулируют. [ 51 ] [ 52 ] Они усиливают транскрипцию гена(ов) под контролем энхансера, с которого они транскрибируются. [ 51 ] [ 53 ]
Регуляторная РНК у прокариот
[ редактировать ]Поначалу регуляторную РНК считали эукариотическим феноменом, что частично объясняло, почему у высших организмов наблюдалось гораздо больше транскрипции, чем предполагалось. Но как только исследователи начали искать возможные регуляторы РНК у бактерий, они появились и там, получившие название малых РНК (мРНК). [ 54 ] [ 47 ] В настоящее время повсеместная природа систем РНК-регуляции генов обсуждается как подтверждение теории мира РНК . [ 46 ] [ 55 ] Есть признаки того, что мРНК энтеробактерий участвуют в различных клеточных процессах и, по-видимому, играют значительную роль в реакциях на стресс, таких как мембранный стресс, стресс голодания, фосфосахарный стресс и повреждение ДНК. Кроме того, было высказано предположение, что мРНК стали играть важную роль в реакциях на стресс из-за их кинетических свойств, которые позволяют быстро реагировать и стабилизировать физиологическое состояние. [ 2 ] Бактериальные малые РНК обычно действуют посредством антисмыслового спаривания с мРНК, подавляя ее трансляцию, либо влияя на стабильность, либо на способность цис-связывания. [ 46 ] рибопереключатели Также были обнаружены . Они представляют собой цис-действующие регуляторные последовательности РНК, действующие аллостерически . Они меняют форму, когда связывают метаболиты , приобретая или теряя способность связывать хроматин для регулирования экспрессии генов. [ 56 ] [ 57 ]
У архей также есть системы регуляторных РНК. [ 58 ] Система CRISPR, недавно используемая для редактирования ДНК in situ , действует через регуляторные РНК в археях и бактериях, обеспечивая защиту от вирусных захватчиков. [ 46 ] [ 59 ]
В процессинге РНК
[ редактировать ]
Многие РНК участвуют в модификации других РНК. Интроны отделяются от с пре-мРНК помощью сплайсосом , которые содержат несколько малых ядерных РНК (мяРНК). [ 5 ] или интроны могут представлять собой рибозимы, которые сплайсируются сами по себе. [ 60 ] РНК также может быть изменена путем модификации ее нуклеотидов на нуклеотиды, отличные A , C , G и U. от У эукариот модификации нуклеотидов РНК обычно направляются небольшими ядрышковыми РНК (мяРНК; 60–300 нт), [ 35 ] Встречается в ядрышках и кахаальных тельцах . мякРНК связываются с ферментами и направляют их к месту на РНК путем спаривания оснований с этой РНК. Эти ферменты затем выполняют модификацию нуклеотидов. рРНК и тРНК сильно модифицируются, но мяРНК и мРНК также могут быть объектом модификации оснований. [ 61 ] [ 62 ] РНК также может быть метилирована. [ 63 ] [ 64 ]
РНК-геномы
[ редактировать ]Как и ДНК, РНК может нести генетическую информацию. РНК-вирусы имеют геномы, состоящие из РНК, которая кодирует ряд белков. Вирусный геном реплицируется некоторыми из этих белков, в то время как другие белки защищают геном, когда вирусная частица перемещается в новую клетку-хозяина. Вироиды — еще одна группа патогенов, но они состоят только из РНК, не кодируют никаких белков и реплицируются полимеразой клетки растения-хозяина. [ 65 ]
В обратной транскрипции
[ редактировать ]Вирусы с обратной транскрипцией реплицируют свои геномы путем обратной транскрипции копий ДНК из их РНК; эти копии ДНК затем транскрибируются в новую РНК. Ретротранспозоны также распространяются путем копирования ДНК и РНК друг у друга. [ 66 ] а теломераза содержит РНК, которая используется в качестве матрицы для построения концов эукариотических хромосом . [ 67 ]
Двухцепочечная РНК
[ редактировать ]
Двухцепочечная РНК (дсРНК) — это РНК с двумя комплементарными цепями, подобная ДНК, встречающейся во всех клетках, но с заменой тимина на урацил и добавлением одного атома кислорода. дсРНК образует генетический материал некоторых вирусов ( вирусов с двухцепочечной РНК ). Двухцепочечная РНК, такая как вирусная РНК или миРНК , может вызывать интерференцию РНК у эукариот , а также интерфероновую реакцию у позвоночных . [ 68 ] [ 69 ] [ 70 ] [ 71 ] У эукариот двухцепочечная РНК (дсРНК) играет роль в активации врожденной иммунной системы против вирусных инфекций. [ 72 ]
Круговая РНК
[ редактировать ]В конце 1970-х годов было показано, что существует одноцепочечная ковалентно-замкнутая, то есть кольцевая форма РНК, экспрессируемая во всем животном и растительном царстве (см. circRNA ). [ 73 ] Считается, что циркРНК возникают в результате реакции «обратного сплайсинга», когда сплайсосома присоединяется к расположенному выше 3'-акцептору к нижестоящему 5'-донному сайту сплайсинга. До сих пор функция circRNAs в значительной степени неизвестна, хотя в нескольких примерах была продемонстрирована спонжирующая активность микроРНК.
Ключевые открытия в биологии РНК
[ редактировать ]
Исследования РНК привели ко многим важным биологическим открытиям и многочисленным Нобелевским премиям . Нуклеиновые кислоты были открыты в 1868 году Фридрихом Мишером , который назвал этот материал «нуклеином», поскольку он был обнаружен в ядре . [ 74 ] Позднее было обнаружено, что прокариотические клетки, не имеющие ядра, также содержат нуклеиновые кислоты. Роль РНК в синтезе белка заподозрили еще в 1939 году. [ 75 ] Северо Очоа получил Нобелевскую премию по медицине 1959 года (совместно с Артуром Корнбергом ) после того, как открыл фермент, способный синтезировать РНК в лаборатории. [ 76 ] Однако позже было показано, что фермент, открытый Очоа ( полинуклеотидфосфорилаза ), отвечает за деградацию РНК, а не за синтез РНК. В 1956 году Алекс Рич и Дэвид Дэвис гибридизовали две отдельные цепи РНК, чтобы сформировать первый кристалл РНК, структуру которого можно было определить с помощью рентгеновской кристаллографии. [ 77 ]
Последовательность из 77 нуклеотидов дрожжевой тРНК была обнаружена Робертом Холли в 1965 году. [ 78 ] получение Холли Нобелевской премии по медицине 1968 года (совместно с Хар Гобиндом Хораной и Маршаллом Ниренбергом ).
В начале 1970-х годов были открыты ретровирусы и обратная транскриптаза , что впервые показало, что ферменты могут копировать РНК в ДНК (противоположно обычному пути передачи генетической информации). За эту работу Дэвид Балтимор , Ренато Дульбекко и Говард Темин были удостоены Нобелевской премии в 1975 году. В 1976 году Уолтер Фирс и его команда определили первую полную нуклеотидную последовательность генома РНК-вируса, бактериофага MS2 . [ 79 ]
В 1977 году интроны и сплайсинг РНК были обнаружены как в вирусах млекопитающих, так и в клеточных генах, что привело к вручению Нобелевской премии 1993 года Филипу Шарпу и Ричарду Робертсу . Каталитические молекулы РНК ( рибозимы ) были открыты в начале 1980-х годов, что привело к Нобелевской премии 1989 года Томасу Чеху и Сидни Альтману . было обнаружено В 1990 году у петунии , что введенные гены могут заглушить аналогичные собственные гены растения, что, как теперь известно, является результатом интерференции РНК . [ 80 ] [ 81 ]
Примерно в то же время было обнаружено, что РНК длиной 22 нуклеотида, теперь называемые микроРНК , играют роль в развитии C. elegans . [ 82 ] Исследования РНК-интерференции принесли Нобелевскую премию Эндрю Файру и Крейгу Мелло в 2006 году, а также еще одну Нобелевскую премию за исследования транскрипции РНК Роджеру Корнбергу в том же году. Открытие регуляторных РНК генов привело к попыткам разработать лекарства из РНК, такие как миРНК , для подавления генов. [ 83 ] Помимо Нобелевской премии за исследования РНК, в 2009 году она была присуждена за выяснение атомной структуры рибосомы Венки Рамакришнану , Томасу А. Стейцу и Аде Йонат . В 2023 году Нобелевская премия по физиологии и медицине была присуждена Каталин Карико и Дрю Вайсману за открытия, касающиеся модифицированных нуклеозидов , которые позволили разработать эффективные мРНК-вакцины против COVID-19. [ 84 ] [ 85 ] [ 86 ]
Актуальность для пребиотической химии и абиогенеза.
[ редактировать ]В 1968 году Карл Вёзе выдвинул гипотезу о том, что РНК может играть роль катализатора, и предположил, что самые ранние формы жизни (самовоспроизводящиеся молекулы) могли полагаться на РНК как для переноса генетической информации, так и для катализа биохимических реакций – мир РНК . [ 87 ] [ 88 ] В мае 2022 года ученые обнаружили, что РНК может спонтанно образовываться на пребиотическом базальтовом лавовом стекле , которое, предположительно, было в изобилии на ранней Земле . [ 89 ] [ 90 ]
Сообщалось, что в марте 2015 года ДНК и РНК нуклеиновые основания , включая урацил , цитозин и тимин , были сформированы в лаборатории в условиях космического пространства с использованием стартовых химических веществ, таких как пиримидин , органическое соединение, обычно встречающееся в метеоритах . Пиримидин , как и полициклические ароматические углеводороды (ПАУ), является одним из наиболее богатых углеродом соединений, обнаруженных во Вселенной , и, возможно, образовался в красных гигантах или в межзвездных пылевых и газовых облаках. [ 91 ] В июле 2022 года астрономы сообщили об огромном количестве пребиотических молекул , включая возможные предшественники РНК, в галактическом центре Галактики Млечный Путь . [ 92 ] [ 93 ]
Медицинские приложения
[ редактировать ]РНК, первоначально считавшаяся непригодной для терапии из-за ее короткого периода полураспада, стала полезной благодаря достижениям в области стабилизации. Терапевтическое применение возникает, когда РНК сворачивается в сложные конформации и связывает белки, нуклеиновые кислоты и небольшие молекулы с образованием каталитических центров. [ 94 ] Считается, что вакцины на основе РНК легче производить, чем традиционные вакцины, полученные из убитых или измененных патогенов, поскольку на выращивание и изучение патогена и определение того, какие молекулярные части следует извлечь, инактивировать и использовать в вакцине, могут потребоваться месяцы или годы. Небольшие молекулы с традиционными терапевтическими свойствами могут воздействовать на структуры РНК и ДНК, тем самым леча новые заболевания. Однако исследований малых молекул, нацеленных на РНК, и одобренных лекарств для лечения заболеваний человека мало. Рибавирин, бранаплам и аталурен в настоящее время являются доступными лекарствами, которые стабилизируют структуры двухцепочечной РНК и контролируют сплайсинг при различных заболеваниях. [ 95 ] [ 96 ]
Кодирующие белок мРНК стали новыми терапевтическими кандидатами, причем замена РНК особенно полезна для кратковременной, но бурной экспрессии белка. [ 97 ] Транскрибируемые in vitro мРНК (IVT-мРНК) использовались для доставки белков для регенерации костей, плюрипотентности и функции сердца на животных моделях. [ 98 ] [ 99 ] [ 100 ] [ 101 ] [ 102 ] SiRNA, короткие молекулы РНК, играют решающую роль во врожденной защите от вирусов и структуре хроматина. Их можно искусственно вводить, чтобы заставить замолчать определенные гены, что делает их ценными для изучения функций генов, проверки терапевтических целей и разработки лекарств. [ 97 ]
мРНК-вакцины стали важным новым классом вакцин, в которых мРНК используется для производства белков, вызывающих иммунный ответ. Их первым успешным крупномасштабным применением стали вакцины против COVID-19 во время пандемии COVID-19 .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Копли С.Д., Смит Э., Марковиц Х.Дж. (декабрь 2007 г.). «Происхождение мира РНК: коэволюция генов и метаболизма». Биоорганическая химия . 35 (6): 430–443. дои : 10.1016/j.bioorg.2007.08.001 . ПМИД 17897696 .
Предположение о том, что жизнь на Земле возникла из мира РНК, широко распространено.
- ^ Перейти обратно: а б «РНК: универсальная молекула» . Университет Юты . 2015.
- ^ «Нуклеотиды и нуклеиновые кислоты» (PDF) . Калифорнийский университет, Лос-Анджелес . Архивировано из оригинала (PDF) 23 сентября 2015 г. Проверено 26 августа 2015 г.
- ^ Шукла Р.Н. (2014). Анализ хромосом . Агротех Пресс. ISBN 978-93-84568-17-7 . [ постоянная мертвая ссылка ]
- ^ Перейти обратно: а б с Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). Биохимия (5-е изд.). WH Фриман и компания. стр. 118–19, 781–808. ISBN 978-0-7167-4684-3 . OCLC 179705944 .
- ^ Тиноко I, Бустаманте С (октябрь 1999 г.). «Как складывается РНК». Журнал молекулярной биологии . 293 (2): 271–81. дои : 10.1006/jmbi.1999.3001 . ПМИД 10550208 .
- ^ Хиггс П.Г. (август 2000 г.). «Вторичная структура РНК: физические и вычислительные аспекты». Ежеквартальные обзоры биофизики . 33 (3): 199–253. дои : 10.1017/S0033583500003620 . ПМИД 11191843 . S2CID 37230785 .
- ^ Перейти обратно: а б Ниссен П., Хансен Дж., Бан Н., Мур П.Б., Стейц Т.А. (август 2000 г.). «Структурные основы активности рибосом при синтезе пептидных связей». Наука . 289 (5481): 920–30. Бибкод : 2000Sci...289..920N . дои : 10.1126/science.289.5481.920 . ПМИД 10937990 .
- ^ Перейти обратно: а б Ли Дж. К., Гутелл Р. Р. (декабрь 2004 г.). «Разнообразие конформаций пар оснований и их появление в структуре рРНК и структурных мотивах РНК». Журнал молекулярной биологии . 344 (5): 1225–49. дои : 10.1016/j.jmb.2004.09.072 . ПМИД 15561141 .
- ^ Барчишевски Дж., Фредерик Б., Кларк С. (1999). Биохимия и биотехнология РНК . Спрингер. стр. 73–87. ISBN 978-0-7923-5862-6 . OCLC 52403776 .
- ^ Салазар М., Федоров О.Ю., Миллер Дж.М., Рибейро Н.С., Рид Б.Р. (апрель 1993 г.). «Нидь ДНК в гибридных дуплексах ДНК.РНК не является ни B-формой, ни A-формой в растворе». Биохимия . 32 (16): 4207–15. дои : 10.1021/bi00067a007 . ПМИД 7682844 .
- ^ Седова А, Банавали Н.К. (февраль 2016 г.). «РНК приближается к B-форме в многоцепочечном динуклеотидном контексте». Биополимеры . 105 (2): 65–82. дои : 10.1002/bip.22750 . ПМИД 26443416 . S2CID 35949700 .
- ^ Герман Т., Патель DJ (март 2000 г.). «Выпуклости РНК как архитектурные и узнаваемые мотивы» . Структура . 8 (3): Р47–54. дои : 10.1016/S0969-2126(00)00110-6 . ПМИД 10745015 .
- ^ Миккола С., Стенман Э., Нурми К., Юсефи-Салакде Э., Стрёмберг Р., Лённберг Х. (1999). «Механизм расщепления фосфодиэфирных связей РНК, стимулируемого ионом металла, включает общий кислотный катализ акво-ионом металла при уходе уходящей группы». Журнал Химического общества, Perkin Transactions 2 (8): 1619–26. дои : 10.1039/a903691a .
- ^ Янковский Ю.А., Полак Ю.М. (1996). Клинический генный анализ и манипуляции: инструменты, методы и устранение неполадок . Издательство Кембриджского университета. п. 14 . ISBN 978-0-521-47896-0 . OCLC 33838261 .
- ^ Ю Кью, компакт-диск Morrow (май 2001 г.). «Идентификация критических элементов в акцепторном стволе тРНК и петле T(Psi)C, необходимых для инфекционности вируса иммунодефицита человека типа 1» . Журнал вирусологии . 75 (10): 4902–6. doi : 10.1128/JVI.75.10.4902-4906.2001 . ПМЦ 114245 . ПМИД 11312362 .
- ^ Эллиотт М.С., Тревин Р.В. (февраль 1984 г.). «Биосинтез инозина в транспортной РНК путем ферментативной вставки гипоксантина» . Журнал биологической химии . 259 (4): 2407–10. дои : 10.1016/S0021-9258(17)43367-9 . ПМИД 6365911 .
- ^ Кантара В.А., Крейн П.Ф., Розенски Дж., Макклоски Дж.А., Харрис К.А., Чжан Х, Вендейкс Ф.А., Фабрис Д., Агрис П.Ф. (январь 2011 г.). «База данных модификации РНК, RNAMDB: обновление 2011 г.» . Исследования нуклеиновых кислот . 39 (Проблема с базой данных): D195-201. дои : 10.1093/нар/gkq1028 . ПМК 3013656 . ПМИД 21071406 .
- ^ Сёлль Д., РаджБхандари У (1995). ТРНК: строение, биосинтез и функции . АСМ Пресс. п. 165. ИСБН 978-1-55581-073-3 . OCLC 183036381 .
- ^ Поцелуй Т (июль 2001 г.). «Посттранскрипционная модификация клеточных РНК под контролем малых ядрышковых РНК» . Журнал ЭМБО . 20 (14): 3617–22. дои : 10.1093/emboj/20.14.3617 . ПМК 125535 . ПМИД 11447102 .
- ^ Тирумалай М.Р., Ривас М., Тран К., Фокс Дж.Э. (ноябрь 2021 г.). «Центр пептидилтрансферазы: окно в прошлое» . Микробиол Мол Биол Rev. 85 (4): e0010421. дои : 10.1128/MMBR.00104-21 . ПМЦ 8579967 . ПМИД 34756086 .
- ^ Кинг Т.Х., Лю Б., Маккалли Р.Р., Фурнье М.Дж. (февраль 2003 г.). «Структура и активность рибосом изменяются в клетках, лишенных мяРНП, которые образуют псевдоуридины в пептидилтрансферазном центре» . Молекулярная клетка . 11 (2): 425–35. дои : 10.1016/S1097-2765(03)00040-6 . ПМИД 12620230 .
- ^ Мэтьюз Д.Х., Дисней, доктор медицинских наук, Чайлдс Дж.Л., Шредер С.Дж., Цукер М., Тернер Д.Х. (май 2004 г.). «Включение ограничений химической модификации в алгоритм динамического программирования для прогнозирования вторичной структуры РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (19): 7287–92. Бибкод : 2004PNAS..101.7287M . дои : 10.1073/pnas.0401799101 . ПМК 409911 . ПМИД 15123812 .
- ^ Бургхардт Б., Хартманн А.К. (февраль 2007 г.). «Дизайн вторичной структуры РНК» . Физический обзор E . 75 (2): 021920. arXiv : физика/0609135 . Бибкод : 2007PhRvE..75b1920B . дои : 10.1103/PhysRevE.75.021920 . ПМИД 17358380 . S2CID 17574854 .
- ^ Тан ZJ, Чен SJ (июль 2008 г.). «Солевая зависимость стабильности шпильки нуклеиновой кислоты» . Биофизический журнал . 95 (2): 738–52. Бибкод : 2008BpJ....95..738T . дои : 10.1529/biophysj.108.131524 . ПМЦ 2440479 . ПМИД 18424500 .
- ^ Фатер А., Клуссманн С. (январь 2015 г.). «Превращение зеркальных олигонуклеотидов в лекарства: эволюция терапии Spiegelmer (®)» . Открытие наркотиков сегодня . 20 (1): 147–55. дои : 10.1016/j.drudis.2014.09.004 . ПМИД 25236655 .
- ^ Нудлер Э., Готтесман М.Е. (август 2002 г.). «Терминация и антитерминация транскрипции в E. coli». Гены в клетки . 7 (8): 755–68. дои : 10.1046/j.1365-2443.2002.00563.x . ПМИД 12167155 . S2CID 23191624 .
- ^ Хансен Дж.Л., Лонг А.М., Шульц С.К. (август 1997 г.). «Структура РНК-зависимой РНК-полимеразы полиовируса» . Структура . 5 (8): 1109–22. дои : 10.1016/S0969-2126(97)00261-X . ПМИД 9309225 .
- ^ Алквист П. (май 2002 г.). «РНК-зависимые РНК-полимеразы, вирусы и подавление РНК». Наука . 296 (5571): 1270–73. Бибкод : 2002Sci...296.1270A . дои : 10.1126/science.1069132 . ПМИД 12016304 . S2CID 42526536 .
- ^ Перейти обратно: а б с Купер Г.К., Хаусман Р.Э. (2004). Клетка: молекулярный подход (3-е изд.). Синауэр. стр. 261–76, 297, 339–44. ISBN 978-0-87893-214-6 . OCLC 174924833 .
- ^ Мэттик Дж.С., Гаген М.Дж. (сентябрь 2001 г.). «Эволюция управляемых многозадачных генных сетей: роль интронов и других некодирующих РНК в развитии сложных организмов» . Молекулярная биология и эволюция . 18 (9): 1611–30. doi : 10.1093/oxfordjournals.molbev.a003951 . ПМИД 11504843 .
- ^ Мэттик Дж.С. (ноябрь 2001 г.). «Некодирующие РНК: архитекторы эукариотической сложности» . Отчеты ЭМБО . 2 (11): 986–91. doi : 10.1093/embo-reports/kve230 . ПМЦ 1084129 . ПМИД 11713189 .
- ^ Мэттик Дж.С. (октябрь 2003 г.). «Бросая вызов догме: скрытый слой некодирующих белок РНК в сложных организмах» (PDF) . Биоэссе . 25 (10): 930–39. CiteSeerX 10.1.1.476.7561 . дои : 10.1002/bies.10332 . ПМИД 14505360 . Архивировано из оригинала (PDF) 6 марта 2009 г.
- ^ Мэттик Дж.С. (октябрь 2004 г.). «Скрытая генетическая программа сложных организмов». Научный американец . 291 (4): 60–67. Бибкод : 2004SciAm.291d..60M . doi : 10.1038/scientificamerican1004-60 . ПМИД 15487671 . [ мертвая ссылка ]
- ^ Перейти обратно: а б с Вирта В (2006). Анализ транскриптома – методы и приложения . Стокгольм: Школа биотехнологии Королевского технологического института. ISBN 978-91-7178-436-0 . OCLC 185406288 .
- ^ Росси Джей-Джей (июль 2004 г.). «Рибозимная диагностика достигает совершеннолетия» . Химия и биология . 11 (7): 894–95. doi : 10.1016/j.chembiol.2004.07.002 . ПМИД 15271347 .
- ^ Шторц Г. (май 2002 г.). «Расширяющаяся вселенная некодирующих РНК». Наука . 296 (5571): 1260–63. Бибкод : 2002Sci...296.1260S . дои : 10.1126/science.1072249 . ПМИД 12016301 . S2CID 35295924 .
- ^ Фатика А, Боццони I (январь 2014 г.). «Длинные некодирующие РНК: новые игроки в дифференцировке и развитии клеток» . Обзоры природы Генетика . 15 (1): 7–21. дои : 10.1038/nrg3606 . ПМИД 24296535 . S2CID 12295847 . [ постоянная мертвая ссылка ]
- ^ Чен Ц, Ян М, Цао Z, Ли X, Чжан Ю, Ши Дж и др. (январь 2016 г.). «ЦРНК спермы способствуют наследованию приобретенного метаболического нарушения из поколения в поколение» (PDF) . Наука . 351 (6271): 397–400. Бибкод : 2016Sci...351..397C . doi : 10.1126/science.aad7977 . ПМИД 26721680 . S2CID 21738301 .
- ^ Вэй Х, Чжоу Б, Чжан Ф, Ту Ю, Ху Ю, Чжан Б, Чжай Ц (2013). «Профилирование и идентификация малых РНК, полученных из рДНК, и их потенциальных биологических функций» . ПЛОС ОДИН . 8 (2): e56842. Бибкод : 2013PLoSO...856842W . дои : 10.1371/journal.pone.0056842 . ПМК 3572043 . ПМИД 23418607 .
- ^ Люрсен К.Р., Николсон Д.Э., Юбэнкс, округ Колумбия, Фокс Дж.Э. (1981). «Архебактериальная 5S рРНК содержит длинную вставочную последовательность». Природа . 293 (5835): 755–756. Бибкод : 1981Natur.293..755L . дои : 10.1038/293755a0 . ПМИД 6169998 . S2CID 4341755 .
- ^ Стэн-Лоттер Х., МакДженити Т.Дж., Легат А., Деннер Э.Б., Глейзер К., Стеттер К.О., Ваннер Г. (1999). «Очень похожие штаммы Halococcus salifodinae обнаружены в географически разделенных пермо-триасовых соляных отложениях» . Микробиология . 145 (Часть 12): 3565–3574. дои : 10.1099/00221287-145-12-3565 . ПМИД 10627054 .
- ^ Тирумалай М.Р., Кельбер Дж.Т., Пак Д.Р., Тран К., Fox GE (август 2020 г.). «Криоэлектронная микроскопия. Визуализация большой вставки в 5S рибосомальной РНК чрезвычайно галофильной археи Halococcus morrhuae » . Открытая биография FEBS . 10 (10): 1938–1946. дои : 10.1002/2211-5463.12962 . ПМЦ 7530397 . ПМИД 32865340 .
- ^ Гено де Новоа П., Уильямс К.П. (январь 2004 г.). «Сайт тмРНК: редуктивная эволюция тмРНК в пластидах и других эндосимбионтах» . Исследования нуклеиновых кислот . 32 (Проблема с базой данных): D104–08. дои : 10.1093/nar/gkh102 . ПМК 308836 . ПМИД 14681369 .
- ^ Джейкоб Ф., Моно Дж. (1961). «Генетические механизмы регуляции синтеза белков». Журнал молекулярной биологии . 3 (3): 318–56. дои : 10.1016/s0022-2836(61)80072-7 . ПМИД 13718526 . S2CID 19804795 .
- ^ Перейти обратно: а б с д и Моррис К., Мэттик Дж. (2014). «Рост регуляторной РНК» . Обзоры природы Генетика . 15 (6): 423–37. дои : 10.1038/nrg3722 . ПМЦ 4314111 . ПМИД 24776770 .
- ^ Перейти обратно: а б Готтесман С (2005). «Микро для микробов: некодирующие регуляторные РНК у бактерий». Тенденции в генетике . 21 (7): 399–404. дои : 10.1016/j.tig.2005.05.008 . ПМИД 15913835 .
- ^ «Нобелевская премия по физиологии и медицине 2006 г.». Нобелевская премия.org. Nobel Media AB 2014. Интернет. 6 августа 2018 г. http://www.nobelprize.org/nobel_prizes/medicine/laureates/2006.
- ^ Огонь и др. (1998). «Мощное и специфическое генетическое вмешательство двухцепочечной РНК в Ceanorhabditis elegans» . Природа . 391 (6669): 806–11. Бибкод : 1998Natur.391..806F . дои : 10.1038/35888 . ПМИД 9486653 . S2CID 4355692 .
- ^ Чжао Дж., Сунь Б.К., Эрвин Дж.А., Сонг Дж.Дж., Ли Дж.Т. (2008). «Белки Polycomb, нацеленные с помощью короткой повторной РНК на Х-хромосому мыши» . Наука . 322 (5902): 750–56. Бибкод : 2008Sci...322..750Z . дои : 10.1126/science.1163045 . ПМЦ 2748911 . ПМИД 18974356 .
- ^ Перейти обратно: а б с д и Ринн Дж.Л., Чанг ХИ (2012). «Регуляция генома длинными некодирующими РНК» . Анну. Преподобный Биохим . 81 : 1–25. doi : 10.1146/annurev-biochem-051410-092902 . ПМЦ 3858397 . ПМИД 22663078 .
- ^ Тафт Р.Дж., Каплан К.Д., Саймонс С., Мэттик Дж.С. (2009). «Эволюция, биогенез и функция РНК, связанных с промотором» . Клеточный цикл . 8 (15): 2332–38. дои : 10.4161/cc.8.15.9154 . ПМИД 19597344 .
- ^ Ором У.А., Дерриен Т., Беринджер М., Гумиредди К., Гардини А. и др. (2010). « Длинные некодирующие РНК с энхансероподобной функцией в клетках человека» . Клетка . 143 (1): 46–58. дои : 10.1016/j.cell.2010.09.001 . ПМК 4108080 . ПМИД 20887892 .
- ^ Э.Г.Г.Вагнер, П.Ромби. (2015). «Маленькие РНК у бактерий и архей: кто они, что они делают и как они это делают». Достижения генетики (т. 90, стр. 133–208).
- ^ Дж. В. Нельсон, RR Breaker (2017) «Потерянный язык мира РНК». наук. Сигнал . 10 , eaam8812 1–11.
- ^ Винклеф WC (2005). «Рибопереключатели и роль некодирующих РНК в бактериальном метаболическом контроле». Курс. Мнение. хим. Биол . 9 (6): 594–602. дои : 10.1016/j.cbpa.2005.09.016 . ПМИД 16226486 .
- ^ Такер Би Джей, Брейкер Р.Р. (2005). «Рибопереключатели как универсальные элементы контроля генов». Курс. Мнение. Структура. Биол . 15 (3): 342–48. дои : 10.1016/j.sbi.2005.05.003 . ПМИД 15919195 .
- ^ Мохика Ф.Дж., Диес-Вильясенор К., Сория Э., Хуэс Дж. (2000). " " "Биологическое значение семейства регулярно расположенных повторов в геномах архей, бактерий и митохондрий" . Мол. Микробиол . 36 (1): 244–46. дои : 10.1046/j.1365-2958.2000.01838.x . ПМИД 10760181 . S2CID 22216574 .
- ^ Браунс С., Джор М.М., Лундгрен М., Вестра Е., Слейкхуис Р., Снейдерс А., Дикман М., Макарова К., Кунин Е., Der Oost JV (2008). «Маленькие РНК CRISPR обеспечивают противовирусную защиту прокариот» . Наука . 321 (5891): 960–64. Бибкод : 2008Sci...321..960B . дои : 10.1126/science.1159689 . ПМЦ 5898235 . ПМИД 18703739 .
- ^ Стейц Т.А., Стейц Дж.А. (июль 1993 г.). «Общий механизм каталитической РНК с двумя ионами металлов» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (14): 6498–502. Бибкод : 1993PNAS...90.6498S . дои : 10.1073/pnas.90.14.6498 . ПМК 46959 . ПМИД 8341661 .
- ^ Се Дж., Чжан М., Чжоу Т., Хуа Х., Тан Л., Ву В. (январь 2007 г.). «Sno/scaRNAbase: тщательно подобранная база данных малых ядрышковых РНК и РНК, специфичных для тела Кахаля» . Исследования нуклеиновых кислот . 35 (Проблема с базой данных): D183–87. дои : 10.1093/нар/gkl873 . ПМК 1669756 . ПМИД 17099227 .
- ^ Омер А.Д., Зиеше С., Декейтер В.А., Фурнье М.Дж., Деннис П.П. (май 2003 г.). «Машины, модифицирующие РНК у архей». Молекулярная микробиология . 48 (3): 617–29. дои : 10.1046/j.1365-2958.2003.03483.x . ПМИД 12694609 . S2CID 20326977 .
- ^ Кавайе Дж., Николозо М., Bachellerie JP (октябрь 1996 г.). «Направленное рибозное метилирование РНК in vivo, управляемое специальными антисмысловыми РНК-гидами» . Природа . 383 (6602): 732–35. Бибкод : 1996Natur.383..732C . дои : 10.1038/383732a0 . ПМИД 8878486 . S2CID 4334683 .
- ^ Кисс-Ласло З., Генри Ю., Бачеллери ЖП, Кайзерг-Феррер М., Кисс Т. (июнь 1996 г.). «Сайт-специфическое метилирование рибозы прерибосомальной РНК: новая функция для малых ядрышковых РНК» . Клетка . 85 (7): 1077–88. дои : 10.1016/S0092-8674(00)81308-2 . ПМИД 8674114 . S2CID 10418885 .
- ^ Дарос Х.А., Елена С.Ф., Флорес Р. (июнь 2006 г.). «Вироиды: нить Ариадны в лабиринте РНК» . Отчеты ЭМБО . 7 (6): 593–98. дои : 10.1038/sj.embor.7400706 . ПМЦ 1479586 . ПМИД 16741503 .
- ^ Календар Р., Висент С.М., Пелег О., Анамтават-Джонссон К., Большой А., Шульман А.Х. (март 2004 г.). «Большие производные ретротранспозонов: многочисленные, консервативные, но неавтономные ретроэлементы ячменя и родственных геномов» . Генетика . 166 (3): 1437–50. дои : 10.1534/генетика.166.3.1437 . ПМК 1470764 . ПМИД 15082561 .
- ^ Подлевский Дж.Д., Блей С.Дж., Омана Р.В., Ци Икс, Чен Дж.Дж. (январь 2008 г.). «База данных теломеразы» . Исследования нуклеиновых кислот . 36 (Проблема с базой данных): D339–43. дои : 10.1093/нар/gkm700 . ПМК 2238860 . ПМИД 18073191 .
- ^ Блевинс Т., Раджешваран Р., Шивапрасад П.В., Бекназарианц Д., Си-Аммур А., Парк Х.С., Васкес Ф., Робертсон Д., Майнс Ф., Хон Т., Пуггин М.М. (2006). «Четыре растительных Dicer опосредуют биогенез вирусных малых РНК и подавление ДНК-вируса, индуцированное» . Исследования нуклеиновых кислот . 34 (21): 6233–46. дои : 10.1093/nar/gkl886 . ПМК 1669714 . ПМИД 17090584 .
- ^ Яна С., Чакраборти С., Нанди С., Деб Дж.К. (ноябрь 2004 г.). «РНК-интерференция: потенциальные терапевтические мишени». Прикладная микробиология и биотехнология . 65 (6): 649–57. дои : 10.1007/s00253-004-1732-1 . ПМИД 15372214 . S2CID 20963666 .
- ^ Вирол, Дж. (май 2006 г.). «Двухцепочечная РНК продуцируется РНК-вирусами с положительной цепью и ДНК-вирусами, но не в обнаруживаемых количествах РНК-вирусами с отрицательной цепью» . Журнал вирусологии . 80 (10): 5059–5064. doi : 10.1128/JVI.80.10.5059-5064.2006 . ПМК 1472073 . ПМИД 16641297 .
- ^ Шульц У., Касперс Б., Стаэхели П. (май 2004 г.). «Система интерферона позвоночных немлекопитающих». Развивающая и сравнительная иммунология . 28 (5): 499–508. дои : 10.1016/j.dci.2003.09.009 . ПМИД 15062646 .
- ^ Уайтхед К.А., Дальман Дж.Э., Лангер Р.С., Андерсон Д.Г. (2011). «Замалчивание или стимуляция? Доставка миРНК и иммунная система». Ежегодный обзор химической и биомолекулярной инженерии . 2 : 77–96. doi : 10.1146/annurev-chembioeng-061010-114133 . ПМИД 22432611 .
- ^ Сюй М.Т., Кока-Прадос М. (июль 1979 г.). «Электронно-микроскопические доказательства кольцевой формы РНК в цитоплазме эукариотических клеток». Природа . 280 (5720): 339–40. Бибкод : 1979Natur.280..339H . дои : 10.1038/280339a0 . ПМИД 460409 . S2CID 19968869 .
- ^ Дам Р. (февраль 2005 г.). «Фридрих Мишер и открытие ДНК». Биология развития . 278 (2): 274–88. дои : 10.1016/j.ydbio.2004.11.028 . ПМИД 15680349 .
- ^ Касперссон Т., Шульц Дж (1939). «Пентозные нуклеотиды в цитоплазме растущих тканей». Природа . 143 (3623): 602–03. Бибкод : 1939Natur.143..602C . дои : 10.1038/143602c0 . S2CID 4140563 .
- ^ Очоа С (1959). «Ферментативный синтез рибонуклеиновой кислоты» (PDF) . Нобелевская лекция .
- ^ Рич А., Дэвис Д. (1956). «Новая двухцепочечная спиральная структура: полиадениловая кислота и полиуридиловая кислота». Журнал Американского химического общества . 78 (14): 3548–49. дои : 10.1021/ja01595a086 .
- ^ Холли Р.В. и др. (март 1965 г.). «Структура рибонуклеиновой кислоты». Наука . 147 (3664): 1462–65. Бибкод : 1965Sci...147.1462H . дои : 10.1126/science.147.3664.1462 . ПМИД 14263761 . S2CID 40989800 .
- ^ Фирс В. и др. (апрель 1976 г.). «Полная нуклеотидная последовательность РНК бактериофага MS2: первичная и вторичная структура гена репликазы». Природа . 260 (5551): 500–07. Бибкод : 1976Natur.260..500F . дои : 10.1038/260500a0 . ПМИД 1264203 . S2CID 4289674 .
- ^ Наполи К, Лемье К, Йоргенсен Р (апрель 1990 г.). «Введение химерного гена халконсинтазы в петунию приводит к обратимой совместной супрессии гомологичных генов в транс» . Растительная клетка . 2 (4): 279–89. дои : 10.1105/tpc.2.4.279 . ПМК 159885 . ПМИД 12354959 .
- ^ Дафни-Елин М., Чунг С.М., Франкман Э.Л., Цфира Т. (декабрь 2007 г.). «Векторы интерференции РНК pSAT: модульная серия для подавления множественных генов у растений» . Физиология растений . 145 (4): 1272–81. дои : 10.1104/стр.107.106062 . ПМК 2151715 . ПМИД 17766396 .
- ^ Рувкун Г. (октябрь 2001 г.). «Молекулярная биология. Взгляд на крошечный мир РНК». Наука . 294 (5543): 797–99. дои : 10.1126/science.1066315 . ПМИД 11679654 . S2CID 83506718 .
- ^ Фишу Ю, Ферек С (декабрь 2006 г.). «Потенциал олигонуклеотидов для терапевтического применения». Тенденции в биотехнологии . 24 (12): 563–70. дои : 10.1016/j.tibtech.2006.10.003 . ПМИД 17045686 .
- ^ «Нобелевская премия по физиологии и медицине 2023 года» . NobelPrize.org . Проверено 03 октября 2023 г.
- ^ «Венгерские и американские учёные получили Нобелевскую премию за открытие вакцины против COVID-19» . Рейтер . 2023-10-02 . Проверено 03 октября 2023 г.
- ^ «Нобелевская премия по физиологии и медицине 2023 года» . NobelPrize.org . Проверено 03 октября 2023 г.
- ^ Зиберт С. (2006). «Свойства общей структуры последовательностей и стабильные области во вторичных структурах РНК» (PDF) . Диссертация, Университет Альберта-Людвига, Фрайбург-им-Брайсгау . п. 1. Архивировано из оригинала (PDF) 9 марта 2012 г.
- ^ Сатмари Э (июнь 1999 г.). «Происхождение генетического кода: аминокислоты как кофакторы в мире РНК». Тенденции в генетике . 15 (6): 223–29. дои : 10.1016/S0168-9525(99)01730-8 . ПМИД 10354582 .
- ^ Джером, Крейг А.; и др. (19 мая 2022 г.). «Каталитический синтез полирибонуклеиновой кислоты на пребиотических каменных стеклах» . Астробиология . 22 (6): 629–636. Бибкод : 2022AsBio..22..629J . дои : 10.1089/ast.2022.0027 . ПМЦ 9233534 . ПМИД 35588195 . S2CID 248917871 .
- ^ Фонд прикладной молекулярной эволюции (3 июня 2022 г.). «Ученые объявляют о прорыве в определении происхождения жизни на Земле и, возможно, на Марсе» . Физика.орг . Проверено 3 июня 2022 г.
- ^ Марлер Р. (3 марта 2015 г.). «НАСА Эймс воспроизводит строительные блоки жизни в лаборатории» . НАСА . Архивировано из оригинала 5 марта 2015 года . Проверено 5 марта 2015 г.
- ^ Старр, Мишель (8 июля 2022 г.). «В центре нашей Галактики обнаружено множество предшественников РНК» . НаукаАлерт . Проверено 9 июля 2022 г.
- ^ Ривилла, Виктор М.; и др. (8 июля 2022 г.). «Молекулярные предшественники мира РНК в космосе: новые нитрилы в молекулярном облаке G + 0,693–0,027» . Границы астрономии и космических наук . 9 : 876870. arXiv : 2206.01053 . Бибкод : 2022FrASS...9.6870R . дои : 10.3389/fspas.2022.876870 .
- ^ Чех, Томас Р.; Стейтц, Джоан А. (март 2014 г.). «Революция некодирующих РНК: отказ от старых правил и создание новых» . Клетка . 157 (1): 77–94. дои : 10.1016/j.cell.2014.03.008 . ISSN 0092-8674 . ПМИД 24679528 . S2CID 14852160 .
- ^ Палачино, Джеймс; Суэлли, Сюзанна Э; Сун, Ченг; Чунг, Этвуд К.; Шу, Лей; Чжан, Сяолу; Ван Хусир, Майлин; Шин, Юнга; Чин, Донован Н; Келлер, Кэролайн Габсер; Бейбель, Мартин; Рено, Николь А; Смит, Томас М; Сальций, Майкл; Ши, Сяоин (01.06.2015). «Модуляторы сплайсинга SMN2 усиливают ассоциацию U1-пре-мРНК и спасают мышей со СМА». Химическая биология природы . 11 (7): 511–517. дои : 10.1038/nchembio.1837 . ISSN 1552-4450 . ПМИД 26030728 .
- ^ Рой, Биджойита; Фризен, Уэстли Дж.; Томизава, Юки; Лешик, Джон Д.; Чжо, Цзинь; Джонсон, Бриана; Дакка, Джумана; Тротта, Кристофер Р.; Сюэ, Сяоцзяо; Мутьям, Венкатешвар; Килинг, Ким М.; Мобли, Джеймс А.; Роу, Стивен М.; Бедвелл, Дэвид М.; Уэлч, Эллен М. (04 октября 2016 г.). «Аталурен стимулирует рибосомальную селекцию почти родственных тРНК, способствуя подавлению нонсенса» . Труды Национальной академии наук . 113 (44): 12508–12513. Бибкод : 2016PNAS..11312508R . дои : 10.1073/pnas.1605336113 . ISSN 0027-8424 . ПМЦ 5098639 . ПМИД 27702906 .
- ^ Перейти обратно: а б Кадир, Мухаммад Имран; Бухат, Шериен; Расул, Сумайра; Мансур, Хамид; Мансур, Маджид (3 сентября 2019 г.). «РНК-терапия: идентификация новых мишеней, ведущих к открытию лекарств». Журнал клеточной биохимии . 121 (2): 898–929. дои : 10.1002/jcb.29364 . ISSN 0730-2312 . ПМИД 31478252 . S2CID 201806158 .
- ^ Балмайор, Элизабет Р.; Гейгер, Йоханнес П.; Анея, Маниш К.; Бережанский, Тарас; Утцингер, Максимилиан; Михайлик, Ольга; Рудольф, Карстен; Планк, Кристиан (май 2016 г.). «Химически модифицированная РНК индуцирует остеогенез стволовых клеток и эксплантатов тканей человека, а также ускоряет заживление костей у крыс». Биоматериалы . 87 : 131–146. doi : 10.1016/j.bimaterials.2016.02.018 . ISSN 0142-9612 . ПМИД 26923361 .
- ^ Плевс, Джордан Р.; Ли, ЦзяньЛян; Джонс, Марк; Мур, Гарри Д.; Мейсон, Крис; Эндрюс, Питер В.; На, Цзе (30 декабря 2010 г.). «Активация генов плюрипотентности в клетках фибробластов человека с помощью нового подхода, основанного на мРНК» . ПЛОС ОДИН . 5 (12): e14397. Бибкод : 2010PLoSO...514397P . дои : 10.1371/journal.pone.0014397 . ISSN 1932-6203 . ПМК 3012685 . ПМИД 21209933 .
- ^ Прески, Дэвид; Эллисон, Томас Ф.; Джонс, Марк; Мамчауи, Камель; Унгер, Кристиан (май 2016 г.). «Синтетически модифицированная мРНК для эффективного и быстрого создания iPS-клеток человека и прямой трансдифференцировки в миобласты». Связь с биохимическими и биофизическими исследованиями . 473 (3): 743–751. дои : 10.1016/j.bbrc.2015.09.102 . ISSN 0006-291X . ПМИД 26449459 .
- ^ Уоррен, Луиджи; Манос, Филип Д.; Ахфельдт, Тим; Ло, Юин-Хан; Ли, Ху; Лау, Фрэнк; Эбина, Ватару; Мандал, Панкадж К.; Смит, Закари Д.; Мейснер, Александр; Дейли, Джордж К.; Брэк, Эндрю С.; Коллинз, Джеймс Дж.; Коуэн, Чад; Шлегер, Торстен М. (ноябрь 2010 г.). «Высокоэффективное перепрограммирование плюрипотентности и направленная дифференцировка клеток человека с помощью синтетической модифицированной мРНК» . Клеточная стволовая клетка . 7 (5): 618–630. дои : 10.1016/j.stem.2010.08.012 . ISSN 1934-5909 . ПМЦ 3656821 . ПМИД 20888316 .
- ^ Элангован, Сатиш; Хорсанд, Бенуш; До, Ань-Ву; Хун, Лю; Деверт, Александр; Корманн, Майкл; Росс, Райан Д.; Рик Самнер, доктор медицинских наук; Алламарго, Шанталь; Салем, Алиасгер К. (ноябрь 2015 г.). «Химически модифицированные матрицы, активированные РНК, улучшают регенерацию костей» . Журнал контролируемого выпуска . 218 : 22–28. дои : 10.1016/j.jconrel.2015.09.050 . ISSN 0168-3659 . ПМЦ 4631704 . ПМИД 26415855 .
Внешние ссылки
[ редактировать ]- Веб-сайт RNA World Коллекция ссылок (структуры, последовательности, инструменты, журналы)
- База данных нуклеиновых кислот Изображения ДНК, РНК и комплексов.
- Семинар Анны Мари Пайл: Структура, функции и распознавание РНК
Виды нуклеиновых кислот |
|---|