Диатомо
| Диатомо Временный диапазон:
| |
|---|---|
| |
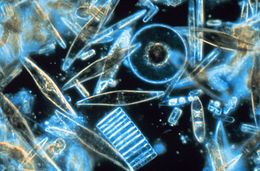
| Световая микроскопия отбора проб морских диатомов, обнаруженных между кристаллами годового морского льда в Антарктике, показывая множество размеров и форм | |
| Научная классификация | |
| Домен: | Эукариота |
| Клада : | Диафоретики |
| Клада : | Сар - |
| Клада : | Стейнамиле |
| Филум: | Гириста |
| Субфилум: | Ochrophytina |
| Инфрафилум: | От диатомного |
| Сорт: | Bacillariophyceae Dangeard , 1933 [ 1 ] |
| Подклассы [ 2 ] | |
| |
| Синонимы | |
| |
диатомо латинская ( нео- диатома ) [ А ] Является ли любой член большой группы, содержащей несколько родов водорослей микроводоросля , в частности, , обнаруженных в океанах, водных путях и почвах мира. Земли Живые диатомомы составляют значительную часть биомассы : они генерируют от 20 до 50 процентов кислорода, производимого на планете каждый год, [ 11 ] [ 12 ] принимать более 6,7 миллиардов тонн кремния каждый год из вод, в которых они живут, [ 13 ] и составляют почти половину органического материала, найденного в океанах. Оболочки Сахары мертвых диатомовых средств могут достигать полмили (800 м) глубиной на дне океана, и весь бассейн Амазонки ежегодно оплодотворяется 27 миллионов тонн пыли с раковиной диатомовой раковины, трансатлантической ветры от африканской , много из этого от депрессии Боделе , которая когда-то была составлена из системы свежевыводных озер. [ 14 ] [ 15 ]
Диатомы являются одноклеточными организмами : они встречаются либо в качестве одиночных клеток, либо в колониях , которые могут принимать форму лент, вентиляторов, зигзагов или звезд. Отдельные ячейки варьируются в размере от 2 до 2000 микрометров. [ 16 ] При наличии адекватных питательных веществ и солнечного света сборка живых диатомов удваивается примерно каждые 24 часа путем бесполого многочисленного деления ; Максимальная продолжительность жизни отдельных клеток составляет около шести дней. [ 17 ] Диатомы имеют две отдельные формы: несколько ( центрических диатомов ) являются радиально симметричными, в то время как большинство ( диатомовые пенсии ) в целом являются двусторонними симметричными.
Уникальная особенность диатомовых заболеваний заключается в том, что они окружены клеточной стенкой из кремнезема (гидратированный диоксид кремния ), называемый ядром . [ 18 ] Эти ящики производят структурную окраску , побуждая их описать как «драгоценности моря» и «живые опалы».
Движение в диатомеях в первую очередь происходит пассивно в результате как океанских течений воды, вызванной ветром , так и турбулентности ; Тем не менее, мужские гаметы центрических диатомов имеют жгутиков , позволяющие активному движению искать женских гамет. Подобно растениям , диатомы превращают энергию света в химическую энергию путем фотосинтеза , но их хлоропласты были получены по -разному. [ 19 ]
Необычно для автотрофных организмов, диатомовые данные обладают циклом мочевины , что они разделяют с животными , хотя этот цикл используется для различных метаболических целей в диатомовых условиях. Семейство Rhopalodiaceae также обладает цианобактериальным эндосимбионтом, называемым сфероидным телом. Этот эндосимбионт утратил свои фотосинтетические свойства, но сохранил свою способность выполнять фиксацию азота , позволяя диатоме фиксировать атмосферный азот . [ 20 ] Другие диатомоды в симбиозе с азотом-фиксирующими цианобактериями являются одними из родов Hemiaulus , Rhizosolenia и Chaetoceros . [ 21 ]
Динотомы - это диатомовые лечения, которые стали эндосимбионтами внутри динофлагеллятов. Исследования динофлагелляции Durinskia Baltica и Glenodinium foliaceum показали, что событие эндосимбионта произошло так недавно, эволюционно говоря, что их органеллы и геном все еще не повреждены с минимальной или отсутствием потери генов. Основное различие между этими и свободными диатомовыми диатомогами состоит в том, что они потеряли свою клеточную стенку кремнезема, что делает их единственными известными раковинами диатомогами. [ 22 ]
Изучение диатомовых заболеваний является ветвью фикологии . Диатомы классифицируются как эукариоты , организмы с ядерным -оболочки ядром клеток , которые отделяют их от прокариот археи и бактерий . Диатомы - это тип планктона , называемый фитопланктоном , наиболее распространенным из типов планктона. Диатомы также растут прикрепленными к бентическим субстратам, плавающим мусорам и на макрофитах . Они составляют неотъемлемый компонент перифитонного сообщества. [ 23 ] Другая классификация делит планктон на восемь типов в зависимости от размера: в этой схеме диатомовые средства классифицируются как микроводоросли. Существуют несколько систем для классификации отдельных видов диатомов .
Факты ископаемых свидетельствуют о том, что диатомы возникли в течение или до раннего периода юрского периода, который составлял около 150-200 миллионов лет назад. Самым старым ископаемым свидетельством диатомовых мышц является образец существующего рода Hemiaulus у позднего юрского периода, выдержанного янтаря из Таиланда. [ 24 ]
Диатомы используются для мониторинга прошлых и настоящих условий окружающей среды и обычно используются в исследованиях качества воды. Диатоматическая земля (диатомит) представляет собой набор раковин диатомовых оболочек, обнаруженных в коре Земли. Они представляют собой мягкие, содержащие кремнезем осадочные породы, которые легко рассыпаются в мелком порошке и обычно имеют размер частиц от 10 до 200 мкм. Диатоматическая земля используется для различных целей, в том числе для фильтрации воды, в качестве мягкого абразива, в кошачьем мусоре и в качестве стабилизатора динамита.
Отображает наложения из четырех флуоресцентных каналов
(B) Cyan: [PLL -A546 флуоресценция] - общая контратака для визуализации поверхностей эукариотических клеток
(c) Синий: [Hoechst Fluorescencess] - ДНК пятна, идентифицирует ядра
(D) Красный: [АВТОФЛУАРЕССОВАНИЕ ХОЛОФИЛЛЯ [ 27 ]
Обзор
[ редактировать ]Диатомы - это протисты, которые образуют массовые ежегодные весенние и осенние цветы в водных средах и, по оценкам, являются ответственными за примерно половину фотосинтеза в глобальных океанах. [ 28 ] Этот предсказуемый годовой динамический цветущий питает более высокие трофические уровни и инициирует доставку углерода в глубокий океанский биом. Диатомы имеют сложные стратегии жизненной истории, которые, как предполагается, способствовали их быстрой генетической диверсификации в ~ 200 000 видов [ 29 ] которые распределены между двумя основными группами диатомовых групп: центрики и выплаты. [ 30 ] [ 31 ]
Морфология
[ редактировать ]Диатомовые данные, как правило, имеют размеры от 20 до 200 микрометров, [ 32 ] с несколькими большими видами. Их желтовато-коричневые хлоропласты , место фотосинтеза, типичны для гетероконтов , имеющие четыре клеточные мембраны и содержащие пигменты , такие как каротиноидный фукомсантин . Людям обычно не хватает жгутиков , но они присутствуют в мужских гаметах ориентированных диатомовых диатомов и имеют обычную гетероконную структуру, включая характеристику волосков ( мастигонемов ) в других группах.
Диатомы часто называют «драгоценностями моря» или «живые опалы» из -за их оптических свойств. [ 33 ] Биологическая функция этой структурной окраски не ясна, но предполагается, что она может быть связана с связью, камуфляжем, тепловым обменом и/или ультрафиолетовой защитой. [ 34 ]
Диатомы строят замысловатые твердые, но пористые клеточные стенки, называемые фрультами, состоящими в основном из кремнезема . [ 35 ] : 25–30 Эта кремнистая стена [ 36 ] может быть очень узорчан с различными пор, ребрами, мельчайшими шипами, маргинальными хребтами и возвышением; Все это может быть использовано для определения родов и видов.
Сама ячейка состоит из двух половинок, каждая из которых содержит по существу плоскую пластину, или клапан, а также маргинальный соединительный, или полоса пояса. Половина, гипотека , немного меньше, чем другая половина, эпитека . Морфология диатома варьируется. Хотя форма клетки обычно является круглой, некоторые клетки могут быть треугольными, квадратными или эллиптическими. Их отличительная особенность - жесткая минеральная оболочка или ядовита, состоящая из опала (гидратированная полимеризованная креминовая кислота).

- Ядро ; держит генетический материал
- Ядра ; расположение хромосом
- Гольджи аппарат ; изменяет белки и отправляет их из клетки
- Клеточная стенка ; внешняя мембрана клетки
- Пиренедоид ; центр углеродной фиксации
- Хроматофор ; пигмент, несущая мембрану
- Вакуоли ; везикула клетки, которая содержит жидкость, связанную мембраной
- Цитоплазматические пряди; Держите ядро
- Митохондрия ; Создать ATP (энергия) для ячейки
- Клапаны/Striae; Разрешить питательные вещества и тратить клетку
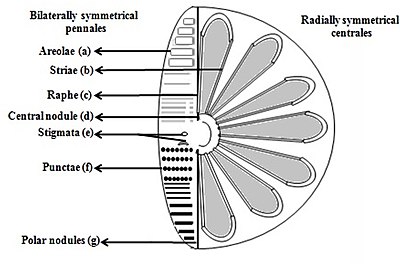
- Ареола (шестиугольная или полигональная коробчатая перфорация с ситом, присутствующим на поверхности диатема)
- Striae (поры, точки, пятна или точки в линии на поверхности)
- Рафе (щель в клапанах)
- Центральный узелок (утолщение стены в средней точке рафе)
- Стигмата (отверстия через поверхность клапана, которая выглядит округлой внешне, но с помощью щели, похожей на внутреннюю)
- Точки (пятна или небольшие перфорации на поверхности)
- Полярные узелки (утолщение стенки на дистальных концах рафе) [ 37 ] [ 38 ]

Диатомы делятся на две группы, которые различаются по форме ящика: центрические диатомовые и типные диатомовые .
Диатомы выписки являются двусторонними симметричными. У каждого из их клапанов есть отверстия, которые прорези вдоль рафе , а их раковины обычно удлинены параллельно этим рафам. Они генерируют движение клеток через цитоплазму, которая течет вдоль рафе, всегда движущихся по твердым поверхностям.
Центрические диатомы радиально симметричны. Они состоят из верхних и нижних клапанов - эпитека и гипотека - каждая из которых состоит из клапана и полосы пояса, которые могут легко скользить друг на друга и расширять, чтобы увеличить содержание клеток по прогрессии диатомовых средств. Цитоплазма центрической диатомы расположена вдоль внутренней поверхности оболочки и обеспечивает пустую подкладку вокруг большой вакуоли, расположенной в центре клетки. Эта большая центральная вакуоль заполняется жидкостью, известной как «клеточный сок», которая похожа на морскую воду, но варьируется в зависимости от определенного содержания ионов. Цитоплазматический слой является домом для нескольких органеллов, таких как хлоропласты и митохондрии. Перед тем, как центричный диатом начинает расширяться, его ядро находится в центре одного из клапанов и начинает двигаться к центру цитоплазматического слоя до завершения деления. Центрические диатомы имеют различные формы и размеры, в зависимости от того, от какой оси простирается оболочка, и если присутствуют колючки.

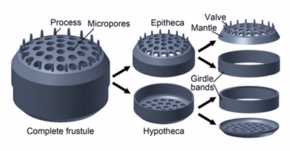
Силикация
[ редактировать ]Диатомовые клетки содержатся в уникальной стенке клеточной клетки кремнезема , известной как ядовитая, состоящая из двух клапанов, называемых Thecae , которые обычно перекрывают друг друга. [ 40 ] Биогенный кремнезем, составляющий клеточную стенку, синтезируется внутриклеточно путем полимеризации из кремниковой кислоты мономеров . Этот материал затем экструдируется на внешнюю часть ячейки и добавляется в стену. У большинства видов, когда диатом делится для получения двух дочерних клеток, каждая клетка сохраняет одну из двух половинок и выращивает меньшую половину в ней. В результате после каждого цикла деления средний размер диатомовых ячеек в популяции становится меньше. Как только такие клетки достигают определенного минимального размера, а не просто делятся, они обращают вспять этот снижение, образуя ауксоспору , обычно через мейоз и сексуальное размножение, но существуют исключения. Auxospore расширяется в размере, чтобы привести к гораздо большей ячейке, которая затем возвращается к подразделениям, димизирующим размер. [ 41 ]



Точный механизм переноса кремнезема , поглощенного диатомом в клеточную стенку, неизвестен. Большая часть секвенирования генов диатомовых генов происходит из-за поиска механизма поглощения кремнезема и осаждения в наномасштабных паттернах в rustule . Самый успех в этой области был получен от двух видов, Thalassiosira Pseudonana , который стал модельными видами, поскольку весь геном был секвенирован и были установлены методы генетического контроля, и цилиндротека Fusiformis , в которых были первыми важными белками осаждения кремнезема Силафины Силафины Силафины и Силика Силафины Силафины обнаруженный. [ 43 ] Салаффины, наборы поликационных пептидов , были обнаружены в клеточных стенках C. fusiformis и могут генерировать сложные структуры кремнезема. Эти структуры продемонстрировали поры размеров, характерные для диатомовых узоров. Когда Т. Псевдонана прошел анализ генома, было обнаружено, что он кодирует цикл мочевины , включая большее количество полиаминов , чем большинство геномов, а также три различных гена транспорта кремнезема. [ 44 ] В филогенетическом исследовании генов транспортировки кремнезема из 8 различных групп диатомовых средств был обнаружен, что транспорт кремнезема обычно группирует с видами. [ 43 ] Это исследование также обнаружило структурные различия между транспортерами кремнезема типа выступающих (двусторонняя симметрия) и центрическими (радиальными симметрией) диатомогами. Последовательности, сравниваемые в этом исследовании, использовались для создания разнообразного фона для определения остатков, которые дифференцируют функцию в процессе осаждения кремнезема. Кроме того, то же самое исследование показало, что ряд регионов сохранялся в видах, вероятно, базовая структура транспорта кремнезема.
Эти белки транспорта кремнезема уникальны для диатомов, без гомологов у других видов, таких как губки или рис. Дивергенция этих генов транспортировки кремнезема также указывает на структуру белка, развивающегося из двух повторных единиц, состоящих из пяти сегментов, связанных с мембраной, что указывает либо на дублирование генов, либо димеризацию . [ 43 ] Предполагается, что осаждение кремнезема, которое происходит из мембранной пузырьки в диатомеях, является результатом активности силафинов и полиаминов длинных цепей. Эта пузырька осаждения кремнезема (SDV) была охарактеризована как кислый компартмент, слитый с везикулами, полученными из Голги. [ 45 ] Было показано, что эти две белковые структуры создают листы стержня кремнезема in vivo с нерегулярными пор по шкале диатомовых плоскостей . Одна гипотеза о том, как работают эти белки для создания сложной структуры, заключается в том, что остатки сохраняются в SDV, что, к сожалению, трудно идентифицировать или наблюдать из -за ограниченного количества доступных различных последовательностей. Хотя точный механизм очень равномерного осаждения кремнезема пока неизвестен, гены псевдонана Thalassiosira, связанные с силафинами, рассматриваются как мишени для генетического контроля наноразмерного осаждения кремнезема.
Способность диатомовых на основе кремнезема стенков была предметом очарования на протяжении веков. Это началось с микроскопического наблюдения анонимного английского дворянина в 1703 году, который наблюдал объект, который выглядел как цепь регулярных параллелограммов и обсуждал, были ли это просто кристаллы соли или растения. [ 46 ] Зритель решил, что это было растение, потому что параллелограммы не отделялись при возбуждении, и при этом они не различались по внешнему виду при высушении или подверженности теплой воде (в попытке растворить «соль»). Незначно, путаница зрителя захватила сущность диатомовых средств - минеральные использующие растения. Не ясно, когда было определено, что диатомовые клеточные стенки изготовлены из кремнезема, но в 1939 году основополагающая ссылка охарактеризовала материал как кремнизную кислоту в «субколлоидальном состоянии» [ 47 ] Идентификация основного химического компонента клеточной стенки побудила исследования того, как она была сделана. Эти исследования были включены и были продвинуты различными подходами, включая микроскопию, химию, биохимию, характеристику материала , молекулярная биология , омика и трансгенные подходы. Результаты этой работы дали лучшее понимание процессов формирования клеточной стенки, установив фундаментальные знания, которые можно использовать для создания моделей, которые контекстуализируют текущие выводы, и прояснить, как работает процесс. [ 48 ]
Процесс создания клеточной стенки на основе минералов внутри клетки, затем экспорта ее снаружи, представляет собой огромное событие, которое должно включать большое количество генов и их белковых продуктов. Акт построения и экзоцитирования этого большого структурного объекта в течение короткого периода времени синхронизировался с прогрессированием клеточного цикла способностей клетки , требует существенных физических движений в ячейке, а также преданности значительной доли биосинтетических . [ 48 ]
Первые характеристики биохимических процессов и компонентов, участвующих в силицификации диатома, были сделаны в конце 1990 -х годов. [ 49 ] [ 50 ] [ 51 ] За ними последовало понимание того, как может возникнуть сборка конструкций кремнезема более высокого порядка. [ 52 ] [ 53 ] [ 54 ] Более поздние сообщения описывают идентификацию новых компонентов, участвующих в процессах более высокого порядка, динамики, документированной с помощью визуализации в реальном времени, и генетических манипуляций с структурой кремнезема. [ 55 ] [ 56 ] Подходы, установленные в этих недавних работах, обеспечивают практические возможности не только идентифицировать компоненты, участвующие в формировании стенки клетки кремнезема, но и для выяснения их взаимодействия и пространственно-временной динамики. Этот тип целостного понимания потребуется для достижения более полного понимания синтеза клеточной стенки. [ 48 ]
Поведение
[ редактировать ]
Великолепно, 1897
Большинство центричных и арафидных диатомов не являются мотильными , и их относительно плотные клеточные стены заставляют их легко погрузиться. Планктонные формы в открытой воде обычно зависят от турбулентного смешивания верхних слоев океанических вод с ветром, чтобы они подвешены в солнечных поверхностных водах. Многие планктонные диатомовые диатомы также развивали особенности, которые замедляют их скорость тонука, такие как шипы или способность расти в колониальных цепях. [ 57 ] Эти адаптации увеличивают их площадь поверхности к объему и сопротивление , что позволяет им дольше оставаться подвешенными в толще воды. Отдельные клетки могут регулировать плавучесть с помощью ионного насоса. [ 58 ]
Некоторые типичные диатомы способны к типу локомоции, называемого «скользящим», что позволяет им перемещаться по поверхностям через клейкую слизь, секретируемую через шва, называемую рафе. [ 59 ] [ 60 ] Для того, чтобы диатомовая ячейка скользила, она должна иметь твердый субстрат для слизи, чтобы придерживаться.
Клетки одиноки или объединены в колонии различных видов, которые могут быть связаны кремнистыми структурами; слизистые прокладки, стебли или трубки; аморфные массы слизи; или по нитям хитина (полисахарида), которые секретируются через распределенные процессы клетки.


Эта проекция стека конфокальных изображений диатомов показывает клеточную стенку (голубой), хлоропласты (красный), ДНК (синий), мембраны и органеллы (зеленые).
Жизненный цикл
[ редактировать ]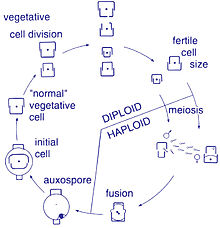
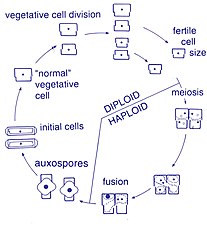
Размножение и размер ячейки
[ редактировать ]Воспроизведение среди этих организмов является бесполым путем бинарного деления , в ходе которого диатом делится на две части, создавая два «новых» диатомовых диатовых данных с идентичными генами. Каждый новый организм получает одну из двух фрукторов - одну большую, другую меньшую - одержимый родителем, который теперь называется эпитекой ; и используется для построения второго, меньшего домочителя, гипотеки . Диатом, получивший больший ярезок, становится тем же размером, как и его родитель, но диатомовая, которая получила меньшую ядовитую, остается меньше, чем его родитель. Это приводит к снижению среднего размера клеток этой популяции диатомы. [ 16 ] Однако наблюдалось, что определенные таксоны обладают способностью делиться, не вызывая уменьшения размера клеток. [ 61 ] Тем не менее, для восстановления размера клеток популяции диатомовой популяции для тех, кто терпит снижение размера, должно произойти сексуальное воспроизведение и образование ауксоспоры . [ 16 ]
Клеточная деление
[ редактировать ]Вегетативные клетки диатомов являются диплоидными (2n), и поэтому мейоз может произойти , производя мужские и женские гаметы, которые затем сливаются, чтобы сформировать зиготу . Зигота проливает свою кремнеземную тека и превращается в большую сферу, покрытую органической мембраной, Auxospore. Новая диатомовая ячейка максимального размера, начальная ячейка, формируется в пределах ауксоспоры, тем самым начинает новое поколение. Споры отдыха также могут быть сформированы как ответ на неблагоприятные условия окружающей среды с прорастанием, возникающим при улучшении условий. [ 35 ]
Определяющей характеристикой всех диатомов является их ограничительная и двухпартийная клеточная стенка кремнезема, которая заставляет их постепенно сокращаться во время бесполого деления клеток. При критическом небольшом размере клеток и в определенных условиях ауксоспоризация реститует размер клеток и предотвращает клональную гибель. [ 62 ] [ 63 ] [ 64 ] [ 65 ] [ 66 ] Весь жизненный цикл только несколько диатомовых средств были описаны, и в окружающей среде редко были запечатлены сексуальные события. [ 31 ]
Сексуальное размножение
[ редактировать ]Большинство эукариот способны к сексуальному размножению , связанному с мейозом . Сексуальное размножение, по -видимому, является обязательной фазой в жизненном цикле диатомовых заболеваний, особенно в том, что размер клеток уменьшается с последовательными вегетативными делениями. [ 67 ] Сексуальное воспроизведение включает в себя производство гаметов и слияние гамет, чтобы сформировать зиготу , в которой восстанавливается максимальный размер клеток. [ 67 ] Сигнализация, которая запускает сексуальную фазу, предпочитается, когда клетки накапливаются вместе, так что расстояние между ними уменьшается, а контакты и/или восприятие химических сигналов облегчено. [ 68 ]
Исследование геномов пяти диатомов и одного транскриптома диатома привело к идентификации 42 генов, потенциально вовлеченных в мейоз. [ 69 ] Таким образом, мейотический инструментарий, по -видимому, консервативный у этих шести видов диатомовых [ 69 ] указывает на центральную роль мейоза в диатомовых условиях, как у других эукариот.
Подвижность спермы
[ редактировать ]Диатомы в основном не мотильны ; Тем не менее, сперма, найденная у некоторых видов, может быть жгутирована , хотя подвижность обычно ограничивается скользящим движением. [ 35 ] В центрических диатомовых лицах маленькие мужские гаметы имеют один жгутичок , в то время как женские гаметы большие и не мотильные ( оогамные ). И наоборот, в Pennate Diatoms у обоим гаметам не хватает жгутиков ( изогам ). [ 16 ] Некоторые виды арафидов, то есть типичные диатомовые данные без рафе (шв), были задокументированы как анизогамные и, следовательно, считаются переходной стадией между центрическими и рафидными диатомогами, диатомовыми с помощью рафе. [ 61 ]
Деградация по микробам
[ редактировать ]Определенные виды бактерий в океанах и озерах могут ускорить скорость растворения кремнезема в мертвых и живых диатомеях, используя гидролитические ферменты для разрушения органического водорослевого материала. [ 70 ] [ 71 ]
Экология
[ редактировать ]| Часть серии на |
| Планктон |
|---|
 |


по сравнению с концентрацией силиката [ 72 ]
Распределение
[ редактировать ]Диатомы являются широко распространенной группой и могут быть найдены в океанах , в пресной воде , в почвах и на влажных поверхностях. Они являются одним из доминирующих компонентов фитопланктона в богатых питательными веществами прибрежными водами и во время цветов океанических весенних цветов, поскольку они могут разделить быстрее, чем другие группы фитопланктона. [ 73 ] Большинство живых пелагически в открытой воде, хотя некоторые живут как поверхностные пленки на границе раздела водяного связки ( бентическая ) или даже в влажных атмосферных условиях. Они особенно важны в океанах, где исследование 2003 года показало, что они вносят около 45% от общего океанического первичного производства органического материала. [ 74 ] Тем не менее, более недавнее исследование 2016 года оценивает, что число ближе к 20%. [ 75 ] Пространственное распределение морских видов фитопланктона ограничено как горизонтально, так и вертикально. [ 76 ] [ 35 ]
Рост
[ редактировать ]Планктонные диатомовые данные в пресной и морской среде, как правило, демонстрируют образ жизни « бум и бюст » (или « цветение и бюст»). Когда условия в верхнем смешанном слое (питательные вещества и свет) благоприятны (как на пружине ), их конкурентный край и быстрый рост [ 73 ] Позволяет им доминировать в сообществах фитопланктона («бум» или «цветение»). Таким образом, они часто классифицируются как оппортунистические R-стратегии (то есть те организмы, экология которых определяется высокими темпами роста, R ).
Влияние
[ редактировать ]пресной воды Гемината , широко известная как Didymo, вызывает серьезную деградацию окружающей среды в водопроводных покрытиях, где она цветут, производя большие количества коричневого желе-впоследствии, называемого «коричневым соном» или «скалистых сопли». Эта диатомовая является родной для Европы и является инвазивным видом как в антиподах , так и в некоторых частях Северной Америки . [ 77 ] [ 78 ] Проблема чаще всего записывается из Австралии и Новой Зеландии . [ 79 ]
Когда условия становятся неблагоприятными, обычно при истощении питательных веществ, диатомовые клетки обычно увеличивают скорость тонука и выходят из верхнего смешанного слоя («бюст»). Это погружение вызвано либо потерей контроля плавучести, синтезом слизи, который соединяет диатологические ячейки, либо производством тяжелых покоящихся споров . Тонут из верхнего смешанного слоя удаляет диатомы из условий, неблагоприятных до роста, включая популяции Grazer и более высокие температуры (что в противном случае увеличит метаболизм клеток ). Клетки, достигающие более глубокой воды или мелкого морского дна, могут затем отдыхать, пока условия снова не станут более благоприятными. В открытом океане многие тонущие клетки теряются в глубине, но убежистые популяции могут сохраняться вблизи термоклина .
В конечном счете, диатомовые клетки в этих популяциях покоя повторно входят в верхний смешанный слой, когда вертикальное смешивание уклоняется от них. В большинстве случаев это смешивание также пополняет питательные вещества в верхнем смешанном слое, устанавливая сцену для следующего раунда цветов диатомовых. В открытом океане (вдали от районов непрерывного подъема [ 80 ] ), этот цикл цветения, бюста, затем возвращается в условия предварительного цвета, как правило, происходит в течение годового цикла, причем диатомовые диатомы распространены только в течение весны и начала лета. Однако в некоторых местах может произойти осеннее цветение, вызванное расщеплением летней стратификации и увлечением питательными веществами, в то время как уровень освещения по -прежнему достаточен для роста. Поскольку вертикальное смешивание увеличивается, а уровень освещения падает при зимних подходах, эти цветы меньше и меньше, чем их пружинные эквиваленты.
В открытом океане цветение диатомовой (пружины) обычно заканчивается нехваткой кремния. В отличие от других минералов, потребность в кремнии является уникальным для диатомовых заболеваний и не регенерируется в экосистеме планктона так эффективно, как, например, азотные или фосфорные питательные вещества. Это можно увидеть на картах концентраций питательных веществ в поверхностных веществах - поскольку питательные вещества снижаются вдоль градиентов, кремний обычно первым измотаны (обычно следовать азотом, затем фосфор).
Из-за этого цикла расцвета, диатомовые данные играют непропорционально важную роль в экспорте углерода из океанических поверхностных вод [ 80 ] [ 81 ] (См. Также биологический насос ). они также играют ключевую роль в регуляции биогеохимического цикла кремния Примечательно, что в современном океане. [ 74 ] [ 82 ]
Причина успеха
[ редактировать ]Диатомы экологически успешны и встречаются практически в каждой среде, которая содержит воду - не только океаны, моря, озера и ручьи, но и на почву и водно -болотные угодья. [ Цитация необходима ] Многие исследователи считают, что использование кремния по диатомом является ключом к этому экологическому успеху. Ворон (1983) [ 83 ] Отметил, что, по сравнению с органическими клеточными стенками , кремнеземные фрукторы требуют меньшей энергии для синтеза (приблизительно 8% сопоставимой органической стенки), что потенциально значительно экономит общий бюджет энергии клеток. В ныне классическом исследовании Egge and Aksnes (1992) [ 72 ] обнаружено, что доминирование диатомовых сообществ мезокосма было напрямую связано с наличием кремниковой кислоты - когда концентрации превышали 2 мкмоль М. −3 Они обнаружили, что диатомовые средства обычно представляют более 70% сообщества фитопланктона. Другие исследователи [ 84 ] предположили, что биогенный кремнезем в клеточных стенках диатомовых клеток действует как эффективный агент PH -буферизации , облегчая преобразование бикарбоната в растворенное CO 2 (который более легко усваивается). В более общем плане, несмотря на эти возможные преимущества, предоставляемые их использованием кремния, диатомовые данные обычно имеют более высокие темпы роста, чем другие водоросли того же соответствующего размера. [ 73 ]
Источники для сбора
[ редактировать ]Диатомы могут быть получены из нескольких источников. [ 85 ] Морские диатомы могут быть собраны путем прямого отбора проб воды, а бентические формы могут быть закреплены путем соскоба сареев , устриц и других раковинов. Диатомы часто присутствуют в виде коричневого, скользкого покрытия на погруженных камнях и палках, и можно увидеть, как «транслируется» с речным током. Поверхностная грязь пруда, канава или лагуны почти всегда даст некоторые диатомовые данные. Живые диатомы часто встречаются в большом количестве в нити, или образуют желатиновые массы на различных затопленных растениях. Cladophora часто покрыта кокконисом , эллиптической диатовой; Vaucheria часто покрыта небольшими формами. Поскольку диатомовые средства образуют важную часть пищи моллюсков , туникатов и рыб , пищевые тракты этих животных часто дают формы, которые нелегко закрепить другими способами. Диатомы могут быть сделаны, чтобы появиться, заполнив банку водой и грязью, завернув ее в черную бумагу и позволив прямым солнечным свети. В течение дня диатомовые ткани будут выйти на вершину в мраке и могут быть изолированы. [ 85 ]
Биогеохимия
[ редактировать ]-
 Современный океанический кремниевый цикл
Современный океанический кремниевый цикл
Потоки в t mol si y −1 (1 тмоль = 28 миллионов метрических тонн кремния)

Цикл кремнезема
[ редактировать ]Диаграмма показывает основные потоки кремния . в нынешнем океане Большая часть биогенного кремнезема в океане ( кремнезем , продуцируемый биологической активностью ) поступает из диатомовых заболеваний. Экстракт диатомов растворил кремнистую кислоту из поверхностных вод по мере их роста, и возвращают его в столб воды, когда они умирают. Входы кремния прибывают сверху через эоловую пыль , от побережья по рекам, и снизу утилизация морского дна , выветривание и гидротермальную активность . [ 82 ]
Хотя диатомовые средства могли существовать с момента триаса , сроки их господства и «поверка» кремниевого цикла произошли в последнее время. До фанерозоя (до 544 млн лет) считается, что микробные или неорганические процессы слабо регулируют кремниевый цикл океана. [ 86 ] [ 87 ] [ 88 ] Впоследствии в цикле появляется доминирование (и более сильно регулируется) радиояльцами и кремнистыми губками , первое как зоопланктон , последняя, как сидячие фильтчики, в первую очередь на континентальных полках . [ 89 ] В течение последних 100 моих, считается, что кремниевый цикл оказался под еще более плотным контролем, и что это происходит от экологического господства диатомовых заболеваний.
Тем не менее, точное время «повернуть» остается неясным, и разные авторы имеют противоречивые интерпретации ископаемой записи. Некоторые доказательства, такие как смещение кремнистых губок с полков, [ 90 ] предполагает, что это поглощение началось в меловом периоде (от 146 млн. До 66 млн. Лет), в то время как доказательства радиояловодов предполагают, что «взрыв» не начались до кайнозоя (66 млн. Лет до настоящего времени). [ 91 ]
-
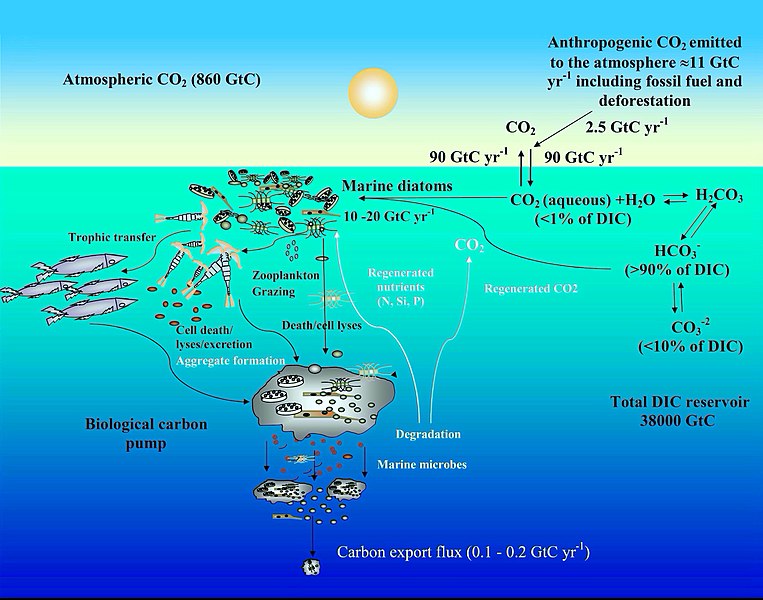 Цикл углерода в океане и механизмы концентрации диоксида углерода [ 92 ]
Цикл углерода в океане и механизмы концентрации диоксида углерода [ 92 ]

Углеродный цикл
[ редактировать ]Диаграмма изображает некоторые механизмы, с помощью которых морские диатомеры способствуют биологическому углеродному насосу и влияют на цикл углерода в океане . Эмиссия антропогенного CO 2 в атмосферу (в основном генерируемое сжиганием и вырубкой лесов) составляет почти 11 Gigatonne Carbon (GTC) в год, из которых почти 2,5 GTC поглощается поверхностным океаном. В поверхностной морской воде ( рН 8,1–8,4), бикарбонат ( HCO −
3 ) и карбонатные ионы ( co 2−
3 ) составляют почти 90 и <10% растворенного неорганического углерода (DIC) соответственно, в то время как растворенный CO 2 (CO 2 -водный) вносит <1%. Несмотря на этот низкий уровень CO 2 в океане и его медленную скорость диффузии в воде, диатомы фиксируют 10–20 GTC в год с помощью фотосинтеза благодаря их механизмам концентрации углекислого газа , что позволяет им поддерживать морские пищевые цепи . Кроме того, 0,1–1% этого органического материала, полученного в эйвфотическом слое, опускается в виде частиц, тем самым перенося поверхностный углерод в глубокий океан и секвестрирующий атмосферный CO 2 в течение тысяч лет или дольше. Оставшееся органическое вещество повсеместно повсеместно . Таким образом, диатомы являются одним из основных игроков в этом биологическом углеродном насосе, который, возможно, является наиболее важным биологическим механизмом в системе Земли, позволяющий CO 2 удалять из углеродного цикла в течение очень длительного периода. [ 93 ] [ 92 ]
-
 Митохондриальный цикл мочевины в общей диатомовой клетке и потенциальных судьбах промежуточных продуктов цикла мочевины [ 94 ]
Митохондриальный цикл мочевины в общей диатомовой клетке и потенциальных судьбах промежуточных продуктов цикла мочевины [ 94 ]

Цикл мочевины
[ редактировать ]Особенностью диатомов является цикл мочевины , который связывает их эволюционно с животными. В 2011 году Allen et al. Установил, что диатомовые данные имеют функционирующий цикл мочевины. Этот результат был значимым, так как до этого, как полагалось, цикл мочевины возник с метазоанами , которые появились за несколько сотен миллионов лет до диатомов. Их исследование показало, что, хотя диатомовые и животные используют цикл мочевины для разных целей, они считаются эволюционно связанными таким образом, что животные и растения нет. [ 95 ]
В то время как часто упускают из виду в фотосинтетических организмах, митохондрии также играют критическую роль в энергетическом балансе. Два пути, связанные с азотом, актуальны, и они также могут измениться в рамках аммония ( NH +
4 ) Питание по сравнению с нитратом ( нет −
3 ) Питание. Во -первых, в диатомовых условиях и, вероятно, в некоторых других водорослях, есть цикл мочевины. [ 96 ] [ 97 ] [ 98 ] Давно известная функция цикла мочевины у животных состоит в том, чтобы выделять избыток азота, продуцируемый аминокислотным катаболизмом ; Как и фотообожищение , цикл мочевины долгое время считался путем отходов. Однако в диатомовых условиях цикл мочевины, по -видимому, играет роль в обмене питательными веществами между митохондриями и цитоплазмой , и потенциально пластид [ 99 ] и может помочь регулировать метаболизм аммония. [ 96 ] [ 97 ] Из-за этого цикла морские диатомы, в отличие от хлорофитов , также приобрели митохондриальный транспортер мочевины и, фактически, на основе биоинформатики полный митохондриальный цикл GS-Gogat . , был вывоен [ 97 ] [ 94 ]
Другой
[ редактировать ]Диатомы в основном фотосинтетические; Однако некоторые из них являются обязательными гетеротрофами и могут жить в отсутствие света при условии, что доступен соответствующий источник органического углерода. [ 100 ] [ 101 ]
Фотосинтетические диатомомы, которые оказываются в окружающей среде, отсутствующей кислорода и/или солнечного света, могут переключиться на анаэробное дыхание, известное как дыхание нитрата (DNRA), и оставаться бездействующими в течение месяцев и десятилетий. [ 102 ] [ 103 ]
Основными пигментами диатомов являются хлорофиллы A и C, бета-каротин , фукомсантин , диатоксантин и диадиноксантин. [ 16 ]
Таксономия
[ редактировать ]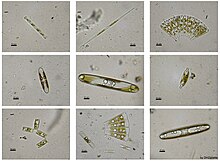



Стефанодиск Ханцша

Истмия Нерерная Нервная
Аурита Одонтелла
Диатомы принадлежат к большой группе протистов, многие из которых содержат пластиды, богатые хлорофиллами a и c. Группу по -разному называли гетероконтами , хризофитами , хромистами или стризенопилами . Многие являются автотрофами , такими как золотые водоросли и водоросли ; и гетеротрофы, такие как водопроводные плесени , опалиниды и актинофрид -гелиозоа. Классификация этой области протистов все еще нерешенная. С точки зрения ранга, с ними рассматривались как разделение , тип , королевство или что -то промежуточное для них. Следовательно, диатомомы ранжируются от класса, обычно называемых Diatomophyceae или Bacillariophyceae , до деления (= phylum), обычно называемого Bacillariophyta , с соответствующими изменениями в рядах их подгрупп.
Роды и виды
[ редактировать ]Предполагается, что существует около 20 000 существующих диатомов видов , из которых около 12 000 были названы на сегодняшний день, согласно Guiry, 2012 [ 104 ] (Другие источники дают более широкий диапазон оценок [ 16 ] [ 105 ] [ 106 ] [ 107 ] ) Было описано около 1000–1300 родов диатома, как существующие, так и ископаемые, [ 108 ] [ 109 ] из которых около 250–300 существует только как окаменелости. [ 110 ]
Классы и заказы
[ редактировать ]В течение многих лет диатомовые данные - обрабатываемые либо классом (Bacillariophyceae), либо тип (Bacillariophyta) - были разделены только на 2 порядка, что соответствует центру и типовым диатомовым лицам ( Centrales и Pennales ). Эта классификация была тщательно пересмотрена в раунде, Кроуфорд и Манн в 1990 году, которые лечили диатомовые данные в более высоком ранге (разделение, соответствующее филому в зоологической классификации) и способствовали основным классовым единицам для классов, сохраняя центрические диатомеры в виде единого класса Coscinodisceae. , но разделение бывших типовых диатомов на 2 отдельных класса, Fragilariophyceae и Bacillariophyceae (последнее более старое имя сохранилось, но с поминенным определением), между ними охватывают 45 заказов, большинство из них новое.
Сегодня (написание в середине 2020 года) признано, что система Round et al. 1990 года. нуждается в пересмотре с появлением более новой молекулярной работы, однако лучшая система для ее замены неясна, а современные системы в широком распространении, такие как водоросли , Всемирный реестр морских видов и ее база данных , и его система , и система для », и система для» Вся жизнь «представлена в Ruggiero et al., 2015, все сохраняют Round et al. Лечение в качестве основы, хотя и с диатомовыми заболеваниями в целом, рассматриваемое как класс, а не как деление/филом, и классы Round et al. Сокращены до подклассов, для лучшего согласия с лечением филогенетически соседних групп и их содержащими таксонами. (Для ссылок ссылаются на отдельные разделы ниже).
Одно из предложений Линды Медлин и коллег, начинающихся в 2004 году, заключается в некоторых из центрических порядков диатомовых условий, которые более тесно связаны с выплатой, которые должны быть разделены как новый класс Mediophyceae, сами по себе более тесно связаны с диатозоми выплаты, чем Остальные центрики. Эта гипотеза-Later обозначала Coscinodiscophyceae-Mediophyceae-bacillariophyceae или Coscinodiscophyceae+(Mediophyceae+Bacillariophyceae) (CMB) гипотеза-был принят DG Mann среди других, которые используют его в качестве основы для классификации диатэмов как в AD. Серия синтезов и др. (2005, 2012, 2019), а также в главе Bacillariophyta Руководства по протистам 2017 года , под редакцией Archibald et al., с некоторыми модификациями, отражающими очевидную немонофилию Medlin et al. Оригинальный "Coscinodiscophyceae". Между тем, группа, возглавляемая EC Theriot, предпочитает другую гипотезу о филогении, которая была названа гипотезой структурной градации (SGH) и не распознает Mediophyceae как монофилетическую группу, в то время как другой анализ Parks et al., 2018,,, обнаруживает, что радиальные центрические диатомовые ткатомы (Coscinodiscophyceae) Medlin et al. Атхей , который является аномальным родом. Обсуждение относительных достоинств этих противоречивых схем продолжается различными вовлеченными сторонами. [ 111 ] [ 112 ] [ 113 ] [ 114 ]
Adl et al., 2019 лечение
[ редактировать ]В 2019 году Adl et al. [ 115 ] представил следующую классификацию диатомов, отмечая: «Этот пересмотр отражает многочисленные достижения в филогении диатомовых заболеваний в течение последнего десятилетия. Из -за нашего бедного отбора проб на таксоне за пределами медиофизии и диатомовых изделий и известного и ожидаемого разнообразия Многие клады появляются на высоком уровне классификации (а классификация более высокого уровня довольно плоская) ». Эта классификация рассматривает диатомовые данные как филом (Diatomeae/Bacillariophyta), принимает класс Mediophyceae of Medlin и коллег, вводит новые субфилы и классы для ряда изолированных родов и переосмысляет ряд ранее установленных таксонов в качестве подкласс, в противном случае, и переосмысляет ряд ранее установленных таксонов, как подкласс, в противном случае изолированные роды и переосмысливают ряд ранее установленных таксонов, как подкласс, в противном случае изолированные роды и перевороты ряда ранее установленных таксонов, и в противном случае изолированные роды и переосмысляют ряд ранее установленных таксонов, а также изолированные роды и переосмысливают ряд ранее установленных таксонов, и в противном случае изолированные роды и переосмысливают ряд ранее установленных таксонов. но не перечисляет заказы или семьи. Предполагаемые ряды были добавлены для ясности (Adl. Et al. Не используйте ранги, но предполагаемые в этой части классификации очевидны из выбора используемых окончаний в рамках используемой системы ботанической номенклатуры).
- Clade Diatomista Derelle et al. 2016, emend. Cavalier-Smith 2017 (diatoms plus a subset of other ochrophyte groups)
- Phylum Diatomeae Dumortier 1821 [= Bacillariophyta Haeckel 1878] (diatoms)
- Subphylum Leptocylindrophytina D.G. Mann in Adl et al. 2019
- Class Leptocylindrophyceae D.G. Mann in Adl et al. 2019 (Leptocylindrus, Tenuicylindrus)
- Class Corethrophyceae D.G. Mann in Adl et al. 2019 (Corethron)
- Subphylum Ellerbeckiophytina D.G. Mann in Adl et al. 2019 (Ellerbeckia)
- Subphylum Probosciophytina D.G. Mann in Adl et al. 2019 (Proboscia)
- Subphylum Melosirophytina D.G. Mann in Adl et al. 2019 (Aulacoseira, Melosira, Hyalodiscus, Stephanopyxis, Paralia, Endictya)
- Subphylum Coscinodiscophytina Medlin & Kaczmarska 2004, emend. (Actinoptychus, Coscinodiscus, Actinocyclus, Asteromphalus, Aulacodiscus, Stellarima)
- Subphylum Rhizosoleniophytina D.G. Mann in Adl et al. 2019 (Guinardia, Rhizosolenia, Pseudosolenia)
- Subphylum Arachnoidiscophytina D.G. Mann in Adl et al. 2019 (Arachnoidiscus)
- Subphylum Bacillariophytina Medlin & Kaczmarska 2004, emend.
- Class Mediophyceae Jouse & Proshkina-Lavrenko in Medlin & Kaczmarska 2004
- Subclass Chaetocerotophycidae Round & R.M. Crawford in Round et al. 1990, emend.
- Subclass Lithodesmiophycidae Round & R.M. Crawford in Round et al. 1990, emend.
- Subclass Thalassiosirophycidae Round & R.M. Crawford in Round et al. 1990
- Subclass Cymatosirophycidae Round & R.M. Crawford in Round et al. 1990
- Subclass Odontellophycidae D.G. Mann in Adl et al. 2019
- Subclass Chrysanthemodiscophycidae D.G. Mann in Adl et al. 2019
- Class Biddulphiophyceae D.G. Mann in Adl et al. 2019
- Subclass Biddulphiophycidae Round and R.M. Crawford in Round et al. 1990, emend.
- Biddulphiophyceae incertae sedis (Attheya)
- Class Bacillariophyceae Haeckel 1878, emend.
- Bacillariophyceae incertae sedis (Striatellaceae)
- Subclass Urneidophycidae Medlin 2016
- Subclass Fragilariophycidae Round in Round, Crawford & Mann 1990, emend.
- Subclass Bacillariophycidae D.G. Mann in Round, Crawford & Mann 1990, emend.
- Class Mediophyceae Jouse & Proshkina-Lavrenko in Medlin & Kaczmarska 2004
- Subphylum Leptocylindrophytina D.G. Mann in Adl et al. 2019
- Phylum Diatomeae Dumortier 1821 [= Bacillariophyta Haeckel 1878] (diatoms)
See taxonomy of diatoms for more details.
Gallery
[edit]- Scanning electron microscope images
-
 Diatom Surirella spiralis
Diatom Surirella spiralis -
 Diatoms Thalassiosira sp. on a membrane filter, pore size 0.4 μm.
Diatoms Thalassiosira sp. on a membrane filter, pore size 0.4 μm. -
 Diatom Paralia sulcata.
Diatom Paralia sulcata. -
 Diatom Achanthes trinodis
Diatom Achanthes trinodis




-
 Stand-alone cell of Bacillaria paxillifer
Stand-alone cell of Bacillaria paxillifer -
 Colonial group of Bacillaria paxillifer
Colonial group of Bacillaria paxillifer


| External videos | |
|---|---|
Three diatom species were sent to the International Space Station, including the huge (6 mm length) diatoms of Antarctica and the exclusive colonial diatom, Bacillaria paradoxa. The cells of Bacillaria moved next to each other in partial but opposite synchrony by a microfluidics method.[116]
Evolution and fossil record
[edit]Origin
[edit]Heterokont chloroplasts appear to derive from those of red algae, rather than directly from prokaryotes as occurred in plants. This suggests they had a more recent origin than many other algae. However, fossil evidence is scant, and only with the evolution of the diatoms themselves do the heterokonts make a serious impression on the fossil record.
Earliest fossils
[edit]The earliest known fossil diatoms date from the early Jurassic (~185 Ma ago),[117] although the molecular clock[117] and sedimentary[118] evidence suggests an earlier origin. It has been suggested that their origin may be related to the end-Permian mass extinction (~250 Ma), after which many marine niches were opened.[119] The gap between this event and the time that fossil diatoms first appear may indicate a period when diatoms were unsilicified and their evolution was cryptic.[120] Since the advent of silicification, diatoms have made a significant impression on the fossil record, with major fossil deposits found as far back as the early Cretaceous, and with some rocks such as diatomaceous earth, being composed almost entirely of them.
Relation to grasslands
[edit]The expansion of grassland biomes and the evolutionary radiation of grasses during the Miocene is believed to have increased the flux of soluble silicon to the oceans, and it has been argued that this promoted the diatoms during the Cenozoic era.[121][122] Recent work suggests that diatom success is decoupled from the evolution of grasses, although both diatom and grassland diversity increased strongly from the middle Miocene.[123]
Relation to climate
[edit]Diatom diversity over the Cenozoic has been very sensitive to global temperature, particularly to the equator-pole temperature gradient. Warmer oceans, particularly warmer polar regions, have in the past been shown to have had substantially lower diatom diversity. Future warm oceans with enhanced polar warming, as projected in global-warming scenarios,[124] could thus in theory result in a significant loss of diatom diversity, although from current knowledge it is impossible to say if this would occur rapidly or only over many tens of thousands of years.[123]
Method of investigation
[edit]The fossil record of diatoms has largely been established through the recovery of their siliceous frustules in marine and non-marine sediments. Although diatoms have both a marine and non-marine stratigraphic record, diatom biostratigraphy, which is based on time-constrained evolutionary originations and extinctions of unique taxa, is only well developed and widely applicable in marine systems. The duration of diatom species ranges have been documented through the study of ocean cores and rock sequences exposed on land.[125] Where diatom biozones are well established and calibrated to the geomagnetic polarity time scale (e.g., Southern Ocean, North Pacific, eastern equatorial Pacific), diatom-based age estimates may be resolved to within <100,000 years, although typical age resolution for Cenozoic diatom assemblages is several hundred thousand years.
Diatoms preserved in lake sediments are widely used for paleoenvironmental reconstructions of Quaternary climate, especially for closed-basin lakes which experience fluctuations in water depth and salinity.
Isotope records
[edit]
When diatoms die their shells (frustules) can settle on the seafloor and become microfossils. Over time, these microfossils become buried as opal deposits in the marine sediment. Paleoclimatology is the study of past climates. Proxy data is used in order to relate elements collected in modern-day sedimentary samples to climatic and oceanic conditions in the past. Paleoclimate proxies refer to preserved or fossilized physical markers which serve as substitutes for direct meteorological or ocean measurements.[126] An example of proxies is the use of diatom isotope records of δ13C, δ18O, δ30Si (δ13Cdiatom, δ18Odiatom, and δ30Sidiatom). In 2015, Swann and Snelling used these isotope records to document historic changes in the photic zone conditions of the north-west Pacific Ocean, including nutrient supply and the efficiency of the soft-tissue biological pump, from the modern day back to marine isotope stage 5e, which coincides with the last interglacial period. Peaks in opal productivity in the marine isotope stage are associated with the breakdown of the regional halocline stratification and increased nutrient supply to the photic zone.[127]

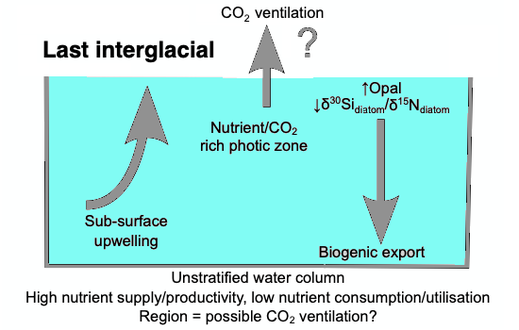
The initial development of the halocline and stratified water column has been attributed to the onset of major Northern Hemisphere glaciation at 2.73 Ma, which increased the flux of freshwater to the region, via increased monsoonal rainfall and/or glacial meltwater, and sea surface temperatures.[128][129][130][131] The decrease of abyssal water upwelling associated with this may have contributed to the establishment of globally cooler conditions and the expansion of glaciers across the Northern Hemisphere from 2.73 Ma.[129] While the halocline appears to have prevailed through the late Pliocene and early Quaternary glacial–interglacial cycles,[132] other studies have shown that the stratification boundary may have broken down in the late Quaternary at glacial terminations and during the early part of interglacials.[133][134][135][136][137][127]
Diversification
[edit]The Cretaceous record of diatoms is limited, but recent studies reveal a progressive diversification of diatom types. The Cretaceous–Paleogene extinction event, which in the oceans dramatically affected organisms with calcareous skeletons, appears to have had relatively little impact on diatom evolution.[138]
Turnover
[edit]Although no mass extinctions of marine diatoms have been observed during the Cenozoic, times of relatively rapid evolutionary turnover in marine diatom species assemblages occurred near the Paleocene–Eocene boundary,[139] and at the Eocene–Oligocene boundary.[140] Further turnover of assemblages took place at various times between the middle Miocene and late Pliocene,[141] in response to progressive cooling of polar regions and the development of more endemic diatom assemblages.
A global trend toward more delicate diatom frustules has been noted from the Oligocene to the Quaternary.[125] This coincides with an increasingly more vigorous circulation of the ocean's surface and deep waters brought about by increasing latitudinal thermal gradients at the onset of major ice sheet expansion on Antarctica and progressive cooling through the Neogene and Quaternary towards a bipolar glaciated world. This caused diatoms to take in less silica for the formation of their frustules. Increased mixing of the oceans renews silica and other nutrients necessary for diatom growth in surface waters, especially in regions of coastal and oceanic upwelling.
Genetics
[edit]
Expressed sequence tagging
[edit]В 2002 году первое понимание свойств репертуара гена PhaeOdactylum Tricornutum была описана с использованием 1000 экспрессируемых метков последовательности (ESTS). [142] Subsequently, the number of ESTs was extended to 12,000 and the diatom EST database was constructed for functional analyses.[143] Эти последовательности были использованы для проведения сравнительного анализа между P. tricornutum и предполагаемыми полными протеомами из зеленой водоросли Chlamydomonas inerhardtii , красной водоросли Cyanidoschyzon Merolae и диатомовой талассиозира псевдонана . [ 144 ] База данных Diatom EST в настоящее время состоит из более чем 200 000 EST из клеток P. tricornutum (16 библиотек) и T. pseudonana (7 библиотек), выращенных в различных условиях, многие из которых соответствуют различным абиотическим стрессам. [ 145 ]
Секвенирование генома
[ редактировать ]
весь геном центрического диатомо . В 2004 году был секвенирован [ 146 ] Следуя в 2008 году с секвенированием Diatom Pennate, Phaeodactylum tricornutum (27,4 МБ). [ 147 ] Сравнения этих двух показывают, что геном P. tricornutum включает в себя меньше генов (10 402 против 11 776), чем Т. Псевдонана ; Не может быть обнаружено никакой основной синтенции (ген -порядок) между двумя геномами. Гены T. pseudonana показывают в среднем ~ 1,52 интрона на ген, в отличие от 0,79 у P. tricornutum , что предполагает недавнее широкое усиление интрона в центрике диатомовой. [ 147 ] [ 148 ] Несмотря на относительно недавнюю эволюционную дивергенцию (90 миллионов лет), степень молекулярной дивергенции между центриками и трудами указывает на быстрой эволюционной скорости в бациллариофике по сравнению с другими эукариотическими группами. [ 147 ] Сравнительная геномика также установила, что специфический класс транспонируемых элементов , диатомоподобные ретротранспозоны (или Codis), был значительно амплифицирован в геноме P. tricornutum по отношению к T. pseudonana , составляющим 5,8 и 1% соответствующих геномов. [ 149 ]
Эндосимбиотический перенос гена
[ редактировать ]Diatom Genomics предоставила много информации о степени и динамике процесса переноса эндосимбиотического гена (EGT). Сравнение белков T. pseudonana с гомологами в других организмах показало, что сотни имеют свои самые близкие гомологи в линии Plantae. EGT к геномам диатомовых геномов можно проиллюстрировать тем фактом, что геном T. pseudonana кодирует шесть белков, которые наиболее тесно связаны с генами, кодируемыми Guillardia Theta ( Cryptomonad ) геномом нуклеоморфа . Четыре из этих генов также обнаружены в геномах красных водорослей пластид, демонстрируя, тем самым демонстрируя последовательные EGT от пластида красного водорослевого ядра до красного водоросля (нуклеоморф) до ядра хозяина гетероконта. [ 146 ] Более поздние филогеномные анализы протеомов диатомовых протеомов предоставили доказательства для празинофитного эндосимбионта в общем предке хромальвеолатов , подтверждаемых тем, что 70% генов диатомовых источников происхождения зеленых линий и то, что такие гены также обнаруживаются в геноме. других стрименопилов . Следовательно, было предложено, что хромальвеолаты являются продуктом серийного вторичного эндосимбиоза , сначала с зелеными водорослями , за которыми следуют вторые с красными водорослями , которые сохранили геномные следы предыдущего, но смещали зеленый пластид. [ 150 ] Тем не менее, филогеномный анализ протеомов диатомовых протеомов и эволюционной истории хромальвевелера, вероятно, будет воспользоваться дополнительными геномными данными из недостаточных линий, таких как красные водоросли.
Горизонтальный перенос генов
[ редактировать ]В дополнение к EGT, горизонтальный перенос генов (HGT) может происходить независимо от эндосимбиотического события. Публикация генома P. tricornutum сообщила, что по меньшей мере 587 генов P. tricornutum , по -видимому, наиболее тесно связаны с бактериальными генами, что составляет более 5% протеома P. tricornutum . Около половины из них также встречаются в геноме T. pseudonana , подтверждая их древнее включение в линию диатомовой линии. [ 147 ]
Генетическая инженерия
[ редактировать ]Чтобы понять биологические механизмы, которые лежат в основе большой важности диатомовых средств в геохимических циклах, ученые использовали Phaeodactylum Tricornutum и Thalassiosira Spp. виды как модельные организмы с 90 -х годов. [ 151 ] В настоящее время доступно несколько инструментов молекулярной биологии для генерации мутантов или трансгенных линий: плазмиды, содержащие трансгены, вставляются в клетки с использованием биолистического метода [ 152 ] или конъюгация бактерий трансмиссии [ 153 ] (с 10 −6 и 10 −4 урожай соответственно [ 152 ] [ 153 ] ) и другие классические методы трансфекции, такие как электропорация или использование ПЭГ, дают результаты с более низкой эффективностью. [ 153 ]
Трансфицированные плазмиды могут быть либо случайным образом интегрированы в хромосомы диатома, либо поддерживаются в качестве стабильных круговых эпизодов (благодаря центромерной последовательности CEN6-Arsh4-His3. [ 153 ] ) Ген устойчивости к флеомицину/ зооцинам обычно используется в качестве маркера отбора, [ 151 ] [ 154 ] и различные трансгены были успешно введены и экспрессируются в диатомовых условиях со стабильными передачами через поколения, [ 153 ] [ 154 ] или с возможностью удалить его. [ 154 ]
Кроме того, эти системы в настоящее время позволяют использовать инструмент Crispr-Cas Genome Edition , что приводит к быстрому производству функциональных нокаутированных мутантов [ 154 ] [ 155 ] и более точное понимание клеточных процессов диатомов.
Человеческое использование
[ редактировать ]-
 Диатоматическая Земля, состоящая из центрических (радиально симметричных) и типичных (двусторонних симметричных) диатомов, подвешенных в воде.
Диатоматическая Земля, состоящая из центрических (радиально симметричных) и типичных (двусторонних симметричных) диатомов, подвешенных в воде.
(Нажмите 3 раза, чтобы полностью увеличить)

Палеонтология
[ редактировать ]Разложение и распад диатомодов приводит к органическому и неорганическому (в форме силикатов ) отложений, неорганический компонент которого может привести к методу анализа прошлых морских средах океанских по этажах или бухте , так как неорганическая вещество вкладывается в Осаждение глин и илтов и образует постоянную геологическую запись таких морских слоев (см. Силисенскую слизную ).
Промышленное
[ редактировать ]Диатомовые средства и их раковины (домочители) в качестве диатомита или диатомовой земли являются важными промышленными ресурсами, используемыми для тонкой полировки и жидкой фильтрации. Сложная структура их микроскопических оболочек была предложена в качестве материала для нанотехнологий. [ 156 ]
Диатомит считается натуральным нано -материалом и имеет много применений и применений, таких как: производство различных керамических продуктов, строительная керамика, рефрактерная керамика, специальная керамика оксида, для производства материалов контроля влажности, используемых в качестве фильтрационного материала, материал в цементе Производственная промышленность, первоначальный материал для производства перевозчиков лекарств с длительным высвобождением, поглощать материал в промышленном масштабе, производство пористой керамики, стеклянная промышленность, используется в качестве поддержки катализатора, в качестве наполнителя в пластиках и красках, очистка промышленных вод, держателя пестицидов, а также для улучшения физических и химических характеристик определенных почв и других целей. [ 157 ] [ 158 ] [ 159 ]
Диатомы также используются для определения происхождения их содержащих материалов, включая морскую воду.
Нанотехнология
[ редактировать ]Осаждение кремнезема по диатомовым вопросам также может оказаться полезным для нанотехнологий . [ 160 ] Диатомовые ячейки неоднократно и надежно изготовляют клапаны различных форм и размеров, потенциально позволяя диатомовым процессам производить микро- или наномасштабные структуры, которые могут использоваться в ряде устройств, включая: оптические системы; полупроводниковая нанолитография ; и даже транспортные средства для доставки наркотиков . При подходящей процедуре искусственного отбора диатомовые средства, которые производят клапаны с определенными формами и размерами, могут быть разработаны для выращивания в хемостата культурах для наноразмерных компонентов массового производства . [ 161 ] Также было предложено, чтобы диатомовые средства могли использоваться в качестве компонента солнечных элементов, заменяя фоточувствительный диоксид титана на диоксид кремния, который обычно используют для создания своих клеточных стен. [ 162 ] Также были предложены солнечные батареи, продуцирующие диатомовые биотопливы. [ 163 ]
- Поддержка и регулирование услуг, предоставляемых морскими диатомогами и некоторыми из их негативных воздействий
-
 CNN = облачные конденсации ядра , DMS = диметилсульфид , DMSP = диметилсульфониопропионат , VOCS = летучие органические соединения
CNN = облачные конденсации ядра , DMS = диметилсульфид , DMSP = диметилсульфониопропионат , VOCS = летучие органические соединения
Пунктирная стрелка: негативное эффект, твердая стрелка: положительные эффекты

Судебно -медицинская экспертиза
[ редактировать ]Основная цель анализа диатомовой криминалистики -дифференцировать смерть от погружения от посмертного погружения тела в воду. Лабораторные испытания могут выявить присутствие диатомовых лиц в организме. Поскольку скелеты на основе кремнезема не легко разлагаются, их иногда можно обнаружить даже в сильно разложенных телах. Поскольку они не встречаются естественным образом в организме, если лабораторные испытания показывают диатомы в трупе, которые имеют тот же вид, что и в воде, где было извлечено тело, то это может быть хорошим доказательством утопления как причины смерти . Смесь видов диатомов, найденных в трупе, может быть одинаковой или отличной от окружающей воды, что указывает, утонула ли жертва в том же месте, в котором было найдено тело. [ 164 ]
История открытия
[ редактировать ]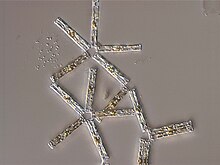
Первые иллюстрации диатомов обнаружены в статье 1703 года в сделках Королевского общества, показывающие безошибочные рисунки Tabellaria . [ 165 ] Хотя публикация была написана неназванным английским джентльменом, есть недавние доказательства того, что он был Чарльзом Кингом Стаффордшира. [ 165 ] [ 166 ] Спустя всего 80 лет мы находим первую формально идентифицированную Diatom, колониальную Bacillaria paxillifera , обнаруженную и описанную в 1783 году датским натуралистом Отто Фридрихом Мюллером . [ 165 ] Как и многие другие после него, он ошибочно думал, что это животное из -за его способности двигаться. Даже Чарльз Дарвин видел, как Диатом остается в пыли, пока на островах Кейп -Верде, хотя он не был уверен, что они были. Только позже они были идентифицированы для него как кремнистыми полигастриками. Инфузория, которую Дарвин позже отметил на лице, краски Fueguinos, местных жителей Тирра -дель -Фуэго в южной части Южной Америки, были позже идентифицированы таким же образом. В течение своей жизни кремнистые полигастрики были уточнены как принадлежащие Diatomaceae , и Дарвин изо всех сил пытался понять причины, лежащие в основе их красоты. Он обменялся мнениями с известным криптогамическим GHK Thwaites по теме. В четвертом издании «Происхождение видов» он заявил, что « мало объектов красивее, чем минутные кремнистые случаи Diatomaceae: они были созданы, что их можно было исследовать и восхищаться под высокими силами микроскопа »? и рассуждал, что их изысканная морфология должна иметь функциональные основы, а не быть созданным исключительно для того, чтобы люди восхищались. [ 167 ]
Смотрите также
[ редактировать ]- Высоковеченные изопреноидные , длинноцепочечные алкены, продуцируемые небольшим количеством морских диатомов
Примечания
[ редактировать ]- ^ От греческого : Διατομή , романизированный : Diatomé , «прорезание, выходное пособие», [ 7 ] от греческого : Διάτομος , романизированный : диатомос , "вырезан пополам, разделен одинаково" [ 8 ] От греческого : Διατέμνω , романизированный : DiaTémno , «чтобы вырезать в двенах». [ 9 ] [ 10 ] : 718
Ссылки
[ редактировать ]- ^ Dangeard, P. (1933). Traite d'algologie. Пол Лечвалье и Филс, Париж, [1] Архивировал 4 октября 2015 года на машине Wayback .
- ^ "Bacillariophyceae" . Черви . Мировой реестр морских видов . 2024 . Получено 9 мая 2024 года .
- ^ Воскресенье, Б.-С. (1822). Ботанические комментарии. Ботанические наблюдения, посвященные Обществу садоводства турниров (PDF ) Турней: гл. Кастерман-бог, Бридж-стрит № 10. стр. , [1, таблица, ошибка. i [ ], [1] -116
- ^ Рабенхорст, Л. Флора Европейская слой воды и сладкого и подводного лодка (1864-1868). Раздел I. Алгас Диатомацес, состоящий из фигур всего Impressis (1864). стр. 1-359. Нью -Йорк [Лейпциг]: в Эдуарде Куммеруме.
- ^ Haeckel, E. (1878). DAS Protistenreich Archived 10 ноября 2014 года на The Wayback Machine .
- ^ Engler, A. & Gilg, E. (1919). Программа семейств растений: обзор всей системы растений с особым рассмотрением медицинских и полезных растений, в дополнение к обзору цветочных и цветочных районов Земли для использования на лекциях и исследованиях по специальной и медицинской фармацевтической ботанике , 8 -е изд.
- ^ Διατομή . Лидделл, Генри Джордж ; Скотт, Роберт ; Грек -английский лексикон в проекте Персея
- ^ Διάτομος . Лидделл, Генри Джордж ; Скотт, Роберт ; Грек -английский лексикон в проекте Персея
- ^ Διατέμνω . Лидделл, Генри Джордж ; Скотт, Роберт ; Грек -английский лексикон в проекте Персея
- ^ Компактный Оксфордский английский словарь . Кларендон Пресс. 1971. ISBN 0918414083 .
- ^ «Воздух, который ты дышишь? Диатом сделал это» . Живая наука . 11 июня 2014 года. Архивировано с оригинала 30 апреля 2018 года . Получено 30 апреля 2018 года .
- ^ "Что такое диатомовые?" Полем Диатомоты Северной Америки. Архивировано из оригинала 25 января 2020 года . Получено 28 января 2020 года .
- ^ Treguer, P.; Нельсон, DM; Ван Беннеком, AJ; Демастер, диджей; Leynaert, A.; Queguiner, B. (1995). «Баланс кремнезема в мировом океане: рецензия». Наука . 268 (5209): 375–9. Bibcode : 1995sci ... 268..375t . doi : 10.1126/science.268.5209.375 . PMID 17746543 . S2CID 5672525 .
- ^ «Королевский колледж Лондон - Лейк Мегачад» . www.kcl.ac.uk. Архивировано с оригинала 27 ноября 2018 года . Получено 5 мая 2018 года .
- ^ Бристоу, CS; Хадсон-Эдвардс, Ка ; Чаппелл А. (2010). «Оборудование Амазонки и экваториальной Атлантики с западноафриканской пылью». Геофий. Резерв Летал 37 (14): L14807. Bibcode : 2010georl..3714807b . doi : 10.1029/2010gl043486 . S2CID 128466273 .
- ^ Jump up to: а беременный в дюймовый и фон HASLE, Grete R.; Syvertsen, Erik E.; Steidinger, Karen A.; Танген, Карл (25 января 1996 г.). «Морские диатомовые» . В Томасе, Кармело Р. (ред.). Идентификация морских диатомовых и динофлагеллятов . Академическая пресса. С. 5–385. ISBN 978-0-08-053441-1 Полем Архивировано из оригинала 3 января 2014 года . Получено 13 ноября 2013 года .
- ^ «Газовые гззлеры» . Архивировано из оригинала 22 мая 2018 года . Получено 22 мая 2018 года .
- ^ «Больше на диатомовых условиях» . Калифорнийский музей палеонтологии . Архивировано с оригинала 4 октября 2012 года . Получено 20 мая 2015 года .
- ^ Van The Hoek, C.; Человек, DG; Jahns, HM (1995). Водоросли: и введение в Phyclooly . Кембридж: издательство Кембриджского университета . стр. 165-218. ISBN 978-0-521-31687-3 .
- ^ Накаяма, Т.; Камикава, Р.; Tanifuji, G.; Kashiyama, Y.; Ohkouchi, N.; Арчибальд, JM; Инагаки, Ю. (2014). «Полный геном нефотосинтетического цианобактериума в диатоме выявляет недавние адаптации к внутриклеточному образу жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (31): 11407–11412. BIBCODE : 2014PNAS..11111407N . doi : 10.1073/pnas.1405222111 . PMC 4128115 . PMID 25049384 .
- ^ Пьелла Карлусич, Хуан Хосе; Пеллетье, Эрик; Ломбард, Фабен; Carsique, Madeline; Дворак, Этьен; Колин, Себастьен; Пичерал, Марк; Корнехо-Кастильо, Франциско М.; Акинас, Сильвия Г.; Пепперкок, Рейнер; Карсанти, Эрик (6 июля 2021 года). «Глобальные модели распределения морских азотных фиксеров с помощью визуализации и молекулярных методов» . Природная связь . 12 (1): 4160. Bibcode : 2021Natco..12.4160p . Doi : 10.1038/s41467-021-24299-y . ISSN 2041-1723 . PMC 8260585 . PMID 34230473 .
- ^ Функциональная связь между динофлагеллятным хозяином и его диатомовым эндосимбионтом | Молекулярная биология и эволюция | Оксфордский академический
- ^ Wehr, JD; Оболочка, RG; Kociolek, JP, eds. (2015). Пресноводные водоросли Северной Америки: экология и классификация (2 -е изд.). Сан -Диего: академическая пресса. ISBN 978-0-12-385876-4 .
- ^ Жирар, Винсент; Святой Мартин, Симона; Buffetout, Eric; Святой Мартин, Жан-Поль; Néraudeau, Didier; Пейро, Даниэль; Костры, Гвидо; Ребята, Евгенио; Suteethorn, Varavudh (2020). "Тайский янтарь: понимание истории ранней диатомы?" Полем BSGF - Бюллетень Земли . 191 : 23. doi : 10.1051/bsgf/2020028 . ISSN 1777-5817 .
- ^ Внутреннее пространство Субарктического Тихого океана Архивировано 27 октября 2020 года в машине Wayback Nasa Earth Expeditions , 4 сентября 2018 года.
 Эта статья включает текст из этого источника, который находится в общественном доступе .
Эта статья включает текст из этого источника, который находится в общественном доступе .
- ^ Rousseaux, Cecile s.; Грегг, Уотсон В. (2015). «Недавние декадальные тенденции в глобальном композиции фитопланктона» . Глобальные биогеохимические циклы . 29 (10): 1674–1688. Bibcode : 2015gbioc..29.1674R . doi : 10.1002/2015GB005139 .
- ^ Colin, S., Coelho, LP, Sunagawa, S., Bowler, C., Karsenti, E., Bork, P., Pepperkok, R. and de Vargas, C. (2017) "Количественное 3D-изображение для клетки Биология и экология экологических микробных эукариот ». Elife , 6 : E26066. два : 10.7554/elife.26066.002 .
 Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивная 16 октября 2017 года на машине Wayback .
Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивная 16 октября 2017 года на машине Wayback .
- ^ Нельсон, Дэвид М.; Трегер, Пол; Brzezinski, Mark A.; Лейнаерт, Ауде; Quéguiner, Bernard (1995). «Производство и растворение биогенного кремнезема в океане: пересмотренные глобальные оценки, сравнение с региональными данными и связь с биогенным седиментацией». Глобальные биогеохимические циклы . 9 (3). Американский геофизический союз (AGU): 359–372. Bibcode : 1995gbioc ... 9..359n . doi : 10.1029/95GB01070 . ISSN 0886-6236 .
- ^ Манн, Дэвид Г. (1999). «Концепция вида в диатомовых заболеваниях». Phycologia . 38 (6). Informa UK Limited: 437–495. Bibcode : 1999phyco..38..437m . doi : 10.2216/i0031-8884-38-6-437.1 . ISSN 0031-8884 .
- ^ Simonsen, R., (1979). «Система диатома: идеи о филогении», Bacillaria , 2 : 9–71.
- ^ Jump up to: а беременный Мур, Эрик Р.; Буллингтон, Бриана С.; Вайсберг, Александра Дж.; Цзян, Юань; Чанг, Джефф; Хэлси, Кимберли Х. (7 июля 2017 г.). «Морфологические и транскриптомные доказательства индукции аммония сексуального размножения в талассиозире псевдонана и других центричных диатомовых заболеваний» . Plos один . 12 (7). Публичная библиотека науки (PLOS): E0181098. BIBCODE : 2017PLOSO..1281098M . doi : 10.1371/journal.pone.0181098 . ISSN 1932-6203 . PMC 5501676 . PMID 28686696 . Модифицированный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Университетский колледж Лондон ( 2002 ) .
- ^ Паркер, Эндрю Р.; Таунли, Хелен Э. (2007). «Биомиметика фотонных наноструктур». Природная нанотехнология . 2 (6): 347–53. Bibcode : 2007natna ... 2..347p . doi : 10.1038/nnano.2007.152 . PMID 18654305 .
- ^ Гордон, Ричард; Лосик, Дусан; Тиффани, Мэри Энн; Надь, Стивен С.; Стерренбург, Фритжоф А.С. (2009). «Стеклянный зверинец: диатомовые средства для новых применений в нанотехнологиях». Тенденции в биотехнологии . 27 (2): 116–27. doi : 10.1016/j.tibtech.2008.11.003 . PMID 19167770 .
- ^ Jump up to: а беременный в дюймовый Рита А. Хорнер (2002). Таксономическое руководство по некоторому общему морскому фитопланктону . Биопресс. С. 25–30. ISBN 978-0-948737-65-7 Полем Архивировано из оригинала 1 августа 2020 года . Получено 13 ноября 2013 года .
- ^ «Стекло в природе» . Музей стекла Корнинг. Архивировано из оригинала 7 марта 2013 года . Получено 19 февраля 2013 года .
- ^ Taylor, JC, Harding, WR и Archibald, C. (2007). Иллюстрированное руководство по некоторым общим видам диатомовых видов из Южной Африки . Гезина: Комиссия по исследованию воды. ISBN 97817770054844 .
- ^ Мишра, М., Аруха, А.П., Башир Т., Ядав Д. и Прасад, Гбкс (2017) «Все новые лица диатомов: потенциальный источник наноматериалов и за его пределами». Границы в микробиологии , 8 : 1239. Два : 10.3389/fmicb.2017.01239 . Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивная 16 октября 2017 года на машине Wayback .
- ^ Jump up to: а беременный Чжан, Д.; Wang, y.; Cai, J.; Пан, Дж.; Цзян, х.; Цзян, Ю. (2012). «Технология биопроизводства на основе диатомовой микро- и наноструктуры» . Китайский научный бюллетень . 57 (30): 3836–3849. Bibcode : 2012Chsbu..57.3836z . doi : 10.1007/s11434-012-5410-x .
- ^ «Диатомовые» . Архивировано с оригинала 2 февраля 2016 года . Получено 13 февраля 2016 года .
- ^ Молекулярная жизнь диатомовых
- ^ Килиас, Эстель С.; Юнгс, Леандро; Шпраха, Лука; Леонард, парень; Metfies, Katja; Ричардс, Томас А. (2020). «Распределение грибов и совместное распространение грибов с диатомогами коррелирует с таянием морского льда в арктическом океане» . Биология связи . 3 (1): 183. doi : 10.1038/s42003-020-0891-7 . PMC 7174370 . PMID 32317738 . S2CID 216033140 .
- ^ Jump up to: а беременный в Thamatrakoln, K.; Алверсон, AJ; Хильдебранд М. (2006). «Сравнительные анализы последовательности транспортеров диатомовых кремния: к механистической модели транспорта кремния». Журнал Phycology . 42 (4): 822–834. Bibcode : 2006jpcgy..42..822t . doi : 10.1111/j.1529-8817.2006.00233.x . S2CID 86674657 .
- ^ Kröger, Nils; Дойцманн, Рейнер; Манфред, Сампер (ноябрь 1999 г.). «Поликационные пептиды из диатомовой биосилики, которые направляют образование наносферы кремнезема». Наука . 286 (5442): 1129–1132. doi : 10.1126/science.286.5442.1129 . PMID 10550045 . S2CID 10925689 . [ Постоянная мертвая ссылка ]
- ^ Kroger, Nils (2007). Справочник по биоминерализации: биологические аспекты и образование структуры . Вайнхайм, Германия: Wiley-VCH Verlag Gmbh. с. Глава 3.
- ^ Аноним (1702). «Два письма от джентльмена в стране, касающиеся письма г -на Левенхука в сделке, № 283.», Филос. Транс. R. Soc. Лонд B , 23 : 1494–1501.
- ^ Rogall, E. (1939). « О тонкой конструкции кремнезема мембраны диатомов» , Planta : 279-291.
- ^ Jump up to: а беременный в Хильдебранд, Марк; Лерч, Сара Дж.Л.; Шрестха, Рошан П. (11 апреля 2018 г.). «Понимание силицификации клеточной стенки диатомовой стенки - продвижение вперед» . Границы в морской науке . 5 Frontiers Media SA. doi : 10.3389/fmars.2018.00125 . ISSN 2296-7745 . Модифицированный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хильдебранд, Марк; Вулкани, Бенджамин Э.; Гассманн, Уолтер; Шредер, Джулиан И. (1997). «Семейство генов кремниевых переносчиков». Природа . 385 (6618). Springer Science and Business Media LLC: 688–689. Bibcode : 1997natur.385..688h . doi : 10.1038/385688b0 . ISSN 0028-0836 . PMID 9034185 . S2CID 4266966 .
- ^ Kröger, Nils; Дойцманн, Рейнер; Сампер, Манфред (5 ноября 1999 г.). «Поликационные пептиды из диатомовой биосилики, которые направляют образование наносферы кремнезема». Наука . 286 (5442). Американская ассоциация по развитию науки (AAAS): 1129–1132. doi : 10.1126/science.286.5442.1129 . ISSN 0036-8075 . PMID 10550045 .
- ^ Kröger, Nils; Дойцманн, Рейнер; Бергсдорф, Кристиан; Сампер, Манфред (5 декабря 2000 г.). «Виды-специфические полиамины из диатомовых контрольных морфологии кремнезема» . Труды Национальной академии наук . 97 (26): 14133–14138. Bibcode : 2000pnas ... 9714133K . doi : 10.1073/pnas.260496497 . ISSN 0027-8424 . PMC 18883 . PMID 11106386 .
- ^ Тессон, Бенуа; Хильдебранд, Марк (10 декабря 2010 г.). «Обширная и интимная ассоциация цитоскелета с образованием кремнезема в диатомовых условиях: контроль над паттерном на мезо- и микромасштабах» . Plos один . 5 (12). Публичная библиотека науки (PLOS): E14300. BIBCODE : 2010PLOSO ... 514300T . doi : 10.1371/journal.pone.0014300 . ISSN 1932-6203 . PMC 3000822 . PMID 21200414 .
- ^ Тессон, Бенуа; Хильдебранд, Марк (23 апреля 2013 г.). «Характеристика и локализация нерастворимых органических матриц, связанных с диатомовыми клеточными стенками: понимание их ролей во время образования клеточной стенки» . Plos один . 8 (4). Публичная библиотека науки (PLOS): E61675. BIBCODE : 2013PLOSO ... 861675T . doi : 10.1371/journal.pone.0061675 . ISSN 1932-6203 . PMC 3633991 . PMID 23626714 .
- ^ Шеффель, Андре; Поулсен, Николь; Шиан, Самуил; Крёгер, Нильс (7 февраля 2011 г.). «Нанопатер -белковые микророры от диатомовой, которая направляет морфогенез кремнезема» . Труды Национальной академии наук . 108 (8): 3175–3180. doi : 10.1073/pnas.1012842108 . ISSN 0027-8424 . PMC 3044418 . PMID 21300899 .
- ^ Коцш, Александр; Грёгер, Филипп; Паволски, Дамиан; Боманс, Пол Х.Х.; Sommerdijk, Nico Ajm; Шлиерф, Майкл; Kröger, Nils (24 июля 2017 г.). «Силиканин-1-это консервативный белок диатомовой мембраны, участвующий в биоминерализации кремнезема» . BMC Biology . 15 (1). Springer Science and Business Media LLC: 65. DOI : 10.1186/S12915-017-0400-8 . ISSN 1741-7007 . PMC 5525289 . PMID 28738898 .
- ^ Тессон, Бенуа; Лерч, Сара Дж.Л.; Хильдебранд, Марк (18 октября 2017 года). «Характеристика нового семейства белков, связанного с мембраной пузырьков кремнезема, обеспечивает генетические манипуляции с диатомовым кремнеземом» . Научные отчеты . 7 (1). Springer Science and Business Media LLC: 13457. Bibcode : 2017natsr ... 713457t . doi : 10.1038/s41598-017-13613-8 . ISSN 2045-2322 . PMC 5647440 . PMID 29044150 .
- ^ Падисак, Джудит; Soróczki-Pintér, Eva; Резнер, Zsuzsanna (2003), Martens, Koen (ed.), «Тонирующие свойства некоторых фитопланктонских форм и связь формы устойчивости к морфологическому разнообразию планктона - экспериментальное исследование» (PDF) , водное биодисность: праздничный объем в честь. Анри Дж. Дюмон , разработки гидробиологии, Springer Netherlands, с. 243–257, doi : 10.1007/978-94-007-1084-9_18 , ISBN 9789400710849 , архивировано из оригинала (PDF) 23 июля 2018 года , получено 4 октября 2019 года
- ^ Андерсон, Ларс WJ; Суини, Беатрис М. (1 мая 1977 г.). «Диэль изменений в характеристиках седиментации Ditylum Brightwelli: изменения клеточных липидов и эффекты ингибиторов дыхания и модификаторов ионного транспорта1» . Лимнология и океанография . 22 (3): 539–552. Bibcode : 1977 Limoc..22..539a . doi : 10.4319/lo.1977.22.3.0539 . ISSN 1939-5590 .
- ^ Поулсен, Николь С.; Спектор, Илан; Сперк, Тимоти П.; Шульц, Томас Ф.; Wetherbee, Richard (1 сентября 1999 г.). «Diatom Glinding является результатом системы подвижности актин-миозина». Клеточная подвижность и цитоскелет . 44 (1): 23–33. doi : 10.1002/(SICI) 1097-0169 (199909) 44: 1 <23 :: AID-CM2> 3.0.CO; 2-D . ISSN 1097-0169 . PMID 10470016 .
- ^ Манн, Дэвид Г. (февраль 2010 г.). «Рафидные диатомовые» . Веб -проект «Дерево жизни» . Архивировано с оригинала 27 сентября 2019 года . Получено 27 сентября 2019 года .
- ^ Jump up to: а беременный Г. Дребс (1 января 1977 г.). «Глава 9: Сексуальность» . В Дитрих Вернер (ред.). Биология диатомовых заболеваний . Ботанические монографии. Тол. 13. Университет Калифорнийского университета. С. 250–283. ISBN 978-0-520-03400-6 Полем Архивировано из оригинала 1 августа 2020 года . Получено 14 ноября 2013 года .
- ^ Lewis, Wm, Jr. (1984). «Половые часы Diatom и его эволюционное значение». Американский натуралист , 123 (1): 73–80
- ^ Chepurnov VA, Mann DG, Sabbe K, Vyverman W. (2004). «Экспериментальные исследования по сексуальному размножению в диатомовых условиях». В Чон К.В., изд. Обзор клеточной биологии . Международный обзор цитологии 237 : 91–154. Лондон
- ^ Дребс (1977) "Сексуальность". В: Werner D, ed. Биология диатомов , ботанические монографии 13 250–283. Оксфорд: Blackwell Scientific Publications.
- ^ «Сокращение размера, репродуктивная стратегия и жизненный цикл центрикального диатомора». Философские транзакции Королевского общества Лондона. Серия B: Биологические науки . 336 (1277). Королевское общество: 191–213. 29 мая 1992 г. DOI : 10.1098/rstb.1992.0056 . ISSN 0962-8436 . S2CID 86332060 .
- ^ Кестер, Джули А.; Brawley, Susan H.; Карп-Босс, Ли; Манн, Дэвид Г. (2007). «Сексуальное воспроизведение в морском центре Diatom Ditylum Brightwellii (Bacillariophyta)» . Европейский журнал Phycology . 42 (4). Informa UK Limited: 351–366. Bibcode : 2007ejphy..42..351k . doi : 10.1080/09670260701562100 . ISSN 0967-0262 . S2CID 80737380 .
- ^ Jump up to: а беременный Mouget JL, Gastineau R, Davidovich O, Gaudin P, Davidovich NA. Свет является ключевым фактором, вызванным сексуальным размножением в Pennate Diatom Haslea Ostrearia. FEMS Microbiol Ecol. 2009 август; 69 (2): 194-201. doi: 10.1111/j.1574-6941.2009.00700.x. Epub 2009 мая. PMID 19486155
- ^ SCCALCO E, STEC K, IUDCONE D, Ferrant MI, Montresor M. Динамика сексуального переворачивания в морской диатоме Pseudo-Nitzschia multistiatta (Bacillaryphyceae). J Phycol. 2014 октябрь; 50 (5): 817-2 Doi: 10.1111/jpy.1 Epub 2014 14 сентября. PMID 26988637
- ^ Jump up to: а беременный Патил С., Моис С., фон Дассоу П., Хуисман М.Дж., Мэйплсон Д., Де Вейлдер Л., Сангес Р., Вюверман В., Монтрезор М., Ферранте М.И. Идентификация мейотического инструментария в диатомовых условиях и исследования гомологов SPO11 и RAD51 в сексуальных видах Pseudo-Nitzschia Multistriata и Seminavis Robusta. BMC Genomics. 2015 14 ноября; 16: 930. doi: 10.1186/s12864-015-1983-5. Ошибка в: BMC Genomics. 2019 июля 5; 20 (1): 544. PMID 26572248; PMCID: PMC4647503
- ^ Азам, Фарук; Бидл, Кей Д. (1999). «Ускоренное растворение диатомового кремнезема морскими бактериальными сообщениями». Природа . 397 (6719): 508–12. Bibcode : 1999natur.397..508b . doi : 10.1038/17351 . S2CID 4397909 . INIST 1755031 .
- ^ Zakharova, Yulia R.; Galachyants, Yuri P.; Kurilkina, Maria I.; Likhoshvay, Alexander V.; Petrova, Darya P.; Shishlyannikov, Sergey M.; Ravin, Nikolai V.; Mardanov, Andrey V.; Beletsky, Alexey V.; Likhoshway, Yelena V. (2013). "The Structure of Microbial Community and Degradation of Diatoms in the Deep Near-Bottom Layer of Lake Baikal" . PLOS ONE . 8 (4): e59977. Bibcode : 2013PLoSO...859977Z . doi : 10.1371/journal.pone.0059977 . PMC 3613400 . PMID 23560063 .
- ^ Jump up to: а беременный Egge, JK; Aksnes, DL (1992). «Силикат как регулирование питательных веществ в конкуренции фитопланктона» . Мар. ЭКОЛ. Прогик Сервис 83 : 281–289. Bibcode : 1992meps ... 83..281e . doi : 10.3354/meps083281 .
- ^ Jump up to: а беременный в Furnas, Miles J. (1990). «Скорости роста на месте морского фитопланктона: подходы к измерениям, темпам роста сообщества и видов». Журнал Plankton Research . 12 (6): 1117–51. doi : 10.1093/plankt/12.6.1117 . INIST 5474600 .
- ^ Jump up to: а беременный Йол, Эндрю; Тиррелл, Тоби (2003). «Роль диатомов в регулировании кремниевого цикла океана». Глобальные биогеохимические циклы . 17 (4): n/a. Bibcode : 2003gbioc..17.1103y . Citeseerx 10.1.1.394.3912 . doi : 10.1029/2002GB002018 . S2CID 16849373 .
- ^ Мальвия, Шрути; Скалько, Элеонора; Audic, Stéphane; Винсент, Флора; Veluchamy, Alaguraj; Пулен, Джули; Винккер, Патрик; Yudicone, Даниэле; де Варгас, Колумбан; Биттнер, Люси; Зингоне, Адриана; Боулер, Крис (29 февраля 2016 г.). «Понимание глобального распределения диатомовых и разнообразия в мировом океане» . Труды Национальной академии наук . 113 (11): E1516 - E1525. BIBCODE : 2016PNAS..113E1516M . doi : 10.1073/pnas.1509523113 . ISSN 0027-8424 . PMC 4801293 . PMID 26929361 .
- ^ Липпс, Джере Х. (1970). «Эволюция планктона». Эволюция 24 (1): 1–22. doi : 10.2307/2406711 . JSTOR 2406711 . PMID 28563010 .
- ^ "Дидимо, инопланетяне среди нас" . Архивировано 7 октября 2015 года на Виртуального музея виртуальной выставке Канады Виртуального музея Виртуального музея Виртуального музея
- ^ «DEP сообщает, что Дидимо обнаружил в реке Западного отделения Фармингтон» . Архивировано из оригинала 16 февраля 2015 года . Получено 27 апреля 2015 года .
- ^ «Обновление заинтересованных сторон Didymo - 31 октября 2008 года» . MAF Biosecurity New Zealand http://www.biosecurity.govt.nz . Архивировано из оригинала 12 февраля 2013 года . Получено 1 декабря 2013 года .
{{cite web}}: Внешняя ссылка в|publisher= - ^ Jump up to: а беременный Dugdale, RC; Wilkerson, FP (1998). «Силикатная регуляция нового производства в экваториальной Тихоокеанской Апвеллинге». Природа . 391 (6664): 270–273. Bibcode : 1998natur.391..270d . doi : 10.1038/34630 . S2CID 4394149 .
- ^ Smetacek, VS (1985). «Роль тонущего в циклах жизни диатомовой истории: экологическое, эволюционное и геологическое значение». Мар. Биол . 84 (3): 239–251. Bibcode : 1985marbi..84..239s . doi : 10.1007/bf00392493 . S2CID 85054779 .
- ^ Jump up to: а беременный Treguer, P.; Нельсон, DM; Ван Беннеком, AJ; Демастер, диджей; Leynaert, A.; Queguiner, B. (1995). «Баланс кремнезема в мировом океане: рецензия». Наука . 268 (5209): 375–9. Bibcode : 1995sci ... 268..375t . doi : 10.1126/science.268.5209.375 . PMID 17746543 . S2CID 5672525 .
- ^ Ворон, JA (1983). «Транспорт и функция кремния в растениях». Биол. Преподобный 58 (2): 179–207. doi : 10.1111/j.1469-185x.1983.tb00385.x . S2CID 86067386 .
- ^ Миллиган, AJ; Морел, FMM (2002). «Роль протонного буферизации для кремнезема в диатомовых условиях». Наука . 297 (5588): 1848–1850. Bibcode : 2002sci ... 297.1848m . doi : 10.1126/science.1074958 . PMID 12228711 . S2CID 206507070 .
- ^ Jump up to: а беременный Чемберлен, CJ (1901) Методы в гистологии растений , Университет Чикагской Прессы.
- ^ Р. Сивер; Стивен Генри Шнайдер; Пенелопа Дж. Бостон (январь 1993 г.). «Кремнезый в океанах: биологическое геологическое взаимодействие» . Ученые на Гайе . MIT Press . С. 287–295 . ISBN 978-0-262-69160-4 Полем Получено 14 ноября 2013 года .
- ^ Киддер, Дэвид Л.; Эрвин, Дуглас Х. (2001). «Светское распределение биогенного кремнезема через фанерозое: сравнение окаменелостей, заполненных кремнезмом и кусочков, на уровне серии». Журнал геологии . 109 (4): 509–22. Bibcode : 2001jg .... 109..509K . doi : 10.1086/320794 . S2CID 128401816 .
- ^ Гренн, Тор; Слэк, Джон Ф. (2003). «Палеозойская и мезозозовая силика, богатая морской водой: доказательства отложений гематитового Chert (Jasper)». Геология . 31 (4): 319–22. Bibcode : 2003geo .... 31..319G . doi : 10.1130/0091-7613 (2003) 031 <0319: Pamsrs> 2,0.co; 2 . INIST 14692468 .
- ^ Ricki, g; Корди, Фабрис (2000). «Радиоларийная палеэкология и радиолариты: ли это настоящее ключом к прошлому?». Земля-наука обзоров . 52 (1): 83–120. Bibcode : 2000esrv ... 52 ... 83r . doi : 10.1016/s0012-8252 (00) 00024-6 .
- ^ Мальдод, Мануэль; Кармонана, М. Кармен; Уриз, Мария Дж.; Crucado, Antonio (1999). «Снижение мезозозовых рифов, объясненных кремниевыми ограничениями» Природа 401 (6755): 785–8 Bibcode : 1999natur.401..785m . Doi : 10.1038/ 4 S2CID 205034177 Inst 1990263
- ^ Харпер, Говард Э.; Нолл, Эндрю Х. (1975). «Силика, диатомовые и кайнозойские радиолоальные эволюции». Геология . 3 (4): 175–177. Bibcode : 1975geo ..... 3..175H . doi : 10.1130/0091-7613 (1975) 3 <175: sdacre> 2.0.co; 2 .
- ^ Jump up to: а беременный Пьелла Карлусич, Хуан Хосе; Боулер, Крис; Бисвас, Хайманти (30 апреля 2021 года). «Механизмы концентрации углекислого газа в естественных популяциях морских диатомов: понимание океанов Тары» . Границы в науке о растениях . 12 Frontiers Media SA: 657821. DOI : 10.3389/fpls.2021.657821 . ISSN 1664-462x . PMC 8119650 . PMID 33995455 . Модифицированный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Фридлингштейн, Пьер; О'Салливан, Майкл; Джонс, Мэтью В.; Эндрю, Робби М.; и др. (11 декабря 2020 г.). «Глобальный углеродный бюджет 2020» . Земля системы науки . 12 (4). Коперник GmbH: 3269–3340. Bibcode : 2020ssd ... 12.3269f . doi : 10.5194/essd-12-3269-2020 . HDL : 10871/126892 . ISSN 1866-3516 .
- ^ Jump up to: а беременный Glibert, Patricia M.; Уилкерсон, Фрэнсис П.; Дугдейл, Ричард С.; Ворон, Джон А.; Дюпон, Кристофер Л.; Ливитт, Питер Р.; и др. (11 октября 2015 г.). «Плюсы и минусы аммония и поглощения и ассимиляции нитратов фитопланктоном и последствиями для продуктивности и состава сообщества, с акцентом на обогащенные азотом условиями». Лимнология и океанография . 61 (1). Уайли: 165–197. doi : 10.1002/lno.10203 . ISSN 0024-3590 . S2CID 16561857 . Модифицированный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Аллен, Эндрю Э.; Дюпон, Кристофер Л.; Обирник, Мирослав; Хорак, Алеш; Nunes-Nesi, Адриано; МакКроу, Джон П.; и др. (2011). «Эволюция и метаболическая значимость цикла мочевины при фотосинтетических диатомовых расставаниях» . Природа . 473 (7346): 203–7. Bibcode : 2011natur.473..203a . doi : 10.1038/nature10074 . PMID 21562560 . S2CID 4350884 . Архивировано из оригинала 19 февраля 2022 года . Получено 20 мая 2021 года .
- «Цикл мочевины, похожий на животных в крошечных диатомовых диатомовых условиях, позволяет морскому фитопланктону использовать углерод и азот из окружающей среды» . ScienceDaily (пресс -релиз). 12 мая 2011 года.
- ^ Jump up to: а беременный Армбруст, Э. Вирджиния; Бергес, Джон А.; Боулер, Крис; Зеленый, Беверли Р.; Мартинес, Диего; Путнэм, Николас Х.; и др. (2004). «Геном диатомовой таласиозиры псевдонаны: экология, эволюция и метаболизм» . Наука . 306 (5693). Американская ассоциация по развитию науки (AAAS): 79–86. Bibcode : 2004sci ... 306 ... 79a . doi : 10.1126/science.1101156 . ISSN 0036-8075 . PMID 15459382 . S2CID 8593895 .
- ^ Jump up to: а беременный в Аллен, Эндрю Э.; Дюпон, Кристофер Л.; Обирник, Мирослав; Хорак, Алеш; Nunes-Nesi, Адриано; МакКроу, Джон П.; и др. (2011). «Эволюция и метаболическая значимость цикла мочевины при фотосинтетических диатомовых расставаниях» . Природа . 473 (7346). Springer Science and Business Media LLC: 203–207. Bibcode : 2011natur.473..203a . doi : 10.1038/nature10074 . ISSN 0028-0836 . PMID 21562560 . S2CID 4350884 .
- ^ Вейман, Филипп Д.; Пиер, Карен; Lefebvre, Stephane C.; Ривера, Йозефа; Маккарти, Джеймс К.; Heuberger, Adam L.; и др. (10 октября 2014 г.). «Инактивация гена уреазы P-heodactylum tricornutum с использованием целевого мутагенеза на основе эффекторной нуклеазы на основе экстрапкции» . Plant Biotechnology Journal . 13 (4). Wiley: 460–470. doi : 10.1111/pbi.12254 . ISSN 1467-7644 . PMID 25302562 .
- ^ Бендер, Сара Дж.; Паркер, Микаэла С.; Armbrust, E. Virginia (2012). «Связанное влияние источника света и азота на цикл мочевины и метаболизм азота на цикл DIL в морской диатоме Thalassiosira Pseudonana». Профист . 163 (2). Elsevier BV: 232–251. doi : 10.1016/j.protis.2011.07.008 . ISSN 1434-4610 . PMID 21873112 .
- ^ Армстронг, E; Роджерсон, а; Leftley, JW (2000). «Использование углерода морских водорослей тремя поверхностными гетеротрофными протистами, стереомикса Ramosa, Nitzschia alba и Labyrinthula sp» . Водная микробная экология . 21 : 49–57. doi : 10.3354/ame021049 . ISSN 0948-3055 .
- ^ Левин, Джойс; Левин, Р.А. (1967). «Культура и питание некоторых апохлоротических диатомов рода Nitzschia» . Микробиология . 46 (3): 361–367. doi : 10.1099/00221287-46-3-361 . ISSN 1350-0872 .
- ^ Камп, А.; De Beer, D.; Nitsch, JL; Lavik, G.; Stief, P. (2011). «Диатомы дышат нитратами, чтобы пережить темные и аноксические условия» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (14): 5649–5654. Bibcode : 2011pnas..108.5649K . doi : 10.1073/pnas.1015744108 . PMC 3078364 . PMID 21402908 .
- ^ Мерц, Элиза; Дик, Грегори Дж.; Пиво, Дирк; Грим, Шарон; Хюбенера, Томас; Литтманн, Стен; и др. (2021). «Нитратные дыхательные и миграционные паттерны диатомовых лиц связаны в отложениях под микробным ковриком» . Экологическая микробиология . 23 (3): 1422–1435. Bibcode : 2021envmi..23.1422m . doi : 10.1111/1462-2920.15345 . HDL : 2027,42/167035 . PMID 33264477 . S2CID 227259897 .
- ^ Guiry, MD (2012). «Сколько там видов водорослей?». Журнал Phycology . 48 (5): 1057–1063. Bibcode : 2012jpcgy..48.1057g . doi : 10.1111/j.1529-8817.2012.01222.x . PMID 27011267 . S2CID 30911529 .
- ^ Раунд, Фрэнк Эрик; Кроуфорд, RM; Манн, Д.Г. (1990). Диатомы: биология и морфология родов . Издательство Кембриджского университета. ISBN 978-0-521-36318-1 Полем Архивировано из оригинала 24 мая 2021 года . Получено 13 ноября 2013 года . [ страница необходима ]
- ^ Canter-Lund, H. and Lund, JWG (1995). Пресноводные водоросли: их микроскопический мир объяснил , Biopress Limited. ISBN 0-948737-25-5 . [ страница необходима ]
- ^ Манн, Дэвид Г. (2005). «Концепция вида в диатомовых условиях: свидетельство морфологически различных симпатрических агодемов у четырех эпипелевых видов». Систематика растения и эволюция . 164 (1/4): 215–37. doi : 10.1007/bf00940439 . JSTOR 23675282 . S2CID 37684109 .
- ^ Fourtanier, Элизабет; Kociolek, J. Patrick (1999). «Каталог родов диатома». Diatom Research . 14 (1): 1–190. Bibcode : 1999diare..14 .... 1f . doi : 10.1080/0269249x.1999.9705462 .
- ^ Всемирный реестр морских видов перечисляет 1 356 названий диатомовых родов из всех мест обитания в июле 2020 года, из которых 1248 «приняты».
- ^ Запросы мирового регистра морских видов, июль 2020 года, возвращение 299 «Только ископаемые» названия родов, из которых 285 «приняты».
- ^ Териот, Эдвард С.; Cannone, Джейми Дж.; Гутелл, Робин Р.; Алверсон, Эндрю Дж. (2009). «Пределы ядерной кодируемой SSU RDNA для разрешения диатомовой филогения» . Европейский журнал Phycology . 4 (3): 277–290. doi : 10.5091/plecevo.2010.418 . PMC 2835975 . PMID 20224747 .
- ^ Териот, Эдвард С.; Эшворт, Мэтт; Рак, Элизабет; Наков, Теофил; Янсен, Роберт К. (2010). «Предварительная мультигенная филогения диатомов (Bacillariophyta): проблемы для будущих исследований» . Экология и эволюция растений . 143 (3): 277–290. doi : 10.1080/09670260902749159 . PMC 2835975 . PMID 20224747 .
- ^ Паркс, Мэтью Б.; Wickett, Norman J.; Алверсон, Эндрю Дж. (2018). «Сигнал, неопределенность и конфликт в филогеномных данных для разнообразной линии микробных эукариот (диатомовые лица, бациллариофита)» . Молекулярная биология и эволюция . 35 (1): 80–93. doi : 10.1093/molbev/msx268 . PMC 5850769 . PMID 29040712 .
- ^ Medlin, LK; Desdevises, Y. (2020). «Обзор филогенетической реконструкции диатомов с использованием молекулярных инструментов с анализом набора данных семи генов с использованием нескольких внешних групп и морфологических данных для общего подхода доказательств» (PDF) . Phycologia . в прессе. Архивировано (PDF) из оригинала 20 июля 2020 года . Получено 20 июля 2020 года .
- ^ Adl, Sina M.; Басс, Дэвид; Лейн, Кристофер Э.; Люкш, Юлиус; Schoch, Conrad L.; Смирнов, Алексей; и др. (26 сентября 2018 г.). «Пересмотр классификации, номенклатуру и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. doi : 10.1111/jeu.12691 . PMC 6492006 . PMID 30257078 .
- ^ Мишра, Мерамбика; Аруха, Ананта П.; Башир, Тафайл; Ядав, Дхананджай; Прасад, GBKS (5 июля 2017 г.). «Все новые лица диатомов: потенциальный источник наноматериалов и за его пределами» . Границы в микробиологии . 8 Frontiers Media SA: 1239. doi : 10.3389/fmicb.2017.01239 . ISSN 1664-302X . PMC 5496942 . PMID 28725218 . Материал был скопирован из этого источника, который доступен в рамках Attribution Creative Commons 4.0 Международная лицензия, архивная 16 октября 2017 года на машине Wayback .
- ^ Jump up to: а беременный Kooistra, Wiebe HCF; Медлин, Линда К. (1996). «Эволюция диатомов (Bacillariophyta)». Молекулярная филогенетика и эволюция . 6 (3): 391–407. doi : 10.1006/mpev.1996.0088 . PMID 8975694 .
- ^ Шибер, Юрген; Кринсли, Дэйв; Риципути, Ли (2000). «Диагенетическое происхождение кварцевого ила в мудруках и последствия для циклического кремнезема». Природа . 406 (6799): 981–5. Bibcode : 2000natur.406..981s . doi : 10.1038/35023143 . PMID 10984049 . S2CID 4417951 .
- ^ Medlin, LK; Kooistra, WHCF; Герсонде, Р.; Симс, Пенсильвания; Wellbrock, U. (1997). «Является ли происхождение диатомовых заболеваний, связанное с исчезновением массы конечной пермины?». Нова Хедвиджия . 65 (1–4): 1–11. doi : 10.1127/nova.hedwigia/65/1997/1 . HDL : 10013/Epic.12689 .
- ^ Ворон, JA; Уэйт, А.М. (2004). «Эволюция силицификации в диатомовых заболеваниях: неизбежное тонущее и тонущее как побег?» Полем Новый фитолог . 162 (1): 45–61. doi : 10.1111/j.1469-8137.2004.01022.x . JSTOR 1514475 .
- ^ Falkowski, PG; Кац, Мириам Э.; Кнолл, Эндрю Х.; Квигг, Антониетта; Ворон, Джон А.; Шофилд, Оскар; Тейлор, FJR (2004). «Эволюция современного эукариотического фитопланктона». Наука . 305 (5682): 354–60. Bibcode : 2004sci ... 305..354f . Citeseerx 10.1.1.598.7930 . doi : 10.1126/science.1095964 . PMID 15256663 . S2CID 451773 .
- ^ Киддер, DL; Gierlowski-Kordesch, EH (2005). «Влияние радиации пастбищ на цикл немориного кремнезема и миоценовый диатомит». Палаис . 20 (2): 198–206. Bibcode : 2005Palai..20..198k . doi : 10.2110/palo.2003.p03-108 . JSTOR 27670327 . S2CID 140584104 .
- ^ Jump up to: а беременный Лазарь, Дэвид; Баррон, Джон; Renaudie, Johan; Дайвер, Патрик; Тюрке, Андреас (2014). «Кенозойское планктонное разнообразие морской диатомовой диатомы и корреляция с изменением климата» . Plos один . 9 (1): E84857. BIBCODE : 2014PLOSO ... 984857L . doi : 10.1371/journal.pone.0084857 . PMC 3898954 . PMID 24465441 .
- ^ IPCC Core Writing Team, 2007. «Изменение климата 2007: отчет о синтезе». 104
- ^ Jump up to: а беременный Scherer, RP; Гладенков, А. Ю.; Баррон, JA (2007). «Методы и применение биостратиграфии кайнозойской морской диатомы». Документы палеонтологического общества . 13 : 61–83. doi : 10.1017/s1089332600001467 .
- ^ такое« прокси »данные? « Что www.ncdc.noaa.gov . Архивировано из оригинала 8 марта 2020 года . Получено 20 октября 2020 года .
- ^ Jump up to: а беременный Сванн, Геа; Snelling, Am (6 января 2015 г.). «Фотография изменений в северо-западе Тихого океана от MIS 4–5E» . Климат прошлого . 11 (1). Коперник GmbH: 15–25. Bibcode : 2015clipa..11 ... 15S . doi : 10.5194/cp-11-15-2015 . ISSN 1814-9332 . Материал был скопирован из этого источника, который доступен в соответствии с Attribution Creative Commons 3.0 Международная лицензия, архивная 23 февраля 2011 года на машине Wayback .
- ^ Сигман, Даниэль М.; Jaccard, Samuel L.; Хауг, Джеральд Х. (2004). «Полярная стратификация океана в холодном климате» . Природа . 428 (6978). Springer Science and Business Media LLC: 59–63. Bibcode : 2004natur.428 ... 59 с . doi : 10.1038/nature02357 . ISSN 0028-0836 . PMID 14999278 . S2CID 4329978 . Архивировано из оригинала 19 февраля 2022 года . Получено 15 декабря 2021 года .
- ^ Jump up to: а беременный Хауг, Джеральд Х.; Ганопольский, Андрей; Сигман, Даниэль М.; Розелл-Меле, Антони; Суонн, Джордж Э.А.; Тидеманн, Ральф; и др. (2005). «Северная тихоокеанская сезонность и оледенение Северной Америки 2,7 миллиона лет назад» . Природа . 433 (7028). Springer Science and Business Media LLC: 821–825. Bibcode : 2005natur.433..821h . doi : 10.1038/nature03332 . ISSN 0028-0836 . PMID 15729332 . S2CID 24116155 . Архивировано из оригинала 19 февраля 2022 года . Получено 15 декабря 2021 года .
- ^ Суонн, Джордж Э.А.; Маслин, Марк А.; Ленг, Мелани Дж.; Слоан, Хилари Дж.; Хауг, Джеральд Х. (24 февраля 2006 г.). «Diatom Δ18O Доказательства развития современной системы галоглина в субарктической северо -западной части Тихого океана в начале оледенения крупного северного полушария» . Палеоооооооография . 21 (1). Американский геофизический союз (AGU): N/A. Bibcode : 2006paloc..21.1009s . doi : 10.1029/2005pa001147 . ISSN 0883-8305 .
- ^ Nie, Junsheng; Король, Джон; Лю, Чженгю; Клеменс, Стив; Прелл, Уоррен; Fang, Xiaomin (2008). «Освещение поверхностной воды: причина начала стратификации в северной части Тихого океана с 2,75 млн. Лет?». Глобальные и планетарные изменения . 64 (1–2). Elsevier BV: 49–52. Bibcode : 2008gpc .... 64 ... 49n . doi : 10.1016/j.gloplacha.2008.08.003 . ISSN 0921-8181 .
- ^ Суонн, Джордж Э.А. (2010). «Изменения засоленности в северо -западной части Тихого океана во время позднего плиоцена/раннего четвертика с 2,73 мА до 2,52 мА» (PDF) . Земля и планетарные научные письма . 297 (1–2). Elsevier BV: 332–338. BIBCODE : 2010E & PSL.297..332S . doi : 10.1016/j.epsl.2010.06.035 . ISSN 0012-821X . Архивировано (PDF) от оригинала 19 июля 2018 года . Получено 15 декабря 2021 года .
- ^ Sarnthein, M.; Gebhardt, H.; Kiefer, T.; Kucera, M.; Кук, м.; Erlenkeuser, H. (2004). «Среднее голоценовое происхождение солености на море низко в субарктической северной части Тихого океана». Кватернарные науки обзоры . 23 (20–22). Elsevier BV: 2089–2099. Bibcode : 2004qsrv ... 23.2089s . doi : 10.1016/j.quascirev.2004.08.008 . ISSN 0277-3791 .
- ^ Jaccard, SL; Гэлбрейт, Эд; Сигман, DM; Haug, GH (2010). «Распространенная связь между антарктическим ледяным ядром и Subarctic Pacific Records за последние 800 кир». Кватернарные науки обзоры . 29 (1–2). Elsevier BV: 206–212. Bibcode : 2010qsrv ... 29..206j . doi : 10.1016/j.quascirev.2009.10.007 . ISSN 0277-3791 .
- ^ Гэлбрейт, Эрик Д.; Кинаст, Маркус; Jaccard, Samuel L.; Педерсен, Томас Ф.; Brunelle, Brigitte G.; Сигман, Даниэль М.; Кифер, Торстен (23 мая 2008 г.). «Последовательная связь между глобальным климатом и использованием поверхностных нитратов в западной части Субарктической Тихоокеанской Тихого океана в течение последних 500 ка» (PDF) . Палеоооооооография . 23 (2). Американский геофизический союз (AGU): N/A. BIBCODE : 2008Paloc..23.2212G . doi : 10.1029/2007PA001518 . ISSN 0883-8305 . S2CID 4082469 . Архивировано (PDF) из оригинала 15 июля 2021 года . Получено 15 декабря 2021 года .
- ^ Brunelle, Brigitte G.; Сигман, Даниэль М.; Jaccard, Samuel L.; Keigwin, Lloyd D.; Плессен, Биргит; Шеттлер, Георг; Кук, Mea S.; Хауг, Джеральд Х. (2010). «Ледниковые/межкльсовые изменения в подаче питательных веществ и стратификации в западной субарктической северной части тихоокеанского океана с момента предпоследнего ледникового максимума». Кватернарные науки обзоры . 29 (19–20). Elsevier BV: 2579–2590. BIBCODE : 2010QSRV ... 29.2579B . doi : 10.1016/j.quascirev.2010.03.010 . ISSN 0277-3791 .
- ^ Кохфельд, Карен Э.; Чейз, Занна (2011). «Контролируют дегласовые изменения в биогенных потоках в северной части Тихого океана». Кватернарные науки обзоры . 30 (23–24). Elsevier BV: 3350–3363. Bibcode : 2011qsrv ... 30.3350K . doi : 10.1016/j.quascirev.2011.08.007 . ISSN 0277-3791 .
- ^ Harwood, DM; Николаев, Вирджиния; Зима, DM (2007). «Цикеты эволюции, радиации и расширения диатомовой эволюции». Документы палеонтологического общества . 13 : 33–59. doi : 10.1017/s1089332600001455 .
- ^ Стрельникова, NI (1990). «Эволюция диатомовых средств в меховые и палеогеновые периоды». В Симоле, Х. (ред.). Материалы десятого международного симпозиума диатома . Koenigstein: Koeltz Scientific Books. С. 195–204. ISBN 3-87429-307-6 .
- ^ Baldauf, JG (1993). «Средний эоцен через ранний миоцен диатомовый оборот». В Протеро, Д.; Berggren, WH (Eds.). Эоцен-олигоцен климатическая и биотическая эволюция . Принстон: издательство Принстонского университета. С. 310–326. ISBN 0-691-02542-8 .
- ^ Баррон, JA (2003). «Внешний вид и вымирание планктонных диатомовых средств в течение последних 18 моих в Тихом и Южном океанах». Diatom Research . 18 : 203–224. doi : 10.1080/0269249x.2003.9705588 . S2CID 84781882 .
- ^ Скала, с.; Калеки, n; Falciatore, A; Chiusano, ML; Боулер, C (2002). «Свойства генома диатомового phaeodactylum tricornutum» . Физиология растений . 129 (3): 993–1002. doi : 10.1104/pp.010713 . PMC 166495 . PMID 12114555 .
- ^ Maheswari, U.; Монтсант, а; Голл, J; Krishnasamy, S; Раджяшри, Кр; Patell, VM; Bowler, C. (2004). «База данных Diatom EST» . Исследование нуклеиновых кислот . 33 (проблема базы данных): D344–7. doi : 10.1093/nar/gki121 . PMC 540075 . PMID 15608213 .
- ^ Монтсант, а.; Джаббари, К; Maheswari, U; Боулер, C (2005). «Сравнительная геномика Pennate Diatom Phaeodactylum tricornutum» . Физиология растений . 137 (2): 500–13. doi : 10.1104/pp.104.052829 . PMC 1065351 . PMID 15665249 .
- ^ Maheswari, U.; Макет, т.; Armbrust, EV; Боулер, C. (2009). «Обновление базы данных Diatom EST: новый инструмент для цифровой транскриптомики» . Исследование нуклеиновых кислот . 37 (проблема базы данных): D1001–5. doi : 10.1093/nar/gkn905 . PMC 2686495 . PMID 19029140 .
- ^ Jump up to: а беременный Armbrust, EV; Бергес, Джон А.; Боулер, Крис; Зеленый, Беверли Р.; Мартинес, Диего; Путнэм, Николас Х.; и др. (2004). «Геном диатомовой таласиозиры псевдонаны: экология, эволюция и метаболизм». Наука . 306 (5693): 79–86. Bibcode : 2004sci ... 306 ... 79a . Citeseerx 10.1.1.690.4884 . doi : 10.1126/science.1101156 . PMID 15459382 . S2CID 8593895 .
- ^ Jump up to: а беременный в дюймовый Боулер, Крис; Аллен, Эндрю Э.; Барсук, Джонатан Х.; Гримвуд, Джейн; Джаббари, Камель; Куо, Алан; и др. (2008). «Геном PhaeOdactylum выявляет эволюционную историю геномов диатомовых геномов» . Природа . 456 (7219): 239–244. Bibcode : 2008natur.456..239b . doi : 10.1038/nature07410 . PMID 18923393 . S2CID 4415177 .
- ^ Рой, SW; Пенни Д. (2007). «Очень высокая доля уникальных позиций интрона в богатых интрон диатомовой таласиозира псевдонана указывает на широко распространенное усиление интрона» . Молекулярная биология и эволюция . 24 (7): 1447–57. doi : 10.1093/molbev/msm048 . PMID 17350938 .
- ^ Моум, Флориан; Аллен, Эндрю Э; Мхири, Коринн; Ху, Ханхуа; Джаббари, Камель; Варди, Ассаф; Дедушка, Мари-Анголь; Боулер, Крис (2009). «Потенциальное влияние активированного стресса ретротранспозонов на эволюцию генома в морской диатоме» . BMC Genomics . 10 : 624. DOI : 10.1186/1471-2164-10-624 . PMC 2806351 . PMID 20028555 .
- ^ Moustafa, A.; Beszteri, B.; Майер, UG; Боулер, C.; Валентин, К.; Бхаттачарья Д. (2009). «Геномные следы загадочного пластидного эндосимбиоза в диатомовых условиях» (PDF) . Наука . 324 (5935): 1724–6. Bibcode : 2009Sci ... 324.1724M . doi : 10.1126/science.1172983 . PMID 19556510 . S2CID 11408339 . Архивировано из оригинала (PDF) 21 апреля 2014 года . Получено 13 января 2019 года .
- ^ Jump up to: а беременный Крот, Питер Г.; Bones, Atle M.; и др. (Октябрь 2018). «Редактирование генома в диатомовых условиях: достижения и цели» . Отчеты растительных ячеек . 37 (10): 1401–1408. doi : 10.1007/s00299-018-2334-1 . HDL : 11250/2590892 . PMID 30167805 . S2CID 52133809 . Архивировано из оригинала 19 февраля 2022 года . Получено 20 мая 2021 года .
- ^ Jump up to: а беременный Фалчаторе, Анжела; Casotti, Raffaella; и др. (Май 2015). «Трансформация генов неэлектаблера в морской диатомере». Морская биотехнология . 1 (3): 239–251. doi : 10.1007/pl00011773 . PMID 10383998 . S2CID 22267097 .
- ^ Jump up to: а беременный в дюймовый и Карас, Богумил Дж.; Закусочная, Рэйчел Э.; Lefebvre, Stephane C.; Маккуэйд, Джефф; Филлипс, Алекс Пр; Noddings, Chari M.; и др. (21 апреля 2015 г.). «Дизайнерские эпизоды Diatom, доставленные бактериальным сопряжением» . Природная связь . 6 (1): 6925. Bibcode : 2015natco ... 6.6925K . doi : 10.1038/ncomms7925 . ISSN 2041-1723 . PMC 4411287 . PMID 25897682 .
- ^ Jump up to: а беременный в дюймовый Slattery, Samuel S.; Даймонд, Эндрю; Ван, Хелен; Терриен, Жасмин А.; Лант, Джереми Т.; Jazey, Teah; и др. (16 февраля 2018 г.). «Расширенный генетический набор инструментов на основе плазмиды позволяет редактировать геном CAS9 и стабильное поддержание синтетических путей в Phaeodactylum tricornutum». ACS Синтетическая биология . 7 (2): 328–338. doi : 10.1021/acssynbio.7b00191 . PMID 29298053 .
- ^ Нимарк, Марианна; Шарма, Амит Кумар; и др. (Июль 2016 г.). «Система CRISPR/CAS9, адаптированная для редактирования генов в морских водорослях» . Научные отчеты . 6 (1): 24951. BIBCODE : 2016NATSR ... 624951N . doi : 10.1038/srep24951 . PMC 4842962 . PMID 27108533 .
- ^ Мишра, м; Arukha, AP; Башир, т; Ядав, D; Gbks, Prasad (2017). «Все новые лица диатомов: потенциальный источник наноматериалов и за его пределами» . Передний микробиол . 8 : 1239. DOI : 10.3389/fmicb.2017.01239 . PMC 5496942 . PMID 28725218 .
- ^ Река, Арианит А.; Павловский, Благо; Макрески, Петр (октябрь 2017 г.). «Новый оптимизированный метод для низкотемпературного гидротермального производства пористой керамики с использованием диатомовой земли» . Ceramics International . 43 (15): 12572–12578. doi : 10.1016/j.ceramint.2017.06.132 . Архивировано из оригинала 19 февраля 2022 года . Получено 8 апреля 2020 года .
- ^ Река, Арианат; Anovski, Todor; Богоевский, Слободан; Павловский, Благо; Бошковски, Бошко (29 декабря 2014 г.). «Физические химические и минералогические петрографические исследования диатомита из месторождения около деревни Рожден, Республика Македония» . Geologica Macedonica . 28 (2): 121–126. ISSN 1857-8586 . Архивировано из оригинала 18 апреля 2020 года . Получено 8 апреля 2020 года .
- ^ Рек, Арианит А.; Павловский, Благо; Адеми, Эгзон; Джашари, Ахмед; Боев, Блазо; Боев, Иван; Макрески, Петр (31 декабря 2019 г.). «Влияние термической обработки обработки трепеля в диапазоне расписаний 800-1200 ̊C » Открытая химия 17 (1): 1235–1 Doi : 10.1515/chem- 2019-0
- ^ Брэдбери, Дж. (2004). «Нанотехнологи природы: раскрыть секреты диатомовых заболеваний» . PLOS Биология . 2 (10): 1512–1515. doi : 10.1371/journal.pbio.0020306 . PMC 521728 . PMID 15486572 .

- ^ Барабан, Райан В.; Гордон, Ричард (2003). «Репликаторы Star Trek и нанотехнологии диатома». Тенденции в биотехнологии . 21 (8): 325–8. doi : 10.1016/s0167-7799 (03) 00169-0 . PMID 12902165 .
- ^ Джонсон, RC (9 апреля 2009 г.). «Диатомы могут тройной эффективности солнечных элементов» . EE времена . Архивировано из оригинала 31 июля 2012 года . Получено 13 апреля 2009 года .
- ^ Рамачандра, ТВ; Махапатра, Дурга Мадхаб; Б, Картик; Гордон, Ричард (2009). «Доильные диатомовые данные для устойчивой энергии: биохимическая инженерия в сравнении с бензиновыми солнечными панелями диатомовых солнечных батарей». Исследования промышленной и инженерной химии . 48 (19): 8769–88. doi : 10.1021/ie900044j .
- ^ Auer, Antti (1991). «Качественный анализ диатомы как инструмент для диагностики утопления». Американский журнал судебной медицины и патологии . 12 (3): 213–8. doi : 10.1097/00000433-199109000-00009 . PMID 1750392 . S2CID 38370984 .
- ^ Jump up to: а беременный в Пьелла Карлусич, Хуан Хосе; Ибарбальц, Федерико М; Боулер, Крис (2020). «Исследование морского фитопланктона: от их исторической оценки эпохи Omics» . Журнал Plankton Research . 42 : 595–612. doi : 10.1093/plankt/fbaa049 . HDL : 11336/143676 .
- ^ Долан, Джон Р. (1 августа 2019 г.). «Раскрытие» старшего сына отца простейших »: Чарльз Кинг» . Профист . 170 (4): 374–384. doi : 10.1016/j.protis.2019.07.002 . ISSN 1434-4610 . PMID 31479910 . S2CID 201207778 .
- ^ Дарвин, Ричард (1866). О происхождении видов посредством естественного отбора: или сохранения любимых рас в борьбе на всю жизнь .
Внешние ссылки
[ редактировать ]- Diatom - это база данных , école normale supériure
- Планктон*сеть , таксономическая база данных, включая изображения видов Diatom
- История жизни и экология диатомов , Музей палеонтологии Калифорнийского университета
- Diatoms: «Мраморы природы» , сайт Eureka, Университет Бергена
- История и экология Diatom циркуляцию микрофоссили , восстановление и
- Diatom Page Archived 8 октября 2009 года в The Wayback Machine , Royal Botanic Garden Edinburgh
- Геометрия и паттерн в природе 3: отверстия в тестах на радиоловых и диатоме
- Diatom QuickFacts , исследовательский институт аквариума Монтерей -Бей
- база данных изображений водорослей Архивирована 14 августа 2011 года в Wayback Matching Академии естественных наук о of Philadelphia (ANSP)
- Diatom таксоны архивировали 3 октября 2011 года в Wayback Machine Академии естественных наук о of Philadelphia (ANSP)
- Введение в микроскопическое исследование диатомов, архивное 5 августа 2022 года на машине Wayback Роберт Б. Маклафлин
