Черепаха
| Черепахи Временной диапазон: Черепахи стволовой группы происходят из среднего триаса.
| |
|---|---|

| |
| Обыкновенная коробчатая черепаха | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | Пантестудины |
| Клэйд : | Черепахи |
| Клэйд : | Перихелидия |
| Заказ: | Черепахи Бач , 1788 г. [ 1 ] |
| Подгруппы | |
| Разнообразие | |
| 14 живых семей | |
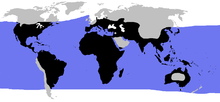
| |
| Синий: морские черепахи, черный: наземные черепахи. | |
| Синонимы [ 2 ] | |
| |
Черепахи — рептилии отряда развитым Testudines , характеризующиеся особым панцирем, преимущественно из их ребер. Современные черепахи делятся на две основные группы: Pleurodira (черепахи с боковой шеей) и Cryptodira (черепахи со скрытой шеей), которые различаются способом втягивания головы. Существует 360 ныне живущих и недавно вымерших видов черепах, включая сухопутных черепах и пресноводных . Они встречаются на большинстве континентов, на некоторых островах и, в случае морских черепах , на большей части океана. Как и другие амниоты (рептилии, птицы и млекопитающие ), они дышат воздухом и не откладывают яйца под водой, хотя многие виды живут в воде или вокруг нее.
Панцири черепах состоят главным образом из костей ; верхняя часть — куполообразный панцирь , а нижняя — более плоский пластрон или брюшная пластина. Его внешняя поверхность покрыта чешуйками из кератина — материала волос, рогов и когтей. Кости панциря развиваются из ребер, которые растут в стороны, и превращаются в широкие плоские пластины, которые соединяются, чтобы покрыть тело. Черепахи являются эктотермными или «хладнокровными», что означает, что их внутренняя температура варьируется в зависимости от их непосредственной среды. Как правило, они всеядны и в основном питаются растениями и животными с ограниченными движениями. Многие черепахи сезонно мигрируют на короткие расстояния. Морские черепахи — единственные рептилии, которые мигрируют на большие расстояния, чтобы отложить яйца на излюбленном пляже.
Черепахи появлялись в мифах и сказках по всему миру. Некоторые наземные и пресноводные виды широко используются в качестве домашних животных. На черепах охотились ради мяса, использования в традиционной медицине и панцирей. Морские черепахи часто случайно погибают в качестве прилова в рыболовных сетях. Места обитания черепах по всему миру уничтожаются. В результате такого давления многие виды вымерли или находятся под угрозой исчезновения.
Именование и этимология
Слово черепаха заимствовано от французского слова tortue или tortre «черепаха, черепаха ». [ 3 ] Это общее название , и его можно использовать без знания таксономических различий. В Северной Америке это может обозначать отряд в целом. В Великобритании это название используется для морских черепах , а не для пресноводных и наземных черепах. В Австралии, где нет настоящих черепах (семейство Testudinidae), неморских черепах традиционно называли черепахами, но в последнее время черепахами стали называть всю группу. [ 4 ]
Название отряда Testudines ( / t ɛ ˈ s tj uː d ɪ n iː z / teh- STEW -din-eez ), основано на латинском слове testudo «черепаха»; [ 5 ] и был придуман немецким натуралистом Августом Бачем в 1788 году. [ 1 ] Орден также исторически известен как Челонии ( Латрей 1800 г.) и Челония (Росс и Макартни 1802 г.). [ 2 ] которые основаны на древнегреческом слове χελώνη ( chelone ) «черепаха». [ 6 ] [ 7 ] Testudines — официальное название заказа, обусловленное принципом приоритета . [ 2 ] Термин хелониан используется как официальное название членов группы. [ 1 ] [ 8 ]
Анатомия и физиология
Размер
Самый крупный из ныне живущих видов черепах (и четвертая по величине рептилия ) — кожистая черепаха , которая может достигать более 2,7 м (8 футов 10 дюймов) в длину и весить более 500 кг (1100 фунтов). [ 9 ] Самой крупной известной черепахой была Archelon ischyros , морская черепаха позднего мелового периода длиной до 4,5 м (15 футов), шириной между кончиками передних ласт 5,25 м (17 футов) и весом более 2200 кг (4900 фунтов). . [ 10 ] Самая маленькая из ныне живущих черепах - Chersobius Signatus из Южной Африки, ее длина не превышает 10 см (3,9 дюйма). [ 11 ] и весом 172 г (6,1 унции). [ 12 ]
Оболочка

Панцирь черепахи уникален среди позвоночных и служит для защиты животного и укрытия от непогоды. [ 13 ] [ 14 ] [ 15 ] Он в основном состоит из 50–60 костей и состоит из двух частей: куполообразного дорсального (заднего) панциря и более плоского вентрального (брюшного) пластрона . Они соединены латеральными (боковыми) отростками пластрона. [ 13 ] [ 16 ]
Панцирь сращен с позвонками и ребрами, а пластрон формируется из костей плечевого пояса , грудины и гастралии (брюшных ребер). [ 13 ] Во время развития ребра растут в стороны, образуя панцирный гребень, уникальный для черепах, и входят в дерму (внутреннюю кожу) спины, чтобы поддерживать панцирь. О развитии локально сигнализируют белки, известные как факторы роста фибробластов , которые включают FGF10 . [ 17 ] Плечевой пояс черепах состоит из двух костей: лопатки и клювовидного отростка . [ 18 ] И плечевой, и тазовый пояс черепах расположены внутри панциря и, следовательно, фактически находятся внутри грудной клетки. Ребра туловища во время развития растут над плечевым поясом. [ 19 ]

Панцирь покрыт эпидермальными (внешними) чешуйками, известными как щитки , которые состоят из кератина — того же вещества, из которого состоят волосы и ногти. Обычно у черепахи 38 щитков на панцире и 16 на пластроне, всего их 54. Щитки панциря делятся на «краевые» по краю и «позвоночные» над позвоночным столбом, хотя щиток, покрывающий шею, называется «шейным». «Плевры» присутствуют между маргинальными и позвонками. [ 20 ] Пластронные щитки включают горло (горло), плечевые кости, грудные кости, брюшные кости и анальные кости. У черепах с боковой шеей дополнительно есть «междузорные» щитки между горлами. [ 16 ] [ 21 ] Щитки черепах обычно имеют структуру мозаичной плитки, но у некоторых видов, например у морской черепахи бисса , щитки на панцире перекрываются. [ 16 ]
Формы панцирей черепах различаются в зависимости от адаптации отдельных видов, а иногда и от пола . Наземные черепахи имеют более куполообразную форму, что делает их более устойчивыми к раздавливанию крупными животными. У водных черепах более плоский и гладкий панцирь, который позволяет им рассекать воду. Морские черепахи, в частности, имеют обтекаемый панцирь, который уменьшает сопротивление и повышает устойчивость в открытом океане. У некоторых видов черепах заостренный или шипастый панцирь обеспечивает дополнительную защиту от хищников и маскировку от листвы. Комки панциря черепахи могут наклонять ее тело, когда ее переворачивают, позволяя ей перевернуться назад. У черепах-самцов кончик пластрона утолщен и используется для бодания и тарана во время боя. [ 22 ]
Оболочки различаются по гибкости. У некоторых видов, таких как коробчатые черепахи , отсутствуют боковые расширения, и вместо этого кости панциря полностью слиты или анкилозированы вместе. У некоторых видов на панцирях, обычно на пластроне, есть шарниры, которые позволяют им расширяться и сжиматься. У черепах софтшелл эластичные края из-за потери костей. В панцире кожистой черепахи почти нет костей, но имеется толстая соединительная ткань и внешний слой кожистой кожи. [ 23 ]
Голова и шея

Череп черепахи уникален среди ныне живущих амниот (включая рептилий, птиц и млекопитающих); он твердый и жесткий, без отверстий для прикрепления мышц ( височные окна ). [ 24 ] [ 25 ] Вместо этого мышцы прикрепляются к углублениям в задней части черепа. Черепа черепах различаются по форме: от длинных и узких черепов софтшеллов до широкого и уплощенного черепа мата-мата . [ 25 ] У некоторых видов черепах большие и толстые головы, что обеспечивает большую мышечную массу и более сильные укусы. [ 26 ]
Черепахи, которые являются плотоядными или дурофагами (питаются животными с твердым панцирем), имеют самые сильные укусы. Например, у дурофага Mesoclemmys nasuta сила укуса составляет 432 фунта-силы (1920 Н). виды Насекомоядные , рыбоядные (рыбоядные) или всеядные имеют меньшую силу укуса. [ 27 ] есть клювы из кератиновых оболочек . У живых черепах нет зубов, но по краям челюстей [ 28 ] [ 13 ] Эти ножны могут иметь острые края для резки мяса, зубцы для обрезки растений или широкие пластины для разламывания моллюсков . [ 29 ] У морских черепах и некоторых вымерших форм развилось костное вторичное небо , которое полностью разделяет ротовую и носовую полости. [ 30 ]
Шеи черепах очень гибкие, возможно, чтобы компенсировать их жесткий панцирь. Некоторые виды, например морские черепахи, имеют короткую шею, а другие, например черепахи со змеешеей , имеют длинную шею. Несмотря на это, все виды черепах имеют восемь шейных позвонков — консистенция, не встречающаяся у других рептилий, но схожая с млекопитающими . [ 31 ] Некоторые черепахи со змеиной шеей имеют как длинную шею, так и большую голову, что ограничивает их способность поднимать их, когда они не находятся в воде. [ 26 ] У некоторых черепах есть складчатые структуры в гортани или голосовой щели , которые вибрируют, издавая звук. У других видов эластином богаты голосовые связки . [ 32 ] [ 33 ]
Конечности и передвижение
Из-за тяжелого панциря черепахи медленно передвигаются по суше. движется Пустынная черепаха всего со скоростью 0,22–0,48 км/ч (0,14–0,30 миль в час). Напротив, морские черепахи могут плавать со скоростью 30 км/ч (19 миль в час). [ 13 ] Конечности черепах приспособлены к различным способам передвижения и повадкам, большинство из них имеют пять пальцев. Черепахи приспособлены к наземной среде обитания и имеют колоннообразные ноги со слоновьими ступнями и короткими пальцами. У черепахи -суслика уплощенные передние конечности, позволяющие рыть субстрат. У пресноводных черепах более гибкие ноги и более длинные пальцы с перепонками , что позволяет им толкаться в воде. Некоторые из этих видов, такие как щелкающие черепахи и грязевые черепахи , в основном ходят по водному дну, как и по суше. Другие, например черепахи, плавают, гребля всеми четырьмя конечностями, переключаясь между противоположными передними и задними конечностями, что сохраняет их направление стабильным. [ 13 ] [ 34 ]

Морские черепахи и свиноносая черепаха наиболее приспособлены к плаванию. Их передние конечности превратились в ласты, а более короткие задние конечности по форме больше напоминают рули направления. Передние конечности обеспечивают большую часть тяги при плавании, а задние служат стабилизаторами. [ 13 ] [ 36 ] Морские черепахи, такие как зеленая морская черепаха, вращают ласты передних конечностей, как крылья птицы, создавая движущую силу как при движении вверх, так и при движении вниз. Это контрастирует с пресноводными черепахами аналогичного размера (в каждом случае измерения проводились на молодых животных), такими как каспийская черепаха , которая использует передние конечности как весла гребной лодки, создавая существенную отрицательную тягу при подъеме в море. каждый цикл. Кроме того, обтекаемость морских черепах снижает сопротивление. В результате морские черепахи производят движущую силу в два раза большую и плавают в шесть раз быстрее, чем пресноводные черепахи. Эффективность плавания молодых морских черепах аналогична эффективности плавания быстроплавающих рыб открытой воды, таких как скумбрия . [ 35 ]
По сравнению с другими рептилиями, черепахи, как правило, имеют уменьшенные хвосты, но они различаются как по длине, так и по толщине в зависимости от вида и пола. У щелкающих черепах и большеголовых черепах более длинные хвосты; последний использует его для равновесия во время лазания. Клоака . находится внизу и у основания, а в самом хвосте находятся репродуктивные органы Следовательно, у самцов более длинные хвосты, в которых находится пенис. У морских черепах хвост длиннее и более цепкий у самцов, которые используют его для захвата самок. У некоторых видов черепах на хвосте есть шипы. [ 37 ] [ 24 ]
Чувства

Черепахи используют зрение , чтобы находить пищу и партнеров, избегать хищников и ориентироваться. палочки фотопигментами Светочувствительные клетки сетчатки включают в себя как для зрения при слабом освещении, так и колбочки различными с тремя для яркого света, обеспечивающие полноцветное зрение. Возможно, существует четвертый тип конуса, который обнаруживает ультрафиолет , поскольку только что вылупившиеся морские черепахи экспериментально реагируют на ультрафиолетовый свет, но неизвестно, могут ли они отличить его от более длинных волн. Пресноводная черепаха, красноухий ползунок , имеет исключительные семь типов колбочек. [ 38 ] [ 39 ] [ 40 ]
Морские черепахи ориентируются на суше ночью, используя визуальные особенности, обнаруживаемые в тусклом свете. Они могут использовать свои глаза в прозрачной поверхностной воде, на илистых берегах, в темноте глубокого океана, а также над водой. В отличие от наземных черепах, роговица (изогнутая поверхность, пропускающая свет в глаз) не помогает фокусировать свет на сетчатке, поэтому фокусировка под водой полностью осуществляется хрусталиком, находящимся за роговицей. Конусные клетки содержат капли масла, которые сдвигают восприятие в сторону красной части спектра, улучшая различение цветов. Острота зрения, изученная у птенцов, самая высокая в горизонтальной полосе, где клетки сетчатки упакованы примерно в два раза плотнее, чем в других местах. Это обеспечивает лучший обзор по всему визуальному горизонту. Морские черепахи, похоже, не используют поляризованный свет для ориентации, как это делают многие другие животные. У ныряющей на глубину кожистой черепахи отсутствуют особые приспособления к слабому освещению, такие как большие глаза, большие линзы или светоотражающий тапетум . Он может полагаться на наблюдение за биолюминесценцией. добычи при охоте на глубокой воде. [ 38 ]
У черепах нет ушных отверстий; барабанная перепонка покрыта чешуей и окружена костной слуховой капсулой , отсутствующей у других рептилий. [ 31 ] Их порог слуха высок по сравнению с другими рептилиями, достигая на воздухе до 500 Гц , но под водой они более приспособлены к более низким частотам. [ 41 ] Экспериментально было показано, что морская черепаха-логгерхед реагирует на тихие звуки с максимальной чувствительностью от 100 до 400 Гц. [ 42 ]
У черепах есть обонятельные (обонятельные) и вомероназальные рецепторы вдоль носовой полости, последние из которых используются для обнаружения химических сигналов. [ 43 ] Эксперименты на зеленых морских черепахах показали, что они могут научиться реагировать на ряд различных пахучих химических веществ, таких как триэтиламин и коричный альдегид , которые обнаруживаются при обонянии носа. Такие сигналы можно было бы использовать в навигации. [ 44 ]
Дыхание

Жесткий панцирь черепах не способен расширяться и освобождать место для легких, как у других амниот, поэтому им пришлось выработать особые приспособления для дыхания. [ 45 ] [ 46 ] [ 47 ] Легкие черепах прикрепляются непосредственно к панцирю сверху, а снизу соединительная ткань прикрепляет их к органам. [ 48 ] У них есть несколько латеральных (боковых) и медиальных (средних) камер (количество которых варьируется у разных видов) и одна терминальная (концевая) камера. [ 49 ]
Легкие вентилируются с помощью определенных групп мышц живота, прикрепленных к органам, которые их тянут и толкают. [ 45 ] В частности, именно большая печень черепахи сдавливает легкие. Под легкими, в целомической полости , печень соединяется с правым легким корнем , а желудок непосредственно прикрепляется к левому легкому, а к печени - брыжейкой . Когда печень опускается вниз, начинается вдох. [ 46 ] Легкие поддерживают стенка или перегородка , которая, как полагают, предотвращает их коллапс. [ 50 ] Во время выдоха сокращение поперечной мышцы живота продвигает органы в легкие и выталкивает воздух. И наоборот, во время вдоха расслабление и уплощение косой мышцы живота тянет поперечную мышцу вниз, позволяя воздуху вернуться в легкие. [ 46 ]
Хотя многие черепахи проводят большую часть своей жизни под водой, все черепахи дышат воздухом и должны регулярно всплывать на поверхность, чтобы наполнить свои легкие. В зависимости от вида продолжительность погружения варьируется от минуты до часа. [ 51 ] Некоторые виды могут дышать через клоаку , которая содержит большие мешочки, выстланные множеством пальцеобразных выступов, которые поглощают растворенный кислород из воды. [ 52 ]
Тираж

Черепахи имеют общую кровеносную и легочную (легочные) системы позвоночных, где трехкамерное сердце перекачивает лишенную кислорода кровь через легкие, а затем перекачивает возвращенную насыщенную кислородом кровь через ткани тела. Сердечно-легочная система имеет как структурные, так и физиологические адаптации, отличающие ее от других позвоночных. Черепахи имеют большой объем легких и могут перемещать кровь через внелегочные кровеносные сосуды, в том числе внутри сердца, чтобы избежать попадания в легкие, пока они не дышат. Они могут задерживать дыхание на гораздо более длительные периоды, чем другие рептилии, и могут переносить низкий уровень кислорода. Они могут смягчать повышение кислотности во время анаэробного (бескислородного) дыхания за счет химической буферизации и могут находиться в состоянии покоя в течение нескольких месяцев в период эстивации или брумации . [ 53 ]
Сердце имеет два предсердия , но только один желудочек . Желудочек подразделяется на три камеры. Мышечный гребень обеспечивает сложную структуру кровотока, так что кровь может быть направлена либо в легкие через легочную артерию , либо в тело через аорту . Способность разделять два оттока варьируется в зависимости от вида. Кожаная кожа имеет мощный мускулистый гребень, позволяющий практически полностью отделить истечения, поддерживая активно плавательный образ жизни. Гребень менее развит у пресноводных черепах типа ползунков ( Trachemys ). [ 53 ]
Черепахи способны переносить периоды анаэробного дыхания дольше, чем многие другие позвоночные. В этом процессе сахара расщепляются не полностью до молочной кислоты , а не до углекислого газа и воды, как при аэробном (кислородном) дыхании . [ 53 ] Они используют скорлупу как источник дополнительных буферных веществ для борьбы с повышенной кислотностью и как поглотитель молочной кислоты. [ 54 ]
Осморегуляция
У морских черепах мочевой пузырь представляет собой одно целое, а у большинства пресноводных черепах — двухлопастное. [ 55 ] Мочевой пузырь морских черепах соединен с двумя небольшими добавочными пузырями, расположенными по бокам от шейки мочевого пузыря и над лобком . [ 56 ] У черепах, живущих в засушливых условиях, есть мочевые пузыри, которые служат запасами воды, сохраняя в жидкости до 20% веса их тела. В жидкостях обычно мало растворенных веществ , но их больше во время засухи, когда рептилия получает соли калия из растительной диеты. Мочевой пузырь хранит эти соли до тех пор, пока черепаха не найдет свежую питьевую воду. [ 57 ] Чтобы регулировать количество соли в организме, морские черепахи и солоноватые ромбовидные черепахи избыток соли в виде густого липкого вещества выделяют из своих слезных желез . Из-за этого морским черепахам может казаться, что они «плачут», находясь на суше. [ 58 ]
Терморегуляция

Черепахи, как и другие рептилии, имеют ограниченную способность регулировать температуру своего тела . Эта способность варьируется в зависимости от вида и размера тела. Маленькие прудовые черепахи регулируют свою температуру, выползая из воды и греясь на солнце, а маленькие наземные черепахи перемещаются между солнечными и затененными местами, чтобы регулировать свою температуру. Крупные виды, как наземные, так и морские, имеют достаточную массу, чтобы придать им значительную тепловую инерцию , а это означает, что они нагреваются или остывают в течение многих часов. Гигантская черепаха Альдабра весит около 60 килограммов (130 фунтов) и способна позволить своей температуре подняться примерно до 33 °C (91 °F) в жаркий день и естественным образом упасть примерно до 29 °C (84 °F). Ф) ночью. Некоторые гигантские черепахи ищут тень, чтобы не перегреться в солнечные дни. На острове Гранд-Терр в глубине суши мало еды, у побережья мало тени, а в жаркие дни черепахи конкурируют за место под несколькими деревьями. Крупные самцы могут вытеснить более мелких самок из тени, а некоторые затем перегреваются и погибают. [ 59 ]
Взрослые морские черепахи также имеют достаточно большие тела, поэтому они могут в некоторой степени контролировать свою температуру. Самая крупная черепаха, кожистая, может плавать в водах Новой Шотландии , температура которых может достигать 8 °C (46 °F), а температура их тела, по измерениям, может быть на 12 °C (22 °F) выше. чем окружающая вода. Чтобы поддерживать температуру на высоком уровне, у них есть система противоточного теплообмена в кровеносных сосудах между ядром тела и кожей ласт. Сосуды, снабжающие голову, изолированы жиром вокруг шеи. [ 59 ]
Поведение
Диета и кормление

Большинство видов черепах являются оппортунистическими всеядными; Наземные виды более травоядны , а водные более плотоядны . [ 26 ] Как правило, не обладая скоростью и ловкостью, большинство черепах питаются либо растительным материалом, либо животными с ограниченными движениями, такими как моллюски, черви и личинки насекомых. [ 13 ] Некоторые виды, такие как африканская шлемофонная черепаха и щелкающие черепахи, едят рыбу, земноводных, рептилий (включая других черепах), птиц и млекопитающих. Они могут захватить их из засады, но также и собрать мусор. [ 60 ] У щелкающей черепахи-аллигатора на языке есть червеобразный придаток, который она использует, чтобы заманивать рыбу в рот. Черепахи — самая травоядная группа, питающаяся травой, листьями и фруктами. [ 61 ] Многие виды черепах, в том числе черепахи, дополняют свой рацион яичной скорлупой, костями животных, волосами и пометом для получения дополнительных питательных веществ. [ 62 ]
Черепахи обычно едят пищу простым способом, хотя у некоторых видов есть особые методы кормления. [ 13 ] Желтопятнистая речная черепаха и раскрашенная черепаха могут фильтровать корм , скользя по поверхности воды с открытым ртом и горлом, чтобы собрать частицы пищи. Когда рот закрывается, горло сжимается, и вода выталкивается через ноздри и щель между челюстями. [ 63 ] Некоторые виды используют «метод зияния и сосания», при котором черепаха открывает челюсти и широко расширяет горло, засасывая добычу. [ 13 ] [ 64 ] [ 65 ]
Рацион особи внутри вида может меняться в зависимости от возраста, пола и сезона, а также может различаться между популяциями. У многих видов молодые особи обычно плотоядны, но по мере взросления становятся более травоядными. [ 13 ] [ 66 ] У черепахи с карты Барбура более крупная самка в основном питается моллюсками, а самец обычно ест членистоногих . [ 13 ] Черепаха Бландинга может питаться в основном улитками или раками, в зависимости от популяции. летом переходит Было зарегистрировано, что европейская прудовая черепаха большую часть года в основном плотоядна, но кувшинки . на [ 67 ] Некоторые виды разработали специализированные диеты, такие как ястреб, питающийся губками , кожистая черепаха, питающаяся медузами , и меконгская черепаха, питающаяся улитками . [ 26 ] [ 13 ]
Связь и интеллект

Хотя черепахи обычно считаются немыми, для общения они издают различные звуки. [ 69 ] [ 70 ] Одно исследование, в котором было зарегистрировано 53 вида, показало, что все они вокализировали. [ 71 ] Черепахи могут реветь во время ухаживания и спаривания. [ 70 ] [ 31 ] Различные виды как пресноводных, так и морских черепах издают короткие низкочастотные звуки с момента нахождения в яйце до взрослой жизни. Эти вокализации могут способствовать сплочению группы при миграции . [ 70 ] особенно У продолговатой черепахи большой голосовой диапазон; издает звуки, описываемые как треск, щелканье, визг, уханье, различные виды щебетания, воя, ууу , хрюканья, рычания, взрывов, завывания и барабанной дроби. [ 68 ]
Игровое поведение было зарегистрировано у некоторых видов черепах. [ 72 ] В лаборатории краснобрюхие кутеры Флориды могут обучаться новым задачам и продемонстрировали долговременную память продолжительностью не менее 7,5 месяцев. [ 73 ] Точно так же гигантские черепахи могут учиться и запоминать задания, а также усваивать уроки гораздо быстрее, если обучаться в группах. [ 74 ] Черепахи, похоже, способны сохранять оперантную обусловленность через девять лет после первоначального обучения. [ 75 ] Исследования показали, что черепахи могут ориентироваться в окружающей среде, используя ориентиры и систему, похожую на карту, что позволяет прокладывать точные прямые маршруты к цели. [ 76 ] Навигация у черепах коррелирует с высокой когнитивной функцией в медиальной области коры головного мозга. [ 76 ] [ 77 ]

Оборона
Почувствовав опасность, черепаха может убежать, замерзнуть или уйти в свой панцирь. Пресноводные черепахи убегают в воду, хотя грязевая черепаха Соноры может найти убежище на суше, поскольку мелкие временные водоемы, в которых они обитают, делают их уязвимыми. [ 78 ] Напуганная черепаха-мягкотел может нырнуть под воду и зарыться под морским дном. [ 79 ] Если хищник упорствует, черепаха может укусить или выбросить из клоаки. Некоторые виды производят зловонные химические вещества из мускусных желез. Другая тактика включает в себя демонстрацию угрозы, а черепаха Белла с шарнирной спиной может притвориться мертвой . При нападении детеныши большеголовых черепах визжат, возможно, пугая хищника. [ 80 ]
Миграция

Черепахи — единственные рептилии, которые мигрируют на большие расстояния, а точнее морские виды, которые могут путешествовать на тысячи километров. Некоторые неморские черепахи, такие как виды Geochelone (наземные), Chelydra (пресноводные) и Malaclemys (устьевые), сезонно мигрируют на гораздо более короткие расстояния, примерно до 27 км (17 миль), чтобы отложить яйца. Такие короткие миграции сравнимы с миграциями некоторых ящериц, змей и крокодилов. [ 81 ] Морские черепахи гнездятся в определенном месте, например, на пляже, оставляя яйца вылупляться без присмотра. Молодые черепахи покидают этот район, мигрируя на большие расстояния в течение многих лет или десятилетий, пока они достигают зрелости, а затем возвращаются, по-видимому, в один и тот же район каждые несколько лет, чтобы спариваться и откладывать яйца, хотя точность варьируется в зависимости от вида и популяции. Это «натальное возвращение» показалось биологам примечательным, хотя сейчас существует множество доказательств этого, в том числе генетических. [ 82 ]
Как морские черепахи добираются до пляжей для размножения, остается неизвестным. Одной из возможностей является импринтинг, как у лосося , когда молодь изучает химическую подпись, то есть запах, своей домашней воды перед отъездом и запоминает это, когда придет время вернуться взрослыми. Другой возможный сигнал — ориентация магнитного поля Земли на натальном пляже. Существуют экспериментальные доказательства того, что черепахи обладают эффективным магнитным чутьем и используют его в навигации . Доказательство того, что происходит хоминг, получено на основе генетического анализа популяций головастых, ястребов, кожистых и оливковых ридли по местам гнездования. Для каждого из этих видов популяции в разных местах имеют свои собственные генетические характеристики митохондриальной ДНК , которые сохраняются на протяжении многих лет. Это показывает, что популяции различны и что возвращение домой должно происходить надежно. [ 82 ]
Размножение и жизненный цикл

Черепахи имеют большое разнообразие брачного поведения, но не образуют парных связей или социальных групп. [ 83 ] У зеленых морских черепах самок обычно больше, чем самцов. [ 84 ] У наземных видов самцы часто крупнее самок, и борьба между самцами устанавливает иерархию доминирования для доступа к партнерам. Для большинства полуводных и донноходных водных видов бои происходят реже. Вместо этого самцы этих видов могут использовать свое преимущество в размерах для насильственного спаривания . У полностью водных видов самцы часто меньше самок и полагаются на ухаживания, чтобы получить доступ к самкам для спаривания. [ 85 ]
Ухаживание и монтаж
Ухаживание варьируется в зависимости от вида и среды обитания. У водных видов, как морских, так и пресноводных, он часто бывает сложным, но проще у полуводных грязевых и щелкающих черепах. Самец черепахи кивает головой, а затем подчиняет самку, кусая и бодя ее, прежде чем сесть на нее. [ 13 ] Самец грязевой черепахи-скорпиона приближается к самке сзади и часто прибегает к агрессивным методам, таким как кусание самки за хвост или задние конечности с последующей садкой на нее. [ 86 ]
Для некоторых видов важен выбор самок, а самки зеленых морских черепах не всегда восприимчивы. Таким образом, они выработали поведение, позволяющее избегать попыток самца совокупления, таких как уплывание, столкновение с самцом с последующим укусом или принятие позиции отказа с вертикальным телом, широко раскинутыми конечностями и пластроном, обращенным к самцу. Если вода слишком мелка для позиции отказа, самки прибегают к выбрасыванию на берег самостоятельно, поскольку самцы не следуют за ними на берег. [ 84 ]

Все черепахи оплодотворяются изнутри; монтаж и совокупление могут быть затруднены. У многих видов самцы имеют вогнутый пластрон, который сцепляется с панцирем самки. У таких видов, как русская черепаха , самец имеет более легкий панцирь и более длинные ноги. Высокая, округлая форма коробчатых черепах является особым препятствием для посадки. Самец восточной коробчатой черепахи наклоняется назад и цепляется за спину пластрона самки. [ 87 ] Водные черепахи садятся в воду, [ 88 ] [ 89 ] а самки морских черепах поддерживают садящегося самца во время плавания и ныряния. [ 90 ] Во время совокупления черепаха-самец выравнивает свой хвост с хвостом самки, чтобы можно было вставить свой пенис в ее клоаку. [ 91 ] Некоторые самки черепах могут хранить сперму от нескольких самцов, а в их яйцевых кладках может быть несколько производителей. [ 92 ] [ 83 ]
Яйца и детеныши

Черепахи, в том числе морские, откладывают яйца на суше, хотя некоторые откладывают яйца вблизи воды, уровень которой поднимается и опускается, погружая яйца в воду. Хотя большинство видов строят гнезда и откладывают яйца там, где добывают пищу, некоторые путешествуют на многие мили. Обыкновенная щелкающая черепаха проходит по суше 5 км (3 мили), а морские черепахи путешествуют еще дальше; кожаный экземпляр проплывает около 12 000 км (7 500 миль) до пляжей, где гнездится. [ 13 ] [ 89 ] Большинство черепах создают гнездо для своих яиц. Самки обычно выкапывают в субстрате камеру, похожую на колбу. Другие виды откладывают яйца в растительности или расщелинах. [ 93 ] Самки выбирают места для гнездования в зависимости от факторов окружающей среды, таких как температура и влажность, которые важны для развития эмбрионов. [ 89 ] В зависимости от вида количество откладываемых яиц варьируется от одного до более 100. Более крупные самки могут откладывать яйца большего количества или большего размера. По сравнению с пресноводными черепахами черепахи откладывают меньше, но более крупные яйца. Самки могут откладывать несколько кладок в течение сезона, особенно у видов, переживающих непредсказуемые муссоны . [ 94 ]

Большинство черепах-матерей не заботятся о своих родителях, а лишь закрывают яйца и немедленно уходят, хотя некоторые виды охраняют свои гнезда в течение нескольких дней или недель. [ 95 ] Яйца различаются между округлыми, овальными, удлиненными, а также с твердой и мягкой скорлупой. [ 96 ] У большинства видов пол определяется температурой . У некоторых видов при более высоких температурах рождаются самки, а при более низких — самцы, в то время как у других при более умеренных температурах рождаются самцы, а при экстремально высоких и холодных температурах рождаются самки. [ 13 ] Существуют экспериментальные доказательства того, что эмбрионы Mauremys reevesii могут перемещаться внутри своих яиц, выбирая лучшую температуру для развития, тем самым влияя на их половую судьбу. [ 97 ] У других видов пол определяется генетически . Продолжительность инкубации яиц черепах варьируется от двух до трех месяцев для видов умеренного климата и от четырех месяцев до более года для тропических видов. [ 13 ] Виды, обитающие в умеренно-теплом климате, могут задерживать свое развитие . [ 98 ]
Вылупившиеся молодые черепахи вырываются из панциря с помощью яичного зуба — острого выступа, который временно существует на их верхней части клюва. [ 13 ] [ 99 ] Птенцы выкапываются из гнезда и находят убежище в растительности или воде. Некоторые виды остаются в гнезде дольше, будь то перезимовка или ожидание дождя, который разрыхлит почву, чтобы их можно было выкопать. [ 13 ] Молодые черепахи очень уязвимы для хищников как в яйце, так и в вылупившемся состоянии. Смертность в этот период высока, но значительно снижается, когда они достигают совершеннолетия. Большинство видов растут быстро в первые годы жизни и замедляются, когда становятся зрелыми. [ 100 ]
Продолжительность жизни
Черепахи могут жить долго. Говорят, что самой старой из ныне живущих черепах и наземных животных является сейшельская гигантская черепаха по имени Джонатан , которой в 2019 году исполнилось 187 лет. [ 101 ] Галапагосская черепаха по имени Харриет была собрана Чарльзом Дарвином в 1835 году; он умер в 2006 году, прожив не менее 176 лет. Большинство диких черепах не достигают этого возраста. Черепахи каждый год отращивают новые щитки под предыдущими, что позволяет исследователям оценить, как долго они живут. [ 102 ] Они также стареют медленно . [ 103 ] Выживаемость взрослых черепах может достигать 99% в год. [ 13 ]
Систематика и эволюция
Ископаемая история

Зоологи пытались объяснить эволюционное происхождение черепах и, в частности, их уникальных панцирей. В 1914 году Ян Верслуис предположил, что костные пластинки в дерме, называемые остеодермами , срослись с ребрами под ними, позже названными Оливье Риппелем «Предком в горошек». [ 19 ] [ 104 ] Теория объясняла эволюцию ископаемых парейазавров от Брадисавра до Антодона , но не объясняла, как ребра могли прикрепиться к костным кожным пластинкам. [ 19 ]
Более поздние открытия нарисовали другой сценарий эволюции панциря черепахи. Стволовые не имели панцирей и пластронов , черепахи Eunotosaurus средней перми , Pappochelys среднего триаса и Eorhynchochelys позднего триаса но имели укороченное туловище, расширенные ребра и удлиненные спинные позвонки. Также в позднем триасе у Odontochelys была частичная раковина, состоящая из полного костного пластрона и неполного панциря. Развитие раковины завершилось у позднетриасового Proganochelys с полностью развитыми панцирем и пластроном. [ 19 ] [ 105 ] Адаптации, которые привели к эволюции панциря, возможно, изначально были связаны с рытьем и ископаемым образом жизни. [ 105 ]
Старейшими известными представителями линии Pleurodira являются Platychelyidae из поздней юры . [ 106 ] Самый старый известный однозначный криптодир — Sinaspideretes , близкий родственник мягкопанцирных черепах, из поздней юры Китая. [ 107 ] В течение позднего мела и кайнозоя представители семейств плевродиров Bothremydidae и Podocnemididae получили широкое распространение в Северном полушарии из-за их прибрежного образа жизни. [ 108 ] [ 109 ] Самые старые из известных черепах с мягким панцирем и морских черепах появились в раннем меловом периоде . [ 110 ] [ 111 ] Черепахи возникли в Азии в эоцене . [ 112 ] Поздно выжившая группа стволовых черепах, Meiolaniidae , выжила в Австралазии в плейстоцене и голоцене . [ 113 ]
Внешние отношения
Точное происхождение черепах является спорным. Считалось, что они были единственной сохранившейся ветвью древнего эволюционного класса Anapsida , в который входят такие группы, как проколофониды и парейазавры. У всех черепов анапсид нет височного отверстия , в то время как у всех других живых амниот височные отверстия есть. [ 114 ] Позже было высказано предположение, что анапсидоподобные черепа черепах могут возникнуть в результате обратной эволюции, а не в результате происхождения анапсид. [ 115 ] Ископаемые свидетельства показали, что ранние стволовые черепахи имели небольшие височные отверстия. [ 105 ]
Некоторые ранние морфологические филогенетические исследования поместили черепах ближе к лепидозавриям ( туатары , ящерицы и змеи ), чем к архозавриям ( крокодилы и птицы). [ 114 ] Напротив, некоторые молекулярные исследования относят черепах либо к архозавриям, либо к архозавриям. [ 116 ] или, что чаще, как сестринскую группу ныне живущим архозаврам, [ 115 ] [ 117 ] [ 118 ] [ 119 ] хотя анализ, проведенный Тайлером Лайсоном и его коллегами (2012), вместо этого обнаружил, что черепахи являются сестринской группой лепидозавров. [ 120 ] Иления Киари и коллеги (2012) проанализировали 248 ядерных генов 16 позвоночных и предположили, что черепахи имеют более недавнего общего предка с птицами и крокодилами. Дата разделения черепах, птиц и крокодилов оценивается в 255 миллионов лет назад, в пермский период. [ 121 ] Посредством (UCE) в масштабе генома филогенетического исследования ультраконсервативных элементов с целью выяснения места черепах среди рептилий Николас Кроуфорд и его коллеги (2012) аналогичным образом обнаружили, что черепахи ближе к птицам и крокодилам. [ 122 ]
Используя черновые (незаконченные) последовательности генома зеленой морской черепахи и китайской мягкопанцирной черепахи , Чжо Ван и его коллеги (2013) пришли к выводу, что черепахи, вероятно, являются сестринской группой крокодилов и птиц. [ 123 ] Внешняя филогения черепах показана на кладограмме ниже. [ 122 ]
| Диапсида |
| ||||||

Внутренние отношения
Современные черепахи и их вымершие родственники с полным панцирем отнесены к кладе Testudinata . [ 124 ] Самый последний общий предок современных черепах, соответствующий расколу между Pleurodira (виды с боковой шеей) и Cryptodira (виды со скрытой шеей), по оценкам, произошел около 210 миллионов лет назад в позднем триасе. [ 125 ] Роберт Томпсон и его коллеги (2021) отмечают, что современные черепахи имеют низкое разнообразие по сравнению с тем, как долго они существовали. Согласно их анализу, разнообразие было стабильным, за исключением одного быстрого увеличения на границе эоцена и олигоцена около 30 миллионов лет назад и крупного регионального вымирания примерно в то же время. Они предполагают, что глобальное изменение климата вызвало оба события, поскольку похолодание и высыхание привели к тому, что земля стала засушливой, а черепахи вымерли, в то время как новые континентальные окраины, открывшиеся в результате изменения климата, предоставили среду обитания для развития других видов. [ 126 ]
Кладограмма Николаса Кроуфорда и его коллег, 2015 г., показывает внутреннюю филогению Testudines вплоть до уровня семей . [ 127 ] [ 128 ] Анализ, проведенный Томпсоном и его коллегами в 2021 году, подтверждает ту же структуру вплоть до уровня семьи. [ 126 ]
| Черепахи |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Различия между двумя подотрядами



Черепахи делятся на два живых подотряда: Cryptodira и Pleurodira. [ 129 ] Эти две группы различаются тем, как шея втягивается для защиты. Плевродиран втягивают шею в сторону и перед плечевым поясом, тогда как криптодираны втягивают шею назад в панцирь. Эти движения обеспечиваются морфологией и расположением шейных позвонков. [ 130 ] [ 131 ] Морские черепахи (относящиеся к криптодирам) по большей части утратили способность втягивать голову. [ 132 ]
Приводящие мышцы нижней челюсти в обеих подгруппах образуют шкивную систему. Однако кости, с которыми сочленяются мышцы, различаются. У Pleurodira шкив образован крыловидными костями неба . , а у Cryptodira шкив образован слуховой капсулой Обе системы помогают вертикально перенаправить приводящие мышцы и сохранить мощный прикус. [ 133 ]
Еще одним отличием подотрядов является прикрепление таза. У Криптодиры таз свободный, соединен с панцирем только связками. У Pleurodira таз пришивается костными соединениями к панцирю и пластрону, образуя пару больших костных столбцов на заднем конце черепахи, соединяющих две части панциря. [ 134 ]
Распространение и среда обитания
Черепахи широко распространены на континентах, океанах и островах мира, встречаются наземные, полностью водные и полуводные виды. Морские черепахи в основном обитают в тропических и субтропических регионах, но кожистых черепах можно встретить и в более холодных районах Атлантического и Тихого океана. [ 135 ] Все живые плевродиры обитают в пресной воде и встречаются только в Южном полушарии. [ 136 ] Криптодиры включают наземные, пресноводные и морские виды, и их распространение более широкое. [ 135 ] Регионы мира, богатые неморскими видами черепах, — это бассейн Амазонки, водосборный бассейн Мексиканского залива в США, а также некоторые районы Южной и Юго-Восточной Азии. [ 137 ]
Распространение черепах в более холодном климате ограничено ограничениями на воспроизводство, которое снижается из-за длительной спячки. Североамериканские виды едва распространены за пределами южной границы Канады. [ 138 ] Некоторые черепахи встречаются на больших высотах, например, вид Terrapene ornata встречается на высоте до 2000 м (6600 футов) в Нью-Мексико. [ 139 ] И наоборот, кожистая морская черепаха может нырять на глубину более 1200 м (3900 футов). [ 140 ] Виды рода Gopherus могут переносить как температуру тела ниже нуля, так и температуру тела выше 40 ° C (104 ° F), хотя наиболее активны они при температуре 26–34 ° C (79–93 ° F). [ 141 ]
Сохранение

Среди отрядов позвоночных черепахи уступают только приматам по проценту видов, находящихся под угрозой исчезновения. С 1500 года нашей эры существовало 360 современных видов. Из них 51–56% считаются находящимися под угрозой исчезновения, а 60% считаются находящимися под угрозой исчезновения или вымершими. [ 144 ] Черепахи сталкиваются со многими угрозами, включая разрушение среды обитания , сбор урожая для потребления, торговлю домашними животными, [ 145 ] [ 146 ] световое загрязнение , [ 147 ] и изменение климата . [ 148 ] Азиатские виды подвергаются особенно высокому риску исчезновения, в первую очередь из-за их долгосрочной нерациональной эксплуатации для производства продуктов питания и лекарств. [ 149 ] и около 83% видов неморских черепах в Азии считаются находящимися под угрозой исчезновения. [ 144 ] По состоянию на 2021 год вымирание черепах прогрессирует гораздо быстрее, чем во время мел-третичного вымирания . При таких темпах все черепахи могут вымереть через несколько столетий. [ 150 ]
Черепаховые инкубаторы могут быть созданы, когда защита от наводнений, эрозии, хищничества или масштабного браконьерства . требуется [ 151 ] [ 152 ] [ 153 ] Китайские рынки стремились удовлетворить растущий спрос на мясо черепах за счет выращенных на фермах черепах. По оценкам, в 2007 году в Китае действовало более тысячи черепаховых ферм. [ 154 ] Тем не менее, диких черепах продолжают ловить и отправлять на рынок в больших количествах, что приводит к тому, что защитники природы назвали «кризисом азиатских черепах». [ 155 ] [ 149 ] По словам биолога Джорджа Амато, охота на черепах «высосала целые виды из районов Юго-Восточной Азии», хотя биологи до сих пор не знали, сколько видов обитает в этом регионе. [ 156 ] В 2000 году все азиатские коробчатые черепахи были внесены в СИТЕС . список исчезающих видов [ 149 ]
Добыча диких черепах разрешена законом в некоторых американских штатах. [ 157 ] и в Китае растет спрос на американских черепах. [ 158 ] [ 159 ] По оценкам Комиссии по охране рыбы и дикой природы Флориды , в 2008 году около 3000 фунтов мягкопанцирных черепах еженедельно экспортировалось через международный аэропорт Тампы . [ 159 ] Однако подавляющее большинство черепах, экспортированных из США в период с 2002 по 2005 год, были выращены на фермах. [ 158 ]
Большое количество морских черепах случайно погибает в ярусах , жаберных и траловых сетях в качестве прилова . Исследование 2010 года показало, что в период с 1990 по 2008 год было убито более 8 миллионов человек; Восточная часть Тихого океана и Средиземноморье были определены как наиболее пострадавшие районы. [ 142 ] С 1980-х годов Соединенные Штаты требуют, чтобы все траулеры для ловли креветок оснащали свои сети устройствами для защиты черепах , которые не позволяют черепахам запутываться в сетях и утонуть. [ 143 ] В более локальном масштабе на морских черепах влияет и другая деятельность человека. В Австралии Квинсленде в в рамках программы отлова акул , в которой используются сети для акул и барабанные лески , в период с 1962 по 2015 год в качестве прилова погибло более 5000 черепах; в том числе 719 черепах-головачей и 33 морских черепахи-ястребов, которые занесены в список находящихся под угрозой исчезновения. [ 160 ]
Местным популяциям черепах также могут угрожать инвазивные виды . Центрально-Североамериканская красноухая черепаха-ползун была включена в список « наихудших инвазивных видов в мире », а домашняя черепаха была выпущена на свободу во всем мире. Похоже, они конкурируют с местными видами черепах в восточной и западной части Северной Америки, Европы и Японии. [ 161 ] [ 162 ]
Человеческое использование
В космических полетах
Советского Союза в сентябре 1968 года на корабле «Зонд-5» Две черепахи участвовали в облете Луны , что сделало их первыми земными живыми существами, совершившими путешествие в окрестности Луны. Черепахи также участвовали на зондах «Зонд-6» (1968 г.) и « Зонд-7» (1969 г.). в облетах Луны [ 163 ] [ 164 ]
В культуре
-

-
 Мир покоится на четырех слонах на спине Мировой Черепахи . Западное изображение «Индуистской Земли», 1877 г.
Мир покоится на четырех слонах на спине Мировой Черепахи . Западное изображение «Индуистской Земли», 1877 г. -

-

-
 Плакат к постановке «Черепахи» 1898 года в Манхэттенском театре на Бродвее
Плакат к постановке «Черепахи» 1898 года в Манхэттенском театре на Бродвее -
 Погремушки на ногах из панциря черепахи, которые носила главная чероки , 20 век. танцовщица
Погремушки на ногах из панциря черепахи, которые носила главная чероки , 20 век. танцовщица





Черепахи присутствовали в человеческих культурах по всему миру с древних времен. Обычно к ним относятся положительно, несмотря на то, что они не являются «приятными» или яркими; их связь с древностью и старостью способствовала созданию их привлекательного имиджа. [ 165 ]
В индуистской мифологии Мировая Черепаха по имени Курма или Каччапа держит на своей спине четырех слонов; они, в свою очередь, несут на своих спинах тяжесть всего мира. [ 166 ] [ 167 ] Черепаха — один из десяти аватаров или воплощений бога Вишну . [ 166 ] Поза йоги Курмасана названа в честь аватара. [ 168 ] [ 169 ] Мировые черепахи встречаются в культурах коренных американцев, включая алгонкинов , ирокезов и ленапе . Они рассказывают множество версий истории создания Черепашьего острова . По одной из версий, Ондатра насыпала землю на спину Черепахи, создав континент Северная Америка. В версии ирокезов ; беременная Небесная Женщина проваливается в дыру в небе между корнями дерева, где ее ловят птицы, которые благополучно приземляют ее на спину Черепахи Земля растет вокруг нее. Черепаха здесь альтруистична, но мир — тяжкое бремя, и черепаха иногда встряхивается, чтобы облегчить бремя, вызывая землетрясения. [ 166 ] [ 170 ] [ 171 ]
Черепаха была символом древнего месопотамского бога Энки начиная с 3-го тысячелетия до нашей эры. [ 172 ] Древнегреческий миф о происхождении гласил, что только черепаха отказалась от приглашения богов Зевса и Геры на их свадьбу, так как предпочла остаться дома. Затем Зевс приказал ему навсегда унести с собой свой дом. [ 173 ] Другой их бог, Гермес , изобрел семиструнную лиру, сделанную из панциря черепахи. [ 174 ] В времен династии Шан китайской практике пластромантии , восходящей к 1200 году до нашей эры, оракулы получали путем написания вопросов на пластронах черепах с использованием древнейшей известной формы китайских иероглифов , сжигания пластрона и интерпретации полученных трещин. Позже черепаха была одним из четырех священных животных в конфуцианстве , а в Хань период стелы устанавливались на вершинах каменных черепах, позже связанных с Биси , сыном Короля Драконов с черепаховым панцирем. [ 175 ] Морские черепахи занимают важное место в искусстве австралийских аборигенов . [ 167 ] Армия Древнего Рима использовала построение тестудо («черепаха») , где солдаты образовывали стену из щитов для защиты. [ 162 ]
В «Баснях» Эзопа « Черепаха и Заяц » рассказывается, как в неравной гонке может выиграть более медленный партнер. [ 176 ] [ 177 ] В книге Льюиса Кэрролла 1865 года «Приключения Алисы в стране чудес» фигурирует ложная черепаха , названная в честь супа, призванного имитировать дорогой суп, приготовленный из настоящего черепашьего мяса. [ 178 ] [ 179 ] [ 180 ] В 1896 году французский драматург Леон Гандилло написал комедию в трех действиях под названием «Тортю» , ставшую «парижской сенсацией». [ 181 ] шел во Франции и появился в Манхэттенском театре на Бродвее, Нью-Йорк, в 1898 году как «Черепаха» . [ 182 ] «Космическая черепаха» и мотив острова вновь появляются в романе Гэри Снайдера 1974 года «Черепаший остров» и снова в Пратчетта Терри серии «Плоский мир» под названием «Великий А'Туин», начиная с романа 1983 года «Цвет волшебства» . Предположительно, это вид Chelys galactica , галактической черепахи с четырьмя слонами на спине, поддерживающими Плоский мир. [ 183 ] Гигантская огнедышащая черепаха по имени Гамера — звезда серии японских фильмов о монстрах в жанре кайдзю , с 1965 по 2006 год снявшаяся в двенадцати фильмах. [ 184 ] Черепахи были показаны в комиксах и мультфильмах, таких как « Черепашки-ниндзя» 1984 года . [ 185 ] [ 186 ]
Как домашние животные
Некоторые черепахи, особенно мелкие наземные и пресноводные виды, содержатся в качестве домашних животных . [ 187 ] [ 188 ] Спрос на домашних черепах увеличился в 1950-х годах, когда США были основным поставщиком, особенно красноухих черепах, выращенных на фермах. Популярность экзотических домашних животных привела к росту незаконной торговли дикими животными . Около 21% стоимости торговли живыми животными приходится на рептилий, а черепахи являются одними из наиболее популярных видов торговли. [ 189 ] Плохое содержание черепах может вызвать хронический ринит (отечность носа), заросшие клювы, гиперпаратиреоз (который размягчает их скелет), запоры , различные репродуктивные проблемы и травмы от собак. [ 187 ] В начале 20-го века люди в Соединенных Штатах организовывали черепашьи бега и играли в них . [ 190 ]
В качестве еды и других целей
Мясо пойманных диких черепах продолжают употреблять в пищу в азиатских культурах. [ 191 ] а черепаховый суп когда-то был популярным блюдом английской кухни . [ 192 ] Рагу из черепахи-суслика популярно среди некоторых групп во Флориде. [ 193 ] Предполагаемые афродизиаки или лечебные свойства черепашьих яиц создали широкую торговлю ими в Юго-Восточной Азии. [ 167 ] Пластроны черепах с твердым панцирем и панцири с мягким панцирем широко используются в традиционной китайской медицине ; Тайвань импортировал около 200 тонн твердых ракушек из своих соседей ежегодно с 1999 по 2008 год. [ 194 ] Популярным лекарственным препаратом на основе трав и панцирей черепах является гулингао . желе [ 195 ] Вещество черепахового панциря , обычно добываемое черепахой-ястребом, веками использовалось для изготовления ювелирных изделий, инструментов и украшений в западной части Тихого океана. [ 167 ] Соответственно, на ястребов охотились из-за их панцирей. [ 196 ] Торговля черепаховым панцирем была запрещена на международном уровне СИТЕС в 1977 году. [ 197 ] Некоторые культуры использовали панцири черепах для создания музыки: шаманы коренных американцев превратили их в церемониальные погремушки, а ацтеки , майя и микстеки сделали айотль . барабаны [ 198 ]
-
 Ловля черепах в Австралии, 1875 год.
Ловля черепах в Австралии, 1875 год. -
 Черепахи продаются в качестве еды в Канаде, 2007 г.
Черепахи продаются в качестве еды в Канаде, 2007 г. -
 Пластроны черепахи для традиционной китайской медицины
Пластроны черепахи для традиционной китайской медицины -
 черепаховый гребешок; материал был дорогим и декоративным и широко использовался для изготовления мелких предметов. [ 199 ]
черепаховый гребешок; материал был дорогим и декоративным и широко использовался для изготовления мелких предметов. [ 199 ] -
 Домашний красноухий ползунок греется на плавучей платформе под солнечной лампой.
Домашний красноухий ползунок греется на плавучей платформе под солнечной лампой. -
 Уведомление в Блуфилдсе , Никарагуа, в 2023 году, указывающее, что отлов и употребление в пищу черепах запрещены.
Уведомление в Блуфилдсе , Никарагуа, в 2023 году, указывающее, что отлов и употребление в пищу черепах запрещены.






См. также
Ссылки
Цитаты
- ^ Перейти обратно: а б с Рабочая группа по таксономии черепах (2017). Черепахи мира: аннотированный контрольный список и атлас таксономии, синонимии, распространения и статуса охраны (PDF) . Челонианские исследовательские монографии (№ 1). Том. 7 (8-е изд.). Челонианский исследовательский фонд: Охрана черепах. стр. 10, 24. doi : 10.3854/crm.7.checklist.atlas.v8.2017 . ISBN 978-1-5323-5026-9 . OCLC 1124067380 . Архивировано (PDF) из оригинала 25 февраля 2021 г. Проверено 20 января 2018 г.
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Перейти обратно: а б с Дюбуа, Алан; Бур, Роджер (2010). «Различие между номенклатурой семейных и классовых серий в зоологической номенклатуре с упором на номинацию, созданную Батчем (1788, 1789), и на высшую номенклатуру черепах» (PDF) . Боннский зоологический бюллетень . 57 (2): 149–171. Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Харпер, Дуглас. «черепаха» . Интернет-словарь этимологии . Проверено 28 октября 2021 г.
- ^ Оренштейн 2012 , с. 9.
- ^ тестудо . Чарльтон Т. Льюис и Чарльз Шорт. Латинский словарь по проекту «Персей» .
- ^ χελώνη . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ «Челония» . Мерриам-Вебстер . Проверено 29 августа 2021 г.
- ^ Франклин 2011 , с. 14.
- ^ Чен, Ирен Х.; Ян, Вэнь; Мейерс, Марк А. (2015). «Панцирь морской черепахи в коже: прочная и гибкая биологическая конструкция» . Акта Биоматериалы . 28 : 2–12. doi : 10.1016/j.actbio.2015.09.023 . ISSN 1742-7061 . ПМИД 26391496 .
- ^ « Архелон » . Институт геологических исследований Блэк-Хиллз. Архивировано из оригинала 12 марта 2016 года . Проверено 23 декабря 2018 г.
- ^ Бонен, Франк; Дево, Бернар; Дюпре, Ален (2006). Черепахи мира . Издательство Университета Джонса Хопкинса. п. 230. ИСБН 978-0-8018-8496-2 .
- ^ Фрейзер, Джек (2020). «Галапогосские черепахи: главные герои спектакля жизни на Земле». В Гиббсе, Джеймс; Кайот, Линда; Агилера, Вашингтон Тапиа (ред.). Галапагосские гигантские черепахи . Академическая пресса. п. 26. ISBN 978-0-12-817554-5 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Айверсон, Джон; Молл, Эдвард О (2002). «Черепахи и черепахи». В Холлидее, Тим; Адлер, Крейг (ред.). Энциклопедия рептилий и амфибий Firefly . Книги Светлячка. стр. 118–129. ISBN 978-1-55297-613-5 .
- ^ Оренштейн 2012 , с. 22.
- ^ Хатчинсон, Дж. Ховард (1996). «Введение в Testudines: Черепахи» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 29 июня 2016 года . Проверено 4 июня 2003 г.
- ^ Перейти обратно: а б с Оренштейн 2012 , с. 16.
- ^ Перейти обратно: а б Себра-Томас, Джудит; Тан, Фрейзер; Систла, Сита; Эстес, Эйлин; Бендер, Гюнеш; Ким, Кристина; Риччио, Пол; Гилберт, Скотт Ф. (2005). «Как черепаха формирует свой панцирь: паракринная гипотеза формирования панциря» (PDF) . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 304Б (6): 558–569. Бибкод : 2005JEZB..304..558C . дои : 10.1002/jez.b.21059 . ISSN 1552-5007 . ПМИД 15968684 . S2CID 2484583 .
- ^ Гаффни, Юджин С. (1990). «Сравнительная остеология триасовой черепахи Proganochelys » . Бюллетень Американского музея естественной истории (194): 1–263. hdl : 2246/884 . OCLC 263164288 . Архивировано из оригинала 16 мая 2021 года . Проверено 16 мая 2021 г.
- ^ Перейти обратно: а б с д и Шох, Райнер Р.; Сьюс, Ханс-Дитер; Бенсон, Роджер (2019). «Происхождение строения тела черепахи: данные окаменелостей и эмбрионов» . Палеонтология . 63 (3): 375–393. дои : 10.1111/пала.12460 . ISSN 0031-0239 .
- ^ Оренштейн 2012 , стр. 16–17.
- ^ Причард, Питер Ч. (2008). «Эволюция и строение панциря черепахи». В Винекене, Жанетт; Белс, В.Л.; Годфри, Мэтью Х. (ред.). Биология черепах . ЦРК Пресс. п. 56. ИСБН 978-0-8493-3339-2 . OCLC 144570900 .
- ^ Оренштейн 2012 , стр. 22–26.
- ^ Оренштейн 2012 , стр. 22–23, 26–27.
- ^ Перейти обратно: а б Франклин 2011 , с. 18.
- ^ Перейти обратно: а б Оренштейн 2012 , с. 33.
- ^ Перейти обратно: а б с д Франклин 2011 , с. 28.
- ^ Херрел, Энтони; О'Рейли, Джеймс С.; Ричмонд, Алан М. (2002). «Эволюция способности укуса черепах». Журнал эволюционной биологии . 15 (6): 1083–1094. CiteSeerX 10.1.1.484.5540 . дои : 10.1046/j.1420-9101.2002.00459.x . S2CID 54067445 .
- ^ Оренштейн 2012 , стр. 33–34.
- ^ Оренштейн 2012 , с. 34.
- ^ Абрамян Дж.; Ричман, Дж. М. (2015). «Недавние открытия о морфологическом разнообразии первичного и вторичного неба амниот» . Динамика развития . 244 (12): 1457–1468. дои : 10.1002/dvdy.24338 . ПМЦ 4715671 . ПМИД 26293818 .
- ^ Перейти обратно: а б с Оренштейн 2012 , с. 36.
- ^ П. Рассел, Энтони; М. Бауэр, Аарон (2020). «Вокализация современных нептичьих рептилий: синтетический обзор фонации и голосового аппарата» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 304 (7): 1478–1528. дои : 10.1002/ar.24553 . ПМИД 33099849 . S2CID 225069598 .
- ^ Кэпшоу, Грейс; Уиллис, Кэти Л.; Хан, Давэй; Бирман, Хилари С. (2020). «Продукция и восприятие звука рептилий». В Розенфельде, Шерил С.; Хоффманн, Фрауке (ред.). Нейроэндокринная регуляция вокализации животных . Академическая пресса. стр. 101–118. ISBN 978-0-12-815160-0 .
- ^ Оренштейн 2012 , с. 38.
- ^ Перейти обратно: а б Давенпорт, Джон; Манкс, Сара А.; Оксфорд, Пи-Джей (22 февраля 1984 г.). «Сравнение плавания морских и пресноводных черепах». Труды Лондонского королевского общества. Серия Б. Биологические науки . 220 (1221): 447–475. Бибкод : 1984РСПСБ.220..447Д . дои : 10.1098/rspb.1984.0013 . ISSN 0080-4649 . JSTOR 35758 . S2CID 84615412 .
- ^ Оренштейн 2012 , стр. 38–40.
- ^ Оренштейн 2012 , с. 40.
- ^ Перейти обратно: а б с Фричес, Керстин А.; Ордер, Эрик Дж. (2013). "Зрение". В Винекене, Жанетт (ред.). Биология морских черепах . ЦРК Пресс. стр. 31–58. ISBN 978-1-4398-7308-3 . OCLC 828509848 .
- ^ Гранда, Алан М.; Дворжак, Чарльз А. (1977). «Видение у черепах». Зрительная система позвоночных . Справочник по сенсорной физиологии. Том. 7 / 5. Берлин, Гейдельберг: Springer Berlin Heidelberg. стр. 451–495. дои : 10.1007/978-3-642-66468-7_8 . ISBN 978-3-642-66470-0 . ISSN 0072-9906 .
- ^ Джейкобс, Джеральд (1981). «Рептилии: Черепаха». Сравнительное цветовое зрение . Нью-Йорк: Академическая пресса. стр. 102–105. ISBN 978-0-12-378520-6 .
- ^ Уиллис, Кэти Л. (2016). «Подводный слух черепах». Влияние шума на водную жизнь II . Достижения экспериментальной медицины и биологии. Том. 875. стр. 1229–1235. дои : 10.1007/978-1-4939-2981-8_154 . ISBN 978-1-4939-2980-1 . ПМИД 26611091 .
- ^ Мартин, Келли Дж.; Алесси, Сара С.; Гаспар, Джозеф К.; Такер, Антон Д.; Бауэр, Гордон Б.; Манн, Дэвид А. (2012). «Подводный слух черепахи-логгерхеда ( Caretta caretta ): сравнение поведенческих и слуховых аудиограмм вызванного потенциала» . Журнал экспериментальной биологии . 215 (17): 3001–3009. дои : 10.1242/jeb.066324 . ISSN 1477-9145 . ПМИД 22875768 . S2CID 459652 .
- ^ Оренштейн 2012 , стр. 35–36.
- ^ Мэнтон, Мэрион; Карр, Эндрю; Эренфельд, Дэвид В. (1972). «Хеморецепция у перелетной морской черепахи Chelonia mydas » . Биологический вестник . 143 (1): 184–195. дои : 10.2307/1540338 . ISSN 0006-3185 . JSTOR 1540338 . Архивировано из оригинала 6 июня 2021 года . Проверено 6 июня 2021 г.
- ^ Перейти обратно: а б Кордейру, Табата Э.Ф.; Абэ, Аугусто С.; Кляйн, Вильфрид (апрель 2016 г.). «Вентиляция и газообмен у двух черепах: Podocnemis unifilis и Phrynops geoffroanus (Testudines: Pleurodira)» (PDF) . Респираторная физиология и нейробиология . 224 : 125–131. дои : 10.1016/j.resp.2014.12.010 . hdl : 11449/158795 . ISSN 1569-9048 . ПМИД 25534144 . S2CID 37446604 . Архивировано из оригинала 24 июля 2021 года . Проверено 25 сентября 2019 г.
- ^ Перейти обратно: а б с Лайсон, Тайлер Р.; Шахнер, Эмма Р.; Бота-Бринк, Дженнифер; Шайер, Торстен М.; Ламбертц, Маркус; Бевер, Г.С.; Рубидж, Брюс С.; де Кейроз, Кевин (7 ноября 2014 г.). «Происхождение уникального дыхательного аппарата черепах» . Природные коммуникации . 5 : 5211. Бибкод : 2014NatCo...5.5211L . дои : 10.1038/ncomms6211 . ISSN 2041-1723 . ПМИД 25376734 .
- ^ Ли, Стелла Ю.; Милсом, Уильям К. (2016). «Метаболические затраты на дыхание у красноухих ползунков: попытка разрешить старый спор». Респираторная физиология и нейробиология . 224 : 114–124. дои : 10.1016/j.resp.2015.10.011 . ISSN 1569-9048 . ПМИД 26524718 . S2CID 5194890 .
- ^ Оренштейн 2012 , с. 41.
- ^ Ламбертц, Маркус; Бёме, Вольфганг; Перри, Стивен Ф. (июль 2010 г.). «Анатомия дыхательной системы Platysternon megacephalum Grey, 1831 (Testudines: Cryptodira) и родственных видов, а также ее филогенетические последствия». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 156 (3): 330–336. дои : 10.1016/j.cbpa.2009.12.016 . ISSN 1095-6433 . ПМИД 20044019 .
- ^ Кляйн, Вильфрид; Кодд, Джонатан Р. (2010). «Дыхание и передвижение: сравнительная анатомия, морфология и функции». Респираторная физиология и нейробиология . 173 : С26–С32. дои : 10.1016/j.resp.2010.04.019 . ISSN 1569-9048 . ПМИД 20417316 . S2CID 28044326 .
- ^ Морера-Бренес, Берналь; Монге-Нахера, Хулиан (2011). «Периоды погружения четырех неотропических черепах» . Исследовательский журнал UNED . 3 (1): 97. дои : 10.22458/urj.v3i1.212 . Архивировано из оригинала 9 августа 2020 года . Проверено 11 июня 2020 г.
- ^ Священник Тони Э.; Франклин, Крейг Э. (декабрь 2002 г.). «Влияние температуры воды и уровня кислорода на поведение при нырянии двух пресноводных черепах: Rheodytes leukops и Emydura macquarii ». Журнал герпетологии . 36 (4): 555–561. doi : 10.1670/0022-1511(2002)036[0555:EOWTAO]2.0.CO;2 . ISSN 0022-1511 . JSTOR 1565924 . S2CID 85279910 .
- ^ Перейти обратно: а б с д Винекен, Жанетт (2008). «Структура сердечно-легочной системы черепах: последствия для поведения и функций». В Винекене, Жанетт; Белс, В.Л.; Годфри, Мэтью Х. (ред.). Биология черепах . ЦРК Пресс. стр. 213–224. ISBN 978-0-8493-3339-2 . OCLC 144570900 .
- ^ Джексон, Дональд С. (2002). «Спячка без кислорода: физиологическая адаптация расписной черепахи» . Журнал физиологии . 543 (Часть 3): 731–737. дои : 10.1113/jphysicalol.2002.024729 . ПМК 2290531 . ПМИД 12231634 .
- ^ Миллер, Джеффри Д.; Динкелакер, Стивен А. (2008). «Репродуктивные структуры и стратегии черепах». В Винекене, Жанетт; Белс, В.Л.; Годфри, Мэтью Х. (ред.). Биология черепах . ЦРК Пресс. п. 234. ИСБН 978-0-8493-3339-2 . OCLC 144570900 .
- ^ Винекен, Жанетт; Уизерингтон, Дон (февраль 2015 г.). «Мочеполовая система» (PDF) . Анатомия морских черепах . 1 : 153–165. Архивировано (PDF) из оригинала 8 июня 2019 г. Проверено 18 мая 2021 г.
- ^ Бентли, Питер Дж. (2013). Эндокринная система и осморегуляция: сравнительный анализ позвоночных . Спрингер. п. 143. ИСБН 978-3-662-05014-9 . Архивировано из оригинала 31 мая 2021 года . Проверено 18 мая 2021 г.
- ^ Франклин 2011 , с. 31.
- ^ Перейти обратно: а б Пау, Ф. Харви; Дженис, Кристин М. (2019). «16. Черепахи». Жизнь позвоночных (10-е изд.). Нью-Йорк: Sinauer Associates. стр. 283–299. ISBN 978-1-60535-607-5 . OCLC 1022979490 .
- ^ Оренштейн 2012 , с. 231.
- ^ Франклин 2011 , стр. 29–30.
- ^ Оренштейн 2012 , с. 237.
- ^ Оренштейн 2012 , с. 235.
- ^ Ван Дамм, Йохан; Аэртс, Питер (1997). «Кинематика и функциональная морфология водного питания австралийских змеешеих черепах (Pleurodira; Chelodina)». Журнал морфологии . 233 (2): 113–125. doi : 10.1002/(SICI)1097-4687(199708)233:2<113::AID-JMOR3>3.0.CO;2-7 . ПМИД 9218349 . S2CID 32906130 .
- ^ Франклин 2011 , с. 30.
- ^ Оренштейн 2012 , с. 239.
- ^ Оренштейн 2012 , с. 229.
- ^ Перейти обратно: а б Джайлз, Жаклин С.; Дэвис, Дженни; МакКоли, Роберт Д.; Кучлинг, Джеральд (2009). «Голос черепахи: подводный акустический репертуар длинношеей пресноводной черепахи Chelodina oblonga » . Журнал Акустического общества Америки . 126 (1): 434–443. Бибкод : 2009ASAJ..126..434G . дои : 10.1121/1.3148209 . ПМИД 19603900 .
- ^ Прайк 2021 , с. 39.
- ^ Перейти обратно: а б с Феррара, Камила Р.; Фогт, Ричард К.; Соуза-Лима, Рената Санторо (2012). «Черепашьи вокализации как первое свидетельство родительской заботы у хелонов после вылупления» (PDF) . Журнал сравнительной психологии . 127 (1): 24–32. дои : 10.1037/a0029656 . ПМИД 23088649 . Архивировано (PDF) из оригинала 2 сентября 2017 г. Проверено 1 сентября 2017 г.
- ^ Йоргевич-Коэн, Г; и др. (2022). «Общее эволюционное происхождение акустической коммуникации у хоановых позвоночных» . Природные коммуникации . 13 (1): 6089. Бибкод : 2022NatCo..13.6089J . дои : 10.1038/s41467-022-33741-8 . ПМЦ 9596459 . ПМИД 36284092 . S2CID 253111242 .
- ^ Бургхардт, Гордон М.; Уорд, Б.; Росско, Роджер (1996). «Проблема игр рептилий: обогащение окружающей среды и игровое поведение содержащихся в неволе нильских черепах с мягким панцирем, Trionyx triunguis ». Зоопарковая биология . 15 (3): 223–238. doi : 10.1002/(SICI)1098-2361(1996)15:3<223::AID-ZOO3>3.0.CO;2-D .
- ^ Дэвис, КМ; Бургхардт, Гордон М. (2007). «Обучение и долговременная память о новой задаче по добыче пищи у черепахи ( Pseudemys nelsoni )». Поведенческие процессы . 75 (2): 225–230. doi : 10.1016/j.beproc.2007.02.021 . ПМИД 17433570 . S2CID 34130920 .
- ^ «Рептилии, известные как «живые камни», демонстрируют удивительные когнитивные способности» . Природа . Поведение животных. 576 (7785): 10. 29 ноября 2019 г. Бибкод : 2019Natur.576...10. . дои : 10.1038/d41586-019-03655-5 . S2CID 208613023 .
- ^ Гутник, Тамар; Вайсенбахер, Антон; Куба, Майкл Дж. (13 ноября 2019 г.). «Недооцененные гиганты: оперантное обучение, зрительная дискриминация и долговременная память гигантских черепах» . Познание животных . 23 (1): 159–167. дои : 10.1007/s10071-019-01326-6 . ISSN 1435-9456 . ПМИД 31720927 . S2CID 207962281 .
- ^ Перейти обратно: а б Салас, Косме; Брольо, Кристина; Родригес, Фернандо (2003). «Эволюция переднего мозга и пространственного познания у позвоночных: сохранение разнообразия» . Мозг, поведение и эволюция . 62 (2): 72–82. дои : 10.1159/000072438 . ISSN 0006-8977 . ПМИД 12937346 . S2CID 23055468 .
- ^ Рейтер, Сэм; Лиау, Хуа-Пэн; Ямаваки, Трейси М.; Науманн, Роберт К.; Лоран, Жиль (2017). «О значении мозга рептилий для картирования эволюции гиппокампа» . Мозг, поведение и эволюция . 90 (1): 41–52. дои : 10.1159/000478693 . ISSN 0006-8977 . ПМИД 28866680 . S2CID 13763864 .
- ^ Оренштейн 2012 , стр. 252–253.
- ^ Франклин 2011 , с. 40.
- ^ Оренштейн 2012 , стр. 252–253, 301.
- ^ Саутвуд, Аманда; Авенс, Лариса (2009). «Физиологические, поведенческие и экологические аспекты миграции рептилий» . Журнал сравнительной физиологии Б. 180 (1): 1–23. дои : 10.1007/s00360-009-0415-8 . ISSN 0174-1578 . ПМИД 19847440 . S2CID 20245401 . Архивировано из оригинала 24 июля 2021 года . Проверено 6 июня 2021 г.
- ^ Перейти обратно: а б Ломанн, Кеннет Дж.; Ломанн, Кэтрин М.Ф.; Братья, Дж. Роджер; Путман, Натан Ф. (2013). «Натальное самонаведение и импринтинг у морских черепах». В Винекене, Жанетт (ред.). Биология морских черепах . ЦРК Пресс. стр. 59–78. ISBN 978-1-4398-7308-3 . OCLC 828509848 .
- ^ Перейти обратно: а б Пирс, Девон Э. (2001). «Системы спаривания черепах: поведение, хранение спермы и генетическое отцовство» . Журнал наследственности . 92 (2): 206–211. дои : 10.1093/jhered/92.2.206 . ПМИД 11396580 .
- ^ Перейти обратно: а б Бут, Джули; Питерс, Джеймс А. (1972). «Поведенческие исследования зеленой черепахи ( Chelonia mydas ) в море». Поведение животных . 20 (4): 808–812. дои : 10.1016/S0003-3472(72)80155-6 .
- ^ Берри, Джеймс Ф.; Шайн, Ричард (1980). «Половой диморфизм размеров и половой отбор черепах (отряд Testudines)». Экология . 44 (2): 185–191. Бибкод : 1980Oecol..44..185B . дои : 10.1007/bf00572678 . ПМИД 28310555 . S2CID 2456783 .
- ^ Берри, Джеймс; Айверсон, Джон (декабрь 2011 г.). « Kinosternon scorpioides (Линней 1766) - Грязевая черепаха-скорпион» (PDF) . Биология сохранения пресноводных черепах и черепах : 063.1–063.15. doi : 10.3854/crm.5.063.scorpioides.v1.2011 . ISBN 978-0-9653540-9-7 . Архивировано (PDF) из оригинала 4 февраля 2019 г. Проверено 20 августа 2019 г.
- ^ Оренштейн 2012 , стр. 270–271.
- ^ Оренштейн 2012 , с. 270.
- ^ Перейти обратно: а б с Франклин 2011 , с. 33.
- ^ Риппл, Дж. (1996). Морские черепахи . Путешественник Пресс. п. 26. ISBN 978-0-89658-315-3 .
- ^ Франклин 2011 , стр. 20–22.
- ^ Оренштейн 2012 , с. 260.
- ^ Оренштейн 2012 , с. 277.
- ^ Оренштейн 2012 , стр. 273, 276.
- ^ Франклин 2011 , с. 37.
- ^ Оренштейн 2012 , с. 274.
- ^ Йе, Инь-Цзы; Ма, Лян; Сунь, Бао-Цзюнь; Ли, Дэн; Ван, Ян; Сияй, Ричард; Ду, Вэй-Го (2019). «Эмбрионы черепах могут влиять на их собственную сексуальную судьбу» . Современная биология . 29 (16): 2597–2603.e4. Бибкод : 2019CBio...29E2597Y . дои : 10.1016/j.cub.2019.06.038 . ISSN 0960-9822 . ПМИД 31378606 .
- ^ Оренштейн 2012 , с. 286.
- ^ Оренштейн 2012 , с. 294.
- ^ Оренштейн 2012 , стр. 301–302.
- ^ Миллуорд, Адам (27 февраля 2019 г.). «Представляем Джонатана, самое старое животное в мире на суше, ему 187 лет» . Книги рекордов Гиннесса . Архивировано из оригинала 9 августа 2020 года . Проверено 14 июля 2020 г.
- ^ Франклин 2011 , стр. 44–45.
- ^ Уорнер, Дэниел А.; Миллер, Дэвид А.В.; Брониковски, Энн М.; Янзен, Фредрик Дж. (2016). «Десятилетия полевых данных показывают, что черепахи стареют в дикой природе» . ПНАС . 113 (23): 6502–6507. Бибкод : 2016PNAS..113.6502W . дои : 10.1073/pnas.1600035113 . ПМЦ 4988574 . ПМИД 27140634 .
- ^ Риппель, Оливье (2017). Черепахи как обнадеживающие монстры: происхождение и эволюция . Издательство Университета Индианы. п. 195. ИСБН 978-0-253-02507-4 . OCLC 962141060 .
- ^ Перейти обратно: а б с Лайсон, Тайлер Р.; Бевер, Габриэль С. (2020). «Происхождение и эволюция строения черепахи» . Ежегодный обзор экологии, эволюции и систематики . 51 (1): 143–166. doi : 10.1146/annurev-ecolsys-110218-024746 . S2CID 225486775 .
- ^ Кадена, Эдвин; Джойс, Уолтер Г. (апрель 2015 г.). «Обзор ископаемых находок черепах клад Platychelyidae и Dortokidae» . Бюллетень Музея естественной истории Пибоди . 56 (1): 3–20. дои : 10.3374/014.056.0101 . ISSN 0079-032X . S2CID 56195415 .
- ^ Эверс, Сережа В.; Бенсон, Роджер Б.Дж. (январь 2019 г.). Смит, Эндрю (ред.). «Новая филогенетическая гипотеза черепах, имеющая значение для времени и количества эволюционных переходов к морскому образу жизни в группе» . Палеонтология . 62 (1): 93–134. Бибкод : 2019Palgy..62...93E . дои : 10.1111/пала.12384 . S2CID 134736808 .
- ^ Перес-Гарсия, Адам (2 сентября 2017 г.). «Новый таксон черепах (Podocnemidoidea, Bothremydidae) раскрывает старейшее известное событие расселения Crown Pleurodira из Гондваны в Лавразию» . Журнал систематической палеонтологии . 15 (9): 709–731. Бибкод : 2017JSPal..15..709P . дои : 10.1080/14772019.2016.1228549 . ISSN 1477-2019 . S2CID 88840423 .
- ^ Феррейра, Габриэль С.; Бандиопадхьяй, Сасвати; Джойс, Уолтер Г. (15 ноября 2018 г.). «Таксономическая переоценка Piramys auffenbergi, забытой черепахи из позднего миоцена острова Пирам, Гуджарат, Индия» . ПерДж . 6 : е5938. дои : 10.7717/peerj.5938 . ISSN 2167-8359 . ПМК 6240434 . ПМИД 30479901 .
- ^ Хираяма, Рен; Исаджи, Синдзи; Хибино, Цуёси (2013). Бринкман, Дональд Б.; Холройд, Патрисия А.; Гарднер, Джеймс Д. (ред.). Дуб Каппачелис род. и др. nov., новая стволовая мягкопанцирная черепаха из раннего мела Японии . Палеобиология и палеоантропология позвоночных. Дордрехт: Springer Нидерланды. стр. 100-1 179–185. дои : 10.1007/978-94-007-4309-0_12 . ISBN 978-94-007-4308-3 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Эверс, Сережа В.; Барретт, Пол М.; Бенсон, Роджер Би Джей (май 2019 г.). «Анатомия Rhinochelys pulchriceps (Protostegidae) и морская адаптация во время ранней эволюции хелониоидов» . ПерДж . 7 : е6811. дои : 10.7717/peerj.6811 . ISSN 2167-8359 . ПМК 6500378 . ПМИД 31106054 .
- ^ Хофмейр, Маргарета Д.; Вамбергер, Мелита; Бранч, Уильям; Шлейхер, Альфред; Дэниэлс, Сэвел Р. (июль 2017 г.). «Радиация черепах (Reptilia, Testudinidae) в Южной Африке от эоцена до современности» . Зоологика Скрипта . 46 (4): 389–400. дои : 10.1111/zsc.12223 . S2CID 88712318 .
- ^ Поропат, Стивен Ф.; Крут, Лесли; Викерс-Рич, Патрисия; Рич, Томас Х. (3 апреля 2017 г.). «Останки древнейшей мейоланиидной черепахи из Австралии: данные из эоценового участка Керосин-Крик в формации Рандл, Квинсленд» . Алчеринга: Австралазийский журнал палеонтологии . 41 (2): 231–239. Бибкод : 2017Алч...41..231П . дои : 10.1080/03115518.2016.1224441 . ISSN 0311-5518 . S2CID 131795055 .
- ^ Перейти обратно: а б Риппель, Оливье; ДеБрага, М. (1996). «Черепахи как диапсидные рептилии» (PDF) . Природа . 384 (6608): 453–455. Бибкод : 1996Natur.384..453R . дои : 10.1038/384453a0 . S2CID 4264378 .
- ^ Перейти обратно: а б Зардоя, Рафаэль; Мейер, Аксель (1998). «Полный митохондриальный геном предполагает сходство черепах с диапсидом» (PDF) . ПНАС . 95 (24): 14226–14231. Бибкод : 1998PNAS...9514226Z . дои : 10.1073/pnas.95.24.14226 . ISSN 0027-8424 . ПМК 24355 . ПМИД 9826682 . Архивировано из оригинала 24 июля 2021 года . Проверено 31 октября 2018 г.
- ^ Маннен, Хидеюки; Ли, Стивен С.-Л. (октябрь 1999 г.). «Молекулярные доказательства существования клада черепах». Молекулярная филогенетика и эволюция . 13 (1): 144–148. дои : 10.1006/mpev.1999.0640 . PMID 10508547 .
- ^ Ивабе, Наоюки; Хара, Юичиро; Кумазава, Ёсинори; Сибамото, Каори; Сайто, Юми; Мията, Такаши; Като, Казутака (декабрь 2004 г.). «Сестринская группа черепах с кладой птиц и крокодилов, выявленная с помощью белков, закодированных ядерной ДНК» . Молекулярная биология и эволюция . 22 (4): 810–813. дои : 10.1093/molbev/msi075 . ПМИД 15625185 .
- ^ Роос, Йонас; Аггарвал, Рамеш К.; Янке, Аксель (ноябрь 2007 г.). «Расширенный митогеномный филогенетический анализ дает новое понимание эволюции крокодилов и их выживания на границе мелового и третичного периода». Молекулярная филогенетика и эволюция . 45 (2): 663–673. дои : 10.1016/j.ympev.2007.06.018 . ПМИД 17719245 .
- ^ Катсу, Ёсинао; Браун, Эдвард Л.; Гийетт, Луис-младший; Игучи, Тайсен (март 2010 г.). «От филогеномики рептилий к геномам рептилий: анализ протоонкогенов c-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. дои : 10.1159/000297715 . ПМИД 20234127 . S2CID 12116018 .
- ^ Лайсон, Тайлер Р.; Сперлинг, Эрик А.; Хеймберг, Алиша М.; Готье, Жак А.; Кинг, Бенджамин Л.; Петерсон, Кевин Дж. (2012). «МикроРНК поддерживают кладу черепаха + ящерица» . Письма по биологии . 8 (1): 104–107. дои : 10.1098/rsbl.2011.0477 . ПМЦ 3259949 . ПМИД 21775315 .
- ^ Кьяри, Иления; Каэ, Винсент; Галтье, Николя; Дельсук, Фредерик (2012). «Филогеномный анализ подтверждает положение черепах как сестринской группы птиц и крокодилов (архозаврии)» . БМК Биология . 10 (65): 65. дои : 10.1186/1741-7007-10-65 . ПМЦ 3473239 . ПМИД 22839781 .
- ^ Перейти обратно: а б Кроуфорд, Николас Г.; Фэрклот, Брант К.; МакКормак, Джон Э.; Брумфилд, Робб Т.; Винкер, Кевин; Глен, Трэвис С. (2012). «Более 1000 ультраконсервативных элементов доказывают, что черепахи являются сестринской группой архозавров» (PDF) . Письма по биологии . 8 (5): 783–786. дои : 10.1098/rsbl.2012.0331 . ПМК 3440978 . ПМИД 22593086 . Архивировано (PDF) из оригинала 10 августа 2012 г. Проверено 21 сентября 2014 г.
- ^ Ван, Чжо; Паскуаль-Аная, Хуан; Задисса, Амонида; и др. (2013). «Проект генома мягкопанцирной черепахи и зеленой морской черепахи дает представление о развитии и эволюции строения тела, специфичного для черепахи» . Природная генетика . 45 (6): 701–706. дои : 10.1038/ng.2615 . ПМК 4000948 . ПМИД 23624526 .
- ^ Джойс, Уолтер Г.; Анкетен, Джереми; Кадена, Эдвин-Альберто; Клод, Жюльен; Данилов Игорь Георгиевич; Эверс, Сережа В.; Феррейра, Габриэль С.; Джентри, Эндрю Д.; Георгалис, Георгиос Л.; Лайсон, Тайлер Р.; Перес-Гарсия, Адам (декабрь 2021 г.). «Номенклатура ископаемых и живых черепах с использованием филогенетически определенных названий клад» . Швейцарский журнал палеонтологии . 140 (1): 5. Бибкод : 2021SwJP..140....5J . дои : 10.1186/s13358-020-00211-x . hdl : 11336/155192 . ISSN 1664-2376 . S2CID 229506832 .
- ^ Бёмер, Кристина; Вернебург, Ингмар (2017). «Глубокий временной взгляд на эволюцию шеи черепахи: в погоне за кодом Хокса по морфологии позвонков» . Научные отчеты . 7 (1): 8939. Бибкод : 2017НатСР...7.8939Б . дои : 10.1038/s41598-017-09133-0 . ПМЦ 5566328 . ПМИД 28827543 .
- ^ Перейти обратно: а б Томсон, Роберт С.; Спинкс, Филип К.; Шаффер, Х. Брэдли (8 февраля 2021 г.). «Глобальная филогения черепах показывает всплеск диверсификации, связанной с климатом, на континентальных окраинах» . Труды Национальной академии наук . 118 (7): e2012215118. Бибкод : 2021PNAS..11812215T . дои : 10.1073/pnas.2012215118 . ISSN 0027-8424 . ПМЦ 7896334 . ПМИД 33558231 .
- ^ Кроуфорд, Николас Г.; Пархэм, Джеймс Ф.; Селлас, Анна Б.; Фэрклот, Брант К.; Гленн, Трэвис С.; Папенфусс, Теодор Дж.; Хендерсон, Джеймс Б.; Хансен, Мэдисон Х.; Симисон, В. Брайан (2015). «Филогеномный анализ черепах». Молекулярная филогенетика и эволюция . 83 : 250–257. дои : 10.1016/j.ympev.2014.10.021 . ISSN 1055-7903 . ПМИД 25450099 .
- ^ Кнаусс, Джорджия Э.; Джойс, Уолтер Г.; Лайсон, Тайлер Р.; Пирсон, Дин (21 сентября 2010 г.). «Новый киностерноид из позднемеловой формации Хелл-Крик в Северной Дакоте и Монтане и происхождение линии Dermatemys mawii ». Палеонтологическая газета . 85 (2). ООО «Спрингер Сайенс энд Бизнес Медиа»: 125–142. дои : 10.1007/s12542-010-0081-x . ISSN 0031-0220 . S2CID 129123961 .
- ^ Джойс, Уолтер Г.; Анкетен, Жереми; Кадена, Эдвин-Альберто; и др. (9 февраля 2021 г.). «Номенклатура ископаемых и живых черепах с использованием филогенетически определенных названий клад» . Швейцарский журнал палеонтологии . 140 (1): 5. Бибкод : 2021SwJP..140....5J . дои : 10.1186/s13358-020-00211-x . hdl : 11336/155192 . ISSN 1664-2384 .
- ^ Вернебург, Ингмар; Уилсон, Лаура AB; Парр, Уильям Ч. Х.; Джойс, Уолтер Г. (1 марта 2015 г.). «Эволюция формы шейных позвонков и ретракции шеи при переходе к современным черепахам: комплексный геометрическо-морфометрический подход» . Систематическая биология . 64 (2): 187–204. дои : 10.1093/sysbio/syu072 . hdl : 1959.4/unsworks_54973 . ISSN 1063-5157 . ПМИД 25305281 .
- ^ Херрел, Энтони; Ван Дамм, Йохан; Аэртс, Питер (2008). «Анатомия и функции шейки матки черепах». В Винекене, Жанетт; Белс, В.Л.; Годфри, Мэтью Х. (ред.). Биология черепах . ЦРК Пресс. стр. 163–186. ISBN 978-0-8493-3339-2 . OCLC 144570900 .
- ^ Оренштейн 2012 , с. 110.
- ^ Феррейра, Габриэль С.; Лаутеншлагер, Стефан; Эверс, Сережа В.; и др. (26 марта 2020 г.). «Биомеханика питания предполагает прогрессивную корреляцию архитектуры черепа и эволюции шеи у черепах» . Научные отчеты . 10 (1). артикул 5505. Бибкод : 2020НатСР..10.5505Ф . дои : 10.1038/s41598-020-62179-5 . ISSN 2045-2322 . ПМК 7099039 . ПМИД 32218478 .
- ^ Уайз, Тейлор Б.; Стейтон, К. Тристан (2017). «Боковая шея и скрытая шея: сравнение морфологии панциря плевродирановых и криптодирановых черепах». Герпетологика . 73 (1): 18–29. doi : 10.1655/HERPETOLOGICA-D-15-00038 . JSTOR 26534349 . S2CID 90226667 .
- ^ Перейти обратно: а б Пау, Ф. Харви (2001). «Рептилии, биоразнообразие». Энциклопедия биоразнообразия . Эльзевир. стр. 146–151. дои : 10.1016/b0-12-226865-2/00233-9 . ISBN 978-0-12-226865-6 .
- ^ Феррейра, Габриэль С.; Бронзати, Марио; Лангер, Макс К.; Стерли, Джулиана (2018). «Филогения, биогеография и закономерности диверсификации бокошейных черепах (Testudines: Pleurodira)» . Королевское общество открытой науки . 5 (3): 171773. Бибкод : 2018RSOS....571773F . дои : 10.1098/rsos.171773 . ISSN 2054-5703 . ПМЦ 5882704 . ПМИД 29657780 .
- ^ Бюльманн, Курт А.; Акре, Томас С.Б.; Айверсон, Джон Б.; и др. (2009). «Глобальный анализ распространения черепах и пресноводных черепах с определением приоритетных охраняемых территорий» (PDF) . Челонианская охрана и биология . 8 (2): 116–149. дои : 10.2744/CCB-0774.1 . S2CID 85942804 . Архивировано (PDF) оригинала 9 мая 2021 г. Проверено 9 мая 2021 г.
- ^ Ульч, Гордон Р. (15 мая 2006 г.). «Экология зимовки черепах: где зимуют черепахи и ее последствия». Биологические обзоры . 81 (3): 339–367. дои : 10.1017/s1464793106007032 . ISSN 1464-7931 . ПМИД 16700968 . S2CID 32939695 .
- ^ Дегенхардт, Уильям Г.; Кристиансен, Джеймс Л. (1975). «Распространение и среда обитания черепах в Нью-Мексико». Юго-западный натуралист . 19 (1): 21–46. дои : 10.2307/3669787 . JSTOR 3669787 .
- ^ Додж, Кара Л.; Галуарди, Бенджамин; Миллер, Тимоти Дж.; Луткаваж, Молли Э. (2014). «Передвижение кожистых черепах, поведение при нырянии и характеристики среды обитания в экорегионах северо-западной части Атлантического океана» . ПЛОС ОДИН . 9 (3): е91726. Бибкод : 2014PLoSO...991726D . дои : 10.1371/journal.pone.0091726 . ПМЦ 3960146 . ПМИД 24646920 .
- ^ Мейер, Рашель (2008). «Gopherus spp.» . Министерство сельского хозяйства США , Лесная служба, Исследовательская станция Роки-Маунтин, Лаборатория пожарных наук. Архивировано из оригинала 16 сентября 2020 года . Проверено 9 мая 2021 г.
- ^ Перейти обратно: а б Уоллес, Брайан П.; Льюисон, Ребекка Л.; Макдональд, Сара Л.; Макдональд, Ричард К.; Кот, Конни Ю.; Келез, Шалейла; Бьоркланд, Рема К.; Финкбайнер, Елена М.; Хельмбрехт, Срай; Краудер, Ларри Б. (5 апреля 2010 г.). «Глобальные закономерности прилова морских черепах» . Письма о сохранении . 3 (3). Уайли: 131–142. Бибкод : 2010ConL....3..131W . дои : 10.1111/j.1755-263x.2010.00105.x . ISSN 1755-263X .
- ^ Перейти обратно: а б Монтгомери, Мэдлин (15 апреля 2021 г.). «Защитники окружающей среды борются против нового закона, который может убить тысячи морских черепах» . ВПЭК . Архивировано из оригинала 17 апреля 2021 года . Проверено 11 мая 2021 г.
- ^ Перейти обратно: а б Роден, Андерс Г.Дж.; Стэнфорд, Крейг Б.; ван Дейк, Питер Пол; Айземберг, Карла Камило; и др. (2018). «Глобальный статус сохранения черепах и черепах (отряд Testudines)» (PDF ) Челонианская охрана и биология . 17 (2): 135–161. дои : 10.2744/CCB-1348.1 . S2CID 91937716 .
- ^ Родин, Андерс Г.Дж.; Уолде, Эндрю Д.; Хорн, Брайан Д.; ван Дейк, Питер Пол; Бланк, Торстен; Хадсон, Рик, ред. (2011). Черепахи в беде: 25+ черепах и пресноводных черепах в мире, находящихся под угрозой исчезновения – 2011 г. Коалиция по сохранению черепах. ОСЛК 711692023 . Архивировано из оригинала 5 марта 2011 года . Проверено 5 марта 2011 г.
- ^ Прайк 2021 , стр. 160–166.
- ^ Прайк 2021 , с. 156.
- ^ Прайк 2021 , с. 157.
- ^ Перейти обратно: а б с ван Дейк, Питер Пол (2002). «Азиатский черепаховый кризис». В Холлидее, Тим; Адлер, Крейг (ред.). Энциклопедия рептилий и амфибий Firefly . Книги Светлячка. стр. 134–135. ISBN 978-1-55297-613-5 .
- ^ МакКаллум, Малькольм (2021). «Утрата биоразнообразия черепах предполагает приближающееся шестое массовое вымирание» . Биоразнообразие и сохранение . 30 (5): 1257–1275. Бибкод : 2021BiCon..30.1257M . дои : 10.1007/s10531-021-02140-8 . ISSN 0960-3115 . S2CID 233903598 . Архивировано из оригинала 24 июля 2021 года . Проверено 12 марта 2021 г.
- ^ Дрейвен, Джеймс (30 мая 2018 г.). «Являются ли черепашьи инкубаторы неэтичными?» . Нэшнл Географик . Архивировано из оригинала 13 июня 2019 года . Проверено 10 мая 2021 г.
- ^ Программы управления пляжами и разведения морских черепах (PDF) . Центр герпетологии / Мадрасский крокодиловый банк, Тамил Наду. 2003. Архивировано (PDF) из оригинала 12 ноября 2020 года . Проверено 7 апреля 2021 г.
- ^ Чакон, Дидиер; Санчес, Хуан; Кальво, Хосе Хоакин; Эш, Дженни (2007). Руководство по управлению и сохранению морских черепах в Коста-Рике; с упором на работу пляжных проектов и питомников [ Руководство по управлению и сохранению морских черепах в Коста-Рике; с упором на эксплуатацию пляжей и детских садов ] (PDF) (на испанском языке). Латиноамериканские морские черепахи и WIDECAST (Большая Карибская сеть морских черепах). Архивировано (PDF) из оригинала 24 июля 2021 г. Проверено 7 апреля 2021 г.
- ^ «Эксперты говорят, что черепаховые фермы угрожают редким видам» . Рыбный фермер. 30 марта 2007. Архивировано из оригинала 18 февраля 2012 года . Проверено 1 ноября 2012 г.
- ^ Чунг, Сзе Ман; Даджен, Дэвид (ноябрь – декабрь 2006 г.). «Количественная оценка азиатского черепахового кризиса: исследования рынка в Южном Китае, 2000–2003 гг.». Охрана водных ресурсов: морские и пресноводные экосистемы . 16 (7): 751–770. Бибкод : 2006ACMFE..16..751C . дои : 10.1002/aqc.803 .
- ^ Амато, Джордж (2007). Разговор в Музее естественной истории (.flv). Точка зрения25. Архивировано из оригинала (видео) 12 ноября 2012 года . Проверено 1 ноября 2012 г.
- ^ Беннетт, Элиза (20 февраля 2020 г.). «Отчет: отказ запретить отлов в ловушки угрожает черепахам в девяти штатах и способствует нелегальной торговле» . Центр биологического разнообразия . Проверено 5 августа 2021 г.
- ^ Перейти обратно: а б Хилтон, Хилари (8 мая 2007 г.). «Не пускать американских черепах в Китай» . Время . Архивировано из оригинала 12 мая 2007 года . Проверено 1 ноября 2012 г.
- ^ Перейти обратно: а б Питтман, Крейг (9 октября 2008 г.). «Китай пожирает флоридских черепах» . «Санкт-Петербург Таймс» . Архивировано из оригинала 20 сентября 2016 года . Проверено 18 августа 2016 г.
- ^ Том, Митчелл (20 ноября 2015 г.). «Программа контроля акул в Квинсленде поймала 84 000 животных» . Акция для дельфинов. Архивировано из оригинала 24 декабря 2020 года . Проверено 25 декабря 2018 г.
- ^ Ламберт, Макс Р.; и др. (2019). «Экспериментальное удаление интродуцированных черепах-ползунков дает новый взгляд на конкуренцию с местной черепахой, находящейся под угрозой исчезновения» . ПерДж . 7 : е7444. дои : 10.7717/peerj.7444 . ПМК 6698372 . ПМИД 31435491 .
- ^ Перейти обратно: а б Прайк 2021 , с. 107.
- ^ Бетц, Эрик (18 сентября 2018 г.). «Первыми землянами, облетевшими Луну, были две советские черепахи» . Обнаружить . Архивировано из оригинала 28 сентября 2019 года . Проверено 14 июля 2019 г.
- ^ Сиддики, Асиф (2018). За пределами Земли: Хроника исследования глубокого космоса, 1958–2016 (PDF) (второе изд.). Офис программы истории НАСА.
- ^ Прайк 2021 , стр. 9–10.
- ^ Перейти обратно: а б с Прайк, 2021 г. , стр. 63–68.
- ^ Перейти обратно: а б с д Маклеллан, Лиз; Никсон, Аманда; Бенн, Джо (июнь 2005 г.). «Сохранение морских черепах в Азиатско-Тихоокеанском регионе» (PDF) . WWF. Архивировано (PDF) из оригинала 24 июля 2021 г. Проверено 22 июля 2021 г.
- ^ Мэллинсон, Джеймс (9 декабря 2011 г.). «Ответ на тело йоги Джеймса Мэллинсона Марка Синглтона» . Академия . Архивировано из оригинала 24 июля 2021 года . Проверено 4 января 2019 г. отредактировано по итогам конференции Американской академии религий, Сан-Франциско, 19 ноября 2011 г.
- ^ Айенгар, Беллур КС (1979) [1966]. Свет на йогу: Йога Дипика . Торсоны. стр. 288–291. ISBN 978-1-85538-166-7 .
- ^ Конверс, Гарриет Максвелл ; Паркер, Артур Касвелл (1908). Мифы и легенды ирокезов штата Нью-Йорк . Университет штата Нью-Йорк. п. 33. Архивировано из оригинала 22 мая 2021 года . Проверено 22 мая 2021 г.
- ^ Филиче, Мишель (6 ноября 2018 г.). «Черепаший остров» . Канадская энциклопедия . Архивировано из оригинала 20 мая 2021 года . Проверено 22 мая 2021 г.
- ^ Прайк 2021 , стр. 44–48.
- ^ Прайк 2021 , с. 56.
- ^ Анонимно; Эвелин-Уайт, Хью Г. (1914). «Гимн 4 Гермесу» . Кембридж, Массачусетс: Издательство Гарвардского университета. Строки 26–65. Архивировано из оригинала 11 мая 2021 года . Проверено 11 мая 2021 г.
- ^ Прайк, 2021 , стр. 49–52.
- ^ «Черепаха и заяц» . Эзопика: Басни Эзопа на английском, латинском и греческом языках . Архивировано из оригинала 16 августа 2019 года . Проверено 11 мая 2021 г.
- ^ Прайк 2021 , с. 139.
- ^ Кэрролл, Льюис (1901) [1865]. «История Якочерепахи». Приключения Алисы в стране чудес . Харпер и братья. п. 128. OCLC 1049742993 .
- ^ Прайк 2021 , с. 135.
- ^ «Суп из черепахи» . Мерриам-Вебстер . Архивировано из оригинала 20 октября 2012 года . Проверено 22 декабря 2020 г.
- ^ Анон (1 апреля 1899 г.). «Бруклинская жизнь [Театр]» . Бруклинская жизнь . п. 31. Архивировано из оригинала 22 мая 2021 года . Проверено 22 мая 2021 г.
это прежде всего очень забавный фарс. Сюжет невелик и касается главным образом пресловутого непостоянства женщины.
- ^ Прайк 2021 , с. 137.
- ^ Прайк 2021 , стр. 118–120.
- ^ Прайк 2021 , стр. 146–148.
- ^ Гринберг, Харви Р. (15 апреля 1990 г.). «Насколько сильны эти черепахи?» . Нью-Йорк Таймс . Архивировано из оригинала 12 июня 2018 года . Проверено 11 мая 2021 г.
- ^ Прайк 2021 , стр. 148–151.
- ^ Перейти обратно: а б Рид, Сиуна А. (2017). «Современные тенденции в животноводстве и ветеринарном уходе за черепахами» (PDF) . Тестудо . 8 (4): 58–68. Архивировано (PDF) из оригинала 31 июля 2019 г. Проверено 31 июля 2019 г.
- ^ Прайк 2021 , с. 181.
- ^ Прайк 2021 , стр. 181–183.
- ^ Прайк 2021 , стр. 120–122.
- ^ Барзик, Джеймс Э. (ноябрь 1999 г.). «Черепахи в кризисе: азиатские продовольственные рынки» . Черепаховый трест. Архивировано из оригинала 22 февраля 2015 года . Проверено 1 ноября 2012 г.
- ^ Кларксон, Джанет (2010). Суп: мировая история . Реакция. п. 115. ИСБН 978-1-86189-774-9 . OCLC 642290114 .
- ^ «Рецепты из другого времени» . Смитсоновский институт . Октябрь 2001 г. Архивировано из оригинала 19 августа 2016 г. Проверено 19 августа 2016 г.
- ^ Чен, Тянь-Си; Чанг, Х.-К.; Лю, Куанг-Янг (2009). «Нерегулируемая торговля черепашьими панцирями для китайской традиционной медицины в Восточной и Юго-Восточной Азии: пример Тайваня». Челонианская охрана и биология . 8 (1): 11–18. дои : 10.2744/CCB-0747.1 . S2CID 86821249 .
- ^ Чжан, Хуан; Ву, Мин-И; Го, Де-Цзянь; и др. (2013). «Гуй-лин-гао (черепашье желе), традиционная китайская функциональная пища, оказывает противовоспалительное действие, ингибируя экспрессию iNOS и провоспалительных цитокинов в спленоцитах, выделенных из мышей BALB/c». Журнал функциональных продуктов питания . 5 (2): 625–632. дои : 10.1016/j.jff.2013.01.004 . hdl : 10397/16357 .
- ^ Кокс, Лиза (12 ноября 2018 г.). «Браконьерство на черепахах-ястребов будет бороться с помощью технологии ДНК» . Хранитель . Проверено 7 августа 2021 г.
Хоксбиллы — единственные морские черепахи, на которых охотятся из-за их панцирей, несмотря на то, что международная торговля продуктами из бисса была запрещена более 20 лет назад.
- ^ «Глобальный статус морской черепахи ястребов: торговля черепаховым панцирем» . Охрана морских черепах. 2007.
- ^ Прайк 2021 , стр. 58–60.
- ^ Страйкер, Гэри (10 апреля 2001 г.). «Запрет на черепаховый панцирь угрожает японской традиции» . Си-Эн-Эн. Архивировано из оригинала 15 декабря 2006 года . Проверено 11 мая 2021 г.
Цитируемые источники
- Франклин, Карл Дж. (2011). Черепаха: необыкновенная естественная история, продолжавшаяся 245 миллионов лет . Крестлайн. ISBN 978-0-7858-2775-7 .
- Оренштейн, Рональд (2012). Черепахи, черепахи и террапины: естественная история . Книги Светлячка. ISBN 978-1-77085-119-1 . OCLC 791162481 .
- Прайк, Луиза (2021). Черепаха . Книги реакции. ISBN 978-1-78914-336-2 . OCLC 1223025640 .
Внешние ссылки
| Базы данных органов управления : Национальные |
|---|