Coronavirus
| Ортокоронавирусы | |
|---|---|
 | |
| Член группы SARS-CoV-2 Ключ иллюстрации: | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Писувирикота |
| Сорт: | Пизонивирицеты |
| Заказ: | Нидовиралес |
| Семья: | Коронавирусы |
| Подсемейство: | Ортокоронавирусы |
| Роды [1] | |
Информация: | |
| Синонимы [2] [3] | |
| |
Коронавирусы представляют собой группу родственных РНК-вирусов , которые вызывают заболевания у млекопитающих и птиц. У людей и птиц они вызывают инфекции дыхательных путей , которые могут варьироваться от легких до летальных. К легким заболеваниям у людей относятся некоторые случаи простуды ( которая также вызывается другими вирусами, преимущественно риновирусами ), тогда как более смертельные разновидности могут вызывать SARS , MERS и COVID-19 . У коров и свиней они вызывают диарею , а у мышей - гепатит и энцефаломиелит .
Коронавирусы составляют подсемейство Orthocoronavirinae , семейство Coronaviridae , порядок Nidovirales и царство Riboviria . [3] [4] Это оболочечные вирусы с одноцепочечной РНК геномом с положительным смыслом и нуклеокапсидом спиральной симметрии. [5] коронавирусов Размер генома колеблется примерно от 26 до 32 килобаз , что является одним из крупнейших среди РНК-вирусов. [6] Они имеют характерные булавовидные шипы , выступающие из их поверхности, которые на электронных микрофотографиях создают изображение, напоминающее звездную корону , от которой и произошло их название. [7]
Этимология
Название «коронавирус» происходит от латинского corona , что означает «корона» или «венок», что само по себе является заимствованием из греческого κορώνη korṓnē , «гирлянда, венок». [8] [9] Название было придумано Джун Алмейда и Дэвидом Тирреллом, которые впервые наблюдали и изучали человеческие коронавирусы. [10] Это слово впервые было использовано в печати в 1968 году неофициальной группой вирусологов в журнале Nature для обозначения нового семейства вирусов. [7] Название относится к характерному внешнему виду вирионов (инфекционной формы вируса) при электронной микроскопии , которые имеют бахрому из крупных выпуклых выступов на поверхности, создающих изображение, напоминающее солнечную корону или гало. [7] [10] Эта морфология создается пепломерами с шипами вируса , которые представляют собой белки на поверхности вируса. [11]
Научное название Coronavirus было принято в качестве названия рода Международным комитетом по номенклатуре вирусов (позже переименованным в Международный комитет по таксономии вирусов ) в 1971 году. [12] По мере увеличения числа новых видов в 2009 году род был разделен на четыре рода, а именно Alphacoronavirus , Betacoronavirus , Deltacoronavirus и Gammacoronavirus . [13] Общее название «коронавирус» используется для обозначения любого члена подсемейства Orthocoronavirinae . [4] По состоянию на 2020 год официально признано 45 видов. [14]
История

Самые ранние сообщения о коронавирусной инфекции у животных произошли в конце 1920-х годов, когда в Северной Америке возникла острая респираторная инфекция домашних кур. [15] Артур Шалк и М.С. Хоун в 1931 году сделали первый подробный отчет, в котором описывалась новая респираторная инфекция кур в Северной Дакоте . Заражение новорожденных птенцов характеризовалось удушьем и вялостью с высокой смертностью 40–90%. [16] Леланд Дэвид Бушнелл и Карл Альфред Брэндли выделили вирус, вызвавший инфекцию, в 1933 году. [17] Вирус тогда был известен как вирус инфекционного бронхита (IBV). Чарльз Д. Хадсон и Фред Роберт Бодетт впервые культивировали вирус в 1937 году. [18] Образец стал известен как штамм Бодетт. В конце 1940-х годов еще два коронавируса животных: JHM, вызывающий заболевание головного мозга (мышиный энцефалит), и вирус мышиного гепатита (MHV), вызывающий гепатит у мышей. были обнаружены [19] В то время еще не осознавалось, что эти три разных вируса связаны между собой. [20] [12]
Коронавирусы человека были открыты в 1960-х годах. [21] [22] используя два разных метода в Великобритании и США. [23] Э. К. Кендалл, Малкольм Байно и Дэвид Тиррел, работавшие в отделении простуды Британского совета медицинских исследований, в 1961 году собрали уникальный вирус простуды , получивший обозначение B814. [24] [25] [26] Вирус невозможно было культивировать с использованием стандартных методов, с помощью которых успешно культивировались риновирусы , аденовирусы и другие известные вирусы простуды. В 1965 году Тиррелл и Байно успешно культивировали новый вирус, серийно пропуская его через культуру органов эмбриональной человека трахеи . [27] Новый метод выращивания был представлен в лаборатории Бертилом Хорном. [28] Изолированный вирус при интраназальном введении добровольцам вызывал простуду и инактивировался эфиром, что указывало на то, что он имел липидную оболочку . [24] [29] Дороти Хамре и Джон Прокноу из Чикагского университета выделили новый вид простуды у студентов-медиков в 1962 году. Они выделили и вырастили вирус в культуре ткани почки , обозначив его как 229E. Новый вирус вызывал у добровольцев простуду и, как и B814, инактивировался эфиром. [30] [31]

Шотландский вирусолог Джун Алмейда из больницы Святого Томаса в Лондоне, сотрудничая с Тирреллом, сравнила структуры IBV, B814 и 229E в 1967 году. [32] [33] С помощью электронной микроскопии было показано, что три вируса морфологически связаны общей формой и характерными булавообразными шипами . [34] Исследовательская группа Национального института здравоохранения в том же году смогла выделить еще одного члена этой новой группы вирусов с помощью культуры органов и назвала один из образцов OC43 (OC для органной культуры). [35] Подобно B814, 229E и IBV, новый вирус простуды OC43 при наблюдении под электронным микроскопом имел характерные булавообразные шипы. [36] [37]
Вскоре было показано, что новые вирусы простуды, подобные IBV, также морфологически родственны вирусу гепатита мышей. [19] Эта новая группа вирусов была названа коронавирусами из-за их отличительного морфологического внешнего вида. [7] Коронавирус человека 229E и коронавирус человека OC43 продолжали изучаться в последующие десятилетия. [38] [39] Штамм коронавируса B814 был утерян. Неизвестно, какой именно у человека это был коронавирус. [40] С тех пор были идентифицированы и другие человеческие коронавирусы, в том числе SARS-CoV в 2003 году, HCoV NL63 в 2003 году, HCoV HKU1 в 2004 году, MERS-CoV в 2013 году и SARS-CoV-2 в 2019 году. [41] С 1960-х годов также было выявлено большое количество коронавирусов животных. [42]
Микробиология
Структура

Коронавирусы представляют собой крупные частицы примерно сферической формы с уникальными выступами на поверхности. [43] Их размер сильно варьируется: средний диаметр от 80 до 120 нм . Известны предельные размеры от 50 до 200 нм в диаметре. [44] Общая молекулярная масса составляет в среднем 40 000 кДа . Они заключены в оболочку, внутри которой находится ряд белковых молекул. [45] Двухслойная липидная оболочка, мембранные белки и нуклеокапсид защищают вирус, когда он находится вне клетки-хозяина. [46]
Оболочка вируса состоит из липидного бислоя , в котором мембраны (М), оболочки (Е) и шипа (S) . структурные белки закреплены [47] Молярное соотношение E:S:M в липидном бислое составляет примерно 1:20:300. [48] Белки E и M представляют собой структурные белки, которые в сочетании с липидным бислоем формируют вирусную оболочку и поддерживают ее размер. [49] S-белки необходимы для взаимодействия с клетками-хозяевами. Но особенность человеческого коронавируса NL63 состоит в том, что его белок М имеет сайт связывания с клеткой-хозяином, а не белок S. [50] Диаметр оболочки составляет 85 нм. Оболочка вируса на электронных микрофотографиях выглядит как отдельная пара электронно-плотных оболочек (оболочек, которые относительно непрозрачны для электронного луча, используемого для сканирования вирусной частицы). [51] [49]
Белок М является основным структурным белком оболочки, обеспечивающим общую форму, и является мембранным белком III типа . Он состоит из 218–263 аминокислотных остатков и образует слой толщиной 7,8 нм. [45] Он имеет три домена: короткий N-концевой эктодомен , тройной трансмембранный домен и С-концевой эндодомен . С-концевой домен образует матричную решетку, которая увеличивает толщину оболочки. Различные виды могут иметь либо N- , либо O -связанные гликаны в аминоконцевом домене своего белка. Белок М имеет решающее значение на этапах сборки, почкования , формирования оболочки и патогенеза жизненного цикла вируса. [52]
Белки E являются второстепенными структурными белками и сильно варьируют у разных видов. [44] В частице коронавируса содержится всего около 20 копий молекулы белка Е. [48] Их размер составляет от 8,4 до 12 кДа, и они состоят из 76–109 аминокислот. [44] Они представляют собой интегральные белки (т.е. встроены в липидный слой) и имеют два домена, а именно трансмембранный домен и внемембранный С-концевой домен. Они почти полностью α-спиральны, с единственным α-спиральным трансмембранным доменом и образуют пентамерные (пятимолекулярные) ионные каналы в липидном бислое. Они ответственны за сборку вирионов, внутриклеточный транспорт и морфогенез (почкование). [45]

Шипы являются наиболее отличительной чертой коронавирусов и отвечают за поверхность, подобную короне или ореолу. В среднем частица коронавируса имеет 74 поверхностных шипа. [53] Каждый шип имеет длину около 20 нм и состоит из тримера S. белка Белок S, в свою очередь, состоит из субъединиц S1 и S2 . Гомотримерный белок S представляет собой слитый белок класса I , который опосредует связывание рецептора и слияние мембран между вирусом и клеткой-хозяином. Субъединица S1 образует головку шипа и имеет рецептор-связывающий домен (RBD). Субъединица S2 образует стебель, который закрепляет шип в оболочке вируса, а при активации протеазы обеспечивает слияние. Две субъединицы остаются нековалентно связанными, пока они находятся на поверхности вируса, пока не прикрепятся к мембране клетки-хозяина. [45] В функционально активном состоянии три S1 прикреплены к двум субъединицам S2. Комплекс субъединиц расщепляется на отдельные субъединицы, когда вирус связывается и сливается с клеткой-хозяином под действием протеаз, таких как семейство катепсинов и трансмембранная протеаза серин 2 (TMPRSS2) клетки-хозяина. [54]

Белки S1 являются наиболее важными компонентами с точки зрения инфекции. Они также являются наиболее вариабельными компонентами, поскольку отвечают за специфичность клетки-хозяина. Они обладают двумя основными доменами, называемыми N-концевым доменом (S1-NTD) и C-концевым доменом (S1-CTD), оба из которых служат рецептор-связывающими доменами. NTD распознают и связывают сахара на поверхности клетки-хозяина. Исключением является NTD MHV , который связывается с белковой молекулой 1 адгезии клеток, связанной с карциноэмбриональным антигеном (CEACAM1). S1-CTD отвечают за распознавание различных белковых рецепторов, таких как ангиотензинпревращающий фермент 2 (ACE2), аминопептидаза N (APN) и дипептидилпептидаза 4 (DPP4). [45]
Подмножество коронавирусов (в частности, представители подгруппы бетакоронавирусов А ) также имеет более короткий шипообразный поверхностный белок, называемый гемагглютининэстеразой (HE). [42] Белки HE встречаются в виде гомодимеров, состоящих примерно из 400 аминокислотных остатков, и имеют размер от 40 до 50 кДа. Они выглядят как крошечные поверхностные выступы длиной от 5 до 7 нм, встроенные между шипами. Они помогают прикрепляться к клетке-хозяину и отсоединяться от нее. [55]
Внутри оболочки находится нуклеокапсид , который образован из нескольких копий белка нуклеокапсида (N), которые связаны с геномом одноцепочечной РНК с положительным смыслом в конформации типа непрерывных бусин на нитке. [49] [56] Белок N представляет собой фосфопротеин размером от 43 до 50 кДа и разделен на три консервативных домена. Большая часть белка состоит из доменов 1 и 2, которые обычно богаты аргининами и лизинами . Домен 3 имеет короткий карбокси-конец и имеет суммарный отрицательный заряд из-за избытка кислотных остатков над основными. [44]
Геном

Коронавирусы содержат геном одноцепочечной РНК с положительным смыслом . коронавирусов Размер генома колеблется от 26,4 до 31,7 тысяч оснований . [6] Размер генома — один из самых больших среди РНК-вирусов. Геном имеет 5'-метилированную кепку и 3'-полиаденилированный хвост . [49]
Организация генома коронавируса следующая: 5'-лидер-UTR -репликаза (ORF1ab)-шип (S)-оболочка (E)-мембрана (M)-нуклеокапсид (N) -3'UTR -поли (A) хвост. Открытые рамки считывания 1a и 1b, занимающие первые две трети генома, кодируют полипротеин репликазы (pp1ab). Полипротеин репликазы самостоятельно расщепляется с образованием 16 неструктурных белков (nsp1–nsp16). [49]
Более поздние рамки считывания кодируют четыре основных структурных белка: шип , оболочку , мембрану и нуклеокапсид . [57] Между этими рамками считывания расположены рамки считывания дополнительных белков. Количество дополнительных белков и их функции уникальны и зависят от конкретного коронавируса. [49]
Цикл репликации
Вход в ячейку
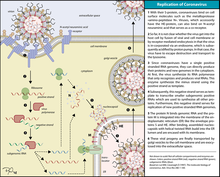
Инфекция начинается, когда вирусный спайковый белок прикрепляется к комплементарному рецептору клетки-хозяина. После прикрепления протеаза клетки-хозяина расщепляет и активирует прикрепленный к рецептору белок-шип. В зависимости от доступной протеазы клетки-хозяина расщепление и активация позволяют вирусу проникнуть в клетку-хозяина путем эндоцитоза или прямого слияния вирусной оболочки с мембраной хозяина . [58]
Коронавирусы могут проникать в клетки либо путем слияния их липидной оболочки с клеточной мембраной на поверхности клетки, либо путем интернализации посредством эндоцитоза. [59]
Перевод генома
При попадании в клетку-хозяина вирусная частица не имеет оболочки , и ее геном попадает в цитоплазму клетки . Геном РНК коронавируса имеет 5'-метилированную кепку и 3'-полиаденилированный хвост, что позволяет ему действовать как информационная РНК и напрямую транслироваться рибосомами клетки-хозяина . Рибосомы хозяина транслируют начальные перекрывающиеся открытые рамки считывания ORF1a и ORF1b вирусного генома в два больших перекрывающихся полипротеина, pp1a и pp1ab. [49]
Более крупный полипротеин pp1ab является результатом сдвига рамки рибосомы -1, вызванного скользкой последовательностью (UUUAAAC) и расположенным ниже псевдоузлом РНК в конце открытой рамки считывания ORF1a. [60] Сдвиг рамки рибосомы обеспечивает непрерывную трансляцию ORF1a, за которой следует ORF1b. [49]
Полипротеины имеют свои собственные протеазы PLpro ( nsp3 ) и 3CLpro (nsp5), которые расщепляют полипротеины в разных специфических сайтах. Расщепление полипротеина pp1ab дает 16 неструктурных белков (от nsp1 до nsp16). Белки-продукты включают различные белки репликации, такие как РНК-зависимая РНК-полимераза ( nsp12 ), РНК-геликаза (nsp13) и экзорибонуклеаза (nsp14). [49]
Репликаза-транскриптаза
Ряд неструктурных белков сливаются, образуя мультибелковый комплекс репликаза-транскриптаза (RTC). Основным белком репликазы-транскриптазы является РНК-зависимая РНК-полимераза (RdRp). Он непосредственно участвует в репликации и транскрипции РНК с цепи РНК. Другие неструктурные белки комплекса участвуют в процессе репликации и транскрипции. Неструктурный белок экзорибонуклеаза , , например, обеспечивает дополнительную точность репликации, обеспечивая функцию корректуры которой нет у РНК-зависимой РНК-полимеразы. [61]
Репликация . Одной из основных функций комплекса является репликация вирусного генома. RdRp непосредственно опосредует синтез геномной РНК с отрицательным смыслом из геномной РНК с положительным смыслом. За этим следует репликация геномной РНК с положительным смыслом из геномной РНК с отрицательным смыслом. [49]
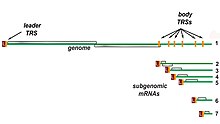
Транскрипция . Другой важной функцией комплекса является транскрипция вирусного генома. RdRp непосредственно опосредует синтез молекул субгеномной РНК с отрицательным смыслом из геномной РНК с положительным смыслом. За этим процессом следует транскрипция этих субгеномных молекул РНК с отрицательным смыслом в соответствующие им мРНК с положительным смыслом . [49] Субгеномные мРНК образуют « вложенный набор », который имеет общую 5'-головку и частично дублирующий 3'-конец. [62]
Рекомбинация . Комплекс репликаза-транскриптаза также способен к генетической рекомбинации , когда в одной и той же инфицированной клетке присутствуют по крайней мере два вирусных генома. [62] Рекомбинация РНК, по-видимому, является основной движущей силой в определении генетической изменчивости внутри вида коронавируса, способности вида коронавируса переходить от одного хозяина к другому и, в редких случаях, в определении появления новых коронавирусов. [63] Точный механизм рекомбинации у коронавирусов неясен, но, вероятно, включает переключение матрицы во время репликации генома. [63]
Сборка и выпуск
Реплицированная геномная РНК с положительным смыслом становится геномом вирусов-потомков . мРНК представляют собой генные транскрипты последней трети генома вируса после начальной перекрывающейся рамки считывания. Эти мРНК транслируются рибосомами хозяина в структурные белки и многие вспомогательные белки. [49] Трансляция РНК происходит внутри эндоплазматической сети . Вирусные структурные белки S, E и M перемещаются по секреторному пути в промежуточный отсек Гольджи . Там белки М управляют большинством межбелковых взаимодействий, необходимых для сборки вируса после его связывания с нуклеокапсидом . Затем вирусы-потомки высвобождаются из клетки-хозяина путем экзоцитоза через секреторные пузырьки. После высвобождения вирусы могут заразить другие клетки-хозяева. [64]
Передача инфекции
Инфицированные носители способны распространять вирусы в окружающую среду. Взаимодействие белка-шипа коронавируса с его комплементарным клеточным рецептором играет центральную роль в определении тропизма ткани , инфекционности и видового диапазона высвободившегося вируса. [65] [66] Коронавирусы в основном поражают эпителиальные клетки . [42] Они передаются от одного хозяина к другому, в зависимости от вида коронавируса, аэрозольным , фомитным или фекально-оральным путем . [67]
Коронавирусы человека поражают эпителиальные клетки дыхательных путей , тогда как коронавирусы животных обычно поражают эпителиальные клетки пищеварительного тракта . [42] Коронавирус SARS , например, заражает эпителиальные клетки легких человека аэрозольным путем. [68] путем связывания с рецептором ангиотензинпревращающего фермента 2 (ACE2). [69] Коронавирус трансмиссивного гастроэнтерита (TGEV) инфицирует эпителиальные клетки пищеварительного тракта свиньи фекально-оральным путем. [67] путем связывания с рецептором аланинаминопептидазы (APN). [49]
Классификация
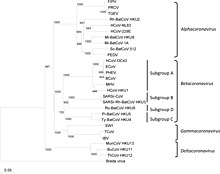
Коронавирусы образуют подсемейство Orthocoronavirinae. [2] [3] [4] которое является одним из двух подсемейств семейства Coronaviridae , порядка Nidovirales и царства Riboviria . [42] [70] Они делятся на четыре рода: Alphacoronavirus , Betacoronavirus , Gammacoronavirus и Deltacoronavirus . Альфакоронавирусы и бетакоронавирусы поражают млекопитающих, а гаммакоронавирусы и дельтакоронавирусы преимущественно поражают птиц. [71] [72]
- Род: Альфакоронавирус ; [67]
- Виды: Альфакоронавирус 1 ( TGEV , Коронавирус кошек , Коронавирус собак ), Коронавирус человека 229E , Коронавирус человека NL63 , Коронавирус летучей мыши Miniopterus 1 , Коронавирус летучей мыши Miniopterus HKU8 , Вирус эпидемической диареи свиней , Коронавирус летучей мыши Rhinolophus HKU2 , Коронавирус летучей мыши Scotophilus 512
- Род Бетакоронавирус ; [68]
- Виды: Бетакоронавирус 1 ( коронавирус крупного рогатого скота , коронавирус человека OC43 ), коронавирус ежей 1 , человеческий коронавирус HKU1 , коронавирус, связанный с ближневосточным респираторным синдромом , мышиный коронавирус , коронавирус летучей мыши Pipistrellus HKU5 , коронавирус летучей мыши Rousettus HKU9 , коронавирус, связанный с тяжелым острым респираторным синдромом ( SARS-CoV-1 , SARS-CoV-2 ), коронавирус летучей мыши Tylonycteris HKU4
- Род Гаммакоронавирус ; [18]
- Род Дельтакоронавирус.
Источник

По оценкам, самый последний общий предок (MRCA) всех коронавирусов существовал совсем недавно, около 8000 г. до н.э. , хотя некоторые модели относят общего предка к 55 миллионам лет или более, что подразумевает долгосрочную коэволюцию с видами летучих мышей и птиц. [73] Самый последний общий предок линии альфакоронавируса датируется примерно 2400 г. до н.э., линии бетакоронавируса - 3300 г. до н.э., линии гаммакоронавируса - 2800 г. до н.э. и линии дельтакоронавируса - примерно 3000 г. до н.э. Летучие мыши и птицы, как теплокровные летающие позвоночные, являются идеальным естественным резервуаром генофонда коронавируса ( летучие мыши являются резервуаром альфа- и бета-коронавирусов, а птицы — резервуаром гамма- и дельтакоронавирусов). Большое количество и глобальный ареал видов летучих мышей и птиц, являющихся носителями вирусов, способствовали обширной эволюции и распространению коронавирусов. [74]
Многие человеческие коронавирусы происходят от летучих мышей. [75] Человеческий коронавирус NL63 имел общего предка с коронавирусом летучих мышей (ARCoV.2) между 1190 и 1449 годами нашей эры. [76] Человеческий коронавирус 229E имел общего предка с коронавирусом летучих мышей (GhanaGrp1 Bt CoV) между 1686 и 1800 годами нашей эры. [77] Совсем недавно, где-то до 1960 года, коронавирус альпаки и человеческий коронавирус 229E разошлись. [78] MERS-CoV возник у человека от летучих мышей через промежуточного хозяина верблюдов. [79] MERS-CoV, хотя и связан с несколькими видами коронавируса летучих мышей, похоже, отошел от них несколько столетий назад. [80] Наиболее близкородственные коронавирус летучих мышей и SARS-CoV разошлись в 1986 году. [81] Предки SARS-CoV первыми заразили летучих мышей-листоносов рода Hipposideridae ; впоследствии они распространились на подковоносов вида Rhinolophidae , затем на азиатских пальмовых циветт и, наконец, на человека. [82] [83]
В отличие от других бетакоронавирусов, считается, что бычий коронавирус вида Betacoronavirus 1 и подрода Embecovirus возник у грызунов , а не у летучих мышей. [75] [84] В 1790-х годах лошадиный коронавирус отделился от бычьего коронавируса после межвидового скачка . [85] Позже, в 1890-х годах, человеческий коронавирус OC43 отделился от бычьего коронавируса после еще одного случая межвидового распространения. [86] [85] Предполагается, что пандемия гриппа 1890 года могла быть вызвана этим побочным событием, а не вирусом гриппа , из-за соответствующего времени, неврологических симптомов и неизвестного возбудителя пандемии. [87] Предполагается, что человеческий коронавирус OC43 не только вызывает респираторные инфекции, но и играет роль в развитии неврологических заболеваний . [88] В 1950-х годах человеческий коронавирус OC43 начал расходиться до своих нынешних генотипов . [89] Филогенетически вирус мышиного гепатита ( мышиный коронавирус мышей ), поражающий печень и центральную нервную систему , [90] относится к человеческому коронавирусу OC43 и бычьему коронавирусу. Человеческий коронавирус HKU1, как и вышеупомянутые вирусы, также происходит от грызунов. [75]
Инфекция у человека

Коронавирусы существенно различаются по фактору риска. , могут убить более 30% инфицированных, Некоторые из них, например MERS-CoV а некоторые относительно безвредны, например, простуда. [49] Коронавирусы могут вызывать простуду с такими серьезными симптомами, как лихорадка и боль в горле из-за опухших аденоидов . [91] Коронавирусы могут вызывать пневмонию (прямую вирусную пневмонию или вторичную бактериальную пневмонию ) и бронхит (прямой вирусный бронхит или вторичный бактериальный бронхит). [92] Человеческий коронавирус, открытый в 2003 году, SARS-CoV , вызывающий тяжелый острый респираторный синдром (ТОРС), имеет уникальный патогенез, поскольку вызывает инфекции как верхних, так и нижних дыхательных путей . [92]
Известно шесть видов коронавирусов человека, причем один вид подразделяется на два разных штамма, всего семь штаммов коронавирусов человека.

Четыре человеческих коронавируса вызывают симптомы, которые, как правило, легкие, хотя утверждается, что в прошлом они могли быть более агрессивными: [93]
- Коронавирус человека OC43 (HCoV-OC43), β-CoV
- Коронавирус человека HKU1 (HCoV-HKU1), β-CoV
- Коронавирус человека 229E (HCoV-229E), α-CoV
- Коронавирус человека NL63 (HCoV-NL63), α-CoV–
Три человеческих коронавируса вызывают потенциально серьезные симптомы:
- Коронавирус тяжелого острого респираторного синдрома (SARS-CoV), β-CoV (выявлен в 2003 г.)
- Коронавирус, связанный с ближневосточным респираторным синдромом (MERS-CoV), β-CoV (выявлен в 2012 г.)
- Коронавирус 2 тяжелого острого респираторного синдрома (SARS-CoV-2), β-CoV (выявлен в 2019 г.)
Они вызывают заболевания, обычно называемые SARS , MERS и COVID-19 соответственно.
Простуда
Хотя простуду обычно вызывают риновирусы , [94] примерно в 15% случаев причиной является коронавирус. [95] Человеческие коронавирусы HCoV-OC43, HCoV-HKU1, HCoV-229E и HCoV-NL63 постоянно циркулируют в человеческой популяции у взрослых и детей во всем мире и вызывают, как правило, легкие симптомы простуды. [88] Четыре легких коронавируса имеют сезонную заболеваемость, возникающую в зимние месяцы в умеренном климате . [96] [97] нет преобладания ни в какое время года В тропическом климате . [98]
Тяжелый острый респираторный синдром (ТОРС)
| БВРС-КоВ | SARS-CoV | SARS-CoV-2 | |
|---|---|---|---|
| Болезнь | ПРОГУЛКА | ОРВИ | COVID-19 |
| Вспышки | Вспышка MERS в 2012 г. | 2002–2004 | COVID-19 пандемия |
| Эпидемиология | |||
| Дата первого выявленный случай | Июнь 2012 | ноябрь 2002 | декабрь 2019 [99] |
| Место первого выявленный случай | Джидда , Саудовская Аравия | Затем , Китай | Ухань , Китай |
| Средний возраст | 56 | 44 [100] [а] | 56 [101] |
| Соотношение полов (М:Ж) | 3.3:1 | 0.8:1 [102] | 1.6:1 [101] |
| Подтвержденные случаи [заболевания | 2494 | 8096 [103] | 676,609,955 [104] [б] |
| Летальные исходы | 858 | 774 [103] | 6,881,955 [104] [б] |
| Уровень смертности | 37% | 9.2% | 1.02% [104] |
| Симптомы | |||
| Высокая температура | 98% | 99–100% | 87.9% [105] |
| Сухой кашель | 47% | 29–75% | 67.7% [105] |
| Одышка | 72% | 40–42% | 18.6% [105] |
| Диарея | 26% | 20–25% | 3.7% [105] |
| Больное горло | 21% | 13–25% | 13.9% [105] |
| Вентиляционное использование | 24.5% [106] | 14–20% | 4.1% [107] |
Примечания
| |||
В 2003 году, после вспышки тяжелого острого респираторного синдрома (ТОРС), которая началась годом ранее в Азии, и вторичных случаев заболевания в других частях мира, Всемирная организация здравоохранения (ВОЗ) выпустила пресс-релиз, в котором говорилось, что новый коронавирус, выявленный несколькими лаборатории был возбудителем атипичной пневмонии. Вирус получил официальное название коронавирус SARS (SARS-CoV). Заразились более 8000 человек из 29 стран и территорий, по меньшей мере 774 умерли. [108] [69]
Ближневосточный респираторный синдром (MERS)
В сентябре 2012 года был идентифицирован новый тип коронавируса, первоначально называвшийся «Новый коронавирус 2012», а теперь официально названный «коронавирус ближневосточного респираторного синдрома» (MERS-CoV). [109] [110] Вскоре после этого Всемирная организация здравоохранения выпустила глобальное предупреждение. [111] В обновленной информации ВОЗ от 28 сентября 2012 года говорится, что вирус, по-видимому, нелегко передается от человека к человеку. [112] Однако 12 мая 2013 года случай передачи вируса от человека человеку во Франции был подтвержден Министерством социальных дел и здравоохранения Франции. [113] Кроме того, о случаях передачи вируса от человека к человеку сообщило Министерство здравоохранения Туниса . Два подтвержденных случая касались людей, которые, судя по всему, заразились от своего покойного отца, который заболел после посещения Катара и Саудовской Аравии. Несмотря на это, похоже, что вирус с трудом передается от человека к человеку, поскольку большинство инфицированных людей не передают вирус. [114] К 30 октября 2013 года в Саудовской Аравии было зарегистрировано 124 случая заболевания и 52 случая смерти. [115]
После того, как голландский Медицинский центр Эразмус секвенировал вирус, вирусу было присвоено новое название — Медицинский центр Коронавируса человека – Эразмус (HCoV-EMC). Окончательное название вируса — коронавирус ближневосточного респираторного синдрома (MERS-CoV). Единственные случаи заболевания в США (оба выжили) были зафиксированы в мае 2014 года. [116]
В мае 2015 года вспышка MERS-CoV произошла в Республике Корея , когда мужчина, посетивший Ближний Восток, посетил четыре больницы в районе Сеула для лечения своего заболевания. Это вызвало одну из крупнейших вспышек MERS-CoV за пределами Ближнего Востока. [117] По состоянию на декабрь 2019 года лабораторными исследованиями подтверждено 2468 случаев заражения БВРС-КоВ, 851 из которых закончились смертельным исходом, уровень смертности составил примерно 34,5%. [118]
Коронавирусное заболевание 2019 (COVID-19)
В декабре 2019 года вспышка пневмонии была зарегистрирована в Ухане . китайском [119] 31 декабря 2019 года вспышка была связана с новым штаммом коронавируса. [120] Всемирная организация здравоохранения присвоила ему временное название 2019-nCoV. [121] [122] [123] позже переименован в SARS-CoV-2 Международным комитетом по таксономии вирусов .
По состоянию на 10 марта 2023 года их было не менее 6 881 955. [104] подтвержденных смертей и более 676 609 955 [104] подтвержденных случаев пандемии COVID-19 . Уханьский штамм был идентифицирован как новый штамм бетакоронавируса из группы 2B, имеющий примерно 70% генетическое сходство с SARS-CoV. [124] Вирус на 96% похож на коронавирус летучих мышей, поэтому многие подозревают, что он также происходит от летучих мышей. [125] [126]
Коронавирус HuPn-2018
В ходе эпиднадзорного исследования архивных образцов малазийских пациентов с вирусной пневмонией вирусологи выявили штамм собачьего коронавируса , которым заразились люди в 2018 году.
Инфекция у животных
Коронавирусы признаны причиной патологических состояний в ветеринарной медицине с 1930-х годов. [19] Они заражают целый ряд животных, включая свиней, крупный рогатый скот, лошадей, верблюдов, кошек, собак, грызунов, птиц и летучих мышей. [127] Большинство коронавирусов, связанных с животными, поражают кишечный тракт и передаются фекально-оральным путем. [128] Значительные исследовательские усилия были сосредоточены на выяснении вирусного патогенеза этих коронавирусов животных, особенно вирусологами, интересующимися ветеринарными и зоонозными заболеваниями. [129]
Сельскохозяйственные животные
Коронавирусы заражают домашних птиц. [130] Вирус инфекционного бронхита (ИБВ), тип коронавируса, вызывает инфекционный бронхит птиц . [131] Вирус вызывает обеспокоенность в птицеводстве из-за высокой смертности от инфекции, быстрого распространения и влияния на производство. [127] Вирус поражает как производство мяса, так и производство яиц и наносит значительный экономический ущерб. [132] У кур вирус инфекционного бронхита поражает не только дыхательные пути, но и урогенитальный тракт . Вирус может распространиться на различные органы курицы. [131] Вирус передается аэрозольным путем и пищевыми продуктами, загрязненными фекалиями. Существуют различные вакцины против ИБК, которые помогли ограничить распространение вируса и его вариантов. [127] Вирус инфекционного бронхита — один из ряда штаммов вида птичьего коронавируса . [133] Другим штаммом птичьего коронавируса является коронавирус индеек (TCV), который вызывает энтерит у индеек . [127]
Коронавирусы также влияют на другие отрасли животноводства, такие как свиноводство и животноводство. [127] Коронавирус синдрома острой диареи свиней (SADS-CoV), родственный коронавирусу летучих мышей HKU2 , вызывает диарею у свиней. [134] Вирус эпидемической диареи свиней (ВЭДС) – это недавно появившийся коронавирус, который также вызывает диарею у свиней. [135] Вирус трансмиссивного гастроэнтерита (TGEV), который относится к виду Alphacoronavirus 1 , [136] — еще один коронавирус, вызывающий диарею у молодых свиней. [137] [138] В животноводстве бычий коронавирус (BCV), который относится к виду Betacoronavirus 1 и связан с HCoV-OC43, [139] является причиной тяжелого профузного энтерита у молодых телят. [127]
Домашние животные
Коронавирусы заражают домашних животных, таких как кошки, собаки и хорьки. [130] Существует две формы кошачьего коронавируса , обе из которых относятся к виду Alphacoronavirus 1 . [136] Кишечный коронавирус кошек является возбудителем незначительного клинического значения, но спонтанная мутация этого вируса может привести к инфекционному перитониту кошек (FIP) — заболеванию с высокой смертностью. [127] Собак заражают два разных коронавируса. Собачий коронавирус (CCoV), принадлежащий к виду Alphacoronavirus 1 , [136] вызывает легкие желудочно-кишечные заболевания. [127] Респираторный коронавирус собак (CRCoV), принадлежащий к виду Betacoronavirus 1 и родственный HCoV-OC43, [139] вызывают респираторные заболевания. [127] Точно так же хорьков заражают два типа коронавируса. [140] Кишечный коронавирус хорьков вызывает желудочно-кишечный синдром, известный как эпизоотический катаральный энтерит (ECE), и более летальную системную версию вируса (например, FIP у кошек), известную как системный коронавирус хорьков (FSC). [141] [142]
Лабораторные животные
Коронавирусы заражают лабораторных животных. [127] Вирус гепатита мышей (MHV), который относится к виду мышиного коронавируса . [143] вызывает эпидемическое заболевание мышей с высокой смертностью, особенно среди колоний лабораторных мышей. [144] До открытия SARS-CoV MHV был наиболее изученным коронавирусом как in vivo , так и in vitro, а также на молекулярном уровне. Некоторые штаммы MHV вызывают у мышей прогрессирующий демиелинизирующий энцефалит , который использовался в качестве мышиной модели рассеянного склероза . [129] Вирус сиалодакриоаденита (SDAV), который представляет собой штамм вида мышиного коронавируса . [143] — высококонтагиозный коронавирус лабораторных крыс, который может передаваться между особями при прямом контакте и опосредованно аэрозольным путем. Кроличий кишечный коронавирус вызывает острые желудочно-кишечные заболевания и диарею у молодых европейских кроликов . [127] Уровень смертности высок. [145]
Профилактика и лечение
ряд вакцин . Против человеческого коронавируса SARS-CoV-2 с использованием различных методов был разработан [146] [147] противовирусные мишени Также были идентифицированы против коронавирусов человека, такие как вирусные протеазы, полимеразы и входные белки. В стадии разработки находятся лекарства , нацеленные на эти белки и на различные этапы репликации вируса. [148] [147]
Доступны вакцины против коронавирусов животных IBV, TGEV и Canine CoV, хотя их эффективность ограничена. В случае вспышек высококонтагиозных коронавирусов животных, таких как PEDV, такие меры, как уничтожение целых стад свиней. для предотвращения передачи инфекции другим стадам могут быть использованы [49]
См. также
Ссылки
- ^ «Таксономия вирусов: выпуск 2018b» . Международный комитет по таксономии вирусов (ICTV) . Март 2019 г. Архивировано из оригинала 04 марта 2018 г. Проверено 24 января 2020 г.
- ^ Перейти обратно: а б «2017.012-015S» (xlsx) . Международный комитет по таксономии вирусов (ICTV) . Октябрь 2018 г. Архивировано из оригинала 14 мая 2019 г. Проверено 24 января 2020 г.
- ^ Перейти обратно: а б с Фань Ю, Чжао К., Ши З.Л., Чжоу П. (март 2019 г.). «Коронавирусы летучих мышей в Китае» . Вирусы . 11 (3): 210. дои : 10.3390/v11030210 . ПМК 6466186 . ПМИД 30832341 .
- ^ Черри Дж., Деммлер-Харрисон Г.Дж., Каплан С.Л., Стейнбах В.Дж., Хотез П.Дж. (2017). Учебник Фейгина и Черри по детским инфекционным болезням . Elsevier Науки о здоровье. п. ПТ6615. ISBN 978-0-323-39281-5 .
- ^ Перейти обратно: а б Ву ПК, Хуан Ю, Лау С.К., Юэнь Кюй (август 2010 г.). «Геномика и биоинформатический анализ коронавируса» . Вирусы . 2 (8): 1804–20. дои : 10.3390/v2081803 . ISSN 1999-4915 . ПМК 3185738 . ПМИД 21994708 .
Коронавирусы обладают самыми большими геномами (от 26,4 т.п.н. (ThCoV HKU12) до 31.7 т.п.н. (SW1)] среди всех известных РНК-вирусов (рис. 1) [2,13,16].
- ^ Перейти обратно: а б с д Алмейда Дж.Д., Берри Д.М., Каннингем CH, Хамре Д., Хофстад М.С., Маллуччи Л. и др. (ноябрь 1968 г.). «Вирусология: Коронавирусы» . Природа . 220 (5168): 650. Бибкод : 1968Natur.220..650. . дои : 10.1038/220650b0 . ПМК 7086490 .
[T] Здесь также имеется характерная «бахрома» выступов длиной 200 А, имеющих округлую или лепестковую форму ... Этот внешний вид, напоминающий солнечную корону, присущ вирусу гепатита мыши и нескольким вирусам, недавно выделенным от человека, а именно штамму В814, 229Е и ряд других.
- ^ «Определение коронавируса по Мерриам-Вебстеру» . Мерриам-Вебстер. Архивировано из оригинала 23 марта 2020 г. Проверено 24 марта 2020 г.
- ^ «Определение короны Мерриам-Вебстером» . Мерриам-Вебстер. Архивировано из оригинала 24 марта 2020 г. Проверено 24 марта 2020 г.
- ^ Перейти обратно: а б Тиррелл Д.А., Филдер М. (2002). Холодные войны: борьба с простудой . Издательство Оксфордского университета. п. 96. ИСБН 978-0-19-263285-2 .
Мы более внимательно рассмотрели внешний вид новых вирусов и заметили, что их окружает своего рода ореол. Обращение к словарю привело к появлению латинского эквивалента «корона», и так родилось название «коронавирус».
- ^ Штурман Л.С., Холмс КВ (1 января 1983 г.). Лауффер М.А., Мараморош К. (ред.). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 28 : 35–112. дои : 10.1016/s0065-3527(08)60721-6 . ISBN 978-0-12-039828-7 . ПМЦ 7131312 . ПМИД 6362367 .
[Эти] вирусы имели характерную бахрому из больших характерных пепломеров или шипов в форме лепестков, которые напоминали корону, как corona spinarum в религиозном искусстве; отсюда и название коронавирусы.
- ^ Перейти обратно: а б Лалчхандама К. (2020). «Хроники коронавирусов: бронхит, гепатит и простуда» . Научное видение . 20 (1): 43–53. дои : 10.33493/scivis.20.01.04 .
- ^ Карстенс Э.Б. (2010). «Ратификационное голосование по таксономическим предложениям Международного комитета по таксономии вирусов (2009 г.)» . Архив вирусологии . 155 (1): 133–46. дои : 10.1007/s00705-009-0547-x . ПМК 7086975 . ПМИД 19960211 .
- ^ «Международный комитет по таксономии вирусов (ICTV)» . talk.ictvonline.org . Проверено 14 сентября 2020 г.
- ^ Эстола Т (1970). «Коронавирусы, новая группа РНК-вирусов животных». Птичьи болезни . 14 (2): 330–336. дои : 10.2307/1588476 . ISSN 0005-2086 . JSTOR 1588476 . ПМИД 4316767 .
- ^ Фабрикант Дж (1998). «Ранняя история инфекционного бронхита». Птичьи болезни . 42 (4): 648–650. дои : 10.2307/1592697 . ISSN 0005-2086 . JSTOR 1592697 . ПМИД 9876830 .
- ^ Бушнелл Л.Д., Брэндли, Калифорния (1933). «Ларинготрахеит у цыплят» . Птицеводство . 12 (1): 55–60. дои : 10.3382/ps.0120055 .
- ^ Перейти обратно: а б Декаро Н. (2011). «Гаммакоронавирус» В Тидоне С., Дарай Дж. (ред.). Гаммакоронавирус‡: Coronaviridae . Индекс вирусов Спрингера Спрингер. стр. 100-1 403–413. дои : 10.1007/978-0-387-95919-1_58 . ISBN 978-0-387-95919-1 . ПМК 7176155 .
- ^ Перейти обратно: а б с Макинтош К. (1974). «Коронавирусы: сравнительный обзор». В: Арбер В., Хаас Р., Хенле В., Хофшнайдер П.Х., Йерне Н.К., Колдовски П., Копровски Х., Маалё О., Ротт Р. (ред.). Актуальные проблемы микробиологии и иммунологии / Результаты микробиологических и иммунологических исследований . Берлин, Гейдельберг: Springer. п. 87. дои : 10.1007/978-3-642-65775-7_3 . ISBN 978-3-642-65775-7 .
- ^ «Жили-были коронавирусы» . Биомедицинские реалии (на французском языке). 27 марта 2020 г. Проверено 18 апреля 2020 г.
- ^ Кан Дж. С., Макинтош К. (ноябрь 2005 г.). «История и последние достижения в открытии коронавируса» . Журнал детских инфекционных заболеваний . 24 (11 Приложение): S223–7, обсуждение S226. дои : 10.1097/01.inf.0000188166.17324.60 . ПМИД 16378050 .
- ^ Махасе Э (апрель 2020 г.). «BMJ в 1965 году» . БМЖ . 369 : м1547. дои : 10.1136/bmj.m1547 . ПМИД 32299810 .
- ^ Монто А.С. (1984). "Коронавирусы". В Эвансе А.С. (ред.). Вирусные инфекции человека . Спрингер США. стр. 151–165. дои : 10.1007/978-1-4684-4727-9_7 . ISBN 978-1-4684-4727-9 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Перейти обратно: а б Кендалл Э.Дж., Байно М.Л., Тиррелл Д.А. (июль 1962 г.). «Выделения вируса от простудных заболеваний, происходящих в школе-интернате» . Британский медицинский журнал . 2 (5297): 82–6. дои : 10.1136/bmj.2.5297.82 . ЧВК 1925312 . ПМИД 14455113 .
- ^ Ричмонд С. (18 июня 2005 г.). «Дэвид Тиррелл» . BMJ: Британский медицинский журнал . 330 (7505): 1451. doi : 10.1136/bmj.330.7505.1451 . ПМК 558394 .
- ^ «Уведомления о некрологе: Малкольм Байон» . Британский медицинский журнал . 2 (5660): 827–829. 28 июня 1969 г. дои : 10.1136/bmj.2.5660.827 . S2CID 220187042 .
- ^ Тиррелл Д.А., Байно М.Л. (июнь 1965 г.). «Культивирование нового типа вируса простуды в органных культурах» . Британский медицинский журнал . 1 (5448): 1467–70. дои : 10.1136/bmj.1.5448.1467 . ПМК 2166670 . ПМИД 14288084 .
- ^ Тиррелл Д.А., Филдер М. (2002). Холодные войны: борьба с простудой . Издательство Оксфордского университета. стр. 93–95. ISBN 978-0-19-263285-2 .
- ^ Хаган В.А., Брунер Д.В., Гиллеспи Дж.Х., Тимони Дж.Ф., Скотт Ф.В., Барлоу Дж.Э. (1988). Микробиология и инфекционные болезни домашних животных Хагана и Брунера: со ссылкой на этиологию, эпизоотологию, патогенез, иммунитет, диагностику и чувствительность к противомикробным препаратам . Издательство Корнельского университета. п. 440. ИСБН 978-0-8014-1896-9 .
- ^ Хамре Д., Процноу Дж. Дж. (январь 1966 г.). «Новый вирус, выделенный из дыхательных путей человека». Труды Общества экспериментальной биологии и медицины . 121 (1): 190–3. дои : 10.3181/00379727-121-30734 . ПМИД 4285768 . S2CID 1314901 .
- ^ Кнапп А. «Тайная история первого коронавируса» . Форбс . Проверено 6 мая 2020 г.
- ^ «Женщина, которая обнаружила первый коронавирус» . Новости Би-би-си . 14 апреля 2020 г.
- ^ Алмейда Дж. (26 июня 2008 г.). «Джун Алмейда (урожденная Харт)» . БМЖ . 336 (7659): 1511.1–1511. дои : 10.1136/bmj.a434 . ISSN 0959-8138 . ПМЦ 2440895 .
- ^ Алмейда JD, Тиррелл Д.А. (апрель 1967 г.). «Морфология трех ранее не охарактеризованных респираторных вирусов человека, растущих в органной культуре» . Журнал общей вирусологии . 1 (2): 175–8. дои : 10.1099/0022-1317-1-2-175 . ПМИД 4293939 .
- ^ Макинтош К., Беккер В.Б., Чанок Р.М. (декабрь 1967 г.). «Рост в мозгу мышей-сосунов вирусов, подобных IBV, у пациентов с заболеваниями верхних дыхательных путей» . Труды Национальной академии наук Соединенных Штатов Америки . 58 (6): 2268–73. Бибкод : 1967PNAS...58.2268M . дои : 10.1073/pnas.58.6.2268 . ПМК 223830 . ПМИД 4298953 .
- ^ Макинтош К., Дис Дж. Х., Беккер В.Б., Капикян А.З., Чанок Р.М. (апрель 1967 г.). «Выявление в культурах органов трахеи новых вирусов от пациентов с респираторными заболеваниями» . Труды Национальной академии наук Соединенных Штатов Америки . 57 (4): 933–40. Бибкод : 1967ПНАС...57..933М . дои : 10.1073/pnas.57.4.933 . ПМК 224637 . ПМИД 5231356 .
- ^ Таймс ХМ-младший (5 мая 1967 г.). «Шесть недавно обнаруженных вирусов могут объяснить простуду; штаммы похожи на микробы, вызывающие бронхиальную инфекцию у кур, которые считаются новой группой» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 25 апреля 2020 г.
- ^ Мьинт С.Х. (1995). «Коронавирусные инфекции человека». В Сидделле С.Г. (ред.). Коронавирусы . Вирусы. Спрингер США. стр. 389–401. дои : 10.1007/978-1-4899-1531-3_18 . ISBN 978-1-4899-1531-3 . S2CID 80726096 .
- ^ Геллер С., Варбанов М., Дюваль Р.Э. (ноябрь 2012 г.). «Коронавирусы человека: понимание устойчивости окружающей среды и ее влияние на разработку новых антисептических стратегий» . Вирусы . 4 (11): 3044–68. дои : 10.3390/v4113044 . ПМЦ 3509683 . ПМИД 23202515 .
- ^ Монто А.С., Коулинг Б.Дж., Пейрис Дж.С. (2014), Каслоу Р.А., Стэнберри Л.Р., Ле Дюк Дж.В. (ред.), «Коронавирусы», Вирусные инфекции человека , Бостон, Массачусетс: Springer США: 199–223, doi : 10.1007/ 978-1-4899-7448-8_10 , ISBN 978-1-4899-7447-1 , PMC 7122465 ,
Другие штаммы OC и B814, которые не удалось адаптировать к мозгу мыши, также сопротивлялись адаптации к клеточной культуре; эти отдельные вирусы с тех пор были утеряны и, возможно, недавно были заново открыты.
- ^ Чжу Н., Чжан Д., Ван В., Ли Х, Ян Б., Сун Дж. и др. (февраль 2020 г.). «Новый коронавирус от пациентов с пневмонией в Китае, 2019 г.» . Медицинский журнал Новой Англии . 382 (8): 727–733. дои : 10.1056/NEJMoa2001017 . ПМК 7092803 . ПМИД 31978945 .
- ^ Перейти обратно: а б с д и де Гроот Р.Дж., Бейкер С.С. , Барик Р., Энхуанес Л., Горбаленья А.Е., Холмс К.В. и др. (2011). «Семейство Coronaviridae » . В King AM, Lefkowitz E, Adams MJ, Carstens EB, Международный комитет по таксономии вирусов, Международный союз микробиологических обществ. Отдел вирусологии (ред.). Девятый доклад Международного комитета по таксономии вирусов . Оксфорд: Эльзевир. стр. 806–28. дои : 10.1016/B978-0-12-384684-6.00068-9 . ISBN 978-0-12-384684-6 . S2CID 212719285 .
- ^ Голдсмит К.С., Татти К.М., Ксиазек Т.Г., Роллин П.Е., Комер Дж.А., Ли У.В. и др. (февраль 2004 г.). «Ультраструктурная характеристика коронавируса SARS» . Новые инфекционные заболевания . 10 (2): 320–6. дои : 10.3201/eid1002.030913 . ПМК 3322934 . ПМИД 15030705 .
Вирионы приобрели оболочку путем отпочкования в цистернах и образовали в основном сферические, иногда плеоморфные частицы, средний диаметр которых составлял 78 нм (рис. 1А).
- ^ Перейти обратно: а б с д Мастера ПС (2006). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 66 : 193–292. дои : 10.1016/S0065-3527(06)66005-3 . ISBN 978-0-12-039869-0 . ПМК 7112330 . ПМИД 16877062 .
- ^ Перейти обратно: а б с д и Лалчхандама К. (2020). «Хроники коронавирусов: электронный микроскоп, пончик и шип» . Научное видение . 20 (2): 78–92. дои : 10.33493/scivis.20.02.03 .
- ^ Нойман Б.В., Кисс Дж., Кундинг А.Х., Бхелла Д., Бакш М.Ф., Коннелли С. и др. (апрель 2011 г.). «Структурный анализ белка М в сборке и морфологии коронавируса» . Журнал структурной биологии . 174 (1): 11–22. дои : 10.1016/j.jsb.2010.11.021 . ПМК 4486061 . ПМИД 21130884 .
См. рисунок 10.
- ^ Лай М.М., Кавана Д. (1997). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 48 : 1–100. дои : 10.1016/S0065-3527(08)60286-9 . ISBN 978-0-12-039848-5 . ПМК 7130985 . ПМИД 9233431 .
- ^ Перейти обратно: а б Годе М., Л'Харидон Р., Вотеро Ж.Ф., Лауд Х. (1992). «Коронавирус TGEV ORF4 кодирует мембранный белок, который встроен в вирионы» . Вирусология . 188 (2): 666–75. дои : 10.1016/0042-6822(92)90521-п . ПМК 7131960 . ПМИД 1316677 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Структура вириона.
- ^ Наскальска А, Домбровска А, Щепански А, Милевска А, Ясик К.П., Пирц К. (октябрь 2019 г.). «Мембранный белок коронавируса человека NL63 отвечает за взаимодействие с рецептором адгезии» . Журнал вирусологии . 93 (19). дои : 10.1128/JVI.00355-19 . ПМК 6744225 . ПМИД 31315999 .
- ^ Нойман Б.В., Адэр Б.Д., Йошиока С., Киспе Дж.Д., Орка Г., Кун П. и др. (август 2006 г.). «Супрамолекулярная архитектура тяжелого острого респираторного синдрома коронавируса, выявленная методом электронной криомикроскопии» . Журнал вирусологии . 80 (16): 7918–28. дои : 10.1128/JVI.00645-06 . ПМЦ 1563832 . ПМИД 16873249 .
Диаметр частиц находился в диапазоне от 50 до 150 нм, исключая шипы, при среднем диаметре частиц от 82 до 94 нм; См. также рисунок 1 для двойной оболочки.
- ^ Шуман Д., Филдинг, Британская Колумбия (май 2019 г.). «Белок оболочки коронавируса: современные знания» . Вирусологический журнал . 16 (1): 69. дои : 10.1186/s12985-019-1182-0 . ПМЦ 6537279 . ПМИД 31133031 .
- ^ Нойман Б.В., Кисс Дж., Кундинг А.Х., Бхелла Д., Бакш М.Ф., Коннелли С. и др. (апрель 2011 г.). «Структурный анализ белка М в сборке и морфологии коронавируса» . Журнал структурной биологии . 174 (1): 11–22. дои : 10.1016/j.jsb.2010.11.021 . ПМК 4486061 . ПМИД 21130884 .
- ^ Дж. Алсаади Э.А., Джонс М.М. (апрель 2019 г.). «Мембраносвязывающие белки коронавирусов» . Будущая вирусология . 14 (4): 275–286. дои : 10.2217/fvl-2018-0144 . ПМК 7079996 . ПМИД 32201500 .
- ^ Цзэн К., Лангерайс М.А., ван Влит А.Л., Хейзинга Э.Г., де Гроот Р.Дж. (июль 2008 г.). «Структура гемагглютинин-эстеразы коронавируса дает представление об эволюции короны и вируса гриппа» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (26): 9065–9. Бибкод : 2008PNAS..105.9065Z . дои : 10.1073/pnas.0800502105 . ПМЦ 2449365 . ПМИД 18550812 .
- ^ Чанг К.К., Хоу М.Х., Чанг К.Ф., Сяо К.Д., Хуан Т.Х. (март 2014 г.). «Белок нуклеокапсида коронавируса SARS — формы и функции» . Противовирусные исследования . 103 : 39–50. дои : 10.1016/j.antiviral.2013.12.009 . ПМЦ 7113676 . ПМИД 24418573 .
См. рисунок 4в.
- ^ Снейдер Э.Дж., Бреденбек П.Дж., Доббе Дж.К., Тиль В., Зибур Дж., Пун Л.Л. и др. (август 2003 г.). «Уникальные и консервативные особенности генома и протеома SARS-коронавируса, раннего отделения от линии коронавируса 2-й группы» . Журнал молекулярной биологии . 331 (5): 991–1004. дои : 10.1016/S0022-2836(03)00865-9 . ПМК 7159028 . ПМИД 12927536 .
См. рисунок 1.
- ^ Симмонс Г., Змора П., Гирер С., Хойрих А., Полманн С. (декабрь 2013 г.). «Протеолитическая активация шипового белка SARS-коронавируса: сокращение ферментов на переднем крае противовирусных исследований» . Противовирусные исследования . 100 (3): 605–14. doi : 10.1016/j.antiviral.2013.09.028 . ПМЦ 3889862 . ПМИД 24121034 .
См. рисунок 2.
- ^ Шлахчич В.Дж., Дабровска А., Милевска А., Зойла Н., Блащик К., Баррето-Дюран Е. и др. (июль 2022 г.). «SARS-CoV-2 заражает in vitro модель развивающейся поджелудочной железы человека посредством эндоцитоза» . iScience . 25 (7): 104594. Бибкод : 2022iSci...25j4594S . дои : 10.1016/j.isci.2022.104594 . ПМЦ 9212970 . ПМИД 35756892 .
- ^ Мастерс ПС (01.01.2006). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 66 . Академическая пресса: 193–292. дои : 10.1016/S0065-3527(06)66005-3 . ISBN 978-0-12-039869-0 . ПМК 7112330 . ПМИД 16877062 .
См. рисунок 8.
- ^ Секстон Н.Р., Смит Э.К., Блан Х., Виньуцци М., Пирсен О.Б., Денисон М.Р. (август 2016 г.). «Идентификация на основе гомологии мутации РНК-зависимой РНК-полимеразы коронавируса, которая придает устойчивость к множественным мутагенам» . Журнал вирусологии . 90 (16): 7415–28. дои : 10.1128/JVI.00080-16 . ПМЦ 4984655 . ПМИД 27279608 .
Наконец, эти результаты в сочетании с результатами предыдущих работ (33, 44) позволяют предположить, что CoV кодируют по крайней мере три белка, участвующих в верности (nsp12-RdRp, nsp14-ExoN и nsp10), поддерживая сборку мультипротеиновой репликазы верности. комплекс, как описано ранее (38).
- ^ Перейти обратно: а б Пейн С (01 января 2017 г.). «Глава 17. Семейство Coronaviridae». Вирусы . Академическая пресса. стр. 149–158. дои : 10.1016/B978-0-12-803109-4.00017-9 . ISBN 978-0-12-803109-4 . S2CID 91572610 .
- ^ Перейти обратно: а б Су С., Вонг Г., Ши В., Лю Дж., Лай А.С., Чжоу Дж. и др. (июнь 2016 г.). «Эпидемиология, генетическая рекомбинация и патогенез коронавирусов» . Тенденции в микробиологии . 24 (6): 490–502. дои : 10.1016/j.tim.2016.03.003 . ПМК 7125511 . ПМИД 27012512 .
- ^ Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Жизненный цикл коронавируса — сборка и выпуск.
- ^ Мастерс ПС (01.01.2006). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 66 . Академическая пресса: 193–292. дои : 10.1016/S0065-3527(06)66005-3 . ISBN 978-0-12-039869-0 . ПМК 7112330 . ПМИД 16877062 .
Тем не менее, взаимодействие между S- белком и рецептором остается основным, если не единственным, определяющим диапазон видов-хозяев коронавируса и тропизм тканей.
- ^ Цуй Дж., Ли Ф., Ши З.Л. (март 2019 г.). «Происхождение и эволюция патогенных коронавирусов» . Обзоры природы. Микробиология . 17 (3): 181–92. дои : 10.1038/s41579-018-0118-9 . ПМК 7097006 . ПМИД 30531947 .
Различные штаммы SARS-CoV, выделенные от нескольких хозяев, различаются по сродству связывания с человеческим ACE2 и, следовательно, по своей инфекционности клеток человека 76, 78 (рис. 6b).
- ^ Перейти обратно: а б с Декаро Н. (2011). «Альфакоронавирус ‡: Coronaviridae». В Тидоне С., Дарай Дж. (ред.). Индекс вирусов Спрингера . Спрингер. стр. 371–383. дои : 10.1007/978-0-387-95919-1_56 . ISBN 978-0-387-95919-1 . ПМК 7176201 .
- ^ Перейти обратно: а б Декаро Н. (2011). «Бетакоронавирус ‡: Coronaviridae». В Тидоне С., Дарай Дж. (ред.). Индекс вирусов Спрингера . Спрингер. стр. 385–401. дои : 10.1007/978-0-387-95919-1_57 . ISBN 978-0-387-95919-1 . ПМК 7176184 .
- ^ Перейти обратно: а б Ли Ф, Ли В, Фарзан М, Харрисон С.С. (сентябрь 2005 г.). «Структура домена, связывающего спайковый рецептор коронавируса SARS, в комплексе с рецептором» . Наука . 309 (5742): 1864–68. Бибкод : 2005Sci...309.1864L . дои : 10.1126/science.1116480 . ПМИД 16166518 . S2CID 12438123 .
- ^ Международный комитет по таксономии вирусов (24 августа 2010 г.). «Основной список видов ICTV 2009 — версия 10» . Архивировано из оригинала (xls) 15 апреля 2013 г.
- ^ Вертхайм Д.О., Чу Д.К., Пейрис Дж.С., Косаковский пруд С.Л., Пун Л.Л. (июнь 2013 г.). «Дело о древнем происхождении коронавирусов» . Журнал вирусологии . 87 (12): 7039–45. дои : 10.1128/JVI.03273-12 . ПМЦ 3676139 . ПМИД 23596293 .
Альфакоронавирусы и бетакоронавирусы встречаются исключительно у млекопитающих, тогда как гаммакоронавирусы и дельтакоронавирусы поражают преимущественно птиц.
- ^ «Nextstrain, филогенетическое древо бета-коронавируса» . nextstrain.org .
- ^ Вертхайм Д.О., Чу Д.К., Пейрис Дж.С., Косаковский пруд С.Л., Пун Л.Л. (июнь 2013 г.). «Дело о древнем происхождении коронавирусов» . Журнал вирусологии . 87 (12): 7039–45. дои : 10.1128/JVI.03273-12 . ПМЦ 3676139 . ПМИД 23596293 .
- ^ Ву ПК, Лау С.К., Лам К.С., Лау CC, Цанг АК, Лау Дж.Х. и др. (апрель 2012 г.). «Обнаружение семи новых коронавирусов млекопитающих и птиц в роде дельтакоронавируса подтверждает, что коронавирусы летучих мышей являются источником генов альфа-коронавируса и бета-коронавируса, а птичьи коронавирусы — источником генов гамма- и дельтакоронавируса» . Журнал вирусологии . 86 (7): 3995–4008. дои : 10.1128/JVI.06540-11 . ПМК 3302495 . ПМИД 22278237 .
- ^ Перейти обратно: а б с Форни Д., Кальяни Р., Клеричи М., Сирони М. (январь 2017 г.). «Молекулярная эволюция геномов коронавируса человека» . Тенденции в микробиологии . 25 (1): 35–48. дои : 10.1016/j.tim.2016.09.001 . ПМК 7111218 . ПМИД 27743750 .
В частности, считается, что все HCoV имеют происхождение от летучих мышей, за исключением бета-CoV линии A, резервуары которых могут иметься у грызунов [2].
- ^ Хьюн Дж., Ли С., Йонт Б., Смит А., Стерджес Л., Олсен Дж.К. и др. (декабрь 2012 г.). «Доказательства, подтверждающие зоонозное происхождение штамма NL63 коронавируса человека» . Журнал вирусологии . 86 (23): 12816–25. дои : 10.1128/JVI.00906-12 . ПМК 3497669 . ПМИД 22993147 .
Если эти прогнозы верны, это наблюдение предполагает, что HCoV-NL63 мог произойти от летучих мышей между 1190 и 1449 годами нашей эры.
- ^ Пфефферле С., Оппонг С., Дрекслер Дж. Ф., Глоза-Рауш Ф., Ипсен А., Зеебенс А. и др. (сентябрь 2009 г.). «Дальние родственники коронавируса тяжелого острого респираторного синдрома и близкие родственники человеческого коронавируса 229E у летучих мышей, Гана» . Новые инфекционные заболевания . 15 (9): 1377–84. дои : 10.3201/eid1509.090224 . ПМК 2819850 . ПМИД 19788804 .
Самый последний общий предок hCoV-229E и GhanaBt-CoVGrp1 существовал примерно в 1686–1800 годах нашей эры.
- ^ Кроссли Б.М., Мок Р.Э., Каллисон С.А., Хиетала С.К. (декабрь 2012 г.). «Идентификация и характеристика нового респираторного коронавируса альпаки, наиболее близкого к человеческому коронавирусу 229E» . Вирусы . 4 (12): 3689–700. дои : 10.3390/v4123689 . ПМЦ 3528286 . ПМИД 23235471 .
- ^ Форни Д., Кальяни Р., Клеричи М., Сирони М. (январь 2017 г.). «Молекулярная эволюция геномов коронавируса человека» . Тенденции в микробиологии . 25 (1): 35–48. дои : 10.1016/j.tim.2016.09.001 . ПМК 7111218 . ПМИД 27743750 .
- ^ Лау С.К., Ли К.С., Цанг А.К., Лам К.С., Ахмед С., Чен Х. и др. (август 2013 г.). «Генетическая характеристика вирусов линии C бетакоронавируса у летучих мышей выявила заметное расхождение последовательностей в белке-шипе коронавируса летучей мыши HKU5 у японского нетопыря: значение для происхождения нового коронавируса ближневосточного респираторного синдрома» . Журнал вирусологии . 87 (15): 8638–50. дои : 10.1128/JVI.01055-13 . ПМЦ 3719811 . ПМИД 23720729 .
- ^ Виджайкришна Д., Смит Г.Дж., Чжан Дж.К., Пейрис Дж.С., Чен Х., Гуань Ю. (апрель 2007 г.). «Эволюционный взгляд на экологию коронавирусов» . Журнал вирусологии . 81 (8): 4012–20. дои : 10.1128/jvi.02605-06 . ПМК 1866124 . ПМИД 17267506 .
- ^ Гуй М.А., Пюхмайль С.Дж., Гонсалес Дж.П., Тилинг Э., Киттаяпонг П., Манугерра Х.К. (октябрь 2011 г.). «Следы предка SARS-коронавируса в колониях летучих мышей Юго-Восточной Азии и теория убежища» . Инфекция, генетика и эволюция . 11 (7): 1690–702. Бибкод : 2011InfGE..11.1690G . дои : 10.1016/j.meegid.2011.06.021 . ПМК 7106191 . ПМИД 21763784 .
- ^ Цуй Дж., Хан Н., Штрейкер Д., Ли Г., Тан Х., Ши З. и др. (октябрь 2007 г.). «Эволюционные взаимоотношения между коронавирусами летучих мышей и их хозяевами» . Новые инфекционные заболевания . 13 (10): 1526–32. дои : 10.3201/eid1310.070448 . ПМК 2851503 . ПМИД 18258002 .
- ^ Лау С.К., Ву ПК, Ли К.С., Цанг А.К., Фан Р.Ю., Лук Х.К. и др. (март 2015 г.). «Обнаружение нового коронавируса, коронавируса China Rattus HKU24, у норвежских крыс подтверждает мышиное происхождение бетакоронавируса 1 и имеет значение для предка линии бетакоронавируса А» . Журнал вирусологии . 89 (6): 3076–92. дои : 10.1128/JVI.02420-14 . ПМЦ 4337523 . ПМИД 25552712 .
- ^ Перейти обратно: а б Бидохти М.Р., Тровен М., Кришна Н.К., Мунир М., Белак С., Алениус С. и др. (сентябрь 2013 г.). «Эволюционная динамика бычьих коронавирусов: закономерность естественного отбора гена спайка предполагает адаптивную эволюцию штаммов» . Журнал общей вирусологии . 94 (Часть 9): 2036–2049 гг. дои : 10.1099/vir.0.054940-0 . ПМИД 23804565 .
См. Таблицу 1.
- ^ Вийген Л., Кейертс Э., Моэс Э., Толен И., Воллантс Э., Леми П. и др. (февраль 2005 г.). «Полная геномная последовательность человеческого коронавируса OC43: анализ молекулярных часов предполагает относительно недавнее событие передачи зоонозного коронавируса» . Журнал вирусологии . 79 (3): 1595–604. doi : 10.1128/jvi.79.3.1595-1604.2005 . ПМК 544107 . ПМИД 15650185 .
- ^ Вийген Л., Кейертс Э., Моэс Э., Толен И., Воллантс Э., Леми П. и др. (февраль 2005 г.). «Полная геномная последовательность человеческого коронавируса OC43: анализ молекулярных часов предполагает относительно недавнее событие передачи зоонозного коронавируса» . Журнал вирусологии . 79 (3): 1595–604. doi : 10.1128/JVI.79.3.1595-1604.2005 . ПМК 544107 . ПМИД 15650185 .
Однако возникает соблазн высказать альтернативную гипотезу, согласно которой пандемия 1889–1890 годов могла быть результатом межвидовой передачи бычьих коронавирусов человеку, что привело к последующему появлению HCoV-OC43.
- ^ Перейти обратно: а б Корман В.М., Мут Д., Нимейер Д., Дростен С. (2018). «Хозяева и источники эндемичных коронавирусов человека» . Достижения в области исследования вирусов . 100 : 163–188. дои : 10.1016/bs.aivir.2018.01.001 . ISBN 978-0-12-815201-0 . ПМК 7112090 . ПМИД 29551135 .
- ^ Лау С.К., Ли П., Цанг АК, Ип CC, Це Х, Ли Р.А. и др. (ноябрь 2011 г.). «Молекулярная эпидемиология человеческого коронавируса OC43 показывает эволюцию различных генотипов с течением времени и недавнее появление нового генотипа в результате естественной рекомбинации» . Журнал вирусологии . 85 (21): 11325–37. дои : 10.1128/JVI.05512-11 . ПМК 3194943 . ПМИД 21849456 .
- ^ Шаумбург CS, Хелд KS, Лейн TE (май 2008 г.). «Инфекция ЦНС вирусом мышиного гепатита: модель защиты, болезней и восстановления» . Границы бионауки . 13 (13): 4393–406. дои : 10.2741/3012 . ПМК 5025298 . ПМИД 18508518 .
- ^ Лю П., Ши Л., Чжан В., Хэ Дж., Лю С., Чжао С. и др. (ноябрь 2017 г.). «Анализ распространенности и генетического разнообразия коронавирусов человека среди трансграничных детей» . Вирусологический журнал . 14 (1): 230. дои : 10.1186/s12985-017-0896-0 . ПМК 5700739 . ПМИД 29166910 .
- ^ Перейти обратно: а б Форги С., Марри Ти Джей (февраль 2009 г.). «Атипичная пневмония, связанная с оказанием медицинской помощи». Семинары по респираторной медицине и медицине интенсивной терапии . 30 (1): 67–85. дои : 10.1055/s-0028-1119811 . ПМИД 19199189 . S2CID 260316838 .
- ^ Король А (02 мая 2020 г.). «Необычная простуда» . Новый учёный . 246 (3280): 32–35. Бибкод : 2020НовыйSc.246...32K . дои : 10.1016/S0262-4079(20)30862-9 . ISSN 0262-4079 . ПМК 7252012 . ПМИД 32501321 .
- ^ Сесил Р.Л., Голдман Л., Шафер А.И. (2012). Goldman's Cecil Medicine, Expert Consult Premium Edition (24-е изд.). Elsevier Науки о здоровье. стр. 2103–. ISBN 978-1-4377-1604-7 . Архивировано из оригинала 4 мая 2016 г.
- ^ Пельчар (2010). Микробиология: прикладной подход . McGraw-Hill Education (India) Pvt Limited. п. 656. ИСБН 978-0-07-015147-5 . Архивировано из оригинала 16 мая 2016 г.
- ^ Чарльтон К.Л., Бабади Э., Гиноккио К.С., Хэтчетт Т.Ф., Джеррис Р.К., Ли Ю и др. (январь 2019 г.). «Практическое руководство для лабораторий клинической микробиологии: Вирусы, вызывающие острые инфекции дыхательных путей» . Обзоры клинической микробиологии . 32 (1). дои : 10.1128/CMR.00042-18 . ПМК 6302358 . ПМИД 30541871 .
См. рисунок 1.
- ^ Монто А.С., ДеДжонг П., Каллеар А.П., Баззи Л.А., Каприола С., Малош Р.Э. и др. (апрель 2020 г.). «Возникновение и передача коронавируса в течение 8 лет в когорте домохозяйств HIVE в Мичигане» . Журнал инфекционных болезней . 222 : 9–16. дои : 10.1093/infdis/jiaa161 . ПМК 7184402 . ПМИД 32246136 .
- ^ Абдул-Расул С., Филдинг, Британская Колумбия (май 2010 г.). «Понимание человеческого коронавируса HCoV-NL63» . Открытый вирусологический журнал . 4 : 76–84. doi : 10.2174/1874357901004010076 (неактивен 16 марта 2024 г.). ПМЦ 2918871 . ПМИД 20700397 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на март 2024 г. ( ссылка ) - ^ Ван С., Хорби П.В., Хайден Ф.Г., Гао Г.Ф. (февраль 2020 г.). «Новая вспышка коронавируса, вызывающая глобальную озабоченность в области здравоохранения» . Ланцет . 395 (10223): 470–473. дои : 10.1016/S0140-6736(20)30185-9 . ПМК 7135038 . ПМИД 31986257 .
- ^ Лау Э.Х., Сюн К.А., Коулинг Б.Дж., Чен Ч., Хо Л.М., Цанг Т. и др. (март 2010 г.). «Сравнительный эпидемиологический анализ атипичной пневмонии в Гонконге, Пекине и Тайване» . БМК Инфекционные болезни . 10:50 . дои : 10.1186/1471-2334-10-50 . ПМЦ 2846944 . ПМИД 20205928 .
- ^ Перейти обратно: а б «Старость, сепсис, связанные с плохими результатами лечения COVID-19, смерть» . CIDRAP , Университет Миннесоты . 10.03.2020 . Проверено 29 марта 2020 г.
- ^ Карлберг Дж., Чонг Д.С., Лай, Вайоминг (февраль 2004 г.). «У мужчин более высокий уровень смертности от тяжелого острого респираторного синдрома, чем у женщин?» . Американский журнал эпидемиологии . 159 (3): 229–31. дои : 10.1093/aje/kwh056 . ПМК 7110237 . ПМИД 14742282 .
- ^ Перейти обратно: а б «Сводка вероятных случаев атипичной пневмонии с началом заболевания с 1 ноября 2002 г. по 31 июля 2003 г.» . Всемирная организация здравоохранения. Апрель 2004 года.
- ^ Перейти обратно: а б с д и «Информационная панель COVID-19 Центра системных наук и инженерии (CSSE) Университета Джона Хопкинса (JHU)» . АркГИС . Университет Джонса Хопкинса . Проверено 10 марта 2023 г.
- ^ Перейти обратно: а б с д и «Отчет совместной миссии ВОЗ и Китая по борьбе с коронавирусной болезнью 2019 г. (COVID-19)» (PDF) . Всемирная организация здравоохранения. февраль 2020.
- ^ О МД, Пак ВБ, Пак СВ, Чхве П.Г., Банг Дж.Х., Сонг К.Х. и др. (март 2018 г.). «Ближневосточный респираторный синдром: что мы узнали из вспышки 2015 года в Республике Корея» . Корейский журнал внутренней медицины . 33 (2): 233–246. дои : 10.3904/kjim.2018.031 . ПМК 5840604 . ПМИД 29506344 .
- ^ Сamendys-Silva SA (март 2020 г.). «Дыхательная поддержка пациентов с инфекцией COVID-19» . «Ланцет». Респираторная медицина . 8 (4): е18. дои : 10.1016/S2213-2600(20)30110-7 . ПМК 7129706 . ПМИД 32145829 .
- ^ Пэсли Дж. «Как атипичная пневмония напугала мир в 2003 году, заразив более 8000 человек и убив 774 человека» . Бизнес-инсайдер . Проверено 8 ноября 2020 г.
- ^ Дуклеф М. (26 сентября 2012 г.). «Ученые углубились в изучение генов SARS-подобного вируса» . Ассошиэйтед Пресс. Архивировано из оригинала 27 сентября 2012 г. Проверено 27 сентября 2012 г.
- ^ Фалько М (24 сентября 2012 г.). «Новый вирус, подобный атипичной пневмонии, представляет собой медицинскую загадку» . CNN Здоровье . Архивировано из оригинала 1 ноября 2013 г. Проверено 16 марта 2013 г.
- ^ «Новый вирус, подобный атипичной пневмонии, обнаружен на Ближнем Востоке» . Аль-Джазира . 24 сентября 2012 г. Архивировано из оригинала 9 марта 2013 г. Проверено 16 марта 2013 г.
- ^ Келланд К. (28 сентября 2012 г.). «Новый вирус нелегко распространяется между людьми: ВОЗ» . Рейтер . Архивировано из оригинала 24 ноября 2012 г. Проверено 16 марта 2013 г.
- ↑ Новый коронавирус — Точка ситуации: Un nouveau cas d'infection submité. Архивировано 8 июня 2013 г. в Wayback Machine (Новый коронавирус — Отчет о состоянии: новый случай подтвержденной инфекции) 12 мая 2013 г., Social-sante.gouv.fr.
- ^ «Передача MERS» . Центры по контролю и профилактике заболеваний (CDC) . 2019-08-02. Архивировано из оригинала 07.12.2019 . Проверено 10 декабря 2019 г.
- ^ "Новая коронавирусная инфекция" . Всемирная ассоциация здравоохранения. 22 мая 2013 г. Архивировано из оригинала 7 июня 2013 г. Проверено 23 мая 2013 г.
- ^ «MERS в США» Центр по контролю заболеваний . 2019-08-02. Архивировано из оригинала 15 декабря 2019 г. Проверено 10 декабря 2019 г.
- ^ Санг-Хун С (8 июня 2015 г.). «Путь вируса MERS: один человек, множество южнокорейских больниц» . Нью-Йорк Таймс . Архивировано из оригинала 15 июля 2017 г. Проверено 01 марта 2017 г.
- ^ «Коронавирус ближневосточного респираторного синдрома (MERS-CoV)» . ВОЗ . Архивировано из оригинала 18 октября 2019 г. Проверено 10 декабря 2019 г.
- ^ Редколлегия (29.01.2020). «Готов ли мир к коронавирусу? Недоверие к науке и институтам может стать серьезной проблемой, если вспышка усугубится» . Нью-Йорк Таймс . Проверено 30 января 2020 г.
- ^ «Заявление ВОЗ относительно кластера случаев пневмонии в Ухане, Китай» . www.who.int . 09.01.2020. Архивировано из оригинала 14 января 2020 г. Проверено 10 января 2020 г.
- ^ «Лабораторное тестирование людей с подозрением на инфекцию нового коронавируса (nCoV). Временные рекомендации, 10 января 2020 г.» (PDF) . Архивировано (PDF) из оригинала 20 января 2020 г. Проверено 14 января 2020 г.
- ^ «Новый коронавирус 2019, Ухань, Китай» . www.cdc.gov (CDC) . 2020-01-23. Архивировано из оригинала 20 января 2020 г. Проверено 23 января 2020 г.
- ^ «Новая коронавирусная инфекция 2019 года (Ухань, Китай): обновленная информация о вспышке» . Канада.ca . 21 января 2020 г.
- ^ Хуэй Д.С., И. Ажар Э., Мадани Т.А., Нтуми Ф., Кок Р., Дар О. и др. (февраль 2020 г.). «Сохраняющаяся эпидемическая угроза новых коронавирусов 2019-nCoV для глобального здравоохранения — последняя вспышка нового коронавируса в 2019 году в Ухане, Китай» . Международный журнал инфекционных заболеваний . 91 : 264–66. дои : 10.1016/j.ijid.2020.01.009 . ПМЦ 7128332 . ПМИД 31953166 .
- ^ Коэн Дж. (26 января 2020 г.). «Рынок морепродуктов в Ухане не может быть источником нового вируса, распространяющегося по всему миру» . ScienceMag Американская ассоциация содействия развитию науки. (АААС) . Архивировано из оригинала 27 января 2020 г. Проверено 29 января 2020 г.
- ^ Эшнер К. (28 января 2020 г.). «Мы до сих пор не уверены, откуда на самом деле взялся COVID-19» . Популярная наука . Архивировано из оригинала 30 января 2020 г. Проверено 30 января 2020 г.
- ^ Перейти обратно: а б с д и ж г час я дж к «Глава 24 – Коронавирусы» . Ветеринарная вирусология Феннера (Пятое изд.). Академическая пресса. 2017. С. 435–461. дои : 10.1016/B978-0-12-800946-8.00024-6 . ISBN 978-0-12-800946-8 . S2CID 219575461 .
- ^ Мерфи Ф.А., Гиббс Э.П., Хорзинек М.К., Студдарт М.Дж. (1999). Ветеринарная вирусология . Бостон: Академическая пресса. стр. 495–508. ISBN 978-0-12-511340-3 .
- ^ Перейти обратно: а б Тиротта Э., Карбахал К.С., Шаумбург К.С., Уитмен Л., Лейн Т.Е. (июль 2010 г.). «Клеточно-заместительная терапия для стимулирования ремиелинизации в вирусной модели демиелинизации» . Журнал нейроиммунологии . 224 (1–2): 101–07. дои : 10.1016/j.jneuroim.2010.05.013 . ПМК 2919340 . ПМИД 20627412 .
- ^ Перейти обратно: а б «Ветеринарное руководство Merck» . Ветеринарное руководство Merck . Проверено 8 июня 2020 г.
- ^ Перейти обратно: а б Банде Ф., Аршад СС, Беджо М.Х., Моейни Х., Омар А.Р. (2015). «Прогресс и проблемы в разработке вакцин против инфекционного бронхита птиц» . Журнал иммунологических исследований . 2015 : 424860. doi : 10.1155/2015/424860 . ПМЦ 4411447 . ПМИД 25954763 .
- ^ Кавана Д. (2007). «Коронавирусный вирус инфекционного бронхита птиц» . Ветеринарное исследование . 38 (2): 281–97. doi : 10.1051/vetres:2006055 . ПМИД 17296157 .

- ^ «Таксономический браузер (Птичий коронавирус)» . www.ncbi.nlm.nih.gov . Проверено 3 июня 2020 г.
- ^ Чжоу П., Фань Х., Лан Т., Ян С.Л., Ши В.Ф., Чжан В. и др. (апрель 2018 г.). «Синдром смертельной острой диареи свиней, вызванный HKU2-родственным коронавирусом летучей мыши» . Природа . 556 (7700): 255–58. Бибкод : 2018Natur.556..255Z . дои : 10.1038/s41586-018-0010-9 . ПМК 7094983 . ПМИД 29618817 .
- ^ Вэй Х, Ше Г, У Т, Сюэ С, Цао Ю (февраль 2020 г.). «ВЭДВ проникает в клетки посредством эндоцитоза, опосредованного клатрином, кавеолами и липидными рафтами, и проникает через эндо-/лизосомный путь» . Ветеринарное исследование . 51 (1): 10. дои : 10.1186/s13567-020-0739-7 . ПМК 7011528 . ПМИД 32041637 .
- ^ Перейти обратно: а б с «Браузер таксономии (Альфакоронавирус 1)» . www.ncbi.nlm.nih.gov . Проверено 8 июня 2020 г.
- ^ Круз Дж.Л., Сола И., Бекарес М., Альберка Б., Плана Дж., Энхуанес Л. и др. (июнь 2011 г.). «Ген 7 коронавируса противодействует защите хозяина и модулирует вирулентность вируса» . ПЛОС Патогены . 7 (6): e1002090. дои : 10.1371/journal.ppat.1002090 . ПМЦ 3111541 . ПМИД 21695242 .
- ^ Круз Х.Л., Бекарес М., Сола И., Оливерос Х.К., Энхуанес Л., Суньига С. (сентябрь 2013 г.). «Белок 7 альфакоронавируса модулирует врожденный иммунный ответ хозяина» . Журнал вирусологии . 87 (17): 9754–67. дои : 10.1128/JVI.01032-13 . ПМК 3754097 . ПМИД 23824792 .
- ^ Перейти обратно: а б «Браузер таксономии (бетакоронавирус 1)» . www.ncbi.nlm.nih.gov . Проверено 8 июня 2020 г.
- ^ «Браузер таксономии (Альфакоронавирус)» . www.ncbi.nlm.nih.gov . Проверено 8 июня 2020 г.
- ^ Мюррей Дж. (16 апреля 2014 г.). «Что нового в FIP-подобной болезни хорьков?» (xls) . Архивировано из оригинала 24 апреля 2014 г. Проверено 24 апреля 2014 г.
- ^ «Инфекционные болезни хорьков – экзотических и лабораторных животных» . Ветеринарное руководство Merck . Проверено 8 июня 2020 г.
- ^ Перейти обратно: а б «Браузер таксономии (Embecovirus)» . www.ncbi.nlm.nih.gov . Проверено 8 июня 2020 г.
- ^ Вайс С.Р., Навас-Мартин С. (декабрь 2005 г.). «Патогенез коронавируса и возникший возбудитель тяжелого острого респираторного синдрома коронавирус» . Обзоры микробиологии и молекулярной биологии . 69 (4): 635–64. дои : 10.1128/MMBR.69.4.635-664.2005 . ПМК 1306801 . ПМИД 16339739 .
- ^ «Кишечный коронавирус» . Болезни экспериментальных животных . Архивировано из оригинала 01 июля 2019 г. Проверено 24 января 2020 г.
- ^ «Отслеживание вакцин и методов лечения COVID-19 (выберите вкладку «Вакцины или методы лечения», примените фильтры для просмотра выбранных данных)» . Институт Милкена. 03.11.2020 . Проверено 3 ноября 2020 г.
- ^ Перейти обратно: а б «Отслеживание вакцины и терапии COVID-19» . БиоРендер. 2020-10-30 . Проверено 3 ноября 2020 г.
- ^ Донг Л., Ху С., Гао Дж. (2020). «Открытие лекарств для лечения коронавирусной болезни 2019 (COVID-19)» . Открытия лекарств и терапия . 14 (1): 58–60. дои : 10.5582/ддт.2020.01012 . ПМИД 32147628 .
Дальнейшее чтение
- Ачесон, Нью-Хэмпшир (2011). «Глава 14: Коронавирусы». Основы молекулярной вирусологии . Хобокен, Нью-Джерси: John Wiley & Sons. стр. 159–171. ISBN 978-0-470-90059-8 .
- Алван А., Маджур Дж., Мемиш З.А. (2013). «Новая коронавирусная инфекция: время быть на шаг впереди» . Восточно-Средиземноморский журнал здравоохранения . 19 (Приложение 1): С3–4. дои : 10.26719/2013.19.supp1.S3 . ПМИД 23888787 . [ мертвая ссылка ]
- Лауд Х., Расшарт Д., Дельмас Б., Годе М., Гельфи Дж., Чарли Б. (июнь 1990 г.). «Молекулярная биология вируса трансмиссивного гастроэнтерита» . Ветеринарная микробиология . 23 (1–4): 147–54. дои : 10.1016/0378-1135(90)90144-К . ПМЦ 7117338 . ПМИД 2169670 .
- Сола И, Алонсо С, Суньига С, Балаш М, Плана-Дуран Х, Энхуанес Л (апрель 2003 г.). «Инженерия генома вируса трансмиссивного гастроэнтерита как вектора экспрессии, индуцирующего лактогенный иммунитет» . Журнал вирусологии . 77 (7): 4357–69. doi : 10.1128/JVI.77.7.4357-4369.2003 . ПМК 150661 . ПМИД 12634392 .
- Тадзима М (1970). «Морфология вируса трансмиссивного гастроэнтерита свиней. Возможный представитель коронавирусов. Краткий отчет» . Архив всех вирусных исследований . 29 (1): 105–08. дои : 10.1007/BF01253886 . ПМК 7086923 . ПМИД 4195092 . S2CID 42104521 .
| Базы данных органов управления : Национальные |
|---|