Вирус
| Вирус | |
|---|---|
 | |
| SARS-CoV-2 , представитель подсемейства Coronavirinae. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Царства | |
Вирус , — субмикроскопический инфекционный агент только внутри живых клеток организма . который размножается [1] Вирусы заражают все формы жизни , от животных и растений до микроорганизмов , включая бактерии и археи . [2] [3] Вирусы встречаются практически во всех экосистемах Земли и являются самым многочисленным типом биологических объектов. [4] [5] Со времени статьи Дмитрия Ивановского 1892 года, описывающей небактериальный патоген, поражающий растения табака, и открытия вируса табачной мозаики Мартинусом Бейеринком в 1898 году, [6] : 4 более 11 000 из миллионов видов вирусов . подробно описано [7] [8] Изучение вирусов известно как вирусология , раздел микробиологии .
При заражении клетка-хозяин часто вынуждена быстро производить тысячи копий исходного вируса. Находясь вне инфицированной клетки или в процессе заражения клетки, вирусы существуют в виде независимых вирусных частиц, или вирионов , состоящих из (i) генетического материала . е. длинных молекул ДНК , т или РНК , кодирующих структуру белки, с помощью которых действует вирус; (ii) белковая оболочка, капсид , которая окружает и защищает генетический материал; и в некоторых случаях (iii) внешняя оболочка из липидов . Формы этих вирусных частиц варьируются от простых спиральных и икосаэдрических форм до более сложных структур. У большинства видов вирусов вирионы слишком малы, чтобы их можно было увидеть в оптический микроскоп , и они составляют одну сотую размера большинства бактерий.
Происхождение вирусов в эволюционной истории жизни до сих пор неясно. Некоторые вирусы, возможно, произошли от плазмид — фрагментов ДНК, способных перемещаться между клетками. Другие вирусы, возможно, произошли от бактерий. В эволюции вирусы являются важным средством горизонтального переноса генов , который увеличивает генетическое разнообразие аналогично половому размножению . [9] считают вирусы Некоторые биологи формой жизни, поскольку они несут генетический материал, размножаются и развиваются посредством естественного отбора , хотя им не хватает ключевых характеристик, таких как клеточная структура, которые обычно считаются необходимыми критериями для определения жизни. Поскольку вирусы обладают некоторыми, но не всеми такими качествами, их называют «организмами на грани жизни». [10] и как репликаторы . [11]
Вирусы распространяются разными способами. Один из путей передачи – через болезнетворные организмы, известные как переносчики : например, вирусы часто передаются от растения к растению насекомыми, питающимися соком растений , такими как тля ; а вирусы животных могут переносить кровососущие насекомые. Многие вирусы распространяются в воздухе при кашле и чихании, включая вирусы гриппа , SARS-CoV-2 , ветряной оспы , оспы и кори . Норовирус и ротавирус , частые причины вирусного гастроэнтерита , передаются фекально-оральным путем , при контакте из рук в рот, через пищу или воду. Инфицирующая доза норовируса, необходимая для заражения человека, составляет менее 100 частиц. [12] ВИЧ – один из нескольких вирусов, передающихся половым путем и при контакте с инфицированной кровью. Разнообразие клеток-хозяев, которые вирус может заразить, называется диапазоном хозяев : он узкий для вирусов, специализирующихся на инфицировании только нескольких видов, или широкий для вирусов, способных заражать многие виды. [13] : 123–124
Вирусные инфекции у животных вызывают иммунный ответ , который обычно уничтожает заражающий вирус. Иммунные реакции также могут быть вызваны вакцинами , которые придают искусственно приобретенный иммунитет к конкретной вирусной инфекции. Некоторые вирусы, в том числе те, которые вызывают ВИЧ/СПИД , инфекцию ВПЧ и вирусный гепатит , уклоняются от этих иммунных реакций и приводят к хроническим инфекциям. несколько классов противовирусных препаратов Разработано .
Этимология
Английское слово «вирус» происходит от латинского vīrus , что означает яд и другие ядовитые жидкости. Вирус происходит от того же индоевропейского корня, что и санскритская виша , авестийская виша и древнегреческий ἰός ( iós ), которые все означают «яд». Первое засвидетельствованное использование слова «вирус» на английском языке появилось в 1398 году в Джона Тревизы переводе » Бартоломеуса Англикуса «De Proprietatibus Rerum . [14] [15] Вирулентный , от латинского virulentus («ядовитый»), датируется ок. 1400 . [16] [17] Значение слова «агент, вызывающий инфекционное заболевание» впервые зафиксировано в 1728 году. [15] задолго до открытия вирусов Дмитрием Ивановским в 1892 году. Английское множественное число — вирусы (иногда также вира ), [18] тогда как латинское слово является массовым существительным , которое не имеет классически подтвержденного множественного числа ( вира используется в неолатинском языке). [19] ). Прилагательное вирусное датируется 1948 годом. [20] Термин вирион (множественное число вирионов ), появившийся в 1959 г., [21] также используется для обозначения одной вирусной частицы, которая высвобождается из клетки и способна инфицировать другие клетки того же типа. [22]
Происхождение
Вирусы встречаются везде, где есть жизнь, и, вероятно, существовали с момента появления живых клеток. [23] Происхождение вирусов неясно, поскольку они не образуют окаменелостей, поэтому молекулярные методы . для выяснения того, как они возникли, используются [24] Кроме того, вирусный генетический материал иногда интегрируется в зародышевую линию организма-хозяина, благодаря чему он может передаваться вертикально потомству хозяина на протяжении многих поколений. бесценный источник информации Это предоставляет палеовирологам для отслеживания древних вирусов, существовавших еще миллионы лет назад.
Существуют три основные гипотезы, призванные объяснить происхождение вирусов: [25]
- Регрессивная гипотеза
- Вирусы, возможно, когда-то представляли собой маленькие клетки, паразитирующие на более крупных клетках. Со временем гены, не необходимые для их паразитизма, были потеряны. Бактерии риккетсии и хламидии представляют собой живые клетки, которые, как и вирусы, могут размножаться только внутри клеток-хозяев. Они поддерживают эту гипотезу, поскольку их зависимость от паразитизма, вероятно, привела к потере генов, которые позволили им выжить вне клетки. Это еще называют «гипотезой вырождения». [6] : 16 [26] : 11 или «гипотеза редукции». [27] : 24
- Гипотеза клеточного происхождения
- Некоторые вирусы могли возникнуть из фрагментов ДНК или РНК, «ускользнувших» от генов более крупного организма. Ускользнувшая ДНК могла произойти из плазмид (кусков обнаженной ДНК, которые могут перемещаться между клетками) или транспозонов (молекул ДНК, которые реплицируются и перемещаются в разные положения внутри генов клетки). [13] : 810 Транспозоны, которые когда-то называли «прыгающими генами», являются примерами мобильных генетических элементов и могут быть источником некоторых вирусов. Они были обнаружены в кукурузе Барбарой МакКлинток в 1950 году. [28] Иногда это называют «гипотезой бродяжничества». [6] : 16 [26] : 11–12 или «гипотеза побега». [27] : 24
- Гипотеза коэволюции
- Это также называется «гипотезой первоочередности вирусов». [27] : 24 и предполагает, что вирусы могли развиться из сложных молекул белка и нуклеиновой кислоты в то же время, когда клетки впервые появились на Земле, и зависели от клеточной жизни в течение миллиардов лет. Вироиды — это молекулы РНК, которые не классифицируются как вирусы, поскольку у них отсутствует белковая оболочка. Они имеют характеристики, общие для нескольких вирусов, и их часто называют субвирусными агентами . [6] : 55 Вироиды – важные патогены растений. [13] : 791 Они не кодируют белки, но взаимодействуют с клеткой-хозяином и используют механизмы хозяина для своей репликации. [29] Вирус гепатита дельта человека имеет геном РНК, сходный с геномом вироидов, но имеет белковую оболочку, полученную из вируса гепатита В, и не может производить собственную. Следовательно, это дефектный вирус. Хотя геном вируса гепатита дельта может реплицироваться независимо внутри клетки-хозяина, ему требуется помощь вируса гепатита В, чтобы обеспечить белковую оболочку, чтобы его можно было передать новым клеткам. [13] : 460 Аналогичным образом вирофаг-спутник зависит от мимивируса , который заражает простейших Acanthamoeba castellanii . [30] Эти вирусы, существование которых зависит от присутствия других видов вирусов в клетке-хозяине, называются « сателлитами » и могут представлять собой эволюционные промежуточные продукты вироидов и вирусов. [26] : 777 > [6] : 55–57
В прошлом со всеми этими гипотезами были проблемы: регрессивная гипотеза не объясняла, почему даже самые маленькие клеточные паразиты никоим образом не похожи на вирусы. Гипотеза побега не объясняла сложные капсиды и другие структуры вирусных частиц. Гипотеза «вирус прежде всего» противоречит определению вирусов, поскольку им необходимы клетки-хозяева. [27] : 24 Сегодня вирусы признаны древними и имеют происхождение, предшествовавшее расколу жизни на три домена . [27] : 28 Это открытие заставило современных вирусологов пересмотреть и переоценить эти три классические гипотезы. [27] : 28
Доказательства существования древнего мира РНК- клеток [27] : 26 а компьютерный анализ последовательностей вирусной ДНК и ДНК хозяина дает лучшее понимание эволюционных взаимоотношений между различными вирусами и может помочь идентифицировать предков современных вирусов. На сегодняшний день такой анализ не доказал, какая из этих гипотез верна. [27] : 26 Кажется маловероятным, что все известные в настоящее время вирусы имеют общего предка, и вирусы, вероятно, возникали в прошлом много раз с помощью одного или нескольких механизмов. [31]
Микробиология
Открытие
Первые доказательства существования вирусов были получены в результате экспериментов с фильтрами, поры которых были достаточно малы, чтобы удерживать бактерии. В 1892 году Дмитрий Ивановский использовал один из этих фильтров, чтобы показать, что сок больного растения табака остается заразным для здоровых растений табака, несмотря на фильтрацию. Мартинус Бейеринк назвал профильтрованное инфекционное вещество «вирусом», и это открытие считается началом вирусологии.Последующее открытие и частичная характеристика бактериофагов Фредериком Твортом и Феликсом д'Эрелем еще больше послужили катализатором развития этой области, и к началу 20 века было открыто множество вирусов. В 1926 году Томас Милтон Риверс определил вирусы как облигатные паразиты. продемонстрировал, что вирусы представляют собой частицы, а не жидкость, Уэнделл Мередит Стэнли а изобретение электронного микроскопа в 1931 году позволило визуализировать их сложные структуры. [32]
Свойства жизни
Научные мнения расходятся во мнениях относительно того, являются ли вирусы формой жизни или органическими структурами, взаимодействующими с живыми организмами. [11] Их описывают как «организмы на грани жизни». [10] поскольку они напоминают организмы в том смысле, что обладают генами , развиваются путем естественного отбора , [33] и размножаются, создавая множество своих копий посредством самосборки. Хотя у них есть гены, у них нет клеточной структуры, которую часто считают основной единицей жизни. Вирусы не имеют собственного метаболизма и требуют клетки-хозяина для производства новых продуктов. Поэтому они не могут естественным образом размножаться вне клетки-хозяина. [34] — хотя некоторые бактерии, такие как риккетсии и хламидии, несмотря на то же ограничение, считаются живыми организмами. [35] [36] Принятые формы жизни используют деление клеток для размножения, тогда как вирусы спонтанно собираются внутри клеток. Они отличаются от автономного роста кристаллов тем, что наследуют генетические мутации , подвергаясь естественному отбору. Самосборка вируса внутри клеток-хозяев имеет значение для изучения происхождения жизни , поскольку она подтверждает гипотезу о том, что жизнь могла зародиться как самоорганизующиеся органические молекулы . [2] Модель вироцеллы , впервые предложенная Патриком Фортером, считает инфицированную клетку «живой формой» вирусов и что вирусные частицы (вирионы) аналогичны спорам . [37] Хотя дебаты о живом и неживом мире продолжаются, модель вироклеток получила некоторое признание. [38]
Структура



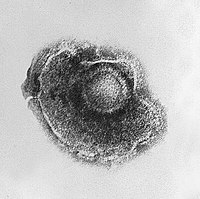
Вирусы имеют большое разнообразие размеров и форм, называемых « морфологиями ». В целом вирусы намного меньше бактерий, и внутри Escherichia coli поместится более тысячи вирусов-бактериофагов. клетки бактерии [39] : 98 Многие изученные вирусы имеют сферическую форму и диаметр от 20 до 300 нанометров . Некоторые филовирусы , представляющие собой нити, имеют общую длину до 1400 нм; их диаметр составляет всего около 80 нм. [26] : 33–55 Большинство вирусов невозможно увидеть в оптический микроскоп сканирующие и просвечивающие электронные микроскопы . , поэтому для их визуализации используются [26] : 33–37 Для увеличения контраста между вирусами и фоном используются электронно-плотные «пятна». Это растворы солей тяжелых металлов, например вольфрама , которые рассеивают электроны из участков, покрытых пятном. Когда вирионы покрыты красителем (положительное окрашивание), мелкие детали не видны. Негативное окрашивание решает эту проблему, окрашивая только фон. [40]
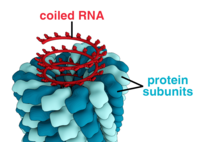
Полная вирусная частица, известная как вирион , состоит из нуклеиновой кислоты, окруженной защитной белковой оболочкой, называемой капсидом . Они образуются из белковых субъединиц, называемых капсомерами . [26] : 40 Вирусы могут иметь липидную «оболочку», полученную из мембраны клетки- хозяина . Капсид состоит из белков, кодируемых вирусным геномом , и его форма служит основой для морфологического различия. [41] [42] Субъединицы белка, кодируемые вирусом, будут самособираться с образованием капсида, что обычно требует присутствия вирусного генома. Сложные вирусы кодируют белки, которые помогают в построении их капсида. Белки, связанные с нуклеиновой кислотой, известны как нуклеопротеины , а ассоциация белков вирусного капсида с вирусной нуклеиновой кислотой называется нуклеокапсидом. Капсид и всю структуру вируса можно механически (физически) исследовать с помощью атомно-силовой микроскопии . [43] [44] В целом выделяют пять основных морфологических типов вирусов:
- спиральный
- Эти вирусы состоят из одного типа капсомеров, расположенных вокруг центральной оси и образующих спиральную структуру, которая может иметь центральную полость или трубку. Такое расположение приводит к образованию вирионов, которые могут представлять собой короткие и очень жесткие палочки или длинные и очень гибкие нити. Генетический материал (обычно одноцепочечная РНК, но в некоторых случаях и одноцепочечная ДНК) связан со спиралью белка за счет взаимодействия между отрицательно заряженной нуклеиновой кислотой и положительными зарядами белка. В целом длина спирального капсида связана с длиной содержащейся в нем нуклеиновой кислоты, а диаметр зависит от размера и расположения капсомеров. Хорошо изученный вирус табачной мозаики. [26] : 37 и иновирус [45] являются примерами спиральных вирусов.
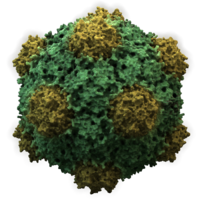
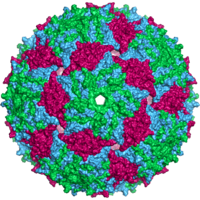
- икосаэдрический
- Большинство вирусов животных имеют икосаэдрическую или почти сферическую форму с хиральной икосаэдрической симметрией . Правильный икосаэдр — оптимальный способ образования замкнутой оболочки из одинаковых субъединиц. Минимальное количество капсомеров, необходимое для каждой треугольной грани, равно 3, что дает 60 для икосаэдра. Многие вирусы, такие как ротавирус, имеют более 60 капсомеров и кажутся сферическими, но сохраняют эту симметрию. Для этого капсомеры на вершинах окружены пятью другими капсомерами и называются пентонами. Капсомеры на треугольных гранях окружены шестью другими и называются гексонами . [26] : 40, 42 Гексоны по своей сути плоские, а пентоны, образующие 12 вершин, изогнуты. Один и тот же белок может выступать в качестве субъединицы как пентамеров, так и гексамеров, или они могут состоять из разных белков. [46]
- Растянуть
- Это икосаэдр, вытянутый по оси пятого порядка и являющийся обычным расположением головок бактериофагов. Эта конструкция состоит из цилиндра с крышками на обоих концах. [47]
- обернутый
- Некоторые виды вирусов окутывают себя модифицированной формой одной из клеточных мембран , либо внешней мембраной, окружающей инфицированную клетку-хозяина, либо внутренними мембранами, такими как ядерная мембрана или эндоплазматическая сеть , таким образом приобретая внешний липидный бислой, известный как вирусная оболочка . Эта мембрана покрыта белками, кодируемыми вирусным геномом и геномом хозяина; сама липидная мембрана и любые присутствующие углеводы полностью происходят от хозяина. Вирус гриппа , ВИЧ (вызывает СПИД ) и коронавирус 2 тяжелого острого респираторного синдрома (вызывает COVID-19 ). [48] используйте эту стратегию. Инфекционность большинства оболочечных вирусов зависит от оболочки. [26] : 42–43
- Сложный
- Эти вирусы обладают капсидом, который не является ни чисто спиральным, ни чисто икосаэдрическим и может иметь дополнительные структуры, такие как белковые хвосты или сложную внешнюю стенку. Некоторые бактериофаги, такие как фаг Enterobacteria T4 , имеют сложную структуру, состоящую из икосаэдрической головки, связанной со спиральным хвостом, который может иметь шестиугольную базовую пластинку с выступающими белковыми хвостовыми волокнами. Эта хвостовая структура действует как молекулярный шприц, прикрепляясь к бактериальному хозяину и затем вводя вирусный геном в клетку. [49]
Поксвирусы — это крупные сложные вирусы с необычной морфологией. Вирусный геном связан с белками внутри структуры центрального диска, известной как нуклеоид . Нуклеоид окружен мембраной и двумя латеральными тельцами неизвестной функции. Вирус имеет внешнюю оболочку с толстым слоем белка, покрывающим ее поверхность. Весь вирион слегка плеоморфен , от яйцевидной до кирпичной формы. [50]
Гигантские вирусы
Мимивирус — один из крупнейших охарактеризованных вирусов, диаметр капсида которого составляет 400 нм. Белковые нити размером 100 нм выступают из поверхности. Под электронным микроскопом капсид кажется шестиугольным, поэтому капсид, вероятно, икосаэдрический. [51] В 2011 году исследователи обнаружили самый крупный из известных на тот момент вирусов в образцах воды, собранных со дна океана у побережья Лас-Крусес, Чили. Его условно называют Megavirus chilensis , и его можно увидеть в простой оптический микроскоп. [52] В 2013 году род Pandoravirus был обнаружен в Чили и Австралии, его геномы примерно в два раза больше, чем у Megavirus и Mimivirus. [53] Все гигантские вирусы имеют геномы дцДНК и делятся на несколько семейств: Mimiviridae , Pithoviridae, Pandoraviridae , Phycodnaviridae . и Mollivirus род [54]
Некоторые вирусы, поражающие архей, имеют сложную структуру, не связанную с какой-либо другой формой вируса, и имеют широкий спектр необычных форм: от веретенообразных структур до вирусов, напоминающих крючковидные стержни, капли или даже бутылки. Другие архейные вирусы напоминают хвостатые бактериофаги и могут иметь несколько хвостовых структур. [55]
Геном
| Свойство | Параметры |
|---|---|
| Нуклеиновая кислота |
|
| Форма |
|
| Неблагополучие |
|
| Смысл |
|
можно наблюдать огромное разнообразие геномных структур У вирусных видов ; Как группа, они содержат больше структурного геномного разнообразия, чем растения, животные, археи или бактерии. Существуют миллионы различных типов вирусов, [8] хотя подробно описано менее 7000 типов. [6] : 49 По состоянию на январь 2021 года база данных геномов вируса NCBI насчитывает более 193 000 полных последовательностей генома. [56] но, несомненно, еще многое предстоит открыть. [57] [58]
Вирус имеет геном ДНК или РНК и называется ДНК-вирусом или РНК-вирусом соответственно. Большинство вирусов имеют геномы РНК. Вирусы растений, как правило, имеют геномы с одноцепочечной РНК, а бактериофаги, как правило, имеют геномы с двухцепочечной ДНК. [26] : 96–99
Вирусные геномы кольцевые, как у полиомавирусов , или линейные, как у аденовирусов . Тип нуклеиновой кислоты не имеет отношения к форме генома. Среди РНК-вирусов и некоторых ДНК-вирусов геном часто разделен на отдельные части, в этом случае его называют сегментированным. У РНК-вирусов каждый сегмент часто кодирует только один белок, и они обычно находятся вместе в одном капсиде. Для того чтобы вирус был инфекционным, не обязательно, чтобы все сегменты находились в одном и том же вирионе, как это продемонстрировали вирус бромовой мозаики и некоторые другие вирусы растений. [26] : 33–35
Вирусный геном, независимо от типа нуклеиновой кислоты, почти всегда либо одноцепочечный (ss), либо двухцепочечный (ds). Одноцепочечные геномы состоят из неспаренной нуклеиновой кислоты, аналогичной половине лестницы, разделенной посередине. Двухцепочечные геномы состоят из двух комплементарных пар нуклеиновых кислот, аналогично лестнице. Вирусные частицы некоторых семейств вирусов, например, принадлежащих к Hepadnaviridae , содержат геном, который частично является двухцепочечным и частично одноцепочечным. [26] : 96–99
Говорят, что для большинства вирусов с геномами РНК и некоторых с геномами одноцепочечной ДНК (оцДНК) одиночные цепи имеют либо положительный смысл (так называемый «плюс-цепь»), либо отрицательный смысл (так называемый «минус-цепь»). ), в зависимости от того, комплементарны ли они вирусной информационной РНК (мРНК). Вирусная РНК с положительным смыслом имеет тот же смысл, что и вирусная мРНК, и, таким образом, по крайней мере ее часть может быть немедленно транслирована клеткой-хозяином. Вирусная РНК с отрицательным смыслом комплементарна мРНК и, следовательно, перед трансляцией должна быть преобразована в РНК с положительным смыслом с помощью РНК-зависимой РНК-полимеразы . Номенклатура ДНК для вирусов с геномной оцДНК аналогична номенклатуре РНК в том, что вирусная оцДНК с положительной цепью идентична по последовательности вирусной мРНК и, таким образом, является кодирующей цепью, тогда как вирусная оцДНК с отрицательным смыслом комплементарна вирусной мРНК и, таким образом, шаблонная прядь. [26] : 96–99 Некоторые типы вирусов оцДНК и оцРНК имеют амбисенсные геномы , поскольку транскрипция может происходить с обеих цепей в двухцепочечном репликативном промежуточном продукте. Примеры включают геминивирусы , которые представляют собой оцРНК-вирусы растений, и аренавирусы , которые представляют собой оцРНК-вирусы животных. [59]
Размер генома
Размер генома сильно различается у разных видов. Самые маленькие — цирковирусы с оцДНК семейства Circoviridae — кодируют только два белка и имеют размер генома всего две тысячи оснований; [60] самые крупные из них — пандоравирусы — имеют размеры генома около двух мегабаз, которые кодируют около 2500 белков. [53] Гены вирусов редко имеют интроны и часто располагаются в геноме таким образом, что перекрываются . [61]
Как правило, РНК-вирусы имеют меньший размер генома, чем ДНК-вирусы, из-за более высокой частоты ошибок при репликации и имеют максимальный верхний предел размера. [24] Помимо этого, ошибки при репликации делают вирус бесполезным или неконкурентоспособным. Чтобы компенсировать это, РНК-вирусы часто имеют сегментированные геномы — геном разделен на более мелкие молекулы — что снижает вероятность того, что ошибка в однокомпонентном геноме выведет из строя весь геном. Напротив, ДНК-вирусы обычно имеют более крупные геномы из-за высокой точности их ферментов репликации. [62] Вирусы с одноцепочечной ДНК являются исключением из этого правила, поскольку частота мутаций в этих геномах может приближаться к пределу, свойственному вирусу с оцРНК. [63]
Генетическая мутация и рекомбинация

Вирусы претерпевают генетические изменения по нескольким механизмам. К ним относится процесс, называемый антигенным дрейфом , при котором отдельные основания ДНК или РНК мутируют в другие основания. Большинство этих точковых мутаций «тихие» — они не меняют белок, который кодирует ген, — но другие могут давать эволюционные преимущества, такие как устойчивость к противовирусным препаратам . [64] [65] Антигенный сдвиг происходит, когда в геноме вируса происходят серьезные изменения. Это может быть результатом рекомбинации или реассортации . Вирус гриппа А очень склонен к реассортации; иногда это приводило к появлению новых штаммов , которые вызывали пандемии . [66] РНК-вирусы часто существуют как квазивиды или группы вирусов одного и того же вида, но с несколько разными нуклеозидными последовательностями генома. Такие квазивиды являются главной мишенью естественного отбора. [67]
Сегментированные геномы дают эволюционные преимущества; разные штаммы вируса с сегментированным геномом могут перемешивать и комбинировать гены и производить вирусы-потомки (или потомки), обладающие уникальными характеристиками. Это называется реассортацией или «вирусным сексом». [68]
Генетическая рекомбинация — это процесс, при котором цепь ДНК (или РНК) разрывается, а затем присоединяется к концу другой молекулы ДНК (или РНК). Это может произойти, когда вирусы одновременно заражают клетки, и исследования вирусной эволюции показали, что у изученных видов широко распространена рекомбинация. [69] Рекомбинация свойственна как РНК-, так и ДНК-вирусам. [70] [71]
Коронавирусы имеют геном однонитевой положительной РНК . Репликация генома катализируется РНК -зависимой РНК-полимеразой . Механизм рекомбинации, используемый коронавирусами, вероятно, включает переключение матрицы полимеразой во время репликации генома. [72] Этот процесс, по-видимому, является адаптацией для борьбы с повреждением генома. [73]
Цикл репликации

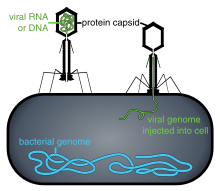
Вирусные популяции не растут за счет деления клеток, поскольку они бесклеточные. Вместо этого они используют механизм и метаболизм клетки-хозяина для производства множества своих копий и собираются в клетке. [74] При заражении клетка-хозяин вынуждена быстро производить тысячи копий исходного вируса. [75]
Их жизненный цикл сильно различается у разных видов, но в их жизненном цикле есть шесть основных стадий: [26] : 75–91
Прикрепление представляет собой специфическое связывание белков вирусного капсида со специфическими рецепторами на поверхности клетки-хозяина. Эта специфичность определяет диапазон хозяев и тип клетки-хозяина вируса. Например, ВИЧ поражает ограниченный круг лейкоцитов человека . Это связано с тем, что его поверхностный белок gp120 специфически взаимодействует с молекулой CD4 — хемокиновым рецептором — который чаще всего встречается на поверхности CD4+ Т-клеток . Этот механизм развился в пользу тех вирусов, которые заражают только те клетки, в которых они способны к репликации. Присоединение к рецептору может вызвать изменения в белке вирусной оболочки, которые приводят к слиянию вирусных и клеточных мембран или изменениям необолоченных поверхностных белков вируса, которые позволяют вирусу проникнуть. [76]
Проникновение или проникновение вируса следует за прикреплением: вирионы проникают в клетку-хозяина посредством рецептор-опосредованного эндоцитоза или слияния мембран . Заражение клеток растений и грибов отличается от заражения клеток животных. У растений жесткая клеточная стенка состоит из целлюлозы , а у грибов — из хитина, поэтому большинство вирусов могут проникнуть внутрь этих клеток только после травмы клеточной стенки. [6] : 70 Почти все вирусы растений (например, вирус табачной мозаики) также могут перемещаться непосредственно от клетки к клетке в виде одноцепочечных нуклеопротеиновых комплексов через поры, называемые плазмодесмами . [77] Бактерии, как и растения, имеют прочные клеточные стенки, которые вирус должен пробить, чтобы заразить клетку. Учитывая, что клеточные стенки бактерий намного тоньше стенок растительных клеток из-за их гораздо меньшего размера, некоторые вирусы развили механизмы, которые вводят свой геном в бактериальную клетку через клеточную стенку, в то время как вирусный капсид остается снаружи. [6] : 71
Снятие покрытия — это процесс удаления вирусного капсида: это может происходить путем разложения вирусными ферментами или ферментами хозяина или простой диссоциацией; конечным результатом является высвобождение вирусной геномной нуклеиновой кислоты. [78]
Репликация вирусов предполагает прежде всего размножение генома. Репликация включает синтез вирусной информационной РНК (мРНК) из «ранних» генов (за исключением РНК-вирусов с положительным смыслом), синтез вирусного белка , возможную сборку вирусных белков, а затем репликацию вирусного генома, опосредованную экспрессией раннего или регуляторного белка. Для сложных вирусов с более крупными геномами за этим может последовать один или несколько дополнительных раундов синтеза мРНК: «поздняя» экспрессия генов, как правило, связана со структурными или вирионными белками. [79]
Сборка . После структурно-опосредованной самосборки вирусных частиц часто происходит некоторая модификация белков. У вирусов, таких как ВИЧ, эта модификация (иногда называемая созреванием) происходит после того, как вирус высвобождается из клетки-хозяина. [80]
Высвобождение . Вирусы могут высвобождаться из клетки-хозяина путем лизиса , процесса, который убивает клетку, разрывая ее мембрану и клеточную стенку, если она присутствует: это особенность многих бактериальных и некоторых вирусов животных. Некоторые вирусы подвергаются лизогенному циклу , при котором вирусный геном встраивается путем генетической рекомбинации в определенное место хромосомы хозяина. Вирусный геном тогда известен как « провирус » или, в случае бактериофагов, « профаг ». [13] : 836 Всякий раз, когда хозяин делится, вирусный геном также реплицируется. Вирусный геном внутри хозяина в основном «молчит». В какой-то момент провирус или профаг могут дать начало активному вирусу, который может лизировать клетки-хозяева. [6] : 243–259 Вирусы с оболочкой (например, ВИЧ) обычно высвобождаются из клетки-хозяина путем отпочкования . В ходе этого процесса вирус приобретает оболочку, которая представляет собой модифицированный кусок плазмы хозяина или другой внутренней мембраны. [6] : 185–187
Репликация генома
Генетический материал внутри вирусных частиц и метод его репликации значительно различаются у разных типов вирусов.
- ДНК-вирусы
- Репликация генома большинства ДНК-вирусов клетки происходит в ядре . Если клетка имеет на своей поверхности соответствующий рецептор, эти вирусы проникают в клетку либо путем прямого слияния с клеточной мембраной (например, герпесвирусы), либо, что чаще, путем рецептор-опосредованного эндоцитоза. Большинство ДНК-вирусов полностью зависят от механизмов синтеза ДНК и РНК и механизмов обработки РНК в клетке-хозяине. Вирусы с более крупными геномами могут сами кодировать большую часть этого механизма. У эукариот вирусный геном должен пересечь ядерную мембрану клетки, чтобы получить доступ к этому механизму, тогда как у бактерий ему достаточно проникнуть в клетку. [13] : 118 [26] : 78
- РНК-вирусы
- Репликация РНК-вирусов обычно происходит в цитоплазме . РНК-вирусы можно разделить на четыре различные группы в зависимости от способов их репликации. Полярность (независимо от того , может ли она использоваться непосредственно рибосомами для создания белков) одноцепочечных РНК-вирусов во многом определяет механизм репликации; другой важный критерий — является ли генетический материал одноцепочечным или двухцепочечным. Все РНК-вирусы используют собственные ферменты РНК-репликазы для создания копий своих геномов. [26] : 79
- Вирусы с обратной транскрипцией
- Вирусы с обратной транскрипцией содержат в своих частицах оцРНК ( Retroviridae , Metaviridae , Pseudoviridae ) или дцДНК ( Caulimoviridae и Hepadnaviridae ). Вирусы с обратной транскрипцией с геномами РНК ( ретровирусы ) используют промежуточную ДНК для репликации, тогда как вирусы с геномами ДНК ( параретровирусы ) используют промежуточную РНК во время репликации генома. Оба типа используют обратную транскриптазу или РНК-зависимый фермент ДНК-полимеразу для преобразования нуклеиновой кислоты. Ретровирусы интегрируют ДНК, полученную путем обратной транскрипции, в геном хозяина в виде провируса как часть процесса репликации; параретровирусы этого не делают, хотя интегрированные копии генома параретровирусов растений, особенно растений, могут давать начало инфекционным вирусам. [81] Они чувствительны к противовирусным препаратам , ингибирующим фермент обратной транскриптазы, например зидовудину и ламивудину . Примером первого типа является ВИЧ, который является ретровирусом. Примерами второго типа являются Hepadnaviridae , включающие вирус гепатита В. [26] : 88–89
Цитопатическое воздействие на клетку-хозяина
Спектр структурного и биохимического воздействия вирусов на клетку-хозяина обширен. [26] : 115–146 Это так называемые « цитопатические эффекты ». [26] : 115 Большинство вирусных инфекций в конечном итоге приводят к гибели клетки-хозяина. Причины смерти включают лизис клеток, изменения поверхностной мембраны клетки и апоптоз . [82] Нередко гибель клетки вызвана прекращением ее нормальной деятельности из-за подавления вирусспецифическими белками, не все из которых входят в состав вирусной частицы. [83] Различие между цитопатическими и безвредными происходит постепенно. Некоторые вирусы, такие как вирус Эпштейна-Барра , могут вызывать пролиферацию клеток, не вызывая злокачественных опухолей. [84] в то время как другие, такие как вирусы папилломы , являются установленными причинами рака. [85]
Спящие и латентные инфекции.
Некоторые вирусы не вызывают видимых изменений в инфицированной клетке. Клетки, в которых вирус находится в латентном состоянии и неактивен, проявляют мало признаков инфекции и часто функционируют нормально. [86] Это вызывает стойкие инфекции, и вирус часто находится в состоянии покоя в течение многих месяцев или лет. Это часто бывает с вирусами герпеса . [87] [88]
Диапазон хостов
Вирусы, безусловно, являются наиболее распространенными биологическими объектами на Земле, и их численность превосходит все остальные вместе взятые. [89] Они заражают все виды клеточной жизни, включая животных, растения, бактерии и грибы . [6] : 49 Различные типы вирусов могут инфицировать лишь ограниченный круг хозяев, и многие из них являются видоспецифичными. Некоторые из них, например , вирус оспы , могут инфицировать только один вид — в данном случае человека. [13] : 643 и, как говорят, имеют узкий круг хозяев . Другие вирусы, такие как вирус бешенства, могут инфицировать различные виды млекопитающих и, как говорят, имеют широкий спектр. [13] : 631 Вирусы, поражающие растения, безвредны для животных, а большинство вирусов, поражающих других животных, безвредны для человека. [6] : 272 Диапазон хозяев некоторых бактериофагов ограничен одним штаммом бактерий, и их можно использовать для отслеживания источника вспышек инфекций с помощью метода, называемого фаговым типированием . [90] Полный набор вирусов в организме или среде обитания называется виромом ; например, все человеческие вирусы составляют человеческий виром . [91]
Новые вирусы
Новый вирус – это тот, который ранее не был зарегистрирован. Это может быть вирус, выделенный из природного резервуара или выделенный в результате распространения к животному или человеку-хозяину , где вирус ранее не был идентифицирован. Это может быть возникший вирус , представляющий собой новый вирус, но это также может быть существующий вирус, который ранее не был идентифицирован . [92] Коронавирус SARS -CoV-2, вызвавший пандемию COVID-19, является примером нового вируса. [93]
Классификация
Классификация направлена на описание разнообразия вирусов путем присвоения им названий и группировки на основе сходства. В 1962 году Андре Львофф , Роберт Хорн и Поль Турнье первыми разработали способ классификации вирусов, основанный на иерархической системе Линнея . [94] Эта система основана на классификации по типу , классу , отряду , семейству , роду и виду . Вирусы были сгруппированы в соответствии с их общими свойствами (а не свойствами их хозяев) и типом нуклеиновой кислоты, образующей их геномы. [95] В 1966 году был создан Международный комитет по таксономии вирусов (ICTV). Система, предложенная Львоффом, Хорном и Турнье, изначально не была принята ICTV, поскольку небольшой размер генома вирусов и высокая скорость их мутаций затрудняли определение их происхождения вне порядка. Таким образом, система классификации Балтимора стала использоваться в качестве дополнения к более традиционной иерархии. [96] Начиная с 2018 года ICTV начал признавать более глубокие эволюционные связи между вирусами, которые были обнаружены с течением времени, и принял 15-ранговую систему классификации, варьирующуюся от области до вида. [97] Кроме того, некоторые виды одного рода объединены в геногруппу . [98] [99]
Классификация ICTV
ICTV разработала действующую систему классификации и написала рекомендации, которые придают больший вес определенным свойствам вируса для поддержания единообразия семейства. Установлена единая таксономия (универсальная система классификации вирусов). [100] Изучена лишь небольшая часть от общего разнообразия вирусов. [101] 6 сфер, 10 царств, 17 типов, 2 подтипа, 40 классов, 72 порядка, 8 подотрядов, 264 семейства, 182 подсемейства , 2818 родов, 84 подрода и 11 273 вида вирусов. По состоянию на 2022 год ICTV определило [7]
Ниже представлена общая таксономическая структура ареалов таксонов и суффиксы, используемые в таксономических названиях. По состоянию на 2022 год ранги подцарства, подцарства и подкласса не используются, тогда как все остальные ранги используются. [7]
- Царство ( -вирия )
- Подцарство ( -вира )
- Королевство ( -virae )
- Подцарство ( -вира )
Балтиморская классификация

Лауреат Нобелевской премии по биологии Дэвид Балтимор разработал систему классификации Балтимора . [102] [103] Система классификации ICTV используется вместе с системой классификации Балтимора в современной классификации вирусов. [104] [105] [106]
Балтиморская классификация вирусов основана на механизме продукции мРНК . Вирусы должны генерировать мРНК из своих геномов, чтобы производить белки и воспроизводить себя, но для достижения этой цели в каждом семействе вирусов используются разные механизмы. Вирусные геномы могут быть одноцепочечными (ss) или двухцепочечными (ds), РНК или ДНК и могут использовать или не использовать обратную транскриптазу (RT). Кроме того, вирусы оцРНК могут быть смысловыми (+) или антисмысловыми (-). Эта классификация делит вирусы на семь групп:
- I: вирусы с дцДНК (например, аденовирусы , герпесвирусы , поксвирусы )
- II: ДНК вирусов оцДНК (+ цепь или «смысловая») ДНК (например, парвовирусов )
- III: вирусы дсРНК (например, реовирусы )
- IV: (+)оцРНК вирусов (+ цепь или смысловая) РНК (например , коронавирусы , пикорнавирусы , тогавирусы )
- В: (-)оцРНК вирусов (-цепь или антисмысловая РНК) (например, ортомиксовирусы , рабдовирусы )
- VI: вирусы оцРНК-RT (+ цепь или смысловая) РНК с промежуточной ДНК в жизненном цикле (например, ретровирусы )
- VII: дцДНК-RT ДНК вирусов с промежуточной РНК в жизненном цикле (например, гепаднавирусы )
Роль в заболеваниях человека

Примеры распространенных заболеваний человека, вызываемых вирусами, включают простуду , грипп , ветряную оспу и герпес . Многие серьезные заболевания, такие как бешенство , болезнь, вызванная вирусом Эбола , СПИД (ВИЧ) , птичий грипп и атипичная пневмония, вызываются вирусами. Относительная способность вирусов вызывать заболевания описывается с точки зрения вирулентности . Другие заболевания исследуются, чтобы выяснить, есть ли у них вирус в качестве возбудителя, например, возможная связь между вирусом герпеса человека 6 (HHV6) и неврологическими заболеваниями, такими как рассеянный склероз и синдром хронической усталости . [108] Существуют разногласия по поводу того, может ли борнавирус , который ранее считался причиной неврологических заболеваний у лошадей, быть ответственным за психические заболевания у людей. [109]
Вирусы имеют разные механизмы, с помощью которых они вызывают заболевания в организме, что во многом зависит от вида вируса. Механизмы на клеточном уровне в первую очередь включают лизис клеток, разрыв и последующую гибель клетки. В многоклеточных организмах , если погибнет достаточное количество клеток, от последствий начнет страдать весь организм. Хотя вирусы вызывают нарушение здорового гомеостаза , приводящее к заболеванию, они могут относительно безвредно существовать внутри организма. Примером может служить способность вируса простого герпеса , вызывающего герпес, оставаться в спящем состоянии в организме человека. Это называется задержка [110] Он характерен для вирусов герпеса, включая вирус Эпштейна-Барра, вызывающий железистую лихорадку, и вирус ветряной оспы , вызывающий ветряную оспу и опоясывающий лишай . Большинство людей были инфицированы хотя бы одним из этих типов вируса герпеса. [111] Эти латентные вирусы иногда могут быть полезными, поскольку присутствие вируса может повысить иммунитет против бактериальных патогенов, таких как Yersinia pestis . [112]
Некоторые вирусы могут вызывать пожизненные или хронические инфекции, при которых вирусы продолжают размножаться в организме, несмотря на защитные механизмы хозяина. [113] Это часто встречается при инфекциях, вызванных вирусом гепатита В и вирусом гепатита С. Хронически инфицированных людей называют носителями, поскольку они служат резервуарами инфекционного вируса. [114] В популяциях с высокой долей носителей болезнь считается эндемической . [115]
Эпидемиология
Вирусная эпидемиология — раздел медицинской науки, занимающийся передачей вирусных инфекций у человека и контролем над ними. Передача вирусов может быть вертикальной, то есть от матери к ребенку, или горизонтальной, то есть от человека к человеку. Примеры вертикальной передачи включают вирус гепатита В и ВИЧ, когда ребенок рождается уже инфицированным вирусом. [116] Другим, более редким примером является вирус ветряной оспы , который, хотя и вызывает относительно легкие инфекции у детей и взрослых, может быть смертельным для плода и новорожденного ребенка. [117]
Горизонтальная передача является наиболее распространенным механизмом распространения вирусов среди населения. [118] Горизонтальная передача может произойти при обмене биологическими жидкостями во время сексуальной активности, при обмене слюной или при употреблении зараженной пищи или воды. Это также может произойти при аэрозолей, вдыхании содержащих вирусы, или от насекомых -переносчиков, например, когда инфицированные комары проникают через кожу хозяина. [118] Большинство типов вирусов ограничены только одним или двумя из этих механизмов, и их называют «респираторными вирусами» или «кишечными вирусами» и так далее. Уровень или скорость передачи вирусных инфекций зависит от таких факторов, как плотность населения, количество восприимчивых людей (т. е. тех, у кого нет иммунитета), [119] качество здравоохранения и погода. [120]
Эпидемиология используется для разрыва цепочки заражения населения во время вспышек вирусных заболеваний . [13] : 264 Применяются меры контроля, основанные на знаниях о том, как передается вирус. Важно найти источник или источники вспышки и идентифицировать вирус. После того как вирус идентифицирован, цепочку передачи иногда можно разорвать с помощью вакцин. Когда вакцины недоступны, санитария и дезинфекция могут быть эффективными. Часто инфицированных людей изолируют от остального сообщества, а тех, кто подвергся воздействию вируса, помещают в карантин . [13] : 894 Для борьбы со вспышкой ящура среди крупного рогатого скота в Великобритании в 2001 году были забиты тысячи голов крупного рогатого скота. [121] Большинство вирусных инфекций человека и других животных имеют инкубационный период , в течение которого инфекция не вызывает никаких признаков или симптомов. [13] : 170 Инкубационный период вирусных заболеваний составляет от нескольких дней до недель, но известен для большинства инфекций. [13] : 170–172 Несколько дублируя, но в основном следующий за инкубационным периодом, существует период коммуникативности — время, когда инфицированный человек или животное заразен и может заразить другого человека или животное. [13] : 170–172 Это также известно для многих вирусных инфекций, и знание продолжительности обоих периодов важно для борьбы со вспышками. [13] : 272 Когда вспышки вызывают необычно высокий процент случаев среди населения, сообщества или региона, их называют эпидемиями. Если вспышки распространяются по всему миру, их называют пандемиями . [13] : 891
Эпидемии и пандемии
Пандемия – это всемирная эпидемия . Пандемия гриппа 1918 года , продолжавшаяся до 1919 года, представляла собой пандемию гриппа 5-й категории , вызванную необычайно тяжелым и смертоносным вирусом гриппа А. Жертвами часто становились здоровые молодые люди, в отличие от большинства вспышек гриппа, от которых преимущественно страдают несовершеннолетние, пожилые или иным образом ослабленные пациенты. [26] : 409–415 По более ранним оценкам, от него погибло 40–50 миллионов человек. [122] в то время как более поздние исследования показывают, что в 1918 году от него могло погибнуть до 100 миллионов человек, или 5% населения мира. [123]
Хотя вирусные пандемии случаются редко, ВИЧ, который развился из вирусов, обнаруженных у обезьян и шимпанзе, является пандемическим, по крайней мере, с 1980-х годов. [124] В течение ХХ века произошло четыре пандемии, вызванные вирусом гриппа, причем те, которые произошли в 1918, 1957 и 1968 годах, были тяжелыми. [125] Большинство исследователей полагают, что ВИЧ зародился в странах Африки к югу от Сахары в 20 веке; [126] сейчас это пандемия: по оценкам, во всем мире с этой болезнью живут 37,9 миллиона человек. [127] В 2018 году от СПИДа умерло около 770 000 человек. [128] По оценкам Объединенной программы Организации Объединенных Наций по ВИЧ/СПИДу (ЮНЭЙДС) и Всемирной организации здравоохранения (ВОЗ), СПИД унес жизни более 25 миллионов человек с момента его первого признания 5 июня 1981 года, что делает его одной из самых разрушительных эпидемий за всю историю человечества. история. [129] В 2007 году было зарегистрировано 2,7 миллиона новых ВИЧ-инфекций и 2 миллиона смертей, связанных с ВИЧ. [130]


Несколько высоколетальных вирусных патогенов относятся к семейству Filoviridae . Филовирусы представляют собой нитевидные вирусы, вызывающие вирусную геморрагическую лихорадку , и включают эболавирусы и марбургвирусы . Вирус Марбург , впервые обнаруженный в 1967 году, привлек широкое внимание прессы в апреле 2005 года из-за вспышки в Анголе . [131] Болезнь, вызванная вирусом Эбола, также вызывала периодические вспышки с высоким уровнем смертности с 1976 года, когда она была впервые выявлена. Самая страшная и последняя из них — эпидемия в Западной Африке в 2013–2016 годах . [132]
За исключением оспы, большинство пандемий вызваны недавно появившимися вирусами. Эти «новые» вирусы обычно представляют собой мутанты менее вредных вирусов, которые ранее циркулировали у людей или других животных. [133]
Тяжелый острый респираторный синдром ( SARS ) и ближневосточный респираторный синдром (MERS) вызываются новыми типами коронавирусов . Известно, что другие коронавирусы вызывают легкие инфекции у людей. [134] поэтому вирулентность и быстрое распространение инфекций атипичной пневмонии, которые к июлю 2003 года стали причиной около 8000 случаев заболевания и 800 смертей, были неожиданными, и большинство стран не были к этому готовы. [135]
Родственный коронавирус, коронавирус тяжелого острого респираторного синдрома 2 (SARS-Cov-2) , предположительно возникший у летучих мышей, появился в Ухане , Китай, в ноябре 2019 года и быстро распространился по всему миру. Заражение вирусом вызвало пандемию COVID-19 , начавшуюся в 2020 году. [93] [136] [137] Беспрецедентные в мирное время ограничения были введены на международные поездки. [138] комендантский час . в ответ на пандемию в нескольких крупных городах мира был введен [139]
Рак
Вирусы являются установленной причиной рака у людей и других биологических видов. Вирусный рак встречается только у меньшинства инфицированных людей (или животных). Раковые вирусы происходят из целого ряда семейств вирусов, включая как РНК-, так и ДНК-вирусы, поэтому не существует единого типа « онковируса » (устаревший термин, первоначально использовавшийся для острой трансформации ретровирусов). Развитие рака определяется множеством факторов, таких как иммунитет хозяина. [140] и мутации в организме хозяина. [141] Вирусы, вызывающие рак человека, включают некоторые генотипы вируса папилломы человека , вируса гепатита В , вируса гепатита С , вируса Эпштейна-Барра , герпесвируса , ассоциированного с саркомой Капоши, и Т-лимфотропного вируса человека . Последний обнаруженный вирус рака человека – это полиомавирус ( полиомавирус клеток Меркеля ), который вызывает большинство случаев редкой формы рака кожи, называемой карциномой клеток Меркеля . [142] Вирусы гепатита могут перерасти в хроническую вирусную инфекцию, приводящую к раку печени . [143] [144] Заражение Т-лимфотропным вирусом человека может привести к тропическому спастическому парапарезу и Т-клеточному лейкозу взрослых . [145] Вирусы папилломы человека являются признанной причиной рака шейки матки , кожи, ануса и полового члена . [146] Внутри Herpesviridae герпесвирус , ассоциированный с саркомой Капоши, вызывает саркому Капоши и лимфому полостей тела , а вирус Эпштейна-Барра вызывает лимфому Беркитта , лимфому Ходжкина , B- лимфопролиферативное заболевание и карциному носоглотки . [147] Полиомавирус клеток Меркеля, тесно родственный SV40 , и полиомавирусы мышей, которые уже более 50 лет используются в качестве животных моделей вирусов рака. [148]
Защитные механизмы хозяина
Первой линией защиты организма от вирусов является врожденная иммунная система . Сюда входят клетки и другие механизмы, которые неспецифически защищают хозяина от инфекции. Это означает, что клетки врожденной системы распознают патогены и реагируют на них общим образом, но, в отличие от адаптивной иммунной системы , она не обеспечивает хозяину долговременного или защитного иммунитета. [149]
РНК-интерференция является важной врожденной защитой от вирусов. [150] Многие вирусы имеют стратегию репликации, включающую двухцепочечную РНК (дцРНК). Когда такой вирус заражает клетку, он высвобождает свою молекулу или молекулы РНК, которые немедленно связываются с белковым комплексом, называемым дайсером , который разрезает РНК на более мелкие кусочки. Активируется биохимический путь — комплекс RISC , который обеспечивает выживание клеток за счет разрушения вирусной мРНК. Ротавирусы эволюционировали, чтобы избежать этого защитного механизма, не снимая полностью покрытие внутри клетки и высвобождая вновь произведенную мРНК через поры во внутреннем капсиде частицы. Их геномная дцРНК остается защищенной внутри ядра вириона. [151] [152]
Когда адаптивная иммунная система позвоночного сталкивается с вирусом, она вырабатывает специфические антитела , которые связываются с вирусом и часто делают его неинфекционным. Это называется гуморальным иммунитетом . Важны два типа антител. Первый, называемый IgM , очень эффективен при нейтрализации вирусов, но вырабатывается клетками иммунной системы лишь в течение нескольких недель. Второй, называемый IgG , вырабатывается неопределенно долго. Присутствие IgM в крови хозяина используется для проверки острой инфекции, тогда как IgG указывает на инфекцию когда-то в прошлом. [153] Антитела IgG измеряются при тестов на иммунитет . проведении [154]
Антитела могут продолжать оставаться эффективным защитным механизмом даже после того, как вирусам удалось проникнуть в клетку-хозяина. Белок, находящийся в клетках, называемый TRIM21 , может прикрепляться к антителам на поверхности вирусной частицы. системы клетки Это запускает последующее разрушение вируса ферментами протеосомной . [155]
Вторая защита позвоночных животных от вирусов называется клеточным иммунитетом и включает в себя иммунные клетки, известные как Т-клетки . Клетки организма постоянно отображают короткие фрагменты своих белков на поверхности клетки, и, если Т-клетка распознает там подозрительный вирусный фрагмент, клетка-хозяин разрушается «Т-киллерами» и вирус-специфичные Т-клетки размножаются. Такие клетки, как макрофаги, являются специалистами в презентации этого антигена . [156] Производство интерферона является важным защитным механизмом хозяина. Это гормон, вырабатываемый организмом при наличии вирусов. Его роль в иммунитете сложна; в конечном итоге он останавливает размножение вирусов, убивая инфицированную клетку и ее ближайших соседей. [157]
Не все вирусные инфекции вызывают такой защитный иммунный ответ. ВИЧ уклоняется от иммунной системы, постоянно изменяя аминокислотную последовательность белков на поверхности вириона. Это известно как «мутация ускользания», поскольку вирусные эпитопы ускользают от распознавания иммунным ответом хозяина. Эти стойкие вирусы уклоняются от иммунного контроля путем секвестрации, блокады представления антигена , устойчивости к цитокинам , уклонения от активности естественных клеток-киллеров , ухода от апоптоза и антигенного сдвига . [158] Другие вирусы, называемые « нейротропными вирусами », распространяются путем нейронного распространения, когда иммунная система может быть не в состоянии достичь их из-за иммунных привилегий . [159]
Профилактика и лечение
Поскольку для репликации вирусы используют жизненно важные метаболические пути внутри клеток-хозяев, их трудно уничтожить без использования лекарств, которые оказывают токсическое воздействие на клетки-хозяева в целом. Наиболее эффективными медицинскими подходами к вирусным заболеваниям являются вакцинация для обеспечения иммунитета к инфекции и противовирусные препараты, избирательно препятствующие репликации вируса.
Вакцина
Вакцинация – дешевый и эффективный способ предотвращения заражения вирусами. Вакцины использовались для предотвращения вирусных инфекций задолго до открытия самих вирусов. Их использование привело к резкому снижению заболеваемости (заболеваний) и смертности (смертности), связанных с вирусными инфекциями, такими как полиомиелит , корь , эпидемический паротит и краснуха . [160] Инфекция оспы ликвидирована. [161] Доступны вакцины для предотвращения более тринадцати вирусных инфекций человека. [162] и другие используются для профилактики вирусных инфекций животных. [163] Вакцины могут состоять из живых ослабленных или убитых вирусов, вирусных белков ( антигенов ) или РНК . [164] [165] Живые вакцины содержат ослабленные формы вируса, которые не вызывают заболевания, но тем не менее придают иммунитет. Такие вирусы называются аттенуированными. Живые вакцины могут быть опасны при введении людям со слабым иммунитетом (которых называют людьми с ослабленным иммунитетом ), поскольку у этих людей ослабленный вирус может вызвать исходное заболевание. [166] Для производства субъединичных вакцин используются методы биотехнологии и генной инженерии. В этих вакцинах используются только капсидные белки вируса. Вакцина против гепатита B является примером вакцины этого типа. [167] Субъединичные вакцины безопасны для пациентов с ослабленным иммунитетом , поскольку они не могут вызвать заболевание. [168] Вакцина против вируса желтой лихорадки , живой аттенуированный штамм под названием 17D, вероятно, является самой безопасной и эффективной вакциной, когда-либо созданной. [169]
Противовирусные препараты
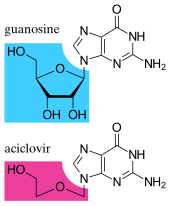
Противовирусные препараты часто представляют собой аналоги нуклеозидов (поддельные строительные блоки ДНК), которые вирусы ошибочно включают в свои геномы во время репликации. [170] Жизненный цикл вируса затем останавливается, поскольку вновь синтезированная ДНК становится неактивной. Это связано с тем, что в этих аналогах отсутствуют гидроксильные группы, которые вместе с атомами фосфора соединяются вместе, образуя прочный «остов» молекулы ДНК. Это называется терминацией цепи ДНК . [171] Примерами аналогов нуклеозидов являются ацикловир при инфекциях, вызванных вирусом простого герпеса, и ламивудин при инфекциях, вызванных вирусом ВИЧ и гепатита В. Ацикловир – один из старейших и наиболее часто назначаемых противовирусных препаратов. [172] Другие используемые противовирусные препараты нацелены на разные стадии жизненного цикла вируса. ВИЧ зависит от протеолитического фермента, называемого протеазой ВИЧ-1 Чтобы стать полностью заразным, . Существует большой класс препаратов, называемых ингибиторами протеазы , которые инактивируют этот фермент. [173] Существует около тринадцати классов противовирусных препаратов, каждый из которых нацелен на разные вирусы или стадии репликации вируса. [170]
Гепатит С вызывается РНК-вирусом. У 80% инфицированных заболевание носит хронический характер, и без лечения они остаются инфицированными на всю оставшуюся жизнь. Существуют эффективные методы лечения, в которых используются противовирусные препараты прямого действия . [174] Лечение хронических носителей вируса гепатита В также было разработано с использованием аналогичных стратегий, включающих ламивудин и другие противовирусные препараты. [175]
Инфекция у других видов
Вирусы заражают всю клеточную жизнь, и, хотя вирусы встречаются повсеместно, каждый вид клеток имеет свой собственный определенный диапазон, который часто заражает только этот вид. [6] : 3 Некоторые вирусы, называемые сателлитами , могут размножаться только внутри клеток, уже зараженных другим вирусом. [30]
Вирусы животных
Вирусы являются важными патогенами домашнего скота. Такие заболевания, как ящур и блютанг , вызываются вирусами. [176] Животные-компаньоны, такие как кошки, собаки и лошади, если не вакцинированы, восприимчивы к серьезным вирусным инфекциям. Собачий парвовирус вызывается небольшим ДНК-вирусом, и инфекции часто приводят к летальному исходу у щенков. [177] Как и все беспозвоночные , медоносная пчела восприимчива ко многим вирусным инфекциям. [178] Большинство вирусов безвредно сосуществуют со своим хозяином и не вызывают никаких признаков или симптомов заболевания. [6] : 4
Вирусы растений

Существует много типов вирусов растений, но зачастую они вызывают лишь потерю урожая , и бороться с ними экономически нецелесообразно. Вирусы растений часто передаются от растения к растению организмами, известными как переносчики . Обычно это насекомые, но переносчиками являются некоторые грибы, нематоды , одноклеточные организмы и растения-паразиты. [179] Когда борьба с вирусными инфекциями растений считается экономичной (например, в отношении многолетних фруктов), усилия концентрируются на уничтожении переносчиков и удалении альтернативных хозяев, таких как сорняки. [13] : 802 Вирусы растений не могут заразить человека и других животных, поскольку могут размножаться только в живых растительных клетках. [13] : 799–807
Родом из Перу, картофель стал основной культурой во всем мире. [180] Картофельный вирус Y вызывает заболевание картофеля и родственных ему видов, включая томаты и перец. В 1980-х годах этот вирус приобрел экономическое значение, когда оказалось, что с ним трудно бороться в посевах семенного картофеля. Передаваемый тлей , этот вирус может снизить урожайность сельскохозяйственных культур до 80 процентов, вызывая значительные потери урожая картофеля. [181]
Растения обладают сложными и эффективными механизмами защиты от вирусов. Одним из наиболее эффективных является наличие так называемых генов устойчивости (R). Каждый ген R придает устойчивость к определенному вирусу, вызывая локализованные области гибели клеток вокруг инфицированной клетки, которые часто можно увидеть невооруженным глазом в виде больших пятен. Это останавливает распространение инфекции. [182] РНК-интерференция также является эффективной защитой растений. [13] : 809 > При заражении растения часто производят естественные дезинфицирующие средства, убивающие вирусы, такие как салициловая кислота , оксид азота и активные молекулы кислорода . [183]
Частицы растительного вируса или вирусоподобные частицы (VLP) находят применение как в биотехнологии, так и в нанотехнологиях . Капсиды большинства растительных вирусов представляют собой простые и надежные структуры и могут производиться в больших количествах либо путем заражения растений, либо путем экспрессии в различных гетерологичных системах. Частицы растительного вируса могут быть модифицированы генетически и химически для инкапсулирования инородного материала и могут быть включены в супрамолекулярные структуры для использования в биотехнологии. [184]
Бактериальные вирусы

Бактериофаги представляют собой распространенную и разнообразную группу вирусов и являются наиболее распространенным биологическим объектом в водной среде: в океанах этих вирусов до десяти раз больше, чем бактерий. [185] достижение уровня 250 000 000 бактериофагов на миллилитр морской воды. [186] Эти вирусы заражают определенные бактерии, связываясь с поверхностными молекулами рецепторов , а затем проникая в клетку. За короткий промежуток времени, в некоторых случаях всего за несколько минут, бактериальная полимераза начинает транслировать вирусную мРНК в белок. Эти белки впоследствии становятся либо новыми вирионами внутри клетки, либо белками-помощниками, которые помогают сборке новых вирионов, либо белками, участвующими в лизисе клеток. Вирусные ферменты способствуют разрушению клеточной мембраны, и, в случае фага Т4 , всего за двадцать минут после инъекции может быть высвобождено более трехсот фагов. [13] : 834–835
Основным способом защиты бактерий от бактериофагов является выработка ферментов, разрушающих чужеродную ДНК. Эти ферменты, называемые эндонуклеазами рестрикции , разрезают вирусную ДНК, которую бактериофаги вводят в бактериальные клетки. [187] Бактерии также содержат систему, которая использует последовательности CRISPR для сохранения фрагментов геномов вирусов, с которыми бактерии ранее контактировали, что позволяет им блокировать репликацию вируса посредством формы РНК-интерференции . [188] [189] Эта генетическая система обеспечивает бактериям приобретенный иммунитет к инфекции. [190]
Некоторые бактериофаги называются « умеренными », поскольку они вызывают латентные инфекции и не сразу уничтожают клетки-хозяева. Вместо этого их ДНК включается в клетку-хозяина в виде профага . Эти скрытые инфекции становятся продуктивными, когда ДНК профагов активируется такими стимулами, как изменения в окружающей среде. [191] В кишечнике животных, в том числе человека, содержатся умеренные бактериофаги, которые активируются различными раздражителями, включая изменения в диете и антибиотики. [192] Хотя впервые это было обнаружено у бактериофагов, известно, что многие другие вирусы образуют провирусы , включая ВИЧ. [191] [193]
Архейные вирусы
Некоторые вирусы размножаются внутри архей : это ДНК-вирусы необычной, а иногда и уникальной формы. [4] [55] Наиболее подробно эти вирусы изучены на термофильных археях, в частности на порядках Sulfolobales и Thermoproteales . [194] Защита от этих вирусов включает вмешательство РНК из повторяющихся последовательностей ДНК в геномах архей, которые связаны с генами вирусов. [195] [196] Большинство архей имеют системы CRISPR-Cas в качестве адаптивной защиты от вирусов. Это позволяет археям сохранять участки вирусной ДНК, которые затем используются для нацеливания и устранения последующих заражений вирусом, используя процесс, аналогичный РНК-интерференции. [197]
Роль в водных экосистемах
Вирусы являются наиболее распространенным биологическим объектом в водной среде. [2] В чайной ложке морской воды их около десяти миллионов. [198] Большинство этих вирусов представляют собой бактериофаги, инфицирующие гетеротрофные бактерии, и цианофаги, инфицирующие цианобактерии, и они необходимы для регуляции морских и пресноводных экосистем. [199] Бактериофаги безвредны для растений и животных и необходимы для регуляции морских и пресноводных экосистем. [200] являются важными агентами смертности фитопланктона , основы пищевой цепи в водной среде. [201] Они заражают и уничтожают бактерии в водных микробных сообществах и являются одним из наиболее важных механизмов переработки углерода и круговорота питательных веществ в морской среде. Органические молекулы, высвобождаемые из мертвых бактериальных клеток, стимулируют рост новых бактерий и водорослей в процессе, известном как вирусный шунт . [202] В частности, было показано, что лизис бактерий вирусами усиливает круговорот азота и стимулирует рост фитопланктона. [203] Вирусная активность может также влиять на биологический насос — процесс, посредством которого в глубинах углерод улавливается океана. [204]
Микроорганизмы составляют более 90% биомассы моря. По оценкам, вирусы убивают примерно 20% этой биомассы каждый день, а в океанах вирусов в 10–15 раз больше, чем бактерий и архей. [205] Вирусы также являются основными агентами, ответственными за разрушение фитопланктона , включая вредоносное цветение водорослей . [206] Число вирусов в океанах уменьшается по мере удаления от берега и глубже в воду, где меньше организмов-хозяев. [204]
В январе 2018 года ученые сообщили, что 800 миллионов вирусов, преимущественно морского происхождения, ежедневно оседают из Земли . атмосферы на каждый квадратный метр поверхности планеты в результате глобального атмосферного потока вирусов, циркулирующих над погодной системой но ниже высоты обычного авиаперелета, распространяя вирусы по планете. [207] [208]
Как любой организм, морские млекопитающие восприимчивы к вирусным инфекциям. В 1988 и 2002 годах тысячи тюленей погибли в Европе от вируса чумы чумы . [209] Многие другие вирусы, в том числе калицивирусы , герпесвирусы , аденовирусы и парвовирусы , циркулируют в популяциях морских млекопитающих. [204]
В декабре 2022 года ученые сообщили о первом наблюдении вироворы в ходе эксперимента с прудовой водой, содержащей хлорвирус , который обычно заражает зеленые водоросли в пресноводной среде. Когда все другие источники микробной пищи были удалены из воды, инфузорий Halteria наблюдалось увеличение численности из-за активного потребления хлоровируса в качестве источника пищи вместо типичного для них бактериоядного рациона. [210] [211]
Роль в эволюции
Вирусы являются важным естественным средством передачи генов между различными видами, что увеличивает генетическое разнообразие и стимулирует эволюцию. [9] [212] [213] Считается, что вирусы играли центральную роль в ранней эволюции, до того, как последний универсальный предок превратился в бактерии, археи и эукариоты. [214] Вирусы по-прежнему являются одним из крупнейших резервуаров неизведанного генетического разнообразия на Земле. [204]
Приложения
Науки о жизни и медицина

Вирусы важны для изучения молекулярной и клеточной биологии , поскольку они представляют собой простые системы, которые можно использовать для манипулирования клетками и исследования их функций. [26] : 8 Изучение и использование вирусов предоставили ценную информацию об аспектах клеточной биологии. [215] Например, вирусы были полезны при изучении генетики и помогли нам понять основные механизмы молекулярной генетики , такие как репликация ДНК , транскрипция , процессинг РНК , трансляция , транспорт белков и иммунология .
Генетики часто используют вирусы в качестве векторов для введения генов в изучаемые клетки. Это полезно для того, чтобы заставить клетку производить чужеродное вещество или изучить эффект от введения нового гена в геном. Точно так же виротерапия использует вирусы в качестве векторов для лечения различных заболеваний, поскольку они могут специфически воздействовать на клетки и ДНК. Он показывает многообещающее применение в лечении рака и в генной терапии . Восточноевропейские ученые уже некоторое время используют фаготерапию в качестве альтернативы антибиотикам, и интерес к этому подходу растет из-за высокого уровня устойчивости к антибиотикам, обнаруженного в настоящее время у некоторых патогенных бактерий. [216] Экспрессия гетерологичных белков вирусами лежит в основе нескольких производственных процессов, которые в настоящее время используются для производства различных белков, таких как вакцинные антигены и антитела. Недавно были разработаны промышленные процессы с использованием вирусных векторов, а несколько фармацевтических белков в настоящее время проходят доклинические и клинические испытания. [217]
Виротерапия
Виротерапия предполагает использование генетически модифицированных вирусов для лечения заболеваний. [218] Ученые модифицировали вирусы, чтобы они размножались в раковых клетках и уничтожали их, но не заражали здоровые клетки. Talimogene laherparepvec (T-VEC), например, представляет собой модифицированный вирус простого герпеса , в котором ген, необходимый вирусам для репликации в здоровых клетках, удален и заменен человеческим геном ( GM-CSF ), который стимулирует иммунитет. Когда этот вирус заражает раковые клетки, он уничтожает их, и при этом наличие гена GM-CSF привлекает дендритные клетки из окружающих тканей организма. Дендритные клетки перерабатывают мертвые раковые клетки и передают их компоненты другим клеткам иммунной системы . [219] После успешных клинических испытаний вирус получил одобрение для лечения меланомы в конце 2015 года. [220] Вирусы, перепрограммированные для уничтожения раковых клеток, называются онколитическими вирусами . [221]
Материаловедение и нанотехнологии
С точки зрения материаловеда, вирусы можно рассматривать как органические наночастицы . [222] На их поверхности имеются специальные инструменты, которые позволяют им преодолевать барьеры клеток-хозяев. Точно определены размеры и форма вирусов, количество и характер функциональных групп на их поверхности. Таким образом, вирусы обычно используются в материаловедении в качестве каркасов для ковалентно связанных модификаций поверхности. Особое качество вирусов заключается в том, что их можно адаптировать путем направленной эволюции. Мощные методы, разработанные науками о жизни, становятся основой инженерных подходов к наноматериалам, открывая широкий спектр приложений, выходящих далеко за рамки биологии и медицины. [223] Благодаря своему размеру, форме и четко определенной химической структуре вирусы использовались в качестве шаблонов для организации материалов на наноуровне. Примеры включают работу в Военно-морской исследовательской лаборатории в Вашингтоне, округ Колумбия, по использованию частиц вируса мозаики вигны (CPMV) для усиления сигналов в датчиках на основе микрочипов ДНК . В этом приложении вирусные частицы отделяют флуоресцентные красители, используемые для передачи сигналов, чтобы предотвратить образование нефлуоресцентных димеров , которые действуют как тушители . [224] Другим примером является использование CPMV в качестве наноразмерного макета для молекулярной электроники. [225]
Синтетические вирусы
Многие вирусы могут быть синтезированы de novo («с нуля»). Первый синтетический вирус был создан в 2002 году. [226] Хотя это некоторое заблуждение, синтезируется не сам вирус, а скорее его ДНК-геном (в случае ДНК-вируса) или кДНК- копия его генома (в случае РНК-вирусов). Для многих семейств вирусов обнаженная синтетическая ДНК или РНК (после ферментативного преобразования обратно из синтетической кДНК) является заразной при введении в клетку. То есть они содержат всю необходимую информацию для создания новых вирусов. Эта технология сейчас используется для исследования новых стратегий вакцинации. [227] Способность синтезировать вирусы имеет далеко идущие последствия, поскольку вирусы больше нельзя считать вымершими, пока известна информация о последовательности их генома и пермиссивные доступны клетки. По состоянию на июнь 2021 года полноразмерные последовательности генома 11 464 различных вирусов, включая оспу, общедоступны в онлайн-базе данных, поддерживаемой Национальными институтами здравоохранения . [228]
Оружие
Способность вирусов вызывать разрушительные эпидемии в человеческом обществе привела к опасениям, что вирусы могут быть использованы в качестве оружия для биологической войны . Дополнительную обеспокоенность вызвало успешное воссоздание гриппа 1918 года . в лаборатории печально известного вируса [229] Вирус оспы опустошал множество обществ на протяжении всей истории, прежде чем был искоренен. В мире есть только два центра, уполномоченных ВОЗ хранить запасы вируса оспы: Государственный научный центр вирусологии и биотехнологии «ВЕКТОР» в России и Центры по контролю и профилактике заболеваний в США. [230] Его можно использовать как оружие, [230] поскольку вакцина от оспы иногда вызывала серьезные побочные эффекты, она больше не используется на регулярной основе ни в одной стране. Таким образом, большая часть современного человечества почти не имеет устойчивой устойчивости к оспе и будет уязвима для этого вируса. [230]
См. также
- Межвидовая передача
- Глоссарий вирусологии
- Закон снижения вирулентности – Опровергнутая гипотеза эпидемиолога Теобальда Смита
- Неклеточная жизнь
- Агент переноса генов
- Ретрозим
- Наименьшие организмы
- Теория вирулентности - теория биолога Пола В. Эвальда.
- Вирусная метагеномика
- Вироплазма
- Зооноз
Ссылки
- ^ Ву К.Дж. (15 апреля 2020 г.). «Во Вселенной больше вирусов, чем звезд. Почему нас заражают лишь некоторые из них? – На Земле существует более квадриллиона квадриллионов отдельных вирусов, но большинство из них не готовы проникнуть в человека. Сможем ли мы найти те, которые есть?» . Национальное географическое общество . Архивировано из оригинала 15 апреля 2020 года . Проверено 18 мая 2020 г.
- ^ Jump up to: а б с Кунин Е.В., Сенкевич Т.Г., Доля В.В. (сентябрь 2006 г.). «Древний мир вирусов и эволюция клеток» . Биология Директ . 1 (1): 29. дои : 10.1186/1745-6150-1-29 . ПМЦ 1594570 . ПМИД 16984643 .
- ^ Циммер С (26 февраля 2021 г.). «Тайная жизнь коронавируса. Маслянистый пузырь генов шириной 100 нанометров убил более двух миллионов человек и изменил мир. Ученые не совсем знают, что с этим делать» . Нью-Йорк Таймс . Архивировано из оригинала 28 декабря 2021 года . Проверено 28 февраля 2021 г.
- ^ Jump up to: а б Лоуренс CM, Менон С., Эйлерс Б.Дж., Ботнер Б., Хаят Р., Дуглас Т. и др. (май 2009 г.). «Структурно-функциональные исследования архейных вирусов» . Журнал биологической химии . 284 (19): 12599–603. дои : 10.1074/jbc.R800078200 . ПМК 2675988 . ПМИД 19158076 .
- ^ Эдвардс Р.А., Ровер Ф. (июнь 2005 г.). «Вирусная метагеномика». Обзоры природы. Микробиология . 3 (6): 504–10. дои : 10.1038/nrmicro1163 . ПМИД 15886693 . S2CID 8059643 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Диммок, Нью-Джерси, Истон А.Дж., Леппард К. (2007). Введение в современную вирусологию (6-е изд.). Издательство Блэквелл. ISBN 978-1-4051-3645-7 .
- ^ Jump up to: а б с «Таксономия вирусов: выпуск 2022 г.» . talk.ictvonline.org . Международный комитет по таксономии вирусов . Проверено 27 апреля 2023 г.
- ^ Jump up to: а б Брейтбарт М. , Ровер Ф. (июнь 2005 г.). «Тут вирус, там вирус, везде один и тот же вирус?». Тенденции в микробиологии . 13 (6): 278–84. дои : 10.1016/j.tim.2005.04.003 . ПМИД 15936660 .
- ^ Jump up to: а б Канчая С., Фурнус Г., Чибани-Шеннуфи С., Диллманн М.Л., Брюссов Х. (август 2003 г.). «Фаг как агент латерального переноса генов». Современное мнение в микробиологии . 6 (4): 417–24. дои : 10.1016/S1369-5274(03)00086-9 . ПМИД 12941415 .
- ^ Jump up to: а б Рыбицкий Е.П. (1990). «Классификация организмов на пороге жизни, или Проблемы вирусной систематики». Южноафриканский научный журнал . 86 : 182–86.
- ^ Jump up to: а б Кунин Е.В., Старокадомский П. (октябрь 2016 г.). «Живы ли вирусы? Парадигма репликатора проливает решающий свет на старый, но ошибочный вопрос» . Исследования по истории и философии биологических и биомедицинских наук . 59 : 125–34. дои : 10.1016/j.shpsc.2016.02.016 . ПМК 5406846 . ПМИД 26965225 .
- ^ Робилотти Э., Дерезинский С., Пинский Б.А. (январь 2015 г.). «Норовирус» . Обзоры клинической микробиологии . 28 (1): 134–64. дои : 10.1128/CMR.00075-14 . ПМЦ 4284304 . ПМИД 25567225 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Шорс Т (2017). Понимание вирусов . Издательство Джонс и Бартлетт. ISBN 978-1-284-02592-7 .
- ^ «Вирус, н.». ОЭД онлайн . Издательство Оксфордского университета. Март 2015.
- ^ Jump up to: а б Харпер Д. (2011). "вирус" . Интернет-словарь этимологии . Проверено 19 декабря 2014 г.
- ^ «Яростный, прил.». ОЭД онлайн . Издательство Оксфордского университета. Март 2015.
- ^ Харпер Д. (2011). «вирулентный» . Интернет-словарь этимологии . Проверено 19 декабря 2014 г.
- ^ Бушард К., Тон Р. (2003). «Модели животных с диабетом». В Хау Дж., Ван Хузьер-младший Г.Л. (ред.). Справочник по лабораторным зоотехникам . Модели животных. Том. II (2-е изд.). ЦРК Пресс. стр. 163, 166.
- ^ Уильям Т. Стерн: Ботаническая латынь. История, грамматика, синтаксис, терминология и словарный запас. Дэвид и Чарльз, 3-е изд., 1983. Цитата: «Вирус: вирус (SN II), gen. Sing. Viri, nom. pl. vira, gen. pl. vīrorum (отличать от virorum , мужчин)».
- ^ Харпер Д. (2011). "популярный" . Интернет-словарь этимологии . Проверено 19 декабря 2014 г.
- ^ Харпер Д. (2011). «вирион» . Интернет-словарь этимологии . Проверено 19 декабря 2014 г.
- ^ Касьенс С (2010). Махи Б.В., Ван Регенмортель М.Х. (ред.). Настольная энциклопедия общей вирусологии . Бостон: Академическая пресса. п. 167. ИСБН 978-0-12-375146-1 .
- ^ Айер Л.М., Баладжи С., Кунин Е.В., Аравинд Л. (апрель 2006 г.). «Эволюционная геномика ядерно-цитоплазматических крупных ДНК-вирусов» . Вирусные исследования . 117 (1): 156–84. doi : 10.1016/j.virusres.2006.01.009 . ПМИД 16494962 .
- ^ Jump up to: а б Санхуан Р., Небот М.Р., Кирико Н., Мански Л.М., Белшоу Р. (октябрь 2010 г.). «Темпы вирусных мутаций» . Журнал вирусологии . 84 (19): 9733–9748. дои : 10.1128/JVI.00694-10 . ПМЦ 2937809 . ПМИД 20660197 .
- ^ Крупович М., Доля В.В., Кунин Е.В. (июль 2019 г.). «Происхождение вирусов: первичные репликаторы, рекрутирующие капсиды от хозяев» (PDF) . Обзоры природы. Микробиология . 17 (7): 449–58. дои : 10.1038/s41579-019-0205-6 . ПМИД 31142823 . S2CID 256744818 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в Коллиер Л., Балоуз А., Сассман М. (1998). Мэхи Б., Коллиер Л.А. (ред.). Микробиология и микробные инфекции Топли и Уилсона . Вирусология. Том. 1 (9-е изд.). ISBN 0-340-66316-2 .
- ^ Jump up to: а б с д и ж г час Мэхи У.Дж., Регенмортель М.Х., ред. (2009). Настольная энциклопедия общей вирусологии . Оксфорд: Академическая пресса. ISBN 978-0-12-375146-1 .
- ^ МакКлинток Б. (июнь 1950 г.). «Происхождение и поведение мутабельных локусов кукурузы» . Труды Национальной академии наук Соединенных Штатов Америки . 36 (6): 344–55. Бибкод : 1950ПНАС...36..344М . дои : 10.1073/pnas.36.6.344 . ПМЦ 1063197 . ПМИД 15430309 .
- ^ Цагрис Э.М., Мартинес де Альба А.Е., Гозманова М., Калантидис К. (ноябрь 2008 г.). «Вироиды» . Клеточная микробиология . 10 (11): 2168–79. дои : 10.1111/j.1462-5822.2008.01231.x . ПМИД 18764915 . S2CID 221581424 .
- ^ Jump up to: а б Ла Скола Б., Десню С., Панье И., Робер С., Баррасси Л., Фурнус Г. и др. (сентябрь 2008 г.). «Вирофаг как уникальный паразит гигантского мимивируса». Природа . 455 (7209): 100–04. Бибкод : 2008Natur.455..100L . дои : 10.1038/nature07218 . ПМИД 18690211 . S2CID 4422249 .
- ^ Диммок, стр. 15–16.
- ^ Зербини FM, Китадзима EW (сентябрь 2022 г.). «От Contagium vivum Liquidum до Рибовирии: Вирус-ориентированная история таксономии вирусов табачной мозаики» . Биомолекулы . 12 (10): 1363. doi : 10.3390/biom12101363 . ПМК 9599303 . ПМИД 36291572 .
- ^ Холмс ЕС (октябрь 2007 г.). «Эволюция вирусов в эпоху генома» . ПЛОС Биология . 5 (10): е278. doi : 10.1371/journal.pbio.0050278 . ЧВК 1994994 . ПМИД 17914905 .
- ^ Виммер Э., Мюллер С., Тумпей Т.М., Таубенбергер Дж.К. (декабрь 2009 г.). «Синтетические вирусы: новая возможность понять и предотвратить вирусные заболевания» . Природная биотехнология . 27 (12): 1163–72. дои : 10.1038/nbt.1593 . ПМК 2819212 . ПМИД 20010599 .
- ^ Хорн М (2008). «Хламидии как симбионты у эукариот». Ежегодный обзор микробиологии . 62 : 113–31. дои : 10.1146/annurev.micro.62.081307.162818 . ПМИД 18473699 .
- ^ Аммерман, Северная Каролина, Бейер-Секстон М., Азад А.Ф. (ноябрь 2008 г.). «Лабораторное содержание Rickettsia rickettsii» . Современные протоколы в микробиологии . 11 (1): 3А.5.1–3А.5.21. дои : 10.1002/9780471729259.mc03a05s11 . ISBN 978-0471729259 . ПМЦ 2725428 . ПМИД 19016440 .
- ^ Фортер, П. Концепция вироцелл и микробиология окружающей среды. ISME J 7, 233–236 (2013). https://doi.org/10.1038/ismej.2012.110
- ^ ДеЛонг Дж.П., Аль-Саммак М.А., Аль-Амели З.Т., Дуниган Д.Д., Эдвардс К.Ф., Фурманн Дж.Дж. и др. (февраль 2022 г.). «К интегративному взгляду на фенотипы вирусов». Обзоры природы. Микробиология . 20 (2): 83–94. дои : 10.1038/s41579-021-00612-w . ПМИД 34522049 .
- ^ Краснер Р. (2014). Микробная проблема: взгляд на общественное здравоохранение . Берлингтон, Массачусетс: Jones & Bartlett Learning. ISBN 978-1-4496-7375-8 . OCLC 794228026 .
- ^ Киселев Н.А., Шерман М.Б., Цупрун В.Л. (1990). «Отрицательное окрашивание белков». Обзоры электронной микроскопии . 3 (1): 43–72. дои : 10.1016/0892-0354(90)90013-I . ПМИД 1715774 .
- ^ Каспар Д.Л., Клюг А. (1962). «Физические принципы построения обычных вирусов». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 27 : 1–24. дои : 10.1101/sqb.1962.027.001.005 . ПМИД 14019094 .
- ^ Крик Ф.Х., Уотсон JD (март 1956 г.). «Строение малых вирусов». Природа . 177 (4506): 473–75. Бибкод : 1956Natur.177..473C . дои : 10.1038/177473a0 . ПМИД 13309339 . S2CID 5740221 .
- ^ Фальво М.Р., Уошберн С., Суперфайн Р., Финч М., Брукс Ф.П., Чи В. и др. (март 1997 г.). «Манипуляции с отдельными вирусами: трение и механические свойства» . Биофизический журнал . 72 (3): 1396–403. Бибкод : 1997BpJ....72.1396F . дои : 10.1016/S0006-3495(97)78786-1 . ПМК 1184522 . ПМИД 9138585 .
- ^ Кузнецов Ю.Г., Малкин А.Дж., Лукас Р.В., Пломп М., Макферсон А. (сентябрь 2001 г.). «Изображение вирусов методом атомно-силовой микроскопии» . Журнал общей вирусологии . 82 (Часть 9): 2025–34. дои : 10.1099/0022-1317-82-9-2025 . ПМИД 11514711 .
- ^ Штраус С.К., Бо Х.Э. (2018). «Белки и сборка нитевидных бактериофагов». Вирусные белки и нуклеопротеиновые комплексы . Субклеточная биохимия. Том. 88. стр. 261–79. дои : 10.1007/978-981-10-8456-0_12 . ISBN 978-981-10-8455-3 . ПМИД 29900501 .
- ^ Уилсон Д.П. (2016). «Выступающие элементы вирусных капсидов сгруппированы на больших икосаэдрических кругах» . ПЛОС ОДИН . 11 (4): e0152319. Бибкод : 2016PLoSO..1152319W . дои : 10.1371/journal.pone.0152319 . ПМЦ 4821576 . ПМИД 27045511 .
- ^ Казенс С. (2009). Настольная энциклопедия общей вирусологии . Бостон: Академическая пресса. стр. 167–74. ISBN 978-0-12-375146-1 .
- ^ Дхама К., Хан С., Тивари Р., Сиркар С., Бхат С., Малик Ю.С. и др. (сентябрь 2020 г.). «Коронавирусная болезнь 2019-COVID-19» . Обзоры клинической микробиологии . 33 (4). дои : 10.1128/CMR.00028-20 . ПМЦ 7405836 . ПМИД 32580969 .
- ^ Россманн М.Г., Месянжинов В.В., Арисака Ф., Лейман П.Г. (апрель 2004 г.). «Машина для инъекции ДНК бактериофага Т4». Современное мнение в области структурной биологии . 14 (2): 171–80. дои : 10.1016/j.sbi.2004.02.001 . ПМИД 15093831 .
- ^ Лонг Г.В., Нобель Дж., Мерфи Ф.А., Херрманн К.Л., Лури Б. (сентябрь 1970 г.). «Опыт применения электронной микроскопии в дифференциальной диагностике оспы» . Прикладная микробиология . 20 (3): 497–504. дои : 10.1128/АЕМ.20.3.497-504.1970 . ПМК 376966 . ПМИД 4322005 .
- ^ Сюзан-Монти М., Ла Скола Б., Рауль Д. (апрель 2006 г.). «Геномные и эволюционные аспекты мимивируса». Вирусные исследования . 117 (1): 145–55. doi : 10.1016/j.virusres.2005.07.011 . ПМИД 16181700 .
- ^ Арслан Д., Лежандр М., Зельцер В., Абергель С., Клавери Дж. М. (октябрь 2011 г.). «Отдаленный родственник мимивируса с более крупным геномом подчеркивает фундаментальные особенности мегавирусов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (42): 17486–91. Бибкод : 2011PNAS..10817486A . дои : 10.1073/pnas.1110889108 . ПМК 3198346 . ПМИД 21987820 .
- ^ Jump up to: а б Филипп Н., Лежандр М., Дутр Дж., Куте Ю., Пуаро О., Леско М. и др. (июль 2013 г.). «Пандоравирусы: амеба-вирусы с геномами до 2,5 МБ, достигающими генома паразитических эукариот» (PDF) . Наука . 341 (6143): 281–86. Бибкод : 2013Sci...341..281P . дои : 10.1126/science.1239181 . ПМИД 23869018 . S2CID 16877147 .
- ^ Брандес Н., Линиал М. (апрель 2019 г.). «Гигантские вирусы – большие сюрпризы» . Вирусы . 11 (5): 404. дои : 10.3390/v11050404 . ПМК 6563228 . ПМИД 31052218 .
- ^ Jump up to: а б Прангишвили Д., Фортер П., Гаррет Р.А. (ноябрь 2006 г.). «Вирусы архей: объединяющий взгляд». Обзоры природы. Микробиология . 4 (11): 837–48. дои : 10.1038/nrmicro1527 . ПМИД 17041631 . S2CID 9915859 .
- ^ «База данных вирусных геномов NCBI» . ncbi.nlm.nih.gov . Проверено 15 января 2017 г.
- ^ Пенниси Э (март 2011 г.). «Микробиология. Виральность: изучение роли вирусов в нашем организме». Наука . 331 (6024): 1513. Бибкод : 2011Sci...331.1513P . дои : 10.1126/science.331.6024.1513 . ПМИД 21436418 .
- ^ Ши М., Линь XD, Тянь Дж.Х., Чен Л.Дж., Чен X, Ли CX и др. (декабрь 2016 г.). «Переопределение виросферы РНК беспозвоночных». Природа . 540 (7634): 539–43. Бибкод : 2016Natur.540..539S . дои : 10.1038/nature20167 . ПМИД 27880757 . S2CID 1198891 .
- ^ Сондерс В.А., Картер Дж. (2007). Вирусология: принципы и приложения . Чичестер: Джон Уайли и сыновья. п. 72. ИСБН 978-0-470-02387-7 .
- ^ Белый В.А., Левин А.Дж., Скалка А.М. (декабрь 2010 г.). «Последовательности предковых одноцепочечных ДНК-вирусов в геномах позвоночных: парвовирусам и цирковирусам более 40–50 миллионов лет» . Журнал вирусологии . 84 (23): 12458–62. дои : 10.1128/JVI.01789-10 . ПМЦ 2976387 . ПМИД 20861255 .
- ^ Брандес Н., Линиал М. (май 2016 г.). «Перекрытие генов и ограничения размера в вирусном мире» . Биология Директ . 11 (1): 26. дои : 10.1186/s13062-016-0128-3 . ПМЦ 4875738 . ПМИД 27209091 .
- ^ Нажатие J, Рианни, округ Колумбия (1984). «Разделенные геномы и внутренний шум» . Журнал молекулярной эволюции . 20 (2): 135–46. Бибкод : 1984JMolE..20..135P . дои : 10.1007/BF02257374 . ПМК 7087551 . ПМИД 6433032 .
- ^ Даффи С., Холмс ЕС (июнь 2009 г.). «Подтверждение высоких показателей нуклеотидных замен в геминивирусах: филогенетические данные восточноафриканских вирусов мозаики маниоки» . Журнал общей вирусологии . 90 (Часть 6): 1539–47. дои : 10.1099/vir.0.009266-0 . ПМК 4091138 . ПМИД 19264617 .
- ^ Сандбулте М.Р., Вестгест К.Б., Гао Дж., Сюй X, Климов А.И., Рассел К.А. и др. (декабрь 2011 г.). «Диссонансный антигенный дрейф нейраминидазы и гемагглютинина у вирусов гриппа H1N1 и H3N2» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (51): 20748–53. Бибкод : 2011PNAS..10820748S . дои : 10.1073/pnas.1113801108 . ПМК 3251064 . ПМИД 22143798 .
- ^ Мосс Р.Б., Дэйви Р.Т., Стейгбигель Р.Т., Фанг Ф. (июнь 2010 г.). «Нацеленность на пандемический грипп: введение в противовирусные препараты против гриппа и лекарственную устойчивость» . Журнал антимикробной химиотерапии . 65 (6): 1086–93. дои : 10.1093/jac/dkq100 . ПМИД 20375034 .
- ^ Хэмпсон А.В., Маккензи Дж.С. (ноябрь 2006 г.). «Вирусы гриппа». Медицинский журнал Австралии . 185 (С10): С39–43. дои : 10.5694/j.1326-5377.2006.tb00705.x . ПМИД 17115950 . S2CID 17069567 .
- ^ Мецнер К.Дж. (декабрь 2006 г.). «Обнаружение и значение квазивидов меньшинства лекарственно-устойчивого ВИЧ-1». Журнал терапии ВИЧ . 11 (4): 74–81. ПМИД 17578210 .
- ^ Гаудсмит, Яап. Вирусный секс . Издательство Оксфордского университета, 1998. ISBN 978-0-19-512496-5
- ^ Воробей М., Холмс ЕС (октябрь 1999 г.). «Эволюционные аспекты рекомбинации РНК-вирусов» . Журнал общей вирусологии . 80 (10): 2535–43. дои : 10.1099/0022-1317-80-10-2535 . ПМИД 10573145 .
- ^ Лукашев А.Н. (2005). «Роль рекомбинации в эволюции энтеровирусов». Обзоры по медицинской вирусологии . 15 (3): 157–67. дои : 10.1002/rmv.457 . ПМИД 15578739 . S2CID 26000112 .
- ^ Умене К. (июль 1999 г.). «Механизм и применение генетической рекомбинации у герпесвирусов». Обзоры по медицинской вирусологии . 9 (3): 171–82. doi : 10.1002/(SICI)1099-1654(199907/09)9:3<171::AID-RMV243>3.0.CO;2-A . ПМИД 10479778 . S2CID 43110533 .
- ^ Су С., Вонг Г., Ши В., Лю Дж., Лай А.С., Чжоу Дж. и др. (июнь 2016 г.). «Эпидемиология, генетическая рекомбинация и патогенез коронавирусов» . Тенденции в микробиологии . 24 (6): 490–502. дои : 10.1016/j.tim.2016.03.003 . ПМК 7125511 . ПМИД 27012512 .
- ^ Барр Дж. Н., Фернс Р. (июнь 2010 г.). «Как РНК-вирусы сохраняют целостность своего генома». Журнал общей вирусологии . 91 (Часть 6): 1373–87. дои : 10.1099/vir.0.020818-0 . ПМИД 20335491 .
- ^ Фрид Э.О. (август 2015 г.). «Сборка, высвобождение и созревание ВИЧ-1» . Обзоры природы. Микробиология . 13 (8): 484–96. дои : 10.1038/nrmicro3490 . ПМК 6936268 . ПМИД 26119571 .
- ^ Инь Дж., Редович Дж. (июнь 2018 г.). «Кинетическое моделирование роста вирусов в клетках» . Обзоры микробиологии и молекулярной биологии . 82 (2). дои : 10.1128/MMBR.00066-17 . ПМЦ 5968458 . ПМИД 29592895 .
- ^ Мас В., Мелеро Х.А. (2013). «Вход вирусов с оболочкой в клетки-хозяева: слияние мембран». Структура и физика вирусов . Субклеточная биохимия. Том. 68. стр. 467–87. дои : 10.1007/978-94-007-6552-8_16 . ISBN 978-94-007-6551-1 . ПМК 7121288 . ПМИД 23737062 .
- ^ Боевинк П., Опарка К.Дж. (август 2005 г.). «Взаимодействие вируса с хозяином в процессах перемещения» . Физиология растений . 138 (4): 1815–21. дои : 10.1104/стр.105.066761 . ПМЦ 1183373 . ПМИД 16172094 .
- ^ Блаас Д. (май 2016 г.). «Пути проникновения вирусов: на примере вирусов простуды» . Wiener Medizinische Wochenschrift . 166 (7–8): 211–26. дои : 10.1007/s10354-016-0461-2 . ПМЦ 4871925 . ПМИД 27174165 .
- ^ Исомура Х., Стински М.Ф. (февраль 2013 г.). «Координация поздней транскрипции гена цитомегаловируса человека с синтезом вирусной ДНК: рекомбинантные вирусы как потенциальные кандидаты на терапевтические вакцины». Мнение экспертов о терапевтических целях . 17 (2): 157–66. дои : 10.1517/14728222.2013.740460 . ПМИД 23231449 . S2CID 11448687 .
- ^ Барман С., Али А., Хуэй Е.К., Адхикари Л., Наяк Д.П. (сентябрь 2001 г.). «Транспорт вирусных белков к апикальным мембранам и взаимодействие матриксного белка с гликопротеинами при сборке вирусов гриппа». Вирусные исследования . 77 (1): 61–69. дои : 10.1016/S0168-1702(01)00266-0 . ПМИД 11451488 .
- ^ Стагиннус С, Рихерт-Пёггелер КР (октябрь 2006 г.). «Эндогенные параретровирусы: двуличные путешественники в геноме растений». Тенденции в науке о растениях . 11 (10): 485–91. Бибкод : 2006TPS....11..485S . doi : 10.1016/j.tplants.2006.08.008 . ПМИД 16949329 .
- ^ Роулстон А., Марселлус Р.К., Брэнтон П.Е. (1999). «Вирусы и апоптоз». Ежегодный обзор микробиологии . 53 : 577–628. дои : 10.1146/аннурев.микро.53.1.577 . ПМИД 10547702 .
- ^ Алвин Дж. К. (2008). «Модуляция стрессовых реакций клеток-хозяев с помощью цитомегаловируса человека». Цитомегаловирус человека . Актуальные темы микробиологии и иммунологии. Том. 325. стр. 263–79. дои : 10.1007/978-3-540-77349-8_15 . ISBN 978-3-540-77348-1 . ПМИД 18637511 .
- ^ Бароцци П., Потенца Л., Рива Г., Валлерини Д., Квадрелли С., Боско Р. и др. (декабрь 2007 г.). «В-клетки и герпесвирусы: модель лимфопролиферации». Обзоры аутоиммунитета . 7 (2): 132–36. дои : 10.1016/j.autrev.2007.02.018 . ПМИД 18035323 .
- ^ Субраманья Д., Гривас П.Д. (ноябрь 2008 г.). «ВПЧ и рак шейки матки: обновленная информация об установившихся отношениях». Последипломное образование по медицине . 120 (4): 7–13. doi : 10.3810/pgm.2008.11.1928 . ПМИД 19020360 . S2CID 1399003 .
- ^ Синклер Дж. (март 2008 г.). «Цитомегаловирус человека: латентный период и реактивация миелоидного ряда». Журнал клинической вирусологии . 41 (3): 180–85. дои : 10.1016/j.jcv.2007.11.014 . ПМИД 18164651 .
- ^ Джордан MC, Джордан Г.В., Стивенс Дж.Г., Миллер Дж. (июнь 1984 г.). «Латентные герпесвирусы человека». Анналы внутренней медицины . 100 (6): 866–80. дои : 10.7326/0003-4819-100-6-866 . ПМИД 6326635 .
- ^ Сиссонс Дж.Г., Бэйн М., Уиллс М.Р. (февраль 2002 г.). «Латентность и реактивация цитомегаловируса человека». Журнал инфекции . 44 (2): 73–77. дои : 10.1053/jinf.2001.0948 . ПМИД 12076064 .
- ^ Кроуфорд Д.Х. (2011). «Глава 2: Вирусы повсюду» . Вирусы: очень краткое введение . Издательство Оксфордского университета, США. п. 16 . ISBN 978-0-19-957485-8 .
- ^ Баггесен Д.Л., Соренсен Г., Нильсен Э.М., Вегенер Х.К. (январь 2010 г.). «Фаготипирование Salmonella Typhimurium – остается ли оно полезным инструментом для надзора и расследования вспышек?» . Евронаблюдение . 15 (4): 19471. PMID 20122382 . Проверено 19 декабря 2014 г.
- ^ Паркер М.Т. (сентябрь 2016 г.). «Экологическая основа человеческого вирома обеспечивает классификацию текущих знаний и определяет области предстоящих открытий» . Йельский журнал биологии и медицины . 89 (3): 339–51. ПМК 5045143 . ПМИД 27698618 .
- ^ Вулхаус М., Скотт Ф., Хадсон З., Хоуи Р., Чейз-Топпинг М. (октябрь 2012 г.). «Человеческие вирусы: открытие и возникновение» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 367 (1604): 2864–71. дои : 10.1098/rstb.2011.0354 . ПМЦ 3427559 . ПМИД 22966141 .
- ^ Jump up to: а б Ашур Х.М., Эльхатиб В.Ф., Рахман М.М., Эльшабрави Х.А. (март 2020 г.). «Информация о недавнем новом коронавирусе 2019 года (SARS-CoV-2) в свете прошлых вспышек коронавируса среди людей» . Патогены . 9 (3): 186. doi : 10.3390/pathogens9030186 . ПМК 7157630 . ПМИД 32143502 .
- ^ Львофф А., Хорн Р.В., Турнье П. (июнь 1962 г.). «[Вирусная система]». Еженедельные отчеты сессий Академии наук (на французском языке). 254 :4225–27. ПМИД 14467544 .
- ^ Львофф А., Хорн Р., Турнье П. (1962). «Система вирусов». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 27 : 51–55. дои : 10.1101/sqb.1962.027.001.008 . ПМИД 13931895 .
- ^ Фоке СМ, Фаржетт Д (август 2005 г.). «Международный комитет по таксономии вирусов и 3142 неустановленных видов» . Вирусологический журнал . 2:64 . дои : 10.1186/1743-422X-2-64 . ПМК 1208960 . ПМИД 16105179 .
- ^ Исполнительный комитет Международного комитета по таксономии вирусов (май 2020 г.). «Новая сфера таксономии вирусов: разделение виросферы на 15 иерархических рангов» . Нат Микробиол . 5 (5): 668–74. дои : 10.1038/s41564-020-0709-x . ПМК 7186216 . ПМИД 32341570 .
- ^ Хан МК, Алам ММ (июль 2021 г.). «Вспышки норовирусного гастроэнтерита, геномное разнообразие и эволюция: обзор». Медицинский журнал Майменсингха . 30 (3): 863–73. ПМИД 34226482 .
- ^ Эберле Дж., Гюртлер Л. (2012). «Типы, группы, подтипы и рекомбинантные формы ВИЧ: ошибки репликации, давление отбора и квазивиды» . Интервирусология . 55 (2): 79–83. дои : 10.1159/000331993 . ПМИД 22286874 . S2CID 5642060 .
- ^ Исполнительный комитет Международного комитета по таксономии вирусов (май 2020 г.). «Новая сфера таксономии вирусов: разделение виросферы на 15 иерархических рангов» . Природная микробиология . 5 (5): 668–674. дои : 10.1038/s41564-020-0709-x . ПМК 7186216 . ПМИД 32341570 .
- ^ Делварт Э.Л. (2007). «Вирусная метагеномика» . Обзоры по медицинской вирусологии . 17 (2): 115–131. дои : 10.1002/rmv.532 . ПМК 7169062 . ПМИД 17295196 .
- ^ Темин Х.М., Балтимор Д. (1972). «РНК-направленный синтез ДНК и РНК-опухолевые вирусы». Достижения в области исследования вирусов . 17 : 129–86. дои : 10.1016/S0065-3527(08)60749-6 . ISBN 978-0120398171 . ПМИД 4348509 .
- ^ Балтимор Д. (1974). «Стратегия РНК-вирусов». Лекции Харви . 70 серия. 70 серия: 57–74. ПМИД 4377923 .
- ^ ван Регенмортель М.Х., Махи Б.В. (январь 2004 г.). «Новые проблемы систематики вирусов» . Новые инфекционные заболевания . 10 (1): 8–13. дои : 10.3201/eid1001.030279 . ПМЦ 3322749 . ПМИД 15078590 .
- ^ Мэйо, Массачусетс (1999). «Достижения в таксономии вирусов растений с момента публикации 6-го отчета ICTV. Международный комитет по таксономии вирусов» . Архив вирусологии . 144 (8): 1659–66. дои : 10.1007/s007050050620 . ПМИД 10486120 . S2CID 33422303 .
- ^ де Вильерс Э.М., Фоке С., Брокер Т.Р., Бернар Х.У., Цур Хаузен Х. (июнь 2004 г.). «Классификация папилломавирусов». Вирусология . 324 (1): 17–27. дои : 10.1016/j.virol.2004.03.033 . ПМИД 15183049 .
- ^ Фишер Б., Харви Р.П., Чамп ПК (2007). «Глава 33 (Краткое описание болезней)». Иллюстрированные обзоры Липпинкотта: Микробиология . Серия иллюстрированных обзоров Липпинкотта. Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. стр. 367–392. ISBN 978-0-7817-8215-9 .
- ^ Комаров А.Л. (декабрь 2006 г.). «Является ли вирус герпеса человека-6 триггером синдрома хронической усталости?». Журнал клинической вирусологии . 37 (Приложение 1): S39–46. дои : 10.1016/S1386-6532(06)70010-5 . ПМИД 17276367 .
- ^ Чен Ч., Чиу Ю.Л., Вэй Ф.К., Кунг Ф.Дж., Лю Х.К., Шоу С.К. и др. (январь 1999 г.). «Высокая распространенность серовирусной инфекции Борна среди больных шизофренией, членов их семей и работников психиатрической службы на Тайване» . Молекулярная психиатрия . 4 (1): 33–38. дои : 10.1038/sj.mp.4000484 . ПМИД 10089006 . S2CID 19830976 .
- ^ Марголис Т.П., Эльфман Ф.Л., Лейб Д., Пакпур Н., Апакупакул К., Имаи Ю. и др. (октябрь 2007 г.). «Спонтанная реактивация вируса простого герпеса типа 1 в латентно инфицированных сенсорных ганглиях мышей» . Журнал вирусологии . 81 (20): 11069–74. дои : 10.1128/JVI.00243-07 . ПМК 2045564 . ПМИД 17686862 .
- ^ Уитли Р.Дж., Ройзман Б. (май 2001 г.). «Инфекции, вызванные вирусом простого герпеса». Ланцет . 357 (9267): 1513–18. дои : 10.1016/S0140-6736(00)04638-9 . ПМИД 11377626 . S2CID 9854903 .
- ^ Бартон Э.С., Уайт Д.В., Кэтлин Дж.С., Бретт-Макклеллан К.А., Энгл М., Даймонд М.С. и др. (май 2007 г.). «Латентность вируса герпеса обеспечивает симбиотическую защиту от бактериальной инфекции». Природа . 447 (7142): 326–29. Бибкод : 2007Natur.447..326B . дои : 10.1038/nature05762 . ПМИД 17507983 . S2CID 4425405 .
- ^ Бертолетти А., Геринг А. (октябрь 2007 г.). «Иммунный ответ и толерантность при хронической инфекции вируса гепатита В». Гепатологические исследования . 37 (Приложение 3): S331–38. дои : 10.1111/j.1872-034X.2007.00221.x . ПМИД 17931183 . S2CID 13386004 .
- ^ Родригес С., Дешмук М., Джейкоб Т., Нукала Р., Менон С., Мехта А. (2001). «Значение ДНК ВГВ с помощью ПЦР по сравнению с серологическими маркерами ВГВ у острых и хронических пациентов». Индийский журнал медицинской микробиологии . 19 (3): 141–44. ПМИД 17664817 .
- ^ Нгуен В.Т., Маклоуз М.Л., Доре Дж.Дж. (декабрь 2007 г.). «Высокоэндемичная инфекция гепатита В в сельских районах Вьетнама». Журнал гастроэнтерологии и гепатологии . 22 (12): 2093–100. дои : 10.1111/j.1440-1746.2007.05010.x . ПМИД 17645465 . S2CID 29885790 .
- ^ Фаулер М.Г., Лампе М.А., Джеймисон Д.Д., Куртис А.П., Роджерс М.Ф. (сентябрь 2007 г.). «Снижение риска передачи вируса иммунодефицита человека от матери к ребенку: прошлые успехи, текущий прогресс и проблемы, а также будущие направления» . Американский журнал акушерства и гинекологии . 197 (3 приложения): С3–9. дои : 10.1016/j.ajog.2007.06.048 . ПМИД 17825648 .
- ^ Зауэрбрей А., Вутцлер П. (декабрь 2000 г.). «Синдром врожденной ветряной оспы». Журнал перинатологии . 20 (8, часть 1): 548–54. дои : 10.1038/sj.jp.7200457 . ПМИД 11190597 . S2CID 7973561 .
- ^ Jump up to: а б Антонович Дж., Уилсон А.Дж., Форбс М.Р., Хауффе Х.К., Каллио Э.Р., Леггетт Х.К. и др. (май 2017 г.). «Эволюция способа передачи» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 372 (1719). дои : 10.1098/rstb.2016.0083 . ПМК 5352810 . ПМИД 28289251 .
- ^ Гарнетт GP (февраль 2005 г.). «Роль коллективного иммунитета в определении эффекта вакцин против болезней, передающихся половым путем» . Журнал инфекционных болезней . 191 (Приложение 1): S97–106. дои : 10.1086/425271 . ПМИД 15627236 .
- ^ Платонов А.Е. (2006). «[Влияние погодных условий на эпидемиологию трансмиссивных болезней на примере лихорадки Западного Нила в России]». Вестник Российской академии медицинских наук (на русском языке) (2): 25–29. ПМИД 16544901 .
- ^ Джуэлл С.П., Килинг М.Дж., Робертс Г.О. (декабрь 2009 г.). «Прогнозирование необнаруженных инфекций во время вспышки ящура в 2007 году» . Журнал Королевского общества, Интерфейс . 6 (41): 1145–51. дои : 10.1098/rsif.2008.0433 . ПМК 2817150 . ПМИД 19091686 .
- ^ Паттерсон К.Д., Пайл Г.Ф. (1991). «География и смертность от пандемии гриппа 1918 года». Бюллетень истории медицины . 65 (1): 4–21. ПМИД 2021692 .
- ^ Джонсон Н.П., Мюллер Дж. (2002). «Обновление отчетов: глобальная смертность от пандемии «испанского» гриппа 1918-1920 годов». Бюллетень истории медицины . 76 (1): 105–15. дои : 10.1353/bhm.2002.0022 . ПМИД 11875246 . S2CID 22974230 .
- ^ Эйзингер Р.В., Фаучи А.С. (март 2018 г.). «1» . Новые инфекционные заболевания . 24 (3): 413–16. дои : 10.3201/eid2403.171797 . ПМК 5823353 . ПМИД 29460740 .
- ^ , Ли XQ, Чжэн Д.Д., Пэн З.Б. и др. (август 2018 г.) «[История пандемий гриппа в Китае за последнее столетие]». Цинь Ю, Чжао М.Ю., Тан Ю. Люсинбинсюэ Зажи (на китайском языке) 39 (8): 1028–31. doi : 10.3760/cma.j.issn.0254-6450.2018.08.003 . PMID 30180422 .
- ^ Гао Ф., Бэйлз Э., Робертсон Д.Л., Чен Ю., Роденбург К.М., Майкл С.Ф. и др. (февраль 1999 г.). «Происхождение ВИЧ-1 у шимпанзе Pan troglodytes troglodytes» . Природа . 397 (6718): 436–41. Бибкод : 1999Natur.397..436G . дои : 10.1038/17130 . ПМИД 9989410 . S2CID 4432185 .
- ^ «Информационный бюллетень» (PDF) . ЮНЭЙДС.орг . 2018 . Проверено 12 декабря 2019 г.
- ^ «ДАННЫЕ ООН ПО СПИДУ 2019» . ЮНЭЙДС.орг . 2019 . Проверено 5 декабря 2019 г.
- ^ Мавар Н., Саха С., Пандит А., Махаджан У (декабрь 2005 г.). «Третья фаза пандемии ВИЧ: социальные последствия стигмы и дискриминации ВИЧ/СПИДа и будущие потребности» (PDF) . Индийский журнал медицинских исследований . 122 (6): 471–84. ПМИД 16517997 . Архивировано из оригинала (PDF) 4 марта 2016 года . Проверено 19 декабря 2014 г.
- ^ «Состояние глобальной эпидемии ВИЧ» (PDF) . ЮНЭЙДС. 2008. Архивировано из оригинала (PDF) 22 ноября 2015 года . Проверено 19 декабря 2014 г.
- ^ Таунер Дж.С., Христова М.Л., Сили Т.К., Винсент М.Дж., Эриксон Б.Р., Бавец Д.А. и др. (июль 2006 г.). «Геномика марбургвируса и связь с крупной вспышкой геморрагической лихорадки в Анголе» . Журнал вирусологии . 80 (13): 6497–516. дои : 10.1128/JVI.00069-06 . ПМЦ 1488971 . ПМИД 16775337 .
- ^ «Отчет Всемирной организации здравоохранения, 24 сентября 2014 г.» (PDF) .
- ^ «Вирусологический журнал» . Вирусологический журнал .
- ^ Вайс С.Р., Лейбовиц Дж.Л. (2011). Патогенез коронавируса . Достижения в области исследования вирусов. Том. 81. С. 85–164. дои : 10.1016/B978-0-12-385885-6.00009-2 . ISBN 978-0-12-385885-6 . ПМК 7149603 . ПМИД 22094080 .
- ^ Вонг А.Т., Чен Х., Лю Ш., Сюй Е.К., Лук К.С., Лай К.К. и др. (май 2017 г.). «От атипичной пневмонии к обеспечению готовности к птичьему гриппу в Гонконге» . Клинические инфекционные болезни . 64 (дополнение_2): S98–S104. дои : 10.1093/cid/cix123 . ПМИД 28475794 .
- ^ Дэн SQ, Пэн HJ (февраль 2020 г.). «Характеристики и ответные меры общественного здравоохранения на вспышку коронавирусной болезни в Китае в 2019 году» . Журнал клинической медицины . 9 (2): 575. дои : 10.3390/jcm9020575 . ПМК 7074453 . ПМИД 32093211 .
- ^ Хан Кью, Линь Кью, Джин С., Ю Л (апрель 2020 г.). «Коронавирус 2019-nCoV: краткий взгляд с передовой» . Журнал инфекции . 80 (4): 373–77. дои : 10.1016/j.jinf.2020.02.010 . ПМК 7102581 . ПМИД 32109444 .
- ^ Лондоньо Э, Ортис А (16 марта 2020 г.). «Ограничения на поездки из-за коронавируса по всему миру» . Нью-Йорк Таймс .
- ^ «США предпринимают более масштабные шаги по реагированию на пандемию; в Европе растет число случаев Covid-19» . ЦИДРАП . 15 марта 2020 г.
- ^ Эйнштейн М.Х., Шиллер Дж.Т., Вискиди Р.П., Стриклер Х.Д., Курсаже П., Тан Т. и др. (июнь 2009 г.). «Руководство для клиницистов по иммунологии вируса папилломы человека: известное и неизвестное». «Ланцет». Инфекционные болезни . 9 (6): 347–56. дои : 10.1016/S1473-3099(09)70108-2 . ПМИД 19467474 .
- ^ Шуда М., Фенг Х., Квун Х.Дж., Розен С.Т., Джоеруп О., Мур П.С. и др. (октябрь 2008 г.). «Мутации Т-антигена являются специфичным для опухоли признаком полиомавируса клеток Меркеля» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (42): 16272–77. Бибкод : 2008PNAS..10516272S . дои : 10.1073/pnas.0806526105 . ПМК 2551627 . ПМИД 18812503 .
- ^ Пулитцеровский член парламента, Амин Б.Д., Бусам К.Дж. (май 2009 г.). «Рак из клеток Меркеля: обзор». Достижения анатомической патологии . 16 (3): 135–44. дои : 10.1097/PAP.0b013e3181a12f5a . ПМИД 19395876 . S2CID 36110778 .
- ^ Койке К. (июнь 2007 г.). «Вирус гепатита С способствует гепатоканцерогенезу путем модуляции метаболических и внутриклеточных сигнальных путей». Журнал гастроэнтерологии и гепатологии . 22 (Приложение 1): S108–11. дои : 10.1111/j.1440-1746.2006.04669.x . ПМИД 17567457 . S2CID 25399220 .
- ^ Ху Дж., Ладгейт Л. (2007). «Коинфекция ВИЧ-ВГВ и ВИЧ-ВГС и развитие рака печени». Вирусный онкогенез, ассоциированный со СПИДом . Лечение и исследования рака. Том. 133. стр. 241–52. дои : 10.1007/978-0-387-46816-7_9 . ISBN 978-0-387-46804-4 . ПМИД 17672044 .
- ^ Беллон М., Никот С. (2007). «Теломераза: решающий игрок в Т-клеточном лейкозе человека, вызванном HTLV-I». Геномика и протеомика рака . 4 (1): 21–25. ПМИД 17726237 .
- ^ Шиффман М., Касл П.Е., Джеронимо Дж., Родригес А.С., Вахолдер С. (сентябрь 2007 г.). «Вирус папилломы человека и рак шейки матки». Ланцет . 370 (9590): 890–907. дои : 10.1016/S0140-6736(07)61416-0 . ПМИД 17826171 . S2CID 20196938 .
- ^ Кляйн Э., Кис Л.Л., Кляйн Г. (февраль 2007 г.). «Вирусная инфекция Эпштейна-Барра у человека: от безобидных до опасных для жизни взаимодействий вируса и лимфоцитов» . Онкоген . 26 (9): 1297–305. дои : 10.1038/sj.onc.1210240 . ПМИД 17322915 .
- ^ Цур Хаузен Х (июль 2008 г.). «Новые полиомавирусы человека – возрождение хорошо известного семейства вирусов как возможных канцерогенов для человека» . Международный журнал рака . 123 (2): 247–50. дои : 10.1002/ijc.23620 . ПМИД 18449881 . S2CID 9482506 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтерс П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк и Лондон: Garland Science. ISBN 0-8153-3218-1 .
- ^ Дин С.В., Войннет О (август 2007 г.). «Противовирусный иммунитет, управляемый малыми РНК» . Клетка . 130 (3): 413–26. дои : 10.1016/j.cell.2007.07.039 . ПМК 2703654 . ПМИД 17693253 .
- ^ Паттон Дж.Т., Васкес-Дель Карпио Р., Спенсер Э. (2004). «Репликация и транскрипция генома ротавируса». Текущий фармацевтический дизайн . 10 (30): 3769–77. дои : 10.2174/1381612043382620 . ПМИД 15579070 .
- ^ Джаярам Х., Эстес М.К., Прасад Б.В. (апрель 2004 г.). «Новые темы проникновения в клетки ротавируса, организации генома, транскрипции и репликации». Вирусные исследования . 101 (1): 67–81. doi : 10.1016/j.virusres.2003.12.007 . ПМИД 15010218 .
- ^ Грир С., Александр Г.Дж. (декабрь 1995 г.). «Вирусная серология и обнаружение». Клиническая гастроэнтерология Байьера . 9 (4): 689–721. дои : 10.1016/0950-3528(95)90057-8 . ПМИД 8903801 .
- ^ Материя Л., Когельшац К., Германн Д. (апрель 1997 г.). «Уровни антител к вирусу краснухи в сыворотке, указывающие на иммунитет: ответ на вакцинацию субъектов с низкими или неопределяемыми концентрациями антител» . Журнал инфекционных болезней . 175 (4): 749–55. дои : 10.1086/513967 . ПМИД 9086126 .
- ^ Маллери Д.Л., Макьюэн В.А., Бидгуд С.Р., Тауэрс Дж.Дж., Джонсон К.М., Джеймс Л.К. (ноябрь 2010 г.). «Антитела опосредуют внутриклеточный иммунитет посредством трехстороннего мотива, содержащего 21 (TRIM21)» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 19985–90. Бибкод : 2010PNAS..10719985M . дои : 10.1073/pnas.1014074107 . ПМЦ 2993423 . ПМИД 21045130 .
- ^ Каскальо М., Платт Дж.Л. (2007). «Новые функции B-клеток». Критические обзоры по иммунологии . 27 (2): 141–51. doi : 10.1615/critrevimmunol.v27.i2.20 . ПМИД 17725500 .
- ^ Ле Пейдж С., Женен П., Бейнс М.Г., Хискотт Дж. (2000). «Активация интерферона и врожденный иммунитет». Обзоры в разделе Иммуногенетика . 2 (3): 374–86. ПМИД 11256746 .
- ^ Хиллеман М.Р. (октябрь 2004 г.). «Стратегии и механизмы выживания хозяина и возбудителя при острых и персистирующих вирусных инфекциях» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (Приложение 2): 14560–66. Бибкод : 2004PNAS..10114560H . дои : 10.1073/pnas.0404758101 . ПМК 521982 . ПМИД 15297608 .
- ^ Либби Дж. Э., Фудзинами Р. С. (2014). «Адаптивный иммунный ответ на вирусные инфекции в центральной нервной системе». Нейровирусология . Справочник по клинической неврологии. Том. 123. С. 225–47. дои : 10.1016/B978-0-444-53488-0.00010-9 . ISBN 978-0444534880 . ПМК 4370180 . ПМИД 25015488 .
- ^ Асария П., МакМахон Э. (октябрь 2006 г.). «Корь в Соединенном Королевстве: сможем ли мы искоренить ее к 2010 году?» . БМЖ . 333 (7574): 890–95. дои : 10.1136/bmj.38989.445845.7C . ПМЦ 1626346 . ПМИД 17068034 .
- ^ Лейн Дж. М. (2006). «Массовая вакцинация и надзор/сдерживание в целях искоренения оспы». Массовая вакцинация: глобальные аспекты — прогресс и препятствия . Актуальные темы микробиологии и иммунологии. Том. 304. стр. 17–29. дои : 10.1007/3-540-36583-4_2 . ISBN 978-3-540-29382-8 . ПМК 7120753 . ПМИД 16989262 .
- ^ Арвин А.М., Гринберг Х.Б. (январь 2006 г.). «Новые вирусные вакцины» . Вирусология . 344 (1): 240–49. дои : 10.1016/j.virol.2005.09.057 . ПМИД 16364754 .
- ^ Пасторет П.П., Шудель А.А., Ломбард М. (август 2007 г.). «Выводы - будущие тенденции в ветеринарной вакцинологии». Revue Scientifique et Technique . 26 (2): 489–94, 495–501, 503–09. дои : 10.20506/rst.26.2.1759 . ПМИД 17892169 .
- ^ Палезе П. (январь 2006 г.). «Создание более качественных вакцин против вируса гриппа?» . Новые инфекционные заболевания . 12 (1): 61–65. дои : 10.3201/eid1201.051043 . ПМК 3291403 . ПМИД 16494719 .
- ^ Ананд П., вице-президент Stahel (май 2021 г.). «Обзор безопасности мРНК-вакцин Covid-19: обзор» . Безопасность пациентов в хирургии . 15 (1): 20. дои : 10.1186/s13037-021-00291-9 . ПМЦ 8087878 . ПМИД 33933145 .
- ^ Томссен Р. (1975). «Живые аттенуированные и убитые вирусные вакцины». Монографии по аллергии . 9 : 155–76. ПМИД 1090805 .
- ^ Маклин А.А. (1986). «Разработка вакцин против гепатита А и гепатита В». Обзоры инфекционных болезней . 8 (4): 591–98. дои : 10.1093/clinids/8.4.591 . ПМИД 3018891 .
- ^ Кассуолл Т.Х., Фишлер Б. (октябрь 2005 г.). «Вакцинация ребенка с ослабленным иммунитетом». Экспертная оценка вакцин . 4 (5): 725–38. дои : 10.1586/14760584.4.5.725 . ПМИД 16221073 . S2CID 40821818 .
- ^ Барнетт Э.Д., Уайлдер-Смит А., Уилсон М.Э. (июль 2008 г.). «Вакцины против желтой лихорадки и международные путешественники». Экспертная оценка вакцин . 7 (5): 579–87. дои : 10.1586/14760584.7.5.579 . ПМИД 18564013 . S2CID 19352868 .
- ^ Jump up to: а б Де Клерк Э., Ли Г (июль 2016 г.). «Утвержденные противовирусные препараты за последние 50 лет» . Обзоры клинической микробиологии . 29 (3): 695–747. дои : 10.1128/CMR.00102-15 . ПМЦ 4978613 . ПМИД 27281742 .
- ^ Магден Дж., Кяарияйнен Л., Ахола Т. (март 2005 г.). «Ингибиторы репликации вирусов: последние разработки и перспективы» . Прикладная микробиология и биотехнология . 66 (6): 612–21. дои : 10.1007/s00253-004-1783-3 . ПМК 7082807 . ПМИД 15592828 .
- ^ Миндель А., Сазерленд С. (сентябрь 1983 г.). «Генитальный герпес – заболевание и его лечение, включая внутривенное введение ацикловира». Журнал антимикробной химиотерапии . 12 (Приложение Б): 51–59. дои : 10.1093/jac/12.suppl_b.51 . ПМИД 6355051 .
- ^ Пальмизано Л., Велла С. (2011). «Краткая история антиретровирусной терапии ВИЧ-инфекции: успехи и проблемы». Анналы Istituto Superiore di Sanità . 47 (1): 44–48. дои : 10.4415/ANN_11_01_10 . ПМИД 21430338 .
- ^ Фаладе-Нвулия О, Суарес-Куэрво С, Нельсон Д.Р., Фрид М.В., Сигал Дж.Б., Сулковски М.С. (май 2017 г.). «Поральная терапия агентами прямого действия при инфекции вируса гепатита С: систематический обзор» . Анналы внутренней медицины . 166 (9): 637–48. дои : 10.7326/M16-2575 . ПМЦ 5486987 . ПМИД 28319996 .
- ^ Нгуен М.Х., Вонг Дж., Гейн Э., Као Дж.Х., Душейко Г. (март 2020 г.). «Вирус гепатита В: достижения в профилактике, диагностике и терапии» . Обзоры клинической микробиологии . 33 (2). дои : 10.1128/CMR.00046-19 . ПМК 7048015 . ПМИД 32102898 .
- ^ Горис Н., Ванденбуше Ф., Де Клерк К. (апрель 2008 г.). «Возможности противовирусной терапии и профилактики для борьбы с РНК-вирусными инфекциями домашнего скота». Противовирусные исследования . 78 (1): 170–78. doi : 10.1016/j.antiviral.2007.10.003 . ПМИД 18035428 .
- ^ Кармайкл Л.Е. (2005). «Аннотированный исторический отчет о собачьем парвовирусе». Журнал ветеринарной медицины. Б. Инфекционные болезни и ветеринарное здравоохранение . 52 (7–8): 303–11. дои : 10.1111/j.1439-0450.2005.00868.x . ПМИД 16316389 .
- ^ Чен Ю, Чжао Ю, Хаммонд Дж, Сюй Х.Т., Эванс Дж, Фельдлауфер М (октябрь – ноябрь 2004 г.). «Множественные вирусные инфекции у медоносных пчел и дивергенция геномов вирусов медоносных пчел» . Журнал патологии беспозвоночных . 87 (2–3): 84–93. Бибкод : 2004JInvP..87...84C . дои : 10.1016/j.jip.2004.07.005 . ПМИД 15579317 .
- ^ Халл Р. (2002). «Глава 12: Передача 2: Механическая, семенная, пыльцевая и эпидемиология» . Вирусология растений Мэтьюза (4-е изд.). Академическая пресса. стр. 555–56. ISBN 978-0123611604 . Проверено 21 февраля 2022 г.
- ^ Захир К., Ахтар М.Х. (2016). «Производство, использование и питание картофеля – обзор». Критические обзоры в области пищевой науки и питания . 56 (5): 711–21. дои : 10.1080/10408398.2012.724479 . ПМИД 24925679 . S2CID 33074838 .
- ^ Фуэнтес С., Джонс Р.А., Мацуока Х., Осима К., Кройце Дж., Гиббс А.Дж. (июль 2019 г.). «Картофельный вирус Y; связь с Андами» . Эволюция вирусов . 5 (2): vez037. дои : 10.1093/ve/vez037 . ПМК 6755682 . ПМИД 31559020 .
- ^ Динеш-Кумар С.П., Там В.Х., Бейкер Б.Дж. (декабрь 2000 г.). «Структурно-функциональный анализ гена N устойчивости к вирусу табачной мозаики» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (26): 14789–94. Бибкод : 2000PNAS...9714789D . дои : 10.1073/pnas.97.26.14789 . ЧВК 18997 . ПМИД 11121079 .
- ^ Соосаар Дж.Л., Берч-Смит Т.М., Динеш-Кумар С.П. (октябрь 2005 г.). «Механизмы устойчивости растений к вирусам». Обзоры природы. Микробиология . 3 (10): 789–98. дои : 10.1038/nrmicro1239 . ПМИД 16132037 . S2CID 27311732 .
- ^ Ломоносов ГП (2011). «Вирусные частицы и использование таких частиц в био- и нанотехнологиях». Последние достижения в вирусологии растений . Кайстер Академик Пресс . ISBN 978-1-904455-75-2 .
- ^ Уоммак К.Е., Колвелл Р.Р. (март 2000 г.). «Вириопланктон: вирусы в водных экосистемах» . Обзоры микробиологии и молекулярной биологии . 64 (1): 69–114. дои : 10.1128/ММБР.64.1.69-114.2000 . ПМК 98987 . ПМИД 10704475 .
- ^ Берг О., Борсхайм К.Ю., Братбак Г., Хелдал М. (август 1989 г.). «Высокое количество вирусов обнаружено в водной среде». Природа . 340 (6233): 467–68. Бибкод : 1989Natur.340..467B . дои : 10.1038/340467a0 . ПМИД 2755508 . S2CID 4271861 .
- ^ Бикл Т.А., Крюгер Д.Х. (июнь 1993 г.). «Биология рестрикции ДНК» . Микробиологические обзоры . 57 (2): 434–50. дои : 10.1128/MMBR.57.2.434-450.1993 . ПМК 372918 . ПМИД 8336674 .
- ^ Баррангу Р., Фремо С., Дево Х., Ричардс М., Бояваль П., Муано С. и др. (март 2007 г.). «CRISPR обеспечивает приобретенную устойчивость прокариот к вирусам». Наука . 315 (5819): 1709–12. Бибкод : 2007Sci...315.1709B . дои : 10.1126/science.1138140 . hdl : 20.500.11794/38902 . ПМИД 17379808 . S2CID 3888761 .
- ^ Браунс С.Дж., Джор М.М., Лундгрен М., Вестра Э.Р., Слиджхуис Р.Дж., Снейдерс А.П. и др. (август 2008 г.). «Маленькие РНК CRISPR обеспечивают противовирусную защиту прокариот» . Наука . 321 (5891): 960–64. Стартовый код : 2008Sci...321..960B . дои : 10.1126/science.1159689 . ПМЦ 5898235 . ПМИД 18703739 .
- ^ Мохика Ф.Д., Родригес-Валера Ф. (сентябрь 2016 г.). «Открытие CRISPR у архей и бактерий». Журнал ФЭБС . 283 (17): 3162–69. дои : 10.1111/февраль 13766 . hdl : 10045/57676 . ПМИД 27234458 . S2CID 42827598 .
- ^ Jump up to: а б Вайс Р.А. (2017). «Обмен генетическими последовательностями между вирусами и хозяевами». Вирусы, гены и рак . Актуальные темы микробиологии и иммунологии. Том. 407. стр. 1–29. дои : 10.1007/82_2017_21 . ISBN 978-3-319-61803-6 . ПМИД 28550453 .
- ^ Ху Дж, Е Х, Ван С, Ван Дж, Хан Д (2021). «Активация профагов в кишечнике: понимание функций и возможных применений» . Границы микробиологии . 12 : 785634. doi : 10.3389/fmicb.2021.785634 . ПМЦ 8710666 . ПМИД 34966370 .
- ^ Расс Э., Иорданский С. (январь 2023 г.). «Эндогенные ретровирусы как модуляторы врожденного иммунитета» . Патогены (Базель, Швейцария) . 12 (2): 162. doi : 10.3390/pathogens12020162 . ПМЦ 9963469 . ПМИД 36839434 .
- ^ Прангишвили Д., Гаррет Р.А. (апрель 2004 г.). «Исключительно разнообразные морфотипы и геномы кренархейных гипертермофильных вирусов» (PDF) . Труды Биохимического общества . 32 (Часть 2): 204–08. дои : 10.1042/BST0320204 . ПМИД 15046572 . S2CID 20018642 .
- ^ Мохика Ф.Х., Диес-Вильясеньор К., Гарсиа-Мартинес Х., Сория Э. (февраль 2005 г.). «Промежуточные последовательности прокариотических повторов с регулярными интервалами происходят от чужеродных генетических элементов». Журнал молекулярной эволюции . 60 (2): 174–82. Бибкод : 2005JMolE..60..174M . дои : 10.1007/s00239-004-0046-3 . ПМИД 15791728 . S2CID 27481111 .
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Ю.И., Кунин Е.В. (март 2006 г.). «Предполагаемая иммунная система прокариот, основанная на РНК-интерференции: вычислительный анализ предсказанного ферментативного механизма, функциональные аналогии с эукариотическими РНКи и гипотетические механизмы действия» . Биология Директ . 1 :7. дои : 10.1186/1745-6150-1-7 . ПМК 1462988 . ПМИД 16545108 .
- ^ ван дер Оост Дж., Вестра Э.Р., Джексон Р.Н., Виденхефт Б. (июль 2014 г.). «Раскрытие структурной и механистической основы систем CRISPR-Cas» . Обзоры природы. Микробиология . 12 (7): 479–92. дои : 10.1038/nrmicro3279 . ПМЦ 4225775 . ПМИД 24909109 .
- ^ Давила-Рамос С., Кастелан-Санчес Х.Г., Мартинес-Авила Л., Санчес-Карбенте М.Д., Перальта Р., Эрнандес-Мендоса А. и др. (2019). «Обзор вирусной метагеномики в экстремальных условиях» . Границы микробиологии . 10 : 2403. дои : 10.3389/fmicb.2019.02403 . ПМК 6842933 . ПМИД 31749771 .
- ^ Чжан Ци, Гуй Дж. Ф. (декабрь 2018 г.). «Разнообразие, эволюционный вклад и экологическая роль водных вирусов». Наука Китай Науки о жизни . 61 (12): 1486–1502. дои : 10.1007/s11427-018-9414-7 . ПМИД 30443861 . S2CID 53564176 .
- ^ Вайц Дж.С., Вильгельм С.В. (2013). «Океан вирусов» . Ученый . 27 (7): 35–39.
- ^ Саттл, Калифорния (сентябрь 2005 г.). «Вирусы в море». Природа . 437 (7057): 356–61. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Вильгельм С.В., Саттл, Калифорния (1999). «Вирусы и циклы питательных веществ в море: вирусы играют решающую роль в структуре и функциях водных пищевых сетей» . Бионаука . 49 (10): 781–88. дои : 10.2307/1313569 . JSTOR 1313569 .
- ^ Шелфорд Э.Дж., Саттл, Калифорния (2018). «Вирус-опосредованный перенос азота от гетеротрофных бактерий к фитопланктону» . Биогеонауки . 15 (3): 809–15. Бибкод : 2018BGeo...15..809S . дои : 10.5194/bg-15-809-2018 .
- ^ Jump up to: а б с д Саттл, Калифорния (октябрь 2007 г.). «Морские вирусы – основные игроки глобальной экосистемы». Обзоры природы. Микробиология . 5 (10): 801–12. дои : 10.1038/nrmicro1750 . ПМИД 17853907 . S2CID 4658457 .
- ^ Вигингтон Ч., Зондереггер Д., Бруссаард К.П., Бьюкен А., Финке Дж.Ф., Фурман Дж.А. и др. (январь 2016 г.). «Повторное исследование взаимосвязи между численностью морских вирусов и микробных клеток» (PDF) . Природная микробиология . 1 (15024): 15024. doi : 10.1038/nmicrobiol.2015.24 . ПМИД 27572161 . S2CID 52829633 .
- ^ Брюссаард КП (2004). «Вирусный контроль популяций фитопланктона – обзор». Журнал эукариотической микробиологии . 51 (2): 125–38. дои : 10.1111/j.1550-7408.2004.tb00537.x . ПМИД 15134247 . S2CID 21017882 .
- ^ Роббинс Дж. (13 апреля 2018 г.). «Триллионы и триллионы вирусов падают с неба каждый день» . Нью-Йорк Таймс . Проверено 14 апреля 2018 г.
- ^ Рече И., Д'Орта Г., Младенов Н., Вингет Д.М., Саттл, Калифорния (апрель 2018 г.). «Скорость осаждения вирусов и бактерий над пограничным слоем атмосферы» . Журнал ISME . 12 (4): 1154–62. Бибкод : 2018ISMEJ..12.1154R . дои : 10.1038/s41396-017-0042-4 . ПМК 5864199 . ПМИД 29379178 .
- ^ Холл А.Дж., Джепсон П.Д., Гудман С.Дж., Харконен Т. (2006). «Вирус чумы чумы пауков в Северном и Европейском морях – данные и модели, природа и воспитание». Биологическая консервация . 131 (2): 221–29. Бибкод : 2006BCons.131..221H . doi : 10.1016/j.biocon.2006.04.008 .
- ^ ДеЛонг Дж.П., Ван Эттен Дж.Л., Аль-Амили З., Агаркова И.В., Дуниган Д.Д. (январь 2023 г.). «Потребление вирусов возвращает энергию в пищевые цепи» . Труды Национальной академии наук Соединенных Штатов Америки . 120 (1): e2215000120. Бибкод : 2023PNAS..12015000D . дои : 10.1073/pnas.2215000120 . ПМЦ 9910503 . ПМИД 36574690 . S2CID 255219850 .
- ^ Ирвинг М. (28 декабря 2022 г.). «Первое открытие «вировора»: организм, питающийся вирусами» . Новый Атлас. Архивировано из оригинала 29 декабря 2022 года . Проверено 29 декабря 2022 г.
- ^ Брокер Ф., Мёллинг К. (2019). «Что вирусы говорят нам об эволюции и иммунитете: помимо Дарвина?» . Анналы Нью-Йоркской академии наук . 1447 (1): 53–68. Бибкод : 2019NYASA1447...53B . дои : 10.1111/nyas.14097 . ПМК 6850104 . ПМИД 31032941 .
- ^ Лауэр, А. (2021). Вирусы – недооцененные факторы экологии и эволюции жизни. В: Херст, CJ (ред.) Микробы: краеугольный камень биосферы. Достижения в области экологической микробиологии, том 8. Springer, Cham. https://doi.org/10.1007/978-3-030-63512-1_28
- ^ Фортер П., Филипп Х (июнь 1999 г.). «Последний универсальный общий предок (LUCA), простой или сложный?». Биологический вестник . 196 (3): 373–75, обсуждение 375–77. дои : 10.2307/1542973 . JSTOR 1542973 . ПМИД 11536914 .
- ^ Лодиш Х., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «Вирусы: структура, функции и использование» . Молекулярно-клеточная биология (4-е изд.). Нью-Йорк: WH Freeman.
- ^ Мацузаки С., Рашель М., Утияма Дж., Сакураи С., Уджихара Т., Курода М. и др. (октябрь 2005 г.). «Бактериофаготерапия: обновленная терапия против бактериальных инфекционных заболеваний». Журнал инфекции и химиотерапии . 11 (5): 211–19. дои : 10.1007/s10156-005-0408-9 . ПМИД 16258815 . S2CID 8107934 .
- ^ Глеба Ю.Ю., Гирич А (2011). «Растительные вирусные векторы для экспрессии белков». Последние достижения в вирусологии растений . Кайстер Академик Пресс. ISBN 978-1-904455-75-2 .
- ^ Джефферсон А., Кадет В.Е., Хильшер А. (сентябрь 2015 г.). «Механизмы использования генетически модифицированных вирусов коровьей оспы для лечения рака». Критические обзоры по онкологии/гематологии . 95 (3): 407–16. doi : 10.1016/j.critrevonc.2015.04.001 . ПМИД 25900073 .
- ^ Каримхани С., Гонсалес Р., Деллавалле Р.П. (август 2014 г.). «Обзор новых методов лечения меланомы». Американский журнал клинической дерматологии . 15 (4): 323–37. дои : 10.1007/s40257-014-0083-7 . ПМИД 24928310 . S2CID 38864550 .
- ^ «FDA одобряет инъекционную иммунотерапию меланомы от Amgen» . Рейтер . 27 октября 2015 г. Проверено 24 января 2020 г.
- ^ Берк Дж., Ниева Дж., Борад М.Дж., Брайтбах К.Дж. (август 2015 г.). «Онколитические вирусы: перспективы клинического развития». Современное мнение в вирусологии . 13 :55–60. дои : 10.1016/j.coviro.2015.03.020 . ПМИД 25989094 .
- ^ Догич З (2016). «Нитевидные фаги как модельная система в физике мягких веществ» . Границы микробиологии . 7 : 1013. дои : 10.3389/fmicb.2016.01013 . ПМЦ 4927585 . ПМИД 27446051 .
- ^ Фишлехнер М., Донат Э. (2007). «Вирусы как строительные блоки материалов и устройств». Ангеванде Хеми . 46 (18): 3184–93. дои : 10.1002/anie.200603445 . ПМИД 17348058 .
- ^ Сото С.М., Блюм А.С., Вора Г.Дж., Лебедев Н., Мидор С.Э., Вон А.П. и др. (апрель 2006 г.). «Усиление флуоресцентного сигнала карбоцианиновых красителей с использованием сконструированных вирусных наночастиц» . Журнал Американского химического общества . 128 (15): 5184–89. дои : 10.1021/ja058574x . ПМИД 16608355 .
- ^ Блюм А.С., Сото СМ, Уилсон К.Д., Брауэр Т.Л., Поллак С.К., Шулл Т.Л. и др. (июль 2005 г.). «Спроектированный вирус как каркас для трехмерной самосборки на наноуровне». Маленький . 1 (7): 702–06. дои : 10.1002/smll.200500021 . ПМИД 17193509 .
- ^ Виолончель Дж., Пол А.В., Виммер Э. (август 2002 г.). «Химический синтез кДНК полиовируса: генерация инфекционного вируса в отсутствие естественной матрицы» . Наука . 297 (5583): 1016–18. Бибкод : 2002Sci...297.1016C . дои : 10.1126/science.1072266 . ПМИД 12114528 . S2CID 5810309 .
- ^ Коулман-младший, Папамихайль Д., Скиена С., Футчер Б., Виммер Э., Мюллер С. (июнь 2008 г.). «Аттенуация вируса за счет изменений в масштабе генома в смещении пары кодонов» . Наука . 320 (5884): 1784–87. Бибкод : 2008Sci...320.1784C . дои : 10.1126/science.1155761 . ПМК 2754401 . ПМИД 18583614 .
- ^ «База данных вирусных геномов НИЗ» . Ncbi.nlm.nih.gov . Проверено 28 июня 2021 г.
- ^ Жилинскас Р.А. (август 2017 г.). «Краткая история программ биологического оружия и использования патогенов животных в качестве боевых биологических агентов». Revue Scientifique et Technique (Международное бюро эпизоотий) . 36 (2): 415–22. дои : 10.20506/rst.36.2.2662 . ПМИД 30152475 .
- ^ Jump up to: а б с Артенштейн А.В., Грабенштайн Дж.Д. (октябрь 2008 г.). «Вакцины от оспы для биозащиты: необходимость и осуществимость» . Экспертная оценка вакцин . 7 (8): 1225–37. дои : 10.1586/14760584.7.8.1225 . ПМК 9709930 . ПМИД 18844596 . S2CID 33855724 .
