Теоретическая экология


Теоретическая экология — это научная дисциплина, посвященная изучению экологических систем с использованием теоретических методов, таких как простые концептуальные модели , математические модели , компьютерное моделирование и расширенный анализ данных . Эффективные модели улучшают понимание мира природы, показывая, как динамика популяций видов часто основана на фундаментальных биологических условиях и процессах. Кроме того, эта область направлена на объединение разнообразного диапазона эмпирических наблюдений, предполагая, что общие механистические процессы порождают наблюдаемые явления среди видов и экологических сред. Основываясь на биологически реалистичных предположениях, экологи-теоретики могут открыть новые, неинтуитивные представления о природных процессах. Теоретические результаты часто проверяются эмпирическими и наблюдательными исследованиями, раскрывающими силу теоретических методов как в предсказании, так и в понимании шумного и разнообразного биологического мира.
Область обширна и включает в себя основы прикладной математики, информатики, биологии, статистической физики, генетики, химии, эволюции и биологии сохранения. Теоретическая экология стремится объяснить широкий спектр явлений в науках о жизни, таких как рост и динамика населения , рыболовство, конкуренция , эволюционная теория, эпидемиология, поведение животных и групповая динамика, пищевые сети , экосистемы, пространственная экология и влияние климата. изменять .
Теоретическая экология еще больше выиграла от появления быстрых вычислительных мощностей, позволяющих анализировать и визуализировать крупномасштабное компьютерное моделирование экологических явлений. Важно отметить, что эти современные инструменты позволяют количественно прогнозировать влияние антропогенных изменений окружающей среды на разнообразные экологические явления, такие как: вторжения видов, изменение климата, влияние рыболовства и охоты на стабильность пищевых сетей и глобальный углеродный цикл .
Подходы к моделированию
[ редактировать ]Как и в большинстве других наук, математические модели составляют основу современной экологической теории.
- Феноменологические модели: выделяют функциональные формы и формы распределения из наблюдаемых закономерностей в данных, или исследователи выбирают функции и распределение, которые достаточно гибки, чтобы соответствовать закономерностям, которые они или другие (полевые или экспериментальные экологи) обнаружили в полевых условиях или посредством экспериментов. [3]
- Механистические модели: моделируют основные процессы напрямую с функциями и распределениями, основанными на теоретических рассуждениях об представляющих интерес экологических процессах. [3]
Экологические модели могут быть детерминистическими или стохастическими . [3]
- Детерминистические модели всегда развиваются одинаково, начиная с заданной отправной точки. [4] Они представляют собой среднее ожидаемое поведение системы, но не имеют случайных вариаций . Многие модели системной динамики являются детерминированными.
- Стохастические модели позволяют напрямую моделировать случайные возмущения, лежащие в основе экологических систем реального мира. Модели цепей Маркова являются стохастическими.
Виды можно моделировать в непрерывном или дискретном времени . [5]
- Непрерывное время моделируется с помощью дифференциальных уравнений .
- Дискретное время моделируется с помощью разностных уравнений . Они моделируют экологические процессы, которые можно описать как происходящие в дискретные временные интервалы. Матричная алгебра часто используется для исследования эволюции популяций с возрастной или стадийной структурой. Матрица Лесли , например, математически представляет дискретное изменение во времени возрастной структуры населения. [6] [7] [8]
Модели часто используются для описания реальных экологических процессов воспроизводства одного или нескольких видов. Их можно смоделировать с помощью стохастических ветвящихся процессов . Примерами могут служить динамика взаимодействующих популяций ( конкуренция хищников и мутуализм ), которую, в зависимости от интересующего вида, лучше всего моделировать в непрерывном или дискретном времени. Другие примеры таких моделей можно найти в области математической эпидемиологии , где динамические отношения, которые необходимо моделировать, представляют собой взаимодействия хозяина и патогена . [5]
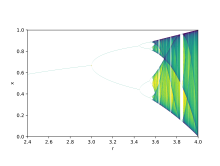
Теория бифуркаций используется для иллюстрации того, как небольшие изменения в значениях параметров могут привести к совершенно разным долгосрочным результатам. Этот математический факт можно использовать для объяснения резких экологических различий, возникающих в качественно очень похожих системах. [9] Логистические карты представляют собой полиномиальные отображения , и их часто называют архетипическими примерами того, как хаотическое поведение может возникнуть из очень простых нелинейных динамических уравнений. Карты были популяризированы в плодотворной статье эколога-теоретика Роберта Мэя , опубликованной в 1976 году . [10] Разностное уравнение предназначено для отражения двух эффектов воспроизводства и голода.
В 1930 году Р. А. Фишер опубликовал свою классическую «Генетическая теория естественного отбора» , в которой представил идею о том, что частотно-зависимая приспособленность привносит стратегический аспект в эволюцию , где выгоды для конкретного организма, возникающие в результате взаимодействия всех соответствующих организмов, — число жизнеспособных потомков этого организма. [11] В 1961 году Ричард Левонтин применил теорию игр к эволюционной биологии в своей книге «Эволюция и теория игр» . [12] за ним внимательно следил Джон Мейнард Смит , который в своей основополагающей статье 1972 года «Теория игр и эволюция боевых действий» [13] определил концепцию эволюционно устойчивой стратегии .
Поскольку экологические системы обычно нелинейны , их часто невозможно решить аналитически, и для получения разумных результатов необходимо использовать нелинейные, стохастические и вычислительные методы. Одним из классов вычислительных моделей, который становится все более популярным, являются модели на основе агентов . Эти модели могут моделировать действия и взаимодействия множества гетерогенных организмов там, где более традиционные аналитические методы не подходят. Прикладная теоретическая экология дает результаты, которые используются в реальном мире. Например, теория оптимального промысла опирается на методы оптимизации, разработанные в области экономики, информатики и исследований операций, и широко используется в рыболовстве . [14]
Популяционная экология
[ редактировать ]Популяционная экология — это раздел экологии , который занимается динамикой видов популяций и тем, как эти популяции взаимодействуют с окружающей средой . [15] Это исследование того, как размеры популяций видов, живущих вместе в группах, меняются во времени и пространстве, и было одним из первых аспектов экологии, которые изучались и моделировались математически.
Экспоненциальный рост
[ редактировать ]Самый простой способ моделирования популяционной динамики — предположить, что скорость роста популяции зависит только от размера популяции в данный момент и скорости роста организма на душу населения. Другими словами, если количество особей в популяции в момент времени t равно N(t), то скорость роста популяции определяется выражением:

где r — темп роста на душу населения или собственный темп роста организма. Его также можно описать как r = bd, где b и d — не зависящие от времени коэффициенты рождаемости и смертности на душу населения соответственно. Это первого порядка линейное дифференциальное уравнение можно решить, чтобы получить решение
- ,

траектория, известная как мальтузианский рост , в честь Томаса Мальтуса , который впервые описал ее динамику в 1798 году. Население, переживающее мальтузианский рост, следует экспоненциальной кривой, где N (0) — начальная численность населения. Популяция растет, когда r > 0, и снижается, когда r < 0. Модель наиболее применима в случаях, когда несколько организмов образовали колонию и быстро растут без каких-либо ограничений или ограничений, препятствующих их росту (например, бактерии, инокулированные в богатую среду). .
Логистический рост
[ редактировать ]Модель экспоненциального роста предполагает ряд допущений, многие из которых часто не верны. Например, на внутреннюю скорость роста влияет множество факторов, которые часто не являются неизменными во времени. Простая модификация экспоненциального роста состоит в том, чтобы предположить, что внутренние темпы роста зависят от размера населения. Это разумно: чем больше численность населения, тем меньше доступных ресурсов, что может привести к снижению рождаемости и повышению смертности. Следовательно, мы можем заменить неизменяемый во времени r на r'(t) = (b –a*N(t)) – (d + c*N(t)), где a и c – константы, модулирующие рождение и смерть. ставки в зависимости от популяции (например, внутривидовая конкуренция ). И a, и c будут зависеть от других факторов окружающей среды, которые мы можем пока считать постоянными в этой приближенной модели. Дифференциальное уравнение теперь имеет вид: [16]

Это можно переписать как: [16]

где r = bd и K = (bd)/(a+c).
Биологическое значение К становится очевидным, если рассматривать устойчивость равновесий системы. Константа K представляет собой пропускную способность населения. Равновесиями системы являются N = 0 и N = K. Если система линеаризована, можно видеть, что N = 0 — неустойчивое равновесие, а K — устойчивое равновесие. [16]
Структурированный рост населения
[ редактировать ]Другое предположение модели экспоненциального роста состоит в том, что все особи в популяции идентичны и имеют одинаковые вероятности выживания и воспроизводства. Это неверное предположение для видов со сложной историей жизни. Для учета этого можно модифицировать модель экспоненциального роста, отслеживая количество особей в разных возрастных группах (например, одно-, двух- и трехлетние дети) или на разных стадиях (подростки, полувзрослые и взрослые особи). ) отдельно и позволяя особям в каждой группе иметь свои собственные показатели выживаемости и воспроизводства. Общий вид этой модели:

где N t — вектор количества особей в каждом классе в момент времени t , а L — матрица , содержащая вероятность выживания и плодовитость для каждого класса. Матрица L называется матрицей Лесли для моделей с возрастной структурой и матрицей Лефковича для с поэтапной структурой . моделей [17]
Если значения параметров в L оцениваются на основе демографических данных о конкретной популяции, то структурированную модель можно затем использовать для прогнозирования, будет ли эта популяция расти или сокращаться в долгосрочной перспективе, и каково будет ожидаемое возрастное распределение внутри популяции . . Это было сделано для ряда видов, включая морских черепах и китов . [18] [19]
Экология сообщества
[ редактировать ]Экологическое сообщество – это группа трофически сходных симпатрических видов, которые фактически или потенциально конкурируют на определенной территории за одни и те же или сходные ресурсы. [20] Взаимодействия между этими видами составляют первые шаги в анализе более сложной динамики экосистем. Эти взаимодействия формируют распределение и динамику видов. Из этих взаимодействий хищничество является одним из наиболее распространенных видов деятельности населения. [21] В самом общем смысле хищничество включает взаимодействия хищник-жертва, хозяин-патоген и хозяин-паразитоид.

Взаимодействие хищник-жертва
[ редактировать ]Взаимодействия хищник-жертва демонстрируют естественные колебания в популяциях как хищника, так и жертвы. [21] В 1925 году американский математик Альфред Дж. Лотка в своей книге по биоматематике разработал простые уравнения взаимодействия хищник-жертва. [22] В следующем году итальянский математик Вито Вольтерра провёл статистический анализ уловов рыбы в Адриатике. [23] и независимо разработали те же уравнения. [24] Это одна из самых ранних и наиболее признанных экологических моделей, известная как модель Лотки-Вольтерры :


где N — жертва, а P — размеры популяции хищников, r — скорость роста добычи, принимаемая экспоненциальной в отсутствие каких-либо хищников, α — уровень смертности жертв при хищничестве на душу населения (также называемый «коэффициентом нападения»). ), c — эффективность превращения добычи в хищника, а d — экспоненциальный уровень смертности хищников в отсутствие какой-либо добычи.
Первоначально Вольтерра использовал эту модель для объяснения колебаний численности рыб и акул после рыболовства сокращения во время Первой мировой войны . Однако впоследствии уравнения стали применяться в более широком смысле. [25] Другие примеры этих моделей включают модель Лотки-Вольтерры зайца -беляка и канадской рыси в Северной Америке. [26] любое моделирование инфекционных заболеваний, таких как недавняя вспышка атипичной пневмонии [27] и биологический контроль калифорнийской красной чешуи путем введения ее Aphytis паразитоида melinus. . [28]
Надежной и простой альтернативой модели «хищник-жертва» Лотки-Вольтерры и ее общим обобщениям, зависящим от добычи, является модель, зависящая от соотношения, или модель Ардити-Гинзбурга . [29] Эти две модели являются крайностями спектра моделей вмешательства хищников. По мнению авторов альтернативной точки зрения, данные показывают, что истинные взаимодействия в природе настолько далеки от крайности Лотки-Вольтерры в интерференционном спектре, что модель можно просто отбросить как неверную. Они намного ближе к экстремуму, зависящему от отношения, поэтому, если нужна простая модель, можно использовать модель Ардити – Гинзбурга в качестве первого приближения. [30]
Взаимодействие хозяина и патогена
[ редактировать ]Второе взаимодействие, взаимодействие хозяина и патогена , отличается от взаимодействия хищник-жертва тем, что патогены намного меньше, имеют гораздо более быстрое время генерации и требуют хозяина для размножения. Следовательно, в моделях хозяин-патоген отслеживается только популяция хозяина. Обычно используются компартментальные модели, которые распределяют популяцию хозяина по таким группам, как восприимчивая, инфицированная и выздоровевшая (SIR). [31]
Взаимодействие хозяина и паразитоида
[ редактировать ]Третье взаимодействие, взаимодействие хозяина и паразитоида , можно проанализировать с помощью модели Николсона-Бейли , которая отличается от моделей Лотки-Вольтерра и SIR тем, что она дискретна во времени. Эта модель, как и модель Лотки-Вольтерры, явно отслеживает обе популяции. Обычно в общей форме он гласит:
![{\displaystyle N_{t+1}=\lambda \ N_{t}\ [1-f(N_{t},P_{t})]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/70346762c7b4aebcd36578c549923bd804870171)

где f(N t , P t ) описывает вероятность заражения (обычно распределение Пуассона ), λ — скорость роста хозяев на душу населения в отсутствие паразитоидов, а c — эффективность конверсии, как в случае Лотки-Вольтерра. модель. [21]
Конкуренция и мутуализм
[ редактировать ]При изучении популяций двух видов система уравнений Лотки-Вольтерра широко использовалась для описания динамики поведения между двумя видами N 1 и N 2 . Примеры включают отношения между D. Discoiderum и E. coli , [32] а также теоретический анализ поведения системы. [33]


Коэффициенты r дают «базовую» скорость роста каждого вида, а коэффициенты K соответствуют несущей способности. Однако что действительно может изменить динамику системы, так это α-члены. Они описывают характер взаимоотношений между двумя видами. Когда α 12 отрицательна, это означает, что N 2 оказывает негативное влияние на N 1 , конкурируя с ним, охотясь на него или используя любое количество других возможностей. Однако когда α 12 положителен, это означает, что N 2 оказывает положительное влияние на N 1 посредством некоторого рода взаимного взаимодействия между ними. Когда оба α 12 и α 21 отрицательны, отношения описываются как конкурентные . В этом случае каждый вид отвлекает друг друга, потенциально создавая чрезмерную конкуренцию за скудные ресурсы. Когда оба α 12 и α 21 положительны, отношения становятся отношениями мутуализма . В этом случае каждый вид приносит пользу другому, так что присутствие одного способствует росту популяции другого.
- См. «Конкурентные уравнения Лотки – Вольтерра» для дальнейшего расширения этой модели.
Нейтральная теория
[ редактировать ]Единая нейтральная теория — это гипотеза, предложенная Стивеном П. Хаббеллом в 2001 году. [20] Гипотеза направлена на объяснение разнообразия и относительного обилия видов в экологических сообществах, хотя, как и другие нейтральные теории в экологии, гипотеза Хаббелла предполагает, что различия между членами экологического сообщества трофически сходных видов «нейтральны» или не имеют отношения к их успеху. . Нейтральность означает, что на данном трофическом уровне пищевой цепи виды эквивалентны по уровням рождаемости, смертности, скорости расселения и видообразования, если измерять их в расчете на душу населения. [34] Это означает, что биоразнообразие возникает случайным образом, поскольку каждый вид совершает случайное блуждание . [35] Это можно считать нулевой гипотезой теории ниш . Гипотеза вызвала споры, и некоторые авторы считают ее более сложной версией других нулевых моделей, которые лучше соответствуют данным.
разрешены сложные экологические взаимодействия Согласно единой нейтральной теории, между отдельными членами экологического сообщества (такие как конкуренция и сотрудничество), при условии, что все люди подчиняются одним и тем же правилам. Асимметричные явления, такие как паразитизм и хищничество , исключены из технического задания; но совместные стратегии, такие как роение , и негативное взаимодействие, такое как конкуренция за ограниченное количество еды или света, разрешены, при условии, что все люди ведут себя одинаково. Теория делает прогнозы, которые имеют значение для управления биоразнообразием , особенно для управления редкими видами. Он предсказывает существование фундаментальной константы биоразнообразия, условно обозначаемой θ , которая, по-видимому, управляет видовым богатством в самых разных пространственных и временных масштабах.
Хаббелл основывался на более ранних нейтральных концепциях, включая Макартура и Уилсона. теорию биогеографии островов [20] и Гулда о симметрии и нулевых моделях. концепции [34]
Пространственная экология
[ редактировать ]Биогеография
[ редактировать ]Биогеография – это наука о распространении видов в пространстве и времени. Его цель - выяснить, где живут организмы, в какой численности и почему они встречаются (или нет) в определенной географической области.
Наиболее остро биогеография наблюдается на островах, что привело к развитию раздела островная биогеография . Эти среды обитания часто являются более управляемыми областями исследования, поскольку они более компактны, чем более крупные экосистемы на материке. В 1967 году Роберт Макартур и Э.О. Уилсон опубликовали «Теорию биогеографии островов» . Это показало, что видовое богатство на территории можно предсказать с точки зрения таких факторов, как площадь обитания, уровень иммиграции и скорость вымирания. [36] Теория считается одной из основ экологической теории. [37] Применение теории островной биогеографии к фрагментам среды обитания стимулировало развитие областей природоохранной биологии и ландшафтной экологии . [38]
r/K-теория отбора
[ редактировать ]Концепция популяционной экологии — это теория отбора r/K, одна из первых прогностических моделей в экологии, используемая для объяснения эволюции жизненного цикла . Предпосылка, лежащая в основе модели отбора r/K, заключается в том, что давление естественного отбора меняется в зависимости от плотности населения . Например, когда остров впервые колонизирован, плотность населения невелика. Первоначальное увеличение численности населения не ограничивается конкуренцией, оставляя изобилие доступных ресурсов для быстрого роста населения. На этих ранних стадиях роста населения действуют независимые от плотности силы естественного отбора, который называется r -отбором. Поскольку население становится более перенаселенным, оно приближается к пропускной способности острова, что вынуждает людей более жестко конкурировать за меньшее количество доступных ресурсов. В условиях скученности популяция испытывает зависящие от плотности силы естественного отбора, называемые K -отбором. [39] [40]

Теория ниши
[ редактировать ]Метапопуляции
[ редактировать ]Пространственный анализ экологических систем часто показывает, что предположения, которые справедливы для пространственно однородных популяций – и, по сути, интуитивно понятны – могут больше не быть верными, когда рассматриваются мигрирующие субпопуляции, перемещающиеся с одного участка на другой. [42] В простой одновидовой формулировке субпопуляция может занять участок, переместиться с одного участка на другой пустой участок или вымереть, оставив после себя пустой участок. В таком случае долю занятых участков можно представить как

где m — скорость колонизации , а e — скорость вымирания . [43] В этой модели, если e < m, устойчивое значение p равно 1 – (e/m), тогда как в другом случае все участки в конечном итоге останутся пустыми. Эту модель можно усложнить путем добавления другого вида несколькими различными способами, включая, помимо прочего, теоретико-игровые подходы, взаимодействие хищник-жертва и т. д. Для простоты мы рассмотрим здесь расширение предыдущей одновидовой системы. Обозначим долю участков, занятых первой популяцией, как p 1 , а вторую — как p 2 . Затем,


В этом случае, если e слишком велико, p 1 и p 2 будут равны нулю в установившемся состоянии. Однако при умеренной скорости вымирания р 1 и р 2 могут стабильно сосуществовать. Стационарное значение p 2 определяется выражением

(p* 1 можно вывести из симметрии). Если e равно нулю, динамика системы благоприятствует виду, который лучше колонизируется (т. е. имеет более высокое значение m). Это приводит к очень важному результату в теоретической экологии, известному как гипотеза промежуточного нарушения , согласно которой биоразнообразие (количество видов, сосуществующих в популяции) максимизируется, когда нарушение (которое здесь является показателем e) не слишком велико или слишком низкий, но на промежуточном уровне. [44]
Форму дифференциальных уравнений, используемых в этом упрощенном подходе к моделированию, можно изменить. Например:
- Колонизация может зависеть от p линейно (m*(1-p)) в отличие от нелинейного режима m*p*(1-p), описанного выше. Этот способ размножения вида называется «дождем из пропагул», при котором в каждом поколении в популяцию попадает множество новых особей. В таком сценарии устойчивое состояние, при котором численность населения равна нулю, обычно нестабильно. [45]
- Угасание может зависеть нелинейно от p (e*p*(1-p)) в отличие от линейного (e*p) режима, описанного выше. Это называется « эффектом спасения », и при таком режиме снова труднее добиться вымирания популяции. [45]
Модель также может быть расширена до комбинаций четырех возможных линейных или нелинейных зависимостей колонизации и вымирания от p, которые более подробно описаны в . [46]
Экосистема экология
[ редактировать ]Введение новых элементов, как биотических , так и абиотических , в экосистемы может иметь разрушительные последствия. В некоторых случаях это приводит к экологическому коллапсу , трофическим каскадам и гибели многих видов внутри экосистемы. Абстрактное понятие экологического здоровья пытается измерить устойчивость и способность экосистемы к восстановлению; т.е. насколько экосистема далека от своего устойчивого состояния. Однако зачастую экосистемы восстанавливаются из-за разрушительного агента. Разница между коллапсом и отскоком зависит от токсичности введенного элемента и устойчивости исходной экосистемы.
Если экосистемы управляются в первую очередь случайными процессами, посредством которых их последующее состояние будет определяться как предсказуемыми, так и случайными действиями, они могут быть более устойчивыми к внезапным изменениям, чем каждый вид в отдельности. В отсутствие баланса природы видовой состав экосистем претерпит изменения, которые будут зависеть от характера изменений, но полный экологический коллапс, вероятно, будет редким событием. В 1997 году Роберт Уланович использовал инструменты теории информации для описания структуры экосистем, подчеркивая взаимную информацию (корреляции) в изучаемых системах. Опираясь на эту методологию и предыдущие наблюдения за сложными экосистемами, Уланович описывает подходы к определению уровней стресса в экосистемах и прогнозированию реакций системы на определенные типы изменений в их условиях (например, увеличение или уменьшение потока энергии) и эвтрофикацию . [47]
Ecopath — это бесплатный пакет программного обеспечения для моделирования экосистем, первоначально разработанный NOAA и широко используемый в управлении рыболовством в качестве инструмента для моделирования и визуализации сложных взаимосвязей, существующих в реальных морских экосистемах.
Пищевые сети
[ редактировать ]Пищевые сети обеспечивают основу, в которой может быть организована сложная сеть взаимодействий хищник-жертва. Модель пищевой сети представляет собой сеть пищевых цепей . Каждая пищевая цепь начинается с первичного производителя или автотрофа — организма, например растения, который способен производить собственную пищу. Следующим в цепочке идет организм, питающийся первичным производителем, и цепочка продолжается, таким образом, в виде цепочки сменяющих друг друга хищников. Организмы в каждой цепи группируются по трофическим уровням в зависимости от того, сколько звеньев они удалены от первичных продуцентов. Длина цепи, или трофический уровень, является мерой количества видов, встречающихся при переходе энергии или питательных веществ от растений к высшим хищникам. [48] Энергия пищи течет от одного организма к другому, к следующему и так далее, при этом некоторая энергия теряется на каждом уровне. На данном трофическом уровне может существовать один вид или группа видов с одинаковыми хищниками и жертвами. [49]
В 1927 году Чарльз Элтон опубликовал влиятельный анализ использования пищевых сетей, в результате которого они стали центральной концепцией экологии. [50] В 1966 году интерес к пищевым цепям возрос после Роберта Пейна , которое позволило предположить, что сложность пищевых сетей является ключом к поддержанию видового разнообразия и экологической стабильности. экспериментального и описательного исследования приливных берегов [51] Это и другие открытия побудили многих экологов-теоретиков, в том числе сэра Роберта Мэя и Стюарта Пимма , изучить математические свойства пищевых сетей. Согласно их анализу, сложные пищевые сети должны быть менее стабильными, чем простые. [1] : 75–77 [2] : 64 Очевидный парадокс между сложностью пищевых сетей, наблюдаемых в природе, и математической хрупкостью моделей пищевых сетей в настоящее время является областью интенсивных исследований и дискуссий. Парадокс может быть частично обусловлен концептуальными различиями между устойчивостью пищевой сети и равновесной стабильностью пищевой сети. [1] [2]
Системная экология
[ редактировать ]Системную экологию можно рассматривать как приложение общей теории систем к экологии. Он использует целостный и междисциплинарный подход к изучению экологических систем, и особенно экосистем. Системная экология особенно озабочена тем, как вмешательство человека может повлиять на функционирование экосистем. Как и другие области теоретической экологии, она использует и расширяет концепции термодинамики и разрабатывает другие макроскопические описания сложных систем. Он также учитывает потоки энергии через различные трофические уровни экологических сетей. Системная экология также учитывает внешнее влияние экологической экономики , которое обычно не учитывается в экологии экосистем. [52] По большей части системная экология является подразделом экологии экосистем.
Экофизиология
[ редактировать ]Это исследование того, как «окружающая среда, как физическая, так и биологическая, взаимодействует с физиологией организма. Оно включает в себя влияние климата и питательных веществ на физиологические процессы как у растений, так и у животных, и уделяет особое внимание тому, как масштабируются физиологические процессы». с размером организма». [53] [54]
Поведенческая экология
[ редактировать ]Поведение роя
[ редактировать ]
Стайное поведение — это коллективное поведение, демонстрируемое животными одинакового размера, которые собираются вместе, возможно, слоняются в одном и том же месте или, возможно, мигрируют в каком-то направлении. Стайное поведение обычно проявляется у насекомых, но оно также встречается при стаях птиц, стайном поведении рыб и стадном поведении четвероногих. Это сложное эмерджентное поведение, которое возникает, когда отдельные агенты следуют простым правилам поведения.
Недавно был открыт ряд математических моделей, объясняющих многие аспекты эмерджентного поведения. Роевые алгоритмы следуют лагранжевому подходу или эйлерову подходу. [56] Эйлеров подход рассматривает рой как поле , работая с плотностью роя и получая средние свойства поля. Это гидродинамический подход, который может быть полезен для моделирования общей динамики крупных роев. [57] [58] [59] Однако большинство моделей работают с лагранжевым подходом, который представляет собой агентную модель, отслеживающую отдельных агентов (точек или частиц), составляющих рой. Модели отдельных частиц могут отслеживать информацию о направлении и расстоянии, которая теряется в эйлеровом подходе. [56] [60] Примеры включают оптимизацию колонии муравьев , самодвижущиеся частицы и оптимизацию роя частиц .
На клеточном уровне отдельные организмы также демонстрировали роевое поведение. В децентрализованных системах люди действуют на основании собственных решений без общего руководства. Исследования показали, что отдельные прилипшие трихоплаксы ведут себя как самодвижущиеся частицы (СПП) и в совокупности демонстрируют фазовый переход от упорядоченного движения к неупорядоченным движениям. [61] Раньше считалось, что соотношение поверхности и объема ограничивает размер животных в эволюционной игре. Учитывая коллективное поведение особей, было высказано предположение, что порядок является еще одним ограничивающим фактором. Было показано, что центральная нервная система жизненно важна для крупных многоклеточных животных на эволюционном пути.
Синхронизация
[ редактировать ]Светлячок Photinus carolinus синхронизирует свои сияющие частоты в коллективной обстановке. По отдельности видимых закономерностей перепрошивки нет. В групповой обстановке периодичность проявляется в сияющем узоре. [62] Сосуществование синхронизации и асинхронности вспышек в системе, состоящей из множества светлячков, можно охарактеризовать как состояния химеры. Синхронизация могла произойти спонтанно. [63] Агентная модель оказалась полезной для описания этого уникального явления. Вспышки отдельных светлячков можно было рассматривать как осцилляторы, а модели глобальной связи были аналогичны тем, которые используются в физике конденсированного состояния .
Эволюционная экология
[ редактировать ]Британский биолог Альфред Рассел Уоллес наиболее известен тем, что независимо предложил теорию эволюции в результате естественного отбора , которая побудила Чарльза Дарвина опубликовать свою собственную теорию. В своей знаменитой статье 1858 года Уоллес предложил естественный отбор как своего рода механизм обратной связи, который поддерживает адаптацию видов и разновидностей к окружающей среде. [64]
Действие этого принципа совершенно аналогично действию центробежного регулятора паровой машины, который проверяет и исправляет любые отклонения почти до того, как они станут очевидными; и точно так же ни один несбалансированный недостаток в животном мире не может когда-либо достичь сколько-нибудь заметной величины, потому что он даст о себе знать на самом первом этапе, сделав существование трудным и почти наверняка вскоре последующим вымиранием. [65]
Кибернетик . и антрополог Грегори Бейтсон заметил в 1970-х годах, что, хотя Уоллес написал это только в качестве примера, «вероятно, сказал самую сильную вещь, которая была сказана в 19 веке» [66] Впоследствии связь между естественным отбором и теорией систем стала областью активных исследований. [64]
Другие теории
[ редактировать ]В отличие от предыдущих экологических теорий, которые считали наводнения катастрофическими явлениями, концепция пульсации речных паводков утверждает, что годовая пульсация паводков является наиболее важным аспектом и наиболее биологически продуктивной особенностью речной экосистемы . [67] [68]
История
[ редактировать ]Теоретическая экология опирается на новаторские работы Дж. Эвелин Хатчинсон и его учеников. Братья Х. Т. Одум и Е. П. Одум общепризнаны основоположниками современной теоретической экологии. Роберт Макартур привнес теорию в экологию сообществ . Дэниел Симберлофф был учеником Э.О. Уилсона , с которым Макартур сотрудничал над «Теорией островной биогеографии» , плодотворной работой в развитии теоретической экологии. [69]
Симберлофф добавил статистической строгости к экспериментальной экологии и был ключевой фигурой в дебатах SLOSS о том, предпочтительнее ли защищать один большой или несколько небольших заповедников. [70] Это привело к тому, что сторонники правил общественных собраний Джареда Даймонда стали защищать свои идеи с помощью анализа нейтральной модели. [70] Симберлофф также сыграл ключевую роль в (все еще продолжающихся) дебатах о полезности коридоров для соединения изолированных резерваций.
Стивен П. Хаббелл и Майкл Розенцвейг объединили теоретические и практические элементы в работах, которые расширили теорию биогеографии островов Макартура и Уилсона: Хаббелл с его Единой нейтральной теорией биоразнообразия и биогеографии и Розенцвейг с его видовым разнообразием в пространстве и времени.
Теоретические и математические экологи
[ редактировать ]Можно провести предварительное различие между экологами-математиками, экологами, применяющими математику к экологическим проблемам, и математиками, разрабатывающими саму математику, возникающую из экологических проблем.
Некоторых известных экологов-теоретиков можно найти в следующих категориях:
Журналы
[ редактировать ]См. также
[ редактировать ]- Эффект бабочки
- Сложная системная биология
- Теория экологических систем
- Модель экосистемы
- Интегроразностное уравнение - широко используется для моделирования расселения и роста популяций.
- Ограничение сходства
- Математическая биология
- Динамика населения
- Популяционное моделирование
- Количественная экология
- Закон Тейлора
- Теоретическая биология
Ссылки
[ редактировать ]- ^ Jump up to: а б с Мэй Р.М. (2001) Стабильность и сложность модельных экосистем Princeton University Press, переиздание издания 1973 года с новым предисловием. ISBN 978-0-691-08861-7 .
- ^ Jump up to: а б с Пимм С.Л. (2002) пищевых сетей , перепечатка издания 1982 года с новым предисловием. Издательство Чикагского университета ISBN 978-0-226-66832-1 .
- ^ Jump up to: а б с Болкер Б.М. (2008) Экологические модели и данные в издательстве R Princeton University Press, страницы 6–9. ISBN 978-0-691-12522-0 .
- ^ Сугихара Г., Мэй Р. (1990). «Нелинейное прогнозирование как способ отличить хаос от ошибки измерения во временных рядах» (PDF) . Природа . 344 (6268): 734–41. Бибкод : 1990Natur.344..734S . дои : 10.1038/344734a0 . ПМИД 2330029 . S2CID 4370167 . Архивировано из оригинала (PDF) 14 августа 2011 г. Проверено 13 мая 2011 г.
- ^ Jump up to: а б Соэтарт К. и Герман П.М.Дж. (2009) Практическое руководство по экологическому моделированию Springer. ISBN 978-1-4020-8623-6 .
- ^ Грант WE (1986) Системный анализ и моделирование в науках о дикой природе и рыболовстве. Уайли, Университет Миннесоты, стр. 223. ISBN 978-0-471-89236-6 .
- ^ Джопп Ф (2011) Моделирование сложной экологической динамики Springer, стр. 122. ISBN 978-3-642-05028-2 .
- ^ Берк А.Р. (2005) Новые тенденции в экологических исследованиях Nova Publishers, стр. 136. ISBN 978-1-59454-379-1 .
- ^ Ма Т и Ван С (2005) Теория и приложения бифуркации World Scientific. ISBN 978-981-256-287-6 .
- ^ Мэй, Роберт (1976). Теоретическая экология: принципы и приложения . Научные издательства Блэквелла. ISBN 978-0-632-00768-4 .
- ^ Фишер, Р.А. (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон пресс.
- ^ Р. К. Левонтин (1961). «Эволюция и теория игр». Журнал теоретической биологии . 1 (3): 382–403. Бибкод : 1961JThBi...1..382L . дои : 10.1016/0022-5193(61)90038-8 . ПМИД 13761767 .
- ^ Джон Мейнард Смит (1974). «Теория игр и эволюция конфликтов животных» (PDF) . Журнал теоретической биологии . 47 (1): 209–21. Бибкод : 1974JThBi..47..209M . дои : 10.1016/0022-5193(74)90110-6 . ПМИД 4459582 .
- ^ Суприатна А.К. (1998) Теория оптимального сбора урожая для метапопуляций хищник-жертва Университет Аделаиды, факультет прикладной математики.
- ^ Одум, Юджин П. (1959). Основы экологии (Второе изд.). Филадельфия и Лондон: WB Saunders Co. p. 546 стр . ISBN 9780721669410 . OCLC 554879 .
- ^ Jump up to: а б с Мосс Р., Уотсон А. и Олласон Дж. (1982) Динамика популяции животных Springer, стр. 52–54. ISBN 978-0-412-22240-5 .
- ^ Хэл Касвелл (2001). Матричные популяционные модели: построение, анализ и интерпретация . Синауэр.
- ^ Д.Т.Крус, Л.Б. Краудер, Х.Касвелл (1987). «Поэтапная популяционная модель морских черепах и последствия для сохранения». Экология . 68 (5): 1412–1423. Бибкод : 1987Ecol...68.1412C . дои : 10.2307/1939225 . JSTOR 1939225 . S2CID 16608658 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ М. Фудзивара; Х. Касвелл (2001). «Демография находящихся под угрозой исчезновения североатлантических китов». Природа . 414 (6863): 537–541. Бибкод : 2001Natur.414..537F . дои : 10.1038/35107054 . ПМИД 11734852 . S2CID 4407832 .
- ^ Jump up to: а б с Хаббелл, СП (2001). «Единая нейтральная теория биоразнообразия и биогеографии (МПБ-32)» . Архивировано из оригинала 18 июля 2011 г. Проверено 16 декабря 2010 г.
- ^ Jump up to: а б с Бонсолл, Майкл Б.; Хассел, Майкл П. (2007). «Взаимодействие хищник-жертва». В мае Роберт; Маклин, Анджела (ред.). Теоретическая экология: принципы и приложения (3-е изд.). Издательство Оксфордского университета. стр. 46–61.
- ^ Лотка, AJ, Элементы физической биологии , Уильямс и Уилкинс , (1925)
- ^ Гоэл, Н.С. и др., «О Вольтерре и других нелинейных моделях взаимодействующих популяций», Academic Press Inc. , (1971).
- ^ Вольтерра, V (1926). «Вариации и колебания численности особей сожительствующих видов животных». Мем. Линкейцы Рим . 2 :31–113.
- ^ Бегон, М.; Харпер, Дж.Л.; Таунсенд, ЧР (1988). Экология: Индивиды, популяции и сообщества . Blackwell Scientific Publications Inc., Оксфорд, Великобритания.
- ^ К.С. Элтон (1924). «Периодические колебания численности животных – Их причины и последствия» . Журнал экспериментальной биологии . 2 (1): 119–163. дои : 10.1242/jeb.2.1.119 .
- ^ Липсич М., Коэн Т., Купер Б., Робинс Дж.М., Ма С., Джеймс Л., Гопалакришна Г., Чу С.К., Тан CC, Самор М.Х., Фисман Д., Мюррей М. (2003). «Динамика передачи и контроль тяжелого острого респираторного синдрома» . Наука . 300 (5627): 1966–70. Бибкод : 2003Sci...300.1966L . дои : 10.1126/science.1086616 . ПМК 2760158 . ПМИД 12766207 .
- ^ Джон Д. Рив; Уильям В. Мердок (1986). «Биологический контроль с помощью паразитоида Aphytis melinus и стабильность популяции по Калифорнийской красной шкале». Журнал экологии животных . 55 (3): 1069–1082. Бибкод : 1986JAnEc..55.1069R . дои : 10.2307/4434 . JSTOR 4434 .
- ^ Ардити, Р.; Гинзбург, Л.Р. (1989). «Связь в динамике хищник-жертва: зависимость от соотношения» (PDF) . Журнал теоретической биологии . 139 (3): 311–326. Бибкод : 1989JThBi.139..311A . дои : 10.1016/s0022-5193(89)80211-5 . Архивировано из оригинала (PDF) 4 марта 2016 г. Проверено 26 июня 2013 г.
- ^ Ардити, Р. и Гинзбург, Л.Р. (2012) Как взаимодействуют виды: изменение стандартного взгляда на трофическую экологию Oxford University Press. ISBN 9780199913831 .
- ^ Гренфелл, Брайан; Килинг, Мэтью (2007). «Динамика инфекционных заболеваний». В мае Роберт; Маклин, Анджела (ред.). Теоретическая экология: принципы и приложения (3-е изд.). Издательство Оксфордского университета. стр. 132–147.
- ^ Его Величество Цучия; Дж. Ф. Дрейк; Дж. Л. Йост и А. Г. Фредриксон (1972). «Взаимодействие хищника и жертвы Dictyostelium discoideum и Escherichia coli в непрерывной культуре1» . Журнал бактериологии . 110 (3): 1147–53. дои : 10.1128/JB.110.3.1147-1153.1972 . ПМЦ 247538 . ПМИД 4555407 .
- ^ Такеучи, Ю. (1989). «Теория кооперативных систем и глобальная устойчивость диффузионных моделей». Acta Applicandae Mathematicae . 14 (1–2): 49–57. дои : 10.1007/BF00046673 . S2CID 189902519 .
- ^ Jump up to: а б Хаббелл, СП (2005). «Нейтральная теория биоразнообразия и биогеографии и Стивен Джей Гулд». Палеобиология . 31 : 122–123. doi : 10.1666/0094-8373(2005)031[0122:TNTOBA]2.0.CO;2 . S2CID 86271294 .
- ^ МакГилл, Би Джей (2003). «Проверка единой нейтральной теории биоразнообразия». Природа . 422 (6934): 881–885. Бибкод : 2003Natur.422..881M . дои : 10.1038/nature01583 . ПМИД 12692564 . S2CID 1627734 .
- ^ Макартур Р.Х. и Уилсон Э.О. (1967) Теория островной биогеографии
- ^ Винс, Джей-Джей; Донохью, MJ (2004). «Историческая биогеография, экология и видовое богатство» (PDF) . Тенденции экологии и эволюции . 19 (12): 639–644. дои : 10.1016/j.tree.2004.09.011 . ПМИД 16701326 .
- ^ Это относится к британским и американским ученым; Ландшафтная экология имеет четкое происхождение среди европейских ученых.
- ^ Впервые представлено в книге Макартура и Уилсона (1967), получившей заметное упоминание в истории и теоретической науке об экологии, «Теория биогеографии островов».
- ^ Резник, Д.; Брайант, MJ; Башей, Ф. (2002). «Возвращение к r- и K-отбору: роль регуляции численности населения в эволюции жизненного цикла» (PDF) . Экология . 83 (6): 1509–1520. doi : 10.1890/0012-9658(2002)083[1509:RAKSRT]2.0.CO;2 . ISSN 0012-9658 . Архивировано из оригинала (PDF) 30 декабря 2010 г. Проверено 5 января 2015 г.
- ^ Гевин В. (2006). «За пределами нейтралитета — экология находит свою нишу» . ПЛОС Биол . 4 (8): 1306–1310. doi : 10.1371/journal.pbio.0040278 . ПМК 1543692 . ПМИД 16895443 .
- ^ Хански I (1999) Экология метапопуляции Oxford University Press. ISBN 978-0-19-854065-6 .
- ^ Хански И., Гилпин М. (1991). «Динамика метапопуляции: краткая история и концептуальная область» (PDF) . Биологический журнал Линнеевского общества . 42 (1–2): 3–16. дои : 10.1111/j.1095-8312.1991.tb00548.x .
- ^ Кокс CB и Мур PD (2010) Биогеография: экологический и эволюционный подход Джон Уайли и сыновья, стр. 146. ISBN 978-0-470-63794-4 .
- ^ Jump up to: а б Вандермеер Дж. Х. и Голдберг Д. Е. (2003) Популяционная экология: основные принципы Princeton University Press, стр. 175–176. ISBN 978-0-691-11441-5 .
- ^ Илкка Хански (1982). «Динамика регионального распространения: гипотеза основных и спутниковых видов». Ойкос . 38 (2): 210–221. Бибкод : 1982Oikos..38..210H . дои : 10.2307/3544021 . JSTOR 3544021 .
- ^ Роберт Уланович (). Экология, восходящая перспектива . Колумбийский университет. Нажимать. ISBN 0-231-10828-1 .
- ^ Пост, DM (1993). «Длинная и короткая длина пищевой цепи». Тенденции экологии и эволюции . 17 (6): 269–277. дои : 10.1016/S0169-5347(02)02455-2 .
- ^ Джерри Боброу, доктор философии; Стивен Фишер (2009). CliffsNotes CSET: Несколько предметов (2-е изд.). Джон Уайли и сыновья. п. 283. ИСБН 978-0-470-45546-3 .
- ^ Элтон CS (1927) Экология животных. Переиздано в 2001 году. Издательство Чикагского университета.
- ^ Пейн РТ (1966). «Сложность пищевой сети и видовое разнообразие». Американский натуралист . 100 (910): 65–75. дои : 10.1086/282400 . S2CID 85265656 .
- ^ Р.Л. Китчинг, Системная экология , University of Queensland Press, 1983, стр.9.
- ^ Экофизиология , Природа . Дата обращения: 9 августа 2017 г.
- ^ Питерс, Р.Х. (1986) Экологические последствия размера тела Издательство Кембриджского университета. ISBN 9780521288866
- ^ Бхаттачарья К. и Вичек Т. (2010) «Коллективное принятие решений в сплоченных стадах»
- ^ Jump up to: а б Ли YX; Люкман Р; Эдельштейн-Кешет Л (2007). «Минимальные механизмы формирования школ самодвижущихся частиц» (PDF) . Физика D: Нелинейные явления . 237 (5): 699–720. Бибкод : 2008PhyD..237..699L . дои : 10.1016/j.physd.2007.10.009 . Архивировано из оригинала (PDF) 1 октября 2011 г.
- ^ Тонер Дж и Ту Ю (1995) «Дальний порядок в двумерной модели xy: как птицы летают вместе» Physical Revue Letters, 75 (23) (1995), 4326–4329.
- ^ Топаз С, Бертоцци А (2004). «Схемы роения в двумерной кинематической модели биологических групп». SIAM J Appl Math . 65 (1): 152–174. Бибкод : 2004APS..MAR.t9004T . CiteSeerX 10.1.1.88.3071 . дои : 10.1137/S0036139903437424 . S2CID 18468679 .
- ^ Топаз С., Бертоцци А., Льюис М. (2006). «Нелокальная континуальная модель биологической агрегации». Бычья Математика Биол . 68 (7): 1601–1623. arXiv : q-bio/0504001 . дои : 10.1007/s11538-006-9088-6 . ПМИД 16858662 . S2CID 14750061 .
- ^ Каррильо, Дж; Форназье, М; Тоскани, Дж. (2010). «Частичные, кинетические и гидродинамические модели роения». Математическое моделирование коллективного поведения в социально-экономических науках и науках о жизни (PDF) . Моделирование и симуляция в науке, технике и технологиях. Том. 3. С. 297–336. CiteSeerX 10.1.1.193.5047 . дои : 10.1007/978-0-8176-4946-3_12 . ISBN 978-0-8176-4945-6 .
- ^ Давидеску, Мирча Р.; Романчук, Павел; Грегор, Томас; Кузин, Иэн Д. (14 марта 2023 г.). «Рост приводит к нарушению координации у Trichoplax adhaerens, животного, лишенного центральной нервной системы» . Труды Национальной академии наук . 120 (11): e2206163120. Бибкод : 2023PNAS..12006163D . дои : 10.1073/pnas.2206163120 . ISSN 0027-8424 . ПМЦ 10089153 . ПМИД 36897970 .
- ^ Сарфати, Рафаэль; Джоши, Кунаал; Мартин, Оуэн; Хейс, Джули С; Айер-Бисвас, Шривидья; Пелег, Орит (13 марта 2023 г.). Джардина, Ирен (ред.). «Эмерджентная периодичность в коллективном синхронном мелькании светлячков» . электронная жизнь . 12 : e78908. doi : 10.7554/eLife.78908 . ISSN 2050-084X . ПМИД 36912783 .
- ^ Сарфати, Рафаэль; Пелег, Орит (18 ноября 2022 г.). «Состояния Химеры среди синхронных светлячков» . Достижения науки . 8 (46): eadd6690. Бибкод : 2022SciA....8D6690S . дои : 10.1126/sciadv.add6690 . ISSN 2375-2548 . ПМЦ 9668303 . ПМИД 36383660 .
- ^ Jump up to: а б Смит, Чарльз Х. «Незаконченное дело Уоллеса» . Сложность (издатель Wiley Periodicals, Inc.) Том 10, № 2, 2004 г. Проверено 11 мая 2007 г.
- ^ Уоллес, Альфред. «О склонности разновидностей к бесконечному отклонению от исходного типа» . Альфред Рассел Уоллес Пейдж, организованный Университетом Западного Кентукки . Проверено 22 апреля 2007 г.
- ^ Брэнд, Стюарт. «Ради бога, Маргарет» . CoEvolutionary Quarterly, июнь 1976 г. Проверено 4 апреля 2007 г.
- ^ Торп, Дж. Х., и Делонг, доктор медицины (1994). Модель речной продуктивности: эвристический взгляд на источники углерода и органическую переработку в крупных речных экосистемах. Ойкос, 305-308
- ^ Бенке, AC, Чаубей, И., Уорд, GM, и Данн, EL (2000). Динамика импульсов паводков в нерегулируемой пойме реки на юго-востоке прибрежной равнины США. Экология, 2730-2741.
- ^ Каддингтон К. и Бейснер Б.Е. (2005) Утраченные экологические парадигмы: пути изменения теории Academic Press. ISBN 978-0-12-088459-9 .
- ^ Jump up to: а б Суле М.Е., Симберлофф Д. (1986). «Что генетика и экология говорят нам о проектировании заповедников?» (PDF) . Биологическая консервация . 35 (1): 19–40. дои : 10.1016/0006-3207(86)90025-X . hdl : 2027.42/26318 .
Дальнейшее чтение
[ редактировать ]- Классический текст — «Теоретическая экология: принципы и приложения » Анжелы Маклин и Роберта Мэя . Издание 2007 года опубликовано издательством Oxford University Press. ISBN 978-0-19-920998-9 .
- Болкер Б.М. (2008) Экологические модели и данные в издательстве R Princeton University Press. ISBN 978-0-691-12522-0 .
- Случай TJ (2000) Иллюстрированное руководство по теоретической экологии Oxford University Press. ISBN 978-0-19-508512-9 .
- Касвелл Х. (2000) Матричные модели популяции: построение, анализ и интерпретация , Sinauer, 2-е изд. ISBN 978-0-87893-096-8 .
- Эдельштейн-Кешет Л. (2005) Математические модели в биологии, Общество промышленной и прикладной математики. ISBN 978-0-89871-554-5 .
- Готелли, Нью-Джерси (2008) Учебник по экологии Sinauer Associates, 4-е изд. ISBN 978-0-87893-318-1 .
- Готелли, Нью-Джерси и Эллисон (2005) Учебник экологической статистики, издательство Sinauer Associates. ISBN 978-0-87893-269-6 .
- Гастингс А. (1996) Популяционная биология: концепции и модели Springer. ISBN 978-0-387-94853-9 .
- Хилборн Р. и М. Кларк (1997) Экологический детектив: сопоставление моделей с данными Princeton University Press.
- Кокко Х (2007) Моделирование для полевых биологов и других интересных людей Издательство Кембриджского университета. ISBN 978-0-521-83132-1 .
- Кот М. (2001) Элементы математической экологии Издательство Кембриджского университета. ISBN 978-0-521-00150-2 .
- Лоутон Дж. Х. (1999). «Есть ли общие законы в экологии?» (PDF) . Ойкос . 84 (2): 177–192. Бибкод : 1999Oikos..84..177L . CiteSeerX 10.1.1.331.1173 . дои : 10.2307/3546712 . JSTOR 3546712 . Архивировано из оригинала (PDF) 11 июня 2010 г.
- Мюррей Дж.Д. (2002) Математическая биология, Том 1 Springer, 3-е изд. ISBN 978-0-387-95223-9 .
- Мюррей Дж. Д. (2003) Математическая биология, Том 2 Springer, 3-е изд. ISBN 978-0-387-95228-4 .
- Пастор Дж. (2008) Математическая экология популяций и экосистем Уайли-Блэквелл. ISBN 978-1-4051-8811-1 .
- Рафгарден Дж. (1998) Основы экологической теории Прентис Холл. ISBN 978-0-13-442062-2 .
- Уланович Р. (1997) Экология: восходящая перспектива Издательство Колумбийского университета.