Полиплоидия

Полиплоидия — состояние, при котором имеют более клетки организма одной пары ( гомологических ) хромосом . Большинство видов, клетки которых имеют ядра ( эукариоты ), диплоидны , то есть имеют два полных набора хромосом, по одному от каждого из двух родителей; каждый набор содержит одинаковое количество хромосом, и хромосомы соединены в пары гомологичных хромосом. Однако некоторые организмы полиплоидны . Полиплоидия особенно распространена у растений. Большинство эукариот имеют диплоидные соматические клетки , но производят гаплоидные гаметы (яйцеклетки и сперматозоиды) путем мейоза . Моноплоид . имеет только один набор хромосом, и этот термин обычно применяется только к клеткам или организмам, которые обычно являются диплоидными самцы пчел и других перепончатокрылых Например, моноплоидны. В отличие от животных, растения и многоклеточные водоросли имеют жизненный цикл с двумя чередующимися многоклеточными поколениями . гаметофитов митоза является гаплоидным и производит гаметы путем Поколение ; Поколение спорофитов диплоидно и образует споры путем мейоза. .
Полиплоидия — результат дупликации всего генома в ходе эволюции вида. Это может произойти из-за аномального деления клеток либо во время митоза, либо, чаще, из-за неспособности хромосом разделиться во время мейоза или из-за оплодотворения яйцеклетки более чем одним сперматозоидом. [ 1 ] Кроме того, его можно индуцировать в растениях и клеточных культурах некоторыми химическими веществами: наиболее известным из них является колхицин , который может привести к удвоению хромосом, хотя его использование может иметь и другие, менее очевидные последствия. Оризалин также удвоит существующее содержание хромосом.
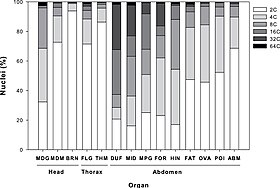
Среди млекопитающих высокая частота полиплоидных клеток обнаруживается в таких органах, как мозг, печень, сердце и костный мозг. [ 2 ] Он также встречается в соматических клетках других животных , например, золотых рыбок , [ 3 ] лосось и саламандры . Распространен среди папоротников и цветковых растений (см. Hibiscus rosa-sinensis ), включая как дикие, так и культурные виды . Пшеница , например, после тысячелетий гибридизации и модификации человеком имеет штаммы, которые являются диплоидными (два набора хромосом), тетраплоидными (четыре набора хромосом) с общим названием твердая или макаронная пшеница и гексаплоидными (шесть наборов хромосом). ) с общим названием хлебная пшеница. Многие важные для сельского хозяйства растения рода Brassica также являются тетраплоидами. Сахарный тростник может иметь уровень плоидности выше, чем октаплоид . [ 4 ]
Полиплоидизация может быть механизмом симпатрического видообразования , поскольку полиплоиды обычно не могут скрещиваться со своими диплоидными предками. Примером может служить растение Erythranthe peregrina . Секвенирование подтвердило, что этот вид произошел от E. × robertsii , стерильного триплоидного гибрида E. Guttata и E. lutea, оба из которых были интродуцированы и натурализованы в Соединенном Королевстве. Новые популяции E. peregrina возникли на материковой части Шотландии и на Оркнейских островах в результате дупликации генома местных популяций E. × robertsii . [ 5 ] Из-за редкой генетической мутации E. peregrina не бесплодна. [ 6 ]
С другой стороны, полиплоидизация может быть и механизмом своего рода «обратного видообразования». [ 7 ] при этом поток генов активируется после события полиплоидии, даже между линиями, которые ранее не испытывали потока генов как диплоиды. Это было подробно описано на геномном уровне у Arabidopsis arenosa и Arabidopsis lyrata . [ 8 ] Каждый из этих видов испытал независимые события автополиплоидии (внутривидовая полиплоидия, описанная ниже), что затем сделало возможным последующий межвидовой поток генов адаптивных аллелей, в данном случае стабилизируя каждую молодую полиплоидную линию. [ 9 ] Такая адаптивная интрогрессия, основанная на полиплоидии, может позволить полиплоидам действовать как «аллельные губки», посредством чего они накапливают загадочные геномные вариации, которые могут быть рекрутированы при столкновении с более поздними экологическими проблемами. [ 10 ]
Терминология
[ редактировать ]Типы
[ редактировать ]
Полиплоидные типы маркируются в зависимости от числа наборов хромосом в ядре . Буква x используется для обозначения количества хромосом в одном наборе:
- гаплоид (один набор; 1 x ), например самцы европейских огненных муравьев
- диплоид (два набора; 2 х ), например человек
- триплоид (три набора; 3 x ), например, стерильные рыжики или арбузы без косточек , также распространены в типе Tardigrada. [ 11 ]
- тетраплоид (четыре набора; 4 х ), например, крыса Plains viscacha , Salmonidae , рыбы [ 12 ] хлопковый Gossypium hirsutum [ 13 ]
- пентаплоид (пять комплектов; 5 х ), например береза Кенай ( Betula kenaica )
- гексаплоид (шесть комплектов; 6 х ), например некоторые виды пшеницы , [ 14 ] киви [ 15 ]
- гептаплоид или септаплоид (семь комплектов; 7 х )
- октаплоид или октоплоид , (восемь комплектов; 8 х ), например Acipenser (род осетровых рыб), георгины
- декаплоид (десять комплектов; 10 х ), например, некоторые сорта клубники
- додекаплоид или дуодекаплоид (двенадцать наборов; 12 х ), например растения Celosia argentea и Spartina anglica. [ 16 ] или амфибия Xenopus ruwenzoriensis .
- тетратетраконтаплоид (сорок четыре комплекта; 44 х ), например шелковица черная . [ 17 ]
Классификация
[ редактировать ]Автополиплоидия
[ редактировать ]Автополиплоиды — это полиплоиды с несколькими наборами хромосом, происходящие от одного таксона .
Двумя примерами природных автополиплоидов являются контрейлерное растение Tolmiea menzisii. [ 18 ] и белый осетр Acipenser transmontanum . [ 19 ] Большинство случаев автополиплоидии являются результатом слияния нередуцированных (2 n ) гамет, в результате чего появляется либо триплоидное ( n + 2 n = 3 n ), либо тетраплоидное (2 n + 2 n = 4 n ) потомство. [ 20 ] Триплоидное потомство обычно стерильно (как в случае с феноменом триплоидного блока ), но в некоторых случаях оно может производить большое количество нередуцированных гамет и, таким образом, способствовать образованию тетраплоидов. Этот путь к тетраплоидии называется триплоидным мостом . [ 20 ] Триплоиды могут также сохраняться посредством бесполого размножения . Фактически, стабильная аутотриплоидия у растений часто связана с апомиктическими системами спаривания. [ 21 ] В сельскохозяйственных системах аутотриплоидия может привести к бессемянности, как у арбузов и бананов . [ 22 ] Триплоидия также используется в разведении лосося и форели для индукции бесплодия. [ 23 ] [ 24 ]
В редких случаях автополиплоиды возникают в результате спонтанного соматического удвоения генома, что наблюдалось у яблони ( Malus Domesticus ) видов . [ 25 ] Это также наиболее распространенный путь искусственно индуцированной полиплоидии, когда такие методы, как слияние протопластов или обработка колхицином , оризалином или ингибиторами митоза , используются для нарушения нормального митотического деления, что приводит к образованию полиплоидных клеток. Этот процесс может быть полезен при селекции растений, особенно при попытке проникновения зародышевой плазмы на плоидные уровни. [ 26 ]
Автополиплоиды обладают по крайней мере тремя гомологичными наборами хромосом, что может приводить к высоким показателям мультивалентного спаривания во время мейоза (особенно у недавно образовавшихся аутополиплоидов, также известных как неополиплоиды) и связанному с этим снижению фертильности из-за образования анеуплоидных гамет. [ 27 ] Естественный или искусственный отбор по фертильности может быстро стабилизировать мейоз у аутополиплоидов за счет восстановления бивалентного спаривания во время мейоза. Быстрая адаптивная эволюция мейотического аппарата, приводящая к снижению уровня мультивалентов (и, следовательно, к стабильному аутополиплоидному мейозу), была зарегистрирована у Arabidopsis arenosa. [ 28 ] и арабидопсис лирата , [ 29 ] со специфическими адаптивными аллелями этих видов, общими только для эволюционировавших полиплоидов. [ 30 ] [ 31 ]
Высокая степень гомологии между дуплицированными хромосомами приводит к тому, что аутополиплоиды проявляют полисомное наследование . [ 32 ] Этот признак часто используется в качестве диагностического критерия для отличия аутополиплоидов от аллополиплоидов, которые обычно демонстрируют дисомное наследование после прохождения стадии неополиплоида. [ 33 ] Хотя большинство полиплоидных видов однозначно характеризуются как автополиплоидные или аллополиплоидные, эти категории представляют собой концы спектра расхождений между родительскими субгеномами. Полиплоиды, находящиеся между этими двумя крайностями, которые часто называют сегментными аллополиплоидами, могут демонстрировать промежуточные уровни полисомного наследования, которые варьируются в зависимости от локуса. [ 34 ] [ 35 ]
Считается, что около половины всех полиплоидов являются результатом аутополиплоидии. [ 36 ] [ 37 ] хотя многие факторы затрудняют оценку этой доли. [ 38 ]
Аллополиплоидия
[ редактировать ]Аллополиплоиды , амфиполиплоиды или гетерополиплоиды — это полиплоиды с хромосомами, происходящими от двух или более разошедшихся таксонов.
Как и при аутополиплоидии, это в первую очередь происходит за счет слияния нередуцированных (2n ) гамет, которое может происходить до или после гибридизации . В первом случае нередуцированные гаметы каждого диплоидного таксона – или редуцированные гаметы двух аутотетраплоидных таксонов – объединяются, образуя аллополиплоидное потомство. В последнем случае один или несколько диплоидных F 1 гибридов производят нередуцированные гаметы, которые сливаются, образуя аллополиплоидное потомство. [ 39 ] Гибридизация с последующей дупликацией генома может быть более распространенным путем к аллополиплоидии, поскольку гибриды F 1 между таксонами часто имеют относительно высокие темпы образования нередуцированных гамет - расхождение между геномами двух таксонов приводит к аномальному спариванию гомеологичных хромосом или нерасхождению во время мейоза. [ 39 ] В этом случае аллополиплоидия может фактически восстановить нормальное двухвалентное мейотическое спаривание, предоставляя каждой гомеологичной хромосоме свой собственный гомолог. Если расхождение между гомеологичными хромосомами происходит даже по двум субгеномам, это теоретически может привести к быстрому восстановлению бивалентного спаривания и дисомного наследования после аллополиплоидизации. Однако многовалентное спаривание часто встречается у многих недавно образовавшихся аллополиплоидов, поэтому вполне вероятно, что большая часть мейотической стабилизации происходит постепенно за счет отбора. [ 27 ] [ 33 ]
Поскольку спаривание между гомеологичными хромосомами редко встречается у укоренившихся аллополиплоидов, им может быть полезна фиксированная гетерозиготность гомеологичных аллелей. [ 40 ] В некоторых случаях такая гетерозиготность может иметь полезные гетеротические эффекты либо с точки зрения приспособленности в природных условиях, либо с точки зрения желательных признаков в сельскохозяйственных условиях. Это могло бы частично объяснить распространенность аллополиплоидии среди видов сельскохозяйственных культур. И мягкая пшеница , и тритикале являются примерами аллополиплоидов с шестью наборами хромосом. Хлопок , арахис или киноа — аллотетраплоиды множественного происхождения. В капустных культурах треугольник U описывает взаимоотношения между тремя распространенными диплоидными растениями Brassica ( B. oleracea , B. rapa B. . nigra ) и тремя аллотетраплоидами ( B. napus , B. juncea и и B. carinata ), полученными из капустных культур гибридизация диплоидных видов. Аналогичная связь существует между тремя диплоидными видами Tragopogon ( T. dubius , T. pratensis и ) и T. porrifolius двумя аллотетраплоидными видами ( T. mirus и T. miscellus ). [ 41 ] Сложные закономерности аллополиплоидной эволюции также наблюдались у животных, например, у лягушек рода Xenopus . [ 42 ]
Анеуплоид
[ редактировать ]Организмы, в которых определенная хромосома или сегмент хромосомы недостаточно или чрезмерно представлены, называются анеуплоидными (от греческих слов, означающих «не», «хороший» и «складной»). Анеуплоидия относится к численному изменению части набора хромосом, тогда как полиплоидия относится к численному изменению всего набора хромосом. [ 43 ]
Эндополиплоидия
[ редактировать ]Полиплоидия встречается в некоторых тканях животных, которые в остальном являются диплоидными, например, в мышечных тканях человека. [ 44 ] Это известно как эндополиплоидия . Виды, клетки которых не имеют ядер, то есть прокариоты , могут быть полиплоидными, как это видно на примере крупной бактерии Epulopiscium fishelsoni . [ 45 ] Следовательно, плоидность определяется по отношению к клетке.
моноплоидный
[ редактировать ]Моноплоид имеет только один набор хромосом, и этот термин обычно применяется только к клеткам или организмам, которые обычно являются диплоидными. Более общий термин для таких организмов — гаплоидный .
Временные условия
[ редактировать ]Неополиплоидия
[ редактировать ]Недавно образовавшийся полиплоид.
Мезополиплоидия
[ редактировать ]В новейшей истории это стало полиплоидным; он не так нов, как неополиплоид, и не так стар, как палеополиплоид. Это полиплоид среднего возраста. Часто это относится к дупликации всего генома с последующим промежуточным уровнем диплоидизации.
Палеополиплоидия
[ редактировать ]
Древние дупликации генома, вероятно, имели место в эволюционной истории всей жизни. События дупликации, которые произошли давным-давно в истории различных эволюционных линий, могут быть трудно обнаружить из-за последующей диплоидизации (например, полиплоид с течением времени начинает вести себя цитогенетически как диплоид), поскольку мутации и трансляции генов постепенно создают одну копию каждой хромосомы в отличие от другой экземпляр. Со временем дублированные копии генов также часто накапливают мутации и становятся неактивными псевдогенами. [ 46 ]
Во многих случаях об этих событиях можно судить только путем сравнения секвенированных геномов . Примеры неожиданных, но недавно подтвержденных дупликаций древнего генома включают пекарские дрожжи ( Saccharomyces cerevisiae ), горчицу/салатовый кресс ( Arabidopsis thaliana ), рис ( Oryza sativa ) и два раунда дупликации всего генома ( гипотеза 2R ) у раннего эволюционного предка. позвоночных линию) и еще один , (включая человеческую близкий к происхождению костистых рыб . [ 47 ] Покрытосеменные ( цветковые растения ) имеют палеополиплоидию в своем происхождении. Все эукариоты , вероятно, в какой-то момент своей эволюционной истории испытали явление полиплоидии.
Другие похожие термины
[ редактировать ]Кариотип
[ редактировать ]Кариотип — это характерный хромосомный набор эукариот видов . [ 48 ] [ 49 ] Получение и изучение кариотипов является частью цитологии , а точнее цитогенетики .
Хотя репликация и транскрипция ДНК у эукариот высоко стандартизированы , этого нельзя сказать об их кариотипах, которые сильно различаются у разных видов по числу хромосом и детальной организации, несмотря на то, что они построены из одних и тех же макромолекул. В некоторых случаях наблюдаются даже значительные различия внутри вида. Эта вариация обеспечивает основу для ряда исследований в области так называемой эволюционной цитологии.
Гомеологичные хромосомы
[ редактировать ]Гомеологичные хромосомы — это хромосомы, собранные вместе в результате межвидовой гибридизации и аллополиплоидизации , взаимоотношения которых были полностью гомологичны у предкового вида. Например, твердая пшеница является результатом межвидовой гибридизации двух диплоидных видов трав Triticum urartu и Aegilops speltoides . Оба диплоидных предка имели два набора по 7 хромосом, сходных по размеру и содержащимся в них генам. Твердая пшеница содержит гибридный геном с двумя наборами хромосом, полученными от Triticum urartu , и двумя наборами хромосом, полученными от Aegilops speltoides . Каждая пара хромосом, полученная от Triticum urartu, родителя гомеологична противоположной паре хромосом, полученной от родителя Aegilops speltoides , хотя каждая пара хромосом сама по себе гомологична .
Примеры
[ редактировать ]Животные
[ редактировать ]Примеры у животных чаще встречаются у беспозвоночных. [ 50 ] такие как плоские черви , пиявки и артемии . Среди позвоночных примеры стабильной полиплоидии включают лососевых и многих карповых (например, карпа ). [ 51 ] Некоторые рыбы имеют до 400 хромосом. [ 51 ] Полиплоидия также часто встречается у амфибий; например, биологически важный род Xenopus включает множество различных видов с 12 наборами хромосом (додекаплоид). [ 52 ] Полиплоидные ящерицы также довольно распространены. Большинство из них бесплодны и размножаются партеногенезом ; [ нужна ссылка ] другие, как Liolaemus chiliensis , поддерживают половое размножение. Полиплоидные кротовые саламандры (в основном триплоиды) все женские особи и размножаются путем клептогенеза . [ 53 ] «кражи» сперматофоров у диплоидных самцов родственных видов, чтобы вызвать развитие яйцеклеток, но без включения ДНК самцов в потомство.
Хотя некоторые ткани млекопитающих, например паренхиматозные клетки печени, полиплоидны, [ 54 ] [ 55 ] редкие случаи полиплоидных млекопитающих известны , но чаще всего они приводят к внутриутробной смерти. октодонтидный , грызун обитающий в районах Аргентины суровых пустынных , известный как равнинная вискача-крыса ( Tympanoctomys barrerae ). Исключением из этого «правила» считается [ 56 ] имеется только две копии каждой хромосомы Однако тщательный анализ с использованием красок хромосом показывает, что у T. barrerae , а не четыре, которые ожидались бы, если бы он действительно был тетраплоидом. [ 57 ] Этот грызун не крыса , а родственник морских свинок и шиншилл . Его «новое» диплоидное число (2 n ) равно 102, поэтому его клетки примерно вдвое превышают нормальный размер. Его ближайшим живым родственником является Octomys mimax , андская вискача-крыса того же семейства, у которой 2 n = 56. Поэтому было высказано предположение, что Octomys -подобный предок произвел тетраплоидное (т. е. 2 n = 4 x = 112) потомство, которое было , в силу своих удвоенных хромосом, репродуктивно изолированных от своих родителей.
Полиплоидию индуцировал у рыб Хар Сваруп (1956) с помощью обработки икры холодовым шоком незадолго до момента оплодотворения, в результате чего были получены триплоидные эмбрионы, которые успешно созрели. [ 58 ] [ 59 ] Также было показано, что холодный или тепловой шок приводит к невосстановлению гамет амфибий, хотя это чаще происходит в яйцеклетках, чем в сперматозоидах. [ 60 ] Джон Гердон (1958) трансплантировал неповрежденные ядра соматических клеток для получения диплоидных яиц у лягушки Xenopus (продолжение работы Бриггса и Кинга в 1952 году), которые были способны развиваться до стадии головастика. [ 61 ] Британский ученый Дж.Б.С. Холдейн высоко оценил потенциальную возможность применения этой работы в медицине и, описывая результаты, стал одним из первых, кто использовал слово « клон » по отношению к животным. Более поздняя работа Шинья Яманака показала, как можно перепрограммировать зрелые клетки, чтобы они стали плюрипотентными, расширяя возможности нестволовых клеток. За эту работу Гурдон и Яманака были совместно удостоены Нобелевской премии в 2012 году. [ 61 ]
Люди
[ редактировать ]
Истинная полиплоидия редко встречается у людей, хотя полиплоидные клетки встречаются в высокодифференцированных тканях , таких как паренхима печени , сердечная мышца, плацента и костный мозг. [ 62 ] [ 63 ] Анеуплоидия встречается чаще.
Полиплоидия встречается у человека в форме триплоидии с 69 хромосомами (иногда называемой 69, ХХХ) и тетраплоидии с 92 хромосомами (иногда называемой 92, ХХХХ). Триплоидия, обычно обусловленная полиспермией , встречается примерно в 2–3% всех беременностей у человека и примерно в 15% выкидышей. [ нужна ссылка ] Подавляющее большинство триплоидных зачатий заканчивается выкидышем ; те, кто доживает до срока, обычно умирают вскоре после рождения. В некоторых случаях выживаемость после рождения может быть увеличена, если имеется миксоплоидия с наличием как диплоидной , так и триплоидной популяции клеток. Был один случай, когда ребенок дожил до семимесячного возраста с синдромом полной триплоидии. У него не было нормального умственного и физического развития в неонатальном периоде, и он умер от инфекции Pneumocystis carinii , что указывает на слабую иммунную систему. [ 64 ]
Триплоидия может быть результатом либо дигинии (дополнительный гаплоидный набор от матери), либо диандрии (дополнительный гаплоидный набор от отца). Диандрия в основном вызвана редупликацией отцовского гаплоидного набора из одного сперматозоида, но также может быть следствием диспермического (два спермия) оплодотворения яйцеклетки. [ 65 ] Дигиния чаще всего вызывается либо отказом одного мейотического деления во время оогенеза, приводящим к образованию диплоидного ооцита , либо неспособностью выдавить одно полярное тельце из ооцита . Диандрия, по-видимому, преобладает среди ранних выкидышей , тогда как дигиния преобладает среди триплоидных зигот, доживающих до внутриутробного периода. [ 66 ] Однако среди ранних выкидышей дигиния также чаще встречается в тех случаях, менее Срок беременности 8 + 1 / 2 недели или те, при которых присутствует эмбрион. Есть также два различных фенотипа у триплоидных плацент и плодов , которые зависят от происхождения дополнительного гаплоидного набора. При дигинии обычно наблюдается асимметричный плохо развитый плод с выраженной надпочечников гипоплазией и очень маленькой плацентой . [ 67 ] частичный пузырный занос . При диандрии развивается [ 65 ] Эти эффекты происхождения родителей отражают эффекты геномного импринтинга . [ нужна ссылка ]
Полная тетраплоидия диагностируется реже, чем триплоидия, но наблюдается в 1–2% ранних выкидышей. Однако некоторые тетраплоидные клетки обычно обнаруживаются при анализе хромосом при пренатальной диагностике , и их обычно считают «безвредными». Неясно, возникают ли эти тетраплоидные клетки просто во время культуры клеток in vitro или они также присутствуют в плацентарных клетках in vivo . Во всяком случае, имеется очень мало клинических сообщений о плодах/младенцах с диагнозом тетраплоидного мозаицизма.
Миксоплоидия довольно часто наблюдается у предимплантационных эмбрионов человека и включает гаплоидные/диплоидные, а также диплоидные/тетраплоидные смешанные популяции клеток. Неизвестно, не удается ли этим эмбрионам имплантироваться и поэтому они редко выявляются при продолжающейся беременности, или это просто селективный процесс, благоприятствующий диплоидным клеткам.
Рыба
[ редактировать ]В стволовой линии костистых рыб произошел случай полиплоидии. [ 47 ]
Растения
[ редактировать ]
Полиплоидия часто встречается у растений; по некоторым оценкам, 30–80% ныне живущих видов растений являются полиплоидными, и многие линии демонстрируют признаки древней полиплоидии ( палеополиплоидии ) в своих геномах. [ 68 ] [ 69 ] [ 70 ] [ 71 ] Огромные взрывы разнообразия видов покрытосеменных , по-видимому, совпали со временем дублирования древних геномов, свойственных многим видам. [ 72 ] Установлено, что 15% видообразований покрытосеменных и 31% папоротников сопровождаются увеличением плоидности. [ 73 ]
Полиплоидные растения могут возникать в природе спонтанно за счет нескольких механизмов, включая мейотические или митотические нарушения и слияние нередуцированных (2n ) гамет. [ 40 ] Оба автополиплоида (например, картофеля [ 74 ] ) и аллополиплоиды (такие как рапс, пшеница и хлопок) можно найти как среди диких, так и среди одомашненных видов растений.
Большинство полиплоидов демонстрируют новые вариации или морфологии по сравнению с их родительскими видами, которые могут способствовать процессам видообразования и эксплуатации экономической ниши. [ 69 ] [ 40 ] Механизмы, приводящие к новым вариациям во вновь образованных аллополиплоидах, могут включать эффекты дозировки генов (в результате более многочисленных копий содержимого генома), воссоединение расходящихся иерархий регуляции генов, хромосомные перестройки и эпигенетическое ремоделирование, все из которых влияют на содержание генов и/или уровни экспрессии. [ 75 ] [ 76 ] [ 77 ] [ 78 ] Многие из этих быстрых изменений могут способствовать репродуктивной изоляции и видообразованию. Однако семена, полученные в результате интерплоидных скрещиваний , например, между полиплоидами и их родительскими видами, обычно имеют аберрантное развитие эндосперма, что снижает их жизнеспособность. [ 79 ] [ 80 ] тем самым способствуя полиплоидному видообразованию . Полиплоиды могут также скрещиваться с диплоидами и давать полиплоидные семена, как это наблюдается в агамных комплексах Crepis . [ 81 ]
Некоторые растения триплоидны. Поскольку мейоз нарушен, эти растения стерильны, причем все растения имеют одинаковую генетическую конституцию: среди них исключительно вегетативно размножаемый шафрановый крокус ( Crocus sativus ). Кроме того, чрезвычайно редкий тасманский кустарник Lomatia tasmanica представляет собой триплоидный стерильный вид.
В природе встречается мало полиплоидных хвойных деревьев . [ 82 ] Одним из примеров является Coast Redwood Sequoia sempervirens , который представляет собой гексаплоид (6 x ) с 66 хромосомами (2 n = 6 x = 66), хотя происхождение неясно. [ 83 ]
Водные растения, особенно однодольные , включают большое количество полиплоидов. [ 84 ]
Культуры
[ редактировать ]Индукция полиплоидии является распространенным методом преодоления стерильности гибридных видов при селекции растений. Например, тритикале — это гибрид пшеницы ( Triticum turgidum ) и ржи ( Secale зерновых ). Он сочетает в себе востребованные характеристики родителей, но исходные гибриды бесплодны. После полиплоидизации гибрид становится плодовитым и, таким образом, может быть далее размножен до тритикале.
В некоторых ситуациях предпочтение отдается полиплоидным культурам, поскольку они стерильны. Например, многие бессемянные сорта плодов являются бессемянными в результате полиплоидии. Такие культуры размножают бесполыми методами, например прививкой .
Полиплоидия у сельскохозяйственных растений чаще всего вызывается обработкой семян химическим веществом колхицином .
Примеры
[ редактировать ]- Триплоидные культуры: некоторые сорта яблони (такие как Бель де Боскоп , Джонаголд , Муцу , Рибстон Пиппин ), бананы , цитрусовые , имбирь , арбуз , [ 85 ] шафрановый крокус , белая мякоть кокоса
- Тетраплоидные культуры: очень мало яблони сортов , твердая или макаронная пшеница , хлопок , картофель , рапс / рапс , лук-порей , табак , арахис , кинноу , пеларгония.
- Гексаплоидные культуры: хризантема , мягкая пшеница , тритикале , овес , киви. [ 15 ]
- Октаплоидные культуры: клубника , георгины , анютины глазки , сахарный тростник , ока ( Oxalis tuberosa ). [ 86 ]
- Додекаплоидные культуры: некоторые сахарного тростника гибриды [ 87 ]
Некоторые культуры встречаются в различных плоидиях: тюльпаны и лилии обычно встречаются как диплоидные, так и триплоидные; лилейники ( сорта Hemerocallis ) доступны как в диплоидном, так и в тетраплоидном виде; яблоки и мандарины кинноу могут быть диплоидными, триплоидными или тетраплоидными.
Грибы
[ редактировать ]Помимо растений и животных, эволюционная история различных видов грибов усеяна прошлыми и недавними событиями полногеномной дупликации (см. Albertin and Marullo 2012). [ 88 ] на рассмотрение). Известно несколько примеров полиплоидов:
- автополиплоид: водные грибы рода Allomyces . [ 89 ] некоторые штаммы Saccharomyces cerevisiae, используемые в хлебобулочных изделиях , [ 90 ] и т. д.
- аллополиплоид: широко распространенный Cyathus stercoreus , [ 91 ] аллотетраплоидные лагерные дрожжи Saccharomyces Pastorianus , [ 92 ] аллотриплоидные винные дрожжи Dekkera bruxellensis , [ 93 ] и т. д.
- палеополиплоид: патоген человека Rhizopus oryzae , [ 94 ] род Saccharomyces , [ 95 ] и т. д.
Кроме того, полиплоидия часто связана с гибридизацией и сетчатой эволюцией, которые, по-видимому, широко распространены в некоторых таксонах грибов. Действительно, гомоплоидное видообразование (гибридное видообразование без изменения числа хромосом ) было доказано для некоторых видов грибов (таких как базидиомикота Microbotryum violaceum) . [ 96 ] ).
Что касается растений и животных, грибные гибриды и полиплоиды демонстрируют структурные и функциональные модификации по сравнению со своими предшественниками и диплоидными аналогами. В частности, структурные и функциональные результаты полиплоидных геномов Saccharomyces поразительно отражают эволюционную судьбу полиплоидных растений. Большие хромосомные перестройки [ 97 ] приводит к образованию химерных хромосом [ 98 ] были описаны, а также более точечные генетические модификации, такие как потеря генов. [ 99 ] Гомоаллели аллотетраплоидных дрожжей S. Pastorianus вносят неравный вклад в транскриптом . [ 100 ] Фенотипическая диверсификация также наблюдается после полиплоидизации и/или гибридизации у грибов. [ 101 ] производство топлива для естественного отбора и последующей адаптации и видообразования.
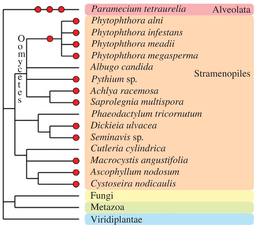
Хромальвеолата
[ редактировать ]Другие таксоны эукариот за свою эволюционную историю испытали один или несколько событий полиплоидизации (см. Albertin and Marullo, 2012). [ 88 ] на рассмотрение). Оомицеты рода , которые не являются настоящими представителями грибов, содержат несколько примеров палеополиплоидных и полиплоидных видов, например, в пределах Phytophthora . [ 102 ] Некоторые виды бурых водорослей ( Fucales , Laminariales). [ 103 ] и диатомовые водоросли [ 104 ] ) содержат очевидные полиплоидные геномы. В группе Alveolata замечательный вид Paramecium Tetraurelia претерпел три последовательных цикла полногеномной дупликации. [ 105 ] и зарекомендовал себя как основная модель для исследований палеополиплоидов.
Бактерии
[ редактировать ]Каждая Deinococcus radiodurans бактерия содержит 4-8 копий своей хромосомы . [ 106 ] Воздействие D. radiodurans облучением рентгеновским или высушиванием может разбить его геном на сотни коротких случайных фрагментов. Тем не менее, D. radiodurans обладает высокой устойчивостью к таким воздействиям. Механизм, с помощью которого геном точно восстанавливается, включает опосредованную RecA гомологичную рекомбинацию и процесс, называемый расширенным зависимым от синтеза отжигом цепи (SDSA) . [ 107 ]
Azotobacter vinelandii может содержать до 80 копий хромосом на клетку. [ 108 ] Однако это наблюдается только в быстрорастущих культурах, тогда как культуры, выращенные на синтетических минимальных средах, не являются полиплоидными. [ 109 ]
Архея
[ редактировать ]Архея . Halobacterium salinarium полиплоидна [ 110 ] и, как и Deinococcus radiodurans , обладает высокой устойчивостью к рентгеновскому облучению и высыханию, условиям, которые вызывают ДНК . двухцепочечные разрывы [ 111 ] Хотя хромосомы разбиты на множество фрагментов, полные хромосомы можно регенерировать, используя перекрывающиеся фрагменты. В этом механизме используется белок, связывающий одноцепочечную ДНК , и, вероятно, представляет собой гомологичную рекомбинационную репарацию. [ 112 ]
См. также
[ редактировать ]- Диплоидизация
- Гибридный геном эукариот
- Плоидность
- Полиплоидный комплекс
- Полисомия
- Взаимное молчание
- Сочувствие
Ссылки
[ редактировать ]- ^ Соломон Э (2014). Соломон/Мартин/Мартин/Берг, Биология . Cengage Обучение. п. 344. ИСБН 978-1285423586 .
- ^ Чжан С., Линь Ю.Х., Тарлоу Б., Чжу Х. (июнь 2019 г.). «Происхождение и функции полиплоидии печени» . Клеточный цикл . 18 (12): 1302–1315. дои : 10.1080/15384101.2019.1618123 . ПМЦ 6592246 . ПМИД 31096847 .
- ^ Оно С., Мурамото Дж., Кристиан Л., Аткин Н.Б. (1967). «Диплоидно-тетраплоидные отношения среди старосветских представителей семейства рыб Cyprinidae». Хромосома . 23 (1): 1–9. дои : 10.1007/BF00293307 . S2CID 1181521 .
- ^
- Манимекалаи Р., Суреш Г., Говинда Куруп Х., Атиаппан С., Кандалам М. (сентябрь 2020 г.). «Роль методов генотипирования NGS и SNP в программах улучшения сахарного тростника». Критические обзоры по биотехнологии . 40 (6). Тейлор и Фрэнсис (T&F): 865–880. дои : 10.1080/07388551.2020.1765730 . ПМИД 32508157 . S2CID 219537026 .
- В этом обзоре цитируется это исследование:
- Вилела М.М., Дель Бем Л.Е., Ван Слейс М.А., де Сетта Н., Китаджима Дж.П., Круз Г.М. и др. (февраль 2017 г.). «Анализ трех гомо/гомеологичных регионов сахарного тростника предполагает независимые события полиплоидизации Saccharum officinarum и Saccharum spontaneum» . Геномная биология и эволюция . 9 (2): 266–278. дои : 10.1093/gbe/evw293 . ПМК 5381655 . ПМИД 28082603 .
- ^ Вальехо-Марин М., Баггс Р.Дж., Кули А.М., Пьюзи-младший (июнь 2015 г.). «Видообразование путем дупликации генома: повторяющееся происхождение и геномный состав недавно образовавшихся аллополиплоидных видов Mimulus peregrinus» . Эволюция; Международный журнал органической эволюции . 69 (6): 1487–1500. дои : 10.1111/evo.12678 . ПМК 5033005 . ПМИД 25929999 .
- ^ Фессенден М. «Освободите место для нового цветения: обнаружен новый цветок» . Научный американец . Проверено 22 февраля 2017 г.
- ^ Шмикл, Розвита; Янт, Леви (апрель 2021 г.). «Адаптивная интрогрессия: как полиплоидия меняет ландшафт потока генов» . Новый фитолог . 230 (2): 457–461. дои : 10.1111/nph.17204 . ISSN 0028-646X . ПМИД 33454987 .
- ^ Марбургер, Сара; Моннахан, Патрик; Сир, Пол Дж.; Мартин, Саймон Х.; Кох, Иордания; Пааянен, Пирита; Богутинска, Магдалена; Хиггинс, Джеймс Д.; Шмикл, Розвита; Янт, Леви (18 ноября 2019 г.). «Межвидовая интрогрессия опосредует адаптацию к полному дупликации генома» . Природные коммуникации . 10 (1): 5218. Бибкод : 2019NatCo..10.5218M . дои : 10.1038/s41467-019-13159-5 . ISSN 2041-1723 . ПМК 6861236 . ПМИД 31740675 .
- ^ Сир, Пол Дж.; Франция, Мартин Г.; Грегори, Кэтрин Л.; Боже мой, Даррен; Шмикл, Розвита; Янт, Леви; Хиггинс, Джеймс Д. (15 июля 2020 г.). «Новый аллель ASY3 связан с большей мейотической стабильностью у аутотетраплоидного Arabidopsis lyrata» . ПЛОС Генетика . 16 (7): e1008900. дои : 10.1371/journal.pgen.1008900 . ISSN 1553-7404 . ПМЦ 7392332 . ПМИД 32667955 .
- ^ Шмикл, Розвита; Янт, Леви (апрель 2021 г.). «Адаптивная интрогрессия: как полиплоидия меняет ландшафт потока генов» . Новый фитолог . 230 (2): 457–461. дои : 10.1111/nph.17204 . ISSN 0028-646X . ПМИД 33454987 .
- ^ Бертолани Р. (2001). «Эволюция репродуктивных механизмов тихоходок: обзор». Зоологический обзор . 240 (3–4): 247–252. Бибкод : 2001ZooAn.240..247B . дои : 10.1078/0044-5231-00032 .
- ^ Стаудер DJ, Биссон П.А., Найман Р.Дж. (1997). Тихоокеанский лосось и его экосистемы: состояние и будущие варианты . Спрингер. стр. 30–31. ISBN 978-0-412-98691-8 . Проверено 9 июля 2013 г.
- ^ Адамс К.Л., Вендел Дж.Ф. (апрель 2005 г.). «Полиплоидия и эволюция генома растений». Современное мнение в области биологии растений . 8 (2): 135–141. Бибкод : 2005COPB....8..135A . дои : 10.1016/j.pbi.2005.01.001 . ПМИД 15752992 .
- ^ «Тритикум — обзор | Темы ScienceDirect» .
- ^ Jump up to: а б Кроухерст Р.Н., Уиттакер Д., Гарднер Р.К. «Генетическое происхождение киви» . Архивировано из оригинала 25 февраля 2021 г. Проверено 30 августа 2008 г.
- ^ Айнуш М.Л., Форчун П.М., Салмон А., Паризод С., Грандбастьен М.А., Фукунага К. и др. (2008). «Гибридизация, полиплоидия и инвазия: уроки Спартины (Poaceae)». Биологические инвазии . 11 (5): 1159–1173. дои : 10.1007/s10530-008-9383-2 . S2CID 25522023 .
- ^
- Хуссейн Ф., Рана З., Шафик Х., Малик А., Хуссейн З. (2017). «Фитофармакологический потенциал различных видов Morus alba и их биоактивных фитохимических веществ: обзор» . Азиатско-Тихоокеанский журнал тропической биомедицины . 7 (10). Медноу : 950–956. дои : 10.1016/j.apjtb.2017.09.015 . ISSN 2221-1691 .
- Аль-Хайри Дж.М., Джайн С.М., Джонсон Д.В. (2018). Аль-Хайри Дж.М., Джайн С.М., Джонсон Д.В. (ред.). Достижения в стратегиях селекции растений: фрукты . Том. 2. Спрингер Интернэшнл Паблишинг АГ . стр. 89–130. дои : 10.1007/978-3-319-91944-7 . ISBN 978-3-319-91943-0 . S2CID 51706250 .
- Этот обзор и книга цитируют это исследование.
- Цзэн Ц, Чен Х, Чжан С, Хань М, Ли Т, Ци Икс и др. (2015). «Определение восьми видов шелковицы рода Morus с помощью внутренней транскрибируемой филогении на основе спейсеров» . ПЛОС ОДИН . 10 (8): e0135411. Бибкод : 2015PLoSO..1035411Z . дои : 10.1371/journal.pone.0135411 . ПМЦ 4534381 . ПМИД 26266951 .
- ^ Солтис Д.Э. (1 января 1984 г.). «Автополиплоидия Tolmiea menziesii (Saxifragaceae)». Американский журнал ботаники . 71 (9): 1171–1174. дои : 10.2307/2443640 . JSTOR 2443640 .
- ^ Драух Шрайер А., Гилле Д., Махарджа Б., Мэй Б. (ноябрь 2011 г.). «Нейтральные маркеры подтверждают октоплоидное происхождение и выявляют спонтанную аутополиплоидию у белого осетра Acipenser transmontanus » . Журнал прикладной ихтиологии . 27 : 24–33. Бибкод : 2011JAPIc..27...24D . дои : 10.1111/j.1439-0426.2011.01873.x . ISSN 1439-0426 .
- ^ Jump up to: а б Бретаньолл Ф., Томпсон Дж. Д. (январь 1995 г.). «Гаметы с соматическим числом хромосом: механизмы их образования и роль в эволюции автополиплоидных растений» . Новый фитолог . 129 (1): 1–22. дои : 10.1111/j.1469-8137.1995.tb03005.x . ПМИД 33874422 .
- ^ Мюнцинг А (март 1936 г.). «Эволюционное значение автополиплоидии». Эредитас . 21 (2–3): 363–378. дои : 10.1111/j.1601-5223.1936.tb03204.x . ISSN 1601-5223 .
- ^ Варокво Ф., Бланвиллен Р., Дельсени М., Галлуа П. (июнь 2000 г.). «Меньше, да лучше: новые подходы к производству фруктов без косточек». Тенденции в биотехнологии . 18 (6): 233–242. дои : 10.1016/s0167-7799(00)01448-7 . ПМИД 10802558 . S2CID 29713625 .
- ^ Коттер Д., О'Донован В., О'Маойлеиди Н., Роган Г., Рош Н., Уилкинс Н.П. (июнь 2000 г.). «Оценка использования триплоидного атлантического лосося ( Salmo salar L.) для минимизации воздействия сбежавшего выращенного лосося на дикие популяции». Аквакультура . 186 (1–2): 61–75. Бибкод : 2000Aquac.186...61C . дои : 10.1016/S0044-8486(99)00367-1 .
- ^ Линкольн РФ, Скотт А.П. (1983). «Производство женской триплоидной радужной форели». Аквакультура . 30 (1–4): 375–380. Бибкод : 1983Aquac..30..375L . дои : 10.1016/0044-8486(83)90179-5 .
- ^ Дермен Х (май 1951 г.). «Тетраплоидные и диплоидные придаточные побеги: от гигантского вида яблок Макинтош». Журнал наследственности . 42 (3): 145–149. doi : 10.1093/oxfordjournals.jhered.a106189 . ISSN 0022-1503 .
- ^ Двиведи С.Л., Упадхьяя HD, Сталкер HT, Блэр М.В., Бертиоли DJ, Нилен С., Ортис Р. (январь 2007 г.). Яник Дж. (ред.). Обзоры селекции растений (PDF) . Джон Уайли и сыновья. стр. 179–230. дои : 10.1002/9780470380130.ch3 . ISBN 9780470380130 .
- ^ Jump up to: а б Джастин Р. (январь 2002 г.). «Неополиплоидия цветковых растений». Ежегодный обзор экологии и систематики . 33 (1): 589–639. doi : 10.1146/annurev.ecolsys.33.010802.150437 . S2CID 41689384 .
- ^ Янт, Леви; Холлистер, Джесси Д.; Райт, Кевин М.; Арнольд, Брайан Дж.; Хиггинс, Джеймс Д.; Франклин, Ф. Крис Х.; Бомблис, Кирстен (ноябрь 2013 г.). «Мейотическая адаптация к дупликации генома Arabidopsis arenosa» . Современная биология . 23 (21): 2151–2156. Бибкод : 2013CBio...23.2151Y . дои : 10.1016/j.cub.2013.08.059 . ISSN 0960-9822 . ПМЦ 3859316 . ПМИД 24139735 .
- ^ Марбургер, Сара; Моннахан, Патрик; Сир, Пол Дж.; Мартин, Саймон Х.; Кох, Иордания; Пааянен, Пирита; Богутинска, Магдалена; Хиггинс, Джеймс Д.; Шмикл, Розвита; Янт, Леви (18 ноября 2019 г.). «Межвидовая интрогрессия опосредует адаптацию к полному дупликации генома» . Природные коммуникации . 10 (1): 5218. Бибкод : 2019NatCo..10.5218M . дои : 10.1038/s41467-019-13159-5 . ISSN 2041-1723 . ПМК 6861236 . ПМИД 31740675 .
- ^ Марбургер, Сара; Моннахан, Патрик; Сир, Пол Дж.; Мартин, Саймон Х.; Кох, Иордания; Пааянен, Пирита; Богутинска, Магдалена; Хиггинс, Джеймс Д.; Шмикл, Розвита; Янт, Леви (18 ноября 2019 г.). «Межвидовая интрогрессия опосредует адаптацию к полному дупликации генома» . Природные коммуникации . 10 (1): 5218. Бибкод : 2019NatCo..10.5218M . дои : 10.1038/s41467-019-13159-5 . ISSN 2041-1723 . ПМК 6861236 . ПМИД 31740675 .
- ^ Сир, Пол Дж.; Франция, Мартин Г.; Грегори, Кэтрин Л.; Боже мой, Даррен; Шмикл, Розвита; Янт, Леви; Хиггинс, Джеймс Д. (15 июля 2020 г.). Грелон, Матильда (ред.). «Новый аллель ASY3 связан с большей мейотической стабильностью у аутотетраплоидного Arabidopsis lyrata» . ПЛОС Генетика . 16 (7): e1008900. дои : 10.1371/journal.pgen.1008900 . ISSN 1553-7404 . ПМЦ 7392332 . ПМИД 32667955 .
- ^ Паризод С., Холдереггер Р., Брохманн С. (апрель 2010 г.). «Эволюционные последствия аутополиплоидии». Новый фитолог . 186 (1): 5–17. дои : 10.1111/j.1469-8137.2009.03142.x . ПМИД 20070540 .
- ^ Jump up to: а б Ле Комбер СК, Айнуш МЛ, Коварик А., Лейтч А.Р. (апрель 2010 г.). «Создание функционального диплоида: от полисомного к дисомному наследованию». Новый фитолог . 186 (1): 113–122. дои : 10.1111/j.1469-8137.2009.03117.x . ПМИД 20028473 .
- ^ Стеббинс Г.Л. (1947). Виды полиплоидов: их классификация и значение . Достижения генетики. Том. 1. С. 403–429. дои : 10.1016/s0065-2660(08)60490-3 . ISBN 9780120176014 . ПМИД 20259289 .
- ^ Стеббинс Г.Л. (1950). Изменчивость и эволюция растений . Издательство Оксфордского университета. [ нужна страница ]
- ^ Рэмси Дж., Шемске Д.В. (январь 1998 г.). «Пути, механизмы и скорость образования полиплоидов у цветковых растений». Ежегодный обзор экологии и систематики . 29 (1): 467–501. дои : 10.1146/annurev.ecolsys.29.1.467 . S2CID 31637733 .
- ^ Баркер М.С., Арриго Н., Баниага А.Е., Ли З., Левин Д.А. (апрель 2016 г.). «Об относительном обилии автополиплоидов и аллополиплоидов» . Новый фитолог . 210 (2): 391–398. дои : 10.1111/nph.13698 . ПМИД 26439879 .
- ^ Дойл Дж. Дж., Шерман-Бройлс С. (январь 2017 г.). «Двойная беда: таксономия и определения полиплоидии» . Новый фитолог . 213 (2): 487–493. дои : 10.1111/nph.14276 . ПМИД 28000935 .
- ^ Jump up to: а б Рэмси Дж. (январь 1998 г.). «Пути, механизмы и скорость образования полиплоидов у цветковых растений». Ежегодный обзор экологии и систематики . 29 (1): 467–501. дои : 10.1146/annurev.ecolsys.29.1.467 . S2CID 31637733 .
- ^ Jump up to: а б с Комай Л. (ноябрь 2005 г.). «Преимущества и недостатки полиплоида». Обзоры природы. Генетика . 6 (11): 836–846. дои : 10.1038/nrg1711 . ПМИД 16304599 . S2CID 3329282 .
- ^ Оунби М. (январь 1950 г.). «Естественная гибридизация и амфиплоидия рода Tragopogon». Американский журнал ботаники . 37 (7): 487–499. дои : 10.2307/2438023 . JSTOR 2438023 .
- ^ Шмид М., Эванс Б.Дж., Богарт Дж.П. (2015). «Полиплоидия у амфибий» . Цитогенетические и геномные исследования . 145 (3–4): 315–330. дои : 10.1159/000431388 . ПМИД 26112701 .
- ^ Гриффитс Эй Джей (1999). Введение в генетический анализ . Сан-Франциско, Калифорния: WH Freeman. ISBN 978-0-7167-3520-5 . [ нужна страница ]
- ^ Пармачек М.С., Эпштейн Дж.А. (июль 2009 г.). «Обновление кардиомиоцитов» . Медицинский журнал Новой Англии . 361 (1): 86–88. doi : 10.1056/NEJMcibr0903347 . ПМЦ 4111249 . ПМИД 19571289 .
- ^ Менделл Дж. Э., Клементс К. Д., Чоат Дж. Х., Ангерт Э. Р. (май 2008 г.). «Чрезвычайная полиплоидия у крупной бактерии» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (18): 6730–6734. Бибкод : 2008PNAS..105.6730M . дои : 10.1073/pnas.0707522105 . ПМЦ 2373351 . ПМИД 18445653 .
- ^ Эджер П.П., Пирес Х.К. (2009). «Дупликация генов и геномов: влияние чувствительности к дозировке на судьбу ядерных генов» . Хромосомные исследования . 17 (5): 699–717. дои : 10.1007/s10577-009-9055-9 . ПМИД 19802709 .
- ^ Jump up to: а б Кларк Дж.Т., Ллойд Г.Т., Фридман М. (октябрь 2016 г.). «Мало доказательств усиленной фенотипической эволюции ранних костистых рыб по сравнению с их живой сестринской группой ископаемых» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (41): 11531–11536. Бибкод : 2016PNAS..11311531C . дои : 10.1073/pnas.1607237113 . ПМК 5068283 . ПМИД 27671652 .
- ^ Белый МЮ (1973). Хромосомы (6-е изд.). Лондон: Чепмен и Холл. п. 28.
- ^ Стеббинс Г.Л. (1950). «Глава XII: Кариотип». Изменчивость и эволюция растений . Нью-Йорк, штат Нью-Йорк: Издательство Колумбийского университета. [ нужна страница ]
- ^ Отто С.П., Уиттон Дж (2000). «Заболеваемость и эволюция полиплоидов». Ежегодный обзор генетики . 34 (1): 401–437. CiteSeerX 10.1.1.323.1059 . дои : 10.1146/annurev.genet.34.1.401 . ПМИД 11092833 .
- ^ Jump up to: а б Смит Л.Е. (октябрь 2012 г.). «Предложение медицинским библиотекарям. 1920 год» . Журнал Ассоциации медицинских библиотек . 100 (4 дополнения): B. doi : 10.1023/B:RFBF.0000033049.00668.fe . ПМЦ 3571666 . ПМИД 23509424 . S2CID 11649126 .
- ^ Каннателла, округ Колумбия, Де Са РО (1993). « Xenopus laevis как модельный организм». Общество биологов-систематиков . 42 (4): 476–507. дои : 10.1093/sysbio/42.4.476 .
- ^ Богарт Дж. П., Би К., Фу Дж., Ноубл Д.В., Недзвецки Дж. (февраль 2007 г.). «Однополые саламандры (род Ambystoma) представляют новый репродуктивный способ для эукариот». Геном . 50 (2): 119–136. дои : 10.1139/g06-152 . ПМИД 17546077 . S2CID 42528911 .
- ^ Эпштейн CJ (февраль 1967 г.). «Размер клеток, содержание ядер и развитие полиплоидии в печени млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 57 (2): 327–334. Бибкод : 1967PNAS...57..327E . дои : 10.1073/pnas.57.2.327 . ПМК 335509 . ПМИД 16591473 .
- ^ Донн Р., Саруль-Айнама М., Кордье П., Селтон-Моризур С., Десдуэ К. (июль 2020 г.). «Полиплоидия в развитии печени, гомеостазе и заболеваниях» (PDF) . Обзоры природы. Гастроэнтерология и гепатология . 17 (7): 391–405. дои : 10.1038/s41575-020-0284-x . ПМИД 32242122 . S2CID 214766734 .
- ^ Галлардо М.Х., Гонсалес Калифорния, Себриан I (август 2006 г.). «Молекулярная цитогенетика и аллотетраплоидия у красной вискачевой крысы, Tympanoctomys barrerae (Rodentia, Octodontidae)» . Геномика . 88 (2): 214–221. дои : 10.1016/j.ygeno.2006.02.010 . ПМИД 16580173 .
- ^ Свартман М., Стоун Дж., Станьон Р. (апрель 2005 г.). «Молекулярная цитогенетика отвергает полиплоидию у млекопитающих». Геномика . 85 (4): 425–430. дои : 10.1016/j.ygeno.2004.12.004 . ПМИД 15780745 .
- ^ Сваруп Х (1956). «Продуцирование гетероплоидии у трехиглой колюшки Gasterosteus aculeatus (L.)». Природа . 178 (4542): 1124–1125. Бибкод : 1956Natur.178.1124S . дои : 10.1038/1781124a0 . S2CID 4226624 .
- ^ Сваруп Х (1959). «Продуцирование триплоидии у Gasterosteus aculeatus (L.)». Журнал генетики . 56 (2): 129–142. дои : 10.1007/BF02984740 . S2CID 42913498 .
- ^ Мейбл Б.К., Александру М.А., Тейлор М.И. (2011). «Дупликация генома у амфибий и рыб: расширенный синтез» . Журнал зоологии . 284 (3): 151–182. дои : 10.1111/j.1469-7998.2011.00829.x . S2CID 58937352 .
- ^ Jump up to: а б «Нобелевская премия по физиологии и медицине 2012 года присуждена за открытие того, что зрелые клетки можно перепрограммировать, чтобы они стали плюрипотентными» . ScienceDaily . 8 октября 2012 г.
- ^ Велицкий П., Мейнхардт Г., Плессл К., Вондра С., Вайс Т., Хаслингер П. и др. (октябрь 2018 г.). «Амплификация генома и клеточное старение являются отличительными чертами развития плаценты человека» . ПЛОС Генетика . 14 (10): e1007698. дои : 10.1371/journal.pgen.1007698 . ПМК 6200260 . ПМИД 30312291 .
- ^ Винкельманн М., Пфитцер П., Шнайдер В. (декабрь 1987 г.). «Значение полиплоидии мегакариоцитов и других клеток для здоровья и опухолевых заболеваний». Клинический еженедельник . 65 (23): 1115–1131. дои : 10.1007/BF01734832 . ПМИД 3323647 . S2CID 23496028 .
- ^ «Триплоидия» . Национальная организация редких заболеваний . Проверено 23 декабря 2018 г.
- ^ Jump up to: а б Бейкер П., Монга А., Бейкер П. (2006). Гинекология десяти учителей . Лондон: Арнольд. ISBN 978-0-340-81662-2 .
- ^ Бранкати Ф, Мингарелли Р, Даллапиккола Б (декабрь 2003 г.). «Рекуррентная триплоидия материнского происхождения» . Европейский журнал генетики человека . 11 (12): 972–974. дои : 10.1038/sj.ejhg.5201076 . ПМИД 14508508 . S2CID 28009278 .
- ^ Уик Дж.Б., Джонсон К.Дж., О'Брайен Дж., Уик М.Дж. (май 2013 г.). «Диагностика триплоидии во втором триместре: серия из четырех случаев» . Об этом сообщает AJP . 3 (1): 37–40. дои : 10.1055/s-0032-1331378 . ПМЦ 3699153 . ПМИД 23943708 .
- ^ Мейерс Л.А., Левин Д.А. (июнь 2006 г.). «О численности полиплоидов у цветковых растений» . Эволюция; Международный журнал органической эволюции . 60 (6): 1198–1206. дои : 10.1111/j.0014-3820.2006.tb01198.x . ПМИД 16892970 .
- ^ Jump up to: а б Ризеберг Л.Х., Уиллис Дж.Х. (август 2007 г.). «Видообразование растений» . Наука . 317 (5840): 910–914. Бибкод : 2007Sci...317..910R . дои : 10.1126/science.1137729 . ПМК 2442920 . ПМИД 17702935 .
- ^ Отто СП (ноябрь 2007 г.). «Эволюционные последствия полиплоидии» . Клетка . 131 (3): 452–462. дои : 10.1016/j.cell.2007.10.022 . ПМИД 17981114 . S2CID 10054182 .
- ^ Инициатива «Тысяча транскриптомов растений» (октябрь 2019 г.). «Тысяча растительных транскриптомов и филогеномика зеленых растений» . Природа . 574 (7780): 679–685. дои : 10.1038/s41586-019-1693-2 . ПМК 6872490 . PMID 31645766 .
- ^ Де Бодт С., Маере С., Ван де Пер Ю. (ноябрь 2005 г.). «Дупликация генома и происхождение покрытосеменных». Тенденции в экологии и эволюции . 20 (11): 591–597. дои : 10.1016/j.tree.2005.07.008 . ПМИД 16701441 .
- ^ Вуд Т.Э., Такебаяши Н., Баркер М.С., Мэйроуз И., Гринспун П.Б., Ризеберг Л.Х. (август 2009 г.). «Частота полиплоидного видообразования у сосудистых растений» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (33): 13875–13879. Бибкод : 2009PNAS..10613875W . дои : 10.1073/pnas.0811575106 . JSTOR 40484335 . ПМЦ 2728988 . ПМИД 19667210 .
- ^ Сюй X, Пан С., Ченг С., Чжан Б., Му Д., Ни П. и др. (июль 2011 г.). «Секвенирование генома и анализ клубнеплодов картофеля» . Природа . 475 (7355): 189–195. дои : 10.1038/nature10158 . ПМИД 21743474 .
- ^ Осборн Т.С., Пирес Дж.К., Бирчлер Дж.А., Огер Д.Л., Чен З.Дж., Ли Х.С. и др. (март 2003 г.). «Понимание механизмов экспрессии новых генов у полиплоидов». Тенденции в генетике . 19 (3): 141–147. дои : 10.1016/S0168-9525(03)00015-5 . ПМИД 12615008 .
- ^ Чен ZJ, Ni Z (март 2006 г.). «Механизмы геномных перестроек и изменения экспрессии генов у полиплоидов растений» . Биоэссе . 28 (3): 240–252. doi : 10.1002/bies.20374 . ЧВК 1986666 . ПМИД 16479580 .
- ^ Чен ZJ (2007). «Генетические и эпигенетические механизмы экспрессии генов и фенотипических вариаций полиплоидов растений» . Ежегодный обзор биологии растений . 58 : 377–406. doi : 10.1146/annurev.arplant.58.032806.103835 . ЧВК 1949485 . ПМИД 17280525 .
- ^ Альбертен В., Баллио Т., Брабант П., Шевр А.М., Эбер Ф., Малосс К., Тиллеман Х. (июнь 2006 г.). «Многочисленные и быстрые нестохастические модификации генных продуктов в недавно синтезированных аллотетраплоидах Brassica napus» . Генетика . 173 (2): 1101–1113. дои : 10.1534/genetics.106.057554 . ПМЦ 1526534 . ПМИД 16624896 .
- ^ Пеннингтон П.Д., Коста Л.М., Гутьеррес-Маркос Х.Ф., Гренландия А.Дж., Дикинсон Х.Г. (апрель 2008 г.). «Когда сталкиваются геномы: аномальное развитие семян после интерплоидных скрещиваний кукурузы» . Анналы ботаники . 101 (6): 833–843. дои : 10.1093/aob/mcn017 . ПМК 2710208 . ПМИД 18276791 .
- ^ фон Вангенхайм К.Х., Петерсон Х.П. (июнь 2004 г.). «Аберрантное развитие эндосперма при интерплоидных скрещиваниях выявляет таймер дифференцировки». Биология развития . 270 (2): 277–289. дои : 10.1016/j.ydbio.2004.03.014 . ПМИД 15183714 .
- ^ Уиттон Дж., Сирс СиДжей, Мэддисон В.П. (декабрь 2017 г.). «Совместное появление родственных бесполых, но не половых линий предполагает, что репродуктивное вмешательство ограничивает сосуществование» . Слушания. Биологические науки . 284 (1868): 20171579. doi : 10.1098/rspb.2017.1579 . ПМК 5740271 . ПМИД 29212720 .
- ^ Халаби К., Шафир А., Мэйроуз I (июнь 2023 г.). «PloiDB: база данных плоидности растений» . Новый фитолог . 240 (3): 918–927. дои : 10.1111/nph.19057 . ПМИД 37337836 .
- ^ Ахуджа М.Р., Нил Д.Б. (2002). «Происхождение полиплоидии прибрежного секвойи ( Sequoia sempervirens (D. Don) Endl.) и связь прибрежного секвойи с другими родами таксодиевых». Сильва Генетика . 51 : 2–3.
- ^ Лес Д.Х., Филбрик, Коннектикут (1993). «Исследования гибридизации и изменения числа хромосом у водных покрытосеменных: эволюционные последствия». Водная ботаника . 44 (2–3): 181–228. Бибкод : 1993AqBot..44..181L . дои : 10.1016/0304-3770(93)90071-4 .
- ^ «Фрукты без косточек делают других ненужными» . Архивировано из оригинала 03 февраля 2019 г. Проверено 7 марта 2022 г.
- ^ Эмшвиллер Э (2006). «Происхождение полиплоидных культур: на примере октаплоидной клубневой культуры Oxalis tuberosa ». Зедер М.А., Декер-Уолтерс Д., Эмшвиллер Э., Брэдли Д., Смит Б.Д. (ред.). Документирование одомашнивания: новые генетические и археологические парадигмы . Беркли, Калифорния: Издательство Калифорнийского университета.
- ^ Ле Кунф Л., Гарсмер О., Рабуан Л.М., Поке Ж., Телисмарт Х., Селви А. и др. (сентябрь 2008 г.). «Диплоидное/полиплоидное синтенное челночное картирование и специфическая для гаплотипа хромосома, идущая к гену устойчивости к ржавчине (Bru1) в высокополиплоидном сахарном тростнике (2n примерно 12x примерно 115)» . Генетика . 180 (1): 649–660. дои : 10.1534/genetics.108.091355 . ПМЦ 2535714 . ПМИД 18757946 .
- ^ Jump up to: а б с Альбертин В., Марулло П. (июль 2012 г.). «Полиплоидия грибов: эволюция после полногеномной дупликации» . Слушания. Биологические науки . 279 (1738): 2497–2509. дои : 10.1098/rspb.2012.0434 . ПМК 3350714 . ПМИД 22492065 .
- ^ Эмерсон Р., Уилсон К.М. (1954). «Межвидовые гибриды, цитогенетика и цитотаксономия эуалломицетов». Микология . 46 (4): 393–434. дои : 10.1080/00275514.1954.12024382 . JSTOR 4547843 .
- ^ Альбертин В., Марулло П., Эгль М., Бурге А., Белый М., Диллманн С. и др. (ноябрь 2009 г.). «Доказательства аутотетраплоидии, связанной с репродуктивной изоляцией у Saccharomyces cerevisiae: на пути к новому одомашненному виду» . Журнал эволюционной биологии . 22 (11): 2157–2170. дои : 10.1111/j.1420-9101.2009.01828.x . ПМИД 19765175 .
- ^ Лу до нашей эры (1964). «Полиплоидия базидиомицета Cyathus stercoreus ». Американский журнал ботаники . 51 (3): 343–347. дои : 10.2307/2440307 . JSTOR 2440307 .
- ^ Либкинд Д., Хиттингер К.Т., Валерио Э., Гонсалвес С., Довер Дж., Джонстон М. и др. (август 2011 г.). «Одомашнивание микробов и идентификация дикого генетического фонда лагерных дрожжей» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (35): 14539–14544. Бибкод : 2011PNAS..10814539L . дои : 10.1073/pnas.1105430108 . ПМК 3167505 . ПМИД 21873232 .
- ^ Борнеман А.Р., Цеппель Р., Чемберс П.Дж., Кертин К.Д. (февраль 2014 г.). «Взгляд на геномный ландшафт Dekkera bruxellensis: сравнительная геномика выявляет различия в плоидности и потенциале использования питательных веществ среди винных изолятов» . ПЛОС Генетика . 10 (2): e1004161. дои : 10.1371/journal.pgen.1004161 . ПМЦ 3923673 . ПМИД 24550744 .
- ^ Ма Л.Дж., Ибрагим А.С., Скори С., Грабхерр М.Г., Бургер Г., Батлер М. и др. (июль 2009 г.). Мадхани HD (ред.). «Геномный анализ гриба базовой линии Rhizopus oryzae выявил дупликацию всего генома» . ПЛОС Генетика . 5 (7): e1000549. дои : 10.1371/journal.pgen.1000549 . ПМК 2699053 . ПМИД 19578406 .
- ^ Вонг С., Батлер Г., Вулф К.Х. (июль 2002 г.). «Эволюция порядка генов и палеополиплоидия у гемиаскомицетных дрожжей» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (14): 9272–9277. Бибкод : 2002PNAS...99.9272W . дои : 10.1073/pnas.142101099 . JSTOR 3059188 . ПМЦ 123130 . ПМИД 12093907 .
- ^ Девиер Б., Агилета Г., Худ М.Э., Жиро Т. (2009). «Использование филогении генов рецепторов феромонов в комплексе видов Microbotryum violaceum для исследования возможного видообразования путем гибридизации». Микология . 102 (3): 689–696. дои : 10.3852/09-192 . ПМИД 20524600 . S2CID 9072505 .
- ^ Данн Б., Шерлок Дж. (октябрь 2008 г.). «Реконструкция происхождения генома и эволюции гибридных лагерных дрожжей Saccharomyces Pastorianus» . Геномные исследования . 18 (10): 1610–1623. дои : 10.1101/гр.076075.108 . ПМЦ 2556262 . ПМИД 18787083 .
- ^ Накао Й., Канамори Т., Ито Т., Кодама Й., Райниери С., Накамура Н. и др. (апрель 2009 г.). «Последовательность генома лагерных пивоваренных дрожжей, межвидового гибрида» . Исследование ДНК . 16 (2): 115–129. дои : 10.1093/dnares/dsp003 . ПМЦ 2673734 . ПМИД 19261625 .
- ^ Сканнелл Д.Р., Бирн К.П., Гордон Дж.Л., Вонг С., Вулф К.Х. (март 2006 г.). «Множественные раунды видообразования, связанные с реципрокной потерей генов у полиплоидных дрожжей». Природа . 440 (7082): 341–345. Бибкод : 2006Natur.440..341S . дои : 10.1038/nature04562 . HDL : 2262/22660 . ПМИД 16541074 . S2CID 94448 .
- ^ Минато Т., Ёсида С., Исигуро Т., Симада Э., Мизутани С., Кобаяши О., Ёсимото Х. (март 2009 г.). «Профиль экспрессии ортологичных генов дрожжей низового брожения Saccharomyces Pastorianus с использованием олигонуклеотидных микрочипов». Дрожжи . 26 (3): 147–165. дои : 10.1002/да.1654 . ПМИД 19243081 . S2CID 23349682 .
- ^ Лидзбарский Г.А., Школьник Т, Нево Е (июнь 2009 г.). Иднурма А (ред.). «Адаптивный ответ на агенты, повреждающие ДНК, в природных популяциях Saccharomyces cerevisiae из «Каньона Эволюции», гора Кармель, Израиль» . ПЛОС ОДИН . 4 (6): е5914. Бибкод : 2009PLoSO...4.5914L . дои : 10.1371/journal.pone.0005914 . ПМЦ 2690839 . ПМИД 19526052 .
- ^ Иоос Р., Андриё А., Марсе Б., Фрей П. (июль 2006 г.). «Генетическая характеристика природного гибридного вида Phytophthora alni, полученная на основе анализа ядерной и митохондриальной ДНК» (PDF) . Грибковая генетика и биология . 43 (7): 511–529. дои : 10.1016/j.fgb.2006.02.006 . ПМИД 16626980 . S2CID 43275127 .
- ^ Филлипс Н., Капраун Д.Ф., Гомес Гаррета А., Рибера Сигуан М.А., Рулл Дж.Л., Сальвадор Солер Н. и др. (2011). «Оценки содержания ядерной ДНК у 98 видов бурых водорослей (Phaeophyta)» . Растения АОБ . 2011 : plr001. дои : 10.1093/aobpla/plr001 . ПМК 3064507 . ПМИД 22476472 .
- ^ Чепурнов В.А., Манн Д.Г., Выверман В., Саббе К., Даниэлидис Д.Б. (2002). «Половое размножение, система спаривания и динамика протопластов Seminavis (Bacillariophyceae)». Журнал психологии . 38 (5): 1004–1019. Бибкод : 2002JPcgy..38.1004C . дои : 10.1046/j.1529-8817.2002.t01-1-01233.x . S2CID 84299819 .
- ^ Аури Дж.М., Жайлон О., Дюре Л., Ноэль Б., Джубин С., Порсель Б.М. и др. (ноябрь 2006 г.). «Глобальные тенденции полногеномных дупликаций, выявленные инфузорией Paramecium Tetraurelia» . Природа . 444 (7116): 171–178. Бибкод : 2006Natur.444..171A . дои : 10.1038/nature05230 . ПМИД 17086204 .
- ^ Хансен М.Т. (апрель 1978 г.). «Множественность эквивалентов генома радиационно-устойчивой бактерии Micrococcus radiodurans» . Журнал бактериологии . 134 (1): 71–75. дои : 10.1128/JB.134.1.71-75.1978 . ПМК 222219 . ПМИД 649572 .
- ^ Заградка К., Слейд Д., Бэйлоне А., Соммер С., Авербек Д., Петранович М. и др. (октябрь 2006 г.). «Повторная сборка разрушенных хромосом у Deinococcus radiodurans». Природа . 443 (7111): 569–573. Бибкод : 2006Natur.443..569Z . дои : 10.1038/nature05160 . ПМИД 17006450 . S2CID 4412830 .
- ^ Нагпал П., Джафри С., Редди М.А., Дас Х.К. (июнь 1989 г.). «Множественные хромосомы Azotobacter vinelandii» . Журнал бактериологии . 171 (6): 3133–3138. дои : 10.1128/jb.171.6.3133-3138.1989 . ПМК 210026 . ПМИД 2785985 .
- ^ Мальдонадо Р., Хименес Х., Касадесус Х. (июль 1994 г.). «Изменения плоидности в ходе цикла роста Azotobacter vinelandii» . Журнал бактериологии . 176 (13): 3911–3919. дои : 10.1128/jb.176.13.3911-3919.1994 . ПМК 205588 . ПМИД 8021173 .
- ^ Соппа Дж. (январь 2011 г.). «Плоидия и конверсия генов у архей». Труды Биохимического общества . 39 (1): 150–154. дои : 10.1042/BST0390150 . ПМИД 21265763 . S2CID 31385928 .
- ^ Коттеманн М., Киш А., Илоануси С., Бьорк С., ДиРуджеро Дж. (июнь 2005 г.). «Физиологические реакции галофильных архей Halobacterium sp., штамм NRC1, на высыхание и гамма-облучение» (PDF) . Экстремофилы . 9 (3): 219–227. дои : 10.1007/s00792-005-0437-4 . ПМИД 15844015 . S2CID 8391234 .
- ^ ДеВо Л.К., Мюллер Дж.А., Смит Дж., Петриско Дж., Уэллс Д.П., ДасСарма С. (октябрь 2007 г.). «Чрезвычайно радиационно-устойчивые мутанты галофильных архей с повышенной экспрессией гена одноцепочечного ДНК-связывающего белка (RPA)» . Радиационные исследования . 168 (4): 507–514. Бибкод : 2007РадР..168..507Д . дои : 10.1667/RR0935.1 . ПМИД 17903038 . S2CID 22393850 .
Дальнейшее чтение
[ редактировать ]- Снустад Д.П., Симмонс М.Дж. (2006). Принципы генетики (4-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 978-0-471-69939-2 .
- Инициатива по геному арабидопсиса (декабрь 2000 г.). «Анализ последовательности генома цветкового растения Arabidopsis thaliana» . Природа . 408 (6814): 796–815. Бибкод : 2000Natur.408..796T . дои : 10.1038/35048692 . ПМИД 11130711 .
- Икин Г.С., Берингер Р.Р. (декабрь 2003 г.). «Развитие тетраплоида у мышей» . Динамика развития . 228 (4): 751–766. дои : 10.1002/dvdy.10363 . ПМИД 14648853 .
- Гаэта РТ, Пирес Х.К., Инигес-Луй Ф., Леон Э., Осборн Т.К. (ноябрь 2007 г.). «Геномные изменения у ресинтезированных Brassica napus и их влияние на экспрессию генов и фенотип» . Растительная клетка . 19 (11): 3403–3417. дои : 10.1105/tpc.107.054346 . ПМК 2174891 . ПМИД 18024568 .
- Грегори Т.Р., Мейбл Б.К. (2005). «Полиплоидия у животных». У Григория Т.Р. (ред.). Эволюция генома . Сан-Диего, Калифорния: Эльзевир. стр. 427–517.
- Жайон О, Ори Дж.М., Брюне Ф., Пети Дж.Л., Штанге-Томанн Н., Маусели Э. и др. (октябрь 2004 г.). «Дупликация генома костистых рыб Tetraodon nigroviridis выявляет ранний протокариотип позвоночных» . Природа . 431 (7011): 946–957. Бибкод : 2004Natur.431..946J . дои : 10.1038/nature03025 . ПМИД 15496914 .
- Патерсон А.Х., Бауэрс Дж.Э., Ван де Пер Ю., Вандеполе К. (март 2005 г.). «Древнее дублирование геномов злаков» . Новый фитолог . 165 (3): 658–661. дои : 10.1111/j.1469-8137.2005.01347.x . ПМИД 15720677 .
- Раес Дж., Вандеполе К., Симиллион С., Саейс Ю., Ван де Пер Ю. (2003). «Исследование древних событий дупликации в геноме арабидопсиса». Журнал структурной и функциональной геномики . 3 (1–4): 117–129. дои : 10.1023/А:1022666020026 . ПМИД 12836691 . S2CID 9666357 .
- Симиллион С., Вандеполе К., Ван Монтегю М.К., Забо М., Ван де Пер Ю. (октябрь 2002 г.). «Скрытое дублирование Arabidopsis thaliana в прошлом» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (21): 13627–13632. Бибкод : 2002PNAS...9913627S . дои : 10.1073/pnas.212522399 . JSTOR 3073458 . ПМК 129725 . ПМИД 12374856 .
- Солтис Д.Э., Солтис П.С. , Шемске Д.В., Хэнкок Дж.Ф., Томпсон Дж.Н., Муж BC, Джадд В.С. (2007). «Аутополиплоидия покрытосеменных: сильно ли мы недооценили количество видов?». Таксон . 56 (1): 13–30. JSTOR 25065732 .
- Солтис Д.Э., Баггс Р.Дж., Дойл Дж.Дж., Солтис П.С. (2010). «Чего мы еще не знаем о полиплоидии» . Таксон . 59 (5): 1387–1403. дои : 10.1002/tax.595006 . JSTOR 20774036 .
- Тейлор Дж.С., Брааш И., Фрики Т., Мейер А., Ван де Пер Ю. (март 2003 г.). «Дублирование генома — черта, присущая 22 000 видам лучепёрых рыб» . Геномные исследования . 13 (3): 382–390. дои : 10.1101/гр.640303 . ПМК 430266 . ПМИД 12618368 .
- Тейт Дж.А., Солтис Д.Э., Солтис П.С. (2005). «Полиплоидия у растений». У Григория Т.Р. (ред.). Эволюция генома . Сан-Диего, Калифорния: Эльзевир. стр. 371–426.
- Ван де Пер Ю., Тейлор Дж. С., Мейер А. (2003). «Все ли рыбы древние полиплоиды?» . Журнал структурной и функциональной геномики . 3 (1–4): 65–73. дои : 10.1023/А:1022652814749 . ПМИД 12836686 . S2CID 14092900 .
- Ван де Пер Ю (2004). «Геном тетраодона подтверждает выводы Такифугу: большинство рыб являются древними полиплоидами» . Геномная биология . 5 (12): 250. doi : 10.1186/gb-2004-5-12-250 . ПМЦ 545788 . ПМИД 15575976 .
- Ван де Пер Ю, Мейер А (2005). «Крупномасштабные дупликации генов и древних геномов». У Григория Т.Р. (ред.). Эволюция генома . Сан-Диего, Калифорния: Эльзевир. стр. 329–368.
- Вулф К.Х., Шилдс, округ Колумбия (июнь 1997 г.). «Молекулярные доказательства древнего дублирования всего генома дрожжей» . Природа . 387 (6634): 708–713. Бибкод : 1997Natur.387..708W . дои : 10.1038/42711 . ПМИД 9192896 . S2CID 4307263 .
- Вулф К.Х. (май 2001 г.). «Вчерашние полиплоиды и тайна диплоидизации». Обзоры природы. Генетика . 2 (5): 333–341. дои : 10.1038/35072009 . ПМИД 11331899 . S2CID 20796914 .
Внешние ссылки
[ редактировать ]- Полиплоидия на страницах биологии Кимбалла
- Портал полиплоидии - это редактируемый сообществом проект, содержащий информацию, исследования, образование и библиографию о полиплоидии.